

Abstract
Skeletal muscle fibers are defined by patterned covariation of key traits that determine contractile and metabolic characteristics. Although the functional properties of most skeletal muscles result from their proportional content of a few conserved muscle fiber types, some, typically craniofacial, muscles exhibit fiber types that appear to lie outside the common phenotypic range. We analyzed gene expression profiles of three putative muscle classes, limb, masticatory, and extraocular muscle (EOM), in adult mice by high-density oligonucleotide arrays. Pairwise comparisons using conservative acceptance criteria identified expression differences in 287 genes between EOM and limb and/or masticatory muscles. Use of significance analysis of microarrays methodology identified up to 400 genes as having an EOM-specific expression pattern. Genes differentially expressed in EOM reflect key aspects of muscle biology, including transcriptional regulation, sarcomeric organization, excitation-contraction coupling, intermediary metabolism, and immune response. These patterned differences in gene expression define EOM as a distinct muscle class and may explain the unique response of these muscles in neuromuscular diseases.
Color has been a simple, yet accurate and durable, means of characterizing skeletal muscle (1). Despite an evolving understanding of cellular and molecular mechanisms that govern contraction speed and metabolic properties, features at the core of fiber typing, the long-standing tripartite red (type I), intermediate (type IIA), and white (IIB) muscle fiber classification scheme (2, 3) remains a central concept in muscle biology. Muscle properties are a direct consequence of fiber type composition, with type I and IIA predominance in red muscles (e.g., soleus) and type IIB predominance in white muscles (e.g., gastrocnemius). The conceptual framework provided by traditional fiber type classifications has proven invaluable for the study of developmental regulatory mechanisms, myofiber structure-function correlations, and muscle response to neuro- or myopathic disease.
Conservation of skeletal myofiber types is a consequence of shared somitic origin, a conserved program of myogenic signal transduction and transcriptional mechanisms, and the trophic influence of only a few, stereotypical motoneuron activity patterns (4, 5). By contrast, some fiber types in specialized craniofacial muscles may lie outside the traditional phenotypic range. The tissue-specific myosin in carnivore muscles of mastication was the basis for the concept of two major skeletal muscle classes, or allotypes—masticatory and limb (6). Extraocular muscle (EOM) was proposed to be a third allotype, also on the basis of myosin expression. Whether there are broader gene expression differences behind muscle allotypes is unknown.
EOM differs from the skeletal muscle stereotype (7, 8). In mammals, only EOM does not fit accepted myofiber classification schemes (7). Adult EOM expresses most known striated muscle myosin isoforms (including EOM-specific and developmental), myofiber myosin expression is heterogeneous, and ≈20% of EOM fibers are multiply innervated. These observations, plus the differential responses of EOM in a wide variety of neuromuscular diseases (8–13), strongly support the idea that it is unique among skeletal muscles.
We hypothesized that the existence of distinct muscle classes should be broadly reflected at the mRNA level. This question was addressed by using high-density oligonucleotide microarrays for expression profiling of murine EOM, jaw, and leg muscles. Here, we report substantial expression differences in gene categories of central importance to skeletal muscle biology and establish the novel molecular signature of EOM.
Materials and Methods
Animals.
Eight-week-old male C57BL/10SnJ mice (The Jackson Laboratory) were killed with CO2, and EOM, masseter, and gastrocnemius/soleus muscles were collected to represent the three putative muscle allotypes. To minimize interanimal variability, muscles from 10 mice were pooled into a single sample/muscle group. A total of 40 mice provided replicate samples of EOM (n = 4), leg (n = 4), and jaw (n = 3) muscle.
Expression Profiling by DNA Microarray.
Total RNA was extracted by using Trizol (GIBCO/BRL). RNA pellets were resuspended at 1 μg RNA/μl diethyl pyrocarbonate-treated water. Total RNA was processed for use on Affymetrix (Santa Clara, CA) murine U74A arrays, according to the manufacturer's protocol. Briefly, 8 μg of total RNA was used in a reverse transcription reaction (SuperScript II; Life Technologies, Rockville, MD) to generate first-strand cDNA. After second-strand synthesis, double-strand cDNA was used in an in vitro transcription (IVT) reaction to generate biotinylated cRNA. After purification and fragmentation, 15 μg of cRNA was used in a 300-μl hybridization mixture containing spiked IVT controls. Approximately 200 μl of mixture was hybridized on chips for 16 h at 45°C. Standard posthybridization wash and double-stain protocols used an Affymetrix GeneChip Fluidics Station 400. Arrays were scanned by using a Hewlett–Packard Gene Array scanner.
Data Analysis Using Affymetrix Software.
