

Abstract
The biosynthesis of lignin in vascular plants is regulated both developmentally and environmentally. In the inflorescence stems of Arabidopsis, lignin is mainly deposited in the walls of xylem cells and interfascicular fiber cells during normal plant growth and development. The mechanisms controlling the spatial deposition of lignin remain unknown. By screening ethyl methanesulfonate-mutagenized populations of Arabidopsis, we have isolated two allelic elp1 (ectopic deposition of lignin in pith) mutants with altered lignin deposition patterns. In elp1 stems, lignin was ectopically deposited in the walls of pith parenchyma cells in addition to its normal deposition in the walls of xylem and fiber cells. Lignin appeared to be deposited in patches of parenchyma cells in the pith of both young and mature elp1 stems. The ectopic deposition of lignin in the pith of elp1 stems was accompanied by an increase in the activities of enzymes in the lignin biosynthetic pathway and with the ectopic expression of caffeoyl coenzyme A O-methyltransferase in pith cells. These results indicate that the ELP1 locus is involved in the repression of the lignin biosynthetic pathway in the pith. Isolation of the elp1 mutants provides a novel means with which to study the molecular mechanisms underlying the spatial control of lignification.
Lignin, a complex phenylpropanoid polymer, is primarily deposited in the walls of cells that have secondary wall thickening, such as tracheary elements and fibers. Lignin provides mechanical strength in the walls of these sclerenchyma cells. It has been shown that lignin is derived from the dehydrogenative polymerization of the monolignols p-coumaryl alcohol, coniferyl alcohol, and sinapyl alcohol. These monolignols are synthesized through a phenylpropanoid biosynthetic pathway (Whetten and Sederoff, 1995). The biosynthesis of lignin is temporally and spatially controlled, and is closely associated with the differentiation of sclerenchyma cells during normal plant growth and development. Lignin biosynthesis can also be induced in response to wounding or infection, a process independent of developmentally regulated lignification. Although the lignin biosynthetic pathway has been intensively investigated in the past several decades, our understanding of the molecular mechanisms underlying the temporal and spatial control of lignin biosynthesis is still in its infancy.
The regulation of lignin biosynthesis has mainly been studied by promoter analysis. A number of genes in the phenylpropanoid pathway have been cloned and the activities of their promoters analyzed. The promoters of genes such as Phe ammonia-lyase (PAL; Bevan et al., 1989; Ohl et al., 1990; Hauffe et al., 1991; Levya et al., 1992), cinnamic acid 4-hydroxylase (Bell-Lelong et al., 1997), hydroxycinnamate:CoA ligase (4CL; Douglas et al., 1991), caffeic acid O-methyltransferase (CAOMT; Capellades et al., 1996), and hydroxycinnamyl alcohol dehydrogenase (Feuillet et al., 1995) all directed the expression of GUS reporter gene in lignifying tissues. Deletion analyses in the promoters of PAL and 4CL genes identified cis-acting elements critical for tissue-specific expression (Hauffe et al., 1991; Levya et al., 1992). These elements, called the AC elements, were also conserved in the promoters of other genes in the phenylpropanoid pathway. In addition to the conserved AC elements, other cis-elements, such as the FP56 element identified in the 4CL promoter, have also been shown to be important for the regulation of gene expression (Neustaedter et al., 1999).
The AC elements appear to be similar to the DNA motifs recognized by plant Myb transcription factors. The first evidence for the role of Myb proteins in the regulation of genes in the phenylpropanoid pathway came from the study of a flower-specific Myb protein from Antirrhinum. It was shown that the Myb protein was able to bind to the AC elements and turn on the expression of a reporter gene driven by the PAL promoter (Sablowski et al., 1994, 1995). Recently, it has been shown that overexpression of two Antirrhinum Myb genes in transgenic tobacco plants down-regulated the expression of certain genes in the phenylpropanoid biosynthetic pathway, which resulted in a reduction in lignin accumulation (Tamagnone et al., 1998). This indicates that Myb-related transcription factors might be involved in regulating the expression of genes in the lignin biosynthetic pathway.
The lignin biosynthesis has also been studied by mutational analysis. A number of natural lignin mutants, called brown midrib (bm) mutants, which show characteristic brown pigmentation in the midrib of the leaf, have been reported in maize, sorghum, and pearl millet (for review, see Campbell and Sederoff, 1996). Analysis of lignin compositions showed that some bm mutants had a decreased syringyl lignin content (Akin et al., 1986). The selective reduction in syringyl lignin in the maize bm3 mutant was most likely the result of decreased CAOMT activity (Grand et al., 1985) due to a mutation in the CAOMT gene (Vignols et al., 1995). In addition to a reduction in CAOMT activity, the sorghum bmr-6 mutant also showed decreased hydroxycinnamyl alcohol dehydrogenase activity, which may result in the incorporation of cinnamaldehydes into the mutant lignin (Pillonel et al., 1991). A close association of the mutation with the hydroxycinnamyl alcohol dehydrogenase gene has recently been demonstrated in the maize bm1 mutant (Halpin et al., 1998). During the screening of mutants lacking sinapoyl malate, Chapple et al. (1992) isolated an Arabidopsis mutant defective in the conversion of ferulic acid into 5-hydroxyferulic acid. Consequently, the mutant lacks syringyl lignin in the lignified fibers of the stems. Recently, the gene encoding ferulic acid 5-hydroxylase which converts ferulic acid into 5-hydroxyferulic acid has been cloned by T-DNA tagging in Arabidopsis (Meyer et al., 1996). In summary, it appears that all lignin mutants studied so far are defective in genes in the lignin biosynthetic pathway, and no mutants affecting the spatial control of lignin deposition have been reported.
