

Abstract
The biotechnology of desert plants is a vast subject. The main applications in this broad field of study comprises of plant tissue culture, genetic engineering, molecular markers and others. Biotechnology applications have the potential to address biodiversity conservation as well as agricultural, medicinal, and environmental issues. There is a need to increase our knowledge of the genetic diversity through the use of molecular genetics and biotechnological approaches in desert plants in the Arabian Gulf region including those in the United Arab Emirates (UAE). This article provides a prospective research for the study of UAE desert plant diversity through DNA fingerprinting as well as understanding the mechanisms of both abiotic stress resistance (including salinity, drought and heat stresses) and biotic stress resistance (including disease and insect resistance). Special attention is given to the desert halophytes and their utilization to alleviate the salinity stress, which is one of the major challenges in agriculture. In addition, symbioses with microorganisms are thought to be hypothesized as important components of desert plant survival under stressful environmental conditions. Thus, factors shaping the diversity and functionality of plant microbiomes in desert ecosystems are also emphasized in this article. It is important to establish a critical mass for biotechnology research and applications while strengthening the channels for collaboration among research/academic institutions in the area of desert plant biotechnology.
Keywords: Biotechnology, Desert plant, Genes, Halophytes, Microbiome, Salinity, UAE
Introduction
Plant genetic resources play a significant role in the sustenance of mankind. The maintenance of sufficient genetic diversity with a known or potential value ensures its availability for utilization by present and future generations, including food and agriculture. In particular, the need for germplasm conservation is recognized for crops and plants with agricultural importance, as in the case of wild plant genetic resources. Due to the adverse effects of various natural and anthropogenic factors, the loss of diversity in wild plant genetic resources is a matter of great concern and requires effective conservation strategies. As a result of plant genome evolution, it is well known that the loss of genes or genetic combinations existing in plants with specific traits is irreversible; hence, plant genetic resources should be conserved to safeguard the botanical heritage. Given the harsh desert environmental conditions, plants face many challenges with respect to soil properties, salinity, high temperatures and water scarcity (Gutterman 2002; McNeely 2003). Therefore, the unique strategies for survival and adaptation of desert plants in such conditions have been the subject of advanced research by scientists of plant science disciplines.
The UAE’s desert plant species may comprise important tolerance and resistance genes against drought, salinity, high temperature and disease, which could be utilized for various purposes including greening the deserts (Alam et al. 2017) and crop improvement programs. The UAE’s flora exhibits several interesting characteristics which have evolved as adaptations to the region’s extreme climatic conditions such as high temperatures, salinity and drought. Thus, the indigenous plant species show specific physiological, behavioral and morphological adaptations to survive under these harsh environmental conditions. At present, the UAE is home to around 830 plant species. This number is gradually increasing as species previously unrecorded in the country are discovered (Gairola et al. 2017). Most of the recent additions to the flora of country are weeds from artificial microhabitats, therefore the actual number of indigenous species is fewer than the 830 previously stated. The vegetation of the UAE includes both halophytic and non-halophytic types. These indigenous desert plants are valuable biodiversity resources and the ecological, socio-economic, and medicinal value of many of these species is well recognized. Recent research shows that halophytes growing in the UAE contain special active phytochemicals, possibly due to their tolerance to severe environmental conditions that previal there (Cybulska et al. 2014).
Importance of desert plant biotechnological research
Desert plant biotechnology is an exciting area for research and innovation. Through biotechnological applications (e.g. genetic engineering), the genome of an organism can be directly manipulated either by adding new exogenous genes, or by reducing the endogenous genes expression. Multigene transfer, site-specific integration and specifically regulated gene expression are important advanced approaches in plant biotechnology (Liu et al. 2013). Thus, recent advances in various biotechnological techniques will pave the way for the introduction of desirable genes and promise to revolutionize biodiversity science. In recent years, researchers have been mainly concentrating on identifying a group of abiotic stress related genes in plants to enhance the plants tolerance to these stresses using genetic modification approaches (Smirnoff 1998; Wang et al. 2003). Currently, stress tolerance traits (particularly tolerance to drought and salinity) are considered significant and are gaining increasing attention. Engineering a salt and drought tolerant plant requires deep understanding of the salt and drought tolerance mechanisms and the associated pathways (Fig. 1). Additionally, the identification of salt and drought stress related genes, transcripts and/or proteins is a crucial step towards this goal. Priority research areas in plant biotechnology include tissue culture and micropropagation, molecular marker technologies, genetic engineering for plant improvement and the “omics” sciences (e.g., genomics, transcriptomics, proteomics, metabolomics etc.) which facilitate the high throughput analysis of plant samples for the identification of candidate genes associated with environmental stresses tolerance (Mir et al. 2012; Jain 2012; Hahn et al. 2013; Gupta and Huang 2014; Garg et al. 2016; Mosa et al. 2017a).
Fig. 1.
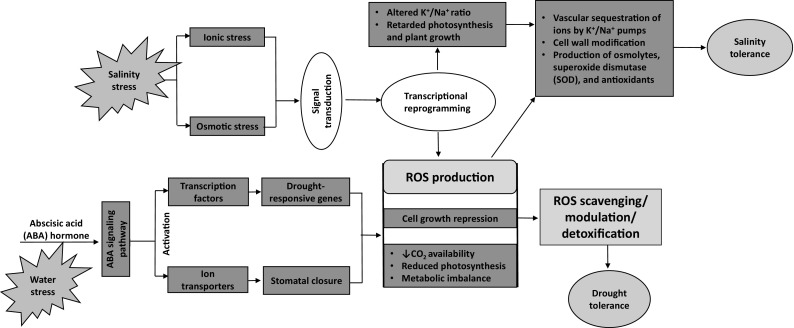
Schematic diagram of salinity tolerance (Partly adapted from Hanumantha Rao et al. 2016; Malhotra et al. 2013 and Onaga and Wydra 2016), and drought tolerance (Partly adapted from Osakabe et al. 2014 and Mosa et al. 2017b) mechanisms in plants
Plant biotechnology has great potential in improving agriculture efficiency, environment and health which will in turn, have a positive impact in the development of countries with arid desert lands (Al-Khalifah 2004). Micropropagation of desert plants (namely Calligonum comosum, Acacia tortilis and A. ehrenbergiana) has been attempted by Al-Khalifah (2004). This study focused on the utilization of desert plants that need less water when planted in the local deserts of Saudi Arabia and other regions with same environmental conditions. A protocol for the in vitro micropropagation of nodal explants of the endangered desert shrub Haloxylon persicum was also developed (Mohamed et al. 2013) and DNA fingerprinting was performed on date palm cultivars grown in the field and produced using tissue cultures (Aljuhani 2016). Phoenix dactylifera (Date Palms) is one of the most important crops in the Arab Gulf Region and neighbouring countries with desert areas. In UAE, the application of micropropagation of date palm and ornamental plants has received increasing attention. Several studies have described early work on date palm regeneration through micropropagation techniques (e.g. Omar et al. 1992; Al-Khalifah 2004; Al-Khayri 2007, 2010) and salinity and drought stress tolerance (Al-Khayri and Ibraheem 2014). In offering an effective propagation means, tissue culture techniques have proven to be applicable in the genetic improvement of date palms through induced mutations and in vitro selection (Al-Khayri 2011). Biotechnology can help in the development of date palm by understanding the genetic basis of these plants, also in helping to produce large numbers of tissue cultured plantlets that will quickly expand planting and replanting of date groves, and finally, to improve fruit yield and increase tolerance to biotic and abiotic stress in the new cultivars using molecular breeding approaches (Johnson 2011; Al-Khayri and Ibraheem 2014).
Desert plants are capable of withstanding very harsh and fluctuating environmental conditions, and are adapted to cope with different stresses in their life cycle. It has been reported that intrinsic metabolic capabilities of plants help them to cope up frequently with such unpredictable and extreme conditions (Simontacchi et al. 2015). Frequent variations in the outside environmental conditions and combination of diverse stresses could disturb plant metabolic activities and consequently force them to acquire some unique genetic and metabolic mechanisms within their cellular system for their survival (Gill and Tuteja 2010; Meena et al. 2017). In recent years, there has been an increasing interest in researching plants from extreme deserts environments for understanding the mechanisms of tolerance to a combination of diverse stress conditions, their survival strategies, and identifying novel stress tolerance determinants (Al-Whaibi 2011; Eshel et al. 2017).