DNA microarray data analysis used Affymetrix microarray suite (version 4.0) and mining tool (version 1.2). These generated an increase/no change/decrease difference call and a fold change value for each probe set. Only difference calls consistent across all replicates (4/4 in EOM vs. leg; 3/3 in EOM vs. jaw and jaw vs. leg) were analyzed further; then only average changes ≥ 2-fold were considered significant. At the time of this writing, Affymetrix announced that 2,611 probe sets arrayed on the MG-U74A chip were based on incorrect GenBank sequences. These GenBank sequences were excluded from the analysis, resulting in a net of 9,977 interrogated genes/chip.
Data Analysis by Significance Analysis of Microarray (SAM).
We also used a statistical method adapted specifically for analysis of microarray data, SAM (14). SAM uses pairwise comparisons of gene expression, with each gene's average difference in expression between tissues calculated over the paired differences across matched sets of samples (ti), standardized by the SD of the paired differences. This result represents an observed average expression difference. An expected difference based on the null hypothesis of no difference is estimated and compared with the observed difference (t̄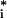 ).
).
We used a bootstrap-based sampling approach (15) in which average paired differences were estimated under the null hypothesis. Basically, each set's paired difference was translated toward a gene mean paired difference of zero. These translated values then were resampled three times with replacement, and standardized average paired difference was estimated. This process was repeated B times (in our case, B = 50), to produce a null sampling distribution. The observed and expected average differences were plotted against one another.
The main difference between our implementation of SAM and that of Tusher et al. (14) is that we did not assume independence across genes when generating the null sampling distributions. Instead, we allowed each gene its own null distribution by using the replicate samples. This approach to sampling has been used previously in microarray analysis by hierarchical statistical modeling (16).
Because thousands of hypothesis tests are needed to scan across all genes, the following analysis was used to compare tissues and account for multiple testing. For some threshold value c > 0, define:
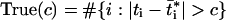 |
1 |
as the number of genes detected to be different for a particular pairwise comparison between two tissues. Also, define average false positive number as:
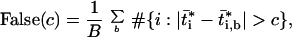 |
2 |
where (t̄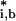 ) is estimated by using all but the bth sample. Finally, the false discovery rate (FDR) for threshold c was defined as:
) is estimated by using all but the bth sample. Finally, the false discovery rate (FDR) for threshold c was defined as:
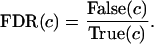 |
3 |
In essence, we aimed to find a threshold value whose FDR is as small as possible, knowing very well that FDR may or may not decrease monotonically as a function of c. Once an optimal threshold value was determined (c*), a SAM plot can be constructed, where observed average differences are plotted against expected ones with a 45° line representing the mean of the null distribution. Error bars that stretch out ± c* can be constructed, and those genes whose observed average difference lies outside these bounds pictorially represent the genes with significantly different average gene expression between tissues of interest.
Results and Discussion
Experimental Strategy.
We determined whether gene expression profiles support the existence of three muscle allotypes—limb, masticatory, and EOM. The percentage of genes on the microarrays reported as present in each muscle group was consistently in the 35–38% range (EOM: 37.4 ± 0.6, leg: 35.1 ± 0.4, jaw: 36.0 ± 1.5; mean ± SD), providing a base of ≈3,500 expressed genes/muscle group for evaluation of putative allotype-specific expression patterns.
We required that muscle group expression differences be robust to validate a distinct allotype. To this end, we used repeated measures and stringent acceptance criteria. By obtaining each set of EOM/leg/jaw muscle samples from 10 mice and restricting muscle group comparisons to within each set, we minimized interanimal variability. In analyses with the Affymetrix algorithm, data were considered valid only when all intermuscle group comparison replicates showed the same qualitative result (increased, decreased, or no change) and the averaged fold difference value was ≥ 2.
Overall Muscle Group Gene Expression Profiles.
Of the genes evaluated, 123 (≈1% on the array and 2.6% appearing as positive) met criteria for significance for both the EOM vs. leg and EOM vs. jaw muscle comparisons. An additional 39 genes (≈0.4% on the array and 0.9% appearing as positive) met criteria for EOM vs. leg only, and 125 genes (≈1% on the array and 2.7% appearing as positive) showed significant differences for EOM vs. jaw only. A prior differential display reverse transcriptase–PCR study identified considerably fewer differences between EOM and leg (17). The substantial structural and functional differences between EOM and other skeletal muscles then are based on gene expression differences that are broader than previously appreciated.
The averaged fold difference distributions for muscle group comparisons are shown in Fig. 1. Similarities in the distributions suggested that EOM was equally different from muscles of somitic (leg) and branchiomeric (jaw) origin. Data did not support jaw as a distinct allotype, as none of the genes surveyed met acceptance criteria in jaw versus leg comparisons. This finding may be a consequence of the similar fiber type composition of mouse masseter and gastrocnemius/soleus muscles, with type IIB predominance in both. Together, data strongly support the notion that EOM is a distinct allotype, but do not validate a separate jaw allotype. We suggest that the muscles of mastication may be an allotype only in carnivores, which alone express the jaw-specific myosin isoform.
