

SUMMARY
The human gastrointestinal tract is resident to a vastly diverse microbial consortium that co-exists through strict rules of invasion, dominance, resilience and succession. While some members possess stronger capabilities for survival than others, each one retains a genome characteristic of their bacterial denomination which subsequently determines survival and ultimately the composition of a human gut microbiome. Collective evidence advocates the concept of gut microbiota modulation via dietary compounds, with or without nutraceutical supplementation. However, consistent reports of strong individuality in responsiveness suggest that initial composition of host microbiota mediates the effect of nutrition modulation. There is also a strong potential for the interaction between mind and microbe to influence responsiveness, although mechanistic understanding of these complex exchanges remains in its infancy at best. Synthetic stool for FMT is a next-generation microbiome-therapy shown effective in treating C.difficile [417] which could provide a feasible alternative to current methods for patients with IBD. Nevertheless, studies investigating optimum timing for FMT administration are essential.
Animal and human studies are only starting to highlight the Pandora of interactions that endure between members of gut microbiome, their associated metabolites, dietary compounds, as well as host neurological and immune systems, all of which characteristic to each individual. Advanced research technologies have excelled the scientific evidence in support of CAM and toward generating NG-CAM systems designed for treatment of specific disease states, such as IBD. While the majority of envisioned NG-CAM strategies presently exist in their experimental and discovery phases, many show promise for future clinical application.
Keywords: Inflammatory bowel disease, Crohn’s disease, ulcerative colitis, gut microbiota, complementary and alternative medicine, nutraceuticals, next-generation
INTRODUCTION
Crohn’s disease (CD) and ulcerative colitis (UC), both subtypes of inflammatory bowel disease (IBD), are chronic relapsing-remitting inflammatory disorders of the gastrointestinal tract associated with a deregulation of the T-cell mediated immune responses toward intestinal bacteria [1–3]. Disease pathogenesis is thought to reflect a complex interaction between genetic susceptibility, a defective immune system, host intestinal microbiota and environmental factors [4]. Despite substantial progress in our mechanistic understanding of chronic intestinal inflammation, including the integral role of enteric bacteria in disease pathogenesis, the precise cause of IBD is still unknown, and available treatment modalities for CD are not curative. Therefore, effective control of IBD, especially CD, is a realistic goal, and an ideal therapy is one that can alter the natural history of the disease in preventing complications while featuring a safe side effect profile and acceptable methods of delivery.
Intestinal bacteria are now considered to be a metabolically active ‘organ’ and the immunologic tone within the intestine should be that of tolerance towards the commensal bacteria, a balance maintained by the innate immune systems’ ability to recognize intestinal antigens and appropriately activate or suppress T cell reactivity to these antigens. It is now recognized that primary colonization of the gut begins in utero, via the umbilical cord, introducing members of the genera Enterocuccus, Streptococcus, Staphlococcus and Propinibacterium [5], and possibly also via the placenta, introducing maternally derived microbes [6]. Further colonization, associated with infant delivery (i.e. vaginal vs cesarean) [7], is followed by a rapid escalation in gut microbial diversity during infancy, consisting of bacteria, archaea, viruses and fungi [8, 9]. 16S rRNA and whole genome sequencing have revealed microbial succession during these early years is nonrandom, potentially implying that early colonization patterns set the stage for bacterial community structures later in life [9]. Age-related factors associated with microbiota composition are of particular interest in IBD, considering the variability in age of onset [10], the phenotypic differences noted between early and late onset [10, 11], and the reality that efficacy in terms of optimum timing for fecal microbiota transplantation (FMT) is not understood.
Despite the fact that over sixty bacterial phyla exist in the world, the gut microbiome of a healthy human primarily consists of bacterial members belonging to two phyla, Firmicutes (~65%) and Bacteroidetes (~25%), thus implying strong underlying constraints in patterns of microbial colonization and succession [8, 12]. The remaining bacterial species are typically distributed among the phyla Actinobacteria (e.g., Bifidobacterium spp.), Proteobacteria (e.g., Escherichia coli) and Verrucomicrobacteria (e.g., Akkermansia muciniphilia), with a possible smaller presence of Fusobacteria and Cyanobacteria [13]. Non-bacterial species, such as archaea, fungi (i.e. mycobiota) and viruses (i.e. virome) also inhabit the human intestinal tract [14], with the human virome largely consisting of bacteriophages, which are numerous, and far more diverse than their bacterial counterparts [8]. Certain viral [15] and fungal species (e.g., Candida albicans) [16–19] identified within the human virome and mycobiota are shown to directly interact with the intestinal immune system and have been portrayed for their role in IBD pathogenesis [20]. Other species, such as the yeast Saccharomyces boulardii, appear protective against intestinal inflammation [21–24]. Notably, intestinal organisms survive in a community governed by a robustness of symbiotic and highly competitive relationships between, and within, host commensals and pathogenic populations. Current research to elucidate these mechanisms takes advantage of the significant, yet fairly recent, technological advancements in next-generation sequencing, synthetic biology, and genetic engineering techniques that are available.
Decades of literature have proposed the concept that gastrointestinal colonization, particularly early in life, plays a key role in development of immune systems and host tolerance to antigens, and it is only over recent years studies have revealed specific microorganisms can directly influence T cell development and maintenance, as well as downstream responses [25–29]. T regulatory cells (Tregs) play an essential role in maintaining gastrointestinal homeostasis through suppression of responses to pathogenic bacteria [30] and food antigens [31]. For example, specific bacterial members belonging to Clostridia [32, 33], and potentially Parabacteroides [34], have been identified to induce Tregs, possibly through production of the short chain fatty acid (SCFA) butyrate [35, 36]. There is also data demonstrating the induction of IL-10-producing Tregs, via polysaccharide A, a Bacteroides fragilis generated metabolite [37]. More recently, specific microbial populations Clostridium ramosum, Bacteroides thetaiotaomicron, Peptostreptococcus magnus and Bacteroides fragilis were identified to control expression of a distinct Rory expressing Treg subset population [38]. Animal studies have revealed segmented filamentous bacteria mediate dendritic cell activity (DCs), via production of serum amyloid A (SAA), leading to a downstream Th17 response [32, 39, 40]. At present, clinical application of microbiota modulation strategies that can target specific immune mechanisms remain forthcoming. Additional studies are essential to delineate the mechanisms by which specific microbiota and their metabolites affect immune cell subtypes, as well as the potential for other factors such as diet and medication to influence these intricate interactions.
COMPLEMENTARY AND ALTERNATIVE MEDICINE (CAM)
Complementary and alternative medicine (CAM) refers to health-care strategies developed separately to that of Western or conventional mainstream medicine, with the term ‘complementary’ implying usage of these therapies in addition to conventional medicine [41]. In the past, many IBD professionals have avoided CAM because of a lack in supporting scientific evidence, despite reports indicating that upwards of 50% of IBD patients utilize non-conventional therapies (i.e. CAM) at some point during their disease course [42–45]. Over recent years however, the body of scientific and clinical evidence in support of CAM has increased dramatically, resulting in a high level of acceptance among medical professionals. Moreover, advanced research technologies are working to reshape the face of many CAM strategies (e.g., genetically engineered probiotics) into those of precise, next-generation CAM (NG-CAM) systems, designed for treatment of specific disease states, including IBD. Still, the integration of CAM/NG-CAM into any medical management plan, whether intended as short- or long-term, should be tailored to each individual patient, and performed by an experienced practitioner capable of evaluating all facets of the available evidence with regard to benefits, safety and cost [41].
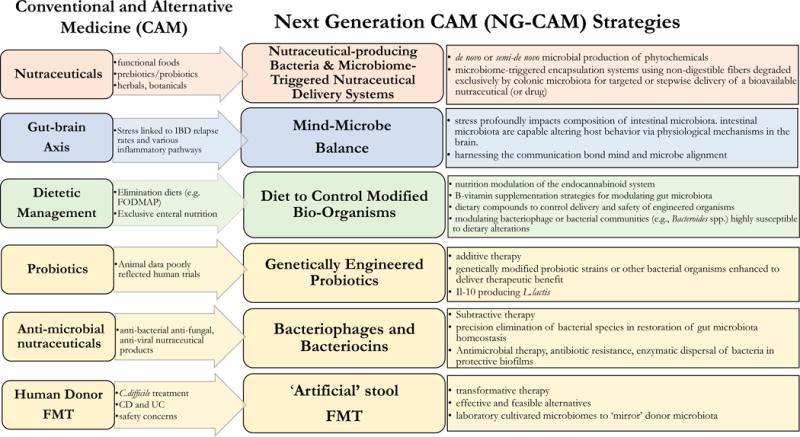
In terms of gut microbiota modulation, CAM/NG-CAM approaches can be classified into one of four major groups; 1) nutraceuticals, 2) mind-microbe balance, 3) dietetic management, and 4) microbiome-therapy. Nutraceuticals refers to an unsuccessfully regulated group of food products ranging from herbals, botanicals, phytochemicals and isolated nutrients, to certain foods and beverages (i.e. ‘functional foods’) stocking specific health benefits based on their ingredients. Recent efforts to overcome the issue of nutraceutical bioavailability have led to advanced microbiome-triggered delivery systems and development of nutraceutical-producing bio-organisms. A growing body of evidence suggests that stress profoundly impacts gut microbiota composition and that the converse is also true, intestinal microorganisms are capable of altering host behavior. Conventionally known as the ‘gut-brain axis’, the mind-microbe balance infers that, to achieve optimal host response from any therapeutic agent, necessitates full consideration of the intimate communication bond between both mind and microbe. Dietetic management encompasses a wide spectrum of dietary regimes characterized by their macronutrient profiles, the nutrient composition or origin of specific foods (e.g. fiber type, gluten-free, saturated vs unsaturated fatty acids), or a combination of these. For example, elimination diets, individual fatty acids, high-fat (HF) diets differing in fat source (animal vs plant), and a range of dietary plant fibers have all been shown to impact gut microbial communities. Microbiome-therapy is primarily gathered into three major paradigms: additive, transformative and subtractive. Additive therapy involves supplementing gut microbiota with individual, or combinations of bacterial strains (i.e. probiotics), whereas transformative therapy entails modulation of the gut microbial community as a whole through fecal microbiota transplantation (FMT). These strategies are now evolving toward genetically engineered probiotic strains designed for specific mechanisms of disease, and the use of ‘artificial’ stool products in FMT. Subtractive therapy refers to selective elimination of pathogenic members of the gut microbiome, with recent studies exploring bacteriophages, a viral species able to kill a precise gamut of bacteria with exquisite specificity, as a potential therapeutic modality for shaping microbiota populations in IBD. Notably, specific dietary compounds have been an important tool for managing activation and biosafety of engineered organisms. Since it is impossible to cover the spectrum of CAM therapies held within the aforementioned groups, this review will focus on CAM and recent NG-CAM that interact with the gut microbiota in context to IBD.
One of the challenges drawing particular attention in the current literature is that baseline (i.e. pretreatment) composition of host intestinal microbiota, namely bacterial diversity, abundance and their functionality, may in fact predict individual responsiveness to therapeutic interventions [46–49]. For instance, one study identified microbiota gene richness at baseline as predictive of inflammatory and metabolic response, following a low-calorie diet intervention [50]. In another diet study involving caloric-restriction in obese individuals, pre-intervention abundance of Akkermansia muciniphila was found inversely related to degree of responsiveness for fasting glucose, body composition and subcutaneous adipocyte diameter [51]. However, further studies are required before individual bacterial species (or microbiome profiles) can be applied as predictive tools for treatment success.
NUTRACEUTICALS
Nutraceuticals are foods, or food products, that provide health and medical benefits in the prevention or treatment of disease [52]. Chemically, nutraceuticals include a range of bioactive substances classified as polyphenolic compounds (e.g., curcumin, resveratrol, ellagitannins, flavenoids), isoprenoids (e.g., terpenoids, carotenoids), minerals (e.g., calcium, zinc), amino acid derivatives (e.g., indoles, choline), carbohydrate derivatives (e.g., oligosaccharides, polysaccharides), fatty acid and structural lipids (e.g., n-3 polyunsaturated fatty acids; PUFAs), and prebiotics [53]. In the United States, there is no specific regulatory framework for nutraceuticals or functional foods [54], and most marketed nutraceuticals do not require clinical trials to support efficacy claims [55, 56]. Although recognized as nutraceuticals, the probiotic arena of today has transcended as a NG-CAM in microbiome-therapy through genetic engineering technology, and thus presented accordingly in this review.
Prebiotics
Prebiotics typically refer to selectively fermented non-digestible compounds that foster growth and activity of health-promoting intestinal microbiota [57]. A review of prebiotic fibers is available elsewhere [58]. Intestinal bacteria rapidly ferment these non-digestible fibers, producing metabolites central to gastrointestinal health such as n-3 fatty acids, metabolic derivatives of tryptophan, and the SCFAs acetate, butyrate and propionate [36, 59]. Ingestion particularly favors growth of beneficial species Lactobacillus and Bifidobacterium, contributing to SCFAs [60, 61]. Gastrointestinal benefits associated with prebiotics have included enhanced mucosal immunity, gut barrier integrity, and epithelial protection from pathogenic bacteria and other metabolites [60, 61]. Bacteria-derived butyrate supports barrier function through two distinct mechanisms, namely, hypoxia-inducible factor activity via increased colonic epithelial cell oxygen consumption [62, 63] and by suppression of histone deacetylases butyrate that can increase tight junction proteins [64].
Dietary components such as fructo-oligosaccharides (FOS), galacto-oligosaccharides (GOS), soya-oligosaccharides, xylo-oligosaccharides, and pyrodextrins have been proposed to increase bacterial diversity [60, 65], as well as selectively enrich Bifidobacterium spp. in the gut [58]. During growth, intake of prebiotic fibers FOS, GOS and soluble corn fiber have been shown to increase absorption of calcium in humans and rats, enhancing bone properties through shifts in gut microbiota [66–69]. There is also evidence that soluble corn fiber increases calcium retention in older individuals who have reached peak bone mass [69], and is capable of altering functional pathways associated with macronutrient and vitamin metabolism [70]. This holds potential clinical utility in IBD considering the high prevalence of suboptimal bone density among patients [71–73]. Both animal and human studies demonstrate increased SCFAs with selective enrichment of Bifidobacterium following ingestion of resistant starches (RS), specifically RS1 and RS2 [74–76]. In humans, RS appears to predominantly elicit colonization by R.bromii, and promote butyrogenic species F.prausnitzii and E.rectale [77, 78]. Another prebiotic dietary fiber, germinated barley foodstuff, provides a glutamine-rich insoluble protein [79] identified to effectively increase luminal butyrate, lactate and acetate production via expansion of Bifidobacterium and Lactobacillus [80]. Clinical studies suggest germinated barley foodstuff may exert therapeutic benefit in UC patients with mild to moderate disease activity, attenuating clinical activity index and mucosal damage, through promotion of Bifidobacterium and butyrate-producing Eubacterium limosum [81–84].
Human prebiotic clinical intervention trials and effects on gut microbiota support the concept that prebiotic feeding increases abundance of certain bacteria and the production of various dietary metabolites. However, strong individuality in responsiveness is observed, underscoring the likelihood that initial composition of host microbiota can mediate effects of nutrition modulation [47]. There is also the possibility that, the structural variations found to exist between a single plant species [85], could prevent Bifidobacterium from degrading certain fiber structures, as bacterium may not possess the appropriate enzymes. Even so, considering the high capacity for microbial horizontal gene transfer in the human gut [86, 87], inter-individual differences would likely persist. Recently, Bindels et al (2015) proposed that prebiotics be redefined as non-digestible compounds that through its metabolization by microorganisms in the gut, modulates composition and activity of the gut microbiota, thus conferring a beneficial physiological effect on the host [65, 88]
Polysaccharides
Polysaccharides refers to a heterogeneous group of structurally diverse carbohydrate molecules, many of which used by the food industry as additives to enhance viscosity and texture of foods. Carrageenans, one of the more widely used food additives, derive from several species of red seaweed (Rhodophyceae) and are unique in cellular structure. Microbial fermentation of carrageenans requires specific gut microbiome-encoded-enzymes, known as carbohydrate active enzymes (‘CAZymes’) [89]. CAZymes have been identified in genomes of marine microbes, but are absent in the human microbiome genome, thus humans are unable to digest carrageenan [86, 87, 90]. Interestingly, metagenomic studies have now uncovered bacterial members of the human gut belonging to the genus Bacteroides (e.g., B. plebeius) that possess CAZyme encoding genes, thus allowing breakdown of specific carbohydrates, and in turn, an additional energy source [87, 91]. This evolutionary adaptation is thought to reflect a natural horizontal gene transfer, in that, bacterial taxa, native to the human gut, have the ability to acquire genes from other microbes typically found outside of the human gut [86, 87]. The extent to which extrinsically acquired genes could influence microbial ecology of the human gut warrants further investigation, especially considering the rising consumption of various traditional seaweeds, such as nori (species of red algae genus Porphyra), traditionally used to prepare sushi throughout many nations outside of Asia.
Squid ink polysaccharide
Mouse models of chemotherapy have reported biological activities of squid ink polysaccharide to include anti-tumour, antioxidant, and anti-coagulation effects, as well as enhancement of both IgA secretion and gut barrier integrity [92–96]. These models suggested that the polysaccharide induced beneficial effects through reversal of chemotherapy-related dysbiosis. One recent animal study reported squid ink polysaccharide decreased abundance of Ruminococcus, Bilophila, Oscillospira, Dorea and, Mucispirillum, bacterium known to thrive during early disruption of the colonic mucosal surface layer [97].
Casein glycomacropeptide
Glycomacropeptide (GMP), a sialic acid rich peptide, releases in whey during cheese making, and from bovine milk, releasing approximately 10-times higher amounts in the casein component of whey (i.e. casein GMP) [98]. Glycomacropeptide is significantly rich in amino acids proline, glutamine, serine, and threonine, as well as the branched chain amino acids (BCAA) isoleucine and valine. There is evidence that certain amino acids act as precursors to SCFA synthesis [99–103]. Specifically, anaerobic bacteria have the ability to metabolize glycine, threonine, glutamate, lysine, ornithine and aspartate, into acetate, which other gut microbiota utilize to generate butyrate [99]. Threonine appears to be the most versatile as it also used to generate propionate [104, 105].
Animal models of induced colitis suggest that casein GMP may attenuate intestinal damage (morphological/histological) via NF-KB/p65 pathway inhibition [106]. Animal models also indicate that Casein GMP could mediate intestinal inflammation via the gut microbiota [107–109], with administration associated with reductions in Proteobacteria, genera Desulfovibrio (from 30-35% to 7%), and increases in acetate, propionate and butyrate levels (cecum) [110]. Recent data proposed that GMP exerts anti-allergenic activity via modulation of gut microbiota [111]. Specifically, the GMP-induced increase in Lactobacillus, Bifidobacterium and Bacteroides, attributed to increased TGG-B production and reductions in mast cells [111]. Clinical trials for active distal colitis in UC patients have reported casein GMP is well-tolerated, exerting similar disease-modifying effects to that of mesalazine [112, 113].
Mushroom Extracts
Mushrooms are recognized for their nutritional value and insoluble fiber content, with evidence supporting their unique range of bioactive metabolite substances [114]. Other compounds include mushroom polysaccharides, all of which with prebiotic properties, such as β-D-glucan polymers, polysaccharopeptides (PSP), polysaccharide proteins (e.g., PSK), chitin, mannans, galactans, and xylans [115]. Isolation of bioactive substances requires the young fruiting body (mycelium with primordia), thus extracted before the mushroom blooms [114], with triterpenes, lipids, and phenols depicted for their immunomodulatory properties [114, 116]. Mushroom bioactive metabolites are shown to stimulate different cells in the immune system, albeit that the ability of certain metabolites promote or suppress immune systems depends on dosage, route and timing of administration, as well as how the mushroom is cultivated (i.e. soil, harvesting, geography) [114]. Nevertheless, various bioactive metabolites are in use clinically [117, 118], and are available commercially worldwide in the form of capsules, food additives, syrups or teas [117].