We have used the model plant Arabidopsis to study how the deposition of lignin is spatially controlled. Histochemical staining of lignin with phloroglucinol-HCl was employed to screen for mutants with altered lignin deposition patterns in the inflorescence stems of Arabidopsis. Lignin in Arabidopsis has been shown to contain both the guaiacyl lignin unit and the syringyl lignin unit (Dharmawardhana et al., 1992). We report the isolation of elp1 (ectopic deposition of lignin in pith) mutants with ectopic deposition of lignin in pith cell walls in addition to its normal deposition in xylem cells and fibers. We propose that the ELP1 locus might be involved in the repression of lignin biosynthesis in pith cells. Isolation of the elp1 mutants will allow us to further study how the coordinate regulation of genes in the lignin biosynthetic pathway is controlled.
RESULTS
Spatial Deposition of Lignin in the Inflorescence Stems of Wild-Type Arabidopsis
To examine the pattern of lignin deposition in the stems of wild-type Arabidopsis plants, we prepared thin sections from the stems and stained them with the lignin-staining dye phloroglucinol-HCl to show lignin (Fig. 1). The use of phloroglucinol-HCl staining was intended to highlight those cell walls with lignin deposition, which was the main focus of this study, although the anatomy of non-lignified cells was not well revealed. In the top part of the stem, where the only sclerenchyma cells present were xylem vessel elements, lignin staining was only seen in the xylem (Fig. 1A). In the middle part of the stem, another type of sclerenchyma cells, interfascicular fibers, were present in addition to xylem cells. Accordingly, lignin was seen in both interfascicular fibers and xylem cells (Fig. 1B). In the lower part of the stem, more xylem cells and interfascicular fibers were formed. Consistently, intense lignin staining was seen in these sclerenchyma cells (Fig. 1C). No lignin staining was detected in the cortical cells or the pith parenchyma cells throughout the stems. This was also true in the stems of mature plants (data not shown). This indicates that lignin is specifically deposited in xylem cells and fibers in Arabidopsis stems during normal plant growth and development.
Figure 1.
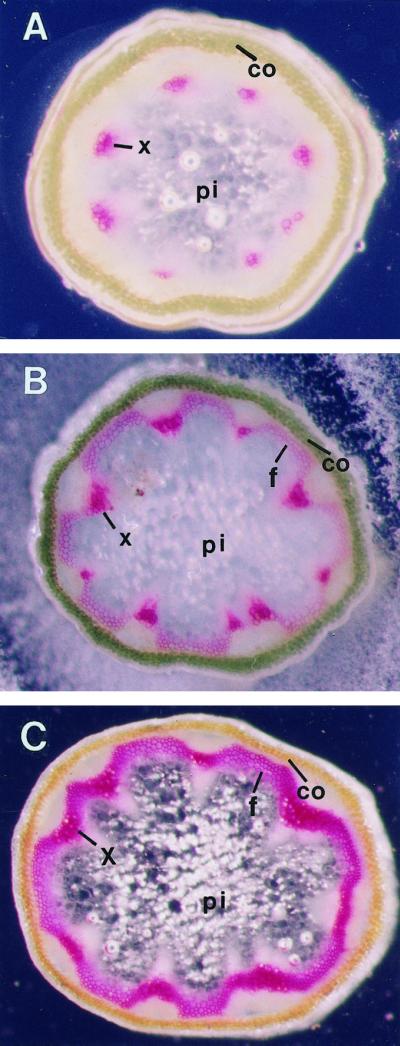
Lignification in the inflorescence stems of wild-type Arabidopsis plants. Inflorescence stems from 6-week-old plants were sectioned and sections were stained for lignin with phloroglucinol-HCl. Lignin staining is red. Pictures were taken under a dissection microscope with dark-field illumination. A, Section from the top part of the stem. Lignin was only seen in xylem cells. B, Section from the middle part of the stem. Lignin staining was heavy in xylem cells but light in interfascicular fibers. C, Section from the basal part of the stem. The lignin staining was heavy in both xylem cells and interfascicular fibers. co, Cortex; f, interfascicular fiber; pi, pith; x, xylem. Magnification, ×52.
Isolation of Mutants with an Altered Lignin Deposition Pattern
To investigate the mechanisms controlling the spatial deposition of lignin, we screened for mutants with altered lignin deposition patterns from the ethyl methanesulfonate-mutagenized M2 populations of Arabidopsis. Free-hand sections of stems were stained for lignin with phloroglucinol-HCl. Of about 10,000 plants screened, we found two mutants with a dramatic alteration in lignin staining pattern. In the stems of both mutants, lignin was ectopically present in the pith cells (Fig. 2) in addition to its normal deposition in xylem cells and fibers. No lignin staining was detected in cortex or epidermal cells (Fig. 2). The presence of lignin in pith cells was not only revealed by staining with phloroglucinol-HCl (Fig. 2, C and E), which stains total lignin, but was also visualized by staining with the Mäule reagent (Fig. 2, D and F), which preferentially stains syringyl lignin. Mäule reagent staining showed that lignin in interfascicular fibers was rich in syringyl lignin, whereas xylem cells lacked syringyl lignin (Fig. 2, B, D, and F). Because the Mäule reagent staining of pith cells was similar to the staining of interfascicular fibers (Fig. 2, D and F), the lignin composition in elp1 pith cells is similar to that in interfascicular fibers. A few lignified phloem fiber cells were present outside the vascular bundles (Fig. 2, C, D, and F). Phloem fiber cells were also observed in mature stems of wild-type plants (data not shown). No alteration in the lignin staining pattern was observed in leaves or roots of the mutants (data not shown).