Figure 1.

Muscle group-specific differences in gene expression identified by using the Affymetrix algorithm. Frequency distribution histogram illustrating distribution of average fold differences for genes expressed in comparisons between EOM, leg, and jaw muscle groups. Only genes consistently reported as increased, decreased, or no change in all intermuscle group comparisons were plotted. Genes not reaching significance fall in the ± 2 bins.
Unique EOM Expression Profile.
The second question was can one identify a unique subset of genes that collectively provide a molecular signature of normal EOM? This question was addressed by using a SAM analysis, combining jaw and leg tissue to compare against EOM. This approach was justified on the basis of the similarity of jaw and leg expression patterns with the Affymetrix algorithm. In this situation, a paired analysis is not feasible, so average expressions were calculated across sets, and the difference between averages was used to compare EOM to the jaw-leg pool. A null distribution was estimated by first translating the jaw-leg samples to have the same mean, and then translating the EOM and jaw-leg samples to have a mean of zero. Resampling data with replacement, and then reconstructing t statistics, allowed us to work up the SAM analysis. Genes identified as having an expression pattern characteristic of EOM are represented in a SAM plot (see Fig. 3, which is published as supporting information on the PNAS web site, www.pnas.org), and the number of genes and associated FDRs at several SAM threshold values are shown in Table 1. The ability to estimate and manage false positives is the strength of SAM in analysis of microarray data. SAM analysis identified 400 genes with patterns characteristic of EOM at the 0.2 threshold (FDR at this threshold predicts < two genes are false positives).
Table 1.
EOM-specific genes by SAM analysis
| Threshold | No. genes detected | FDR |
|---|---|---|
| 0.2 | 400 | 0.0029 |
| 0.4 | 135 | 0.0013 |
| 0.6 | 55 | 0.001 |
| 0.8 | 25 | 0 |
| 1.0 | 17 | 0 |
SAM analysis compared EOM vs. pooled jaw-leg data, increasing the statistical power of the analysis. Number of genes detected are those identified as EOM-specific at different SAM threshold values, with the FDR at each threshold.
Functional Annotation of Differentially Expressed Genes.
Functional classes were assigned to all genes with significant differences between muscle groups. The general distribution of genes across functional classes includes a broad range of central importance to muscle biology and is shown in Table 2. The expression patterns of several genes of special interest are highlighted in Table 3. The 26 genes with EOM-specific expression patterns as identified by SAM analysis, at thresholds with zero FDRs, are shown in Table 4, which is published as supporting information on the PNAS web site. Some differences in gene expression were previously identified for EOM versus other skeletal muscles (7, 8), whereas most had not been described.
Table 2.
Genes exhibiting significant expression differences for EOM versus leg and/or jaw muscle by gene class
| Gene class | No. of genes by Affymetrix (%) | No. of genes by SAM (%) |
|---|---|---|
| Transcription factor | 17 (5.9) | 5 (3.7) |
| Signal transduction | 45 (15.7) | 10 (7.4) |
| Cell cycle/apoptosis | 15 (5.2) | 5 (3.7) |
| Intermediary metabolism, total | 34 (11.8) | 25 (18.5) |
| Glucose | 9 (3.1) | 11 (8.2) |
| Lipid | 10 (3.5) | 4 (3.0) |
| Other | 15 (5.2) | 10 (7.4) |
| Sarcomeric protein/excitation-contraction coupling | 13 (4.5) | 18 (13.3) |
| Channel/transporter | 23 (8.0) | 11 (8.2) |
| Extracellular matrix/cytoskeleton | 25 (8.7) | 10 (7.4) |
| Immunology/cell defense | 14 (4.9) | 1 (0.7) |
| Other | 30 (10.5) | 20 (14.8) |
| Expressed sequence tag | 71 (24.7) | 30 (22.2) |
For Affymetrix analysis, the number of genes (n = 287) equals those exhibiting ≥ 2-fold change in EOM versus leg or jaw. For SAM analysis, the number of genes (n = 135) represents those differing between EOM and the jaw-leg pool by threshold values ≥ 0.4.
Table 3.
