

Abstract
Auxin/indole-3-acetic acid (Aux/IAA) genes encode short-lived transcription factors that are induced as a primary response to the plant growth hormone IAA or auxin. Gain-of-function mutations in Arabidopsis genes, SHY2/IAA3, AXR3/IAA17, and AXR2/IAA7 cause pleiotropic phenotypes consistent with enhanced auxin responses, possibly by increasing Aux/IAA protein stability. Semidominant mutations shy2-1D, shy2-2, axr3-1, and axr2-1 induce ectopic light responses in dark-grown seedlings. Because genetic studies suggest that the shy2-1D and shy2-2 mutations bypass phytochrome requirement for certain aspects of photomorphogenesis, we tested whether SHY2/IAA3 and related Aux/IAA proteins interact directly with phytochrome and whether they are substrates for its protein kinase activity. Here we show that recombinant Aux/IAA proteins from Arabidopsis and pea (Pisum sativum) interact in vitro with recombinant phytochrome A from oat (Avena sativa). We further show that recombinant SHY2/IAA3, AXR3/IAA17, IAA1, IAA9, and Ps-IAA4 are phosphorylated by recombinant oat phytochrome A in vitro. Deletion analysis of Ps-IAA4 indicates that phytochrome A phosphorylation occurs on the N-terminal half of the protein. Metabolic labeling and immunoprecipitation studies with affinity-purified antibodies to IAA3 demonstrate increased in vivo steady-state levels of mutant IAA3 in shy2-2 plants and phosphorylation of the SHY2-2 protein in vivo. Phytochrome-dependent phosphorylation of Aux/IAA proteins is proposed to provide one molecular mechanism for integrating auxin and light signaling in plant development.
Indole-3-acetic acid (IAA or auxin) profoundly affects a plethora of cellular activities and is therefore indispensable for plant growth and development (Thimann, 1977). At the molecular level the hormone rapidly activates transcription of a select set of early genes that are thought to mediate, at least in part, the various effects of auxin (Guilfoyle, 1986; Theologis, 1986). Primary auxin response genes provide bidirectional molecular probes for elucidating auxin action and comprise three major classes known as the SAUR, GH3-like, and Aux/IAA gene families (Abel and Theologis, 1996). Functional promoter analyses of members of each class revealed conserved auxin response elements that led to the identification of their cognate DNA-binding proteins, termed auxin response factors (ARFs). On the other hand, little is known about the function and post-translational regulation of early auxin-induced gene products. However, recent molecular and genetic studies suggest Aux/IAA proteins play a central role in auxin signaling (Guilfoyle et al., 1998; Walker and Estelle, 1998).
Expression of Aux/IAA genes is stringently regulated at multiple levels, which is a hallmark of genes coding for regulatory proteins with transitory roles (Abel and Theologis, 1996). Large families of Aux/IAA genes (>25 members in Arabidopsis; P. Overvoorde and A. Theologis, personal communication) encode short-lived nuclear proteins that homo- and heterodimerize in vitro and in vivo (Abel et al., 1994; Kim et al., 1997). Aux/IAA proteins share four domains of conserved primary structure (Abel et al., 1995; see Fig. 1). Domains II and IV contain functional nuclear localization signals (Abel and Theologis, 1995) and domain III is predicted to adopt a βαα-fold that is similar to the DNA-binding domain of certain oligomeric transcription factors in prokaryotes (Abel et al., 1994). Biophysical and biochemical studies of recombinant peptides demonstrated that domain III is folded and heterodimerizes in vitro (Morgan et al., 1999). It is interesting that the C-terminal regions of most ARF proteins contain Aux/IAA-like domains III and IV, which mediate intra- and interfamily protein-to-protein interactions of ARF and Aux/IAA family members (Kim et al., 1997; Ulmasov et al., 1997a). As shown in a transient expression system, several Aux/IAA proteins repress auxin-dependent activation of auxin response element-mediated reporter genes, likely by interacting with endogenous ARFs (Ulmasov et al., 1997b). Because ARF genes are apparently not induced by auxin (Ulmasov et al., 1999), it is thus conceivable that Aux/IAA proteins are intricately involved in (auto) regulatory circuits of auxin-dependent gene expression.
Figure 1.

Structural and functional domains of Aux/IAA proteins. The primary structure conserved in the core of domain II is given above the designated box. Uppercase letters denote invariant amino acid residues. Positions affected by missense gain-of-function mutations of the indicated Aux/IAA genes are underlined. Bars below the structural domains delineate regions important to various protein activities or modifications. These include functionally identified signals for nuclear localization (NLS; Abel and Theologis, 1995) and protein degradation (Worley et al., 2000), as well as regions shown to mediate homo- and heterodimerization (Kim et al., 1997; Morgan et al., 1999) and to be modified by phytochrome A phosphorylation (this study).
The cloning of genes in Arabidopsis for semidominant mutations shy2 (suppressor of hy2 or short hypocotyl), axr3 (auxin resistant), and axr2 provided compelling genetic evidence for the importance of Aux/IAA genes in plant development. Missense mutations in SHY2/IAA3 (Tian and Reed, 1999), AXR3/IAA17 (Rouse et al., 1998), and AXR2/IAA7 (Nagpal et al., 2000) alter invariant amino acids in domain II (see Fig. 1). The associated pleiotropic phenotypes are consistent with enhanced auxin responses (Timpte et al., 1994; Leyser et al., 1996; Kim et al., 1996, 1998; Tian and Reed, 1999). It is interesting that the gain-of-function mutation in domain II of AXR3/IAA17 resides in a transferable protein degradation signal and stabilizes otherwise labile Aux/IAA::luciferase fusion proteins in heterologous expression systems (Worley et al., 2000). Thus missense mutations in domain II are likely to be hypermorphic because they may increase steady-state levels of Aux/IAA proteins in vivo.
Semidominant shy2-1D (Kim et al., 1996) and shy2-2 (Reed et al., 1998) mutations were independently isolated as extragenic suppressors of the long hypocotyl phenotype of hy2 and hy3(phyB-1) mutants, respectively. The hy2 mutation blocks synthesis of the chromophore, phytochromobilin, and likely causes defects in all holophytochromes, whereas the phyB-1 mutation is a null allele of PHYB, the gene coding for the red/far-red sensing photoreceptor, phytochrome B (Parks and Quail, 1991; Reed et al., 1993). The semidominant SHY2/IAA3 alleles cause photomorphogenic characteristics in dark-grown seedlings such as true leaf development, cotyledon expansion, and reduced hypocotyl growth. A similar de-etiolated phenotype of dark-grown seedlings is observed as a consequence of the axr3 and axr2 mutations (Nagpal et al., 2000). Moreover, shy2-1D plants express light-inducible genes, CAB and PSBA, in darkness (Kim et al., 1998). The collective data suggest that at least some Aux/IAA proteins may normally participate in light responses. When grown in light, young seedlings carrying loss-of-function alleles of SHY2/IAA3 and AXR2/IAA7 consistently have slightly longer hypocotyls than wild-type plants (Tian and Reed, 1999; Nagpal et al., 2000).
