

Abstract
Iron is an essential nutrient for all living organisms with critical roles in many biological processes. The mammalian host maintains the iron requirements by dietary intake, while the invading pathogenic bacteria compete with the host to obtain those absorbed irons. In order to limit the iron uptake by the bacteria, the human host employs numerous iron binding proteins and withholding defense mechanisms that capture iron from the microbial invaders. To counteract, the bacteria cope with the iron limitation imposed by the host by expressing various iron acquisition systems, allowing them to achieve effective iron homeostasis. The armamentarium used by the human host and invading bacteria, leads to the dilemma of who wins the ultimate war for iron.
Keywords: Iron, Host–pathogen, Iron homeostasis, Host defense, Siderophore, Microbial iron acquisition
Introduction
Iron is the fourth most abundant element on the Earth, approximately 32% of the planet’s mass; it is a crucial micronutrient for all forms of life including humans, other vertebrates, and pathogens. However, some microorganisms such as Streptococcus sanguis [1], Lactobacillus plantarum [2], Borrelia burgdorferi [3] and Treponema pallidum [4] utilize manganese in the place of iron. According to Virulence Factor of Pathogenic Bacteria (VFDB) [5], iron acquisition systems among medically significant bacteria have tremendously evolved in recent years. In this review, a chess game analogy was used to understand the interaction between human host and invading pathogen in the battle for obtaining iron for their survival.
In general, chess is a well-known two-player abstract strategy board game played on a chequered chessboard consisting 64 squares arranged in an 8 × 8 grid. The opposing pieces are designated white and black, lined up on either side initially, which in this scenario represented by the human host (white pieces) and invading bacteria (black pieces) as shown in Fig. 1. The board game consists of sixteen pieces each sides; a king, a queen, two rocks, two knights, two bishops and eight pawns. Overall, the ultimate aim of the game is: (1) to protect the king; (2) not to lose the pieces carelessly as each of them are valuable; and (3) to make sure all the pieces take part in defending the opponent. Using the same strategy as research model, we compare and discuss the ‘army in battle array’ employed by both human host and invading bacteria and their characteristics in biological context. Besides, the aspect of host–pathogen interaction in infectious diseases such as tuberculosis, malaria and HIV were also reviewed in this study.
Fig. 1.

A representative chess pieces during the ‘checkmate’ for iron (Fe). Iron (king) is the key element for both the mammalian host and invading pathogen for survival. The pathogen secretes various iron and/or heme chelator (bishop) such as siderophore (S) and hemophore (H) during iron starvation. Upon invasion, the bacterial cell experiences a phenomenon called ‘nutritional immunity’ imposed by the human immune system (rook), to prevent the access of extracellular iron. Numerous iron withholding components (pawn) such as lactoferrin (Lf), transferrin (Tf), ferritin and heme stores the free available iron. However, in some circumstance, the bacterial chelator escapes these first line defense and captures the iron sources. The human host also employs various iron withholding defense mechanisms (knight) such as hepcidin (Hp) and siderocalin (Sn) that block (mark X) the iron uptake into the pathogen by attaching to ferroportin and siderophore respectively. Finally, successful pathogen utilizes the captured iron sources through the production of ‘stealth siderophore’ and iron acquisition system ensuring its continual survival (the strategy of the chess game is highlighted in the numbered box [1–6])
The Chess Pieces: The ‘Army in Battle Array’
A ‘Precious’ Metal: ‘King’ The Iron in Human Host
The ‘king’ (
 ) resembles the bioavailable iron in the human host. If the bacteria ‘checkmates’ the iron, the host loses the ‘war’ and in this case disease process will be initiated. Iron plays a central role in wide variety of physiological processes, ranging from respiration, tricarboxylic acid cycle (TCA), oxygen transport, ATP generation, cellular growth, functionality of immune cells, gene regulation, and DNA biosynthesis as outlined in Table 1 [6]. On an average, the human adult contains approximately 3–4 g of iron and the largest pool is held within circulating erythrocytes (1800 mg), followed by tissue pool—predominately the liver (1000 mg), reticuloendothelial cells (200–1000 mg), bone marrow (300 mg) and myoglobin (300 mg) [7]. Since the daily dietary intake of iron is very low, the pool continuously turned over largely by iron recycling from senescent erythrocytes, through degradation of heme-containing iron.
) resembles the bioavailable iron in the human host. If the bacteria ‘checkmates’ the iron, the host loses the ‘war’ and in this case disease process will be initiated. Iron plays a central role in wide variety of physiological processes, ranging from respiration, tricarboxylic acid cycle (TCA), oxygen transport, ATP generation, cellular growth, functionality of immune cells, gene regulation, and DNA biosynthesis as outlined in Table 1 [6]. On an average, the human adult contains approximately 3–4 g of iron and the largest pool is held within circulating erythrocytes (1800 mg), followed by tissue pool—predominately the liver (1000 mg), reticuloendothelial cells (200–1000 mg), bone marrow (300 mg) and myoglobin (300 mg) [7]. Since the daily dietary intake of iron is very low, the pool continuously turned over largely by iron recycling from senescent erythrocytes, through degradation of heme-containing iron.
Table 1.
Role of iron in human host
| Functions | Examples |
|---|---|
| Metabolism | Production of red blood cells through hematopoeisis Carrier of oxygen to the tissues from the lungs by red blood cell containing haemoglobin Conversion of blood glucose to energy which allows muscle to work at their optimum Regulation of the cell cycle by activating the cyclin/cyclin-dependent kinase complexes, favouring the progression to the S phase Iron participates as coenzyme in the respiratory chains enzymes (i.e.: cytochromes) and in the formation of heme, which is incorporated in the other heme-containing proteins (i.e.: hemoglobin and myoglobin) Influence the mRNA translational process |
| Immune function | Proliferation and activation of immune cells (i.e.: T-, B- and natural killer cells) and interacts with cell-mediated immune effector pathways and cytokines activities Important role in phagocyte activation and function in killing microorganisms Development of proteins with high affinity to trivalent iron, such as transferrin (Tf) and lactoferrin (Lf) Ferroportin (Fp), an iron efflux protein, strongly influences host response to infection Hepcidin, the allosteric inhibitor of Fp, regulates intracellular iron levels by interacting with, and promoting Fp degradation Ferritin bind to T lymphocytes, suppression of the delayed-type hypersensitivity, and antibody production by B lymphocytes, and impairment of phagocytosis by the granulocytes |
| Enzyme activity | Peroxidases, catalases and certain forms of superoxide dismutase (SOD) that prevent the attack of free reactive oxygen species (ROS) depend on iron Iron-containing enzymes (i.e.: cytochrome P450) include the synthesis of steroid hormones and bile acids, detoxification of foreign substances in the liver, and signal controlling in some neurotransmitters |
| Inflammation and neoplasia | Manifestation of serositris and various hematological disturbances in rheumatoid arthritis, systemic lupus erythematosous, Still’s disease, dermatomyositis, and other collagen diseases are strongly correlated with serum and tissue ferritin levels Ferritin and iron homeostasis are implicated in the pathogenesis of many other disorders, including atherosclerosis, Parkinson’s disease, Alzheimer disease, and restless leg syndrome Synthesis of myelin and severely iron deficient patients exhibit impaired myelin formation, among patients with multiple sclerosis Interferes with cell proliferation, by enhancing c-myc expression crucial for the maintenance of cellular homeostasis |
Iron is absorbed in the proximal duodenum and upper jejunum, whereby ferric iron (Fe3+) is reduced to ferrous iron (Fe2+) by intestinal ferric reductase and duodenal cytochrome B (DcytB) at the apical brush border [7, 8]. The ferrous iron is then transported by divalent metal ion transporter 1 (DMT1) into the enterocytes. Once inside the cells, the iron complexes with the apoferritin and stored in ferritin for cellular usage. Some ferrous iron may exit through the basolateral membrane via the ferroportin (FPN1) transporter. Iron can be subsequently distributed via the bloodstream by loading onto transferrin by either through ferroxidase hephaestin iron oxidation or caeruloplasmin in the macrophages. In short, regulation of intestinal iron absorption serves as the primary homeostatic mechanism, however, iron transportation between sites of utilization and storage constitute some significant internal homeostatic mechanisms.