In a mouse model of diet-induced obesity, administration of a water extract of Ganoderma lucidum mycelium (WEGL) was found to reverse HF-diet induced gut dysbiosis, noted by decreased Firmicutes-to-Bacteroidetes ratio and Proteobacteria levels, with a 10-fold increase in butyrate-producing species Roseburia [119]. The anti-inflammatory and gastroprotective effects identified were recapitulated following FMT (WEGL microbiota inoculum) into WEGL naïve recipients fed a HF-diet [119]. In a recent report, administration of G. lucidum (100mg/kg rat body weight, as oral suspension) improved barrier function via upregulation of occludin expression and increased ileal IgA [120]. Change in immune function correlated with improved microbiota richness, decreased Firmicutes-to-Bacteroidetes ratio and reduced relative abundance of Proteobacteria (cecal content). However, a 2015 Cochrane review of randomized controlled clinical trials (5 trials, N=398) investigating effectiveness of WEGL (as anti-obesity) and G. lucidum for treatment of pharmacologically modifiable risk factors of cardiovascular disease (CVD), did not support use in CVD or type 2 diabetes [121]. In addition, participants who took G. lucidum for four months were 1.67 times (RR=1.67; 95%CI 0.86 to 3.24) more likely to experience an adverse event vs placebo. Adverse effects included nausea, diarrhea or constipation.
Phytochemicals
Phytochemicals are naturally occurring plant chemicals that provide plants with color, odor and flavor. Thousands of structurally heterogeneous phytochemical compounds have been identified thus far, many of which shown to influence chemical processes of the host, including several pathways related to IBD pathogenesis [122]. Supplementation strategies in animal models of IBD seem to ameliorate intestinal inflammation, with or without changes to gut microbiota, however, data has poorly reflected in human clinical trials [123]. Differences in study design, and reports of conflicting or harmful effects have limited generalization of findings. Details of this are outside the scope of the present review and are discussed extensively elsewhere [124–126].
Approximately 90-95% of total dietary polyphenols reach the colon unabsorbed [127]. Microbial modulation has been reported in animal or human studies, or both, by; epigallocatechin gallate (EGCG, main catechin of green tea) [128–131], ellagic acid and ellagitannis (pomegranate, raspberries, blackberries, strawberries and chestnuts) [132, 133], ginseng saponins (ginsenodises) [134, 135], and resveratrol (red wine) [136, 137]. In vitro evidence has shown that naringenin (a flavone) can inhibit growth and adhesion of Gram-negative pathogen Salmonella typhimurium, yet enhance proliferation of the anti-inflammatory probiotic strain L. rhamnosus [138, 139]. Recently, EGCG supplementation was shown to decrease DNA damage in mice fed a HF diet, with reversal of tissue-specific gene expression and methylation patterns of DNA methyltransferase 1 and MutL homologue 1, implying potential for epigenetic consequences [140]. The ratio of Firmicutes to Bacteroidetes in supplemented mice was also significantly lower to that of controls.
The ability for phytochemicals to influence composition and metabolic activity of colonic bacteria definitely exists, but efficacy intimately reflects dosage, timing and route of administration. Bioavailability is also an important issue, and depends on phytochemical origin (whole food vs extract), overall diet composition [141], as well as host colonic microbiome via bacterial metabolism of dietary polyphenols, in turn affecting bioavailability and derived metabolites [122, 142, 143]. While certain colonic bacteria can enhance bioavailability of dietary [124, 143–147], recent data revealed microbial metabolism of certain phytochemicals (e.g., quercetin), could indirectly influence the effects (enhance or suppress) of other dietary compounds, such as n-3 PUFAs [148].
Microbiome triggered delivery
In an effort to overcome bioavailability issues, several groups have focused on colon-targeted delivery systems, utilizing non-digestible fibers (e.g. resistant starch) degraded exclusively by colonic microbiota, for encapsulation and delivery of bioavailable nutraceuticals [149–151]. This method of encapsulation has proven clinically effective for delivery of 1, 25-dihydroxyvitamin D3-25-b-glucuronide (hormonally active vitamin D) in treating localized colonic inflammation, without risk of hypercalcemia or intestinal loss [152, 153]. In another study, mesalamine linked with L-glutamine via azo linkage, a bond specifically cleaved by the colonic microbiome, was reported to reach 84.7% targeted drug and nutraceutical delivery in the colon [154].
Colon-targeted drug delivery systems intended for IBD patients have implemented food grade materials including soy protein, β-lacto globulin (whey), chitosan and zein [126]. To improve encapsulation and pH controlled release of nutraceuticals, smart pH nanoparticles namely carboxymethyl chitosan and calcium pectinate cross-linked with PEI, have also been established [155]. The latter coating method is degraded only by pectinases in the colonic environment, and was reported to protect resveratrol activity until desired release into the lower bowel [156]. Other colon-targeted carrier systems of curcumin have proven successful in vivo [157–159]. Several groups have effectively implemented co-encapsulation, essentially, two nutraceuticals in a single carrier.
Nutraceutical co-encapsulations have comprised; curcumin/resveratrol [160], curcumin/catechin [161], curcumin/piperine [162], gallic acid/curcumin co-administered with ascorbic acid/quercetin [163], curcumin/flax seed oil [164], and prebiotic/probiotic co-delivery [165]. Efforts to develop multilayered particulate drug delivery systems for controlled, sustained drug release, including co-encapsulation of drug and nutraceutical while preventing pharmacological cross-reactivity has shown immense promise as a NG-CAM approach in nutraceutical and drug delivery [126, 166].
Medical Cannabis
Cannabinoids, derived from the cannabis plant C.sativa, include over 60 aromatic hydrocarbons, of which Δ9-tetrahydrocannabinol (THC) is the primary psychoactive component. The endocannabinoid system is ubiquitously expressed throughout the human and rodent body, modulating intestinal peristalsis, gastric acid secretion, hunger (including fat-rich intake) [167–169], gut barrier integrity and intestinal inflammation, with gut microbiota interactions identified [170–175]. The inhibitory effects on gastrointestinal motility may benefit patients with diarrhea and intestinal inflammation, as activation of cannabinoid receptors appears to reduce inflammation-associated hyper-motility [67, 176, 177]. Comprehensive reviews of these mechanisms are available elsewhere [41, 173].
Epithelial cells, immune cells (B cells, NK cells, and mast cells) [178, 179], and the enteric nervous system are all cellular targets of the endocannabinoid system [169, 180–182]. Molecular targets include cannabinoid receptor (CBR) type 1 (CB1R) and CBR type 2 (CB2R), as well as transient receptor potential vanilloid 1 receptors (TRPV1s, best known as the receptor for capsaicin), peroxisome proliferator-activated receptor alpha (PPARαs) and the orphan G-protein coupled receptors (GPCRs), GPR55 and GPR119 [169, 183].
Endogenously derived bioactive lipids that activate CBRs include members of the N-acylethanolamine (NAE) family, namely 2-arachidonoylglycerol (2-AG), a promotor of gut barrier integrity, and N-arachidonoylethanolamine (AEA; also termed anandamide), a promotor of epithelial permeability [184], as well as N-palmitoylethanolamine (PEA), N-oleoylethanolamine (OEA), N-stearoylethanolamine (SEA) and N‐linoleylethanolamine (LEA) [185]. These lipids can act as natural ligands for PPARα (OEA and PEA) [186, 187] TRPV1 (AEA and OEA) [169, 188, 189]. Other bioactive lipids belonging to the acylglycerol family, palmitoyl-glycerol (2-PG) and oleoylglycerol (2-OG), are also considered protective of barrier function [190–193].
Mechanistic studies (both in vitro experiments and animal models) have suggested endocannabinoids, namely CB1R and CB2R mediate gut barrier function, epithelial barrier permeability and inflammatory processes, and thus may offer protective effects in IBD [194–198]. Conversely, intestinal biopsies of UC and CD patients revealed enhanced endocannabinoid or bioactive lipid levels, with or without increased CB receptor expression, [189, 199–206] implying dysregulation of the endocannabinoid system plays a role in IBD pathogenesis. In addition, there is now evidence supporting a link between host intestinal microbiota and endocannabinoids in mediating intestinal integrity [207] .
Lactobacillus acidophilus was the first bacteria specifically shown to modulate the expression of CBRs and μ-opioid receptors in murine intestinal cells [208]. The effects of intestinal bacteria on CBR expression was well illustrated in an elaborate study implementing several models of gut microbiota modulation (HF-diet, prebiotics, probiotics or antibiotics), and mice bearing specific mutations involved in bacterial recognition (TLR, Myd88; myeloid differentiation primary response gene 88). Results indicated that gut microbiota modulation could activate colonic Cnr1 expression, the gene encoding for CB1R, which in turn influences gut permeability and plasma lipopolysaccharide (LPS) levels [207, 209]. Everard et al. (2013) observed improvements in gut barrier function correlated with increased intestinal levels of 2-PG, 2-OG and 2-AG in genetically susceptible obese and type 2 diabetic mice administered Akkermansia muciniphila and fed a HF-diet [207, 209]. More recently, the same group showed that intestinal epithelial Myd88 deletion in mice (IEC Myd99 KO) partially protected against inflammation and gut barrier disruption induced by a HF-diet via mechanisms directly involving gut microbiota, and that in the absence of IEC Myd88, levels of AEA decreased, whereas both 2-AG and 2-OG increased. The authors suggested that activation of GPR119 via 2-OG stimulates intestinal L-cell release of glucagon-like peptide-2, a peptide involved in barrier function [210]. Overall, crosstalk between the gut microbiota, endocannabinoid system and metabolism of the host appears capable of inducing specific changes in the intestinal innate immune system, and vise versa.
In a 2011 survey conducted in Mount Sinai Hospital, Toronto Canada evaluating cannabis usage among IBD patients (N=291), 50% of CD and 33% of UC patients reported having used cannabis as a CAM method to relieve IBD-related symptoms such as pain, appetite loss and diarrhea, at some point during their disease course [211]. One retrospective study (N=30, CD) [212] and two prospective studies (N=13; IBD, N=21; CD) [213, 214] have reported beneficial effects of cannabis usage for IBD symptom control, although study limitations included lack of standardization, dosing inconsistency and diversity in cannabis plant strain. In addition, side effects ranged from addiction, dry mouth, drowsiness, increased appetite, to a sensation of “high”. To this end, authorities now forbid motor vehicle use of for at least 3-4 hours after smoking, and 6 hours post oral ingestion [215]. Studies evaluating pharmacological modulation of the endocannabinoid system are currently underway (experimental and clinical trials), but few of these account for the potential interaction between the gut microbiome or endogenous bioactive lipids [216].
Nutrition-based modulation of the endocannabinoid system tone by reducing ratio of n-6 to n-3 has proven effective in some obesity studies [217–219]. For example, docosahexaenoic acid (DHA, C22:0) administration was associated with decreased levels of AEA and 2-AG, and reductions in inflammation and fat mass in mice [220]. In obese individuals, daily supplementation with n-3 PUFAs from krill oil reduced plasma levels of 2-AG [217]. The implication of these findings in IBD is unclear considering the gastro-protective properties of 2-AG. Nutrition modulation of the endocannabinoid system, particularly in IBD, remains in the exploratory and discovery phase.
MIND-MICROBE BALANCE
Numerous studies advocate the neural, hormonal and immune-related communication pathways that exist between the gut and brain of the host [221–224]. In IBD patients, various inflammatory mechanisms affecting epithelial integrity and pro-inflammatory cytokine production [225] have been proposed in explanation of the long-reported correlation between stress and increased relapse of disease [226–230]. However, it is possible that some of these proposed mechanisms are in fact consequences of altered gut bacteria, as stress has now been discovered to profoundly influence gut microbial communities [231, 232]. Yet the converse is also true, with animal models showing that changes to intestinal microorganisms can indeed lead to alterations in behavior. (For review, refer to Abautret-Daly 2017).
Additional animal studies are required and should provide and invaluable first line approach to understanding the physiological changes that occur in the brain in response to intestinal microorganisms, in context to the internal and external milieu (host-specific) that constantly shapes both microbiome and brain. To this end, maintaining the host mind-microbe balance, via an ongoing consideration of this multifaceted communication pathway, could prove quintessential in attaining maximum efficacy of any treatment modality.
DIETETIC MANAGEMENT
Diet is recognized as one of the primary driving forces in shaping the composition of gut bacteria and metabolite production [233–235]. Notably, intestinal microbiota also play an important role in the synthesis of several vitamins (B-vitamins, vitamin K, vitamin A) [236, 237], and can affect the absorption of essential minerals such as iron, (Kau A 2011) magnesium and calcium in the host [238]. Literature supports dietary intervention as a means of promoting beneficial host-bacteria interactions [59, 216, 239], with new evidence revealing how single food ingredients (e.g., turmeric) can interact with functional traits of intestinal microbiota to regulate host physiology [240]. However, the potential interactions between dietary compounds and host-specific microbial communities have not been investigated in most human diet intervention studies performed to date. A recent review of the known interactions between mucosal immunity, host genetics, gut microbiome and diet is available [241].
Macronutrients
Even small dietary changes have the potential to alter the composition of gut microbiota, in as little as a day [242, 243]. Short-term dietary interventions, especially those devoid of carbohydrates appear to have the most profound effect [233], but taxonomic changes are not consistent among individuals [235], perhaps a reflection of baseline microbiota composition. Fungal abundance has been linked to a carbohydrate-rich diet, with candida abundance positively correlated with carbohydrate intake, and negatively correlated with total saturated fatty acids [244]. Long-term dietary changes tend to impact ratios of Bacteriodes, Prevotella, and Firmicutes, with the most profound change in microbiota composition and bacterial metabolites resulting from diets high in red meat and fat, namely PUFAs [235, 245, 246]. Studies have suggested dietary fiber promotes Prevotella, whereas higher protein and fat intakes promote Bacteroides dominance [235, 245, 246].
In IL-10−/− mice, significant blooms in the ‘pathobiont’ Bilophila wadsworthia correlated with feeding a HF-diet rich in saturated milk fat [25]. The sulfite-reducing properties of B.wadsworthia led to an abundance of hydrogen sulfide production, a molecule disruptive to epithelial barrier function [247, 248]. Other animal models have shown that a HF-diet, in combination with high sugar, increases adherent invasive E.coli (AIEC) [249], while others show that certain fibers (pectin, guar gum) can potentially mediate the pro-inflammatory effects of a dietary lipid [250]. Various IBD mouse models have suggested medium chain triglycerides (MCTs) can exert anti-inflammatory effects, thereby reducing intestinal inflammation [251–253]. In these studies, MCTs consisted of various saturated fatty acids sourced from coconut oil.
A predominantly meat-based diet was reported to decrease Clostridiales, a bacterium involved in plant-based fiber metabolism, while increasing abundance of bile-tolerant Alistipes [233]. In professional athletes, microbial diversity is markedly impacted by disproportionately high dietary intakes of protein, particularly within the genus Akkermansia [254]. Animal-based foods (i.e. red meat) deliver L-carnitine to certain gut bacteria producing trimethylamine N-oxide (TMAO) [255], a molecule identified to promote atherosclerosis in animal models (Lang 1998) and humans [256]. In mice, potato resistant-starch appeared to attenuate detrimental effects of a meat-based diet, partly by promoting members the beneficial, Lactobacillus spp. [257, 258].
Mediterranean diets, which are inherently low in red meat, has been suggested to beneficially impact gut microbiota [259], whereas an energy restricted diet, in combination with dietary fiber, was shown to increase microbial diversity an upwards of 25%, in individuals with low diversity [260]. Of interest, omnivores appear to produce more TMAO from dietary L-carnitine than that of vegans or vegetarians [256]. One plausible explanation could be archaeal lineages underlying the human gut microbiome. Certain strains of methanogens in the human gut, such as Methanomassiliicoccus luminyensis and Methanosarcina barkeri strictly use methyl-based compounds, including TMA, to enable their growth, thus could readily deplete TMA levels [261]. For instance, M.luminyensis encodes a rare proteinogenetic amino acid pyrrolysine (Pyl), a characteristic shared by few other bacteria [262]. This is a truly unique characteristic since methanogenesis of TMA (a methylated amine) is only possible in the presence of pyrrolysine in active catalytic site [263]. Notably, studies exploring the abundance and activity of archaeal taxa between human populations have identified significant differences in organism groups based on geographic location, as well as the consumption of specific food items such as salt-fermented seafood [262, 264–268].
B-Vitamins
The human gut microbiota supplies the host with B-vitamins, including niacin (B3), riboflavin (B2), cobalamin (B12), biotin (B8), folate (B9), thiamin (B1), pantothenate (B5) and pyridoxine (B7). The host relies largely on microbiota-derived B-vitamins, as well as those obtained from dietary sources, as human cells alone do not produce sufficient quantities of B-vitamins [269]. A comprehensive genome assessment of 256 common human gut bacteria revealed a diverse distribution in the presence and absence of B-vitamin biosynthesis pathways, as well as that gut microbes actively exchange B-vitamins among each other, and in doing so, enable organisms which are deficient in any of these vital biosynthesis pathways, to survive [236]. Three distinct genome patterns, were identified, namely; 1) Actinobacteria are limited to niacin, pyridoxine and thiamin pathways, 2) except for niacin, all pathways are lacking in six Firmicutes and two Actinobacteria genomes, and 3) five Firmicutes and three Proteobacteria genomes lack all pathways except for biotin and folate biosynthesis. Inverse patterns were also noted, implying complementary relationships between microbiota. For instance, the essential roles for folate metabolism were missing in all four F.prausnitzii genomes. Findings indicate that perturbations to gut microbiota may impact not only individual B-vitamin requirements, but that deficiency in one or more B-vitamin, can lead to unfavorable blooms in pro-inflammatory organisms[236]. On the other hand, supplementation strategies using single or various combinations of B-vitamins may provide a novel avenue for manipulating the gut microbiota.
Vitamin D
The vitamin D receptor (VDR) is abundantly expressed throughout the intestine and in all immune cells, and is both directly and indirectly targeted by the bioactive forms of vitamin D, 1,25-Dihydroxyvitamin D (1,25[OH]₂D and 1,25[OH]₂D₃) [270–272]. There is evidence to support the interaction between vitamin D, the VDR and gut microbial communities in immune-mediated disorders. For instance, compared to WT mice, VDR null mice have a depletion in Alistipes and Odoribacter with markedly increased levels of Clostridium, Bacteroides, and Eggerthella (cecum) [273], the latter bacterium implicated in UC and CD [274]. In a model of colitis-induced colorectal cancer, increased VDR expression following administration of VSL#3 was associated with lower colon damage and decreased in richness and diversity of mucosally adherent bacteria [275]. Low vitamin D status is frequently observed in IBD, especially in CD patients [276]. One randomized controlled trial reported oral supplementation with Lactobacillus reuteri NCIMB 30242 increased circulating 25(OH)D concentrations [277].
Elimination Diets
Elimination diets consist of an assortment of restrictive dietary regimens entailing the avoidance of specific foods or food groups that may, or may not, precede re-introduction of single food types for identification of those that initiate symptoms. Most elimination diets are not supported by gastroenterology organizations [278–280] although approximately 70% of IBD patients report implementing some form of elimination diet while in remission [281]. Often, these self-imposed diets are based on non-medical resources, such as the internet or IBD support groups, with long-term avoidance of major food groups resulting in severe nutrient deficiencies and malnutrition [125, 282]. Recent data shows malnutrition can alter microbial composition, and intraepithelial lymphocyte phenotype of the small intestine [283].