Some recent studies on detecting regulations of stress-related genes using RNA-Seq analysis are available from the Gulf region. For example, Shokry et al. (2014) have utilized an RNA-seq approach to identify a USP-like gene in Calotropis procera, which is generally involved in response to a number of environmental stresses like heat and/or drought. Mutwakil et al. (2017) studied the transcriptomic and metabolic response of Calotropis procera when treated with polyethylene glycol and sodium chloride (drought and osmotic stress). As a result, a sharp increase of myo-inositol, a key signaling molecule was detected. This indicates that inositol signaling has a critical part in regulating stress response. An increase of amino acid concentration and polyphenols were also detected in stressed samples. In addition, key plant hormones such as cytokinins were synthesized and induced multiple downstream effects like closing the stomatal aperture to reduce water loss, reducing cell wall rigidity and accumulation of metabolites to balance osmotic pressure. Hence, the data obtained from this study provided recent insights on adaptation strategies of C. procera to the hyper-arid environments and can be helpful for future studies.
Obaid et al. (2016) studied the regulatory mechanisms of thermotolerance in an Arabian Desert plant Rhazya stricta. Using RNA-Seq analysis, apical and mature leaves of this species were analyzed for the expression of heat responsive genes. This study revealed that R. stricta exhibits metabolic strategies to cope with heat stress. Such insights on biochemical and molecular strategies will help in better understanding the regulatory mechanisms of thermotolerance in R. stricta and other native plants of the Arabian deserts. Plants detect heat stress through increase in membrane fluidity, which induces the activation of several pathways/ transcription factors/ and genes to facilitate the thermotolerance mechanisms as shown in Fig. 2. Another study by Hajrah et al. (2017) examined the salinity tolerance mechanism of Rhazya stricta by performing high-throughput RNA-Seq analysis. Conclusions were drawn showing that the different mechanisms adapted by R. stricta is for avoiding protein misfolding, aggregation or denaturation during salt stress to protect the RNA editing patterns from changes and the efficient production of several key secondary metabolites, like flavonoids, and ROS that helps the plant to withstand the affecting stress. Bahieldin et al. (2015) used the mRNA-Seq approach to analyze range of genes that get expressed under salt stress in Hordeum spontaneum leaves. These authors concluded that the salt stress responsive genes were involved in several biological processes, including reactive oxygen species (ROS) scavenging, electron transport and exchanger mechanisms, protein refolding, flavonoid biosynthesis, ethylene production, and signaling network. Drought is another serious limitation for the growth of plants. However, plants of the Arabian Peninsula exhibit a remarkable capability in surviving the dryness of the desert. An example of drought tolerance can be seen in C. procera, which grows in harsh, arid and stressful conditions. Ramadan et al. (2014) analyzed the metabolomic and lipidomic response with respect to rehydration of C. procera. The results indicated that this plant survive the harsh conditions by rapidly changing its metabolism according to the presence or absence of water. It was found that the availability of water increased the levels of amino acids and membrane lipids, while decreasing sucrose, raffinose, maltitol and storage lipids (triacylglycerols). Therefore, the data suggested that the ability of Calotropis procera to tolerate drought is dependent upon its rapid adaptation to water availability and water loss.
Fig. 2.
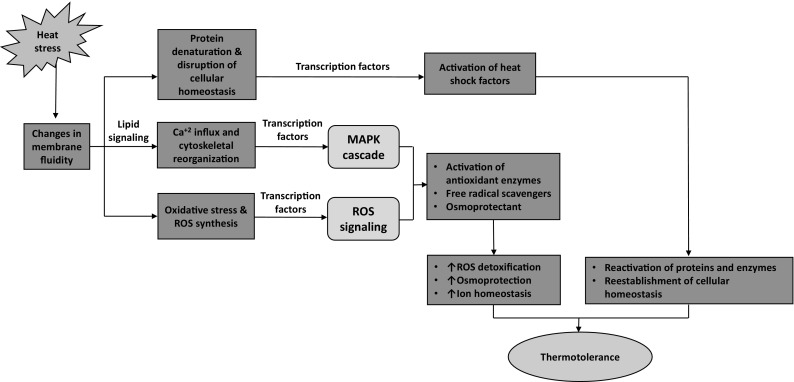
Schematic diagram of heat tolerance in plants (Partly adapted from Hanumantha Rao et al. 2016). Plants detect heat stress through increase in membrane fluidity, which induces lipid signaling. This in turn activates calcium channels to release calcium ions into the cytosol. Heat stress also causes the release of reactive oxygen species (ROS) and the denaturation of proteins. Transcription factors will then activate heat responsive genes, thereby regulating the expression of osmoprotectants, detoxifying enzymes and molecular chaperons to facilitate protein folding
Molecular marker techniques such as amplified fragment length polymorphism (AFLP), random amplified polymorphic DNA (RAPD), simple sequence repeat (SSR), single nucleotide polymorphism (SNP), and especially whole genome sequencing techniques are extensively used in the study of biological diversity, genetic transformation and the conservation of the genetic resources of desert plants (Khan et al. 2012). A recent attempt to use single nucleotide polymorphism (SNP) analysis to characterize date palm biodiversity was carried out by Sabir et al. (2014). In this study, mitochondrial and plastid genomes of nine Saudi Arabian cultivars were sequenced. For each cultivar, sequences were aligned against their respective reference genomes, and SNPs were identified. Cultivar-specific SNPs for eight out of the nine cultivars were successfully identified. However, using the plastid and mitochondrial genomes required removing regions of transferred plastid sequences to avoid mistakes in SNP identification. This is because the sequences are highly conserved. These authors have suggested that the use of nuclear genome in the identification of SNPs will be more accurate.
Creating biological barcodes using molecular taxonomy to identify organisms is the central goal of DNA barcoding. The use of DNA barcoding during floristic studies may help in identification of species, especially when plant specimens lack diagnostic floral or fruit characteristics at the time of their collection. Recently, next-generation sequencing, i.e. genomics and transcriptomics, has been used to study the mechanism of adaptation seen in desert plants such as Rhazya stricta (Yates et al. 2014), Reaumuria soongarica (Shi et al. 2013), and Ammopiptanthus mongolicus (Zhou et al. 2012). As a primary feature of desert plants is the ability for extensive and unique adaptation, such findings allows for a better understanding of their abilities to adapt to many types of stress factors prevailing in desert environments (Zhao et al. 2014). The development of expressed sequence tags (ESTs) and cDNA libraries using various genomic approaches such as suppressive subtractive hybridization (SSH), serial analysis of gene expression (SAGE) and cDNA microarray (Breyne and Zabeau 2001) can create databases for understanding the genetic network involved in abiotic stress tolerance mechanisms of halophytes (Popova et al. 2008; Kore-eda et al. 2004; Wang and Showalter 2004). Additionally, in employing such approaches, stress responsive genes can be identified and isolated which could be used to develop enhanced stress tolerant plants with improved crop yield (Lokhande and Suprasanna 2012).
Potential manipulation of desert halophytes
Different plant biotechnological approaches are currently utilized to solve the major challenges in agriculture such as abiotic stresses including salt, drought, high temperature etc. Soil salinity, for instance, is a big challenge since the majority of agricultural crop plants are salt-sensitive glycophytes that are usually lost due to an increase in soil salinity levels (Ventura et al. 2014). Halophytes, on the other hand, are salt tolerant plants that could be of great potential in industrial, ecological and agricultural fields (Koyro et al. 2013). The Arabian Gulf region possesses several important halophyte species such as Zygophyllum mandavillei, Salicornia europaea, Suaeda maritima, Suaeda vermiculata, Salsola imbricata, Sesuvium verrucosum, Arthrocnemum macrostachyum, Atriplex farinosa, Halopeplis perfoliata etc. (Gairola et al. 2015).
The Salicornia/Sarcocornia species have been extensively studied to showcase their multipurpose applications that include biodiesel, oil (Glenn et al. 1991, 2013), vegetable, ornamental (Ventura and Sagi 2013), bioremediation (Webb et al. 2012, 2013; Shpigel et al. 2013), fodder (Glenn et al. 1992; Imai et al. 2004) and probiotic (Sarker et al. 2010) usages. Mesembryanthemum crystallinum is another well studied halophyte and considered to be model plant for studying abiotic stresses. It activates the change from the C3 to CAM photosynthetic mechanism (Bohnert and Cushman 2000). In addition, it can be used in ornamental gardening (Jessop 1986). The Suaeda, Atriplex and Thellungiella halophila species (homologous to the glycophyte model, A. thaliana) (Wang and Showalter 2004; Amtmann 2009) are also great models for studying the molecular basis of stress tolerance mechanisms in halophytes (Lokhande and Suprasanna 2012). Several genes encoding for ion transporters facilitating the influx, efflux, compartmentation and translocation of different toxic ions which results in enhanced salt stress tolerance in plants have been identified (Kumar and Mosa 2015). In one attempt, salt tolerant grass has been developed using Agrobacterium-mediated transformation by inserting rice vacuolar membrane Na+/H+ antiporter gene in the grass DNA. The transformed plants demonstrated the ability to tolerate salt stress more strongly than non-transformed plants (Wu et al. 2005). An additional advantage of transgenic salt tolerant plants is their resistance to other abiotic stresses like heat, drought and freezing (Zhu 2001).