Rise of the Microbial Invader: ‘Queen’ The Bacteria
The ‘queen’ (
 ) represents the pathogens invading the human host for iron sources. In bacteria, approximately 10−6 to 10−7 molar of iron is required per cell to support their various biological functions as outlined in Table 2 [9]. Various metabolic products such as porphyrins, toxins, vitamins, cytochromes, siderophores, and aromatic compounds secreted depending upon the level of iron concentrations [10]. Many bacteria secure iron within its iron storage proteins to cope with iron demand and protect their cells from free radical attack. Iron storage proteins usually take up the ferrous iron which is later deposited as an oxidized ferric form in its central cavity. Ferritin A (FtnA) accumulates iron during post-exponential growth in the presence of iron-rich condition and utilized stored iron during subsequent growth under iron-limited condition. Bacterioferritin (Bfr) are heme-containing protein whereas DNA-binding proteins (Dps) protect the DNA against the combined action of ferrous and H2O2 in the creation of hydroxyl radicals [11, 12]. Most microorganisms has no known heme storage proteins, however, bacteria that obtain heme exogenously such storage proteins may be presented [13].
) represents the pathogens invading the human host for iron sources. In bacteria, approximately 10−6 to 10−7 molar of iron is required per cell to support their various biological functions as outlined in Table 2 [9]. Various metabolic products such as porphyrins, toxins, vitamins, cytochromes, siderophores, and aromatic compounds secreted depending upon the level of iron concentrations [10]. Many bacteria secure iron within its iron storage proteins to cope with iron demand and protect their cells from free radical attack. Iron storage proteins usually take up the ferrous iron which is later deposited as an oxidized ferric form in its central cavity. Ferritin A (FtnA) accumulates iron during post-exponential growth in the presence of iron-rich condition and utilized stored iron during subsequent growth under iron-limited condition. Bacterioferritin (Bfr) are heme-containing protein whereas DNA-binding proteins (Dps) protect the DNA against the combined action of ferrous and H2O2 in the creation of hydroxyl radicals [11, 12]. Most microorganisms has no known heme storage proteins, however, bacteria that obtain heme exogenously such storage proteins may be presented [13].
Table 2.
Role of iron in bacteria cells
| Functions | Examples |
|---|---|
| Cellular composition |
Mycobacterium smegmatis shows decreased DNA and RNA levels under iron-limited conditions Escherichia coli needed iron for the synthesis of the deoxyribotides essential for DNA biosynthesis Bacillus subtilis iron depletion blocks DNA biosynthesis |
| Metabolism | Production of oxidative metabolism enzymes (i.e.: aconitase) of the tricarboxylic acid (TCA) cycle Cytochromes and non-haem iron electron carriers of the electron transport chain (ETC) Prokaryotic nitrogenase enzyme complex consists of an iron-molybdenum protein and an iron-protein, reduce dinitrogen to ammonia compounds 3-deoxy-D-arabino-heptulosonate-7-phosphate (DAHP) synthase of Escherichia coli that is involved in the aromatic pathway contains one atom of iron |
| Metabolic products | Many bacteria produce siderophores (i.e.: hydroxamate- and phenolates type) under iron-limited conditions to solubilize and transport iron for survival Aquatic bacteria contain iron in the form of magnetite (Fe3O4) that confers on them magnetotactic features |
| Enzyme activity | Peroxidases, catalases and certain forms of superoxide dismutase (SOD) that prevent the attack of free radical oxygen species (ROS) depend on iron Glutamate synthase responsible for formation of glutamate from a-ketoglutarate requires iron for it to catalyse the reaction |
Five strategies had been outlined on iron homeostasis in bacteria which include: (1) expression of high-affinity iron transport system allowing iron to be scavenged, in different forms, from different conditions; (2) deposition of intracellular iron stores that provide source of iron upon external iron depletion; (3) employment of redox stress resistance systems such as reactive oxygen species (ROS); (4) control of iron consumption by down-regulating the expression of iron-containing proteins, through regulator system such as ferric uptake regulator (Fur) under iron-limited condition, and (5) an over-arching iron-responsive regulatory system that co-ordinates the expression of above iron homeostatic machinery according to iron availability [11].
Stopping Bacteria From Stealing the Iron: ‘Rook’ The Host Immune System
The ‘rook’ (
 ) represents the ‘castle’s wall’ that acts as protecting or immune barrier from the microbial invader. During the bacterial invasion, the vertebrate host reduces the intestinal iron assimilation and sequesters the circulating iron into storage proteins, ensuring the pathogen encounter a period of iron starvation in a process called ‘nutritional immunity’ [14]. The human hosts obtain access to iron largely by nutritional intake, thus malnutrition contributes to infectious diseases in a susceptible individual, as micronutrients are mainly essential for the development of defense system. However, excess intake of nutrition and associated predisposing metabolic disorders may promote infective stage, as these nutrients are also required by the pathogen for replication [15]. Thus, in context of immunity iron may be regarded as ‘double-edged sword’ as both iron deficiency and iron overload could potentially confer microbial infections in the human host.
) represents the ‘castle’s wall’ that acts as protecting or immune barrier from the microbial invader. During the bacterial invasion, the vertebrate host reduces the intestinal iron assimilation and sequesters the circulating iron into storage proteins, ensuring the pathogen encounter a period of iron starvation in a process called ‘nutritional immunity’ [14]. The human hosts obtain access to iron largely by nutritional intake, thus malnutrition contributes to infectious diseases in a susceptible individual, as micronutrients are mainly essential for the development of defense system. However, excess intake of nutrition and associated predisposing metabolic disorders may promote infective stage, as these nutrients are also required by the pathogen for replication [15]. Thus, in context of immunity iron may be regarded as ‘double-edged sword’ as both iron deficiency and iron overload could potentially confer microbial infections in the human host.
