

The distribution of global high seas longline fishing is predictable across space and time using environmental variables.
Abstract
International interest in the protection and sustainable use of high seas biodiversity has grown in recent years. There is an opportunity for new technologies to enable improvements in management of these areas beyond national jurisdiction. We explore the spatial ecology and drivers of the global distribution of the high seas longline fishing fleet by creating predictive models of the distribution of fishing effort from newly available automatic identification system (AIS) data. Our results show how longline fishing effort can be predicted using environmental variables, many related to the expected distribution of the species targeted by longliners. We also find that the longline fleet has seasonal environmental preferences (for example, increased importance of cooler surface waters during boreal summer) and may only be using 38 to 64% of the available environmentally suitable fishing habitat. Possible explanations include misclassification of fishing effort, incomplete AIS coverage, or how potential range contractions of pelagic species may have reduced the abundance of fishing habitats in the open ocean.
INTRODUCTION
The high seas [or areas beyond national jurisdiction (ABNJ)] encompass more than 45% of the world’s surface area and 90% of the ocean’s volume. Before the 1950s, limitations in fisheries technologies predominantly restricted global marine fisheries to coastal and shelf waters. However, technological advancements after World War II, such as improved refrigeration, increased engine power, and acoustic sonars, prompted a rapid expansion of marine fisheries into ever more remote high seas waters (1). Consequently, high seas fisheries catch increased by 10-fold, from 450,000 metric tons (MT) in 1950 to about 6,000,000 MT by 2014 (2). As of 2015, high seas fisheries represented 6% of the global annual marine fisheries catch by mass and 8% by fishing revenue (3). Tuna and billfish make up the majority of the reported high seas catch by longliners and purse seiners and, by 2012, represented 9.3% of global annual marine fisheries catch (4, 5). This expansion also entailed novel impacts on oceanic and deep-sea systems (6, 7). While the importance of the high seas for the global seafood industry has continued to grow, the regulatory frameworks and monitoring mechanisms necessary to support their sustainable use have lagged (7).
The current governance frameworks for management of marine life in ABNJ were established in 1982 by the third United Nations Convention on the Law of the Sea and were further developed by the 1995 UN Fish Stocks Agreement (UNFSA) through the establishment and consolidation of regional fisheries management organizations (RFMOs). RFMOs have the legal responsibility to manage high seas fish stocks, but also nonfish species [UNFSA Article 5(g)], and biodiversity [UNFSA Article 5(f)]. The performance of these bodies in protecting biodiversity beyond their target commercial species has been questioned recently (8, 9). According to the UN Food and Agriculture Organization, migratory and straddling stocks harvested in ABNJ are overfished or are experiencing overfishing at twice the rate of stocks found within national waters (64% versus 28.8%)(4). A separate assessment of the status of the stocks managed by the world’s RFMOs concluded that 67% of these were either overfished or depleted (8) and that several of these have experienced range contractions due to overharvesting (10).
Some of the existing concerns about RFMO management include insufficient monitoring and weak implementation of ecosystem-based management measures due to the consensus-based RFMO governance process (9). As an example, the fisheries observer coverage of some pelagic longline fleets is as low ~5%, and can be even lower (11), which means that most longline fishing remains unmonitored. Novel forms of electronic monitoring help to address challenges related to the monitoring of catch and bycatch, reporting of fishing effort, and vessel distribution (12). These new technologies include vessel tracking systems such as the vessel monitoring system (VMS) or the automatic identification system (AIS), which can help with the surveillance and monitoring of marine fisheries (13, 14) even in remote waters. Not all vessels are required to carry AIS devices onboard, and regulations change between vessel type, size, and nationality as well as where vessels are fishing. For example, the United States requires that all self-propelled fishing vessels of 20 m or more in length must carry an AIS device onboard, but only while fishing in nearshore waters (Code of Federal Regulations, § 164.46). The International Maritime Organization (IMO) requires all passenger vessels or those larger than 300 gross MT to carry AIS devices. A growing number of programs have recently emerged using satellite-based AIS geolocation data to track and monitor fishing at sea. Some monitoring programs such as the Pew Charitable Trusts’ Eyes on the Sea project or the FISH-i Africa project (www.fish-i-africa.org) focus on identifying illegal and unreported fishing, while other programs such as Global Fishing Watch (GFW; www.globalfishingwatch.org) classify the behaviors of fishing vessels, providing open access data on the global distribution of fishing effort across the main gear types, and are continuously improving their ability to detect, classify, and quantify fishing effort estimates (12, 15).
Ecosystem-based fisheries management must address the impacts of fishing, such as habitat destruction and alterations of biological communities, via techniques that can monitor current activities and predict and manage future ones. Many of the management and conservation conflicts addressed by the ecosystem-based fisheries management framework are the result of a lack of information on the spatiotemporal distribution of resources and resource users. One mechanism to understand existing impacts on high seas biodiversity is to compare the distribution of fishing effort with bycatch species such as sharks (16) or seabirds (17). While these studies are very useful for gaining an initial understanding of the overlap of fisheries and their associated species (which RFMOs are required to manage), they are retrospective and do not capture the underlying dynamic oceanographic processes that result in the spatiotemporal overlap. To understand potential future interactions, mechanistic or correlative statistical models that explore the distribution or density of species in relation to environmental predictors are necessary [for example, (18–20)]. These studies have been conducted on many marine vertebrate species, yet very few studies (21) have analyzed the environmental correlates of human (for example, fisheries) distribution and attempted to understand their impacts through this lens. One example of such a study accurately predicted the distribution of fishing effort a year in advance (21). Fisheries managers can use information regarding the predicted distribution of fishing effort and bycatch species to provide information on the likely location of bycatch [for example, (22)], targeted observer coverage or enforcement (23), and partial closures or zoning (24). Further, models of the distribution of fishing effort can be run under different climate scenarios to help understand how fishing effort may shift in the future and affect fishing communities (25).