Oh et al. (2009) investigated the role of ThSOS1, a Salt Overly Sensitive 1 (SOS1) homolog in Thellungiella. SOS1 has an important function in the ion homeostasis mechanism (Shi et al. 2000). The knockdown of this gene made Thellungiella sensitive to salt stress like Arabidopsis. It was also found that ThSOS1 is responsible for limiting Na+ accumulation and the distribution of Na+ ions (Oh et al. 2009). This precise response to environmental stresses indicated that Thellungiella has the potential to be used as a model plant for abiotic stress studies. A proteomics analysis of Thellungiella leaves under salinity stress identified important salt responsive proteins which are mainly involved in starch and sucrose metabolism, suggesting its possible role in salt tolerance (Wang et al. 2013).
Prospective on desert plant microbiome research
Plant-associated microorganisms can assist plants in many ways, including stimulating growth and promoting stress resistance (Lugtenberg and Kamilova 2009; Yang et al. 2009). Increasing evidence suggests that microbial interactions play a key role in plant health and productivity and are important determinants of plant biodiversity (Berg et al. 2015). In desert environments, plant distribution and diversity are mainly restricted by environmental stresses (e.g. low water availability, high temperatures and salinity). In such habitats, plant microbiome research efforts are mainly focused on divulging the mechanisms by which the microbiome permits the plant to survive various environmental stresses. Advancements in the application of molecular, omics and novel microscopic techniques in microbial ecology has allowed for broad and inclusive studies of plant microbiomes (Jansson et al. 2012; Berg et al. 2015). Berg and Smalla (2009) and Hartmann et al. (2009) described plant species-specific rhizosphere microbial communities via molecular fingerprints and molecular strain analysis. Information on the diversity and abundance of plant associated microbiomes has paramount importance in promoting plant growth, and the use of molecular methods in studying these communities. Beside the profound impact of symbiotic bacteria and fungi respectively on plant growth, productivity and adaptation to stresses, the plant microbiome can also increase the yield of crops with agronomical value (Turner et al. 2013; Panke-Buisse et al. 2015; Coleman-Derr et al. 2016).
The Rhizosphere contains soil microorganisms that are in direct proximity with plants, the Phyllosphere, on the other hand, contains microbial species (epiphytes) that grow on the plant’s surface. In addition, some microbial populations can be also found inside the plant itself and are known as endophytes (Ruiz-Pérez et al. 2016). Plant roots can be associated with a variety of endosymbiotic microbes including mycorrhizal fungi, rhizosphere bacteria and endophyte microorganisms (Dighton et al. 2005; González-Teuber et al. 2017). It has also been highlighted that through the regulation of gene expression, bacteria that are associated with plants, specifically endophytes, may help in conferring elevated osmotic and drought stress tolerance (Compant et al. 2010). However, the information on the extent of influence of microorganisms on plant gene regulation, as well as, the effect of climate change on such plant-microbe interactions is still inadequate and requires further research. The UAE desert flora needs to be explored more to provide insight regarding the structure and function of endophyte community which has effect in plant functions. As stated by Kaplan et al. (2013), the extent plant communities regulate and are regulated by their microbiomes is an area that needs to be explored along with mechanisms for acclimating and adapting to stress. Investigation of the endophyte community associated with the UAE’s native desert plants will serve as baseline research that can help in understanding the relationship between target plants and their endosymbiotic microbes.
Coleman-Derr et al. (2016) carried out a comprehensive and extensive investigation of microbial communities associated with species of Agave plants, which are native to semiarid and arid regions of Central and North America. Using Illumina amplicon sequencing, different plant compartments (rhizosphere, phyllosphere, leaf and root endosphere) were inspected for both prokaryotic and fungal communities. They also analyzed soil samples taken from areas that are proximal and distal to the cultivated and native agaves. With the help of phylogenetic profiling, results revealed a difference within the sample types and among the different plant compartments which concludes that the composition of prokaryotic communities can be influenced by these two factors. The same conclusions were previously drawn for agaves by Desgarennes et al. (2014) and were further confirmed by using next generation sequencing (NGS) in this study. However, the geographical distribution of host species chiefly influenced the constitution of the fungal communities. Native agaves revealed higher prokaryotic diversity than the cultivated A. tequilana, although the microbial diversity remained the same in the endosphere. Thus, it can be concluded from this study that a group of prokaryotic and fungal organisms living inside agaves helps in stimulating plant growth and increasing plant tolerance to abiotic stresses such as drought. In another study conducted by Fonseca-García et al. (2016), two native and sympatric Cacti species (Myrtillocactus geometrizans and Opuntia robusta) were analyzed in terms of composition, diversity and potential factors influencing above- and below-ground bacterial, archaeal and fungal communities. Plant compartment as revealed by phylogenetic profiling, is what influenced the constitution of microbial communities. While fungi found in the stem part of Cacti species displayed specific connection to their host, bacterial and archaeal diversity found to be much less in the rhizosphere part when compared to the phyllosphere of Cacti. In addition, comparison of microbial diversity revealed that the semi-arid soils featured the highest level while stem endosphere featured the lowest level. Regardless of their taxonomic distance, both species shared most microbial taxa in all analyzed compartments. The authors also carried out biochemical and molecular analysis of Cacti seed-borne bacteria. Results obtained proposed that vertical inheritance of these microbial symbionts can be achieved along with promoting plant development and tolerating water scarcity (Fonseca-García et al. 2016). Such information is valuable to promote and sustain agriculture in arid and semi-arid regions.
In arid lands, reverse desertification can be applied to sustain soil stability and fertility. Hence, desert farming sounds promising in promoting productivity and preventing desertification. Marasco et al. (2012) intended to analyze the effect of desert farming on plant-microbe association in the drought-sensitive pepper plant (Capsicum annuum L.) cultivated in the arid conditions of a traditional Egyptian farm. The root system was dissected into the endosphere, the rhizosphere and the root-surrounding soil. To gain an insight regarding the bacterial community structure and diversity, a 16S rRNA gene PCR-DGGE analysis was done. It revealed a difference in community fingerprinting according to the microhabitat which suggests a selective pressure determined by plant activity. A total of 299 bacterial strains were cultured from the root system. 68% of the isolates were of the Bacillus species recovered from the endosphere. The Klebsiella species were found to dominate both the rhizosphere (61% of the isolates) and the root surrounding soil fractions (44% of the isolates) (Marasco et al. 2012). Plant Growth Promoting (PGP) activities and tolerances to abiotic stress analysis revealed that most of the isolates (95%) showed such capabilities, although a difference in the distribution was found over the root system fractions that were analyzed (with enhanced abilities for Bacillus and the rhizobacteria strains). The conclusion drawn was that the rhizosphere of C. annuum enriched populations of PGP bacteria which played a crucial rule in overcoming abiotic stress (Marasco et al. 2012). Yuan et al. (2016) hypothesized the existence of a tight correlation between the halo-tolerant coastal plant Suaeda salsa (Amaranthaceae) and below ground microbial communities that has an impact on plant productivity and stress tolerance. Phylogenetic framework analysis focused on traits was also performed depending on the bacterial 16S rRNA gene and fungal nuclear rRNA internal transcribed spacer profiling. The investigations revealed 2 dominant classes (α-proteobacteria and γ-proteobacteria) of communities reside in bulk soil and root endosphere while both were found to be more phylogenetically clustered and at the same time exhibiting phylogenetic over-dispersion in the rhizosphere. With the assistance of the PICRUSt tool for predicting bacterial community functions, they found that the genomes of both the rhizospheric and endophytic bacteria contained genes contributing to nutrient solubilization, salt stress acclimatization, and competitive root colonization. This interpretation was supported by the existence of a great variety of well-known halotolerant rhizobacteria species.