Figure 2.

Ectopic deposition of lignin in the pith of elp1 stems. Basal parts of the stems of 6-week-old plants were sectioned and sections were stained for lignin with phloroglucinol-HCl (left column) and the Mäule reagent (right column). Phloroglucinol-HCl stains total lignin; the Mäule reagent stains syringyl lignin. Lignin was stained red with both reagents. A and B, Sections from a wild-type stem. Lignin was present in both xylem cells and interfascicular fibers but absent in the pith. Note that xylem cells were stained orange with the Mäule reagent due to the lack of syringyl lignin. C and D, Sections from an elp1-1 stem. Lignin was seen in the pith in addition to xylem cells and fibers. Like the lignin in the interfascicular fibers, the lignin in the pith was rich in syringyl lignin units, as revealed by staining with the Mäule reagent. E and F, Sections from an elp1-2 stem. Lignin was evident in the pith in addition to xylem cells and fibers. Similar to the elp1-1 mutant, the lignin in the pith was rich in syringyl lignin units, as revealed by staining with the Mäule reagent. co, Cortex; f, interfascicular fiber; pf, phloem fiber; pi, pith; x, xylem. Magnification, ×52.
To determine whether these two mutants were allelic, they were crossed with each other. Stems of the resulting F1 plants were examined for the lignin deposition pattern. Of the 20 F1 plants examined, they all showed lignin staining in the pith cells in addition to xylem cells and interfascicular fibers (data not shown), indicating that these two mutants could not complement each other and therefore were allelic. The mutant locus was named elp1.
Lignin Deposition Patterns in the Pith of the elp1 Mutants
We noticed that lignin staining was not present in all pith cells of elp1 stems (Fig. 2). We were then prompted to examine the lignin staining pattern throughout the mutant stems. An elp1-2 stem was divided into five equal segments, and sections from each segment were stained for lignin with phloroglucinol-HCl. As seen in Figure 3, although lignin staining in the pith was obvious in all sections, no common staining pattern was seen. In the top segment of the stem, a few patches of intense lignin staining were evident in the pith (Fig. 3A). Two sections from the second segment showed a stronger lignin staining in the pith (Fig. 3, B and C) compared with the section from the top segment (Fig. 3A). However, patches of lignin staining were present in different locations in the pith in these two sections (Fig. 3, B and C). Similar random occurrence of lignin staining in the pith was also observed in the sections from the lower segments (Fig. 3, D–F). Occasionally, some sections showed lignin staining in nearly all pith cells (Fig. 3E). No diffusion of lignin staining was noticed. The lignin staining pattern throughout elp1-1 stems was similar to that in the elp1-2 mutant (data not shown). These results indicate that the ectopic deposition of lignin in the pith of the elp1 mutants is mosaic with no common patterns even within the same stem.
Figure 3.

Lignification pattern in the pith throughout an elp1-2 stem. A stem from the elp1-2 mutant plant was divided into five equal segments, and thin sections from each segment were stained for lignin with phloroglucinol-HCl. Lignin staining is red. A, Section from the top segment of the stem. B and C, Sections with 1-cm apart from the second segment of the stem. D, Section from the third segment of the stem. E. Section from the fourth segment of the stem. F, Section from the bottom segment of the stem. Although lignin was seen in the pith cells in all sections, lignin distribution in the pith appeared to be mosaic. f, Interfascicular fiber; pf, phloem fiber; pi, pith; x, xylem. Magnification, ×52.
Lignin Content and Activities of Enzymes in the Lignin Pathway Increase in elp1 Stems
To confirm that the positive staining in pith by both phloroglucinol-HCl and Mäule reagent was a result of ectopic deposition of lignin, we analyzed Klason lignin content in stems of the wild-type and the elp1 mutants. We reasoned that lignin content could be increased in the mutants due to the extra lignin present in the pith. When walls of the inflorescence stems were used for Klason lignin analysis, it was found that the elp1 stems had about 20% more lignin than the wild-type stems (Fig. 4). The result was consistent with the histological staining showing the presence of extra lignin in the pith of the mutants.
Figure 4.

Lignin content and enzyme activity in elp1 stems. Mature inflorescence stems of 50 individual plants were pooled and assayed for Klason lignin content. Klason lignin in the mutants was expressed as a percentage of the wild type. Data for Klason lignin are the means ± se from three separate assays. Inflorescence stems from 6-week-old plants were collected and assayed for the activities of PAL, CCoAOMT, and CCR. The enzyme activity in the wild type was taken as 100, and the activity in the mutants was expressed as a percentage of the wild type. Data for enzyme activity are the means ± se from 10 plants.
During lignin biosynthesis, all of the genes in the lignin biosynthetic pathway have to be turned on. To investigate whether there was any change in the activities of lignin pathway enzymes in the elp1 mutants, we assayed the activities of PAL, CCoAOMT, and CCR, the enzymes involved in the lignin biosynthetic pathway. The specific activities of these enzymes in elp1 mutants were about 50% higher than those in the wild type (Fig. 4), which was consistent with the presence of extra lignin in the pith of elp1 mutants.