Many underlying diseases may contribute to increasing of iron storage in host systems. The primary cause of iron overload is contributed by hereditary hemochromatosis, whereby too much of iron is accumulated causing damage to liver, heart, pancreas, and joints [16]. Numerous bacterial infections are associated with hemochromatosis which includes Escherichia coli, Listeria monocytogenes, Plesiomonas shigelloides, Vibrio vulnificus, Vibrio cholera, Yersinia pseudotuberculosis and Yersinia enterocolitica [17]. On the other hand, African iron overload (also known as Bantu siderosis) is a type of dietary iron overload caused by high levels of iron in food or drinks. A study reported, the traditional beer prepared at home from local grains in non-galvanized steel containers has a high ferrous iron level which contributes toward significant Mycobacterium tuberculosis pulmonary infection [18].
Therapeutically, iron supplementation, either oral or parenteral route are used largely for treatment of iron deficiency anemia. It was elucidated that, IV iron administration may exacerbate existing infections and this is largely dependent on larger dose compared to the length of therapy [19]. In-vitro experiments suggested that increased transferrin saturation (TSAT) exceeds a critical threshold (normal range for adult 20–50%) may result in polymorphonuclear neutrophils (PMNs) dysfunction and increased bacterial growth such as E. coli, Klebsiella spp., Pseudomonas spp., and Salmonella spp [20]. Blood transfusion for an anemic patient admitted to the medical-surgical-trauma intensive care unit (ICU) was found to be associated with nosocomial infections such as pneumonia, bacteremia, sepsis, and cystitis [21]. In occupational activities, such as welding, cutting of metal, iron ore mining, fettling and metal-related working may result in exposure to iron-containing dust. It was noted in Welder’s siderosis, increased hazard applies to pneumococcal infections may extend to other infectious agents such as Legionella, Mycoplasma and Haemophilus influenzae [22].
First Line Army: ‘Pawn’ the Human Iron Withholding Components
The ‘pawns’ (
 ) are the only pieces to stand in the front row and serve as the primary line of defense in the human host. The mammalian hosts employ iron constitutive components termed ‘siderophilins’ such as transferrin (Tf) and lactoferrin (Lf) [23, 24]. Tf is an extracellular iron-binding glycoprotein that is present in various biological fluids, and responsible for transporting iron from sites of absorption (intestinal mucosal cells), sites of utilization (immature erythroid cells), sites of storage (liver) and sites of hemoglobin degradation (macrophages). [25, 26]. Structurally, Tf possess a molecular weight of 76,000–81,000 kDa, appear prolate ellipsoidal in shape, consist of two similar but not identical binding sites capable of binding reversibly with two atoms of ferric iron in the presence of bicarbonate anion [27].
) are the only pieces to stand in the front row and serve as the primary line of defense in the human host. The mammalian hosts employ iron constitutive components termed ‘siderophilins’ such as transferrin (Tf) and lactoferrin (Lf) [23, 24]. Tf is an extracellular iron-binding glycoprotein that is present in various biological fluids, and responsible for transporting iron from sites of absorption (intestinal mucosal cells), sites of utilization (immature erythroid cells), sites of storage (liver) and sites of hemoglobin degradation (macrophages). [25, 26]. Structurally, Tf possess a molecular weight of 76,000–81,000 kDa, appear prolate ellipsoidal in shape, consist of two similar but not identical binding sites capable of binding reversibly with two atoms of ferric iron in the presence of bicarbonate anion [27].
On the other hand, Lf is an iron-binding monomeric glycoprotein found in secretions of lacrimal, mammary glands (milk), respiratory, gastrointestinal and genital tracts. Lf is a multifunctional protein responsible for many cellular processes such as iron absorption, anti-inflammatory and immunomodulatory [28]. In human adult, synthesis of Lf takes place in glandular epithelial cells followed by secreting into mucosal fluids that soak the cellular surfaces [29, 30]. Apolactoferrin is synthesized and stored within secondary granules of polymorphonuclear neutrophils (PMNs) are released upon stimulation during inflammation or infection to scavenge the iron from septic sites [31].
Ferritin (Fn) remains as the largest iron storage protein in the host cells, and capable of storing iron in bioavailable and non-toxic forms [32]. Structurally, a typical 24-mer ferritin forms a hollow, folded in a 4-helical bundle, spherical in shape and filled up to 4000–4500 Fe atoms as an inorganic complex core [33]. Mammalian ferritin is composed of two subunits, H (heavy) and L (light) subunits which varies according to organ and iron requirements. H subunits are found in the heart and brain which possesses anti-oxidant activity, while L subunits are common in iron storage organs such as spleen and liver [34–37]. Apart from that, in the vertebrate host, the most abundant source of iron is in the form of heme (Hm) as a cofactor of hemoproteins [38]. The total body intracellular iron pool (4 g) consists of Hm that is bound to hemoglobin (Hb) (2.67 g), myoglobin (0.2 g), some Hm enzymes (0.08 g), insoluble ferric irons that are stored in ferritin (1 g) and a poorly defined labile iron compartment (0.07 g) [39]. Distinctly, a unit of packed red blood cells (RBCs) contains approximately 200 mg of Fe, as these component act as a nutrient source for microbial survival [40].
Attackers Never Stop: ‘Bishops’ The Bacterial Iron Scavenger
The bishop (
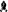 ) represents the iron chelator secreted by bacteria under iron starvation. Successful pathogens have evolved numerous iron acquisition mechanisms to scavenge the iron from their potential hosts, such as siderophore and hemophore systems [41]. Particularly, siderophores are secreted by bacteria to chelate environmental iron and deliver it for pathogen uptake [42, 43]. Typically, microbial siderophores are classified into different classes depending upon moieties of chemical nature. These include catecholate type such as enterobactin produced by enteric bacteria and vibriobactin by V. chloreae, phenolate type such as yersiniabactin by Yersinia spp. or pyochelin by Pseudomonas aeruginosa, hydroxymate type include alcaligin secreted by Bordetella spp. and carboxylate type produced by Staphylococcus spp. New chemistry of siderophores has emerged recently, which led to a more complex classification of integrated chemical features, resulting in “mixed-type” for example phenolate-hydroxamate (mycobactin T), citrate-hydroxamate (aerobactin) and citrate-catecholate (petrobactin) [43]. Interestingly, some bacterial species could evade the defense strategies, by producing or acquiring ‘stealth siderophores’. This strategy allows the siderophore petrobactin produced by Bacillus anthracis to remain in the bloodstream and scavenge available iron for its growth [44]. Some bacteria such as P. aeruginosa able to utilize different siderophores from other microorganisms (xenosiderophore) through ‘siderophore piracy’ [45]. On the other hand, hemophores are family of highly specialized extracellular proteins that are capable of capturing iron-containing Hm from various sources [46]. Hemophores extract Hm from hemoglobin (Hb), hemoglobin–haptoglobin (Hb–Hpt) and heme–hemopexin (Hm–Hpx) complexes.