Plant microbiome research in the UAE is necessary to be attempted in greater detail. Vegetation in the country is mainly dominated by arid zone species. Soils are generally poor in nutrients, lacking particularly nitrogen. Stressful desert conditions, low water availability or water deficit, temperature stress and salinity are serious threats to rhizobium-legume symbiosis (Sharma et al. 2013). All these factors affect soil productivity in numerous ways, including lack of organic matter in the soil and increase in soil salinity, which is worse in hyper-arid desert climates and consequently leads to the deterioration of agricultural crops (Ksiksi and El-Shaigy 2012). Legume plants can play a critical role when integrated as a part of native or agricultural ecosystems with low fertility quality, for instance saline soil, by increasing nitrogen. The salinity tolerance of rhizobia isolated from the root nodules of Sesbania sesban, Lablab purpureus and Cajanus cajan were analyzed. It has been reported that the desert soils isolated rhizobia can tolerate and grow under high salt concentrations as well as nodulate their leguminous plants. Sharma et al. (2013) highlighted the ability of rhizobia to tolerate and survive under extreme desert conditions which enables them to become highly valuable inoculums to aid in enhancing the productivity of leguminous plants cultivated under harsh environments. Further research on the identification of the rhizobial strains through molecular characterization, evaluation of growth performance, symbiotic efficiency and nodulating ability against temperature, pH and heavy metals will provide a better understanding of survival strategies under stressful desert conditions (Sharma et al. 2013). To enhance legume productivity under symbiosis, a promising approach can be implemented by inoculating it with salt-tolerant strains of rhizobia (Keneni et al. 2010). A comprehensive review by Laranjo et al. (2014) focuses on the genus Mesorhizobium which includes species with high geographical dispersion and the ability to nodulate a range of economically vital food and forage legume crops. The review highlighted the importance of using biotechnological approaches and further investigating symbiosis genes in addition to stress response genes in order to develop more effective mesorhizobia inoculants.
Rising biotechnology research in UAE
Encouragingly, some universities, research centers and institutes have recently initiated biotechnology education and research programs in the UAE. For example, the University of Sharjah offers a Bachelor of Sciences in Biotechnology which was initiated in the academic year 2008/2009. The UAE also recognized the significance of plant biotechnology when Khalifa Center for Genetic Engineering and Biotechnology (KCGEB) opened in 2014. The center focuses on expanding the usage of desert plant biotechnological approaches in the UAE along with assisting in providing a sustainable desert agro-ecosystem. The research work in this center focuses on increasing the ability of plants to tolerate drought, heat and salt (www.uaeu.ac.ae/en/kcgeb). Similarly, in 2014, the International Center for Biosaline Agriculture (ICBA) established its plant biotechnology laboratory with the intention to implement genetic engineering in order to grow novel crops by coupling genetics and genomic approaches. The main focus of this research was to understand the molecular mechanisms underlying plant responses to harsh desert environments such as soil salinity (www.biosaline.org/content/molecular-biology). Similarly, the Masdar institute is also promoting industrial biotechnology research in the UAE by focusing on the development of biofuels from halophytes.
Researchers are using a number of biotechnological approaches for the conservation and improvement of plant species for desired traits (Khan et al. 2012). In particular, biotechnological tools are necessary for the conservation and improvement of economically and medicinally important rare and endangered plant species. Dutta et al. (2012) developed a protocol for Agrobacterium-mediated genetic transformation of the desert shrub Leptadenia pyrotechnica using hypocotyl explants from 5 days old seedlings. The method was further enhanced by the use of sonication. For reliability purposes, the species were transformed using two reporter genes, GUS and GFP. Their presence was confirmed by PCR analysis and southern blots. A recent study by Enan et al. (2017) on the barcoding of ten medicinal plant species found in the UAE suggested that the three candidate barcodes (rbcL, matK, and rpoC1) tested were successfully amplified with the universal primer pairs. The authors also highlighted the difficulty in extracting DNA from desert plants as many of them contain high levels of secondary metabolites. Therefore, there is a need to carry out extensive DNA barcoding work on the UAE flora in order to standardize and modify the protocols for obtaining genomic DNA of higher quality and quantity from various plant types. However, the UAE still has a long way to go in terms of advancing their biotechnology sector and fully utilizing DNA barcoding in desert plants. Primarily, there is a need to establish a comprehensive DNA barcode library of the UAE’s plant species for taxonomic clarification and other conservation and research purposes. The Sharjah Research Academy (SRA) is currently in the process of establishing a state of the art plant molecular biology laboratory with a focus on the documentation and conservation of the country’s plant genetic resources. This would be the first priority in permitting in-depth research on the use of DNA from the UAE’s native plants in different applications. This research would also focus on studying plant response mechanisms to various abiotic stresses (e.g. drought, salinity, temperature, etc.) at the molecular level and in the fields of microevolution (population genetics) and macroevolution (phylogenetics) of important desert plants. The molecular biology laboratory will offer an opportunity for research, education and training in desert plant molecular biology and biotechnology. Beside seed banks and other conservation measures, biotechnological approaches coupled with molecular tools can be implemented for the conservation of various rare and threatened UAE plant species. For several desert species, seed conservation specifically in germplasm banks is achievable. However, when some species have very low sexual reproductive success, micropropagation techniques are generally applied to rapidly overcome the reproductive problems. Thus, the use of in vitro and plant tissue culture techniques for ex situ conservation purposes is particularly important when there is low or no possibility of seed conservation.
Biotechnology also plays a great role in research involving the identification of heavy metal tolerant and sensitive genotypes as well as the characterization of genes involved in heavy metal tolerance. In fact, biotechnology has opened new horizons in phytoremediation research by enabling the direct gene transfer to improve plant capabilities to cleanup environmental pollutants (Ibañez et al. 2015; Mosa et al. 2016). Some of the halophytes found in the Arabian Gulf region are reported to be better salt/metal accumulators (Almahasheer et al. 2013). Thus, halophytic plants represent an emerging trend in phytoremediation research and are expected to receive considerable attention in the near future (Shabala 2013). Recently, the phytochemical investigation of some UAE halophytes revealed that these plants are a source of unique active phytochemicals, concluding that the phenomena might be likely occurring due to the tolerance properties by the halophytes to harsh local environmental conditions plants (Cybulska et al. 2014). Hence, these plants are of great research interest due to their salt tolerance properties in conjunction with potential ecological, economic and medicinal importance.
Plants are exposed to either individual or multiple environmental stresses, and depending on the type of stress, plants can activate certain resistance pathways (Alcázar and Parker 2011). However, the type of plant’s response is dependent on whether a single or multiple stresses are applied. This was confirmed by microarray analyses which revealed that individual stress elicited a transcriptional response that was different from when the plant was exposed to combinations of biotic or abiotic stresses (Rizhsky et al. 2004; Voelckel and Baldwin 2004; Matsui et al. 2008). By comparing different stress responses, various changes at the genetic and molecular levels can be identified (Rizhsky et al. 2004; Mittler and Blumwald 2010). Consequently, this process guides the production of genetically engineered plants with ability to tolerate several stresses. As such, a recent study carried out by researchers from the United Arab Emirates University analyzed the responses of the Arabidopsis plant to both biotic and abiotic stresses (Sham et al. 2014). This was accomplished by analyzing earlier microarray data from studies on the transcriptomic responses of Arabidopsis induced by biotic stress (infection with Botrytis cinerea) as well as abiotic stresses (cold, drought and oxidative stresses). The authors were able to identify genes that were uniquely expressed to each stress in addition to those expressed commonly amongst all the types of stresses studied. Hence, this study and other similar types of studies can help in determining the frequently expressed genes under stressful environmental conditions. Moreover, to enhance the plant resistance to several stresses, additional analysis on the functions of encoded products may lead to the identification of possible targets to be genetically modified.
One of the most important potential applications of biotechnology is in the area of medicinal plants research. Medicinal plants are receiving more attention in recent years due to their use in traditional remedies for a wide variety of ailments and the commercialization of herbal formulations. In the UAE, a total of 132 plant species were reported to have medicinal value (Sakkir et al. 2012). There is a need to promote propagation and sustainable utilization of these underutilized plants through biotechnology. Biotechnological approaches have been increasingly applied for mass propagation of medicinal plants, conservation of their germplasm, study and production of secondary metabolites and genetic improvement. Such research may contribute to the growing trend in the field of pharmacognosy, i.e. to utilize medicinal plants as bio-factories for the production of bioactive substances with possible therapeutic uses (Hussain et al. 2012; Tasheva and Kosturkova 2013).
Future perspective
Advanced biotechnological applications that may be utilized in relation to medicine, the environment and agriculture, need to be explored in regards to desert plants. As shown in Fig. 3, the stress responsive and disease resistance genes, transcripts, proteins, metabolites and microbiome of desert plants can provide benefits for various sectors such as conservation, environment, agriculture and medicine with the aid of plant biotechnology. Thus, it is important and essential to set potential thrust areas for plant biotechnological research. The research work should focus more on applications including marker-assisted selection, tissue culture, genetic engineering, genomics and other related areas. In our opinion, most of the research areas mentioned in this article should be considered in a collaborative manner. It is also essential to strengthen plant biotechnology and molecular biology research and education by setting up new research centers of excellence and offering attractive fellowship programs. When considering the potential importance of halophytes, there is a need to establish a ‘Halophyte Biotechnology Center’ and ‘Stress Physiology laboratory’ to address research in these prospective areas. Moreover, better coordination between academic and research institutions across the country could be promoted to create more effective biotechnology and molecular biology research programs.