FODMAP
A low-FODMAP (acronym of “Fermentable Oligo-, Di-, Mono and Polyols”) diet encompasses the dietary restriction of fermentable oligosaccharides (many kinds of vegetables including ‘onion family’, legumes, wheat/rye), disaccharides (lactose-based dairy; milk, yoghurt, cheeses), monosaccharides (many kinds of fruits, dried fruit, fruit juice) and polyols (honey, corn syrup, fructose, sweeteners ending in ‘-ol’) [284]. In principal, the diet may reduce symptoms of bloating and abdominal pain through the avoidance of short-chain carbohydrates (insoluble fibers) which undergo rapid fermentation by colonic bacteria [284, 285]. In patients with existing disease or intestinal narrowing (strictures) avoidance of high-FODMAP foods (i.e. insoluble fiber) thus may be appropriate [281]. However, the majority of excluded FODMAP foods are indeed prebiotics, capable of beneficially modulating microbiota communities [286, 287]. In IBS patients, one randomized controlled trial reported a low-FODMAP diet to significantly reduce Bifidobacterium spp [287], with a second trial reporting greater bacterial diversity of butyrate-producing microbiota clusters in the high-FODMAP group, and reduced total bacterial abundance in the low-FODMAP group [286]. The high-FODMAP diet increased the relative abundance of anti-inflammatory Akkermansia muciniphila and the butyrate-producing Clostridium cluster XIVa [286]. Oligosaccharides (soluble fiber) were reported to block adherence of pathogenic bacteria to epithelia in vitro, including mucosally associated AIEC, a bacterium frequently observed within the mucosa of CD patients [288, 289]. Of the soluble plant fibers, soluble plantain fiber was reported the most consistently effective, inhibiting adherence of Salmonella spp., Shigella spp, Enterotoxigenic E.coli and C.difficile [290, 291]. Many of the oligosaccharide-containing cruciferous vegetables excluded from the FODMAP diet (e.g., brussel sprouts, cabbage, collard greens, kale) are also rich in sulfur containing compounds called glucosinolates, converted by gut microbiota to biologically active, anti-inflammatory compounds such as indoles, nitriles and isothiocyanates [241, 292].
Specific Carbohydrate Diet (SCD)
The specific carbohydrate diet (SCD) involves elimination of most dietary carbohydrates, primarily grains, starches, dairy, and sugars/sweeteners (except honey). Evidence to support effectiveness of SCD to induce or maintain remission in IBD is only available through small case series in CD and UC [293–296]. The proposed mechanisms underlying SCD beneficial effects include; altered macronutrient ratio (protein, fat), reduced dietary gluten intake, or reduced intake of food additives (e.g., emulsifiers), shown to influence gut microbiota and barrier function [249, 297, 298]. Animal models of gluten sensitivity support the modulatory role of gut microbiota in host responses to gluten [299], with non-celiac, gluten-sensitive IBD patients reporting improvement in clinical symptoms following a gluten-free diet [300].
Exclusive elemental nutrition (EEN)
Exclusive enteral nutrition (EEN) is a formula based therapy recommended as first line therapy for induction of remission in pediatric CD, with 85% efficacy rate [301]. Lower efficacy is noted in adult CD patients, possibly because of poor compliance or longer exposure to immunosuppressive treatments [239]. Some pediatric CD studies show EEN-induced alterations to Bacteroides-Prevotella correlate with therapeutic response [302, 303] whereas less impressive changes are observed in samples of adult CD patients post-treatment with enteral nutrition [303]. There is also literature suggesting that intestinal inflammation, relative to CD, may induce some changes in microbiota, such as reduced diversity and increased Proteobacteria [288]. Taken together, additional studies assessing the effects of EEN and enteral nutrition in adult CD patients are required.
MICROBIOME-THERAPY
Microbiome-therapy such as additive (e.g., probiotics), subtractive (e.g., selective antibiotics, nutraceuticals with anti-microbial properties), or transformative (i.e. FMT) are proving effective, although individual responsiveness varies substantially.
Subtractive therapy
Subtractive therapy is the removal of specific bacterial species, or groups of species, from the gut to correct gut disease-associated dysbiosis, thereby restoring host microbial homeostasis. Elimination of host pathogenic gut bacteria though conventional broad-spectrum antibiotics or non-conventional nutraceutical products with anti-microbial activity [304], leads to a concomitant reduction in host commensals, possibly increasing host susceptibility to other infectious agents (e.g., C.difficile). Next-generation subtractive therapies have overturned the concept of therapeutic probiotics, to that of therapeutic bacteriocins, a bacterially derived antimicrobial peptide with toxic activity [305]. Metagenomic and in vivo efforts demonstrate bacteriocin producers have a strong competitive/fitness advantage and increased protection against pathogens [58, 306–309]. Subtractive therapy has also gone viral, via bacteriophage therapy, a subset of viral species with the potential to bind and kill a narrow range of bacteria with impeccable specificity [310–312]. Bacteriophages, or phages, are highly abundant, naturally occurring organic entities. Viral genomes have been identified in both healthy and diseased individuals [313–315], with healthy humans harboring an estimated 1200 viral genotypes [316]. Some IBD studies demonstrate reduced bacterial diversity with concomitant increase in bacteriophage richness, specifically in Caudovirales [314], whereas others have reported decreased overall diversity [317–319]. Intestinal virome populations seem to be unique to each individual [313, 315], and phage community structure is highly susceptible to diet [313], including other factors such as geographical location [314]. Evidence indicates that viromes are also acquired via diet [320], and it has been suggested that ingestion of viral communities occurs at a much higher rate than internal production [321].
Current efforts to enhance, or engineer bacteriophages, for host therapeutic benefit, have focused on; enzymatic dispersal of bacteria in protective biofilms [322], antibiotic resistance [322, 323], antimicrobial activity [324], and modulation of gut microbial communities via cell death [325–330]. Therapeutic deployment of bacteriophage therapy in IBD has utilized naturally occurring, or genetically engineered viral parasites [325]. Of interest, it now proposed that phages could exert immunogenic effects [331], with several studies demonstrating direct effects of intestinal phages on the immune system, inding pro- and anti-inflammatory responses [316, 332–341]. Research exploring bacteriophage therapy remains in its early stages; however, the prospective that natural or engineered phages (i.e. augmented anti-bacterial capability) can intimately control microbial population structure, while mediating mucosal immunity, offers an intriguing prospect in the management of IBD patients.
Additive therapy
Additive therapy entails supplementation of host microbiota with either individual, or a combination of bacterial strains either natural (i.e.probiotics) or genetically engineered in origin, such as probiotic strains with enhanced function.
Probiotics
Probiotics are live microorganisms that exert health benefits to the host when administered in sufficient quantity, with strains isolated from the human intestine suggested as preferential [310]. Animals and humans investigations of various probiotic regimens in IBD have yielded controversial results, and animal data inconsistently reflecting in human IBD trials [305]. Two strains of probiotics Escherichia coli Nissle 1917 and VSL#3 have shown some efficacy in UC and pouchitis [307, 342–346]; however, no recommendations exist for probiotic administration in CD. Clinical intervention studies investigating their effects on the microbiota and human health have been limited by differences in probiotic formulation, timing of administration, and dosage, as well as variability in patient diet and medication use (For reviews see: Vindigni 2016, Uranga 2016, Abautret 2017, Yadav 2016) [125, 226, 347, 348]. To date, no IBD focused research has explored the potential therapeutic benefit of probiotics in alleviating anxiety or depression, although this has been explored in other disorders [226].
Genetically Engineered Probiotics
Recent efforts are focusing on genetic modification of probiotic strains as a NG-CAM strategy. Advantages of enhanced bio-organisms can include stability of colonization (i.e. bioavailability, longevity) and dynamic correction of disease-dysbiosis related perturbations to prevent or resolve inflammation [305]. Application of cellular engineering in augmentation of prophylactic probiotics such as E. coli Nissle 1917 and Lactobacillus jensenii has been successful in overcoming natural colonization resistance conferred by host resident gut microbiota [349]. Today, Lactococcus lactis is one of the most commonly applied bacterial chassis, a nonpathogenic, non-colonizing Gram-positive bacterium used extensively throughout the dairy industry [350]. In mice, L. lactis has been successfully engineered to secrete biologically active murine IL-10, a potent anti-inflammatory cytokine, relevant to IBD [350, 351]. In addition, when synthesized in vivo, lower doses of IL-10 were required compared to systemic IL-10 administration [351]. Phase I clinical trials using IL-10 secreting L.lactis have been well tolerated in CD patients, although efficacy was modest [352]. A nanobody-secreting modified strain of L. lactis, delivering active anti-mTNF nanobodies locally to the colonic mucosa, has proven highly efficacious in DSS-induced colitis and established enterocolitis IL-10 null mice [353]. Described for its anti-inflammatory properties, elafin is associated with restoring barrier function of damaged intestinal epithelia [354–356], with reduced expression and subsequent defective elastolytic activity reported in patients with IBD [357–359]. Two strains of elafin-secreting bacteria, L.lactis and L. casei, were shown to decrease intestinal inflammation in mice, and to protect cultured human epithelial cells from increased epithelial permeability [360]. Other modifications of L. lactis for treatment of diabetes and autoimmune disease [361, 362] have been investigated, namely strains-producing auto-antigen proinsulin [363] and glutamic acid [291]. Administration of NAEs directly to the intestinal lumen to promote endocannabinoid function is also under investigation. Using a genetically modified strain of E. coli Nissle 1917 to produce NAPE-synthesizing enzymes, precursors to the NAE family of lipids, mice had lower food intake, insulin resistance, adiposity, and hepatosteatosis, with increases in hepatic NAE [364]. However, the effects on other bacterial members in the gut were not investigated.
To date, almost every type of known polyphenolic compound have been produced de novo or semi-de novo by genetically engineered strains of E.coli or Saccharomyces cerevisiae [365]. Modified strains have also been used to produce alkaloids (amino acid-derived compounds) [283, 366–370], polysaccharides [371], and various forms of terpenoids (e.g., lutein, lycopene, carotenes and carotenoids), a group of phytochemicals typically present in green foods and soy plants [372–376]. Recently, tryptophan-derived indolyguco-sinolate was produced in a strain of S.cerevisae after genome insertion with eight plant genes [377]. These remarkable progresses have also ensued development of novel or non-natural polyphenolic compounds [378, 379].
One of the drawbacks to genetically engineered therapies is the very real concern of biosafety and environmental contamination [351]. The fact is, when administering a live genetically modified organism, that organism is also released into the environment. Once released, there is a possibility of unintentional colonization of individuals, or food industries (e.g., dairy). Moreover, conditions in the human gastrointestinal tract favor natural horizontal gene transfer between bacterial members [380, 381]. Engineered bacterium should thus inherently include a biological containment system such as auxotrophy or other gene defect that necessitates supplementation with an essential metabolite or intact gene [382]. In addition, recombinant therapies designed on bacterial and probiotic strains currently listed by the U.S. Food and Drug Administration as generally regarded as safe (GRAS) should be subject to strict reevaluation protocols via regulatory frameworks [351].
To address the important issues of environmental safety, a modified strain of Bacteriodes ovatus (human commensal) was developed to deliver human transforming growth factor beta (TGF-β1) but under the strict control dietary xylan (i.e. sole energy source) [353, 383, 384]. In a DSS model of colitis, xylan supplementation induced B. ovatus secretion of human TGF-β1, as well as significantly improved DSS-induced colitis, in some cases superior to that of steroid treatment [384]. Additional studies investigating specific dietary compounds and functional capacity as a means of controlling therapy delivery and biosafety of genetically modified bacteria are highly warranted.
Predicting probiotic-microbiota interactions in the host
In practice, a truly effective recombinant therapy would likely require long-term gastrointestinal colonization, and thus, should be capable of self-activation (or not) in response to specific environmental cues, over time in the host. This requires comprehensive understanding of the parameters governing competitive microbial colonization [385], preferred areas of local microbial colonization (i.e. small vs large bowel) [386], as well as how diet, environmental stimuli and mucosal immunity influence microbial ecosystems, or possible latent effects of stimulated immune pathways on microbiota composition. For instance, Bacteroides spp. appear to be resistant against antimicrobial peptides [387] and their abilities for colonization and resilience are largely determined by the presence of carbohydrates [388–390].
In order to predict the complex interactions between probiotic strains (natural or modified), mucosal immune systems, the gut microbiota (intestinal vs surface attached) [391–393] and host genetics, sophisticated in vitro systems, capable of dynamic, physiologically relevant environmental changes are essential [305, 394]. At present, various in and ex vitro models, testing inter-bacterial interactions have been applied, namely, single [395] and multi-stage chemostat models [394], synthetic 3-D tissue scaffolds with villous features [396], mouse ileal organoids [397] and a human gut-on-a-chip microdevice [218]. Assessment of recombinant therapies in gene circuits allows for detection of disease biomarkers and sometimes drug production, but are shown to result in high cellular burden and in turn, their condensed timeframes (i.e. 24 hours) may not adequately gage evolutionary stability of a recombinant therapy [305]. Stability of engineered therapies is a noted limitation as in vitro evolution experiments have shown bacteriophage functionality can reduce rapidly over time [398, 399]. Moving forward, efforts to enhance both robustness and long-term durability of engineered therapies and their testing models should help in the realization of an environmentally autonomous ‘intelligent’ engineered bio-organism as a NG-CAM strategy.
Transformative therapy
Representing a somewhat more intrusive form of additive-type therapy, FMT focuses on changing intestinal microbiota communities as a whole. Procedures encompass infusion of fecal material sourced from a healthy donor, into the gut of an individual (i.e. the recipient) for the treatment of disease-specific dysbiosis, with the aim of restoring microbial balance. Fecal microbiota inoculum is delivered either to the upper gastrointestinal tract via nasogastric/nasojejunal tube or upper endoscopy, or to the lower intestinal area via enema or colonoscopy [400]. For treatment of recurrent C.difficile infections, FMT boasts 90% clinical success, proving twice as effective as antibiotic therapy alone [401]. Mechanisms as to exactly why FMT is effective are not fully understood, although abundance of butyrate-producing species Lachnospiraceae in donor microbiota was recently proposed [28, 402].
FMT
Cumulative evidence seems to indicate that FMT-induced remission is possible in a subset of both UC and CD patients [403–405], although only two randomized controlled studies, bearing conflicting results, have evaluated FMT efficacy in UC patients, and no randomized controlled trial data is published for CD to date [347, 348, 403, 406]. In a 2014 meta-analysis of nine studies (included 79 UC and 39 CD patients), a remission rate of 36.2% in UC patients (pooled proportion) and 60.5% in CD patients (pooled estimate on subgroup analysis) was reported [406, 407]. These effects however, are not universal, nor are they sustained in either of the two IBD groups.
Failure of FMT in clinical studies may be attributed to various factors including, patient population (severe vs medically refractory), mechanism of action, stool donors, administration route and dosage, or other confounding factors such as smoking, diet and medication use [404, 406–409]. A recent study comparing lyophilized FMT product with fresh or frozen products (using same donors) revealed lyophilized product had slightly lower efficacy (vs fresh), whereas no difference between fresh and frozen fecal product was identified [410]. The hygiene hypothesis, one of the longest standing theoretical frameworks associated with the pathogenesis of IBD [4, 411–414], also happens to underpin the single most unexplored aspects of FMT administration, namely optimum timing for microbiota modulation. It is plausible that, remarkable efficacy may be observed if administered early in life, during certain key windows of plasticity in immune development. Equally, manipulation of microbial communities, perhaps early in diagnosis, could greatly improve long-term outcomes by attenuating disease progression.
One decidedly important issue concerning FMT therapy is the overall safety. The gut microbiome is a dynamic and living organism that constantly evolves over time, and a vast number of human-associated microbial and non-microbial (viruses, fungi) species remain undiscovered [415]. While short-term infectious risks of FMT appear to be definable and quantifiable, the theoretical risk of introducing pathogenic organisms that could later exacerbate disease conditions [407, 416], or the potential for unknown long-term consequences [408], are two caveats that continue to dampen enthusiasm in FMT clinical treatment. In an effort to mitigate these risks, several groups are currently investigating use of an ‘artificial’, synthetic stool mixture, designed to contain clinically active microbes [417, 418]. The stool substitute appears capable of curing antibiotic resistant C.difficile colitis [417] and should be pursued as a feasible alternative. Overall, the concept of FMT holds strong potential within the repertoire of CAM and NG-CAM for management of IBD. In the future, the most effective FMT modalities will like encompass defined microbial communities [419] infused into pre-characterized IBD recipients, pertaining to their gut microbiota community, diet, lifestyle and medication intake.
Figure 1.
Conventional and Alternative Medicine Strategies.
KEY POINTS.
The human gut microbiome exerts a major impact on human health and disease, and therapeutic gut microbiota modulation is now a well-advocated strategy in the management of many diseases, including inflammatory bowel disease (IBD).
Scientific and clinical evidence in support of complementary and alternative medicine (CAM), in targeting intestinal dysbiosis among patients with IBD, or other disorders, has increased dramatically over past years. The more recent scientific investigations have excelled through advancements in research technologies to allow redesign of many CAM strategies into that of precision, next-generation CAM (NG-CAM) systems, intended for the treatment of specific disease states.
Intriguing NG-CAM strategies, showing promise in IBD, include; 1) nutraceutical-producing bio-organisms, 2) genetically enhanced probiotic strains and usage of dietary compounds for controlling activation and biosafety, 3) microbiome-triggered encapsulation of bioavailable nutraceuticals (or drugs) for targeted, stepwise delivery, 4) ‘artificial’ stool for fecal material transplantation (FMT), and 5) bacteriophage delivery for precise elimination of specific microorganisms to reshape microbial populations. While the majority presently exist in their experimental and discovery phases, these envisioned NG-CAM strategies offer an intriguing prospect to the future management of IBD patients.
Delivery of ‘artificial’ stool replacements for FMT could provide an effective, safer alternative to that of human donor stool. Nevertheless, optimum timing of FMT administration in IBD remains unexplored, and future investigations to this end are essential.
As a prerequisite, future studies must consider host initial microbiome, as baseline composition of gut microbiota plays a key role in an individual responsiveness to nutrition modulation.
Animal and human studies continue to uncover the Pandora of interactions that endure between members of gut microbiome, their associated metabolites, dietary compounds, as well as the neurological and immune systems of the host, all of which characteristic to each individual. Exploitation of the communication bond between mind and microbe (i.e. mind-microbe balance) may prove to be a quintessential component for development of a truly effective therapeutic agent in IBD.
Acknowledgments
Financial research support for this publication comes from the National Institute of Diabetes And Digestive And Kidney Diseases of the National Institutes of Health (NIH) under Award number P30DK097948 as part of the Digestive Diseases Research Core Centers program, as well as NIH awards DK091222, DK042191, DK055812, and DK097948 to FC.
Footnotes
CONFLICT OF INTEREST
All authors declare no conflict of interest.