CCoAOMT Is Expressed in the Pith Cells of elp1 Stems
To further confirm the correlation between the increase in enzyme activity and the ectopic deposition of lignin in the pith of elp1 mutants, we examined the distribution of CCoAOMT in the elp1 stems by immunolocalization. In the sections of elp1 stems, the distribution of CCoAOMT was not only evident in xylem cells and fibers, but also very intense in some pith cells, which was directly correlated with the localization of lignin shown as red autofluorescence (Fig. 5, A and B). In contrast, a section from a wild-type stem showed the CCoAOMT signal only in xylem cells and fibers (Fig. 5C), the only cell types showing lignin autofluorescence. A control section from an elp1-2 stem incubated with preimmune serum did not show any CCoAOMT signal (Fig. 5D). These results indicate that the expression of the lignin pathway enzyme CCoAOMT in the pith of elp1 stems directly correlates with the ectopic deposition of lignin.
Figure 5.

Immunolocalization of CCoAOMT in elp1 stems. Stem sections were first probed with polyclonal antibodies against zinnia CCoAOMT, and then incubated with gold-labeled secondary antibody. After silver enhancement, the signal was visualized under UV epifluorescent illumination. The CCoAOMT signal is seen as yellow under UV epifluorescence. Lignin shows red autofluorescence under UV light. A and B, Stem sections from the elp1-1 (A) and elp1-2 (B) mutants, respectively, probed with the CCoAOMT antibodies. Note that the CCoAOMT signal was evident in lignified pith cells. C, Stem section from the wild type probed with the CCoAOMT antibodies. Note that the CCoAOMT signal was seen only in xylem cells and fibers but not in pith cells. D, Stem section from the elp1-2 mutant incubated with preimmune serum. No signal was observed in any cells. f, Interfascicular fiber; pi, pith; x, xylem. Magnification, ×625.
Walls of the Lignified Pith Cells in the elp1 Mutants Are Not Thickened
Because lignin deposition was normally associated with sclerenchyma cells in the wild type, we investigated whether lignification in the pith cells of elp1 stems was also accompanied with secondary wall thickening. The wall thickness of pith cells in both the wild-type and the elp1 mutants was examined under electron microscope. After scanning through the walls of all pith cells in sections from different stems of the elp1 mutants, no secondary wall thickening was observed in any pith cells of elp1 stems. Representatives of the walls of pith cells from both the wild-type and the elp1-2 mutant are shown in Figure 6. The wall thickness of pith cells is similar to that of parenchyma cells reported in Arabidopsis (Freshour et al., 1996). This indicates that lignification in the pith of elp1 mutants is not accompanied by secondary wall thickening.
Figure 6.

Electron microscopy of the walls of pith cells. A, Primary walls of the pith cells from a wild-type stem. B, Primary walls of the pith cells from an elp1-2 stem. No secondary wall thickening was observed in the pith cells of the elp1-2 mutant. pw, Primary wall. Magnification, ×12,000.
Genetic Analysis of the elp1 Mutants
To perform genetic analysis, both the elp1-1 and the elp1-2 mutants were backcrossed with wild-type Arabidopsis ecotype Columbia. The resulting F1 plants did not show any lignin staining in the pith cells of the stems, indicating that the elp1 mutations were recessive. The F1 plants were selfed, and the resulting F2 generation analyzed for segregation of the elp1 mutations. Of 323 F2 plants from the cross between the elp1-1 mutant and the wild type, 75 plants showed lignin staining in pith cells. Similarly, of 261 F2 plants from the cross between the elp1-2 mutant and the wild type, 58 plants showed lignin staining in pith cells. Thus, a segregation ratio of 3.3:1 (χ2 = 0.545, P > 10%) and 3.5:1 (χ2 = 1.0, P > 10%) was observed for the elp1-1 and elp1-2 mutations, respectively. This indicates that both the elp1-1 and elp1-2 mutations are most likely due to a single gene mutation. The elp1 mutants did not show dramatic morphological changes compared with the wild-type plants, except they grew slower and had shorter inflorescence stems (data not shown).
To determine the genetic location of the elp1 mutation, the elp1-2 mutant was crossed with wild-type Arabidopsis ecotype Landsberg erecta. The resulting F1 plants were selfed and the F2 plants examined for segregation of the elp1-2 mutation, which showed ectopic staining of lignin in the pith. F2 plants with the elp1 mutant phenotype were used as mapping plants to analyze for linkage with CAPS markers located on each of the five chromosomes (Konieczny and Ausubel, 1993). No linkage was observed with markers on chromosomes 2 to 5. However, a close linkage was found with markers PVV4 and NCC1 on chromosome 1. Of 464 F2 mapping plants analyzed, 73 plants showed crossovers between PVV4 and the elp1 locus. This placed the elp1 locus in a region 7.9 centimorgans (cM) away from PVV4 (Fig. 7). Further mapping of the elp1 locus with NCC1 indicated that the elp1 locus was located between PVV4 and NCC1. The mapping data with NCC1 indicated that the elp1 locus was 7 cM away from NCC1 (Fig. 7).
Figure 7.

Genetic mapping of the elp1 locus. A total of 464 F2 mapping plants were used for mapping with markers on the left side of the elp1 locus, and a total of 449 F2 mapping plants were used with markers on the right side of the elp1 locus. All markers used for mapping were CAPS markers. The markers shown on chromosome 1 were not positioned on scale.