Because gain-of-function mutations in SHY2/IAA3 induce ectopic photomorphogenesis in the dark and bypass phytochrome requirement for inhibiting hypocotyl elongation in the light, we tested whether Aux/IAA proteins directly interact with phytochrome and, if so, whether they are substrates for phytochrome protein kinase activity (Yeh and Lagarias, 1998). Here, we demonstrate that SHY2/IAA3, AXR3/IAA17, and other Aux/IAA proteins interact with and are phosphorylated by recombinant phytochrome A in vitro. We further show that the SHY2-2 allele causes higher steady-state levels of the encoded protein and that SHY2-2 is a phosphoprotein in vivo. We propose phytochrome-mediated phosphorylation of Aux/IAA proteins may be a regulatory event at which light and auxin signaling converge.
RESULTS
IAA Proteins Interact with Phytochrome A in Vitro
Genetic characterization of shy2-1D and shy2-2 mutations derived from hy2 and phyB suppressor screens, respectively, suggests interaction of SHY2/IAA3 and PHY genes (Kim et al., 1998; Tian and Reed, 1999). Therefore, we tested whether SHY2/IAA3 and related Aux/IAA proteins such as AXR3/IAA17 from Arabidopsis and Ps-IAA4 from pea (Pisum sativum) interact directly with phytochrome in vitro. To perform in vitro interaction assays we expressed Aux/IAA proteins as (His)6-tagged fusion proteins in Escherichia coli and Strep-tagged phyA from oat (Avena sativa) in the yeast Saccharomyces cerevisiae. Recombinant Aux/IAA proteins were affinity-purified and functional phyA was prepared as described in “Materials and Methods.” In our in vitro assay, interaction of both protein partners is revealed by cosedimentation of recombinant Aux/IAA proteins and Strep-tagged phyA with streptavidin agarose beads after co-incubation and repeated washes of the protein agarose complexes with detergent-containing buffer.
Protein analysis of control incubations by SDS-PAGE demonstrated quantitative binding of Strep-tagged phyA to streptavidin agarose. Recombinant oat phyA cosedimented with the agarose matrix, but was not detectable in the supernatant fraction (Fig. 2A). On the other hand, purified recombinant Ps-IAA4 and IAA17 protein did not bind to streptavidin agarose and remained in the supernatant (Fig. 2, B and C). Purified IAA3 had considerable non-specific affinity to streptavidin agarose, which precluded further experimentation. Is is important to note that when Ps-IAA4 or IAA17 and Strep-tagged phyA were combined in the same binding reactions, Ps-IAA4 and IAA17 cosedimented with immobilized phyA as evidenced by their presence in the agarose bead fraction after repeated washes (Fig. 2, D and E). The data suggest that at least some members of the Aux/IAA protein family have the ability to interact with recombinant oat phyA in vitro.
Figure 2.

Aux/IAA proteins and phytochrome A interact in vitro. Recombinant oat phyA (AsPhyA) and IAA proteins were mixed with streptavidin agarose beads, incubated at room temperature, and treated as described in “Materials and Methods.” Shown are SDS-PAGE resolved proteins of the binding reactions after Coomassie Blue staining for AsPhyA (A) and IAA protein (B and C) control incubations, as well as for the test incubations containing AsPhyA and IAA protein (D and E). S indicates the entire supernatant fraction after the first sedimentation step. P denotes the entire pellet fraction after the third washing step with detergent-containing buffer.
Phytochrome A Phosphorylates IAA Proteins in Vitro
Biochemical data demonstrate that phytochromes are light-regulated, autophosphorylating Ser/Thr protein kinases (Yeh and Lagarias, 1998; Fankhauser et al., 1999). Since we have shown that recombinant oat phyA interacts with Aux/IAA proteins in vitro, we further examined whether recombinant Aux/IAA proteins are substrates for the protein kinase activity of phyA. We performed kinase activity assays with recombinant oat phyA in the Pr and Pfr form and with recombinant Aux/IAA proteins, SHY2/IAA3, AXR3/IAA17, IAA1, IAA9, and Ps-IAA4, as potential protein kinase substrates.
In agreement with previous studies (Yeh and Lagarias, 1998; Fankhauser et al., 1999), recombinant oat phyA exhibited light-regulated autophosphorylation, which was about 2-fold higher for the Pfr form than for the Pr form (Fig. 3). On the other hand, the IAA proteins alone showed little or no autophosphorylation or non-specific binding of [32P]ATP or [32P]orthophosphate. However, phosphorylation of IAA proteins increased at least 10-fold upon addition of phyA, irrespective of its spectral form, indicating that all Aux/IAA proteins tested are substrates for the protein kinase activity of phyA in vitro. The amount of IAA17 and Ps-IAA4 phosphorylation was about three to five times higher than the amount of phyA-dependent phosphorylation of IAA1 or IAA3. Unlike the light-regulated autophosphorylation activity of phyA, phosphorylation of the IAA protein substrates by recombinant phyA was similar for the Pr form and Pfr from, suggesting that Aux/IAA phosphorylation by phyA is not light-dependent under the conditions tested (Fig. 3).
Figure 3.

Phytochrome A phosphorylates Aux/IAA proteins in vitro. Recombinant oat phyA (AsPhyA) in the Pr form or Pfr form was used in kinase assays with recombinant IAA proteins as described in “Materials and Methods.” To detect any autophosphorylation of IAA proteins, assays were also performed without AsPhyA. Shown are autoradiographs of the kinase reactions after SDS-PAGE and protein transfer to membranes (top), and Coomassie Blue staining of the transferred proteins (bottom). Amounts of phyA autophosphorylation and of Aux/IAA phosphorylation by phyA are expressed relative to the reactions containing the Pr form of phyA and are given below the phyA protein bands and below the top panels, respectively.
Phytochrome A Phosphorylates the N-Terminal Region of Ps-IAA4
We tested a small set of Ps-IAA4 deletions as substrates to locate, in a first approximation, phosphorylation sites of phyA on Ps-IAA4. We prepared various truncated, recombinant polypeptides encompassing different conserved domains in Ps-IAA4, including domains I-II, I-III, II-IV, and III-IV. Unlike the protein kinase assays described above, we used phyA pre-assembled in white light for the deletion analysis. Again the data show absence of autophosphorylation for the full-length Ps-IAA4 protein and its truncated forms (Fig. 4). A C-terminal polypeptide of Ps-IAA4, which contains several Ser and Thr residues in conserved domains III and IV, as well as intervening and adjacent sequences, is not phosphorylated to detectable levels by recombinant oat phyA. The C-terminal region provides a negative control and thus support for the specificity of the phyA kinase activity toward Aux/IAA proteins in vitro. Likewise, a recombinant polypeptide corresponding to domain III of IAA3 (Morgan et al., 1999) is not a substrate of oat phyA (data not shown). On the other hand, the N-terminal region-encompassing domain I and II is a very potent substrate that is phosphorylated to high extent (Fig. 4). Consistent with this data, recombinant polypeptides spanning domains I through III and domains II through IV are also phosphorylated (data not shown). Experiments currently in progress will elucidate phosphorylation site(s) of phyA on Aux/IAA proteins.
Figure 4.

Deletion analysis of Ps-IAA4. Kinase assays were conducted in white light with (+) or without (−) recombinant oat phyA (AsPhyA) using full-length (I-IV) Ps-IAA4 (amino acids 1–189) and Ps-IAA4 deletions, domains I-II (amino acids 1–90), and domains III-IV (amino acids 86–189), as protein substrates. Shown are autoradiographs and Coomassie Blue protein staining of the resolved kinase reactions. The arrowhead and bar indicate the positions of oat phyA and of truncated PS-IAA4 polypeptides, respectively.