References
- 1.Bamias G, Pizarro TT, Cominelli F. Pathway-based approaches to the treatment of inflammatory bowel disease. Transl Res. 167(1):104–15. doi: 10.1016/j.trsl.2015.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Sartor RB. Microbial influences in inflammatory bowel diseases. Gastroenterology. 2008;134(2):577–94. doi: 10.1053/j.gastro.2007.11.059. [DOI] [PubMed] [Google Scholar]
- 3.Sartor RB, Wu GD. Roles for Intestinal Bacteria, Viruses, and Fungi in Pathogenesis of Inflammatory Bowel Diseases and Therapeutic Approaches. Gastroenterology. 152(2):327–339 e4. doi: 10.1053/j.gastro.2016.10.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Molodecky NA, Kaplan GG. Environmental risk factors for inflammatory bowel disease. Gastroenterol Hepatol (N Y) 6(5):339–46. [PMC free article] [PubMed] [Google Scholar]
- 5.Jimenez E, et al. Isolation of commensal bacteria from umbilical cord blood of healthy neonates born by cesarean section. Curr Microbiol. 2005;51(4):270–4. doi: 10.1007/s00284-005-0020-3. [DOI] [PubMed] [Google Scholar]
- 6.Veldhoen M, Ferreira C. Influence of nutrient-derived metabolites on lymphocyte immunity. Nat Med. 21(7):709–18. doi: 10.1038/nm.3894. [DOI] [PubMed] [Google Scholar]
- 7.Dominguez-Bello MG, et al. Development of the human gastrointestinal microbiota and insights from high-throughput sequencing. Gastroenterology. 140(6):1713–9. doi: 10.1053/j.gastro.2011.02.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Cultrone A, et al. Metagenomics of the human intestinal tract: from who is there to what is done there. Current Opinion in Food Science. 4:64–68. [Google Scholar]
- 9.Koenig JE, et al. Succession of microbial consortia in the developing infant gut microbiome. Proc Natl Acad Sci U S A. 108(Suppl 1):4578–85. doi: 10.1073/pnas.1000081107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Silverberg MS, et al. Toward an integrated clinical, molecular and serological classification of inflammatory bowel disease: report of a Working Party of the 2005 Montreal World Congress of Gastroenterology. Can J Gastroenterol. 2005;19(Suppl A):5A–36A. doi: 10.1155/2005/269076. [DOI] [PubMed] [Google Scholar]
- 11.Kostic AD, Xavier RJ, Gevers D. The microbiome in inflammatory bowel disease: current status and the future ahead. Gastroenterology. 146(6):1489–99. doi: 10.1053/j.gastro.2014.02.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ley RE, et al. Evolution of mammals and their gut microbes. Science. 2008;320(5883):1647–51. doi: 10.1126/science.1155725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Walker AW, et al. Dominant and diet-responsive groups of bacteria within the human colonic microbiota. ISME J. 5(2):220–30. doi: 10.1038/ismej.2010.118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Tang J, et al. Mycobiome: Approaches to analysis of intestinal fungi. J Immunol Methods. 421:112–21. doi: 10.1016/j.jim.2015.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kernbauer E, Ding Y, Cadwell K. An enteric virus can replace the beneficial function of commensal bacteria. Nature. 516(7529):94–8. doi: 10.1038/nature13960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Iliev ID, et al. Interactions between commensal fungi and the C-type lectin receptor Dectin-1 influence colitis. Science. 336(6086):1314–7. doi: 10.1126/science.1221789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Richard ML, et al. Gut fungal microbiota: the Yin and Yang of inflammatory bowel disease. Inflamm Bowel Dis. 21(3):656–65. doi: 10.1097/MIB.0000000000000261. [DOI] [PubMed] [Google Scholar]
- 18.Sokol H, et al. Card9 mediates intestinal epithelial cell restitution, T-helper 17 responses, and control of bacterial infection in mice. Gastroenterology. 145(3):591–601 e3. doi: 10.1053/j.gastro.2013.05.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Sokol H, et al. Fungal microbiota dysbiosis in IBD. Gut. doi: 10.1136/gutjnl-2015-310746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Odds FC. Candida and candidosis: a review and bibliography. Bailliere Tindall 1988 [Google Scholar]
- 21.Chen X, et al. Probiotic yeast inhibits VEGFR signaling and angiogenesis in intestinal inflammation. PLoS One. 8(5):e64227. doi: 10.1371/journal.pone.0064227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Jawhara S, Poulain D. Saccharomyces boulardii decreases inflammation and intestinal colonization by Candida albicans in a mouse model of chemically-induced colitis. Med Mycol. 2007;45(8):691–700. doi: 10.1080/13693780701523013. [DOI] [PubMed] [Google Scholar]
- 23.Jawhara S, et al. Colonization of mice by Candida albicans is promoted by chemically induced colitis and augments inflammatory responses through galectin-3. J Infect Dis. 2008;197(7):972–80. doi: 10.1086/528990. [DOI] [PubMed] [Google Scholar]
- 24.Zwolinska-Wcislo M, et al. Effect of Candida colonization on human ulcerative colitis and the healing of inflammatory changes of the colon in the experimental model of colitis ulcerosa. J Physiol Pharmacol. 2009;60(1):107–18. [PubMed] [Google Scholar]
- 25.Devkota S, et al. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10−/− mice. Nature. 487(7405):104–8. doi: 10.1038/nature11225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gkouskou KK, et al. The gut microbiota in mouse models of inflammatory bowel disease. Front Cell Infect Microbiol. 4:28. doi: 10.3389/fcimb.2014.00028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hill DA, et al. Metagenomic analyses reveal antibiotic-induced temporal and spatial changes in intestinal microbiota with associated alterations in immune cell homeostasis. Mucosal Immunol. 3(2):148–58. doi: 10.1038/mi.2009.132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Natividad JM, et al. Ecobiotherapy Rich in Firmicutes Decreases Susceptibility to Colitis in a Humanized Gnotobiotic Mouse Model. Inflamm Bowel Dis. 21(8):1883–93. doi: 10.1097/MIB.0000000000000422. [DOI] [PubMed] [Google Scholar]
- 29.Thorburn AN, et al. Evidence that asthma is a developmental origin disease influenced by maternal diet and bacterial metabolites. Nat Commun. 6:7320. doi: 10.1038/ncomms8320. [DOI] [PubMed] [Google Scholar]
- 30.Sun M, et al. Regulatory immune cells in regulation of intestinal inflammatory response to microbiota. Mucosal Immunol. 8(5):969–78. doi: 10.1038/mi.2015.49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Pabst O, Mowat AM. Oral tolerance to food protein. Mucosal Immunol. 5(3):232–9. doi: 10.1038/mi.2012.4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Atarashi K, et al. Th17 Cell Induction by Adhesion of Microbes to Intestinal Epithelial Cells. Cell. 163(2):367–80. doi: 10.1016/j.cell.2015.08.058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Atarashi K, et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature. 500(7461):232–6. doi: 10.1038/nature12331. [DOI] [PubMed] [Google Scholar]
- 34.Kverka M, et al. Oral administration of Parabacteroides distasonis antigens attenuates experimental murine colitis through modulation of immunity and microbiota composition. Clin Exp Immunol. 163(2):250–9. doi: 10.1111/j.1365-2249.2010.04286.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Arpaia N, et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature. 504(7480):451–5. doi: 10.1038/nature12726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Furusawa Y, et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature. 504(7480):446–50. doi: 10.1038/nature12721. [DOI] [PubMed] [Google Scholar]
- 37.Round JL, Mazmanian SK. The gut microbiota shapes intestinal immune responses during health and disease. Nature Reviews Immunology. 2009;9(5):313–323. doi: 10.1038/nri2515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sefik E, et al. MUCOSAL IMMUNOLOGY. Individual intestinal symbionts induce a distinct population of RORgamma(+) regulatory T cells. Science. 349(6251):993–7. doi: 10.1126/science.aaa9420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ivanov II, et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell. 2009;139(3):485–98. doi: 10.1016/j.cell.2009.09.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kanther M, et al. Commensal microbiota stimulate systemic neutrophil migration through induction of serum amyloid A. Cell Microbiol. 16(7):1053–67. doi: 10.1111/cmi.12257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Yanai H, Salomon N, Lahat A. Complementary Therapies in Inflammatory Bowel Diseases. Curr Gastroenterol Rep. 18(12):62. doi: 10.1007/s11894-016-0537-6. [DOI] [PubMed] [Google Scholar]
- 42.Koning M, et al. Use and predictors of oral complementary and alternative medicine by patients with inflammatory bowel disease: a population-based, case-control study. Inflamm Bowel Dis. 19(4):767–78. doi: 10.1097/MIB.0b013e31827f27c8. [DOI] [PubMed] [Google Scholar]
- 43.Opheim R, et al. Use of complementary and alternative medicine in patients with inflammatory bowel disease: results of a cross-sectional study in Norway. Scand J Gastroenterol. 47(12):1436–47. doi: 10.3109/00365521.2012.725092. [DOI] [PubMed] [Google Scholar]
- 44.Opheim R, et al. Complementary and alternative medicine in patients with inflammatory bowel disease: the results of a population-based inception cohort study (IBSEN) J Crohns Colitis. 6(3):345–53. doi: 10.1016/j.crohns.2011.09.007. [DOI] [PubMed] [Google Scholar]
- 45.Sirois FM. Health-related self-perceptions over time and provider-based Complementary and Alternative Medicine (CAM) use in people with inflammatory bowel disease or arthritis. Complement Ther Med. 22(4):701–9. doi: 10.1016/j.ctim.2014.07.003. [DOI] [PubMed] [Google Scholar]
- 46.Canfora EE, Jocken JW, Blaak EE. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat Rev Endocrinol. 11(10):577–91. doi: 10.1038/nrendo.2015.128. [DOI] [PubMed] [Google Scholar]
- 47.Dore J, Blottiere H. The influence of diet on the gut microbiota and its consequences for health. Curr Opin Biotechnol. 32:195–9. doi: 10.1016/j.copbio.2015.01.002. [DOI] [PubMed] [Google Scholar]
- 48.Fischbach MA, Sonnenburg JL. Eating for two: how metabolism establishes interspecies interactions in the gut. Cell Host Microbe. 10(4):336–47. doi: 10.1016/j.chom.2011.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Krishnan S, Alden N, Lee K. Pathways and functions of gut microbiota metabolism impacting host physiology. Curr Opin Biotechnol. 36:137–45. doi: 10.1016/j.copbio.2015.08.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Le Chatelier E, et al. Richness of human gut microbiome correlates with metabolic markers. Nature. 500(7464):541–6. doi: 10.1038/nature12506. [DOI] [PubMed] [Google Scholar]
- 51.Dao MC, et al. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: relationship with gut microbiome richness and ecology. Gut. 65(3):426–36. doi: 10.1136/gutjnl-2014-308778. [DOI] [PubMed] [Google Scholar]
- 52.Biesalski HK, et al. 26th Hohenheim Consensus Conference, September 11, 2010 Scientific substantiation of health claims: evidence-based nutrition. Nutrition. 27(10 Suppl):S1–20. doi: 10.1016/j.nut.2011.04.002. [DOI] [PubMed] [Google Scholar]
- 53.Sharma G, et al. Phytochemicals of nutraceutical importance. CAB International; Wallingford: Phytochemicals of nutraceutical importance: do they defend against diseases; pp. 1–19. [Google Scholar]
- 54.Coppens P, Da Silva MF, Pettman S. European regulations on nutraceuticals, dietary supplements and functional foods: a framework based on safety. Toxicology. 2006;221(1):59–74. doi: 10.1016/j.tox.2005.12.022. [DOI] [PubMed] [Google Scholar]
- 55.da Costa JoP. A current look at Nutraceuticals–key concepts and future prospects. Trends in Food Science & Technology [Google Scholar]
- 56.Taylor CL. Regulatory frameworks for functional foods and dietary supplements. Nutr Rev. 2004;62(2):55–9. doi: 10.1111/j.1753-4887.2004.tb00024.x. [DOI] [PubMed] [Google Scholar]
- 57.Scaldaferri F, et al. Gut microbial flora, prebiotics, and probiotics in IBD: their current usage and utility. Biomed Res Int. 2013:435268. doi: 10.1155/2013/435268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Kommineni S, et al. Bacteriocin production augments niche competition by enterococci in the mammalian gastrointestinal tract. Nature. 526(7575):719–22. doi: 10.1038/nature15524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Thorburn AN, Macia L, Mackay CR. Diet, metabolites, and “western-lifestyle” inflammatory diseases. Immunity. 40(6):833–42. doi: 10.1016/j.immuni.2014.05.014. [DOI] [PubMed] [Google Scholar]
- 60.Gibson GR, Roberfroid MB. Dietary modulation of the human colonic microbiota: introducing the concept of prebiotics. J Nutr. 1995;125(6):1401–12. doi: 10.1093/jn/125.6.1401. [DOI] [PubMed] [Google Scholar]
- 61.Macfarlane S, Macfarlane GT, Cummings JH. Review article: prebiotics in the gastrointestinal tract. Aliment Pharmacol Ther. 2006;24(5):701–14. doi: 10.1111/j.1365-2036.2006.03042.x. [DOI] [PubMed] [Google Scholar]
- 62.Cushing K, Alvarado DM, Ciorba MA. Butyrate and Mucosal Inflammation: New Scientific Evidence Supports Clinical Observation. Clin Transl Gastroenterol. 6:e108. doi: 10.1038/ctg.2015.34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Kelly CJ, et al. Crosstalk between Microbiota-Derived Short-Chain Fatty Acids and Intestinal Epithelial HIF Augments Tissue Barrier Function. Cell Host Microbe. 17(5):662–71. doi: 10.1016/j.chom.2015.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Bordin M, et al. Histone deacetylase inhibitors up-regulate the expression of tight junction proteins. Mol Cancer Res. 2004;2(12):692–701. [PubMed] [Google Scholar]
- 65.Bindels LB, Walter J, Ramer-Tait AE. Resistant starches for the management of metabolic diseases. Curr Opin Clin Nutr Metab Care. 18(6):559–65. doi: 10.1097/MCO.0000000000000223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Abrams SA, et al. A combination of prebiotic short-and long-chain inulin-type fructans enhances calcium absorption and bone mineralization in young adolescents. The American journal of clinical nutrition. 2005;82(2):471–476. doi: 10.1093/ajcn.82.2.471. [DOI] [PubMed] [Google Scholar]
- 67.Kimball ES, et al. Agonists of cannabinoid receptor 1 and 2 inhibit experimental colitis induced by oil of mustard and by dextran sulfate sodium. Am J Physiol Gastrointest Liver Physiol. 2006;291(2):G364–71. doi: 10.1152/ajpgi.00407.2005. [DOI] [PubMed] [Google Scholar]
- 68.Weaver CM, et al. Galactooligosaccharides improve mineral absorption and bone properties in growing rats through gut fermentation. Journal of agricultural and food chemistry. 59(12):6501–6510. doi: 10.1021/jf2009777. [DOI] [PubMed] [Google Scholar]
- 69.Whisner CM, et al. Galacto-oligosaccharides increase calcium absorption and gut bifidobacteria in young girls: a double-blind cross-over trial. British Journal of Nutrition. 110(07):1292–1303. doi: 10.1017/S000711451300055X. [DOI] [PubMed] [Google Scholar]
- 70.Jakeman SA, et al. Soluble corn fiber increases bone calcium retention in postmenopausal women in a dose-dependent manner: a randomized crossover trial. The American journal of clinical nutrition. 104(3):837–843. doi: 10.3945/ajcn.116.132761. [DOI] [PubMed] [Google Scholar]
- 71.Guz-Mark A, et al. Pediatric-onset inflammatory bowel disease poses risk for low bone mineral density at early adulthood. Dig Liver Dis. doi: 10.1016/j.dld.2017.01.151. [DOI] [PubMed] [Google Scholar]
- 72.Schüle S, et al. Prediction of low bone mineral density in patients with inflammatory bowel diseases. United European Gastroenterology Journal. 4(5):669–676. doi: 10.1177/2050640616658224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Zhao X, et al. Efficacy and safety of medical therapy for low bone mineral density in patients with Crohn disease: A systematic review with network meta-analysis. Medicine. 96(11) doi: 10.1097/MD.0000000000006378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Conlon MA, et al. Resistant starches protect against colonic DNA damage and alter microbiota and gene expression in rats fed a Western diet. J Nutr. 142(5):832–40. doi: 10.3945/jn.111.147660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Kleessen B, et al. Feeding resistant starch affects fecal and cecal microflora and short-chain fatty acids in rats. J Anim Sci. 1997;75(9):2453–62. doi: 10.2527/1997.7592453x. [DOI] [PubMed] [Google Scholar]
- 76.Wang X, et al. Manipulation of colonic bacteria and volatile fatty acid production by dietary high amylose maize (amylomaize) starch granules. J Appl Microbiol. 2002;93(3):390–7. doi: 10.1046/j.1365-2672.2002.01704.x. [DOI] [PubMed] [Google Scholar]
- 77.Leitch EC, et al. Selective colonization of insoluble substrates by human faecal bacteria. Environ Microbiol. 2007;9(3):667–79. doi: 10.1111/j.1462-2920.2006.01186.x. [DOI] [PubMed] [Google Scholar]
- 78.Ze X, et al. Ruminococcus bromii is a keystone species for the degradation of resistant starch in the human colon. ISME J. 6(8):1535–43. doi: 10.1038/ismej.2012.4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Kanauchi O, Agata K, Fushiki T. Mechanism for the increased defecation and jejunum mucosal protein content in rats by feeding germinated barley foodstuff. Biosci Biotechnol Biochem. 1997;61(3):443–8. doi: 10.1271/bbb.61.443. [DOI] [PubMed] [Google Scholar]
- 80.Kanauchi O, et al. Treatment of ulcerative colitis by feeding with germinated barley foodstuff: first report of a multicenter open control trial. J Gastroenterol. 2002;37(Suppl 14):67–72. doi: 10.1007/BF03326417. [DOI] [PubMed] [Google Scholar]
- 81.Faghfoori Z, et al. Effects of an oral supplementation of germinated barley foodstuff on serum tumour necrosis factor-alpha, interleukin-6 and -8 in patients with ulcerative colitis. Ann Clin Biochem. 48(Pt 3):233–7. doi: 10.1258/acb.2010.010093. [DOI] [PubMed] [Google Scholar]
- 82.Hanai H, et al. Germinated barley foodstuff prolongs remission in patients with ulcerative colitis. Int J Mol Med. 2004;13(5):643–7. [PubMed] [Google Scholar]
- 83.Kanauchi O, et al. Effect of germinated barley foodstuff administration on mineral utilization in rodents. J Gastroenterol. 2000;35(3):188–94. doi: 10.1007/s005350050329. [DOI] [PubMed] [Google Scholar]
- 84.Mitsuyama K, et al. Treatment of ulcerative colitis with germinated barley foodstuff feeding: a pilot study. Aliment Pharmacol Ther. 1998;12(12):1225–30. doi: 10.1046/j.1365-2036.1998.00432.x. [DOI] [PubMed] [Google Scholar]