Using the DNA sequence of Arabidopsis yeast artificial chromosome clone yUP8H12, we have developed two new CAPS markers, JGB8 and 8H12-1. Mapping with these markers showed that the elp1 locus was on the right side of JGB8 and 8H12-1 with a genetic distance of 1.3 and 0.65 cM, respectively (Fig. 7). Further mapping with the PAI1 marker indicated that the elp1 locus was on the left side of PAI1, with a genetic distance of 2.4 cM. The region between the markers 8H12-1 and PAI1 is covered by two Arabidopsis yeast artificial chromosome clones, and this region has been targeted for sequencing by the Arabidopsis genome sequencing group. Therefore, we will be able to develop more CAPS markers for fine-mapping and cloning of the ELP1 gene in the near future.
DISCUSSION
A Recessive Single Gene Mutation Results in Lignin Deposition in Pith
Lignin deposition is generally associated with sclerified cells during normal plant growth and development. It may also occur in parenchyma cells in response to wounding or infection. Because all genes in the phenylpropanoid pathway and all those involved in the polymerization of monolignols need to be turned on to synthesize lignin, it is obvious that genes involved in lignin biosynthesis have to be coordinately regulated in response to developmental or environmental cues. It has been suggested that the developmental or environmental cues might turn on common regulatory proteins, which in turn regulate the coordinate expression of genes in the lignin biosynthetic pathway. The coordinate regulation of certain genes in the phenylpropanoid pathway has recently been demonstrated in transgenic studies. It was found that two Myb transcription factors from Antirrhinum could selectively regulate expression of some genes in the phenylpropanoid pathway (Tamagnone et al., 1998). Therefore, Myb-related proteins are likely to be one group of regulatory proteins involved in coordinating the expression of genes in the phenylpropanoid pathway. Our isolation of the elp1 mutants further supports the hypothesis that genes in the lignin biosynthetic pathway are coordinately regulated, because a recessive single gene mutation could result in lignin biosynthesis in the pith cells that are normally not lignified.
The elp1 mutation might have occurred in a gene involved in the control of the coordinate expression of lignin pathway genes. Because pith cell walls in Arabidopsis stems are not lignified during normal plant growth and development, it seems likely that the ELP1 gene might act as a repressor which normally represses the expression in the pith cells of lignin pathway genes. Mutation of the ELP1 gene might turn on genes in the lignin biosynthetic pathway, which leads to ectopic deposition of lignin in the pith. Once the ELP1 gene is cloned, it will be interesting to find the target genes regulated by ELP1.
Ectopic Deposition of Lignin in the elp1 Mutants Is Pith Specific and Independent of Sclerification
Similar to the tissue-specific deposition of lignin in xylem and fibers in the stems of wild-type plants, the ectopic deposition of lignin in the stems of the elp1 mutants appears to be pith specific. No ectopic deposition of lignin was observed in the leaves, roots, or cortex of the stems in the elp1 mutants. This indicates that the ELP1 gene is only involved in the repression of lignin deposition in the pith of stems. It will be interesting to investigate whether a similar mechanism might be involved in the repression of lignin deposition in other non-lignified tissues.
The ectopic deposition of lignin in the pith cells of elp1 stems is not age-dependent, because lignin is observed not only in the pith of old stems, but also in the pith of young, elongating stems (Fig. 3). The ectopic deposition of lignin also appears to be independent of secondary wall thickening. This is very different from the lignin deposition in the sclerified parenchyma cells observed in some other plants. For example, some pith parenchyma cells in stems of alfalfa and forsythia are sclerified, and sclerification is accompanied by a deposition of lignin in walls of these cells (Ye, 1997). In these plants, the sclerification of pith cells only occurs in the non-elongating mature stems, and generally only cells right next to the vascular tissues are sclerified. Thus, sclerification and lignification of pith cells in stems of alfalfa and forsythia are developmentally regulated processes different from the mechanism controlling the ectopic deposition of lignin in elp1 pith cells.
The ectopic deposition of lignin in the pith of elp1 stems appears to be similar to the lignification process in response to wounding or infection. Neither the wounding-induced lignification nor the elp1-induced lignification accompany secondary wall thickening. It is possible that ELP1 is involved in repression of the wounding- or infection-induced lignification pathway under normal plant growth. Mutation of the ELP1 gene may thus lead to constitutive operation of the lignification pathway in the pith. Once the ELP1 gene is cloned, it will be interesting to determine whether ELP1 is involved in wounding- or infection-induced lignification.
Lignin Deposited in the Pith of the elp1 Stems Is Rich in Syringyl Units
It has been shown that the lignin composition differs between xylem and interfascicular fibers in the inflorescence stems of Arabidopsis (Chapple et al., 1992; Meyer et al., 1996). The xylem is rich in guaiacyl lignin but lacks syringyl lignin, whereas the interfascicular fibers are rich in both guaiacyl lignin and syringyl lignin. This difference is most likely caused by the preferential expression of the ferulate 5-hydroxylase gene in the interfascicular fibers but not in the xylem. Therefore, it is obvious that different regulatory proteins are involved in the coordinate expression of different sets of genes in the lignin biosynthetic pathway to control lignin heterogeneity in different tissues. It is interesting that the lignin in the pith of the elp1 mutants is also rich in syringyl lignin, indicating that the lignin composition in the pith of the elp1 mutants is similar to that in the interfascicular fibers. This suggests that the elp1 mutation might lead to the coordinate expression of genes involved in the synthesis of syringyl-rich lignin.