For instance, Kroodsma et al. (26) explore the influence of primary productivity and surface temperature estimates on the intensity of global fishing effort across multiple types of gear and flag States derived from AIS data. They conclude that other socioeconomic factors are much more influential in explaining the intensity of fishing effort than either environmental variable; however, they also note that longline fishing effort in the Indian Ocean was correlated with surface temperature (26). We consider that the lack of a global response of fishing intensity across gear types to either environmental variable is to be expected given the wide range of fishing strategies that were assessed jointly. We find that their conclusions of a correlation between longline fishing and temperature are evidence that, when assessed individually, individual gear types may show well-delineated responses to certain environmental predictors.
Here, we explore the spatial ecology and drivers of the global distribution of the pelagic longline fishing fleet in the high seas by creating environmental predictive models of fishing distribution from satellite-based AIS data from GFW. We build environmental niche models using a boosted regression tree (BRT) modeling approach that relates the location of fishing events to different environmental conditions and compares them to a set of pseudoabsence points (areas of no observed fishing). By comparing the conditions where fishing was observed to locations where fishing was not observed, we hope to decipher which environmental conditions seem to be preferred by longliners. Our model used 14 environmental variables: sea surface temperature (SST), temperature at 400 m (T400), turbulent kinetic energy (TKE), particulate organic carbon (POC), net primary productivity (NPP), mixed-layer depth (MLD), surface oxygen concentration (SOC), oxygen concentration at 400 m (O400), sea surface salinity (SSS), salinity at 400 m (S400), euphotic depth (ZEU), bathymetry (BATH), distance to continental shelf (DCS), and distance to seamount (DSM). By applying concepts that were originally developed to explore the ecological niches of terrestrial and marine animals, we aim to better understand the environmental preferences of longline fishing fleets in ABNJ and to shed light on the factors shaping their distributions at large scales, which opens new avenues for predictive forecasting of future spatial patterns of global longline fishing effort and concomitant stresses on the high seas. We are aware that other socioeconomic factors play important roles in the decision-making process of marine fishing activities and hope that our analysis informs future work that also includes these variables.
RESULTS
High seas longline fleet composition and distribution
After analyzing all satellite-based AIS fishing effort data from GFW, we found that longline fishing effort in the high seas accounted for 84 to 87% of the fishing effort (by hour) across gears during the study period (fig. S1). While longline fishing effort is lower in ABNJ, it represents a major top-down pressure on oceanic ecosystems (27). Of the high seas longline fishing effort, 88.9% (2015) and 80.4% (2016) were attributable to five fishing States or territories: China, Japan, South Korea, Spain, and Taiwan (fig. S2). Taiwan dominates global longline fishing effort (by hour) in the high seas, followed by Japan, Spain, China, and South Korea. Our analysis focuses on these top-five fishing States or territories. AIS-derived fishing effort data show that the distribution of longline fishing effort in the high seas changes across space (Fig. 1) and time (Fig. 2). During 2015 and 2016, the tropical (23.5°N to −23.5°S) and temperate (66.5°N to 24.5°N and −24.5°S to −66.5°S) regions contained 64.6 and 35.3% of the global fishing effort, respectively. On average, the intensity of fishing effort in the high seas is higher during the boreal summers and peaks in July and August during 2015 and 2016, respectively (Fig. 2). The overall increase in fishing effort data between years is likely driven by an increase in the number of orbiting satellites capable of detecting AIS signals, as well as an increase in the capability of detecting and classifying longline fishing effort by the GFW group. Despite the increase in fishing effort intensity, the seasonal pattern where global longline fishing effort increases during the boreal summer months seems to be preserved between the two years. Untangling the drivers of the observed seasonal patterns of fishing effort requires a regional, fleet-specific approach that includes information about target species, fishing seasons, and quotas. All fishing effort data needed to evaluate the conclusions in this paper are available from GFW.
Fig. 1. Distribution of global pelagic drifting longline fishing in ABNJ in 2015 and 2016.
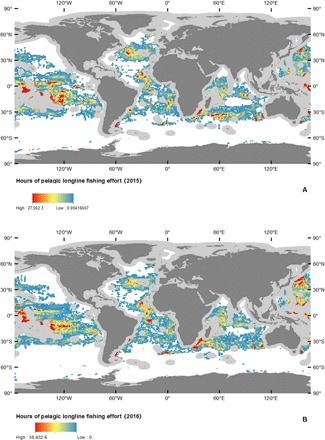
(A) 2015. (B) 2016. Light gray areas depict exclusive economic zones (EEZs) that were excluded from this study. Fishing effort (hours) as calculated by GFW using satellite-based AIS data. Given the differences in quantified fishing effort between 2015 and 2016, the scales were maintained separate to showcase how, despite changes in intensity, the main trends in longline fishing effort are maintained. Gray areas around coastlines depict EEZs excluded from this study. Data are from GFW.