Expression patterns of selected genes for EOM versus leg and jaw muscles
| Accession no. | Gene | Affymetrix average fold difference
|
SAM threshold | |
|---|---|---|---|---|
| EOM vs. leg | EOM vs. jaw | |||
| Transcription factor | ||||
| U70132 | Paired-like homeodomain transcription factor 2 (Pitx2) | 28.5 | 32.9 | 0.2 |
| X56182 | Myogenic factor 5 (Myf5) | ns | 4.1 | — |
| X80338 | Sine oculis-related homeobox 2 (Six2) | −20.7 | −23.3 | 0.2 |
| Sarcomeric protein/excitation-contraction coupling | ||||
| X56518 | Acetylcholinesterase (Ache) | 13.9 | ns | — |
| M17640 | Acetylcholine receptor α-subunit (Chrna1) | 9.9 | 6.5 | 0.4 |
| L47600 | Cardiac troponin T isoform A3b (Tnt2) | 3 | 3.7 | — |
| M14537 | Acetylcholine receptor β-subunit (Chrnb1) | 2.1 | ns | — |
| M15501 | Actin, alpha, cardiac (Actc1) | 2 | 2.1 | 1.0 |
| AJ006306 | Skeletal muscle calcium channel, γ-subunit (Cacng1) | −32.8 | −29.4 | 1.0 |
| AJ001038 | M-protein (Myom2) | −68.7 | −66 | 0.4 |
| U93291 | Calsequestrin (Casq1) | ns | 4.7 | 1.0 |
| AF068244 | Cardiac calsequestrin (Casq) | ns | 2.7 | 0.2 |
| AB010144 | Mitsugumin29 (Mg29) | ns | −2.4 | 0.4 |
| Muscle disease-related protein | ||||
| X53928 | Biglycan (Bgn) | 2.7 | 2.5 | 0.6 |
| X86000 | N-glycan alpha 2,8-sialyltransferase (Siat8d) | 2.2 | ns | — |
| X92523 | Calpain 3 (Capn3) | −2.4 | −2.6 | 0.2 |
| U36579 | Caveolin 3 (Cav3) | ns | −2.4 | 0.4 |
| Immune response | ||||
| *M29009 | Complement factor H-related protein | 12.3 | 5.8 | 0.2 |
| *M29008 | Complement factor H-related protein | ns | 5.4 | — |
| D63679 | Decay accelerating factor 1 (Daf1) | −5 | −7 | — |
| M12660 | Complement component factor h (Cfh) | ns | 4.3 | 0.2 |
| U60473 | CD59 antigen (Cd59a) | ns | 2 | 0.2 |
| L41365 | Decay accelerating factor 2 (Daf2) | ns | −10.7 | — |
Positive values for average fold difference indicate higher gene expression in EOM, negative values indicate lower expression, and ns indicates comparison did not meet criteria for significance. ∗ denotes results obtained with two different array probes. SAM threshold denotes the threshold at which EOM expression value differs from those of both leg and jaw muscles. Dash indicates gene did not meet SAM threshold at ≥ 0.2.
Genes Meeting Acceptance Criteria.
For a list of all genes meeting acceptance criteria by the Affymetrix algorithm (n = 287) and/or SAM (at 0.13% FDR, n = 135), see Table 5, which is published as supporting information on the PNAS web site.
Transcription Factors and Cell Signaling.
Approximately 22% of differentially expressed genes function in transcriptional regulation or cell signaling (Table 2). Although myogenic regulatory factor cascades are highly conserved, identification of craniofacial muscle group-specific Myf5 enhancers suggested that inductive signals might differ among muscle precursor cell populations (18, 19). This mechanism could account for differential development and maintenance of the terminally differentiated muscle phenotypes (20). As muscle group-specific genes, such as the EOM-specific myosin (21, 22) and four uncharacterized EOM expressed sequence tags (17), are identified, 5′ untranslated regions may reveal binding sites for one or more of the transcription factors preferentially expressed in EOM.
Several transcription factors with muscle group differences identified by microarray have been previously linked to myogenesis (e.g., Pitx2, Idb2, Zpf97, Myf5, Six2; Tables 3–5). Pitx2 is expressed by embryonic day 8–12.5 in mouse eye, EOM, and skeletal muscle (23), but its role in myogenesis is unknown. Pitx2 mutations produce the ocular defects in Rieger's syndrome, and knockout mice also exhibit EOM dysgenesis (23). Adult EOM Pitx2 expression was ≈30-fold greater than both limb and jaw and met SAM criteria as part of an EOM-specific expression pattern (Table 3). Beyond an established developmental role, we suggest that Pitx2 may act in maintenance of the adult EOM phenotype. Pitx2 or other differentially expressed transcription factors may be critical determinants of the EOM phenotype, as unlike other myosins, there is a paucity of typical muscle-specific transcription factor sites upstream of EOM-specific myosin (24). By contrast, EOM expression of another gene shared in eye and skeletal muscle development, sine-oculus-related homeobox 2 (Six2), was 20-fold less than jaw or leg (Table 3). Six2, a transcriptional regulator in retina, is expressed in limb muscle precursors and retained in adult tendon (25). Low or absent Six2 in EOM may relate to the neural crest origins of orbital connective tissues (26) and to differences in gene expression profiles of adult tendon fibroblasts.