- 85.Salminen S, Salminen E. Lactulose, lactic acid bacteria, intestinal microecology and mucosal protection. Scand J Gastroenterol Suppl. 1997;222:45–8. doi: 10.1080/00365521.1997.11720717. [DOI] [PubMed] [Google Scholar]
- 86.Hehemann JH, et al. Transfer of carbohydrate-active enzymes from marine bacteria to Japanese gut microbiota. Nature. 464(7290):908–912. doi: 10.1038/nature08937. [DOI] [PubMed] [Google Scholar]
- 87.Hehemann JH, et al. Bacteria of the human gut microbiome catabolize red seaweed glycans with carbohydrate-active enzyme updates from extrinsic microbes. Proceedings of the National Academy of Sciences. 109(48):19786–19791. doi: 10.1073/pnas.1211002109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Bindels LB, et al. Towards a more comprehensive concept for prebiotics. Nat Rev Gastroenterol Hepatol. 12(5):303–10. doi: 10.1038/nrgastro.2015.47. [DOI] [PubMed] [Google Scholar]
- 89.Michel G, et al. Bioconversion of red seaweed galactans: a focus on bacterial agarases and carrageenases. Applied Microbiology and Biotechnology. 2006;71(1):23–33. doi: 10.1007/s00253-006-0377-7. [DOI] [PubMed] [Google Scholar]
- 90.Thomas F, et al. Environmental and gut bacteroidetes: the food connection. Front Microbiol. 2:93. doi: 10.3389/fmicb.2011.00093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Martens EC, et al. Recognition and degradation of plant cell wall polysaccharides by two human gut symbionts. PLoS Biol. 9(12):e1001221. doi: 10.1371/journal.pbio.1001221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.He W, et al. Production of chondroitin in metabolically engineered E. coli. Metab Eng. 27:92–100. doi: 10.1016/j.ymben.2014.11.003. [DOI] [PubMed] [Google Scholar]
- 93.Zuo T, et al. The squid ink polysaccharides protect tight junctions and adherens junctions from chemotherapeutic injury in the small intestinal epithelium of mice. Nutr Cancer. 67(2):364–71. doi: 10.1080/01635581.2015.989369. [DOI] [PubMed] [Google Scholar]
- 94.Zuo T, et al. Dietary squid ink polysaccharide could enhance SIgA secretion in chemotherapeutic mice. Food Funct. 5(12):3189–96. doi: 10.1039/c4fo00569d. [DOI] [PubMed] [Google Scholar]
- 95.Zuo T, et al. Dietary squid ink polysaccharide induces goblet cells to protect small intestine from chemotherapy induced injury. Food Funct. 6(3):981–6. doi: 10.1039/c4fo01191k. [DOI] [PubMed] [Google Scholar]
- 96.Zuo T, et al. The dietary polysaccharide from Ommastrephes bartrami prevents chemotherapeutic mucositis by promoting the gene expression of antimicrobial peptides in Paneth cells. Journal of Functional Foods. 12:530–539. [Google Scholar]
- 97.Lu S, et al. High throughput sequencing analysis reveals amelioration of intestinal dysbiosis by squid ink polysaccharide. Journal of Functional Foods. 20:506–515. [Google Scholar]
- 98.Furlanetti AaM, Prata LF. Free and total GMP (glycomacropeptide) contents of milk during bovine lactation. Food Science and Technology (Campinas) 2003;23:121–125. [Google Scholar]
- 99.Barker HA. Amino acid degradation by anaerobic bacteria. Annu Rev Biochem. 1981;50:23–40. doi: 10.1146/annurev.bi.50.070181.000323. [DOI] [PubMed] [Google Scholar]
- 100.Mortensen PB, et al. The degradation of amino acids, proteins, and blood to short-chain fatty acids in colon is prevented by lactulose. Gastroenterology. 1990;98(2):353–60. doi: 10.1016/0016-5085(90)90825-l. [DOI] [PubMed] [Google Scholar]
- 101.Neis EP, Dejong CH, Rensen SS. The role of microbial amino acid metabolism in host metabolism. Nutrients. 7(4):2930–46. doi: 10.3390/nu7042930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Nordgaard I, Mortensen PB, Langkilde AM. Small intestinal malabsorption and colonic fermentation of resistant starch and resistant peptides to short-chain fatty acids. Nutrition. 1995;11(2):129–37. [PubMed] [Google Scholar]
- 103.Rasmussen HS, Holtug K, Mortensen PB. Degradation of amino acids to short-chain fatty acids in humans. An in vitro study. Scand J Gastroenterol. 1988;23(2):178–82. doi: 10.3109/00365528809103964. [DOI] [PubMed] [Google Scholar]
- 104.Davila AM, et al. Re-print of “Intestinal luminal nitrogen metabolism: role of the gut microbiota and consequences for the host”. Pharmacol Res. 69(1):114–26. doi: 10.1016/j.phrs.2013.01.003. [DOI] [PubMed] [Google Scholar]
- 105.Davila AM, et al. Intestinal luminal nitrogen metabolism: role of the gut microbiota and consequences for the host. Pharmacol Res. 68(1):95–107. doi: 10.1016/j.phrs.2012.11.005. [DOI] [PubMed] [Google Scholar]
- 106.Requena P, et al. Bovine glycomacropeptide ameliorates experimental rat ileitis by mechanisms involving downregulation of interleukin 17. Br J Pharmacol. 2008;154(4):825–32. doi: 10.1038/bjp.2008.138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Brody EP. Biological activities of bovine glycomacropeptide. Br J Nutr. 2000;84(Suppl 1):S39–46. doi: 10.1017/s0007114500002233. [DOI] [PubMed] [Google Scholar]
- 108.Idota T, Kawakami H, Nakajima I. Growth-promoting effects of N-acetylneuraminic acid-containing substances on bifidobacteria. Bioscience, biotechnology, and biochemistry. 1994;58(9):1720–1722. [Google Scholar]
- 109.Yakabe T, Kawakami H, Idota T. Growth stimulation agent for bifidus and lactobacillus. Japanese patent. 1994;7 [Google Scholar]
- 110.Sawin EA, et al. Glycomacropeptide is a prebiotic that reduces Desulfovibrio bacteria, increases cecal short-chain fatty acids, and is anti-inflammatory in mice. Am J Physiol Gastrointest Liver Physiol. 309(7):G590–601. doi: 10.1152/ajpgi.00211.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Jimenez M, et al. Novel mechanisms underlying the therapeutic effect of glycomacropeptide on allergy: change in gut microbiota, upregulation of TGF-Î2, and inhibition of mast cells. International Archives of Allergy and Immunology. 171(3–4):217–226. doi: 10.1159/000453035. [DOI] [PubMed] [Google Scholar]
- 112.Hvas CL, et al. Casein glycomacropeptide for active distal ulcerative colitis: a randomized pilot study. Eur J Clin Invest. 46(6):555–63. doi: 10.1111/eci.12634. [DOI] [PubMed] [Google Scholar]
- 113.Wernlund PG, et al. MON-PP058: Randomised Clinical Trial: Casein Glycomacropeptide for Active Distal Ulcerative Colitis–A Pilot Study. Clinical Nutrition. 34:S149. doi: 10.1111/eci.12634. [DOI] [PubMed] [Google Scholar]
- 114.Lull C, Wichers HJ, Savelkoul HF. Antiinflammatory and immunomodulating properties of fungal metabolites. Mediators Inflamm. 2005;2005(2):63–80. doi: 10.1155/MI.2005.63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Valverde ME, Hernandez-Perez T, Paredes-Lopez O. Edible mushrooms: improving human health and promoting quality life. Int J Microbiol. 2015:376387. doi: 10.1155/2015/376387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Wasser SP. Medicinal mushroom science: Current perspectives, advances, evidences, and challenges. Biomed J. 37(6):345–56. doi: 10.4103/2319-4170.138318. [DOI] [PubMed] [Google Scholar]
- 117.Cui J, Chisti Y. Polysaccharopeptides of Coriolus versicolor: physiological activity, uses, and production. Biotechnol Adv. 2003;21(2):109–22. doi: 10.1016/s0734-9750(03)00002-8. [DOI] [PubMed] [Google Scholar]
- 118.Ooi VE, Liu F. Immunomodulation and anti-cancer activity of polysaccharide-protein complexes. Curr Med Chem. 2000;7(7):715–29. doi: 10.2174/0929867003374705. [DOI] [PubMed] [Google Scholar]
- 119.Chang CJ, et al. Ganoderma lucidum reduces obesity in mice by modulating the composition of the gut microbiota. Nat Commun. 6:7489. doi: 10.1038/ncomms8489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Jin M, et al. Effects of polysaccharide from mycelia of Ganoderma lucidum on intestinal barrier functions of rats. International Journal of Biological Macromolecules. 94:1–9. doi: 10.1016/j.ijbiomac.2016.09.099. [DOI] [PubMed] [Google Scholar]
- 121.Klupp NL, et al. Ganoderma lucidum mushroom for the treatment of cardiovascular risk factors. Cochrane Database Syst Rev. (2):CD007259. doi: 10.1002/14651858.CD007259.pub2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Williamson G, Clifford MN. Role of the small intestine, colon and microbiota in determining the metabolic fate of polyphenols. Biochem Pharmacol. doi: 10.1016/j.bcp.2017.03.012. [DOI] [PubMed] [Google Scholar]
- 123.Tomás-Barberán FA, Selma MaV, Espin JC. Interactions of gut microbiota with dietary polyphenols and consequences to human health. Current Opinion in Clinical Nutrition & Metabolic Care. 19(6):471–476. doi: 10.1097/MCO.0000000000000314. [DOI] [PubMed] [Google Scholar]
- 124.Dueñas M, et al. A survey of modulation of gut microbiota by dietary polyphenols. BioMed research international. 2015 doi: 10.1155/2015/850902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Uranga JA, et al. Food, nutrients and nutraceuticals affecting the course of inflammatory bowel disease. Pharmacol Rep. 68(4):816–26. doi: 10.1016/j.pharep.2016.05.002. [DOI] [PubMed] [Google Scholar]
- 126.Yang N, Sampathkumar K, Loo SC. Recent advances in complementary and replacement therapy with nutraceuticals in combating gastrointestinal illnesses. Clin Nutr. doi: 10.1016/j.clnu.2016.08.020. [DOI] [PubMed] [Google Scholar]
- 127.Cardona F, et al. Benefits of polyphenols on gut microbiota and implications in human health. J Nutr Biochem. 24(8):1415–22. doi: 10.1016/j.jnutbio.2013.05.001. [DOI] [PubMed] [Google Scholar]
- 128.Ikarashi N, et al. Epigallocatechin gallate induces a hepatospecific decrease in the CYP3A expression level by altering intestinal flora. Eur J Pharm Sci. 100:211–218. doi: 10.1016/j.ejps.2017.01.022. [DOI] [PubMed] [Google Scholar]
- 129.Oz HS, Chen T, de Villiers WJ. Green Tea Polyphenols and Sulfasalazine have Parallel Anti-Inflammatory Properties in Colitis Models. Front Immunol. 4:132. doi: 10.3389/fimmu.2013.00132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Steinmann J, et al. Anti-infective properties of epigallocatechin-3-gallate (EGCG), a component of green tea. Br J Pharmacol. 168(5):1059–73. doi: 10.1111/bph.12009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Unno T, Sakuma M, Mitsuhashi S. Effect of dietary supplementation of (-)-epigallocatechin gallate on gut microbiota and biomarkers of colonic fermentation in rats. J Nutr Sci Vitaminol (Tokyo) 60(3):213–9. doi: 10.3177/jnsv.60.213. [DOI] [PubMed] [Google Scholar]
- 132.Henning SM, et al. Pomegranate ellagitannins stimulate the growth of Akkermansia muciniphila in vivo. Anaerobe. 43:56–60. doi: 10.1016/j.anaerobe.2016.12.003. [DOI] [PubMed] [Google Scholar]
- 133.Landete JM, et al. Bioactivation of Phytoestrogens: Intestinal Bacteria and Health. Crit Rev Food Sci Nutr. 56(11):1826–43. doi: 10.1080/10408398.2013.789823. [DOI] [PubMed] [Google Scholar]
- 134.Chen L, Tai WCS, Hsiao WLW. Dietary saponins from four popular herbal tea exert prebiotic-like effects on gut microbiota in C57BL/6 mice. Journal of Functional Foods. 17:892–902. [Google Scholar]
- 135.Li Z, et al. Pomegranate extract induces ellagitannin metabolite formation and changes stool microbiota in healthy volunteers. Food Funct. 6(8):2487–95. doi: 10.1039/c5fo00669d. [DOI] [PubMed] [Google Scholar]
- 136.Barroso E, et al. Phylogenetic profile of gut microbiota in healthy adults after moderate intake of red wine. Molecular Nutrition & Food Research. 61(3) doi: 10.1002/mnfr.201600620. [DOI] [PubMed] [Google Scholar]
- 137.Cueva C, et al. An Integrated View of the Effects of Wine Polyphenols and Their Relevant Metabolites on Gut and Host Health. Molecules. 22(1) doi: 10.3390/molecules22010099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Ghouri YA, et al. Systematic review of randomized controlled trials of probiotics, prebiotics, and synbiotics in inflammatory bowel disease. Clin Exp Gastroenterol. 7:473–87. doi: 10.2147/CEG.S27530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Parkar SG, Stevenson DE, Skinner MA. The potential influence of fruit polyphenols on colonic microflora and human gut health. Int J Food Microbiol. 2008;124(3):295–8. doi: 10.1016/j.ijfoodmicro.2008.03.017. [DOI] [PubMed] [Google Scholar]
- 140.Remely M, et al. EGCG Prevents High Fat Diet-Induced Changes in Gut Microbiota, Decreases of DNA Strand Breaks, and Changes in Expression and DNA Methylation of Dnmt1 and MLH1 in C57BL/6J Male Mice. Oxidative Medicine and Cellular Longevity. 2017 doi: 10.1155/2017/3079148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Sheflin AM, et al. Linking dietary patterns with gut microbial composition and function. Gut Microbes. 8(2):113–129. doi: 10.1080/19490976.2016.1270809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Smoliga JM, Blanchard O. Enhancing the delivery of resveratrol in humans: if low bioavailability is the problem, what is the solution? Molecules. 19(11):17154–72. doi: 10.3390/molecules191117154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Stevens JF, Maier CS. The Chemistry of Gut Microbial Metabolism of Polyphenols. Phytochem Rev. 15(3):425–444. doi: 10.1007/s11101-016-9459-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Duda-Chodak A. The inhibitory effect of polyphenols on human gut microbiota. J Physiol Pharmacol. 63(5):497–503. [PubMed] [Google Scholar]
- 145.Duda-Chodak A, et al. Interaction of dietary compounds, especially polyphenols, with the intestinal microbiota: a review. Eur J Nutr. 54(3):325–41. doi: 10.1007/s00394-015-0852-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Hervert-Hernandez D, Goñi I. Dietary polyphenols and human gut microbiota: a review. Food Reviews International. 27(2):154–169. [Google Scholar]
- 147.Laparra JMs, Sanz Y. Interactions of gut microbiota with functional food components and nutraceuticals. Pharmacological Research. 61(3):219–225. doi: 10.1016/j.phrs.2009.11.001. [DOI] [PubMed] [Google Scholar]
- 148.Camuesco D, et al. Intestinal anti-inflammatory activity of combined quercitrin and dietary olive oil supplemented with fish oil, rich in EPA and DHA (n-3) polyunsaturated fatty acids, in rats with DSS-induced colitis. Clin Nutr. 2006;25(3):466–76. doi: 10.1016/j.clnu.2005.12.009. [DOI] [PubMed] [Google Scholar]
- 149.Li W, et al. Combined use of vitamin D3 and metformin exhibits synergistic chemopreventive effects on colorectal neoplasia in rats and mice. Cancer Prev Res (Phila) 8(2):139–48. doi: 10.1158/1940-6207.CAPR-14-0128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Patten GS, et al. Site specific delivery of microencapsulated fish oil to the gastrointestinal tract of the rat. Digestive diseases and sciences. 2009;54(3):511–521. doi: 10.1007/s10620-008-0379-7. [DOI] [PubMed] [Google Scholar]
- 151.Zator ZA, et al. Pretreatment 25-hydroxyvitamin D levels and durability of anti-tumor necrosis factor-alpha therapy in inflammatory bowel diseases. JPEN J Parenter Enteral Nutr. 38(3):385–91. doi: 10.1177/0148607113504002. [DOI] [PubMed] [Google Scholar]
- 152.Goff JP, et al. Targeted delivery of vitamin D to the colon using beta-glucuronides of vitamin D: therapeutic effects in a murine model of inflammatory bowel disease. Am J Physiol Gastrointest Liver Physiol. 302(4):G460–9. doi: 10.1152/ajpgi.00156.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Woloszynska-Read A, Johnson CS, Trump DL. Vitamin D and cancer: clinical aspects. Best Pract Res Clin Endocrinol Metab. 25(4):605–15. doi: 10.1016/j.beem.2011.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Dhaneshwar SS, Kandpal M, Vadnerkar G. L-glutamine conjugate of meselamine: a novel approach for targeted delivery to colon. Journal of Drug Delivery Science and Technology. 2009;19(1):67–72. [Google Scholar]
- 155.Teng Z, Luo Y, Wang Q. Carboxymethyl chitosan-soy protein complex nanoparticles for the encapsulation and controlled release of vitamin D(3) Food Chem. 141(1):524–32. doi: 10.1016/j.foodchem.2013.03.043. [DOI] [PubMed] [Google Scholar]
- 156.Das S, Ng KY. Colon-specific delivery of resveratrol: optimization of multi-particulate calcium-pectinate carrier. Int J Pharm. 385(1–2):20–8. doi: 10.1016/j.ijpharm.2009.10.016. [DOI] [PubMed] [Google Scholar]
- 157.Almouazen E, et al. Nano-encapsulation of vitamin D3 active metabolites for application in chemotherapy: formulation study and in vitro evaluation. Pharmaceutical research. 30(4):1137–1146. doi: 10.1007/s11095-012-0949-4. [DOI] [PubMed] [Google Scholar]
- 158.Beloqui A, et al. pH-sensitive nanoparticles for colonic delivery of curcumin in inflammatory bowel disease. Int J Pharm. 473(1–2):203–12. doi: 10.1016/j.ijpharm.2014.07.009. [DOI] [PubMed] [Google Scholar]
- 159.Sareen R, et al. Curcumin loaded microsponges for colon targeting in inflammatory bowel disease: fabrication, optimization, and in vitro and pharmacodynamic evaluation. Biomed Res Int. 2014:340701. doi: 10.1155/2014/340701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Coradini K, et al. A novel approach to arthritis treatment based on resveratrol and curcumin co-encapsulated in lipid-core nanocapsules: In vivo studies. Eur J Pharm Sci. 78:163–70. doi: 10.1016/j.ejps.2015.07.012. [DOI] [PubMed] [Google Scholar]
- 161.Aditya NP, et al. Co-delivery of hydrophobic curcumin and hydrophilic catechin by a water-in-oil-in-water double emulsion. Food Chem. 173:7–13. doi: 10.1016/j.foodchem.2014.09.131. [DOI] [PubMed] [Google Scholar]
- 162.Li Q, et al. Curcumin-piperine mixtures in self-microemulsifying drug delivery system for ulcerative colitis therapy. Int J Pharm. 490(1–2):22–31. doi: 10.1016/j.ijpharm.2015.05.008. [DOI] [PubMed] [Google Scholar]
- 163.Tavano L, et al. Co-encapsulation of antioxidants into niosomal carriers: gastrointestinal release studies for nutraceutical applications. Colloids Surf B Biointerfaces. 114:82–8. doi: 10.1016/j.colsurfb.2013.09.058. [DOI] [PubMed] [Google Scholar]
- 164.Ganta S, Amiji M. Coadministration of Paclitaxel and curcumin in nanoemulsion formulations to overcome multidrug resistance in tumor cells. Mol Pharm. 2009;6(3):928–39. doi: 10.1021/mp800240j. [DOI] [PubMed] [Google Scholar]
- 165.Okuro PK, et al. Co-encapsulation of Lactobacillus acidophilus with inulin or polydextrose in solid lipid microparticles provides protection and improves stability. Food Research International. 53(1):96–103. [Google Scholar]
- 166.Lee WL, Widjaja E, Loo SC. One-step fabrication of triple-layered polymeric microparticles with layer localization of drugs as a novel drug-delivery system. Small. 6(9):1003–11. doi: 10.1002/smll.200901985. [DOI] [PubMed] [Google Scholar]