Fig. 3.
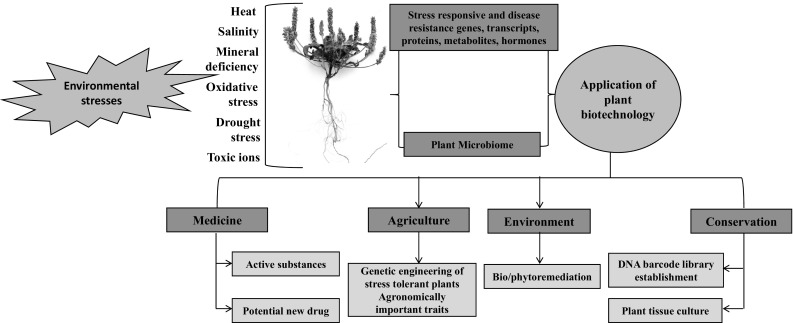
Conceptual framework summarizing the potential biotechnology applications of desert plants. Stress responsive and disease resistance genes, transcripts, proteins, metabolites and microbiomes of desert plants can provide benefits for various sectors such as conservation, the environment, agriculture and medicine with the aid of plant biotechnology
The adverse effects of climate change (mainly fluctuating temperatures and frequent drought events) on crop productivity are expected to increase in the coming decade. In this context, agricultural biotechnologies have major potential in facilitating and promoting sustainable agriculture, and the power of biotechnologies to generate useful genotypes should be beneficially harnessed. Growing evidence suggests that biotechnology allows plant breeders to select genes that produce beneficial traits and transfer them from one organism to another. Similarly, desired agronomic traits can be transferred into target crops. Additionally, current genome-editing technologies such as ZFNs (zinc finger nucleases), TALENs (transcription activator like effectors nucleases) or CRISPR-Cas9 (clustered regularly interspaced short palindromic repeats) allows the genetic modification of targeted genes by editing the targeted plant native genome itself without the need of transferring foreign genes (Xiong et al. 2015). Several reported crop plants such as rice, wheat, corn, and tomato have been developed with engineered traits via genome editing approaches (Song et al. 2016). At present, biotechnology for enhanced abiotic stress tolerance in crop plants is gaining increasing attention in research. In the arid desert region, research in generating novel abiotic stress tolerant genotypes via the introduction of gene constructs can potentially enhance drought, heat and salt stress tolerance and is of prime importance. Biotechnology coupled with molecular biology tools play an important role in determining plant responses to drought and salinity via the identification and characterization of the genes involved, and help improve the yield under unfavorable climate conditions. The heat and /or drought and salinity stress responses have been successfully characterized by utilizing RNA-Seq in some desert plants of the Gulf region (e.g., Shokry et al. 2014; Bahieldin et al. 2015; Obaid et al. 2016). The recent insights obtained from these RNA-seq studies would improve our understanding of genes involved in diverse stress tolerance mechanisms of desert plants. Thus, utilizing next generation sequencing, especially the RNA-seq approaches for molecular discovery in desert plants of the UAE is required.
Research networking is an effective means for enhancing scientific cooperation and maximizing information exchange at various levels. Building platforms for conducting biotechnological research of desert plants in institutions across the UAE will allow biotechnologists to investigate the different mechanisms of salinity, heat and drought tolerance as well as examine the functional ecology and genetics involved while enhancing their molecular understanding of plant microbiomes. Additionally, developing regional research collaborations with other countries of the Arabian Peninsula with similar floral diversity and environmental stresses (such as Oman, Kuwait and also Saudi Arabia which has rich and diverse flora) is critical. Such collaborations will also help the training of research scientists and the development of expertise in local biotechnology research programs.
Acknowledgements
The authors thank Dr. Amr Abdel-Hamid, Director General of the Sharjah Research Academy, for his encouragement and support.
Compliance with ethical standards
Conflict of interest
The authors declare that there is no conflict of interest regarding the publication of this paper.
Contributor Information
Sanjay Gairola, Email: sgairola@sharjah.ac.ae.
Kareem A. Mosa, Email: kmosa@sharjah.ac.ae
References
- Alam H, Khattak JZK, Ppoyil SBT, Kurup S, Ksiksi TS. Landscaping with native plants in the UAE: a review. Emir J Food Agric. 2017;29(10):729–741. [Google Scholar]
- Alcázar R, Parker JE. The impact of temperature on balancing immune responsiveness and growth in Arabidopsis. Trends Plant Sci. 2011;16(12):666–675. doi: 10.1016/j.tplants.2011.09.001. [DOI] [PubMed] [Google Scholar]
- Aljuhani WS. Genetic diversity and the impact of geographical location on the relationships between Phoenix dactylifera L. germplasms grown in Saudi Arabia. Hered Genet. 2016;5:172. [Google Scholar]
- Al-Khalifah NS (2004) The role of Biotechnology in developing plant resources in deserts environment. In: Proceedings of international conference on water resources and arid environment (WRAE04). King Abdulatziz City, KSA, pp 1–16
- Al-Khayri JM. Date Palm Phoenix Dactylifera L. Micropropagation. In: Jain SM, Haggman H, editors. Protocols for micropropagation of woody trees and fruits. Berlin: Springer; 2007. pp. 509–526. [Google Scholar]
- Al-Khayri JM. Somatic embryogenesis of date palm (Phoenix dactylifera L.) improved by coconut water. Biotech. 2010;9(4):477–484. [Google Scholar]
- Al-Khayri JM. Basal salt requirements differ according to culture stage and cultivar in date palm somatic embryogenesis. Am J Biochem Biotechnol. 2011;7(1):32–42. [Google Scholar]
- Al-Khayri JM, Ibraheem Y (2014) In vitro selection of abiotic stress tolerant date palm (Phoenix dactylifera L.): A review. Emir J Food Agric 26(11):921-933. 10.9755/ejfa.v26i11.18975
- Almahasheer HB, Al-Taisan WA, Mohamed MK. Metals accumulation in grey mangrove (Avicennia marina (Forsk.) Vierh.) inhabiting Tarut Bay. Eastern Saudi Arabia. J Argic Sci. 2013;6:137. [Google Scholar]
- Al-Whaibi MH. Plant heat-shock proteins: a mini review. J King Saud Univ Sci. 2011;23(2):139–150. [Google Scholar]
- Amtmann A. Learning from evolution: Thellungiella generates new knowledge on essential and critical components of abiotic stress tolerance in plants. Mol Plant. 2009;2(1):3–12. doi: 10.1093/mp/ssn094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bahieldin A, Atef A, Sabir JSM, Gadalla NO, Edris S, Alzohairy AM, Radhwan NA, Baeshen MN, Ramadan AM, Eissa HF, Hassan SM, Baeshen NA, Abuzinadah O, Al-Kordy MA, El-Domyati FM, Jansen RK. RNA-Seq analysis of the wild barley (H. spontaneum) leaf transcriptome under salt stress. C R Biol. 2015;338:285–297. doi: 10.1016/j.crvi.2015.03.010. [DOI] [PubMed] [Google Scholar]
- Berg G, Smalla K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol Ecol. 2009;68(1):1–13. doi: 10.1111/j.1574-6941.2009.00654.x. [DOI] [PubMed] [Google Scholar]
- Berg G, Rybakova D, Grube M, Köberl M. The plant microbiome explored: implications for experimental botany. J Exp Bot. 2015;67(4):995–1002. doi: 10.1093/jxb/erv466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bohnert HJ, Cushman JC. The ice plant cometh: lessons in abiotic stress tolerance. J Plant Growth Regul. 2000;19:334–346. [Google Scholar]
- Breyne P, Zabeau M. Genome-wide expression analysis of plant cell cycle modulated genes. Curr Opin Plant Biol. 2001;4:136–142. doi: 10.1016/s1369-5266(00)00149-7. [DOI] [PubMed] [Google Scholar]
- Coleman-Derr D, Desgarennes D, Fonseca-Garcia C, Gross S, Clingenpeel S, Woyke T, North G, Visel A, Partida-Martinez LP, Tringe SG. Plant compartment and biogeography affect microbiome composition in cultivated and native Agave species. New Phytol. 2016;209(2):798–811. doi: 10.1111/nph.13697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Compant S, Heijden MG, Sessitsch A. Climate change effects on beneficial plant–microorganism interactions. FEMS Microbiol Ecol. 2010;73(2):197–214. doi: 10.1111/j.1574-6941.2010.00900.x. [DOI] [PubMed] [Google Scholar]