Muscle-Specific Proteins and the EOM Phenotype.
The absence of significant differences in expression profiles of jaw and hindlimb is consistent with the nearly universal conservation of the three basic skeletal muscle fiber types in mammals. Recent expression profiling of red and white muscles (27) showed only modest differences among the traditional fiber types. There is, however, considerable heterogeneity in striated muscles when the phenotypic range of mammalian cardiac and amphibian/avian slow fiber types is considered. Prior findings, plus our expression profiles, show that EOM combines traits of fast skeletal, cardiac, and phylogenetically primitive slow-tonic fiber types. EOM is characterized by a wide physiologic operating range; fast, but low force, contractions (28); the presence of multiply innervated, nontwitch fiber types (29); and expression of nearly all skeletal muscle myosin isoforms (for review, see ref. 8). Commensurate with its distinct physiology, sarcomeric proteins and excitation-contraction coupling components accounted for 4.5% of the gene profile differences between EOM and other skeletal muscles (Table 2). By SAM, this gene category was more heavily represented, accounting for ≈13% of gene expression differences. Cardiac α-actin, cardiac troponin T isoform a3b, and α-cardiac myosin heavy chain expression were elevated in EOM, as were other heart muscle markers (Tables 3–5). The M-line and the myomesin protein (Myom2) are ubiquitous features of mammalian skeletal and cardiac muscle, stabilizing the alignment of myosin filaments in the sarcomere and anchoring other cellular components to this site, including creatine kinase. However, Myom2 expression was low in adult EOM (Table 3), suggesting the expression of an alternative protein at the EOM M-line.
Key components of skeletal muscle excitation-contraction coupling also were differentially expressed in EOM. Elevated expression of acetylcholine receptor subunits and acetylcholinesterase in EOM (Table 3) may relate to high innervation density. Expression of the γ-subunit of the dihydropyridine receptor (DHPR) was 30-fold less in EOM, despite extensive T-tubules and sarcoplasmic reticulum. The oligomeric DHPR functions as a voltage sensor for excitation-contraction coupling and signals ryanodine receptor calcium channel opening. Deletion of the DHPR γ-subunit alters calcium current and channel inactivation properties (30); low or absent expression of the γ-subunit suggests that EOM uses an alternative isoform with potentially different kinetics. Likewise, mitsugumin 29 (Mg29), a component of skeletal muscle triads, was reduced in EOM (Table 3). Although EOM expresses the known skeletal muscle SERCA1 and SERCA2 isoforms (31, 32), SERCA1B (a neonatal isoform) and alternative isoforms of ryanodine receptor not represented on the Affymetrix array have been reported in fish EOM (33, 34). We did observe a higher level of the sarcoplasmic reticulum calcium binding proteins, calsequestrin and cardiac calsequestrin, in EOM (Table 3). Collectively, these findings show that EOM exhibits diversity at the level of the triad and sarcoplasmic reticulum. This, plus evidence that force-pCa curves in EOM differ from other skeletal muscle (35), suggests that it may diverge from established skeletal muscle excitation-contraction coupling mechanisms.
EOM Gene Profile and Neuromuscular Disease.
Alternative constitutive proteins or adaptive mechanisms unique to the EOM phenotype may be responsible for the known sparing of this muscle group in several neuromuscular diseases. Prior studies described a potentially compensatory increase in EOM utrophin expression in dystrophin deficiency (10, 36), but we observed no constitutive differences between normal adult EOM and other skeletal muscles in expression of dystrophin-glycoprotein complex components. Biglycan (Bgn), an extracellular matrix chondroitin sulfate proteoglycan, binds to the muscle transmembrane dystrophin-glycoprotein complex and is elevated in the dystrophin-deficient mdx mouse (37). Thus, constitutively high Bgn expression in EOM (Table 3) might contribute toward EOM sparing in muscular dystrophy. Two proteins linked to limb-girdle muscular dystrophy, calpain 3 and caveolin-3, were expressed at lower levels in EOM. Finally, EOM expressed higher levels of Siat8d (Table 3), a marker of muscle development or degeneration/regeneration. EOM expression of Siat8d may relate to the retention of the developmental pattern of muscle polysialyated NCAM on some EOM fiber types (38).
Intermediary Metabolism.
The EOM phenotype also was characterized by substantial differences in expression of intermediary metabolism genes, accounting for 11.6% of muscle group differences. A group of genes encoding proteins involved in glucose metabolism, particularly enzymes and regulators associated with glycogenolysis and gluconeogenesis, were expressed at lower levels in EOM (see Table 5). Because glycogen represents a major energy source for skeletal muscle, we more closely examined genes associated with carbohydrate metabolism by mapping expression data onto key metabolic pathways (Fig. 2).
Figure 2.