) represents the iron chelator secreted by bacteria under iron starvation. Successful pathogens have evolved numerous iron acquisition mechanisms to scavenge the iron from their potential hosts, such as siderophore and hemophore systems [41]. Particularly, siderophores are secreted by bacteria to chelate environmental iron and deliver it for pathogen uptake [42, 43]. Typically, microbial siderophores are classified into different classes depending upon moieties of chemical nature. These include catecholate type such as enterobactin produced by enteric bacteria and vibriobactin by V. chloreae, phenolate type such as yersiniabactin by Yersinia spp. or pyochelin by Pseudomonas aeruginosa, hydroxymate type include alcaligin secreted by Bordetella spp. and carboxylate type produced by Staphylococcus spp. New chemistry of siderophores has emerged recently, which led to a more complex classification of integrated chemical features, resulting in “mixed-type” for example phenolate-hydroxamate (mycobactin T), citrate-hydroxamate (aerobactin) and citrate-catecholate (petrobactin) [43]. Interestingly, some bacterial species could evade the defense strategies, by producing or acquiring ‘stealth siderophores’. This strategy allows the siderophore petrobactin produced by Bacillus anthracis to remain in the bloodstream and scavenge available iron for its growth [44]. Some bacteria such as P. aeruginosa able to utilize different siderophores from other microorganisms (xenosiderophore) through ‘siderophore piracy’ [45]. On the other hand, hemophores are family of highly specialized extracellular proteins that are capable of capturing iron-containing Hm from various sources [46]. Hemophores extract Hm from hemoglobin (Hb), hemoglobin–haptoglobin (Hb–Hpt) and heme–hemopexin (Hm–Hpx) complexes.
Furthermore, iron uptake from Tf is mediated by two transferrin-binding proteins (TbpA and TbpB) in bacteria [47, 48], particularly in pathogenic Neisseria such as Neisseria meningitidis and Neiserria gonorrhoeae [49, 50] as well as H. influenzae [51, 52]. Certain bacteria such as H. influenzae and Actinobacillus pleuropneumonia are capable of using transferrin- or lactoferrin-bound iron directly without secreting siderophore [53, 54]. The two lactoferrin-binding proteins (Lbp) of N. meningitidis (LbpA and LbpB) are largely regulated by the presence of iron and closely resemble the meningococcal transferrin-binding protein complexes which display high sensitivity for human lactoferrin [55–57].
Second Line Army: ‘Knight’ The Human Iron Withholding Defense
The knight (
 ) piece depicts second line defense in the human host with complex ‘armour, weapon, and the war horse’ strategies. Multiple innate immune mechanisms restrict the iron availability to invading microorganisms in response to infection [58]. Siderocalin (Scn) was discovered in uterine secretions, serum, and epithelial cells in response to infection and inflammation. Scn is released through degranulation of neutrophils, resulting in capturing the siderophores such as enterobactin secreted by E. coli under iron starvation [14]. Besides that, natural resistance-associated macrophage protein-1 (Nramp1) found primarily in macrophages and polymorphonuclear neutrophils (PMNs), restricts microbial access to iron within the phagosome of phagocytes [59]. The function of Nramp1 was originally noted when the inbred mice strains were resistant to infection caused by Salmonella enterica serovar Typhimurium, Mycobacterium bovis and Leishmania donovani [60, 61].
) piece depicts second line defense in the human host with complex ‘armour, weapon, and the war horse’ strategies. Multiple innate immune mechanisms restrict the iron availability to invading microorganisms in response to infection [58]. Siderocalin (Scn) was discovered in uterine secretions, serum, and epithelial cells in response to infection and inflammation. Scn is released through degranulation of neutrophils, resulting in capturing the siderophores such as enterobactin secreted by E. coli under iron starvation [14]. Besides that, natural resistance-associated macrophage protein-1 (Nramp1) found primarily in macrophages and polymorphonuclear neutrophils (PMNs), restricts microbial access to iron within the phagosome of phagocytes [59]. The function of Nramp1 was originally noted when the inbred mice strains were resistant to infection caused by Salmonella enterica serovar Typhimurium, Mycobacterium bovis and Leishmania donovani [60, 61].
A liver-associated hormone known as hepcidin, was discovered to regulate iron homeostasis through three main sources of iron which include dietary absorption from enterocytes, the release of recycled iron from macrophages as well as the release of stored iron from hepatocytes and Kupffer cells. Functionally, hepcidin primarily inhibits the absorption of iron by binding to the receptor called ferroportin. Ferroportin is a transmembrane protein expressed in the mammalian host, such as in duodenal enterocytes, splenic macrophages, hepatocytes, lung, renal tubules and erythrocyte precursors in bone marrow [62, 63]. Hepcidin expression is down-regulated during hypoxia, anaemia or when there is increased demand for erythropoiesis, while largely up-regulated when there is increased iron storage in the body or during infection or inflammation process mediated by the release of interleukin 6 (IL-6) [64, 65]. When hepcidin binds to ferroportin, the complex is internalized through the clathrin-coated pits. Once internalized, phosphates are removed and ferroportin is tagged with ubiquitin, that signals the protein-transport machinery and subsequently get degraded by the proteasome, resulting in diminished cellular iron transport to plasma.
Iron and Microbial Infections
During clinical infections, iron is freely available from three main sources such as iron overload due to altered metabolic diseases, haemolysis of erythrocytes and normal bodily fluid from an altered homeostatic condition [66]. In view of the increasing worldwide epidemic of tuberculosis (TB), malaria and HIV/AIDS, it may be crucial to decipher the complete machinery employed by both human and microbial invader, in the struggle for iron in these infectious diseases. A comparative representation of the pathogenesis of TB, malaria, and HIV/AIDS in relation to iron withholding defense mechanisms is illustrated in Fig. 2.
Fig. 2.

A comparison schematic representation on the pathogenesis of tuberculosis, malaria, and HIV/AIDS in relation to iron withholding defense mechanisms, particularly hepcidin (Hp) and NRAMP1. During TB infection, MTB infected macrophage induces hepcidin production from liver. Hepcidin potentially exert antimicrobial activity against mycobacteria, by promote localization of mycobacteria containing phagosome and cause cellular damages. NRAMP1 efflux pump potentially removes phagosomal iron out of the macrophages or cause iron influx resulting in killing of mycobacteria through ROS reaction. In malaria infection, infected RBC induces macrophages or liver to release hepcidin, triggered by IL-6 and IL-10 respectively. Higher hepcidin concentration in blood potentially increase iron stored in macrophages and resulting in diminished serum iron. Inhibition of ferroportin by hepcidin in HIV/AIDS could have detrimental effects, as increase cellular iron storage would ultimately speed up the HIV progression. Increased iron concentration in CD4+ T cell enhance reverse transcription, HIV-1 gene expression and capsid assembly
Iron, Mycobacterium tuberculosis and Tuberculosis
Mycobacterium tuberculosis (MTB) is an intracellular pathogen that resides within the phagosome of the macrophages, that have higher affinity to iron [67]. It was reported the supplementation of iron induce the growth of TB bacilli within macrophages, while limiting the supply potentially inhibits its multiplication [68]. Furthermore, an experimental study in iron-loaded female Balb/C mice showed significantly enhanced multiplication of bacilli in spleen and lungs, thus could worsen the prognosis of human tuberculosis. Mycobacteria produced two distinct siderophore, namely intracellular mycobactins and the extracellular exochelins/carboxymycobactin, in which mycobactin is found to be the virulence factor for MTB [69]. The iron-dependent regulator protein (IdeR) protein acts as the global iron-dependent transcriptional regulator, particularly IdeR-Fe complexes regulates the synthesis of exochelin and mycobactin [70].