Fig. 2. Monthly distribution of pelagic longline fishing effort in ABNJ by the top five fishing States or territories, and all other countries combined.
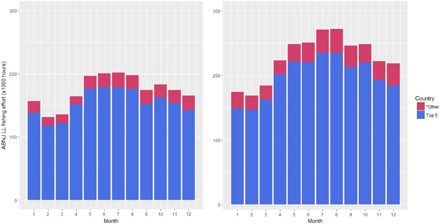
The total calculated fishing effort between the years increases between 2015 and 2016, with China and Taiwan experiencing the largest increases in quantified fishing effort. ”*Other” represents a total of 45 other fishing nations deployed longline (LL) gear in ABNJ between 2015 and 2016.
Model performance and prediction
We assessed the accuracy of our models using various metrics that measured the degree to which we can predict the raw fishing effort observations using our environmental suitability models. Our results demonstrate how the global distribution of longline fishing effort in the high seas can be predicted with high levels of accuracy across months and years using BRTs to explore the environmental conditions in which fishing observations occurred. By comparing four different model performance metrics across years and threshold types (n = 16), we were able to determine that predictions from monthly models outperformed the temporally averaged model that used the data across all months. Using a Wilcoxon signed-rank test, we found statistically significant differences in the distribution of the accuracy metrics in 14 of the 16 model performance comparisons (table S5), and the average performance scores were superior for the monthly model in 15 of the 16 cases. The predictive accuracy and correct classification scores were high throughout the entire period studied (figs. S3 and S4 and tables S1 and S2) and support the use of environmental modeling for understanding the spatial patterns and environmental drivers of the human fisheries footprint in the ABNJ. We found that the predictive accuracy of the monthly environmental niche models after projecting them onto future environments (1, 6, and 12 months in advance) remained high. The mean predictive accuracy was lowest for the 6-month prediction (~74%), and both the 1- and 12-month predictions showed similar mean accuracy rates at ~82% (fig. S5).
The model outputs were projected onto geographic surfaces, where the likelihoods of observing longline fishing effort were displayed as probability estimates between 0 and 1. Post-processing of the model outputs required a definition of “suitable fishing habitat,” which was done by selecting a probability cutoff value threshold for each map. Lower thresholds will classify more areas of the study region as suitable, while higher thresholds will be more restrictive. Here, we assessed the implications of applying two different types of threshold on our model outputs: receiver operator characteristic (ROC) curve and mean probability distribution (MPD) thresholds. Our results show how the choice of threshold provides slightly different estimates of longline suitable fishing habitat, although the differences in model performance are small (tables S1 and S2 and figs. S3 and S4). The thresholds derived from the ROC curve resulted in higher cutoff values, while the MPD thresholds were lower on average. Higher thresholds minimized the overprediction of suitable fishing grounds, while lower thresholds resulted in higher classification rates of observed fishing effort; this is reflected in the specificity and sensitivity values presented in the Supplementary Materials (tables S1 and S2).
Monthly persistence maps provide a visual representation of the global changes of fishing habitat suitability throughout the year (Figs. 3 and 4) and help identify areas of the high seas where favorable environmental conditions for longline fishing are most stable. The monthly persistence maps also help identify areas of the high seas that are not classified as environmentally suitable for longline fishing throughout the year, which provides valuable information about which areas may be experiencing less longline fishing pressure. The variability of environmental suitability to fishing in the high seas was assessed by mapping the average coefficient of variation of predicted high seas fishing suitability for each year (Fig. 5), which combines the 24 monthly predictions and identifies the areas where we can expect the highest changes in suitability. Tropical latitudes were found to be the most stable year-round fishing grounds after assessing both persistence maps and estimates of variability.
Fig. 3. The monthly persistence of suitable habitat in ABNJ for 2015.
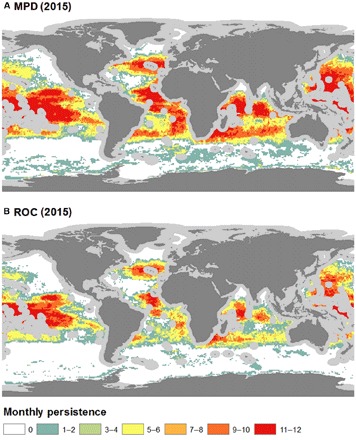
These persistence estimates were calculated using two different probability distribution cutoff thresholds: (A) MPD and (B) ROC. Gray areas around coastlines depict EEZs excluded from this study. Data are from GFW.
Fig. 4. The monthly persistence of suitable habitat in ABNJ for 2016.
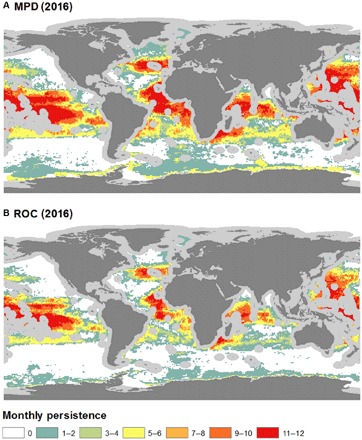
These persistence estimates were calculated using two different probability distribution cutoff thresholds: (A) MPD and (B) ROC. Gray areas around coastlines depict EEZs excluded from this study. Data are from GFW.