Lignin Deposition Pattern in the Pith of the elp1 Stems Is Mosaic
The mosaic pattern of lignin deposition in the pith of the elp1 mutants is intriguing. It is obvious that all pith cells have the potential to become lignified, as revealed in some sections of the elp1 stems (Fig. 3). However, in most cases, not all cells in the pith of the elp1 mutants were lignified (Fig. 3). The mosaic mutant phenotype raises the possibility that it is caused by an epigenetic control mechanism, as are the mosaic phenotypes of some Arabidopsis mutants. For example, Bender and Fink (1995) found that the mosaic blue fluorescence in the leaves of an Arabidopsis mutant was due to epigenetic control of the phosphoribosylanthranilate isomerase gene family. Once the ELP1 gene is cloned, we might be able to figure out the mechanisms causing this mosaic lignin deposition phenotype.
To make lignin, both the genes in the phenylpropanoid pathway and the genes involved in the polymerization of monolignols have to be coordinately expressed. Because in the elp1 mutants lignin is ectopically deposited in the pith cells, which are normally not lignified, it is reasonable to propose that the mutant gene is involved in the control of genes in the phenylpropanoid pathway, and probably also of genes involved in the polymerization of monolignols. Further characterization of the elp1 mutants will help further our understanding of the mechanisms controlling lignification. Our successful isolation of the elp1 mutants demonstrates the feasibility of studying lignification by genetic analysis. Further screening for other potential lignin mutants would conceivably lead to a better understanding of how the temporal and spatial control of lignification is regulated.
MATERIALS AND METHODS
Mutant Screening
M2 plants of ethyl methanesulfonate-mutagenized populations of Arabidopsis ecotype Columbia (Lehle Seeds, Round Rock, TX) were grown in the greenhouse. Inflorescence stems of 6-week-old plants were free-hand sectioned with a razor blade. Sections were stained with phloroglucinol-HCl (1% [w/v] phloroglucinol in 6 n HCl) and observed under a dissection microscope. Plants with abnormal lignin staining patterns were saved and grown to maturity for seed production. Putative mutants were backcrossed with wild-type Columbia three times to reduce unlinked background mutations.
Syringyl lignin was stained with the Mäule reagent as described by Chapple et al. (1992). Sections were incubated in 0.5% (w/v) KMnO4 for 10 min and then in 4 n HCl for 5 min. Stained sections were mounted in the presence of concentrated NH4OH and observed under a dissection microscope.
Assay of Klason Lignin
Lignin content was quantitatively measured using the Klason method (Kirk and Obst, 1988). Stems were ground into fine powder. After being extracted four times in methanol and vacuum-dried, 200 mg of the sample was hydrolyzed in 4 mL of 72% (v/v) H2SO4 at 30°C for 1 h. The hydrolysate was diluted with addition of 112 mL of water, and then autoclaved for 1 h. The sample solution was filtered through a fritted glass crucible. After being washed and dried, lignin was measured and expressed as a percentage of the original weight of the cell wall residues.
Assay of Enzyme Activity
Inflorescence stems of 6-week-old plants were homogenized in the extraction buffer (50 mm Tris-HCl, pH 7.5, 0.2 mm MgCl2, 2 mm dithiothreitol [DTT], 10% [v/v] glycerol, 0.2 mm phenylmethylsulfonyl fluoride, 10 mg/mL leupeptin, and 10 mg/mL aprotinin) with a mortar and pestle. After homogenization, the extracts were centrifuged at 12,000g for 15 min. The supernatants were saved for the assay of PAL (Lin and Northcote, 1990), caffeoyl coenzyma A O-methyltransferase (CCoAOMT) (Zhong et al., 1998), and cinnamoyl-coenzyme A:NADP oxidoreductase (CCR; Goffner et al., 1994) activities. Each data point was the mean of two separate assays. Protein concentration was determined using the Bradford method (Bradford, 1976) with bovine serum albumin (BSA) as the standard protein.
Immunolocalization by Light Microscopy
Inflorescence stem segments were fixed in the fixation solution (4% [v/v] freshly depolymerized paraformaldehyde in 0.1 m Na-phosphate, pH 7.0) for 12 h at 4°C after vacuum infiltration for 10 min. After dehydration through a gradient series of ethanol, the tissue segments were embedded in paraffin. Thin sections (15 μm) were cut from paraffin-embedded tissues with a microtome, and mounted on glass slides coated with poly-Lys. After rehydration, sections were blocked in 5% (w/v) BSA in phosphate-buffered saline (PBS) solution (10 mm Na-phosphate, pH 7.2, 138 mm NaCl, and 3 mm KCl) for 1 h, and then incubated with polyclonal antibodies against zinnia CCoAOMT fusion protein (1:500 dilution in PBS containing 0.1% [w/v] BSA; Ye, 1997) for 4 h. After washing, the sections were incubated with gold-conjugated goat anti-rabbit polyclonal antibodies (AuroProbe LM GAR, Amersham-Pharmacia Biotech, Uppsala; 1:40 dilution in PBS solution containing 0.1% [w/v] BSA) for 1 h. Bound gold particles were revealed by silver enhancement using a kit (IntenSe, Amersham-Pharmacia Biotech). After being counter-stained with 0.5% (w/v) safranine O for 1 min, sections were dehydrated through a gradient series of ethanol and embedded in Paramount medium (Fisher Scientific, Loughborough, Leicestershire, UK). The signals were observed under a compound microscope with epipolarized illumination.