Evidence suggests that stimulation of BALB/c mice lung macrophages with mycobacteria alone or combination of IFN-γ and M. tuberculosis induced hepcidin mRNA [71]. Furthermore, the human alveolar A549 epithelial cells, mycobacteria’s mannose-capped lipoarabinomannan and phosphatidylmyoinositol mannosides, were found to induce hepcidin mRNA. Peripheral blood mononuclear cells (PBMC) analysis revealed CD11c+ myeloid dendritic cells stimulated with killed MTB and live Mycobacterium bovis produced hepcidin, suggest hepcidin as potent antimicrobial peptide against mycobacteria. Similarly, expression of hepcidin mRNA and protein was noted when mouse RAW24.7 macrophage cell line and mouse bone-marrow derived macrophages were infected with Mycobacterium avium and MTB [72]. Moreover, hepcidin was found to localise mycobacteria containing phagososme, which potentially inhibit growth and cause structural damage to the mycobacteria. Besides that, NRAMP transporters either cause iron influx into MTB-containing phagosome resulting in bactericidal by forming ROS or iron efflux restrict bacterial growth by iron depletion [67]. In an animal study, deletion of Nramp1 gene were found to be as resistant as wild-type mice to infected with virulent MTB strain H37Rv, suggesting NRAMP is of limited value in resistance to TB in mice, but deciphering the resistance or susceptibility to tuberculosis in certain human populations would be important [73].
Iron, Plasmodium falciparum and Malaria
Plasmodium falciparum multiplication requires iron, both during the hepatic stage of growth and the disease-associated erythrocytic stages. There are five possible reasons that prelude the relationship of iron concentration in human host and susceptibility to malarial infections: (1) plasmodia does not appear to be utilizing the available iron in abundance supplies in the erythrocyte haemoglobin, but detoxifies through formation of the hemazoin complex, which comprises approximately 2.2 mol/L of iron; (2) plasmodia depends on the very small pool of labile iron in the cytoplasm, possibly an intraerythocytic pool of ferrous iron; (3) iron supplementation enhance susceptibility of human host to malaria, by stimulating erythropoiesis, as plasmodia have higher preference to reticulocytes; (4) zinc protoporhyrin produced during iron deficient erythropoiesis, could potentially inhibit formation of hemozoin, ultimately create unfavourable environmental for parasite’s growth; and (5) relationship between iron concentration and human immune system against malaria infections [74, 75]. The Pemba, Zanzibar randomized trial with routine supplementation with iron and folic acid in preschool children in a population with high rates of malaria showed an increased risk of severe illness and death [76].
Hepcidin up-regulation could potentially increase iron accumulation in macrophages and resulting in decrease serum iron, resulting in dyserythropoiesis and anemia [77]. Co-culturing P. falciparum infected RBC with PBMC resulting in increasing hepcidin mRNA synthesis by PBMC after 3 h of co-culture [78]. Even though, IL-6 has been shown to be correlated with hepcidin production, the infected RBC did not strongly induce IL-6 mRNA, suggesting there could be other factors apart from IL-6 in responsible for hepcidin production in the context of malaria [79]. The expression of hepcidin in livers of mice infected with Plasmodium berghei, could be potentially regulated by other factors such as inflammation and erythropoietin (EPO), rather than the iron-sensing pathway [80]. A recent study found that, activin B, which signal through the bone morphogenetic protein/sons of mothers against decapentaplegic (BMP/SMAD) pathway and has been associated with increased hepcidin during inflammation [81].
Iron, HIV and AIDS
HIV infection virtually affects all the organ functions of human body causing numerous metabolic imbalance and alteration of the immune system, particularly involving iron metabolism. The increasing iron concentration in HIV infection occurs due to accumulation in heart, liver, pancreas, macrophages, depletion of CD4+ T cell population and antioxidant storage [82]. Similarly, HIV-positive patients in Nnewi, South East Nigeria were reported with significant ferritin and transferrin iron loading [83]. Clinical observations on the effect of iron concentration on the prognosis of HIV-1 infection showed: (1) the rate of HIV progression was significantly accelerated in individuals with higher serum ferritin, (2) administration of low doses of oral iron with dapsone revealed higher rate of mortality among HIV-positive patients, (3) the haptoglobin 2-2 polymorphism among HIV-positive patients is linked with higher iron concentration and reduced rate of survival compared to normal polymorphism phenotypes, and (4) HIV-1 infected patients have shortened survival with higher iron stores in bone marrow [84].
In HIV infection, cells with ferroportin were found to inhibit HIV-1 transcription and inhibition of ferroportin, through action of hepcidin resulting in increasing HIV-1 transcription and viral protein production [85]. Blockage of ferroprotin by hepcidin, resulting in an increase cellular iron storage in HIV-infected macrophages, thereby promote rapid progression of HIV-infection [86]. Iron overload within CD4+ T cell population resulting in increased HIV life cycle such as reverse transcription, HIV-1 gene expression and capsid assembly. In such phenomenon, iron chelation would be able to reduce the amount of nucleic acids, viral replication and release and enhance downregulation the expression of adhesion molecules for HIV attachment and internalization [87].
The Finale: Who is Winning?
As both human and bacteria fairly exert their scavenging strategies especially during iron starvation, debating the winner of ‘iron-tug-of-war’ seems implausible. It is rather important to develop future therapeutic strategies such as designing novel antimicrobial agents that could potentially block the access of iron to the invading pathogen. This includes discovering antibodies (anti-Ig) against bacterial iron-regulated outer membrane protein (IROMP), which inhibit the binding of iron and/or heme complexes to the microbial receptor proteins. Apart from that, the iron transport abilities of siderophores could be utilized to carry drugs into cells by preparing conjugates called ‘sideromycins’ [14, 42]. Siderophore deferoxamine B produced by Streptomyces pilosus could be used therapeutically for treatment of acute iron intoxication and chronic iron overload diseases [43]. Unravelling the mechanisms involved in the struggle for iron could provide fundamental understanding of the host–pathogen interactions in infectious diseases.
Funding
This work was supported by Ministry of Higher Education, Malaysia through Fundamental Research Grant Scheme [04-01-14-53FR] and Universiti Putra Malaysia through Geran Putra—Inisiatif Putra Siswazah (IPS) [GP-IPS/2016/9478200].
Compliance of Ethical Standards
Conflicts of interest
The authors declare that they have no competing interests.