Fig. 5. The average coefficient of variation of predicted high seas fishing suitability for 2015 and 2016.
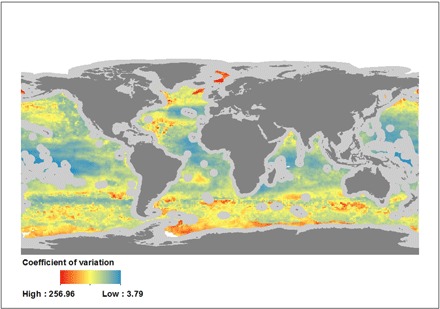
Tropical latitudes show, on average, more predictive stability throughout the study period, whereas temperate and subpolar waters show higher degrees of variability of suitable habitat. Gray areas around coastlines depict EEZs excluded from this study. Data are from GFW.
We used the binary estimates of suitable fishing habitat to calculate the proportion of the predicted fishing grounds where longliners were observed to obtain estimates of the global suitable fishing habitat that is occupied (table S6). These estimates are important to understand the realized niche of global longliners, that is, the amount of suitable fishing habitat that is actually fished. Results show how the global fleet is not occupying large proportions of the fishing grounds that our models classify as potentially environmentally suitable for fishing. In 2015, the average proportion of occupied suitable fishing habitat was estimated to be 38 to 55%, whereas estimates were slightly higher for 2016 at 47 to 64%; the differences within each year were partly related to the choice of threshold. We briefly explored the distribution of false-negative and false-positive classification for two months (January and July) in 2015 to explore potential seasonal effects or patterns and the influence of using different cutoff thresholds (fig. S6). We consider that, while there might be a slight seasonal effect on the distribution of unfished areas that were classified as suitable—higher latitudes earlier in the year and lower latitudes in the boreal summer months—the distribution of false negatives (unsuitable fished habitat) did not seem to follow any patterns associated with latitude, longitude, or environmental gradients when these areas were overlaid with various environmental predictors, including SST, T400, O400, and DCS.
Environmental predictors of fleet distribution
The relative explanatory variable importance (VI) of the environmental variables used in the BRT models fluctuates on a monthly and interannual basis (Fig. 6 and tables S7 and S8), with different environmental variables explaining the distribution of fishing effort during different times of the year. The VI scores obtained from the monthly models (Fig. 6) show (i) how the environmental preferences of the high seas longline fleet can be characterized by a few environmental variables, namely, SST, DCS, T400 and O400; and (ii) how the correlates of fishing effort distribution show both intra- and interannual variability. The four variables shown to be consistently important throughout the year had annual average VI scores >10. Our results also showed how other environmental predictors with lower average VI scores (that is, NPP, SOC, POC, MLD, and S000) gain importance during certain times of the year (figs. S3 and S4), although these may be difficult to interpret given their weak signals.
Fig. 6. Radar plots of the average quarterly VI scores in 2015 and 2016.
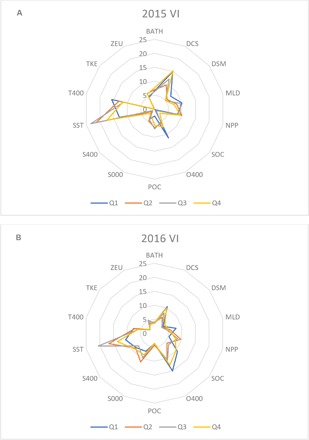
(A) 2015. (B) 2016. The monthly VI scores for each of the two years assessed were averaged by quarter (Q) to capture the seasonal changes in the importance of each of the environmental predictors: Q1, January–March; Q2, April–June; Q3, July–September; Q4, October–December.
We further explored the explanatory power of static and dynamic predictors by comparing various iterations of the 2016 monthly models by using (i) only static predictors, (ii) only dynamic predictors, and (iii) a combination of the two. The model performance metrics (table S9) demonstrate how the model that included both static and dynamic predictors outperformed both the static and dynamic models. The performance of the model with static predictors was worse than that of the other two.
We explored the preferences of the longline fishing fleet further by assessing graphical visualizations of how increases or decreases in an environmental variable (for example, higher or lower temperatures) affect the probability of longline fishing; these figures are termed partial dependence plots (figs. S7 and S11). For instance, the relationship between longline fishing and SST, which was the most important environmental predictor in both years, shows a two-state response, where higher temperatures lead to higher suitability between January and February, and then progressively transitions to a response favoring a broader range of temperatures (including ~15° to 20°C surface waters). This second temperature state is most apparent in the months of June and July. We assessed the partial dependence plots for the environmental predictors that appeared to be persistently important throughout 2015 and 2016: SST, O400, T400, and DCS.