Detection of SHY2/IAA3 in Vivo
To detect authentic IAA3 and to probe its phosphorylation status in vivo we raised polyclonal antibodies to recombinant IAA3, which were subsequently purified by affinity chromatography as described in “Materials and Methods.” Because Aux/IAA proteins are encoded by a large gene family in Arabidopsis and share four domains of conserved primary structure (Abel et al., 1995), we used a series of shy2 mutant alleles to expedite identification of the IAA3 protein in vivo. Based on the genetic characterization of shy2-2 plants (Tian and Reed, 1999) and on the molecular characterization of mutations in conserved domain II of AXR3/IAA17 (Worley et al., 2000), the shy2-2 gain-of-function mutation is hypothesized to stabilize IAA3, a protein with a predicted molecular mass of 21.5 kD. On the other hand, the loss-of-function alleles, SHY2-22 and SHY2-24, are predicted to encode truncated IAA3 proteins of 19.5 and 7 kD, respectively (Tian and Reed, 1999).
Aux/IAA proteins from pea, Ps-IAA4 and Ps-IAA6, are of extremely low abundance and are difficult to detect by immunoblot analysis (Abel et al., 1994; Oeller and Theologis, 1995). To identify the Arabidopsis IAA3 protein and to analyze its steady-state levels in vivo we used metabolic labeling of wild-type and the various shy2 mutant plants with [35S]Met followed by immunoprecipitation of IAA3 from total labeled proteins. Affinity-purified IAA3 antibodies precipitate three major proteins within the range of molecular mass expected for IAA3 synthesized in vivo of 27, 23, and 19 kD (Fig. 5). The following observations suggest that the 23-kD protein represents endogenous IAA3: (a) Steady-state levels are auxin-inducible in wild-type seedlings (compare lane 3 and lane 4); (b) steady-state levels are appreciably higher in untreated shy2-2 seedlings than in untreated or auxin-treated wild-type plants (compare lane 2 with lane 3 and lane 4); and (c) as expected, the 23-kD protein fails to be immunoprecipitated from shy2-24 and shy2-22 extracts (see lane 5 and lane 6). The specificity of the immunoprecipitation reactions was confirmed by competition with recombinant (His)6-tagged IAA3 protein (Fig. 5, lane 7) and by the absence of the auxin-inducible 23-kD polypeptide in immunoprecipitations with preimmune serum (Fig. 5, lane 1).
Figure 5.

Immunoprecipitations with anti-IAA3 of [35S]Met-labeled proteins synthesized in vivo. Etiolated seedlings of the indicated genotypes were metabolically labeled, treated with auxin, and extracted as described in “Materials and Methods.” Shown is the autoradiograph (8 d of exposure) for resolved proteins after immunoprecipitations of the extracts with affinity-purified antibodies to IAA3 (lanes 2–7) or with preimmune serum (lane 1). For each immunoprecipitation reaction, 2.1 × 108 cpm of trichloroacetic acid-precipitable material were used. Incorporation of [35S]Met into trichloroacetic acid-precipitable material was between 31% to 37% of the radiolabel measured in total tissue extracts. The arrowhead indicates a 23-kD protein identified as endogenous IAA3.
Because the shy2-2 mutation causes increased steady-state levels of the 23-kD protein (Fig. 5, lane 2), we tested whether its concentrations are sufficiently high to be detected by immunoblot analysis. Using 300 μg of total protein extract from 5-d-old dark-grown seedlings, affinity-purified IAA3 antibodies reproducibly recognize a protein of approximately 23 kD in extracts of shy2-2 plants, but not in extracts prepared from shy2-22 and shy2-24 seedlings (Fig. 6). For wild-type extracts the amount of the 23-kD protein was near the detection limit and detection was not consistently achieved. It is interesting that the IAA3 protein accumulates to higher steady-state levels in shy2-2 than in wild-type seedlings. In summary, the data indicate specific detection by metabolic labeling and immunoprecipitation of IAA3 in vivo.
Figure 6.

Detection of IAA3 proteins by immunoblot analysis. Seedlings of the indicated genotypes were grown for 5 d in darkness and 300 μg of total proteins were resolved by SDS-PAGE, transferred to a membrane, and probed with affinity-purified IAA3 antibodies as described in “Materials and Methods.” Detection by enhanced chemiluminescence was for 20 min of exposure time. The arrowhead indicates a 23-kD protein identified as endogenous IAA3.
SHY2-2 Is a Phosphoprotein in Vivo
In a first attempt to examine if the observed in vitro phosphorylation of IAA3 by phytochrome is functionally relevant we tested whether the IAA3 protein is phosphorylated in vivo. To directly demonstrate in vivo phosphorylation, we used metabolic labeling with [32P]orthophosphate followed by immunoprecipitation analysis (Fig. 7). Wild-type, shy2-2, and shy2-24 seedlings were germinated for 5 d in the dark and labeled for 3 h in ambient white light in the presence (wild type, shy2-24) or absence (shy2-2) of auxin. After immunoprecipitation of total protein extracts and resolution of the precipitated protein complexes by SDS-PAGE, a protein of approximately 23 kD was detectable by autoradiography for the shy2-2 extract (Fig. 7, lane 3). As expected, a [32P]labeled protein of 23 kD could not be immunoprecipitated from shy2-24 extracts, lending support for the specificity of the immunoprecipitation observed for shy2-2 extracts (Fig. 7, lane 2). However, repeated attempts failed to demonstrate [32P] labeling of IAA3 in wild-type seedlings treated with (Fig. 7, lane 1) or without (data not shown) auxin, possibly due to the lower steady-state level of IAA3 in wild-type seedlings than of the mutant IAA3 protein in shy2-2 plants. Nonetheless, the data suggest that at least the mutant IAA3 protein, SHY2-2, is phosphorylated in vivo.
Figure 7.
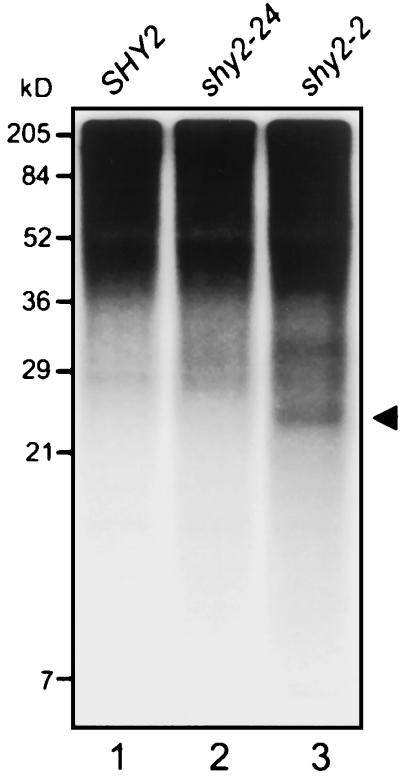
Immunoprecipitations of [32P]orthophosphate-labeled IAA3 proteins synthesized in vivo. Etiolated seedlings of the indicated genotypes were radioactively labeled, treated with auxin (wild-type and shy2-24), and extracted as described in “Materials and Methods.” Shown is the autoradiograph of resolved proteins after immunoprecipitations of the extracts with affinity-purified antibodies to IAA3 (10 d of exposure). The arrowhead indicates a 23-kD protein, likely representing IAA3. The labeled protein bands seen in the region above 36 kD are due to non-specific binding to protein A agarose beads (data not shown).