References
- 1.DiGuiseppi J, Fridovich I. Oxygen toxicity in Streptococcus sanguis. J Biol Chem. 1982;257:4046–4051. [PubMed] [Google Scholar]
- 2.Archibald F. Lactobacillus plantarum, an organism not requiring iron. FEMS Microbiol Lett. 1983;19:29–32. doi: 10.1111/j.1574-6968.1983.tb00504.x. [DOI] [Google Scholar]
- 3.Posey JE, Gherardini FC (2000) Lack of role for iron in lyme disease pathogen. Science (80) 288:1651–1653. 10.1126/science.288.5471.1651 [DOI] [PubMed]
- 4.Posey JE, Hardham JM, Norris SJ, Gherardini FC. Characterization of a manganese-dependent regulatory protein, TroR, from Treponema pallidum. Proc Natl Acad Sci USA. 1999;96:10887–10892. doi: 10.1073/pnas.96.19.10887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chen L, Zheng D, Liu B, et al. VFDB 2016: hierarchical and refined dataset for big data analysis—10 years on. Nucleic Acids Res. 2016;44:D694–D697. doi: 10.1093/nar/gkv1239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Symeonidis A, Marangos M (2012) Iron and microbial growth. In: Insight and control of Infectious disease in global scenario. InTech, pp. 289–330. 10.5772/34760
- 7.Von Drygalski A, Adamson JW. Iron metabolism in man. J Parenter Enter Nutr. 2013;37:599–606. doi: 10.1177/0148607112459648. [DOI] [PubMed] [Google Scholar]
- 8.Andrews NC, Schmidt PJ. Iron homeostasis. Annu Rev Physiol. 2007;69:69–85. doi: 10.1146/annurev.physiol.69.031905.164337. [DOI] [PubMed] [Google Scholar]
- 9.Schaible UE, Kaufmann SH. Iron and microbial infection. Nat Rev Microbiol. 2004;2:946–953. doi: 10.1038/nrmicro1046. [DOI] [PubMed] [Google Scholar]
- 10.Messenger AJ, Barclay R. Bacteria, iron and pathogenicity. Biochem Educ. 1983;11:54–63. doi: 10.1016/0307-4412(83)90043-2. [DOI] [Google Scholar]
- 11.Andrews SC, Robinson AK, Rodríguez-Quiñones F. Bacterial iron homeostasis. FEMS Microbiol Rev. 2003;27:215–237. doi: 10.1016/S0168-6445(03)00055-X. [DOI] [PubMed] [Google Scholar]
- 12.Cornelis P, Wei Q, Andrews SC, Vinckx T. Iron homeostasis and management of oxidative stress response in bacteria. Metallomics. 2011;3:540–549. doi: 10.1039/c1mt00022e. [DOI] [PubMed] [Google Scholar]
- 13.Wandersman C, Delepelaire P. Bacterial iron sources: from siderophores to hemophores. Annu Rev Microbiol. 2004;58:611–647. doi: 10.1146/annurev.micro.58.030603.123811. [DOI] [PubMed] [Google Scholar]
- 14.Hennigar SR, McClung JP. Nutritional immunity: starving pathogens of trace minerals. Am J Lifestyle Med. 2016 doi: 10.1177/1559827616629117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Indriati Hood M, Skaar EP. Nutritional immunity: transition metals at the pathogen-host interface. Nat Rev Microbiol. 2012 doi: 10.1038/nrmicro2836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.McLaren GD, Muir WA, Kellermeyer RW, Jacobs A. Iron overload disorders: natural history, pathogenesis, diagnosis, and therapy. CRC Crit Rev Clin Lab Sci. 1983;19:205–266. doi: 10.3109/10408368309165764. [DOI] [PubMed] [Google Scholar]
- 17.Khan FA, Fisher MA, Khakoo RA. Association of hemochromatosis with infectious diseases: expanding spectrum. Int J Infect Dis. 2007;11:482–487. doi: 10.1016/j.ijid.2007.04.007. [DOI] [PubMed] [Google Scholar]
- 18.Gangaidzo IT, Moyo VM, Mvundura E, et al. Association of pulmonary tuberculosis with increased dietary iron. J Infect Dis. 2001;184:936–939. doi: 10.1086/323203. [DOI] [PubMed] [Google Scholar]
- 19.Canziani MEF, Yumiya ST, Rangel EB, et al. Risk of bacterial infection in patients under intravenous iron therapy: dose versus length of treatment. Artif Organs. 2001;25:866–869. doi: 10.1046/j.1525-1594.2001.06894.x. [DOI] [PubMed] [Google Scholar]
- 20.Maynor L, Brophy DF. Risk of infection with intravenous iron therapy. Ann Pharmacother. 2007;41:1476–1480. doi: 10.1345/aph.1K187. [DOI] [PubMed] [Google Scholar]
- 21.Taylor RW, Manganaro L, O’Brien J, et al. Impact of allogenic packed red blood cell transfusion on nosocomial infection rates in the critically ill patient. Crit Care Med. 2002;30:2249–2254. doi: 10.1097/00003246-200210000-00012. [DOI] [PubMed] [Google Scholar]
- 22.Palmer KT, Poole J, Ayres JG, et al. Exposure to metal fume and infectious pneumonia. Am J Epidemiol. 2003;157:227–233. doi: 10.1093/aje/kwf188. [DOI] [PubMed] [Google Scholar]
- 23.Weinberg ED. Iron availability and infection. Biochim Biophys Acta—Gen Subj. 2009;1790:600–605. doi: 10.1016/j.bbagen.2008.07.002. [DOI] [PubMed] [Google Scholar]
- 24.Kumar V, Choudhry VP. Iron deficiency and infections. Indian J Pediatr. 2010;77:789–793. doi: 10.1007/s12098-010-0120-3. [DOI] [PubMed] [Google Scholar]
- 25.Chung MC-M. Structure and function of transferrin. Biochem Educ. 1984;12:146–154. doi: 10.1016/0307-4412(84)90118-3. [DOI] [Google Scholar]
- 26.Otto BR, Verweij-van Vught AM, MacLaren DM. Transferrins and heme-compounds as iron sources for pathogenic bacteria. Crit Rev Microbiol. 1992;18:217–233. doi: 10.3109/10408419209114559. [DOI] [PubMed] [Google Scholar]
- 27.Thorstensen K, Romslo I. The role of transferrin in the mechanism of cellular iron uptake. Biochem J. 1990;271:1–9. doi: 10.1042/bj2710001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Farnaud S, Evans RW. Lactoferrin—a multifunctional protein with antimicrobial properties. Mol Immunol. 2003;40:395–405. doi: 10.1016/S0161-5890(03)00152-4. [DOI] [PubMed] [Google Scholar]
- 29.Ward PP, Conneely OM. Lactoferrin: role in iron homeostasis and host defense against microbial infection. Biometals. 2004;17:203–208. doi: 10.1023/B:BIOM.0000027693.60932.26. [DOI] [PubMed] [Google Scholar]
- 30.Ward PP, Paz E, Conneely OM. Multifunctional roles of lactoferrin: a critical overview. Cell Mol Life Sci. 2005;62:2540–2548. doi: 10.1007/s00018-005-5369-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Vorland LH. Lactoferrin: a multifmctional glycoprotein. Acta Pathol Microbiol Immunol Scand. 1999;107:971–981. doi: 10.1111/j.1699-0463.1999.tb01499.x. [DOI] [PubMed] [Google Scholar]