Muscle group differences in carbohydrate metabolism. Alterations in expression patterns of genes known to participate in glycolysis, gluconeogenesis, glycogen synthesis and catabolism, and glycerol metabolism were mapped onto metabolic pathways. GLUT4, solute carrier family 2 (facilitated glucose transporter), member 4; GLUT1, solute carrier family 2 (facilitated glucose transporter), member 1; G6pt1, glucose-6-phosphatase, transport protein 1; PGM, phosphoglucomutase; Gly Syn, glycogen synthase; Gly Phos, glycogen phosphorylase; Phak1, Phosphorylase kinase alpha 1; Phkb, phosphorylase kinase β; Phkg, phosphorylase kinase gamma; Calm, calmodulin; glu 6-P isomerase, glucose 6-phosphate isomerase; Pkffb, 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase; fru 2,6-P, fructose 2,6-bisphospate; glyceraldehyde 3-P DH, glyceraldehyde 3-phosphate dehydrogenase; glycerol 3-P DH, glycerol 3-phosphate dehydrogenase; TPI, triose phosphate isomerase; PK, pyruvate kinase; LDH (B), lactic dehydrogenase, type B; pyrl Cxl, pyruvate carboxylase; pep CK, phosphoenolpyruvate carboxykinase; fruc 6-Pase, fructose 6-phosphotase.
In EOM, several glycolytic enzymes were expressed at levels higher than other skeletal muscles (Fig. 2; see Table 5). By contrast, key regulators of glycogen metabolism (several phosphorylase kinases) were expressed at low levels in EOM. No muscle group differences in citrate cycle enzymes were detected. Our data suggest that EOM generation of glucose by gluconeogenesis or glycogen breakdown is considerably reduced relative to other skeletal muscles, and that EOM instead relies on glucose obtained directly from an extensive microvascular network in these muscles. These data are supported by findings that glycogen content in EOM is very low and that the typical association between fast-fatigable fibers (type IIB) and high glycogen content is not seen in these muscles (7). Together, the lack of reliance on glycogen by a highly active muscle like EOM stands in contrast to other skeletal muscles.
Immune Response.
A number of genes associated with the immune response were differentially expressed in EOM (4.9% of muscle group expression differences), with particular differences in both classical and alternative complement-mediated immune response pathways (Tables 3 and 4). In the classical pathway, Cd59a, an inhibitor of complement deposition on the cell surface, was expressed at higher levels in EOM (Table 3). By contrast, EOM expression for both of the known decay accelerating factors (Daf1and Daf2), complement regulatory proteins that inhibit the central C3 amplification convertases of the classical complement cascade, was substantially lower than other skeletal muscles (Table 3). EOM expressed higher levels of negative regulators of the alternative pathway of complement activation, Cfh and complement factor H-related protein. Complement system activation by both classical and alternative pathways has been implicated in muscle disease (39–41). Whether this specific expression pattern of complement system regulators relates to the greater sensitivity of EOM to autoimmune disorders like myasthenia gravis and Graves' disease is unknown.
Overview and Conclusions.
The diversity that typifies mammalian skeletal muscle provides for fibers with task-specific contractile properties, while maintaining peak energy efficiency. Skeletal muscle fiber type classification schemes have proven durable because they are based on patterned variations in basic features responsible for contraction and energy metabolism. Most skeletal muscles execute limited functional repertoires and achieve role specificity by simple differences in proportions of the traditional fiber types. DNA microarray has previously identified gene expression differences between red (soleus) and white (quadriceps) skeletal muscles (27). Here, we show that leg and jaw muscles comprised of similar fiber type proportions exhibit no differences in gene expression, an expected finding if fiber types are conserved.
In contrast to traditional skeletal muscles, EOM operates through a wide dynamic range in serving five distinct eye movement classes. Demands placed on EOM then require utilization of a wider range of options for muscle biology. Our study represents an attempt to understand, on a genomic scale, the expression profiles that underlie the broad skeletal muscle classes or allotypes. We show that EOM is a fundamentally distinct skeletal muscle class or allotype, with its own unique gene expression signature. Jaw and leg belong to a separate allotype. Because we have used a conservative approach likely to underestimate muscle diversity, our gene profile is restricted to an estimated 25–35% of the mouse genome, and our data include a moderate percentage of unknown/uncharacterized genes or expressed sequence tags (≈35%), these data represent a conservative estimate of the divergence of EOM from other skeletal musculature.