- 167.DiPatrizio NV, Piomelli D. Intestinal lipid-derived signals that sense dietary fat. J Clin Invest. 125(3):891–8. doi: 10.1172/JCI76302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.DiPatrizio NV, Piomelli D. The thrifty lipids: endocannabinoids and the neural control of energy conservation. Trends Neurosci. 35(7):403–11. doi: 10.1016/j.tins.2012.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Izzo AA, Sharkey KA. Cannabinoids and the gut: new developments and emerging concepts. Pharmacol Ther. 126(1):21–38. doi: 10.1016/j.pharmthera.2009.12.005. [DOI] [PubMed] [Google Scholar]
- 170.Adami M, et al. Gastric antisecretory role and immunohistochemical localization of cannabinoid receptors in the rat stomach. Br J Pharmacol. 2002;135(7):1598–606. doi: 10.1038/sj.bjp.0704625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Adami M, et al. Gastric antisecretory effects of synthetic cannabinoids after central or peripheral administration in the rat. Brain Res Bull. 2004;64(4):357–61. doi: 10.1016/j.brainresbull.2004.09.004. [DOI] [PubMed] [Google Scholar]
- 172.Coruzzi G, et al. Effects of cannabinoid receptor agonists on rat gastric acid secretion: discrepancy between in vitro and in vivo data. Dig Dis Sci. 2006;51(2):310–7. doi: 10.1007/s10620-006-3130-2. [DOI] [PubMed] [Google Scholar]
- 173.DiPatrizio NV. Endocannabinoids in the Gut. Cannabis Cannabinoid Res. 1(1):67–77. doi: 10.1089/can.2016.0001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Izzo AA, et al. Defaecation, intestinal fluid accumulation and motility in rodents: implications of cannabinoid CB1 receptors. Naunyn Schmiedebergs Arch Pharmacol. 1999;359(1):65–70. doi: 10.1007/pl00005325. [DOI] [PubMed] [Google Scholar]
- 175.Izzo AA, et al. The role of cannabinoid receptors in intestinal motility, defaecation and diarrhoea in rats. Eur J Pharmacol. 1999;384(1):37–42. doi: 10.1016/s0014-2999(99)00673-1. [DOI] [PubMed] [Google Scholar]
- 176.Izzo AA, et al. Cannabinoid CB1-receptor mediated regulation of gastrointestinal motility in mice in a model of intestinal inflammation. Br J Pharmacol. 2001;134(3):563–70. doi: 10.1038/sj.bjp.0704293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Mathison R, et al. Effects of cannabinoid receptor-2 activation on accelerated gastrointestinal transit in lipopolysaccharide-treated rats. Br J Pharmacol. 2004;142(8):1247–54. doi: 10.1038/sj.bjp.0705889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Klein TW, et al. The cannabinoid system and immune modulation. J Leukoc Biol. 2003;74(4):486–96. doi: 10.1189/jlb.0303101. [DOI] [PubMed] [Google Scholar]
- 179.Samson MT, et al. Differential roles of CB1 and CB2 cannabinoid receptors in mast cells. J Immunol. 2003;170(10):4953–62. doi: 10.4049/jimmunol.170.10.4953. [DOI] [PubMed] [Google Scholar]
- 180.Kulkarni-Narla A, Brown DR. Localization of CB1-cannabinoid receptor immunoreactivity in the porcine enteric nervous system. Cell Tissue Res. 2000;302(1):73–80. doi: 10.1007/s004410000261. [DOI] [PubMed] [Google Scholar]
- 181.Lynn AB, Herkenham M. Localization of cannabinoid receptors and nonsaturable high-density cannabinoid binding sites in peripheral tissues of the rat: implications for receptor-mediated immune modulation by cannabinoids. J Pharmacol Exp Ther. 1994;268(3):1612–23. [PubMed] [Google Scholar]
- 182.Trautmann SM, Sharkey KA. The Endocannabinoid System and Its Role in Regulating the Intrinsic Neural Circuitry of the Gastrointestinal Tract. Int Rev Neurobiol. 125:85–126. doi: 10.1016/bs.irn.2015.10.002. [DOI] [PubMed] [Google Scholar]
- 183.Pertwee RG. Pharmacology of cannabinoid CB1 and CB2 receptors. Pharmacol Ther. 1997;74(2):129–80. doi: 10.1016/s0163-7258(97)82001-3. [DOI] [PubMed] [Google Scholar]
- 184.Devane WA, et al. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science. 1992;258(5090):1946–9. doi: 10.1126/science.1470919. [DOI] [PubMed] [Google Scholar]
- 185.Cani PD, et al. Endocannabinoids–at the crossroads between the gut microbiota and host metabolism. Nat Rev Endocrinol. 12(3):133–43. doi: 10.1038/nrendo.2015.211. [DOI] [PubMed] [Google Scholar]
- 186.De Petrocellis L, Di Marzo V. Non-CB1, non-CB2 receptors for endocannabinoids, plant cannabinoids, and synthetic cannabimimetics: focus on G-protein-coupled receptors and transient receptor potential channels. J Neuroimmune Pharmacol. 5(1):103–21. doi: 10.1007/s11481-009-9177-z. [DOI] [PubMed] [Google Scholar]
- 187.Syed SK, et al. Regulation of GPR119 receptor activity with endocannabinoid-like lipids. Am J Physiol Endocrinol Metab. 303(12):E1469–78. doi: 10.1152/ajpendo.00269.2012. [DOI] [PubMed] [Google Scholar]
- 188.Capasso R, et al. Palmitoylethanolamide normalizes intestinal motility in a model of post-inflammatory accelerated transit: involvement of CB(1) receptors and TRPV1 channels. Br J Pharmacol. 171(17):4026–37. doi: 10.1111/bph.12759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Izzo AA, et al. Peripheral endocannabinoid dysregulation in obesity: relation to intestinal motility and energy processing induced by food deprivation and re-feeding. Br J Pharmacol. 2009;158(2):451–61. doi: 10.1111/j.1476-5381.2009.00183.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Ben-Shabat S, et al. An entourage effect: inactive endogenous fatty acid glycerol esters enhance 2-arachidonoyl-glycerol cannabinoid activity. Eur J Pharmacol. 1998;353(1):23–31. doi: 10.1016/s0014-2999(98)00392-6. [DOI] [PubMed] [Google Scholar]
- 191.Cani PD, et al. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes. 2008;57(6):1470–81. doi: 10.2337/db07-1403. [DOI] [PubMed] [Google Scholar]
- 192.Drucker DJ. Glucagon-like peptides. Diabetes. 1998;47(2):159–69. doi: 10.2337/diab.47.2.159. [DOI] [PubMed] [Google Scholar]
- 193.Lambert DM, Di Marzo V. The palmitoylethanolamide and oleamide enigmas: are these two fatty acid amides cannabimimetic? Curr Med Chem. 1999;6(8):757–73. [PubMed] [Google Scholar]
- 194.Alhamoruni A, et al. Cannabinoids mediate opposing effects on inflammation-induced intestinal permeability. Br J Pharmacol. 165(8):2598–610. doi: 10.1111/j.1476-5381.2011.01589.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Massa F, et al. The endogenous cannabinoid system protects against colonic inflammation. J Clin Invest. 2004;113(8):1202–9. doi: 10.1172/JCI19465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Storr MA, et al. Targeting endocannabinoid degradation protects against experimental colitis in mice: involvement of CB1 and CB2 receptors. J Mol Med (Berl) 2008;86(8):925–36. doi: 10.1007/s00109-008-0359-6. [DOI] [PubMed] [Google Scholar]
- 197.Storr MA, et al. Activation of the cannabinoid 2 receptor (CB2) protects against experimental colitis. Inflamm Bowel Dis. 2009;15(11):1678–85. doi: 10.1002/ibd.20960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Turcotte C, et al. Regulation of inflammation by cannabinoids, the endocannabinoids 2-arachidonoyl-glycerol and arachidonoyl-ethanolamide, and their metabolites. J Leukoc Biol. 97(6):1049–70. doi: 10.1189/jlb.3RU0115-021R. [DOI] [PubMed] [Google Scholar]
- 199.Alhouayek M, Muccioli GG. Harnessing the anti-inflammatory potential of palmitoylethanolamide. Drug Discov Today. 19(10):1632–9. doi: 10.1016/j.drudis.2014.06.007. [DOI] [PubMed] [Google Scholar]
- 200.Borrelli F, et al. Palmitoylethanolamide, a naturally occurring lipid, is an orally effective intestinal anti-inflammatory agent. Br J Pharmacol. 172(1):142–58. doi: 10.1111/bph.12907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.D’Argenio G, et al. Overactivity of the intestinal endocannabinoid system in celiac disease and in methotrexate-treated rats. J Mol Med (Berl) 2007;85(5):523–30. doi: 10.1007/s00109-007-0192-3. [DOI] [PubMed] [Google Scholar]
- 202.Di Marzo V, Izzo AA. Endocannabinoid overactivity and intestinal inflammation. Gut. 2006;55(10):1373–6. doi: 10.1136/gut.2005.090472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Esposito G, et al. Palmitoylethanolamide improves colon inflammation through an enteric glia/toll like receptor 4-dependent PPAR-alpha activation. Gut. 63(8):1300–12. doi: 10.1136/gutjnl-2013-305005. [DOI] [PubMed] [Google Scholar]
- 204.Guagnini F, et al. Neural contractions in colonic strips from patients with diverticular disease: role of endocannabinoids and substance P. Gut. 2006;55(7):946–53. doi: 10.1136/gut.2005.076372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Ligresti A, et al. Possible endocannabinoid control of colorectal cancer growth. Gastroenterology. 2003;125(3):677–87. doi: 10.1016/s0016-5085(03)00881-3. [DOI] [PubMed] [Google Scholar]
- 206.Smid SD. Gastrointestinal endocannabinoid system: multifaceted roles in the healthy and inflamed intestine. Clin Exp Pharmacol Physiol. 2008;35(11):1383–7. doi: 10.1111/j.1440-1681.2008.05016.x. [DOI] [PubMed] [Google Scholar]
- 207.Muccioli GG, et al. The endocannabinoid system links gut microbiota to adipogenesis. Mol Syst Biol. 6:392. doi: 10.1038/msb.2010.46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Rousseaux C, et al. Lactobacillus acidophilus modulates intestinal pain and induces opioid and cannabinoid receptors. Nat Med. 2007;13(1):35–7. doi: 10.1038/nm1521. [DOI] [PubMed] [Google Scholar]
- 209.Everard A, et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc Natl Acad Sci U S A. 110(22):9066–71. doi: 10.1073/pnas.1219451110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Everard A, et al. Intestinal epithelial MyD88 is a sensor switching host metabolism towards obesity according to nutritional status. Nat Commun. 5:5648. doi: 10.1038/ncomms6648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Lal S, et al. Cannabis use amongst patients with inflammatory bowel disease. Eur J Gastroenterol Hepatol. 23(10):891–6. doi: 10.1097/MEG.0b013e328349bb4c. [DOI] [PubMed] [Google Scholar]
- 212.Naftali T, et al. Treatment of Crohn’s disease with cannabis: an observational study. Isr Med Assoc J. 13(8):455–8. [PubMed] [Google Scholar]
- 213.Lahat A, Lang A, Ben-Horin S. Impact of cannabis treatment on the quality of life, weight and clinical disease activity in inflammatory bowel disease patients: a pilot prospective study. Digestion. 85(1):1–8. doi: 10.1159/000332079. [DOI] [PubMed] [Google Scholar]
- 214.Naftali T, et al. Cannabis induces a clinical response in patients with Crohn’s disease: a prospective placebo-controlled study. Clin Gastroenterol Hepatol. 11(10):1276–1280 e1. doi: 10.1016/j.cgh.2013.04.034. [DOI] [PubMed] [Google Scholar]
- 215.Kahan M, et al. Prescribing smoked cannabis for chronic noncancer pain: preliminary recommendations. Can Fam Physician. 60(12):1083–90. [PMC free article] [PubMed] [Google Scholar]
- 216.Cani PD, Everard A. Talking microbes: When gut bacteria interact with diet and host organs. Mol Nutr Food Res. 60(1):58–66. doi: 10.1002/mnfr.201500406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Banni S, et al. Krill oil significantly decreases 2-arachidonoylglycerol plasma levels in obese subjects. Nutr Metab (Lond) 8(1):7. doi: 10.1186/1743-7075-8-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Kim HJ, et al. Contributions of microbiome and mechanical deformation to intestinal bacterial overgrowth and inflammation in a human gut-on-a-chip. Proc Natl Acad Sci U S A. 113(1):E7–15. doi: 10.1073/pnas.1522193112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Kim J, Li Y, Watkins BA. Fat to treat fat: emerging relationship between dietary PUFA, endocannabinoids, and obesity. Prostaglandins Other Lipid Mediat. 104–105:32–41. doi: 10.1016/j.prostaglandins.2012.11.005. [DOI] [PubMed] [Google Scholar]
- 220.Brown I, et al. Cannabinoid receptor-dependent and -independent anti-proliferative effects of omega-3 ethanolamides in androgen receptor-positive and -negative prostate cancer cell lines. Carcinogenesis. 31(9):1584–91. doi: 10.1093/carcin/bgq151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Daniels GE. Psychiatric aspects of ulcerative colitis. New England Journal of Medicine. 1942;226(5):178–184. [Google Scholar]
- 222.Murray CD. Psychogenic factors in the etiology of ulcerative colitis and bloody diarrhea. The American Journal of the Medical Sciences. 1930;180(2):239–247. [Google Scholar]
- 223.Straker M. Ulcerative colitis: recovery of a patient with brief psychiatric treatment. Can Med Assoc J. 1960;82:1224–7. [PMC free article] [PubMed] [Google Scholar]
- 224.Sullivan AJ, Chandler CA. Ulcerative Colitis of Psychogenic Origin: A Report of Six Cases. Yale J Biol Med. 1932;4(6):779–96. [PMC free article] [PubMed] [Google Scholar]
- 225.Goldsmith DR, Rapaport MH, Miller BJ. A meta-analysis of blood cytokine network alterations in psychiatric patients: comparisons between schizophrenia, bipolar disorder and depression. Mol Psychiatry. 21(12):1696–1709. doi: 10.1038/mp.2016.3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Abautret-Daly A, et al. Gut-brain actions underlying comorbid anxiety and depression associated with inflammatory bowel disease. Acta Neuropsychiatr. :1–22. doi: 10.1017/neu.2017.3. [DOI] [PubMed] [Google Scholar]
- 227.Camilleri M, et al. Intestinal barrier function in health and gastrointestinal disease. Neurogastroenterol Motil. 24(6):503–12. doi: 10.1111/j.1365-2982.2012.01921.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Matsunaga H, et al. Physiological stress exacerbates murine colitis by enhancing proinflammatory cytokine expression that is dependent on IL-18. Am J Physiol Gastrointest Liver Physiol. 301(3):G555–64. doi: 10.1152/ajpgi.00482.2010. [DOI] [PubMed] [Google Scholar]
- 229.Melgar S, et al. Psychological stress reactivates dextran sulfate sodium-induced chronic colitis in mice. Stress. 2008;11(5):348–62. doi: 10.1080/10253890701820166. [DOI] [PubMed] [Google Scholar]
- 230.Singh S, Graff LA, Bernstein CN. Do NSAIDs, antibiotics, infections, or stress trigger flares in IBD? Am J Gastroenterol. 2009;104(5):1298–313. doi: 10.1038/ajg.2009.15. quiz 1314. [DOI] [PubMed] [Google Scholar]
- 231.Bendtsen KMB, et al. Gut microbiota composition is correlated to grid floor induced stress and behavior in the BALB/c mouse. PLoS One. 7(10):e46231. doi: 10.1371/journal.pone.0046231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Lutgendorff F, Akkermans LM, Soderholm JD. The role of microbiota and probiotics in stress-induced gastro-intestinal damage. Curr Mol Med. 2008;8(4):282–98. doi: 10.2174/156652408784533779. [DOI] [PubMed] [Google Scholar]
- 233.David LA, et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature. 505(7484):559–63. doi: 10.1038/nature12820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Sonnenburg ED, et al. Diet-induced extinctions in the gut microbiota compound over generations. Nature. 529(7585):212–5. doi: 10.1038/nature16504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Wu GD, et al. Linking long-term dietary patterns with gut microbial enterotypes. Science. 334(6052):105–8. doi: 10.1126/science.1208344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Magnusdottir S, et al. Systematic genome assessment of B-vitamin biosynthesis suggests co-operation among gut microbes. Front Genet. 6:148. doi: 10.3389/fgene.2015.00148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Zempleni J, et al. Handbook of vitamins. CRC Press [Google Scholar]
- 238.Wallace TC, et al. New Frontiers in Fibers: Innovative and Emerging Research on the Gut Microbiome and Bone Health. J Am Coll Nutr. :1–5. doi: 10.1080/07315724.2016.1257961. [DOI] [PubMed] [Google Scholar]
- 239.Lee D, et al. Diet in the pathogenesis and treatment of inflammatory bowel diseases. Gastroenterology. 148(6):1087–106. doi: 10.1053/j.gastro.2015.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Dey N, et al. Regulators of gut motility revealed by a gnotobiotic model of diet-microbiome interactions related to travel. Cell. 163(1):95–107. doi: 10.1016/j.cell.2015.08.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Basson A, et al. Mucosal Interactions between Genetics, Diet, and Microbiome in Inflammatory Bowel Disease. Front Immunol. 7:290. doi: 10.3389/fimmu.2016.00290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Donohoe DR, et al. The microbiome and butyrate regulate energy metabolism and autophagy in the mammalian colon. Cell Metab. 13(5):517–26. doi: 10.1016/j.cmet.2011.02.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Wanders D, Graff EC, Judd RL. Effects of high fat diet on GPR109A and GPR81 gene expression. Biochem Biophys Res Commun. 425(2):278–83. doi: 10.1016/j.bbrc.2012.07.082. [DOI] [PubMed] [Google Scholar]
- 244.Hoffmann C, et al. Archaea and fungi of the human gut microbiome: correlations with diet and bacterial residents. PLoS One. 8(6):e66019. doi: 10.1371/journal.pone.0066019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.De Filippo C, et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proceedings of the National Academy of Sciences. 107(33):14691–14696. doi: 10.1073/pnas.1005963107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246.Yatsunenko T, et al. Human gut microbiome viewed across age and geography. Nature. 486(7402):222–7. doi: 10.1038/nature11053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Attene-Ramos MS, et al. Evidence that hydrogen sulfide is a genotoxic agent. Mol Cancer Res. 2006;4(1):9–14. doi: 10.1158/1541-7786.MCR-05-0126. [DOI] [PubMed] [Google Scholar]
- 248.Baron EJ, et al. Bilophila wadsworthia, gen. nov. and sp. nov., a unique gram-negative anaerobic rod recovered from appendicitis specimens and human faeces. J Gen Microbiol. 1989;135(12):3405–11. doi: 10.1099/00221287-135-12-3405. [DOI] [PubMed] [Google Scholar]
- 249.Martinez-Medina M, et al. Western diet induces dysbiosis with increased E coli in CEABAC10 mice, alters host barrier function favouring AIEC colonisation. Gut. 63(1):116–24. doi: 10.1136/gutjnl-2012-304119. [DOI] [PubMed] [Google Scholar]
- 250.Jakobsdottir G, et al. High-fat diet reduces the formation of butyrate, but increases succinate, inflammation, liver fat and cholesterol in rats, while dietary fibre counteracts these effects. PLoS One. 8(11):e80476. doi: 10.1371/journal.pone.0080476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251.Mane J, et al. Partial replacement of dietary (n-6) fatty acids with medium-chain triglycerides decreases the incidence of spontaneous colitis in interleukin-10-deficient mice. J Nutr. 2009;139(3):603–10. doi: 10.3945/jn.108.101170. [DOI] [PubMed] [Google Scholar]