- 32.Orino K, Watanabe K. Molecular, physiological and clinical aspects of the iron storage protein ferritin. Vet J. 2008;178:191–201. doi: 10.1016/j.tvjl.2007.07.006. [DOI] [PubMed] [Google Scholar]
- 33.Reif DW. Ferritin as a source of iron for oxidative damage. Free Radic Biol Med. 1992;12:417–427. doi: 10.1016/0891-5849(92)90091-T. [DOI] [PubMed] [Google Scholar]
- 34.Arosio P, Levi S. Ferritin, iron homeostasis, and oxidative damage. Free Radic Biol Med. 2002;33:457–463. doi: 10.1016/S0891-5849(02)00842-0. [DOI] [PubMed] [Google Scholar]
- 35.Arosio P, Ingrassia R, Cavadini P. Ferritins: a family of molecules for iron storage, antioxidation and more. Biochim Biophys Acta. 2009;1790:589–599. doi: 10.1016/j.bbagen.2008.09.004. [DOI] [PubMed] [Google Scholar]
- 36.Theil EC. Ferritin: at the crossroads of iron and oxygen metabolism. J Nutr. 2003;133:1549–1553. doi: 10.1093/jn/133.5.1549S. [DOI] [PubMed] [Google Scholar]
- 37.Carrondo MA. Ferritins, iron uptake and storage from the bacterioferritin viewpoint. EMBO J. 2003;22:1959–1968. doi: 10.1093/emboj/cdg215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Anzaldi LL, Skaar EP. Overcoming the heme paradox: heme toxicity and tolerance in bacterial pathogens. Infect Immun. 2010;78:4977–4989. doi: 10.1128/IAI.00613-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Tong Y, Guo M. Bacterial heme-transport proteins and their heme-coordination modes. Arch Biochem Biophys. 2009;481:1–15. doi: 10.1016/j.abb.2008.10.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Pieracci FM, Barie PS. Iron and the risk of infection. Surg Infect (Larchmt) 2005;6:41–46. doi: 10.1089/sur.2005.6.s1-41. [DOI] [PubMed] [Google Scholar]
- 41.Parrow NL, Fleming RE, Minnick MF. Sequestration and scavenging of iron in infection. Infect Immun. 2013;81:3503–3514. doi: 10.1128/IAI.00602-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Saha R, Saha N, Donofrio RS, Bestervelt LL. Review microbial siderophores: a mini review. J Basic Microbiol. 2013;53:303–317. doi: 10.1002/jobm.201100552. [DOI] [PubMed] [Google Scholar]
- 43.Miethke M, Marahiel MA. Siderophore-based iron acquisition and pathogen control. Microbiol Mol Biol Rev. 2007;71:413–451. doi: 10.1128/MMBR.00012-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Hotta K, Kim CY, Fox DT, Koppisch AT. Siderophore-mediated iron acquisition in Bacillus anthracis and related strains. Microbiology. 2010;156:1918–1925. doi: 10.1099/mic.0.039404-0. [DOI] [PubMed] [Google Scholar]
- 45.Cornelis P, Dingemans J. Pseudomonas aeruginosa adapts its iron uptake strategies in function of the type of infections. Front Cell Infect Microbiol. 2013;3:75. doi: 10.3389/fcimb.2013.00075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Skaar EP. The battle for iron between bacterial pathogens and their vertebrate hosts. PLoS Pathog. 2010;6:1–2. doi: 10.1371/journal.ppat.1000949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Evans RW, Oakhill JS. Transferrin-mediated iron acquisition by pathogenic Neisseria. Biochem Soc. 2002;30:705–707. doi: 10.1042/bst0300705. [DOI] [PubMed] [Google Scholar]
- 48.Noinaj N, Easley NC, Oke M, et al. Structural basis for iron piracy by pathogenic Neisseria. Nature. 2012;483:53–58. doi: 10.1038/nature10823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Genco CA, Desai PJ. Iron acquisition in the pathogenic Neisseria. Trends Microbiol. 1996;4:179–184. doi: 10.1016/0966-842X(96)10029-9. [DOI] [PubMed] [Google Scholar]
- 50.Schryvers AB, Stojiljkovic I. Iron acquisition systems in the pathogenic Neisseria. Mol Microbiol. 1999;32:1117–1123. doi: 10.1046/j.1365-2958.1999.01411.x. [DOI] [PubMed] [Google Scholar]
- 51.Gray-Owen SD, Loosmore S, Schryvers AB. Identification and characterization of genes encoding the human transferrin-binding proteins from Haemophilus influenzae. Infect Immun. 1995;63:1201–1210. doi: 10.1128/iai.63.4.1201-1210.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Loosmore SM, Yang YP, Coleman DC, et al. Cloning and expression of the Haemophilus influenzae transferrin receptor genes. Mol Microbiol. 1996;19:575–586. doi: 10.1046/j.1365-2958.1996.406943.x. [DOI] [PubMed] [Google Scholar]
- 53.Herrington DA, Sparling PF. Haemophilus influenzae can use human transferrin as a sole source for required iron. Infect Immun. 1985;48:248–251. doi: 10.1128/iai.48.1.248-251.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Gonzalez GC, Caamano DL, Schryvers AB. Identification and characterization of a porcine-specific transferrin receptor in Actinobacillus pleuropneumoniae. Mol Microbiol. 1990;4:1173–1179. doi: 10.1111/j.1365-2958.1990.tb00692.x. [DOI] [PubMed] [Google Scholar]
- 55.Perkins-Balding D, Ratliff-Griffin M, Stojiljkovic I. Iron transport systems in Neisseria meningitidis. Microbiol Mol Biol Rev. 2004;68:154–171. doi: 10.1128/MMBR.68.1.154-171.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Criado MT, Pintor M, Ferreirós CM. Iron uptake by Neisseria meningitidis. Res Microbiol. 1993;144:77–82. doi: 10.1016/0923-2508(93)90217-P. [DOI] [PubMed] [Google Scholar]
- 57.Rohde KH, Dyer DW. Mechanisms of iron acquisition by the human pathogens Neisseria Meningitidis and Neisseria Gonorrhoeae. Front Biosci. 2003;8:1186–1218. doi: 10.2741/1133. [DOI] [PubMed] [Google Scholar]
- 58.Ganz T. Iron in innate immunity: starve the invaders. Curr Opin Immunol. 2009;21:63–67. doi: 10.1016/j.coi.2009.01.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Wyllie S, Seu P, Goss JA. The natural resistance-associated macrophage protein 1 Slc11a1 (formerly Nramp1) and iron metabolism in macrophages. Microbes Infect. 2002;4:351–359. doi: 10.1016/S1286-4579(02)01548-4. [DOI] [PubMed] [Google Scholar]