Electron Microscopy
Stem slices were fixed in 4% (w/v) paraformaldehyde and 1% (v/v) glutaraldehyde in 0.1 m phosphate buffer, pH 7.2, for 4 h at room temperature and then postfixed in 2% (w/v) uranyl acetate. After dehydration through a gradient series of acetone, the tissue slices were infiltrated with K4M embedding medium and polymerized overnight under UV light at −20°C. Ultrathin sections (80 nm thick) were prepared and visualized under a transmission electron microscope.
Genetic Analysis
The mutants were backcrossed with wild-type Columbia. Stem sections of the F1 plants were stained with phloroglucinol-HCl to examine lignin deposition patterns. After selfing of the F1 plants, the F2 plants were analyzed for segregation of the mutation through examination of the lignin deposition patterns in stem sections.
For genetic mapping, the mutants were crossed with Arabidopsis ecotype Landsberg erecta. The resulting F1 plants were selfed, and the F2 plants were analyzed for segregation of the elp1 mutation. Leaves from the F2 plants with the homozygous mutant phenotype were collected for isolation of genomic DNA, as described by Cocciolone and Cone (1993). Linkage of the mutation with markers on individual chromosomes was determined using co-dominant CAPS markers developed by Konieczny and Ausubel (1993). Conditions for PCR reactions and restriction enzyme digestions were essentially the same as described (Konieczny and Ausubel, 1993). The information on CAPS markers PAI1 (Li et al., 1995), PVV4, and NCC1 (Konieczny and Ausubel, 1993) was from the Arabidopsis database. The CAPS markers JGB8 and 8H12-1 were developed during our mapping work.
ACKNOWLEDGMENTS
We thank D.L. Droste and J.J. Taylor for participation of mutant screening, and two anonymous reviewers for their suggestions.
LITERATURE CITED
- Akin DE, Hanna WW, Snook ME, Himmelsbach DS, Barton FE, Windham WR. Normal-12 and brown midrib-12 sorghum: II. Chemical variations and digestibility. Agron J. 1986;78:832–837. [Google Scholar]
- Bell-Lelong DA, Cusumano JC, Meyer K, Chapple C. Cinnamate-4-hydroxylase expression in Arabidopsis: regulation in response to development and the environment. Plant Physiol. 1997;113:729–738. doi: 10.1104/pp.113.3.729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bender J, Fink GR. Epigenetic control of an endogenous gene family is revealed by a novel blue fluorescent mutant of Arabidopsis. Cell. 1995;83:725–734. doi: 10.1016/0092-8674(95)90185-x. [DOI] [PubMed] [Google Scholar]
- Bevan M, Shufflebottom D, Edwards K, Jefferson R, Schuch W. Tissue- and cell-specific activity of a phenylalanine ammonia-lyase promoter in transgenic plants. EMBO J. 1989;8:1899–1906. doi: 10.1002/j.1460-2075.1989.tb03592.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradford M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- Campbell MM, Sederoff RR. Variation in lignin content and composition: mechanisms of control and implications for the genetic improvement of plants. Plant Physiol. 1996;110:3–13. doi: 10.1104/pp.110.1.3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Capellades M, Torres MA, Bastisch I, Stiefel V, Vignols F, Bruce WB, Peterson D, Puigdoménech P, Rigau J. The maize caffeic acid O-methyltransferase gene promoter is active in transgenic tobacco and maize plant tissues. Plant Mol Biol. 1996;31:307–322. doi: 10.1007/BF00021792. [DOI] [PubMed] [Google Scholar]
- Chapple CCS, Vogt T, Ellis BE, Somerville CR. An Arabidopsis mutant defective in the general phenylpropanoid pathway. Plant Cell. 1992;4:1413–1424. doi: 10.1105/tpc.4.11.1413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cocciolone SM, Cone KC. Pl-Bh, an anthocyanin regulatory gene of maize that leads to variegated pigmentation. Genetics. 1993;135:575–588. doi: 10.1093/genetics/135.2.575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dharmawardhana DP, Ellis BE, Carlson JE. Characterization of vascular lignification in Arabidopsis thaliana. Can J Bot. 1992;70:2238–2244. [Google Scholar]
- Douglas CJ, Hauffe KD, Ites-Morales M-E, Ellard M, Paszkowski U, Hahlbrock K, Dangl JL. Exonic sequences are required for elicitor and light activation of a plant defense gene, but promoter sequences are sufficient for tissue specific expression. EMBO J. 1991;10:1767–1775. doi: 10.1002/j.1460-2075.1991.tb07701.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feuillet C, Lauvergeat V, Deswarte C, Pilate G, Boudet A, Grima-Pettenati J. Tissue- and cell-specific expression of a cinnamyl alcohol dehydrogenase promoter in transgenic poplar plants. Plant Mol Biol. 1995;27:651–667. doi: 10.1007/BF00020220. [DOI] [PubMed] [Google Scholar]