DISCUSSION
Here, we demonstrate how environmental niche models can be used to explain and predict the distribution of longline fishing effort in the high seas. What ecological niches do these new “fisheries apex predators” occupy in the high seas? Studying fisheries using analyses similar to those used to study marine animals has been suggested previously (28) and may provide opportunities to understand and predict the dynamics of fishing fleets. We suggest that models like the ones presented here be used in management to (i) identify likely areas of interaction between fisheries and bycatch species, allowing for spatial management approaches to be used to mitigate interactions; (ii) anticipate changes in the distribution of fishing effort by using the existing model output to predict fishing effort 12 months out or by running this type of model under various climate scenarios as has been done for many species (18); and (iii) better focus monitoring and surveillance efforts of longlining in open-ocean pelagic environments within EEZs by directing authorities to areas of likely fishing (23), and should be applicable to similar management of living marine resources in the high seas.
Model accuracy
The high model performance metrics (figs. S3 and S4 and tables S1 and S2) demonstrate how the distribution of pelagic longline fishing effort in the high seas is environmentally structured and can be explained and projected using predictive models combined with information on the environment surrounding fishing observations. The mean accuracy values (0.84) and area under the curve (AUC) scores (0.86) throughout the study period were high and exhibited slight average increases in 2016. Improvements in the availability of fishing effort data due to increased numbers of AIS-capable satellites launched over the past years as well as increased accuracy of the neural net detecting and classifying longline fishing effort due to more data available may have contributed to this slight improvement. The average correct classification (sensitivity) scores for both years were also high (0.93) and showed slight increases (6 to 11%) in correct classification when applying an MPD threshold. Conversely, the average false-positive classification (specificity) values were lower, suggesting that our model can correctly classify most of the observed fishing effort observations but slightly overpredicts fishing in some areas where no fishing effort was observed. The small differences in sensitivity and specificity scores between cutoff thresholds were expected as the MPD thresholds were, on average, lower than the thresholds derived from ROC curves. Lower probability distribution thresholds translate to larger areas of the high seas being classified as suitable for longline fishing effort, thus capturing more observed presence points (explaining the higher sensitivity scores) and including more areas with no fishing observations (which explains the higher overprediction of fishing or false positives). Regardless of the choice of threshold, our BRT models were able to explain most of the high seas longline fishing effort observations, which we consider to be a meaningful step toward understanding the current and future human use of the high seas. Potential explanations for the overprediction of the models into areas where no fishing effort was observed follow later in the discussion.
Variability and persistence of suitable fishing habitat
Through mapping the habitat suitability of the longline fleet in ABNJ across months, we identify areas of the ocean with higher intra-annual variability of environmental suitability for fishing; these predominantly occur in the peripheries of the more stable year-round fishing grounds. These areas of high intra-annual variability may correspond to waters where oceanographic conditions show strong seasonal variability throughout the year, such as boundary currents in the peripheries of oceanic gyres. The latitudinal poleward spread of some variables—such as surface temperature or dissolved oxygen, which decreases and increases, respectively, as you move away from the equator—during the boreal and austral summers likely causes the temporary increase in suitability of areas within the temperate and subpolar latitudinal bands. As previously mentioned, some target species show stable north-south seasonal movements, which correspond well to the seasonal increases in habitat suitability in those waters.
Persistent suitable habitat for longline fishing (that is, areas suitable for 6 to 12 months) is contained within the tropical and temperate latitudes, though there are longitudinal differences (Figs. 3 to 5) and the lower latitudes are the most stable and hold most of the persistent suitable habitat. These areas seem consistent with the global latitudinal habitat preferences displayed by the top-six tuna target species (29, 30), which are among the main target species of longliners in ABNJ. However, further work is required to assess the degree of overlap by longline fisheries and target, both in geographic space and in environmental space; this may help better understand the areas of high persistence of fishing suitability seen in Figs. 3 and 4.
Environmental predictors of fleet distribution
Tuna and billfish species comprise 81.2% of the global longline landing estimates in ABNJ between 1950 and 2014 (2). While there seems to be a clear geographic overlap between the preferred habitat of the main tuna species and suitable fishing habitat for pelagic longliners in the high seas, our models also offer the opportunity to compare their environmental preferences. Our results suggest that longliners in the high seas show similar preferences to those of the species they are targeting. In 2015, the fleet showed strong preference for areas where the temperature at 400-m depth was between 8° and 18°C (fig. S8); this preference was stable throughout the year and consistent with the temperature preferences of some commercially exploited tunas (31). The response of fishing to different oxygen concentrations at 400-m depth (fig. S10) shows that longline fishing effort in ABNJ is more commonly found in waters where the dissolved oxygen concentration at that depth is between 1 and 5 ml O2 liter−1; this is also consistent with findings on the physiological preferences and thresholds of tuna. For instance, studies have found oxygen concentration tolerances as low as 3.5 ml O2 liter−1 for skipjack tuna (Katsuwonus pelamis) and yellowfin tuna (Thunnus albacares) and 1.5 ml O2 liter−1 for bigeye tuna (Thunnus obesus) (32). We consider that the response to the DCS variable is partially masked by the 200-nm jurisdictional buffer that was used to exclude any within-EEZ fishing in this study; a separate analysis including coastal fishing may be required to interpret the true influence of this variable on longline fishing effort distribution. From the DCS partial dependence plot (fig. S9), we can infer that the probability of fishing in the high seas increases with DCS during some months of the year, which is likely driven by the high amounts of longline fishing in the central Pacific Ocean. The overall preference for warmer waters described by the model likely results from the fact that many of the species targeted by the global pelagic longline fleet year-round are tropical or subtropical (for example, T. albacares or T. obesus). These results (figs. S7 and S11) agree with those of Kroodsma et al. (26), who found that longline fishing effort intensity was highest between the 16° and 19°C isotherms. The bimodal response to SST that can be observed during the boreal summer months may be caused by the northward movement of some targeted species into more temperate waters during these months, as seen in swordfish (Xiphias gladius) (33) or Atlantic bluefin tuna (Thunnus thynnus) (34). These findings are aligned with the conclusions of the study by Arrizabalaga et al. (30), where sea temperature and dissolved oxygen were the variables that explained the most deviance when modeling tuna logarithmic catch per unit effort. Despite differences in the extents between our studies, as well as some variations in the environmental variables, the similarities in the environmental preferences of tunas and those displayed by longliners suggest that the high seas longline fleet is tracking many of the same environmental cues as their main target species. Given the wide range of species targeted by the global pelagic longline fleet, the environmental preferences of the fleet are not expected to bear exact resemblance with any one given taxon. Further, the high seas longline fleet may not have a static environmental niche but instead may adapt its distribution and “environmental preferences” to maximize overlap with its multiple target species throughout the year, in a potentially consistent and predictable manner.