- Cybulska I, Brudecki G, Alassali A, Thomsen M, Brown JJ. Phytochemical composition of some common coastal halophytes of the United Arab Emirates. Emir J Food Agric. 2014;26(12):1046–1056. [Google Scholar]
- Desgarennes D, Garrido E, Torres-Gomez MJ, Peña-Cabriales JJ, Partida-Martinez LP. Diazotrophic potential among bacterial communities associated with wild and cultivated Agave species. FEMS Microbiol Ecol. 2014;90:844–857. doi: 10.1111/1574-6941.12438. [DOI] [PubMed] [Google Scholar]
- Dighton J, White JF, Oudemans P. The fungal community: its organization and role in the ecosystem. Boca Raton: CRC Press; 2005. [Google Scholar]
- Dutta I, Kottackal M, Tumimbang E, Tajima H, Zaid A, Blumwald E. Sonication-assisted efficient Agrobacterium-mediated genetic transformation of the multipurpose woody desert shrub Leptadenia pyrotechnica. Plant Cell Tiss Organ Cult. 2012;112(3):289–301. [Google Scholar]
- Enan MR, Palakkott AR, Ksiksi TS. DNA barcoding of selected UAE medicinal plant species: a comparative assessment of herbarium and fresh samples. Physiol Mol Biol Plants. 2017;23(1):221–227. doi: 10.1007/s12298-016-0412-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eshel G, Shaked R, Kazachkova Y, Khan A, Eppel A, Cisneros A, Acuna T, Gutterman Y, Tel-Zur N, Rachmilevitch S, Fait A. Anastatica hierochuntica, an Arabidopsis Desert relative, is tolerant to multiple abiotic stresses and exhibits species-specific and common stress tolerance strategies with its halophytic relative, Eutrema (Thellungiella) salsugineum. Front Plant Sci. 2017;7:1992. doi: 10.3389/fpls.2016.01992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fonseca-García C, Coleman-Derr D, Garrido E, Visel A, Tringe SG, Partida-Martínez LP. The Cacti microbiome: interplay between habitat-filtering and host-specificity. Front Microbiol. 2016;7:150. doi: 10.3389/fmicb.2016.00150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gairola S, Bhatt A, El-Keblawy A. A perspective on potential use of halophytes for reclamation of salt-affected lands. Wulfenia. 2015;22:88–97. [Google Scholar]
- Gairola S, Mahmoud T, Shabana H, El-Keblawy A. Growing knowledge about the floral diversity of United Arab Emirates: new additions and conservation through seed banking. Tribulus. 2017;24:136–143. [Google Scholar]
- Garg R, Shankar R, Thakkar B, Kudapa H, Krishnamurthy L, Mantri N, Varshney RK, Bhatia S, Jain M. Transcriptome analyses reveal genotype-and developmental stage-specific molecular responses to drought and salinity stresses in chickpea. Sci Rep. 2016;6(1):19228. doi: 10.1038/srep19228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gill SS, Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem. 2010;48(12):909–930. doi: 10.1016/j.plaphy.2010.08.016. [DOI] [PubMed] [Google Scholar]
- Glenn EP, O’Leary JW, Watson MC, Thompson LT, Kuehl RO. Salicornia bigelovii Torr.: an oilseed halophyte for seawater irrigation. Science. 1991;251:1065–1067. doi: 10.1126/science.251.4997.1065. [DOI] [PubMed] [Google Scholar]
- Glenn EP, Coates WE, Riley JJ, Kuehl RO, Swingle RS. Salicornia bigelovii Torr.: a seawater-irrigated forage for goats. Anim Feed Sci Technol. 1992;40:21–30. [Google Scholar]
- Glenn EP, Anday T, Chaturvedi R, Martinez-Garcia R, Pearlstein S, Soliz D, Nelson SG, Felger RS. Three halophytes for saline-water agriculture: an oilseed, a forage and a grain crop. Environ Exp Bot. 2013;92:110–121. [Google Scholar]
- González-Teuber M, Vilo C, Bascuñán-Godoy L. Molecular characterization of endophytic fungi associated with the roots of Chenopodium quinoa inhabiting the Atacama Desert, Chile. Genom Data. 2017;11:109–112. doi: 10.1016/j.gdata.2016.12.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gupta B, Huang B. Mechanism of salinity tolerance in plants: physiological, biochemical, and molecular characterization. Int J Genom. 2014;2014:1–18. doi: 10.1155/2014/701596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gutterman Y. Minireview: survival adaptations and strategies of annuals occurring in the Judean and Negev Deserts of Israel. Isr J Plant Sci. 2002;50(3):165–175. [Google Scholar]
- Hahn A, Kilian J, Mohrholz A, Ladwig F, Peschke F, Dautel R, Harter K, Berendzen KW, Wanke D. Plant core environmental stress response genes are systemically coordinated during abiotic stresses. Int J Mol Sci. 2013;14(4):7617–7641. doi: 10.3390/ijms14047617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hajrah NH, Obaid AY, Atef A, Ramadan AM, Arasappan D, Nelson CA, Edris S, Mutwakil MZ, Alhebshi A, Gadalla NO, Makki RM, Al-Kordy MA, El-Domyati FM, Sabir JSM, Khiyami MA, Hall N, Bahieldin A, Jansen RK. Transcriptomic analysis of salt stress responsive genes in Rhazya stricta. PLoS ONE. 2017;12(5):e0177589. doi: 10.1371/journal.pone.0177589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanumantha Rao B, Nair R, Nayyar H. Salinity and high temperature tolerance in mungbean [Vigna radiata (L.) Wilczek] from a physiological perspective. Front Plant Sci. 2016;7:957. doi: 10.3389/fpls.2016.00957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartmann A, Schmid M, Van Tuinen D, Berg G. Plant-driven selection of microbes. Plant Soil. 2009;321(1–2):235–257. [Google Scholar]
- Hussain MS, Fareed S, Ansari S, Rahman MA, Ahmad IZ, Saeed M. Current approaches toward production of secondary plant metabolites. J Pharm Bioallied Sci. 2012;4(1):10–20. doi: 10.4103/0975-7406.92725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ibañez S, Talano M, Ontañon O, Suman J, Medina MI, Macek T, Agostini E. Transgenic plants and hairy roots: exploiting the potential of plant species to remediate contaminants. New Biotechnol. 2015;33:625–635. doi: 10.1016/j.nbt.2015.11.008. [DOI] [PubMed] [Google Scholar]
- Imai H, Kinoshita M, Ohishi M. Chemical characterization of glycerolipids and cerebrosides in a halophytic plant, Salicornia europaea L. J Oleo Sci. 2004;53(7):337–341. [Google Scholar]
- Jain M. Next-generation sequencing technologies for gene expression profiling in plants. Brief Funct Genom. 2012;11(1):63–70. doi: 10.1093/bfgp/elr038. [DOI] [PubMed] [Google Scholar]
- Jansson JK, Neufeld JD, Moran MA, Gilbert JA. Omics for understanding microbial functional dynamics. Environ Microbiol. 2012;14:1–3. doi: 10.1111/j.1462-2920.2011.02518.x. [DOI] [PubMed] [Google Scholar]
- Jessop JP. Family—Aizoaceae (Ficoidaceae, Mesembryanthemaceae, Molluginaceae, Tetragoniaceae) In: Jessop JP, Toelken HR, editors. Flora of South Australia Part I, Lycopodiaceae—Rosaceae. 4. Adelaide: South Australian Government Publishing Division; 1986. p. 415. [Google Scholar]
- Johnson DV. Introduction: date palm biotechnology from theory to practice. In: Jain SM, Al-Khayri JM, Johnson DV, editors. Date palm biotechnology. Dordrecht: Springer; 2011. pp. 1–11. [Google Scholar]
- Kaplan D, Maymon M, Agapakis CM, Lee A, Wang A, Prigge BA, Volkogon M, Hirsch AM. A survey of the microbial community in the rhizosphere of two dominant shrubs of the Negev Desert highlands, Zygophyllum dumosum (Zygophyllaceae) and Atriplex halimus (Amaranthaceae), using cultivation-dependent and cultivation-independent methods. Am J Bot. 2013;100:1713–1725. doi: 10.3732/ajb.1200615. [DOI] [PubMed] [Google Scholar]
- Keneni A, Assefa F, Prabu PC. Characterization of acid and salt-tolerant rhizobial strains isolated from faba bean fields of Wollo, Northern Ethiopia. J Agric Sci Technol. 2010;12(3):365–376. [Google Scholar]