DISCUSSION
Aux/IAA Proteins Are Substrates of Phytochrome Kinase Activity in Vitro
Molecular and genetic studies suggest that Aux/IAA proteins play a central role in auxin signal transduction by regulating gene expression. However, the structures, biochemical functions, and regulation of Aux/IAA polypeptides are largely unknown. In this communication we provide first experimental evidence for post-translational modification of Aux/IAA gene products. Using various recombinant Aux/IAA proteins from Arabidopsis and pea, which partially reflect the structural diversity of the Aux/IAA superfamily, we have demonstrated in vitro phosphorylation of Aux/IAA polypeptides by recombinant phyA from oat.
Recent biochemical studies have firmly established that eukaryotic phytochromes belong to a growing class of autophosphorylating eukaryotic Ser/Thr kinases with His kinase ancestry (Yeh et al., 1997; Lapko et al., 1999). The phytochrome protein kinase activity is stimulated by red light, which has been demonstrated for recombinant oat phyA (Yeh and Lagarias, 1998). To date, only a few plant proteins have been identified genetically or by yeast two-hybrid screens that are phosphorylated by phytochromes in a light-modulated manner (Reed, 1999). Among the substrates of recombinant oat phyA are the blue light photoreceptors, cry1 and cry2. Although the phosphorylation in vitro is not light regulated, red light stimulates cry1 phosphorylation in vivo (Ahmad et al., 1998). PhyA-mediated in vitro phosphorylation of phytochrome kinase substrate 1, a protein of unknown function, is modulated by light. The degree of phytochrome kinase substrate 1 phosphorylation in vivo is increased upon red light illumination and in plants overexpressing phyB, suggesting that phyB is also a protein kinase (Fankhauser et al., 1999).
Our in vitro data on Aux/IAA protein phosphorylation by recombinant oat phyA are reminiscent of in vitro studies with cry1 and cry2 (Ahmad et al., 1998). Although we have reproduced light-modulated autophosphorylation of phyA, phosphorylation of Aux/IAA proteins by recombinant oat phyA in vitro is apparently not modulated by light in our conditions. However, the prospect arises that biochemical activities of at least some Aux/IAA proteins are post-translationally regulated by phytochrome-mediated phosphorylation or that other factors in the nucleus are necessary for light specificity. Phosphorylation of additional Aux/IAA proteins by phytochromes in vitro is presently under investigation to examine regulation of the modification reaction by light and to assess specificity of Aux/IAA proteins as phytochrome kinase substrates.
IAA3 Is Overexpressed and Phosphorylated in shy2-2 Plants
If in vitro phosphorylation of Aux/IAA proteins by phytochrome reflects biological relevance, it is to be expected that at least some Aux/IAA polypeptides are phosphoproteins in vivo. To investigate the phosphorylation status of select Aux/IAA proteins in vivo we initially focused on SHY2/IAA3 for the following reasons. First, the genetic data pointing to a possibly direct interaction of Aux/IAA proteins and phytochromes is most indicative for shy2/iaa3 mutations (Kim et al., 1996, 1998; Reed et al., 1998; Tian and Reed, 1999). Second, loss-of-function alleles of SHY2/IAA3 are available that may facilitate identification of endogenous IAA3 by immunochemistry.
Metabolic labeling of seedlings with [35S]Met followed by immunoprecipitation with affinity-purified antibodies to IAA3 proved to be the only method for reliably detecting IAA3 in plant extracts. Immunoblot analysis did not consistently recognize the IAA3 polypeptide in up to 300 μg of total wild-type protein. These observations are in agreement with previous reports describing biochemical studies of Aux/IAA proteins from pea, Ps-IAA4, and Ps-IAA6, which were found to be of extremely low abundance in vivo (Abel et al., 1994; Oeller and Theologis, 1995). As expected, the SHY2-2 gain-of-function allele, predicted to stabilize the encoded mutant IAA3 protein (Worley et al., 2000), and the SHY2-22 and SHY2-24 loss-of-function alleles, predicted to encode truncated IAA3 polypeptides (Tian and Reed, 1999), allowed for unambiguous identification of IAA3 in vivo. IAA3 synthesized in planta migrates in SDS-PAGE as a 23-kD protein, which closely matches the predicted molecular mass of 21.5 kD. Despite the complexity of the Aux/IAA family in Arabidopsis, only three major proteins with molecular weights in the range expected for IAA3 were immunoprecipitated with affinity-purified IAA3 antibodies. Again these data are reminiscent of immunoprecipitation studies in pea (Oeller and Theologis, 1995). Although Aux/IAA proteins share four conserved domains and are encoded by large gene families in higher plants (Abel et al., 1995), affinity-purified polyclonal antibodies to Ps-IAA4 and Ps-IAA6 recognize a limited set of proteins in vivo and are specific for the cognate Aux/IAA protein in immunoprecipitation assays (Oeller and Theologis, 1995). In summary, our data suggest specific detection in vivo of the Arabidopsis IAA3 polypeptide.
It is interesting that we found in vivo steady-state levels of [35S]-labeled IAA3 to be significantly higher in shy2-2 than in wild-type plants, exceeding even steady-state concentrations of IAA3 in auxin-treated wild-type seedlings. Moreover, mutant IAA3 protein accumulated in shy2-2 plants to levels high enough to be detected by immunoblot analysis. Our biochemical data provide the first direct evidence for the hypothesis that gain-of-function mutations in domain II of AXR3/IAA17 (Rouse et al., 1998), SHY2/IAA3 (Tian and Reed, 1999), and AXR2/IAA7 (Nagpal et al., 2000) may stabilize the short-lived proteins. Furthermore, our data are in agreement with and support studies demonstrating that conserved domain II of Aux/IAA proteins constitutes a transferable protein degradation signal and that shy2-2-like mutations therein increase the stability of reporter fusion proteins (Worley et al., 2000; J. Callis, personal communication). In collection, these studies provide additional compelling evidence for the importance of protein degradation in auxin responses (Gray and Estelle, 2000). Direct measurement of the metabolic half-lives of wild-type and mutant IAA3 proteins will be necessary to understand the consequences of these semidominant mutations on the stability and activity of IAA3 and of other Aux/IAA proteins.
Detection of endogenous IAA3 by metabolic labeling and immunoprecipitation assays enabled us to directly test whether IAA3 is phosphorylated in vivo. Labeling of shy2-2 seedlings with [32P]orthophosphate followed by immunoprecipitation with anti-IAA3 revealed a phosphoprotein of 23 kD. Failure to detect a [32P]-labeled protein of similar size in immunoprecipitates from shy2-24 extracts suggests specific precipitation of mutant IAA3 from shy2-2 extracts. Although considered to be unlikely, we cannot entirely rule out the possibility that the 23-kD protein represents an Aux/IAA polypeptide that is highly similar to IAA3 and whose expression is enhanced in the shy2-2 background. We were unable to detect [32P] labeling of IAA3 in untreated or auxin-treated wild-type seedlings, which may be explained by lower steady-state levels of IAA3 in wild-type than of mutant IAA3 in shy2-2 plants. In an alternate manner, it is possible that auxin treatment or the shy2-2 mutation (P69S) per se affect turnover of phosphate ester groups in IAA3 and thus [32P] labeling in vivo. Although we have shown that at least the SHY2-2 allele of IAA3 encodes a phosphoprotein in vivo, more powerful analytical tools such as two-dimensional gel electrophoresis will be required to probe the phosphorylation status of immunoprecipitated, [35S]-labeled wild-type IAA3. Different degrees of phosphorylation may be revealed by differences in the electrophoretic mobilities of post-translationally modified IAA3 isoforms.