Given the global scale of our analysis, the spatial and temporal resolution of our predictors was coarse, which limited our ability to capture the influence of mesoscale oceanographic features, such as oceanic frontal zones and eddies, on the distribution of fishing effort. However, given the spatial scale of our study, we consider that our models successfully explain the broad environmental patterns that shape the distribution of the global longline fishing fleet in the high seas. We also see great promise in the use of environmental niche models for predicting the distribution of future fishing effort, which could bring us a step closer to designing and implementing precautionary spatiotemporal management measures based on future oceanographic conditions. While these efforts would have to be tailored for specific regional fleets, the promising predictive accuracy estimates that we obtained from our models indicate that estimating the future distribution of fishing pressure based on oceanography is likely feasible.
Interpreting low occupancy rates
Our models predict significantly more area as being suitable for fishing than was observed to have fishing effort. The unfished parts of ABNJ that were classified as suitable fishing grounds are not closed to fishing by RFMOs and are not further away from commercial fishing ports than those areas fished in ABNJ. While these areas could be the result of classification errors in our models, ecological theory provides an alternative explanation: decreased occupancy by target species of their fundamental niche. Multiple factors may influence whether a species occupies its fundamental niche, including limitations to dispersal, predator avoidance, exclusion by interspecific or intraspecific competition, or lack of resources. Our results show that the average proportion of fishing ground occupancy for high seas longliners fluctuated by year and threshold method, with a maximum of 55 and 64% in 2015 and 2016, respectively. Just as an animal would avoid habitats with limited resources, unfished areas with appropriate environmental conditions may be avoided by the high seas pelagic longline fleet due to reduced overlap with target species or insufficient abundances despite environmentally suitable conditions. A recent study demonstrated how several pelagic target species have experienced contractions in their ecological range because of decreasing abundance (10). It is therefore conceivable that environmentally suitable fishing areas are avoided by longliners given changes in the distribution and abundance of pelagic target species, such as the range loss in the South Atlantic seen for bluefin tuna (10). Have the last six decades of pelagic fishing led to the overfishing of a significant proportion of the suitable fishing grounds in ABNJ? Or are longline fleets just following changes of prey abundance influenced by other factors such as decadal oscillations or climate change? Limitations in fleet capacity or fuel cost seem unlikely reasons to explain the absence of fishing effort in 36 to 62% of suitable fishable habitat in ABNJ, as the high seas fishing fleet is supported by subsidies and the extent of their distribution suggests that no region of the ocean is too distant to be fished. We run a linear regression between the intensity of longline fishing effort and DCS and found no correlation between the two (fig. S12); this suggests that any lack of fishing is not due to the remoteness of those areas despite the high fuel costs. The suite of explanatory variables used in our models is limited to the biophysical and physiographic dimensions, and therefore, do not take into consideration socioeconomic factors that may be crucial for explaining observed patterns of fleet distribution. Fishermen are not subject to the same physiological and dispersal limitations as are marine species; their limitations are more likely to be political and economic ones. The distribution of high seas longliners is therefore likely to also be influenced by socioeconomic factors, including, but not limited to, catch quotas, market prices of commercial species, fleet communication, or selection of landing sites. Regional bioeconomic models of the distribution of fishing effort could be used to understand differences in the drivers of distributions of fishers and nontarget species and allow for the development of dynamic management measures based on the environmental and economic correlates (33); we believe that our models could represent a meaningful component of these wider frameworks.
Additional factors may explain the lack of observed occurrence of fishing activities in areas predicted to be suitable for fishing, including (i) misclassification and thus missed observations of longline vessels or fishing effort, (ii) poor spatiotemporal satellite coverage, (iii) intentionally switching AIS transponders off, and (iv) fishing events by fishing States or territories not included in our analysis. We addressed the last factor by assessing how much of the unfished suitable habitat may have been fished by longliners from the 26 States or territories not included in our analysis. We found that none of these countries fished the unoccupied suitable fishing areas; most (65%) of the 1267 fishing events by these fleets were predicted by our model. We think that a more detailed understanding of the behavior of fishing vessels out at sea may help us identify the general areas of expected fishing vessel activity, thus bringing us one step closer to abating illegal fishing by designating enforcement resources more strategically.