- Khan S, Al-Qurainy F, Nadeem M. Biotechnological approaches for conservation and improvement of rare and endangered plants of Saudi Arabia. Saudi J Biol Sci. 2012;19(1):1–11. doi: 10.1016/j.sjbs.2011.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kore-eda S, Cushman MA, Akselrod I, Bufford D, Fredrickson M, Clark E, Cushman JC. Transcript profiling of salinity stress responses by large-scale expressed sequence tag analysis in Mesembryanthemum crystallinum. Gene. 2004;341:83–92. doi: 10.1016/j.gene.2004.06.037. [DOI] [PubMed] [Google Scholar]
- Koyro H, Hussain T, Huchzermeyer B, Khan MA. Photosynthetic and growth responses of a perennial halophytic grass Panicum turgidum to increasing NaCl concentrations. Environ Exp Bot. 2013;91:22–29. [Google Scholar]
- Ksiksi TS, El-Shaigy NO. Growth responses of nutrient-stressed Cenchrus ciliaris under carbon dioxide enrichment. J Earth Sci Clim Change. 2012 [Google Scholar]
- Kumar K, Mosa KA (2015) Ion transporters: a decisive component of salt stress tolerance in plants. In: Wani SH, Anwar Hossain A (eds) Managing salt tolerance in plants: molecular and genomic perspectives, CRC Press, pp 373–390
- Laranjo M, Alexandre A, Oliveira S. Legume growth-promoting rhizobia: an overview on the Mesorhizobium genus. Microbiol Res. 2014;169(1):2–17. doi: 10.1016/j.micres.2013.09.012. [DOI] [PubMed] [Google Scholar]
- Liu W, Yuan JS, Stewart CN., Jr Advanced genetic tools for plant biotechnology. Nat Rev Genet. 2013;14:781–793. doi: 10.1038/nrg3583. [DOI] [PubMed] [Google Scholar]
- Lokhande VH, Suprasanna P. Prospects of halophytes in understanding and managing abiotic stress tolerance. In: Prasad MNV, Ahmad P, editors. Environmental adaptations and stress tolerance of plants in the era of climate change. New York: Springer; 2012. pp. 29–56. [Google Scholar]
- Lugtenberg B, Kamilova F. Plant-growth-promoting rhizobacteria. Annu Rev Microbiol. 2009;63:541–556. doi: 10.1146/annurev.micro.62.081307.162918. [DOI] [PubMed] [Google Scholar]
- Malhotra K, Chhabra GK, Jain R, Sharma V, Kumar S. Drought and salinity tolerant biofuel crops for the Thar Desert. In: Tuteja N, Singh Gill S, editors. Plant Acclimation to environmental stress. New York: Springer; 2013. [Google Scholar]
- Marasco R, Rolli E, Ettoumi B, Vigani G, Mapelli F, Borin S, et al. A drought resistance-promoting microbiome is selected by root system under desert farming. PLoS ONE. 2012;7(10):e48479. doi: 10.1371/journal.pone.0048479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsui A, Ishida J, Morosawa T, Mochizuki Y, Kaminuma E, Endo TA, Okamoto M, Nambara E, Nakajima M, Kawashima M, Satou M, Kim JM, Kobayashi N, Toyoda T, Shinozaki K, Seki M. Arabidopsis transcriptome analysis under drought, cold, high-salinity and ABA treatment conditions using a tiling array. Plant Cell Physiol. 2008;49(8):1135–1149. doi: 10.1093/pcp/pcn101. [DOI] [PubMed] [Google Scholar]
- McNeely JA. Biodiversity in arid regions: values and perceptions. J Arid Environ. 2003;54(1):61–70. [Google Scholar]
- Meena KK, Sorty AM, Bitla UM, Choudhary K, Gupta P, Pareek A, Singh DP, Prabha R, Sahu PK, Gupta VK, Singh HB. Abiotic stress responses and microbe-mediated mitigation in plants: the omics strategies. Front Plant Sci. 2017;8:172. doi: 10.3389/fpls.2017.00172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mir RR, Zaman-Allah M, Sreenivasulu N, Trethowan R, Varshney RK. Integrated genomics, physiology and breeding approaches for improving drought tolerance in crops. Theor Appl Genet. 2012;125(4):625–645. doi: 10.1007/s00122-012-1904-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mittler R, Blumwald E. Genetic engineering for modern agriculture: challenges and perspectives. Annu Rev Plant Biol. 2010;61:443–462. doi: 10.1146/annurev-arplant-042809-112116. [DOI] [PubMed] [Google Scholar]
- Mohamed MA, Assaeed AM, Yousuf HN. Micropropagation of the endangered desert shrub Haloxylon persicum. Aust J Crop Sci. 2013;7(2):255–260. [Google Scholar]
- Mosa KA, Saadoun I, Kumar K, Helmy M, Dhankher OP. Potential biotechnological strategies for the cleanup of heavy metals and metalloids. Front Plant Sci. 2016;7:303. doi: 10.3389/fpls.2016.00303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mosa KA, Ismail A, Helmy M. Functional genomics combined with other omics approaches for better understanding abiotic stress tolerance in plants. In: Mosa KA, Ismail A, Helmy M, editors. Plant stress tolerance: an integrated omics approach. Basel: Springer; 2017. pp. 55–73. [Google Scholar]
- Mosa KA, Ismail A, Helmy M. Introduction to Plant Stresses. In: Mosa KA, Ismail A, Helmy M, editors. Plant stress tolerance: an integrated omics approach. Switzerland: Springer; 2017. pp. 1–16. [Google Scholar]
- Mutwakil M, Hajrah N, Atef A, Edris S, Sabir M, Al-Ghamdi A, et al. Transcriptomic and metabolic responses of Calotropis procera to salt and drought stress. BMC Plant Biol. 2017;17(1):231. doi: 10.1186/s12870-017-1155-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Obaid AY, Sabir JSM, Atef A, Liu X, Edris S, El-Domyati FM, Mutwakil MZ, Gadalla NO, Hajrah NH, Al-Kordy MA, Hall N, Bahieldin A, Jansen RK. Analysis of transcriptional response to heat stress in Rhazya stricta. BMC Plant Biol. 2016;16:252. doi: 10.1186/s12870-016-0938-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oh D-H, Leidi E, Zhang Q, Hwang S-M, Li Y, Quintero FJ, Jiang X, D’Urzo MP, Sang Lee Y, Zhao Y, Bahk JD, Bressan RA, Yun D-J, Pardo JM, Bohnert HJ. Loss of halophytism by interference with SOS1 expression. Plant Physiol. 2009;15:210–222. doi: 10.1104/pp.109.137802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Omar, Hameed MK, Al-Rawi . Micropropagation of date palm (Phoenix dactylifera L.) In: Bajaj YPS, editor. Biotechnology in agriculture and forestry, vol. 18 high-tech and micropropagation II. Berlin: Springer; 1992. pp. 471–490. [Google Scholar]
- Onaga G, Wydra K (2016). Advances in plant tolerance to abiotic stresses. In: Abdurakhmonov IY (ed), Plant genomics. InTech. 10.5772/64351
- Osakabe Y, Osakabe K, Shinozaki K, Tran L. Response of plants to water stress. Front Plant Sci. 2014 doi: 10.3389/fpls.2014.00086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panke-Buisse K, Poole AC, Goodrich JK, Ley RE, Kao-Kniffin J. Selection on soil microbiomes reveals reproducible impacts on plant function. ISME J. 2015;9(4):980–989. doi: 10.1038/ismej.2014.196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Popova OV, Yang O, Dietz KJ, Golldack D. Differential transcript regulation in Arabidopsis thaliana and the halotolerant Lobularia maritima indicates genes with potential function in plant salt adaptation. Gene. 2008;423:142–148. doi: 10.1016/j.gene.2008.07.017. [DOI] [PubMed] [Google Scholar]
- Ramadan A, Sabir JSM, Alakilli SYM, Shokry AM, Gadalla NO, Edris S, Al-Kordy MA, Al-Zahrani HS, El-Domyati FM, Bahieldin A, Baker NR, Willmitzer L, Irgang S. Metabolomic response of Calotropis procera growing in the desert to changes in water availability. PLoS ONE. 2014;9(2):e87895. doi: 10.1371/journal.pone.0087895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rizhsky L, Liang HJ, Shuman J, Shulaev V, Davletova S, Mittler R. When defense pathways collide. The response of Arabidopsis to a combination of drought and heat stress. Plant Physiol. 2004;134(4):1683–1696. doi: 10.1104/pp.103.033431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruiz-Pérez CA, Restrepo S, Zambrano MM. Microbial and functional diversity within the Phyllosphere of Espeletia species in an Andean High-Mountain ecosystem. Appl Environ Microbiol. 2016;82(6):1807–1817. doi: 10.1128/AEM.02781-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sabir JSM, Arasappan D, Bahieldin A, Abo-Aba S, Bafeel S, Zari TA, Edris S, Shokry AM, Gadalla NO, Ramadan AM, Atef A, Al-Kordy MA, El-Domyati FM, Jansen RK. Whole mitochondrial and plastid genome SNP analysis of nine date palm cultivars reveals plastid heteroplasmy and relationships among cultivars. PLoS ONE. 2014;9(4):e94158. doi: 10.1371/journal.pone.0094158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakkir S, Kabshawi M, Mehairbi M. Medicinal plants diversity and their conservation status in the United Arab Emirates (UAE) J Med Plants Res. 2012;6(7):1304–1322. [Google Scholar]