Functional Implications
Although the in vivo significance of the observed in vitro phosphorylation of Aux/IAA proteins by phyA remains to be further addressed, several observations are consistent with a functional role of Aux/IAA protein phosphorylation in vivo. First, genetic characterization of gain- and loss-of-function shy2 mutations suggests functional interactions between PHY and SHY2/IAA3 gene products (Kim et al., 1996, 1998; Reed et al., 1998; Tian and Reed, 1999). Higher steady-state levels of IAA3 in shy2-2 plants indicate that the gain-of-function mutation is hypermorphic and that IAA3 may normally activate de-etiolation in response to light. Second, since most plant cells express PHYA-PHYE mRNAs at some level during the entire life cycle of Arabidopsis (Somers and Quail, 1995; Goosey et al., 1997), spatio-temporal co-expression of PHY and Aux/IAA genes is most likely determined by tissue-specific expression of Aux/IAA genes (Abel et al., 1995; Wong et al., 1996). Third, phytochromes and Aux/IAA proteins are localized to the cell nucleus in light-grown plants (Abel et al., 1994; Sakamoto and Nagatani, 1996). Although nuclear transport of phytochromes is regulated by light (Kirchner et al., 1999; Yamaguchi et al., 1999; Hisada et al., 2000), nuclear localization of Aux/IAA proteins appears to be constitutive, although the dynamics of the translocation process remains to be studied (Abel et al., 1994; Abel and Theologis, 1995). Thus cellular co-expression and nuclear colocalization of phytochrome and Aux/IAA proteins provide opportunity for phytochrome-mediated phosphorylation of Aux/IAAs proteins to occur in vivo, which may be regulated by light-induced nuclear transport of the photoreceptors.
We are left to speculate about the biochemical consequences of potential phosphorylation of Aux/IAA proteins by phytochrome in vivo. As implicated by genetic and biochemical studies of semidominant shy2-2 and axr3-1 mutations (Kim et al., 1998; Tian and Reed, 1999; Worley et al., 2000), phytochrome-mediated phosphorylation of SHY2/IAA3 would be most consistent with achieving a higher specific activity and/or nuclear concentration of IAA3. As expected, the axr3-1 mutation does not change the ability of IAA17 to localize to the nucleus or to form homo- and heterodimers (Quellet et al., 2000). It is therefore conceivable that phosphorylation of IAA3 may regulate nuclear localization of IAA3, its metabolic stability, its ability to (auto) regulate transcription by interacting with other proteins, including Aux/IAAs and ARFs, its DNA-binding properties (A. Colón-Carmona and S. Abel, unpublished data), or combinations thereof. Various biochemical functions of transcription factors are affected by phosphorylation (Hunter and Karim, 1992), and multiple phosphorylation sites can play distinct and separable roles in regulating transcription factor activity (Komeili and O'Shea, 1999). It is interesting that phyA phosphorylation site(s) map to the N-terminal one-half of Ps-IAA4 (see Fig. 1). The corresponding region of all Aux/IAA proteins, encompassing conserved domains I and II, is Ser/Thr-rich and contains functionally identified, conserved signals for nuclear localization (Abel and Theologis, 1995) and protein degradation (Worley et al., 2000). We are currently mapping phytochrome phosphorylation sites on select Aux/IAA proteins as a prerequisite for studying the function of Aux/IAA phosphorylation in vivo.
Concluding Remarks
An increasing body of evidence suggests intimate interactions between phytochrome and hormone signaling (Neff et al., 2000). As for auxin, in addition to the cloning of SHY2/IAA3 (Tian and Reed, 1999), a number of physiological and genetic studies indicate its involvement in photomorphogenesis. For example, auxin transport is affected in a light-dependent manner (Jensen et al., 1998), which has been implicated in playing a role in the shade avoidance response (Morelli and Ruberti, 2000); loss of NPH4/ARF7 results in a number of differential growth defects, including phytochrome-dependent hypocotyl curvature (Stowe-Evans et al., 1998; Harper et al., 2000) or characterization of hy5, which affects a bZIP transcription factor acting downstream of cryptochrome and phytochrome signaling, strongly suggests a link between light and auxin signal transduction (Oyama et al., 1997). The existence of such a link is further indicated by the phenotypic analyses of the far-red insensitive219 mutation, suggesting that it defines a novel phyA signaling component. It is also interesting that the FIN219 gene is auxin-inducible and highly similar to GH3, a primary auxin response gene (Hsieh et al., 2000). As discussed above, the collective data suggest that SHY2/IAA3 and additional Aux/IAA proteins are bona fide substrates of phytochrome. However, it remains to be seen whether phytochrome-mediated phosphorylation of Aux/IAA proteins provides a mechanism for integrating auxin and light signals in plant development.
MATERIALS AND METHODS
Plant Material and Growth Conditions
Arabidopsis (ecotype Landsberg erecta) was obtained from the Arabidopsis Biological Resource Center at Ohio State University. Seeds of mutant lines shy2-2, shy2-22, and shy2-24, derived from Landsberg erecta (Tian and Reed, 1999), were kindly provided by Dr. J. Reed (University of North Carolina, Chapel Hill). Seeds were surface sterilized and plated on solid medium containing 8 g L−1 phytagar (Gibco-BRL, Gaithersburg, MD), 15 g L−1 Suc, and 2.15 g L−1 (0.5×) Murashige-Skoog salts (Gibco-BRL), pH 5.6. For [32P]orthophosphate-labeling experiments, seedlings were grown on low-phosphate agar medium as previously described (Chen et al., 2000). After stratification at 4°C for 2 to 3 d, seeds were germinated in a controlled environmental chamber at 22°C under illumination with fluorescent and incandescent light at a photon fluence rate of approximately 60 μmol m−2 s−1 for 16 h daily. Prior to growth in darkness, seeds were induced to germinate by treatment with 6 to 12 h of ambient white light.
Plasmid Construction and Purification of Recombinant Proteins
The full-length cDNAs of IAA1, IAA3, IAA9, and IAA17 from Arabidopsis, and Ps-IAA4 from pea (Pisum sativum) were generated by PCR and subcloned into pQE plasmids (Qiagen, Valencia, CA) to produce recombinant, C-terminally (His)6-tagged proteins. Plasmid construction was essentially performed as described by Kim et al. (1997) and Morgan et al. (1999). Expression vectors coding for various C-terminally (His)6-tagged, truncated Ps-IAA4 derivatives, domain I-II (amino acids 1–90), domain I-III (amino acids 1–138), domain II-IV (amino acids 66–189), and domain III-IV (amino acids 86–189), were constructed accordingly. All subcloned cDNA inserts were verified by DNA sequencing. Recombinant proteins were expressed in Escherichia coli M15[pREP] (Qiagen) and purified by affinity chromatography as described (Kim et al., 1997). Expression in the yeast Saccharomyces cerevisiae of Strep-tagged phyA from oat (Avena sativa), purification of the recombinant protein, and chromophore attachment were performed as described (Wu and Lagarias, 1996; Murphy and Lagarias, 1997).