Reflections on the use of predictive models of fishing effort
While the distribution of some sectoral activities, such as deep-sea mining or oil and gas exploration, is well mapped and static, the dynamism of open-ocean fisheries makes understanding their spatiotemporal distribution difficult and, consequently, monitoring, control, and surveillance a challenge. The models we present could become a useful tool for managers to focus their efforts on areas of likely fishing activity. As high-resolution satellite imagery is increasingly being used to look for fishing vessels, narrowing the areas of the ocean where there is a high portability of fishing activities may help streamline the process of detecting legal or illegal fishing vessel activity. The spatial and temporal resolution of our predictors was coarse, which limited our ability to capture the influence of mesoscale oceanographic features, such as oceanic frontal zones and eddies, on the distribution of fishing effort. However, given the global scale of our analysis, we consider that our models successfully explain the broad environmental patterns that shape the distribution of the global longline fishing fleet in the high seas.
We see great potential in improving the predictive ability of global longline fishing effort models through the development of future region-specific models that capture the regional relationships between fishing effort and environment, and predator-prey dynamics in more detail. Additionally, partitioning the niche models by fishing State may also be required to tease apart distinct environmental correlations and behaviors, leading to more accurate predictions. As the spatiotemporal coverage of satellites capable of recording AIS signals improves and fishing effort classification algorithms become more accurate, so will the estimates of fishing effort between years, and behavior classification errors will decrease. Furthermore, with more years of data and improving spatiotemporal satellite coverage, the differences in environmental preferences between years will be attributable more to changes in fleet behavior than to biases in fishing effort observations.
CONCLUSIONS
As we combine an improved understanding of open-ocean fleet behavior with knowledge of the drivers of distribution of target and nontarget marine taxa, our ability to predict the co-occurrence of fishing with sensitive species or ecosystems will improve, as will the efficacy of related management measures. As the intensity and overlap of human uses of ABNJ continue to grow, ocean governance structures will have to rely more heavily on different forms of dynamic spatial management to accommodate all users and activities (35), which, in turn, rely on open-access remote sensing data and collaborations between researchers, fishers, and the management community (36). Our research demonstrates how the global pelagic longline fleet exhibits predictable environmental preferences for various biophysical and physiographic predictors, which can be used to explore the current and future distributions of fishing fleets. Improvements in remote sensing and oceanographic forecasting for variables (for example, SST) open new opportunities for the implementation of adaptable ocean management measures that match the dynamics and distributions of ocean biological resources and resource users. As we grapple with rapidly changing oceans and ocean uses, advancements in predictive modeling, aided by new technologies, will help us move away from reliance on retrospective tactics in area-based management and toward more dynamic approaches capable of delivering ecosystem-based management.
MATERIALS AND METHODS
Here, we used a form of classification model known as BRT to characterize the distribution of longline fishing in the high seas (as reported by GFW) from environmental variables primarily obtained from increasingly available remote sensing sources (36). We used the processed AIS geolocation data from GFW in the form of gridded fishing hour estimates for 2015 and 2016 as observations to fit the environmental niche models.
AIS fishing effort data
GFW analyzes and provides online interactive maps of the behavior of fishing vessels from global AIS and VMS data. AIS was originally designed as a tool to avoid collisions at sea as part of the IMO Safety of Life At Sea Treaty [SOLAS Treaty, Chapter V; (37)]. Vessels equipped with an AIS transponder signal their position and vessel identification data such as IMO number, maritime mobile security information number, call sign, ship type, speed and course over ground, and other information to ships nearby carrying the transponders as well as to receiving ground stations and low-orbit satellites. Signal transmission frequencies vary with speeds between a few seconds and a few minutes. These high-resolution tracking data are then analyzed by GFW to assess ship movements and behavior, using neural network algorithms and logistic models to classify different fishing gear types as well as the points in space and time where individual vessels deploy their fishing gear (16). Data used for this study were derived from the logistic regression model 1.1 (http://globalfishingwatch.io/fishing__logistic_1_1.html). It is worth noting, however, that GFW only uses satellite-based AIS data, which have limitations such as a maximum number of individual signals that can be detected simultaneously, heterogenous satellite spatiotemporal coverage, or gaps near coastlines, where shore-based stations receive the signal that the satellite can no longer detect. It is unlikely for areas in the high seas to experience satellite channel saturation, and vessel AIS signals are also unlikely to be detected by shore-based stations. Fishing effort is detected and calculated, as hours of fishing, for individual fishing gear types: (i) pelagic longlines, (ii) trawls, (iii) purse seines, (iv) fixed gear, and (v) other types of fishing gear. However, each of these is subject to behavior classification errors. For this study, estimates of global pelagic fishing effort for the years 2015 and 2016 were extracted from the GFW database, including vessels from 114 countries and territories.