- 252.Ohta N, et al. A comparison of the effects of medium- and long-chain triglycerides on neutrophil stimulation in experimental ileitis. J Gastroenterol. 2003;38(2):127–33. doi: 10.1007/s005350300021. [DOI] [PubMed] [Google Scholar]
- 253.Tsujikawa T, et al. Medium-chain triglycerides modulate ileitis induced by trinitrobenzene sulfonic acid. J Gastroenterol Hepatol. 1999;14(12):1166–72. doi: 10.1046/j.1440-1746.1999.02024.x. [DOI] [PubMed] [Google Scholar]
- 254.Clarke SF, et al. Exercise and associated dietary extremes impact on gut microbial diversity. Gut. 63(12):1913–20. doi: 10.1136/gutjnl-2013-306541. [DOI] [PubMed] [Google Scholar]
- 255.Wang Z, et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature. 472(7341):57–63. doi: 10.1038/nature09922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 256.Koeth RA, et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat Med. 19(5):576–85. doi: 10.1038/nm.3145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257.Paturi G, et al. Dietary combination of potato resistant starch and red meat up-regulates genes involved in colonic barrier function of rats. International Journal of Food Science & Technology. 48(11):2441–2446. [Google Scholar]
- 258.Paturi G, et al. Effects of potato fiber and potato-resistant starch on biomarkers of colonic health in rats fed diets containing red meat. J Food Sci. 77(10):H216–23. doi: 10.1111/j.1750-3841.2012.02911.x. [DOI] [PubMed] [Google Scholar]
- 259.De Filippis F, et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut. 65(11):1812–1821. doi: 10.1136/gutjnl-2015-309957. [DOI] [PubMed] [Google Scholar]
- 260.Cotillard A, et al. Dietary intervention impact on gut microbial gene richness. Nature. 500(7464):585–8. doi: 10.1038/nature12480. [DOI] [PubMed] [Google Scholar]
- 261.Brugere JF, et al. Archaebiotics: proposed therapeutic use of archaea to prevent trimethylaminuria and cardiovascular disease. Gut Microbes. 5(1):5–10. doi: 10.4161/gmic.26749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262.Horz HP. Archaeal Lineages within the Human Microbiome: Absent, Rare or Elusive? Life (Basel) 5(2):1333–45. doi: 10.3390/life5021333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.Gaci N, et al. Archaea and the human gut: new beginning of an old story. World J Gastroenterol. 20(43):16062–78. doi: 10.3748/wjg.v20.i43.16062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 264.Marchand LcL, et al. Breath hydrogen and methane in populations at different risk for colon cancer. International journal of cancer. 1993;55(6):887–890. doi: 10.1002/ijc.2910550603. [DOI] [PubMed] [Google Scholar]
- 265.Nam YD, et al. Bacterial, archaeal, and eukaryal diversity in the intestines of Korean people. J Microbiol. 2008;46(5):491–501. doi: 10.1007/s12275-008-0199-7. [DOI] [PubMed] [Google Scholar]
- 266.Nava GM, et al. Hydrogenotrophic microbiota distinguish native Africans from African and European Americans. Environ Microbiol Rep. 4(3):307–15. doi: 10.1111/j.1758-2229.2012.00334.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 267.Segal I, et al. Breath methane and large bowel cancer risk in contrasting African populations. Gut. 1988;29(5):608–13. doi: 10.1136/gut.29.5.608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 268.Tyakht AV, et al. Human gut microbiota community structures in urban and rural populations in Russia. Nat Commun. 4:2469. doi: 10.1038/ncomms3469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 269.Rucker RB, Steinberg FM, Johnston CS. Handbook of Vitamins. Fourth. CRC Press; 2007. Ascorbic acid. [Google Scholar]
- 270.Cantorna MT, Mahon BD. D-hormone and the immune system. J Rheumatol Suppl. 2005;76:11–20. [PubMed] [Google Scholar]
- 271.Cantorna MT, et al. Vitamin D status, 1,25-dihydroxyvitamin D3, and the immune system. Am J Clin Nutr. 2004;80(6 Suppl):1717S–20S. doi: 10.1093/ajcn/80.6.1717S. [DOI] [PubMed] [Google Scholar]
- 272.Lim WC, Hanauer SB, Li YC. Mechanisms of disease: vitamin D and inflammatory bowel disease. Nat Clin Pract Gastroenterol Hepatol. 2005;2(7):308–15. doi: 10.1038/ncpgasthep0215. [DOI] [PubMed] [Google Scholar]
- 273.Jin D, et al. Lack of vitamin D receptor causes dysbiosis and changes the functions of the murine intestinal microbiome. Clinical therapeutics. 37(5):996–1009.e7. doi: 10.1016/j.clinthera.2015.04.004. [DOI] [PubMed] [Google Scholar]
- 274.Rehman A, et al. Transcriptional activity of the dominant gut mucosal microbiota in chronic inflammatory bowel disease patients. J Med Microbiol. 59(Pt 9):1114–22. doi: 10.1099/jmm.0.021170-0. [DOI] [PubMed] [Google Scholar]
- 275.Appleyard CB, et al. Pretreatment with the probiotic VSL#3 delays transition from inflammation to dysplasia in a rat model of colitis-associated cancer. Am J Physiol Gastrointest Liver Physiol. 301(6):G1004–13. doi: 10.1152/ajpgi.00167.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 276.Basson A. Vitamin D and Crohn’s disease in the adult patient: a review. JPEN J Parenter Enteral Nutr. 38(4):438–58. doi: 10.1177/0148607113506013. [DOI] [PubMed] [Google Scholar]
- 277.Jones ML, Martoni CJ, Prakash S. Oral supplementation with probiotic L. reuteri NCIMB 30242 increases mean circulating 25-hydroxyvitamin D: a post hoc analysis of a randomized controlled trial. J Clin Endocrinol Metab. 98(7):2944–51. doi: 10.1210/jc.2012-4262. [DOI] [PubMed] [Google Scholar]
- 278.Kornbluth A, Sachar DB. Ulcerative colitis practice guidelines in adults (update): American College of Gastroenterology, Practice Parameters Committee. Am J Gastroenterol. 2004;99(7):1371–85. doi: 10.1111/j.1572-0241.2004.40036.x. [DOI] [PubMed] [Google Scholar]
- 279.Lichtenstein GR, Hanauer SB, Sandborn WJ. Management of Crohn’s disease in adults. Am J Gastroenterol. 2009;104(2):465–83. doi: 10.1038/ajg.2008.168. quiz 464, 484. [DOI] [PubMed] [Google Scholar]
- 280.Turner D, et al. Management of pediatric ulcerative colitis: joint ECCO and ESPGHAN evidence-based consensus guidelines. J Pediatr Gastroenterol Nutr. 55(3):340–61. doi: 10.1097/MPG.0b013e3182662233. [DOI] [PubMed] [Google Scholar]
- 281.Owczarek D, et al. Diet and nutritional factors in inflammatory bowel diseases. World J Gastroenterol. 22(3):895–905. doi: 10.3748/wjg.v22.i3.895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 282.Hou JK, Lee D, Lewis J. Diet and inflammatory bowel disease: review of patient-targeted recommendations. Clin Gastroenterol Hepatol. 12(10):1592–600. doi: 10.1016/j.cgh.2013.09.063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 283.Brown S, et al. De novo production of the plant-derived alkaloid strictosidine in yeast. Proc Natl Acad Sci U S A. 112(11):3205–10. doi: 10.1073/pnas.1423555112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 284.Basson A. Nutrition management in the adult patient with Crohn’s disease. South African Journal of Clinical Nutrition. 25(4):164–172. [Google Scholar]
- 285.Donnellan CF, Yann LH, Lal S. Nutritional management of Crohn’s disease. Therap Adv Gastroenterol. 6(3):231–42. doi: 10.1177/1756283X13477715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 286.Halmos EP, et al. Diets that differ in their FODMAP content alter the colonic luminal microenvironment. Gut. 64(1):93–100. doi: 10.1136/gutjnl-2014-307264. [DOI] [PubMed] [Google Scholar]
- 287.Staudacher HM, et al. Fermentable carbohydrate restriction reduces luminal bifidobacteria and gastrointestinal symptoms in patients with irritable bowel syndrome. J Nutr. 142(8):1510–8. doi: 10.3945/jn.112.159285. [DOI] [PubMed] [Google Scholar]
- 288.Flanagan P, Campbell BJ, Rhodes JM. Bacteria in the pathogenesis of inflammatory bowel disease. Biochem Soc Trans. 39(4):1067–72. doi: 10.1042/BST0391067. [DOI] [PubMed] [Google Scholar]
- 289.Martin HM, et al. Enhanced Escherichia coli adherence and invasion in Crohn’s disease and colon cancer. Gastroenterology. 2004;127(1):80–93. doi: 10.1053/j.gastro.2004.03.054. [DOI] [PubMed] [Google Scholar]
- 290.Parsons BN, et al. Dietary supplementation with soluble plantain non-starch polysaccharides inhibits intestinal invasion of Salmonella typhimurium in the chicken. PLoS One. 9(2):e87658. doi: 10.1371/journal.pone.0087658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 291.Roberts CL, et al. Soluble plantain fibre blocks adhesion and M-cell translocation of intestinal pathogens. J Nutr Biochem. 24(1):97–103. doi: 10.1016/j.jnutbio.2012.02.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292.Zelante T, et al. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity. 39(2):372–85. doi: 10.1016/j.immuni.2013.08.003. [DOI] [PubMed] [Google Scholar]
- 293.Cohen SA, et al. Clinical and mucosal improvement with specific carbohydrate diet in pediatric Crohn disease. J Pediatr Gastroenterol Nutr. 59(4):516–21. doi: 10.1097/MPG.0000000000000449. [DOI] [PubMed] [Google Scholar]
- 294.Kakodkar S, et al. The Specific Carbohydrate Diet for Inflammatory Bowel Disease: A Case Series. J Acad Nutr Diet. 115(8):1226–32. doi: 10.1016/j.jand.2015.04.016. [DOI] [PubMed] [Google Scholar]
- 295.Obih C, et al. Specific carbohydrate diet for pediatric inflammatory bowel disease in clinical practice within an academic IBD center. Nutrition. 32(4):418–25. doi: 10.1016/j.nut.2015.08.025. [DOI] [PubMed] [Google Scholar]
- 296.Suskind DL, et al. Nutritional therapy in pediatric Crohn disease: the specific carbohydrate diet. J Pediatr Gastroenterol Nutr. 58(1):87–91. doi: 10.1097/MPG.0000000000000103. [DOI] [PubMed] [Google Scholar]
- 297.Chassaing B, et al. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature. 519(7541):92–6. doi: 10.1038/nature14232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 298.Nickerson KP, Chanin R, McDonald C. Deregulation of intestinal anti-microbial defense by the dietary additive, maltodextrin. Gut Microbes. 6(1):78–83. doi: 10.1080/19490976.2015.1005477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 299.Galipeau HJ, et al. Intestinal microbiota modulates gluten-induced immunopathology in humanized mice. Am J Pathol. 185(11):2969–82. doi: 10.1016/j.ajpath.2015.07.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 300.Herfarth HH, et al. Prevalence of a gluten-free diet and improvement of clinical symptoms in patients with inflammatory bowel diseases. Inflamm Bowel Dis. 20(7):1194–7. doi: 10.1097/MIB.0000000000000077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 301.Critch J, et al. Use of enteral nutrition for the control of intestinal inflammation in pediatric Crohn disease. J Pediatr Gastroenterol Nutr. 54(2):298–305. doi: 10.1097/MPG.0b013e318235b397. [DOI] [PubMed] [Google Scholar]
- 302.Lionetti P, et al. Enteral nutrition and microflora in pediatric Crohn’s disease. JPEN J Parenter Enteral Nutr. 2005;29(4 Suppl):S173–5. doi: 10.1177/01486071050290S4S173. discussion S175-8, S184-8. [DOI] [PubMed] [Google Scholar]
- 303.Shiga H, et al. Changes of faecal microbiota in patients with Crohn’s disease treated with an elemental diet and total parenteral nutrition. Dig Liver Dis. 44(9):736–42. doi: 10.1016/j.dld.2012.04.014. [DOI] [PubMed] [Google Scholar]
- 304.Ambrosio CMS, et al. Antimicrobial activity of several essential oils on pathogenic and beneficial bacteria. Industrial Crops and Products. 97:128–136. [Google Scholar]
- 305.Mimee M, Citorik RJ, Lu TK. Microbiome therapeutics - Advances and challenges. Adv Drug Deliv Rev. 105(Pt A):44–54. doi: 10.1016/j.addr.2016.04.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 306.Corr SC, et al. Bacteriocin production as a mechanism for the antiinfective activity of Lactobacillus salivarius UCC118. Proc Natl Acad Sci U S A. 2007;104(18):7617–21. doi: 10.1073/pnas.0700440104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 307.Millette M, et al. Capacity of human nisin- and pediocin-producing lactic Acid bacteria to reduce intestinal colonization by vancomycin-resistant enterococci. Appl Environ Microbiol. 2008;74(7):1997–2003. doi: 10.1128/AEM.02150-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 308.Walsh CJ, et al. In silico identification of bacteriocin gene clusters in the gastrointestinal tract, based on the Human Microbiome Project’s reference genome database. BMC Microbiol. 15:183. doi: 10.1186/s12866-015-0515-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 309.Zheng J, et al. Diversity and dynamics of bacteriocins from human microbiome. Environ Microbiol. 17(6):2133–43. doi: 10.1111/1462-2920.12662. [DOI] [PubMed] [Google Scholar]
- 310.McCarville JL, Caminero A, Verdu EF. Novel perspectives on therapeutic modulation of the gut microbiota. Therap Adv Gastroenterol. 9(4):580–93. doi: 10.1177/1756283X16637819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 311.Reyes A, et al. Gnotobiotic mouse model of phage-bacterial host dynamics in the human gut. Proc Natl Acad Sci U S A. 110(50):20236–41. doi: 10.1073/pnas.1319470110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 312.Rodriguez-Valera F, et al. Explaining microbial population genomics through phage predation. Nat Rev Microbiol. 2009;7(11):828–36. doi: 10.1038/nrmicro2235. [DOI] [PubMed] [Google Scholar]
- 313.Minot S, et al. The human gut virome: inter-individual variation and dynamic response to diet. Genome Res. 21(10):1616–25. doi: 10.1101/gr.122705.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 314.Norman JM, et al. Disease-specific alterations in the enteric virome in inflammatory bowel disease. Cell. 160(3):447–60. doi: 10.1016/j.cell.2015.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 315.Reyes A, et al. Viruses in the faecal microbiota of monozygotic twins and their mothers. Nature. 466(7304):334–8. doi: 10.1038/nature09199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 316.Mills S, et al. Movers and shakers: influence of bacteriophages in shaping the mammalian gut microbiota. Gut Microbes. 4(1):4–16. doi: 10.4161/gmic.22371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 317.Lepage P, et al. Dysbiosis in inflammatory bowel disease: a role for bacteriophages? Gut. 2008;57(3):424–5. doi: 10.1136/gut.2007.134668. [DOI] [PubMed] [Google Scholar]
- 318.Perez-Brocal V, et al. Study of the viral and microbial communities associated with Crohn’s disease: a metagenomic approach. Clin Transl Gastroenterol. 4:e36. doi: 10.1038/ctg.2013.9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 319.Wagner J, et al. Bacteriophages in gut samples from pediatric Crohn’s disease patients: metagenomic analysis using 454 pyrosequencing. Inflamm Bowel Dis. 19(8):1598–608. doi: 10.1097/MIB.0b013e318292477c. [DOI] [PubMed] [Google Scholar]
- 320.Zhang T, et al. RNA viral community in human feces: prevalence of plant pathogenic viruses. PLoS Biol. 2006;4(1):e3. doi: 10.1371/journal.pbio.0040003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 321.Letarov A, Kulikov E. The bacteriophages in human- and animal body-associated microbial communities. J Appl Microbiol. 2009;107(1):1–13. doi: 10.1111/j.1365-2672.2009.04143.x. [DOI] [PubMed] [Google Scholar]
- 322.Lu TK, Collins JJ. Engineered bacteriophage targeting gene networks as adjuvants for antibiotic therapy. Proc Natl Acad Sci U S A. 2009;106(12):4629–34. doi: 10.1073/pnas.0800442106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 323.Edgar R, et al. Reversing bacterial resistance to antibiotics by phage-mediated delivery of dominant sensitive genes. Appl Environ Microbiol. 78(3):744–51. doi: 10.1128/AEM.05741-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 324.Westwater C, et al. Use of genetically engineered phage to deliver antimicrobial agents to bacteria: an alternative therapy for treatment of bacterial infections. Antimicrob Agents Chemother. 2003;47(4):1301–7. doi: 10.1128/AAC.47.4.1301-1307.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 325.Ando H, et al. Engineering Modular Viral Scaffolds for Targeted Bacterial Population Editing. Cell Syst. 1(3):187–196. doi: 10.1016/j.cels.2015.08.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 326.Bikard D, et al. Exploiting CRISPR-Cas nucleases to produce sequence-specific antimicrobials. Nat Biotechnol. 32(11):1146–50. doi: 10.1038/nbt.3043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 327.Citorik RJ, Mimee M, Lu TK. Sequence-specific antimicrobials using efficiently delivered RNA-guided nucleases. Nat Biotechnol. 32(11):1141–5. doi: 10.1038/nbt.3011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 328.Gibson DG, et al. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat Methods. 2009;6(5):343–5. doi: 10.1038/nmeth.1318. [DOI] [PubMed] [Google Scholar]
- 329.Hagens S, et al. Therapy of experimental pseudomonas infections with a nonreplicating genetically modified phage. Antimicrob Agents Chemother. 2004;48(10):3817–22. doi: 10.1128/AAC.48.10.3817-3822.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 330.Kiro R, Shitrit D, Qimron U. Efficient engineering of a bacteriophage genome using the type I-E CRISPR-Cas system. RNA Biol. 11(1):42–4. doi: 10.4161/rna.27766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 331.Riley PA. Bacteriophages in autoimmune disease and other inflammatory conditions. Med Hypotheses. 2004;62(4):493–8. doi: 10.1016/j.mehy.2003.12.016. [DOI] [PubMed] [Google Scholar]
- 332.Dabrowska K, et al. Possible association between phages, Hoc protein, and the immune system. Arch Virol. 2006;151(2):209–15. doi: 10.1007/s00705-005-0641-7. [DOI] [PubMed] [Google Scholar]
- 333.Dabrowska K, et al. Hoc protein regulates the biological effects of T4 phage in mammals. Arch Microbiol. 2007;187(6):489–98. doi: 10.1007/s00203-007-0216-y. [DOI] [PubMed] [Google Scholar]
- 334.Gorski A, et al. Bacteriophages and transplantation tolerance. Transplant Proc. 2006;38(1):331–3. doi: 10.1016/j.transproceed.2005.12.073. [DOI] [PubMed] [Google Scholar]
- 335.Gorski A, et al. New insights into the possible role of bacteriophages in transplantation. Transplant Proc. 2003;35(6):2372–3. doi: 10.1016/s0041-1345(03)00811-x. [DOI] [PubMed] [Google Scholar]
- 336.Gorski A, et al. Bacteriophage translocation. FEMS Immunol Med Microbiol. 2006;46(3):313–9. doi: 10.1111/j.1574-695X.2006.00044.x. [DOI] [PubMed] [Google Scholar]
- 337.Gorski A, Weber-Dabrowska B. The potential role of endogenous bacteriophages in controlling invading pathogens. Cell Mol Life Sci. 2005;62(5):511–9. doi: 10.1007/s00018-004-4403-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 338.Kniotek M, et al. Phages as immunomodulators of antibody production. Immunology. 2004:33–37. [Google Scholar]