Antibody Production and Purification
Antibodies to recombinant (His)6-tagged IAA3 protein were produce in New Zealand White rabbits (Harlow and Lane, 1988). IAA3 rabbit antiserum and preimmune serum were purified by sequential affinity chromatography (Harlow and Lane, 1988) using total proteins from E. coli M15[pREP] and purified (His)6-tagged IAA3 protein immobilized on CNBr-activated Sepharose-4B (Amersham-Pharmacia Biotech, Uppsala). To obtain antibodies preferentially recognizing variable spacer regions of the IAA3 protein, affinity-purified antibodies were passed over a Sepharose-4B column containing immobilized recombinant (His)6-tagged IAA1 protein.
In Vitro Interaction Assays
Purified recombinant proteins, 2 μg of IAA protein, and 1 μg of Strep-tagged oat phyA were mixed in a total volume of 30 μL of kinase buffer (25 mm Tris-HCl, pH 7.5, 0.2 mm EDTA, and 5 mm MgCl2) with 5 μL (bed volume) of kinase buffer-washed streptavidin agarose beads. Binding reactions were incubated at room temperature for 45 min and mixed every 5 min. Agarose beads were sedimented by centrifugation and washed three times with 500 μL of detergent-containing buffer (50 mm Tris-HCl, pH 7.9, 150 mm NaCl, 1 mm EDTA, and 0.1% [v/v] NP-40). The attached proteins were solubilized by boiling in 1× SDS sample buffer. The proteins of the pellet and initial supernatant (entire fractions) were resolved on 12.5% (w/v) SDS-polyacrylamide gels (Laemmli, 1970). Proteins were transferred to poly(vinylidene difluoride) (PVDF) membranes (Osmonics, Westborough, MA) before visualizing by Coomassie Blue staining.
Phytochrome Kinase Assays
Protein phosphorylation experiments were essentially performed as described (Yeh et al., 1997; Yeh and Lagarias, 1998). The reaction mixtures (total volume of 25 μL) contained kinase buffer (25 mm Tris-HCl, pH 7.5, 0.2 mm EDTA, and 5 mm MgCl2), 0.1 mm ATP, 1 μg of purified recombinant oat phyA in the Pr or Pfr form, and 2 μg of purified recombinant IAA protein (IAA1, IAA3, IAA9, IAA17, PS-IAA4, or truncated forms of PS-IAA4). Kinase reactions were started by adding 10 μCi of [γ-32P]ATP (ICN, 3,000 Ci mmol−1) and were incubated at room temperature for 25 min. Reactions were stopped by the addition of 6 μL of 5× SDS sample buffer. Proteins were resolved on 12.5% (w/v) SDS-polyacrylamide gels and transferred to PVDF membranes for autoradiographic analysis using a Storm 860 PhosphorImager (Molecular Dynamics, Sunnyvale, CA) and for subsequent Coomassie Blue protein staining to verify protein loading.
Metabolic Labeling
For in vivo labeling experiments, 5-d-old dark-grown wild-type and mutant seedlings (1 g fresh weight) were submerged in 7 mL of incubation buffer consisting of 15 mm Suc, 1 mm KCl, 1 mm sodium citrate, and 1 mm 2-(N-morpholino) ethane sulfonic acid, pH 5.6 (Theologis et al., 1985). The seedlings were incubated at room temperature in ambient light under moderate shaking with or without 20 μm IAA in the presence of 1 mCi [35S]Met (Trans label, ICN, >1,000 Ci mmol−1) or 10 mCi [32P]orthophosphate (ICN, 400–800 Ci mmol−1) as follows: for the detection of [35S]Met-labeled IAA3 proteins in vivo, seedlings were incubated with 1 mCi [35S]Met for 4 h. Where indicated, treatment with 20 μm IAA was initiated after 2 h of metabolic labeling. For [32P] labeling experiments, wild-type and mutant seedlings were incubated with 10 mCi [32P]orthophosphate for 3 h.
Protein Extraction and Immunoprecipitation of IAA3
After the indicated incubation regimes, the radioactively labeled seedlings were frozen in liquid nitrogen, ground to a fine powder using a mortar and a pestle, and lyophilized overnight. Proteins were extracted with 1.0 mL of boiling extraction buffer (100 mm Na2HPO4-NaH2PO4, pH 7.0, 140 mm β-mercaptoethanol, and 1% [w/v] SDS) to inactivate endogenous proteases (Vierstra and Quail, 1982). After re-extraction of the plant residues with 0.5 mL of buffer, the combined extract was cleared by centrifugation (20,000g for 20 min at 4°C). The supernatant was diluted 10-fold with buffer A, consisting of 20 mm Na2HPO4-NaH2PO4, pH 7.0, 250 mm NaCl, 5 mm EDTA, 1% (v/v) Triton X-100, and a protease inhibitor cocktail (Complete, Boehringer Mannheim, Basel), or with buffer B (buffer A containing 5 mm EGTA, 50 mm β-glycerophosphate, 50 mm NaF, 10 mm Na3VO4, 10 mm Na4P2O7, 100 μg mL−1 RNase A, and 200 units mL−1 RNase T1) to inactivate phosphatases and to degrade [32P]-labeled RNA (Lanker et al., 1996). The diluted extracts were first precleared with 50 μL of protein A-agarose beads (Santa Cruz Biotechnologies, Santa Cruz, CA) for 1 h. After removal of the agarose beads, affinity-purified IAA3 antibodies were added corresponding to 100 μL of crude antiserum and incubated with agitation for 5 to 6 h at 4°C. Fifty microliters of protein A agarose beads were subsequently added and incubated for 2 h at 4°C. The immunoprecipitate-agarose bead complexes were collected by centrifugation and washed essentially according to Oeller and Theologis (1995). In brief, the agarose beads were sedimented four times in 10 mL of wash buffer (20 mm Tris-HCl, pH 7.5, 10 mm EDTA, 150 mm NaCl, and 1% [v/v] Triton X-100), followed by treatments with 1 mL each of wash buffer containing 0.4 m LiCl and 0.2 m LiCl, wash buffer, and wash buffer containing 1% (w/v) sodium deoxycholate and 0.1% (w/v) SDS. The beads were then washed twice with 20 mm Na2HPO4-NaH2PO4, pH 7.4, 150 mm NaCl, and once with deionized water. Immunocomplexes were eluted into boiling 2× SDS loading buffer and resolved on a 12.5% (w/v) SDS-polyacrylamide gel. Dried gels were analyzed by phosphorimaging (Storm 860, Molecular Dynamics) and by autoradiography (Oeller and Theologis, 1995) using BioMax film (Kodak, Rochester, NY).
Immunoblot Analysis
Proteins were extracted from lyophilized plant tissues as described above. Total proteins were separated by electrophoresis on 12.5% (w/v) SDS-polyacrylamide gels and transferred to PVDF membranes. Membranes were blocked with 10% (w/v) skim milk in 20 mm Tris-HCl, pH 7.6, 150 mm NaCl, and 0.2% (v/v) Tween 20 for 45 min at room temperature. Blots were probed with affinity-purified IAA3 antibodies (2 h) followed by incubation with anti-rabbit IgG conjugated with horse radish peroxidase (Amersham, Buckinghamshire, UK) for 1 h at room temperature. Signals were detected using the enhanced chemiluminescence system (SuperSignal, Pierce, Rockford, IL).
ACKNOWLEDGMENTS
We are indebted to Clark Lagarias for his generous help and inspiring discussions and to Jason Reed for providing various shy2 seed stock. We thank Judy Callis for frequent discussions and Clark Lagarias and John Wiseley for critical reading of the manuscript.