- Sarker SK, Park S-R, Kim G-M, Yang C-J. Hamcho (Salicornia herbacea) with probiotics as alternative to antibiotic for broiler production. J Med Plant Res. 2010;4(5):415–420. [Google Scholar]
- Shabala S. Learning from halophytes: physiological basis and strategies to improve abiotic stress tolerance in crops. Ann Bot. 2013;112(7):1209–1221. doi: 10.1093/aob/mct205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sham A, Al-Azzawi A, Al-Ameri S, Al- Mahmoud B, Awwad F, Al-Rawashdeh A, Iratni R, AbuQamar S. Transcriptome analysis reveals genes commonly induced by Botrytis cinerea infection, cold, drought and oxidative stresses in Arabidopsis. PLoS ONE. 2014;9(11):e113718. doi: 10.1371/journal.pone.0113718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma SR, Rao NK, Gokhale TS, Ismail S. Isolation and characterization of salt-tolerant rhizobia native to the desert soils of United Arab Emirates. Emir J Food Agric. 2013;25(2):102. [Google Scholar]
- Shi H, Ishitani M, Kim C, Zhu JK. The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc Natl Acad Sci USA. 2000;97(12):6896–6901. doi: 10.1073/pnas.120170197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi Y, Yan X, Zhao P, Yin H, Zhao X, Xiao H, Li X, Chen G, Ma X-F. Transcriptomic analysis of a tertiary relict plant, extreme Xerophyte Reaumuria soongorica to identify genes related to drought adaptation. PLoS ONE. 2013;8(5):e63993. doi: 10.1371/journal.pone.0063993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shokry AM, Al-Karim S, Ramadan A, Gadallah N, Al Attas SG, Sabir JS, Hassan SM, Madkour MA, Bressan R, Mahfouz M, Bahieldin A. Detection of a Usp-like gene in Calotropis procera plant from the de novo assembled genome contigs of the high-throughput sequencing dataset. C R Biol. 2014;337(2):86–94. doi: 10.1016/j.crvi.2013.12.008. [DOI] [PubMed] [Google Scholar]
- Shpigel M, Ben-Ezra D, Shauli L, Sagi M, Ventura Y, Samocha T, Lee JJ. Constructed wetland with Salicornia as a biofilter for mariculture effluents. Aquac. 2013;412–413:52–63. [Google Scholar]
- Simontacchi M, Galatro A, Ramos-Artuso F, Santa-Maria GE. Plant survival in a changing environment: the role of nitric oxide in plant responses to abiotic stress. Front Plant Sci. 2015;6:977. doi: 10.3389/fpls.2015.00977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smirnoff N. Plant resistance to environmental stress. Curr Opin Biotechnol. 1998;9(2):214–219. doi: 10.1016/s0958-1669(98)80118-3. [DOI] [PubMed] [Google Scholar]
- Song G, Jia M, Chen K, Kong X, Khattak B, Xie C, et al. CRISPR/Cas9: a powerful tool for crop genome editing. Crop J. 2016;4(2):75–82. [Google Scholar]
- Tasheva K, Kosturkova G. Role of biotechnology for protection of endangered medicinal plants. In: Petre M, editor. Environmental biotechnology—new approaches and prospective applications. Croatia: InTech; 2013. pp. 235–238. [Google Scholar]
- Turner TR, Ramakrishnan K, Walshaw J, Heavens D, Alston M, Swarbreck D, Poole PS. Comparative metatranscriptomics reveals kingdom level changes in the rhizosphere microbiome of plants. ISME J. 2013;7(12):2248–2258. doi: 10.1038/ismej.2013.119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ventura Y, Sagi M. Halophyte crop cultivation: the case for Salicornia and Sarcocornia. Environ Exp Bot. 2013;92:144–153. [Google Scholar]
- Ventura Y, Eshel A, Pasternak D, Sagi M. The development of halophyte-based agriculture: past and present. Ann Bot. 2014;115(3):529–540. doi: 10.1093/aob/mcu173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Voelckel C, Baldwin IT. Herbivore-induced plant vaccination. Part II. Array-studies reveal the transience of herbivore-specific transcriptional imprints and a distinct imprint from stress combinations. Plant J. 2004;38(4):650–663. doi: 10.1111/j.1365-313X.2004.02077.x. [DOI] [PubMed] [Google Scholar]
- Wang LW, Showalter AM. Cloning and salt-induced, ABA-independent expression of choline monooxygenase in Atriplex prostrate. Physiol Plant. 2004;120:405–412. doi: 10.1111/j.0031-9317.2004.00247.x. [DOI] [PubMed] [Google Scholar]
- Wang W, Vinocur B, Altman A. Plant responses to drought, salinity and extreme temperatures: towards genetic engineering for stress tolerance. Planta. 2003;218(1):1–14. doi: 10.1007/s00425-003-1105-5. [DOI] [PubMed] [Google Scholar]
- Wang X, Chang L, Wang B, Wang D, Li P, Wang L, et al. Comparative proteomics of Thellungiella halophila leaves from plants subjected to salinity reveals the importance of chloroplastic starch and soluble sugars in halophyte salt tolerance. Mol Cell Proteomics. 2013;12(8):2174–2195. doi: 10.1074/mcp.M112.022475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webb JM, Quinta R, Papadimitriou S, Norman L, Rigby M, Thomas D, Vay LL. Halophyte filter beds for treatment of saline wastewater from aquaculture. Water Res. 2012;46(16):5102–5114. doi: 10.1016/j.watres.2012.06.034. [DOI] [PubMed] [Google Scholar]
- Webb JM, Quinta R, Papadimitriou S, Norman L, Rigby M, Thomas D, Vay LL. The effect of halophyte planting density on the efficiency of constructed wetlands for the treatment of wastewater from marine aquaculture. Ecol Eng. 2013;61:145–153. [Google Scholar]
- Wu YY, Chen QJ, Chen M, Chen J, Wang XC. Salt-tolerant transgenic perennial ryegrass (Loliumperenne L.) obtained by Agrobacterium tumefaciens-mediated transformation of the vacuolar Na+/H+ antiporter gene. Plant Sci. 2005;169(1):65–73. [Google Scholar]
- Xiong JS, Ding J, Li Y. Genome-editing technologies and their potential application in horticultural crop breeding. Hortic Res. 2015;2:15019. doi: 10.1038/hortres.2015.19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang J, Kloepper JW, Ryu CM. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009;14(1):1–4. doi: 10.1016/j.tplants.2008.10.004. [DOI] [PubMed] [Google Scholar]
- Yates SA, Chernukhin I, Alvarez-Fernandez R, Bechtold U, Baeshen M, Baeshen N, Mutwakil MZ, Sabir J, Lawson T, Mullineaux PM. The temporal foliar transcriptome of the perennial C3 desert plant Rhazya stricta in its natural environment. BMC Plant Biol. 2014;14:2. doi: 10.1186/1471-2229-14-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan Z, Druzhinina IS, Labbé J, Redman R, Qin Y, Rodriguez R, Zhang C, Tuskan GA, Lin F. Specialized microbiome of a halophyte and its role in helping non-host plants to withstand salinity. Sci Rep. 2016 doi: 10.1038/srep32467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao P, Capella-Gutiérrez S, Shi Y, Zhao X, Chen G, Gabaldón T, Ma XF. Transcriptomic analysis of a psammophyte food crop, sand rice (Agriophyllum squarrosum) and identification of candidate genes essential for sand dune adaptation. BMC Genom. 2014;15(1):872. doi: 10.1186/1471-2164-15-872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou Y, Gao F, Liu R, Feng J, Li H. De novo sequencing and analysis of root transcriptome using 454 pyrosequencing to discover putative genes associated with drought tolerance in Ammopiptanthus mongolicus. BMC Genom. 2012;13:266. doi: 10.1186/1471-2164-13-266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu JK. Plant salt tolerance. Trends Plant Sci. 2001;6(2):66–71. doi: 10.1016/s1360-1385(00)01838-0. [DOI] [PubMed] [Google Scholar]