Supplementary Material
Acknowledgments
We thank Marty Veigl for DNA microarray assistance, Richard Hanson, Robert F. Spencer, and Elizabeth C. Engle for helpful discussions, and Chris Nuss and Pat Porter for help with data analysis. This work was supported by grants from The Research Institute of University Hospitals of Cleveland (to J.D.P.), Research to Prevent Blindness Departmental Grant; Senior Scientific Investigator Award (to J.D.P.), the Knights-Templar Eye Research Foundation (to S.K.), the Muscular Dystrophy Association USA (to J.D.P. and F.H.A.), The Evenor Armington Fund (to J.D.P. and F.H.A.), and National Institutes of Health Grants EY09834 (to J.D.P.), EY12779 (to J.D.P.), EY11998 (to H.J.K.), EY12998 (to F.H.A.) and P30 EY11373. J.D.P. also is supported by the Carl F. Asseff, M.D. Professorship in Ophthalmology.
Abbreviations
- EOM
extraocular muscle
- FDR
false discovery rate
- SAM
significance analysis of microarray
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
References
- 1.Ranvier L. Arch Physiol Norm Pathol. 1874;1:5. [Google Scholar]
- 2.Brooke M H, Kaiser K K. Arch Neurol. 1970;23:369–379. doi: 10.1001/archneur.1970.00480280083010. [DOI] [PubMed] [Google Scholar]
- 3.Peter J B, Barnard R J, Edgerton V R, Gillespie C A, Stempel K E. Biochemistry. 1972;11:2627–2633. doi: 10.1021/bi00764a013. [DOI] [PubMed] [Google Scholar]
- 4.Hauschka S D. In: Myology. Engle A G, Franzini-Armstrong C, editors. Vol. 1. New York: McGraw–Hill; 1994. pp. 3–73. [Google Scholar]
- 5.Burke R E, Levine D N, Zajac F E D. Science. 1971;174:709–712. doi: 10.1126/science.174.4010.709. [DOI] [PubMed] [Google Scholar]
- 6.Hoh J F, Hughes S, Hoy J F. J Muscle Res Cell Motil. 1988;9:59–72. doi: 10.1007/BF01682148. [DOI] [PubMed] [Google Scholar]
- 7.Spencer R F, Porter J D. Rev Oculomot Res. 1988;2:33–79. [PubMed] [Google Scholar]
- 8.Porter J D, Baker R S. Neurology. 1996;46:30–37. doi: 10.1212/wnl.46.1.30. [DOI] [PubMed] [Google Scholar]
- 9.Kaminski H J, al-Hakim M, Leigh R J, Katirji M B, Ruff R L. Ann Neurol. 1992;32:586–588. doi: 10.1002/ana.410320418. [DOI] [PubMed] [Google Scholar]
- 10.Porter J D, Rafael J A, Ragusa R J, Brueckner J K, Trickett J I, Davies K E. J Cell Sci. 1998;111:1801–1811. doi: 10.1242/jcs.111.13.1801. [DOI] [PubMed] [Google Scholar]
- 11.Engle E C, Marondel I, Houtman W A, de Vries B, Loewenstein A, Lazar M, Ward D C, Kucherlapati R, Beggs A H. Am J Hum Genet. 1995;57:1086–1094. [PMC free article] [PubMed] [Google Scholar]
- 12.Heufelder A E, Joba W. Strabismus. 2000;8:101–111. [PubMed] [Google Scholar]
- 13.Porter J D, Merriam A P, Hack A A, Andrade F H, McNally E M. Neuromuscul Disord. 2001;11:197–207. doi: 10.1016/s0960-8966(00)00171-1. [DOI] [PubMed] [Google Scholar]
- 14.Tusher V G, Tibshirani R, Chu G. Proc Natl Acad Sci USA. 2001;98:5116–5121. doi: 10.1073/pnas.091062498. . (First Published April 17, 2001; 10.1073/pnas.091062498) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Efron B, Tibshirani R. An Introduction to the Bootstrap. New York: Chapman and Hall; 1993. [Google Scholar]
- 16.Newton N M, Kendziorski C M, Richmond C S, Blattner F R, Tsui K W. J Comput Biol. 2001;8:37–52. doi: 10.1089/106652701300099074. [DOI] [PubMed] [Google Scholar]
- 17.Niemann C U, Krag T O, Khurana T S. J Neurol Sci. 2000;179:76–84. doi: 10.1016/s0022-510x(00)00384-1. [DOI] [PubMed] [Google Scholar]
- 18.Summerbell D, Ashby P R, Coutelle O, Cox D, Yee S, Rigby P W. Development (Cambridge, UK) 2000;127:3745–3757. doi: 10.1242/dev.127.17.3745. [DOI] [PubMed] [Google Scholar]