Footnotes
This work was supported by the U.S. Department of Agriculture National Research Initiative Competitive Grants Program (grant no. 9801409 to S.A.).
LITERATURE CITED
- Abel S, Nguyen D, Theologis A. The PS-IAA4/5-like family of early auxin-inducible mRNAs in Arabidopsis thaliana. J Mol Biol. 1995;251:533–549. doi: 10.1006/jmbi.1995.0454. [DOI] [PubMed] [Google Scholar]
- Abel S, Oeller PW, Theologis A. Early auxin-induced genes encode short-lived nuclear proteins. Proc Natl Acad Sci USA. 1994;91:326–330. doi: 10.1073/pnas.91.1.326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abel S, Theologis A. A polymorphic bipartite motif signals nuclear targeting of early auxin-induced proteins related to PS-IAA4 from pea (Pisum sativum) Plant J. 1995;8:87–96. doi: 10.1046/j.1365-313x.1995.08010087.x. [DOI] [PubMed] [Google Scholar]
- Abel S, Theologis A. Early genes and auxin action. Plant Physiol. 1996;111:9–17. doi: 10.1104/pp.111.1.9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ahmad M, Jarillo JA, Smirnova O, Cashmore AR. The CRY1 blue light photoreceptor of Arabidopsis interacts with phytochrome A in vitro. Mol Cell. 1998;1:939–948. doi: 10.1016/s1097-2765(00)80094-5. [DOI] [PubMed] [Google Scholar]
- Chen DL, Delatorre CA, Bakker A, Abel S. Conditional identification of phosphate-starvation-response mutants in Arabidopsis thaliana. Planta. 2000;211:13–22. doi: 10.1007/s004250000271. [DOI] [PubMed] [Google Scholar]
- Fankhauser C, Yeh K-C, Lagarias JC, Zhang H, Elich TD, Chory J. PKS1, a substrate phosphorylated by phytochrome that modulates light signaling in Arabidopsis. Science. 1999;284:1539–1541. doi: 10.1126/science.284.5419.1539. [DOI] [PubMed] [Google Scholar]
- Goosey L, Palecanda L, Sharrock RA. Differential patterns of expressions of the Arabidopsis PHYB, PHYD, and PHYE genes. Plant Physiol. 1997;115:959–969. doi: 10.1104/pp.115.3.959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gray WM, Estelle M. Function of the ubiquitin-proteasome pathway in auxin response. Trends Biochem. 2000;25:133–138. doi: 10.1016/s0968-0004(00)01544-9. [DOI] [PubMed] [Google Scholar]
- Guilfoyle T. Auxin-regulated gene expression in higher plants. CRC Crit Rev Plant Sci. 1986;4:247–276. [Google Scholar]
- Guilfoyle T, Hagen G, Ulmasov T, Murfett J. How does auxin turn on genes? Plant Physiol. 1998;118:341–347. doi: 10.1104/pp.118.2.341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harlow D, Lane E. Antibodies: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1988. [Google Scholar]
- Harper RM, Stowe-Evans EL, Luesse DR, Muto H, Tatematsu K, Watahiki M, Yamamoto K, Liscum E. The NPH4 locus encodes the auxin response factor ARF7, a conditional regulator of differential growth in aerial Arabidopsis tissue. Plant Cell. 2000;12:757–770. doi: 10.1105/tpc.12.5.757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hisada A, Hanzawa H, Weller JL, Nagatani A, Reid JB, Furuya M. Light-induced nuclear translocation of endogenous pea phytochrome A visualized by immunocytochemical procedures. Plant Cell. 2000;12:1063–1078. doi: 10.1105/tpc.12.7.1063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsieh H-L, Okamoto H, Wang M, Ang L-H, Matsui M, Goodman H, Deng XW. FIN219, an auxin-regulated gene, defines a link between phytochrome A and the downstream regulator COP1 in light control of Arabidopsis development. Genes Dev. 2000;14:1958–1970. [PMC free article] [PubMed] [Google Scholar]
- Hunter T, Karim M. The regulation of transcription by phosphorylation. Cell. 1992;70:375–387. doi: 10.1016/0092-8674(92)90162-6. [DOI] [PubMed] [Google Scholar]
- Jensen PJ, Hangarter RP, Estelle M. Auxin transport is required for hypocotyl elongation in light-grown but not dark-grown Arabidopsis. Plant Physiol. 1998;116:975–979. doi: 10.1104/pp.116.2.455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim BC, Soh MS, Kang BJ, Furuya M, Nam HG. Two dominant photomorphogenic mutations of Arabidopsis thaliana identified as suppressor mutations of hy2. Plant J. 1996;9:441–456. doi: 10.1046/j.1365-313x.1996.09040441.x. [DOI] [PubMed] [Google Scholar]
- Kim BC, Soh MS, Kang BJ, Furuya M, Nam HG. Photomorphogenic development of the Arabidopsis shy2-1D mutation and its interaction with phytochromes in darkness. Plant J. 1998;15:61–68. doi: 10.1046/j.1365-313x.1998.00179.x. [DOI] [PubMed] [Google Scholar]
- Kim J, Harter K, Theologis A. Protein-protein interactions among the Aux/IAA proteins. Proc Natl Acad Sci USA. 1997;94:11786–11791. doi: 10.1073/pnas.94.22.11786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirchner S, Kozma-Bognar L, Kim L, Adam E, Harter K, Schäfer E, Nagy F. Light quality dependent nuclear import of the plant photoreceptors phytochrome A and B. Plant Cell. 1999;11:1445–1456. doi: 10.1105/tpc.11.8.1445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Komeili A, O'Shea EK. Roles of phosphorylation sites in regulating activity of the transcription factor Pho4. Science. 1999;284:977–980. doi: 10.1126/science.284.5416.977. [DOI] [PubMed] [Google Scholar]
- Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- Lanker S, Valdivieso H, Wittenberg C. Rapid degradation of the G1 cyclin Cln2 induced by CDK-dependent phosphorylation. Science. 1996;271:1597–1601. doi: 10.1126/science.271.5255.1597. [DOI] [PubMed] [Google Scholar]
- Lapko VN, Jiang XY, Smith DL, Song PS. Mass spectrometric analysis of oat phytochrome A: isoforms and post-translational modifications. Protein Sci. 1999;8:1032–1044. doi: 10.1110/ps.8.5.1032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leyser HMO, Pickett FB, Dharmasiri S, Estelle M. Mutation in the AXR3 gene of Arabidopsis results in altered auxin response including ectopic expression from the SAUR-AC1 promoter. Plant J. 1996;10:403–413. doi: 10.1046/j.1365-313x.1996.10030403.x. [DOI] [PubMed] [Google Scholar]
- Morelli G, Ruberti I. Shade avoidance responses: driving auxin along lateral routes. Plant Physiol. 2000;122:621–626. doi: 10.1104/pp.122.3.621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgan KE, Zarembinski TI, Theologis A, Abel S. Biochemical characterization of recombinant polypeptides corresponding to the predicted βαα fold in Aux/IAA proteins. FEBS Lett. 1999;454:283–287. doi: 10.1016/s0014-5793(99)00819-4. [DOI] [PubMed] [Google Scholar]
- Murphy JT, Lagarias JC. Purification and characterization of recombinant affinity peptide-tagged oat phytochrome A. Photochem Photobiol. 1997;65:750–758. doi: 10.1111/j.1751-1097.1997.tb01920.x. [DOI] [PubMed] [Google Scholar]