- 339.Miedzybrodzki R, et al. Bacteriophage preparation inhibition of reactive oxygen species generation by endotoxin-stimulated polymorphonuclear leukocytes. Virus Res. 2008;131(2):233–42. doi: 10.1016/j.virusres.2007.09.013. [DOI] [PubMed] [Google Scholar]
- 340.Pajtasz-Piasecka E, et al. Bacteriophages support anti-tumor response initiated by DC-based vaccine against murine transplantable colon carcinoma. Immunol Lett. 2008;116(1):24–32. doi: 10.1016/j.imlet.2007.11.004. [DOI] [PubMed] [Google Scholar]
- 341.Przerwa A, et al. Effects of bacteriophages on free radical production and phagocytic functions. Med Microbiol Immunol. 2006;195(3):143–50. doi: 10.1007/s00430-006-0011-4. [DOI] [PubMed] [Google Scholar]
- 342.Gionchetti P, et al. Oral bacteriotherapy as maintenance treatment in patients with chronic pouchitis: a double-blind, placebo-controlled trial. Gastroenterology. 2000;119(2):305–9. doi: 10.1053/gast.2000.9370. [DOI] [PubMed] [Google Scholar]
- 343.Henker J, et al. Probiotic Escherichia coli Nissle 1917 (EcN) for successful remission maintenance of ulcerative colitis in children and adolescents: an open-label pilot study. Z Gastroenterol. 2008;46(9):874–5. doi: 10.1055/s-2008-1027463. [DOI] [PubMed] [Google Scholar]
- 344.Kruis W, et al. Maintaining remission of ulcerative colitis with the probiotic Escherichia coli Nissle 1917 is as effective as with standard mesalazine. Gut. 2004;53(11):1617–23. doi: 10.1136/gut.2003.037747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 345.Mimura T, et al. Once daily high dose probiotic therapy (VSL#3) for maintaining remission in recurrent or refractory pouchitis. Gut. 2004;53(1):108–14. doi: 10.1136/gut.53.1.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 346.Sood A, et al. The probiotic preparation, VSL#3 induces remission in patients with mild-to-moderately active ulcerative colitis. Clin Gastroenterol Hepatol. 2009;7(11):1202–9. 1209 e1. doi: 10.1016/j.cgh.2009.07.016. [DOI] [PubMed] [Google Scholar]
- 347.Vindigni SM, et al. The intestinal microbiome, barrier function, and immune system in inflammatory bowel disease: a tripartite pathophysiological circuit with implications for new therapeutic directions. Therap Adv Gastroenterol. 9(4):606–25. doi: 10.1177/1756283X16644242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 348.Yadav V, et al. Inflammatory bowel disease: exploring gut pathophysiology for novel therapeutic targets. Transl Res. 176:38–68. doi: 10.1016/j.trsl.2016.04.009. [DOI] [PubMed] [Google Scholar]
- 349.Duan F, March JC. Engineered bacterial communication prevents Vibrio cholerae virulence in an infant mouse model. Proc Natl Acad Sci U S A. 107(25):11260–4. doi: 10.1073/pnas.1001294107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 350.Lagenaur LA, et al. Prevention of vaginal SHIV transmission in macaques by a live recombinant Lactobacillus. Mucosal Immunol. 4(6):648–57. doi: 10.1038/mi.2011.30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 351.Steidler L, et al. Treatment of murine colitis by Lactococcus lactis secreting interleukin-10. Science. 2000;289(5483):1352–5. doi: 10.1126/science.289.5483.1352. [DOI] [PubMed] [Google Scholar]
- 352.Braat H, et al. A phase I trial with transgenic bacteria expressing interleukin-10 in Crohn’s disease. Clin Gastroenterol Hepatol. 2006;4(6):754–9. doi: 10.1016/j.cgh.2006.03.028. [DOI] [PubMed] [Google Scholar]
- 353.Vandenbroucke K, et al. Orally administered L. lactis secreting an anti-TNF Nanobody demonstrate efficacy in chronic colitis. Mucosal Immunol. 3(1):49–56. doi: 10.1038/mi.2009.116. [DOI] [PubMed] [Google Scholar]
- 354.Bergstrom A, et al. Establishment of intestinal microbiota during early life: a longitudinal, explorative study of a large cohort of Danish infants. Appl Environ Microbiol. 80(9):2889–900. doi: 10.1128/AEM.00342-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 355.Maurice CF, Haiser HJ, Turnbaugh PJ. Xenobiotics shape the physiology and gene expression of the active human gut microbiome. Cell. 152(1–2):39–50. doi: 10.1016/j.cell.2012.10.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 356.Morgan XC, et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 13(9):R79. doi: 10.1186/gb-2012-13-9-r79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 357.Carmody RN, et al. Diet dominates host genotype in shaping the murine gut microbiota. Cell Host Microbe. 17(1):72–84. doi: 10.1016/j.chom.2014.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 358.Voreades N, Kozil A, Weir TL. Diet and the development of the human intestinal microbiome. Front Microbiol. 5:494. doi: 10.3389/fmicb.2014.00494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 359.Zhang C, et al. Structural resilience of the gut microbiota in adult mice under high-fat dietary perturbations. ISME J. 6(10):1848–57. doi: 10.1038/ismej.2012.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 360.Motta JP, et al. Food-grade bacteria expressing elafin protect against inflammation and restore colon homeostasis. Sci Transl Med. 4(158):158ra144. doi: 10.1126/scitranslmed.3004212. [DOI] [PubMed] [Google Scholar]
- 361.Bermudez-Humaran LG, et al. A novel mucosal vaccine based on live Lactococci expressing E7 antigen and IL-12 induces systemic and mucosal immune responses and protects mice against human papillomavirus type 16-induced tumors. J Immunol. 2005;175(11):7297–302. doi: 10.4049/jimmunol.175.11.7297. [DOI] [PubMed] [Google Scholar]
- 362.Daniel C, et al. Recombinant lactic acid bacteria as mucosal biotherapeutic agents. Trends Biotechnol. 29(10):499–508. doi: 10.1016/j.tibtech.2011.05.002. [DOI] [PubMed] [Google Scholar]
- 363.Takiishi T, et al. Reversal of autoimmune diabetes by restoration of antigen-specific tolerance using genetically modified Lactococcus lactis in mice. J Clin Invest. 122(5):1717–25. doi: 10.1172/JCI60530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 364.Chen Z, et al. Incorporation of therapeutically modified bacteria into gut microbiota inhibits obesity. The Journal of clinical investigation. 124(8):3391–3406. doi: 10.1172/JCI72517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 365.Wang J, et al. Microbial production of value-added nutraceuticals. Current opinion in biotechnology. 37:97–104. doi: 10.1016/j.copbio.2015.11.003. [DOI] [PubMed] [Google Scholar]
- 366.Fossati E, et al. Reconstitution of a 10-gene pathway for synthesis of the plant alkaloid dihydrosanguinarine in Saccharomyces cerevisiae. Nat Commun. 5:3283. doi: 10.1038/ncomms4283. [DOI] [PubMed] [Google Scholar]
- 367.Hawkins KM, Smolke CD. Production of benzylisoquinoline alkaloids in Saccharomyces cerevisiae. Nat Chem Biol. 2008;4(9):564–73. doi: 10.1038/nchembio.105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 368.Minami H, et al. Microbial production of plant benzylisoquinoline alkaloids. Proc Natl Acad Sci U S A. 2008;105(21):7393–8. doi: 10.1073/pnas.0802981105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 369.Nakagawa A, et al. (R,S)-tetrahydropapaveroline production by stepwise fermentation using engineered Escherichia coli. Sci Rep. 4:6695. doi: 10.1038/srep06695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 370.Nakagawa A, et al. A bacterial platform for fermentative production of plant alkaloids. Nat Commun. 2:326. doi: 10.1038/ncomms1327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 371.Zhang C, et al. Metabolic engineering of Escherichia coli BL21 for biosynthesis of heparosan, a bioengineered heparin precursor. Metab Eng. 14(5):521–7. doi: 10.1016/j.ymben.2012.06.005. [DOI] [PubMed] [Google Scholar]
- 372.Jain N, Ramawat KG. Natural Products. Springer; Nutraceuticals and antioxidants in prevention of diseases; pp. 2559–2580. [Google Scholar]
- 373.Li XR, et al. Metabolic engineering of Escherichia coli to produce zeaxanthin. Journal of industrial microbiology & biotechnology. 42(4):627–636. doi: 10.1007/s10295-014-1565-6. [DOI] [PubMed] [Google Scholar]
- 374.Marienhagen J, Bott M. Metabolic engineering of microorganisms for the synthesis of plant natural products. J Biotechnol. 163(2):166–78. doi: 10.1016/j.jbiotec.2012.06.001. [DOI] [PubMed] [Google Scholar]
- 375.Mora-Pale M, et al. Metabolic engineering and in vitro biosynthesis of phytochemicals and non-natural analogues. Plant science. 210:10–24. doi: 10.1016/j.plantsci.2013.05.005. [DOI] [PubMed] [Google Scholar]
- 376.Zhao J, et al. Engineering central metabolic modules of Escherichia coli for improving Î2-carotene production. Metabolic engineering. 17:42–50. doi: 10.1016/j.ymben.2013.02.002. [DOI] [PubMed] [Google Scholar]
- 377.Mikkelsen MD, et al. Microbial production of indolylglucosinolate through engineering of a multi-gene pathway in a versatile yeast expression platform. Metab Eng. 14(2):104–11. doi: 10.1016/j.ymben.2012.01.006. [DOI] [PubMed] [Google Scholar]
- 378.Bhan N, et al. Expanding the chemical space of polyketides through structure-guided mutagenesis of Vitis vinifera stilbene synthase. Biochimie. 115:136–43. doi: 10.1016/j.biochi.2015.05.019. [DOI] [PubMed] [Google Scholar]
- 379.Bhan N, et al. Enzymatic formation of a resorcylic acid by creating a structure-guided single-point mutation in stilbene synthase. Protein Sci. 24(2):167–73. doi: 10.1002/pro.2600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 380.Juhas M. Horizontal gene transfer in human pathogens. Crit Rev Microbiol. 41(1):101–8. doi: 10.3109/1040841X.2013.804031. [DOI] [PubMed] [Google Scholar]
- 381.Soucy SM, Huang J, Gogarten JP. Horizontal gene transfer: building the web of life. Nat Rev Genet. 16(8):472–82. doi: 10.1038/nrg3962. [DOI] [PubMed] [Google Scholar]
- 382.Sola-Oladokun B, Culligan EP, Sleator RD. Engineered Probiotics: Applications and Biological Containment. Annu Rev Food Sci Technol. 8:353–370. doi: 10.1146/annurev-food-030216-030256. [DOI] [PubMed] [Google Scholar]
- 383.Hamady ZZ, et al. Xylan-regulated delivery of human keratinocyte growth factor-2 to the inflamed colon by the human anaerobic commensal bacterium Bacteroides ovatus. Gut. 59(4):461–9. doi: 10.1136/gut.2008.176131. [DOI] [PubMed] [Google Scholar]
- 384.Hamady ZZ, et al. Treatment of colitis with a commensal gut bacterium engineered to secrete human TGF-beta1 under the control of dietary xylan 1. Inflamm Bowel Dis. 17(9):1925–35. doi: 10.1002/ibd.21565. [DOI] [PubMed] [Google Scholar]
- 385.Seedorf H, et al. Bacteria from diverse habitats colonize and compete in the mouse gut. Cell. 159(2):253–66. doi: 10.1016/j.cell.2014.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 386.Donaldson GP, Lee SM, Mazmanian SK. Gut biogeography of the bacterial microbiota. Nat Rev Microbiol. 14(1):20–32. doi: 10.1038/nrmicro3552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 387.Cullen TW, et al. Gut microbiota. Antimicrobial peptide resistance mediates resilience of prominent gut commensals during inflammation. Science. 347(6218):170–5. doi: 10.1126/science.1260580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 388.Lee SM, et al. Bacterial colonization factors control specificity and stability of the gut microbiota. Nature. 501(7467):426–9. doi: 10.1038/nature12447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 389.Sonnenburg ED, et al. Specificity of polysaccharide use in intestinal bacteroides species determines diet-induced microbiota alterations. Cell. 141(7):1241–52. doi: 10.1016/j.cell.2010.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 390.Wu M, et al. Genetic determinants of in vivo fitness and diet responsiveness in multiple human gut Bacteroides. Science. 350(6256):aac5992. doi: 10.1126/science.aac5992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 391.Earle KA, et al. Quantitative Imaging of Gut Microbiota Spatial Organization. Cell Host Microbe. 18(4):478–88. doi: 10.1016/j.chom.2015.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 392.Li H, et al. The outer mucus layer hosts a distinct intestinal microbial niche. Nat Commun. 6:8292. doi: 10.1038/ncomms9292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 393.Nava GM, Friedrichsen HJ, Stappenbeck TS. Spatial organization of intestinal microbiota in the mouse ascending colon. ISME J. 5(4):627–38. doi: 10.1038/ismej.2010.161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 394.Van den Abbeele P, et al. Butyrate-producing Clostridium cluster XIVa species specifically colonize mucins in an in vitro gut model. ISME J. 7(5):949–61. doi: 10.1038/ismej.2012.158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 395.Auchtung JM, Robinson CD, Britton RA. Cultivation of stable, reproducible microbial communities from different fecal donors using minibioreactor arrays (MBRAs) Microbiome. 3:42. doi: 10.1186/s40168-015-0106-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 396.Costello CM, et al. 3-D intestinal scaffolds for evaluating the therapeutic potential of probiotics. Molecular pharmaceutics. 11(7):2030. doi: 10.1021/mp5001422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 397.Lukovac S, et al. Differential modulation by Akkermansia muciniphila and Faecalibacterium prausnitzii of host peripheral lipid metabolism and histone acetylation in mouse gut organoids. MBio. 5(4):e01438–14. doi: 10.1128/mBio.01438-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 398.Gladstone EG, I, Molineux J, Bull JJ. Evolutionary principles and synthetic biology: avoiding a molecular tragedy of the commons with an engineered phage. Journal of biological engineering. 6(1):13. doi: 10.1186/1754-1611-6-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 399.Springman R, et al. Evolutionary stability of a refactored phage genome. ACS synthetic biology. 1(9):425–430. doi: 10.1021/sb300040v. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 400.Borody TJ, Campbell J. Fecal microbiota transplantation: techniques, applications, and issues. Gastroenterol Clin North Am. 41(4):781–803. doi: 10.1016/j.gtc.2012.08.008. [DOI] [PubMed] [Google Scholar]
- 401.van Nood E, et al. Duodenal infusion of donor feces for recurrent Clostridium difficile. N Engl J Med. 368(5):407–15. doi: 10.1056/NEJMoa1205037. [DOI] [PubMed] [Google Scholar]
- 402.Moayyedi P, et al. Fecal Microbiota Transplantation Induces Remission in Patients With Active Ulcerative Colitis in a Randomized Controlled Trial. Gastroenterology. 149(1):102–109 e6. doi: 10.1053/j.gastro.2015.04.001. [DOI] [PubMed] [Google Scholar]
- 403.Cammarota G, et al. The involvement of gut microbiota in inflammatory bowel disease pathogenesis: potential for therapy. Pharmacol Ther. 149:191–212. doi: 10.1016/j.pharmthera.2014.12.006. [DOI] [PubMed] [Google Scholar]
- 404.Pigneur B, Sokol H. Fecal microbiota transplantation in inflammatory bowel disease: the quest for the holy grail. Mucosal Immunol. 9(6):1360–1365. doi: 10.1038/mi.2016.67. [DOI] [PubMed] [Google Scholar]
- 405.Reinisch W. Fecal Microbiota Transplantation in Inflammatory Bowel Disease. Dig Dis. 35(1–2):123–126. doi: 10.1159/000449092. [DOI] [PubMed] [Google Scholar]
- 406.Colman RJ, Rubin DT. Fecal microbiota transplantation as therapy for inflammatory bowel disease: a systematic review and meta-analysis. Journal of Crohn’s and Colitis. 8(12):1569–1581. doi: 10.1016/j.crohns.2014.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 407.Rubin DT. Curbing our enthusiasm for fecal transplantation in ulcerative colitis. Am J Gastroenterol. 108(10):1631–3. doi: 10.1038/ajg.2013.279. [DOI] [PubMed] [Google Scholar]
- 408.Angelberger S, et al. Temporal bacterial community dynamics vary among ulcerative colitis patients after fecal microbiota transplantation. Am J Gastroenterol. 108(10):1620–30. doi: 10.1038/ajg.2013.257. [DOI] [PubMed] [Google Scholar]
- 409.Kump PK, et al. Alteration of intestinal dysbiosis by fecal microbiota transplantation does not induce remission in patients with chronic active ulcerative colitis. Inflamm Bowel Dis. 19(10):2155–65. doi: 10.1097/MIB.0b013e31829ea325. [DOI] [PubMed] [Google Scholar]
- 410.Jiang ZD, et al. Randomised clinical trial: faecal microbiota transplantation for recurrent Clostridum difficile infection - fresh, or frozen, or lyophilised microbiota from a small pool of healthy donors delivered by colonoscopy. Aliment Pharmacol Ther. 45(7):899–908. doi: 10.1111/apt.13969. [DOI] [PubMed] [Google Scholar]
- 411.Cholapranee A, Ananthakrishnan AN. Environmental Hygiene and Risk of Inflammatory Bowel Diseases: A Systematic Review and Meta-analysis. Inflamm Bowel Dis. 22(9):2191–9. doi: 10.1097/MIB.0000000000000852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 412.Gent AE, et al. Inflammatory bowel disease and domestic hygiene in infancy. Lancet. 1994;343(8900):766–7. doi: 10.1016/s0140-6736(94)91841-4. [DOI] [PubMed] [Google Scholar]
- 413.Guarner F. Hygiene, microbial diversity and immune regulation. Curr Opin Gastroenterol. 2007;23(6):667–72. doi: 10.1097/MOG.0b013e3282eeb43b. [DOI] [PubMed] [Google Scholar]
- 414.Koloski NA, Bret L, Radford-Smith G. Hygiene hypothesis in inflammatory bowel disease: a critical review of the literature. World J Gastroenterol. 2008;14(2):165–73. doi: 10.3748/wjg.14.165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 415.Lloyd-Price J, Abu-Ali G, Huttenhower C. The healthy human microbiome. Genome Med. 8(1):51. doi: 10.1186/s13073-016-0307-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 416.Robert S, et al. Oral delivery of glutamic acid decarboxylase (GAD)-65 and IL10 by Lactococcus lactis reverses diabetes in recent-onset NOD mice. Diabetes. 63(8):2876–87. doi: 10.2337/db13-1236. [DOI] [PubMed] [Google Scholar]
- 417.Petrof EO, et al. Stool substitute transplant therapy for the eradication of Clostridium difficile infection: ‘RePOOPulating’ the gut. Microbiome. 1(1):3. doi: 10.1186/2049-2618-1-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 418.Petrof EO, Khoruts A. From stool transplants to next-generation microbiota therapeutics. Gastroenterology. 146(6):1573–82. doi: 10.1053/j.gastro.2014.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 419.Ianiro G, et al. Fecal microbiota transplantation in inflammatory bowel disease: beyond the excitement. Medicine (Baltimore) 93(19):e97. doi: 10.1097/MD.0000000000000097. [DOI] [PMC free article] [PubMed] [Google Scholar]