- Freshour G, Clay RP, Fuller MS, Albersheim P, Darvill AG, Hahn MG. Developmental and tissue-specific structural alterations of the cell-wall polysaccharides of Arabidopsis thaliana roots. Plant Physiol. 1996;110:1413–1429. doi: 10.1104/pp.110.4.1413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goffner D, Campbell MM, Campargue C, Clastre M, Borderies G, Boudet A, Boudet AM. Purification and characterization of cinnamoyl-coenzyme A:NADP oxidoreductase in Eucalyptus gunnii. Plant Physiol. 1994;106:625–632. doi: 10.1104/pp.106.2.625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grand C, Parmentier P, Boudet A, Boudet AM. Comparison of lignins and enzymes involved in lignification in normal and brown midrib (bm3) mutant corn seedlings. Physiol Veg. 1985;23:905–911. [Google Scholar]
- Halpin C, Holt K, Chojecki J, Oliver D, Chabbert B, Monties B, Edwards K, Barakate A, Foxon GA. Brown-midrib maize (bm1): a mutation affecting the cinnamyl alcohol dehydrogenase gene. Plant J. 1998;14:545–553. doi: 10.1046/j.1365-313x.1998.00153.x. [DOI] [PubMed] [Google Scholar]
- Hauffe KD, Paszkowski U, Schulze-Lefert P, Hahlbrock K, Dangl JL, Douglas CJ. A parsley 4CL-1 promoter fragment specifies complex expression patterns in transgenic tobacco. Plant Cell. 1991;3:435–443. doi: 10.1105/tpc.3.5.435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirk TK, Obst JR. Lignin determination. Methods Enzymol. 1988;161:87–101. [Google Scholar]
- Konieczny A, Ausubel FM. A procedure for mapping Arabidopsis mutations using co-dominant ecotype-specific PCR-based markers. Plant J. 1993;4:403–410. doi: 10.1046/j.1365-313x.1993.04020403.x. [DOI] [PubMed] [Google Scholar]
- Levya A, Liang X, Pintor-Toro JA, Dixon RA, Lamb CJ. cis-Element combinations determine phenylalanine ammonia-lyase gene tissue-specific expression patterns. Plant Cell. 1992;4:263–271. doi: 10.1105/tpc.4.3.263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li J, Zhao J, Rose AB, Schmidt R, Last RL. Arabidopsis phosphoribosylanthranilate isomerase: molecular genetic analysis of triplicate tryptophan pathway genes. Plant Cell. 1995;7:447–461. doi: 10.1105/tpc.7.4.447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin Q, Northcote DH. Expression of phenylalanine ammonia-lyase gene during tracheary-element differentiation from cultured mesophyll cells of Zinnia elegans L. Planta. 1990;182:591–598. doi: 10.1007/BF02341036. [DOI] [PubMed] [Google Scholar]
- Meyer K, Cusumano JC, Somerville C, Chapple CCS. Ferulate-5-hydroxylase from Arabidopsis thaliana defines a new family of cytochrome P450-dependent monooxygenase. Proc Natl Acad Sci USA. 1996;93:6869–6874. doi: 10.1073/pnas.93.14.6869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neustaedter DA, Lee SP, Douglas CJ. A novel parsley 4CL1 cis-element is required for developmentally regulated expression and protein-DNA complex formation. Plant J. 1999;18:77–88. doi: 10.1046/j.1365-313x.1999.00430.x. [DOI] [PubMed] [Google Scholar]
- Ohl S, Hedrick SA, Chory J, Lamb CJ. Functional properties of a phenylalanine ammonia-lyase promoter from Arabidopsis. Plant Cell. 1990;2:837–848. doi: 10.1105/tpc.2.9.837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pillonel C, Mulder MM, Boon JJ, Forster B, Binder A. Improvement of cinnamyl-alcohol dehydrogenase in the control of lignin formation in Sorghum bicolor L. Moench Planta. 1991;185:538–544. doi: 10.1007/BF00202964. [DOI] [PubMed] [Google Scholar]
- Sablowski RWM, Baulcombe DC, Bevan M. Expression of a flower-specific Myb protein in leaf cells using a viral vector causes ectopic activation of a target promoter. Proc Natl Acad Sci USA. 1995;92:6901–6905. doi: 10.1073/pnas.92.15.6901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sablowski RWM, Moyano E, Culianez-Macia FA, Schuch W, Martin C, Bevan M. A flower-specific Myb protein activates transcription of phenylpropanoid biosynthetic genes. EMBO J. 1994;13:128–137. doi: 10.1002/j.1460-2075.1994.tb06242.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tamagnone L, Merida A, Parr A, Mackay S, Culianez-Macia FA, Roberts K, Martin C. The AmMYB308 and AmMYB330 transcription factors from Antirrhinum regulate phenylpropanoid and lignin biosynthesis in transgenic tobacco. Plant Cell. 1998;10:135–154. doi: 10.1105/tpc.10.2.135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vignols F, Rigau J, Torres MA, Capellades M, Puigdomenech P. The brown-midrib3 (bm3) mutation in maize occurs in the gene encoding caffeic acid O-methyltransferase. Plant Cell. 1995;7:407–416. doi: 10.1105/tpc.7.4.407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whetten R, Sederoff R. Lignin biosynthesis. Plant Cell. 1995;7:1001–1013. doi: 10.1105/tpc.7.7.1001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ye Z-H. Association of caffeoyl coenzyme A 3-O-methyltransferase expression with lignifying tissues in several dicot plants. Plant Physiol. 1997;115:1341–1350. doi: 10.1104/pp.115.4.1341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhong R, Morrison WH, Negrel J, Ye Z-H. Dual methylation pathways in lignin biosynthesis. Plant Cell. 1998;10:2033–2045. doi: 10.1105/tpc.10.12.2033. [DOI] [PMC free article] [PubMed] [Google Scholar]