- 60.Cellier MF, Courville P, Campion C. Nramp1 phagocyte intracellular metal withdrawal defense. Microbes Infect. 2007;9:1662–1670. doi: 10.1016/j.micinf.2007.09.006. [DOI] [PubMed] [Google Scholar]
- 61.Govoni G, Gros P. Macrophage NRAMP1 and its role in resistance to microbial infections. Inflamm Res. 1998;47:277–284. doi: 10.1007/s000110050330. [DOI] [PubMed] [Google Scholar]
- 62.Ganz T, Nemeth E. Hepcidin and iron homeostasis. Biochim Biophys Acta—Mol Cell Res. 2012;1823:1434–1443. doi: 10.1016/j.bbamcr.2012.01.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Rishi G, Wallace DF, Subramaniam VN. Hepcidin: regulation of the master iron regulator. Biosci Rep. 2015;35:e00192. doi: 10.1042/BSR20150014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Rossi E. Hepcidin–the iron regulatory hormone. Clin Biochem Rev. 2005;26:47–49. [PMC free article] [PubMed] [Google Scholar]
- 65.Collins JF, Wessling-Resnick M, Knutson MD. Hepcidin regulation of iron transport. J Nutr. 2008;138:2284–2288. doi: 10.3945/jn.108.096347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Bullen JJ, Rogers HJ, Spalding PB, Ward CG. Natural resistance, iron and infection: a challenge for clinical medicine. J Med Microbiol. 2006;55:251–258. doi: 10.1099/jmm.0.46386-0. [DOI] [PubMed] [Google Scholar]
- 67.Boelaert JR, Vandecasteele SJ, Appelberg R, Gordeuk VR. The effect of the host’s iron status on tuberculosis. J Infect Dis. 2007;195:1745–1753. doi: 10.1086/518040. [DOI] [PubMed] [Google Scholar]
- 68.Lounis N, Truffot-Pernot C, Grosset J, et al. Iron and Mycobacterium tuberculosis infection. J Clin Virol. 2001;20:123–126. doi: 10.1016/S1386-6532(00)00136-0. [DOI] [PubMed] [Google Scholar]
- 69.Sritharan M. Iron as a candidate in virulence and pathogenesis in mycobacteria and other microorganisms. World J Microbiol Biotechnol. 2000;16:769–780. doi: 10.1023/A:1008995313232. [DOI] [Google Scholar]
- 70.Pandey R, Rodriguez GM. IdeR is required for iron homeostasis and virulence in Mycobacterium tuberculosis. Mol Microbiol. 2014;91:98–109. doi: 10.1111/mmi.12441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Sow FB, Nandakumar S, Velu V, et al. Mycobacterium tuberculosis components stimulate production of the antimicrobial peptide hepcidin. Tuberculosis. 2011;91:314–321. doi: 10.1016/j.tube.2011.03.003. [DOI] [PubMed] [Google Scholar]
- 72.Sow FB, Florence WC, Satoskar AR, et al. Expression and localization of hepcidin in macrophages: a role in host defense against tuberculosis. J Leukoc Biol. 2007;82:934–945. doi: 10.1189/jlb.0407216. [DOI] [PubMed] [Google Scholar]
- 73.North RJ, Lacourse R, Ryan L, et al. Consequence of Nramp1 deletion to Mycobacterium tuberculosis infection in mice. Infect Immun. 1999;67:5811–5814. doi: 10.1128/iai.67.11.5811-5814.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Prentice AM. Iron metabolism, malaria, and other infections: what is all the fuss about? Symp A Q J Mod Foreign Lit. 2008 doi: 10.3945/jn.108.098806. [DOI] [PubMed] [Google Scholar]
- 75.Prentice AM, Ghattas H, Doherty C, Cox SE. Iron metabolism and malaria. Food Nutr Bull. 2007;28:524–539. doi: 10.1177/15648265070284S406. [DOI] [PubMed] [Google Scholar]
- 76.Sazawal S, Black R, Ramsan M, et al. Effect of routine prophylactic supplementation with iron and folic acid on admission to hospital and mortality in preschool children in a high malaria transmission setting: community based, randomised, placebo-controlled trial. Lancet. 2006;367:133–143. doi: 10.1016/S0140-6736(06)67962-2. [DOI] [PubMed] [Google Scholar]
- 77.Spottiswoode N, Duffy PE, Drakesmith H. Iron, anemia and hepcidin in malaria. Front Pharmacol. 2014;5:1–12. doi: 10.3389/fphar.2014.00125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Armitage AE, Pinches R, Eddowes LA, et al. Plasmodium falciparum infected erythrocytes induce hepcidin (HAMP) mRNA synthesis by peripheral blood mononuclear cells. Br J Haematol. 2009;147:769–771. doi: 10.1111/j.1365-2141.2009.07880.x. [DOI] [PubMed] [Google Scholar]
- 79.Prentice AM. Hepcidin and iron-mediated resistance to malaria. EMBO Mol Med. 2011;3:620–622. doi: 10.1002/emmm.201100170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Wang H-Z, He Y-X, Yang C-J, et al. Hepcidin is regulated during blood-stage malaria and plays a protective role in malaria infection. J Immunol. 2011;187:6410–6416. doi: 10.4049/jimmunol.1101436. [DOI] [PubMed] [Google Scholar]
- 81.Spottiswoode N, Armitage AE, Williams AR, et al. Role of activins in hepcidin regulation during malaria. Infect Immun. 2017;85:1–17. doi: 10.1128/IAI.00191-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Banjoko SO, Oseni FA, Togun RA, et al. Iron status in HIV-1 infection: implications in disease pathology. BMC Clin Pathol. 2012;12:1. doi: 10.1186/1472-6890-12-26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Manafa PO, Aneke JC, Okocha CE, et al. The human immunodeficiency virus infection is associated with positive iron balance among subjects in Nnewi, South East Nigeria. J HIV Hum Reprod. 2017;4:8–12. [Google Scholar]
- 84.Gordeuk VR, Delanghe JR, Langlois MR, Boelaert JR. Iron status and the outcome of HIV infection: an overview. J Clin Virol. 2001;20:111–115. doi: 10.1016/S1386-6532(00)00134-7. [DOI] [PubMed] [Google Scholar]
- 85.Xu M, Kashanchi F, Foster A, et al. Hepcidin induces HIV-1 transcription inhibited by ferroportin. Retrovirology. 2010 doi: 10.1186/1742-4690-7-104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Nekhai S, Kumari N, Dhawan S. Role of cellular iron and oxygen in the regulation of HIV-1 infection. Future Virol. 2013;8:301–311. doi: 10.2217/fvl.13.6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Chang H-C, Bayeva M, Taiwo B, et al. High cellular iron levels are associated with increased HIV infection and replication. AIDS Res Hum Retroviruses. 2015;31:305–312. doi: 10.1089/aid.2014.0169. [DOI] [PMC free article] [PubMed] [Google Scholar]