- Nagpal P, Walker LM, Young JC, Sonawala A, Timpte C, Estelle M, Reed JW. AXR2 encodes a member of the Aux/IAA protein family. Plant Physiol. 2000;123:563–573. doi: 10.1104/pp.123.2.563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neff MM, Fankhauser C, Chory J. Light: an indicator of time and place. Genes Dev. 2000;14:257–271. [PubMed] [Google Scholar]
- Oeller PW, Theologis A. Induction kinetics of the nuclear proteins encoded by the early indoacetic acid-inducible genes, PS-IAA4/5 and PS-IAA6, of pea (Pisium sativum L.) Plant J. 1995;7:37–48. doi: 10.1046/j.1365-313x.1995.07010037.x. [DOI] [PubMed] [Google Scholar]
- Oyama T, Shimura Y, Okada K. The Arabidopsis HY5 gene encodes a bZIP protein that regulates stimulus-induced development of root and hypocotyl. Genes Dev. 1997;11:2983–2995. doi: 10.1101/gad.11.22.2983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parks BM, Quail PH. Phytochrome-deficient hy1 and hy2 long hypocotyl mutants of Arabidopsis are defective in phytochrome chromophore biosynthesis. Plant Cell. 1991;3:1177–1186. doi: 10.1105/tpc.3.11.1177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quellet F, Overvoorde PJ, Theologis A (2000) IAA17/AXR3: biochemical insight into an auxin mutant phenotype. Presented at the 11th International Conference on Arabidopsis Research, June 24–28, 2000, Madison, WI. Abstract No. 229
- Reed JW. Phytochromes are Pr-ipatetic kinases. Curr Opin Plant Biol. 1999;2:393–397. doi: 10.1016/s1369-5266(99)00011-4. [DOI] [PubMed] [Google Scholar]
- Reed JW, Elumalai RP, Chory J. Suppressors of an Arabidopsis thaliana phyB mutation identify genes that control light signaling and hypocotyl elongation. Genetics. 1998;148:1295–1310. doi: 10.1093/genetics/148.3.1295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reed JW, Nagpal P, Poole DS, Furuya M, Chory J. Mutations in the gene for the red-far-red light receptor phytochrome B alter cell elongation and physiological responses throughout Arabidopsis development. Plant Cell. 1993;5:147–157. doi: 10.1105/tpc.5.2.147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rouse D, Mackay P, Stirnberg P, Estelle M, Leyser O. Changes in auxin response from mutations in an Aux/IAA gene. Science. 1998;279:1371–1373. doi: 10.1126/science.279.5355.1371. [DOI] [PubMed] [Google Scholar]
- Sakamoto K, Nagatani A. Nuclear localization activity of phytochrome B. Plant J. 1996;10:859–868. doi: 10.1046/j.1365-313x.1996.10050859.x. [DOI] [PubMed] [Google Scholar]
- Somers DE, Quail PH. Temporal and spatial expression patterns of PHYA and PHYB genes in Arabidopsis. Plant Physiol. 1995;7:413–427. doi: 10.1046/j.1365-313x.1995.7030413.x. [DOI] [PubMed] [Google Scholar]
- Stowe-Evans EL, Harper RM, Motchoulski AV, Liscum E. NPH4, a conditional modulator of auxin dependent differential growth responses in Arabidopsis. Plant Physiol. 1998;118:1265–1275. doi: 10.1104/pp.118.4.1265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Theologis A. Rapid gene regulation by auxin. Annu Rev Plant Physiol. 1986;37:407–438. [Google Scholar]
- Theologis A, Huynh TV, Davis RW. Rapid induction of specific mRNAs by auxin in pea epicotyl tissue. J Mol Biol. 1985;183:53–68. doi: 10.1016/0022-2836(85)90280-3. [DOI] [PubMed] [Google Scholar]
- Thimann KV. Hormone Action in the Whole Life of Plants. Amherst, MA: University of Massachusetts Press; 1977. [Google Scholar]
- Tian Q, Reed JW. Control of auxin-regulated root development by the Arabidopsis thaliana SHY2/IAA3 gene. Development. 1999;126:711–721. doi: 10.1242/dev.126.4.711. [DOI] [PubMed] [Google Scholar]
- Timpte C, Wilson AK, Estelle M. The axr2-1 mutation of Arabidopsis thaliana is a gain-of-function mutation that disrupts an early step in auxin response. Genetics. 1994;138:1239–1249. doi: 10.1093/genetics/138.4.1239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ulmasov T, Hagen G, Guilfoyle TJ. ARF1, a transcription factor that binds to auxin response elements. Science. 1997a;276:1865–1868. doi: 10.1126/science.276.5320.1865. [DOI] [PubMed] [Google Scholar]
- Ulmasov T, Hagen G, Guilfoyle TJ. Dimerization and DNA binding of auxin responsive factors. Plant J. 1999;19:309–319. doi: 10.1046/j.1365-313x.1999.00538.x. [DOI] [PubMed] [Google Scholar]
- Ulmasov T, Murfett J, Hagen G, Guilfoyle T. Aux/IAA proteins repress expression of reporter genes containing natural and highly active synthetic auxin response elements. Plant Cell. 1997b;9:1963–1971. doi: 10.1105/tpc.9.11.1963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vierstra RD, Quail PH. Native phytochrome: inhibition of proteolysis yields a homogenous monomer of 124 kD from Avena. Proc Natl Acad Sci USA. 1982;79:5272–5276. doi: 10.1073/pnas.79.17.5272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walker L, Estelle M. Molecular mechanisms of auxin action. Curr Opin Plant Biol. 1998;1:434–439. doi: 10.1016/s1369-5266(98)80269-0. [DOI] [PubMed] [Google Scholar]
- Wong L-M, Abel S, Shen N, de la Foata M, Mall Y, Theologis A. Differential activation of the primary auxin response genes, PS-IAA4/5 and PS-IAA6, during early plant development. Plant J. 1996;9:587–599. doi: 10.1046/j.1365-313x.1996.9050587.x. [DOI] [PubMed] [Google Scholar]
- Worley CK, Zenser N, Ramos J, Rouse D, Leyser O, Theologis A, Callis J. Degradation of Aux/IAA proteins is essential for normal auxin signaling. Plant J. 2000;21:553–562. doi: 10.1046/j.1365-313x.2000.00703.x. [DOI] [PubMed] [Google Scholar]
- Wu SH, Lagarias JC. The methylotrophic yeast Pichia pastoris synthesizes a functionally active chromophore precursor of the plant photoreceptor phytochrome. Proc Natl Acad Sci USA. 1996;93:8989–8994. doi: 10.1073/pnas.93.17.8989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaguchi R, Nakamura M, Mochizuki N, Kay SA, Nagatani A. Light-dependent translocation of a phytochrome B-GFP fusion protein to the nucleus in transgenic Arabidopsis. J Cell Biol. 1999;145:437–445. doi: 10.1083/jcb.145.3.437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeh K-C, Lagarias JC. Eukaryotic phytochromes: light-regulated serine/threonine protein kinases with histidine kinase ancestry. Proc Natl Acad Sci USA. 1998;95:13976–13981. doi: 10.1073/pnas.95.23.13976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeh K-C, Wu S-H, Murphy JT, Lagarias JC. A cyanobacterial two-component light sensory system. Science. 1997;227:1505–1508. doi: 10.1126/science.277.5331.1505. [DOI] [PubMed] [Google Scholar]