- 19.Hadchouel J, Tajbakhsh S, Primig M, Chang T H, Daubas P, Rocancourt D, Buckingham M. Development (Cambridge, UK) 2000;127:4455–4467. doi: 10.1242/dev.127.20.4455. [DOI] [PubMed] [Google Scholar]
- 20.Noden D M, Marcucio R, Borycki A G, Emerson C P., Jr Dev Dyn. 1999;216:96–112. doi: 10.1002/(SICI)1097-0177(199910)216:2<96::AID-DVDY2>3.0.CO;2-6. [DOI] [PubMed] [Google Scholar]
- 21.Wieczorek D F, Periasamy M, Butler-Browne G S, Whalen R G, Nadal-Ginard B. J Cell Biol. 1985;101:618–629. doi: 10.1083/jcb.101.2.618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Winters L M, Briggs M M, Schachat F. Genomics. 1998;54:188–189. doi: 10.1006/geno.1998.5558. [DOI] [PubMed] [Google Scholar]
- 23.Kitamura K, Miura H, Miyagawa-Tomita S, Yanazawa M, Katoh-Fukui Y, Suzuki R, Ohuchi H, Suehiro A, Motegi Y, Nakahara Y, et al. Development (Cambridge, UK) 1999;126:5749–5758. doi: 10.1242/dev.126.24.5749. [DOI] [PubMed] [Google Scholar]
- 24.Briggs M M, Schachat F. J Exp Biol. 2000;203:2485–2494. doi: 10.1242/jeb.203.16.2485. [DOI] [PubMed] [Google Scholar]
- 25.Oliver G, Wehr R, Jenkins N A, Copeland N G, Cheyette B N, Hartenstein V, Zipursky S L, Gruss P. Development (Cambridge, UK) 1995;121:693–705. doi: 10.1242/dev.121.3.693. [DOI] [PubMed] [Google Scholar]
- 26.Noden D M. Am J Anat. 1983;168:257–276. doi: 10.1002/aja.1001680302. [DOI] [PubMed] [Google Scholar]
- 27.Campbell W G, Gordon S E, Carlson C J, Pattison J S, Hamilton M T, Booth F W. Am J Physiol. 2001;280:C763–C768. doi: 10.1152/ajpcell.2001.280.4.C763. [DOI] [PubMed] [Google Scholar]
- 28.Frueh B R, Hayes A, Lynch G S, Williams D A. J Physiol (London) 1994;475:327–336. doi: 10.1113/jphysiol.1994.sp020073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Hess A, Pilar G. J Physiol (London) 1963;169:780–798. doi: 10.1113/jphysiol.1963.sp007296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Freise D, Held B, Wissenbach U, Pfeifer A, Trost C, Himmerkus N, Schweig U, Freichel M, Biel M, Hofmann F, et al. J Biol Chem. 2000;275:14476–14481. doi: 10.1074/jbc.275.19.14476. [DOI] [PubMed] [Google Scholar]
- 31.Tullis A, Block B A. Am J Physiol. 1996;271:R262–R275. doi: 10.1152/ajpregu.1996.271.1.R262. [DOI] [PubMed] [Google Scholar]
- 32.Porter J D, Karathanasis P. Biochem Biophys Res Commun. 1999;257:678–683. doi: 10.1006/bbrc.1999.0536. [DOI] [PubMed] [Google Scholar]
- 33.O'Brien J, Valdivia H H, Block B A. Biophys J. 1995;68:471–482. doi: 10.1016/S0006-3495(95)80208-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Londraville R L, Cramer T D, Franck J P, Tullis A, Block B A. Comp Biochem Physiol B Biochem Mol Biol. 2000;127:223–233. doi: 10.1016/s0305-0491(00)00256-x. [DOI] [PubMed] [Google Scholar]
- 35.Briggs M M, Jacoby J, Davidowitz J, Schachat F H. J Muscle Res Cell Motil. 1988;9:241–247. doi: 10.1007/BF01773894. [DOI] [PubMed] [Google Scholar]
- 36.Matsumura K, Ervasti J M, Ohlendieck K, Kahl S D, Campbell K P. Nature (London) 1992;360:588–591. doi: 10.1038/360588a0. [DOI] [PubMed] [Google Scholar]
- 37.Bowe M A, Mendis D B, Fallon J R. J Cell Biol. 2000;148:801–810. doi: 10.1083/jcb.148.4.801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.McLoon L K, Wirtschafter J D. Invest Ophthalmol Visual Sci. 1996;37:318–327. [PubMed] [Google Scholar]
- 39.Gasque P, Morgan B P, Legoedec J, Chan P, Fontaine M. J Immunol. 1996;156:3402–3411. [PubMed] [Google Scholar]
- 40.Lang T J, Shin M L. J Neuroimmunol. 1993;44:185–192. doi: 10.1016/0165-5728(93)90041-v. [DOI] [PubMed] [Google Scholar]
- 41.Navenot J M, Villanova M, Lucas-Heron B, Malandrini A, Blanchard D, Louboutin J P. Muscle Nerve. 1997;20:92–96. doi: 10.1002/(sici)1097-4598(199701)20:1<92::aid-mus12>3.0.co;2-3. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.