We filtered the GFW fishing effort estimates spatially to only include longlining events in the high seas. Within the high seas, fishing effort by pelagic longliners accounted for 88.9 and 80.4% of the quantified fishing effort (hours) in ABNJ across all gear groups in 2015 and 2016, respectively (fig. S1). The dominance of longline fishing effort in ABNJ and its known negative impacts on multiple nontarget species (38) underscore the importance of understanding the potential drivers of its global distribution (Fig. 1). Hence, we focused on longlines only in our modeling efforts, particularly the distribution of fishing events rather than fishing intensity.
According to GFW fishing effort estimates, 45 to 50 fishing States and territories deployed longlines in ABNJ throughout 2015 and 2016. We refined the list of countries to only include those that accounted for the >80% of the observed fishing effort; this reduced the list of fishing States and territories to five (fig. S2). We further selected the fishing effort data used to build the environmental niche models by only including these five major fishing States and territories. The fishing effort applied by these countries was aggregated spatially to 1° by 1° cells for 2015 and 2016 (Fig. 1) given the global extent of the analysis (39) and then partitioned temporally into 24 months (Fig. 2). Environmental data layers specific to each month were then used to run each of the 24 monthly environmental niche models. The use of monthly averages and monthly climatologies for certain environmental variables inevitably resulted in the loss of some fine-scale environmental features (for example, mesoscale oceanic eddies and frontal zones) that may influence the distribution of fishing effort at submonthly time steps. Future analysis at finer spatiotemporal resolution may allow the inclusion of more information on dynamic oceanographic features. For the purpose of this study, we focused on the monthly environmental variability on the distribution of fishing effort.
Environmental predictor variables
The environmental variables selected for modeling the ecological niche of global longline fishing effort in ABNJ included both static (physiographic) and dynamic predictors (biophysical). Various habitat-modeling studies support the inclusion of biophysical and physiographic predictors across spatial and temporal scales for studying the ecology of species of commercial interest (30, 33). The dynamic variables were extracted by month and consisted of SST, T400, TKE, POC, NPP, MLD, SOC, O400, SSS, S400, and ZEU. The static physiographic variables included BATH, DCS, and DSM. All variables were extracted at 1° by 1° spatial resolution cells or aggregated as necessary and had different temporal resolutions; some are monthly estimates while others are climatological (that is, averages for the month across many years; see table S9).
Environmental niche model fitting, validation, and projection
All BRTs were fitted to the fishing effort data using RStudio, a development environment for the open-access statistical software R. The models were fitted to the number of monthly fishing effort presence points derived from GFW estimates and double the number background (pseudoabsence) points from the high seas region; a low number of points is recommended for modeling approaches such as BRTs (40). Background points were created on a monthly basis by randomly selecting from the unfished areas of ABNJ (tables S11 and S12). Randomly selecting background pseudoabsence points from anywhere in the high seas, including polar and subpolar regions, where almost no longline fishing effort occurs, biased the results and exaggerated the importance of latitudinally structured variables such as SST and SOC (fig. S13). The distribution of background pseudoabsence points was therefore constrained to areas that had SST values within the observed temperature range of observed fishing. In addition to the 24 monthly models, we computed a temporally averaged model that included the data from all 24 months to assess which of the two approaches performed better. After fitting the classification models, model outputs were mapped onto geographic space by projecting them using layers of the same environmental predictors. The resulting two-dimensional map represents a probability distribution surface where each grid cell in ABNJ was assigned a value between 0 and 1. Confusion matrices were then computed to assess how well each of the monthly models could predict the distribution of longline fishing effort. Various model performance indices were calculated, including the AUC, κ statistic (a measure of categorical agreement describing the difference between the observed and chance agreements), sensitivity (the proportion of actual presence that is accurately predicted), specificity (the proportion of actual absences that are accurately predicted), and accuracy values (tables S1 to S4 and figs. S3 and S4). We then used a nonparametric Wilcoxon signed-rank test to assess whether the performance metrics of the monthly models are statistically dissimilar from those of the temporally averaged model (table S5).
We also explored the explanatory accuracy of the monthly models at predicting the distribution of future fishing effort by projecting monthly models onto the oceanographic conditions 1 (n = 23), 6 (n = 180), and 12 (n = 12) months in advance and assessing how accurately we could predict the distribution of observed longline fishing effort in those months (fig. S5). We further explored the influence of the environmental variables by running two additional monthly models for all the months of 2016, one of which only included static (n = 3) variables and the other was run using only dynamic variables (n = 11) (table S10).
There are multiple possible approaches for selecting a probability distribution threshold to convert probability maps into binary maps. Here, we explored the influence of two separate methods of selecting thresholds for obtaining binary habitat suitability maps. Areas with a monthly probability distribution above the set threshold were considered as suitable habitat for the studied organism. First, we calculated monthly thresholds based on ROC curves, which show the relationship between the true-positive (sensitivity) and false-positive (specificity) rates. The second type of threshold that we calculated was based on the MPD of the monthly models. While both methods are widely accepted procedures (41) for establishing cutoff threshold values, the resulting binary habitat suitability landscapes can differ, and results must not be interpreted as final, but instead as different scenarios of pelagic longline fishing suitability in the high seas. Additional information about how BRTs were fitted and projected is available in Supplementary Materials and Methods.
Supplementary Material
Acknowledgments
This study is a product of the Nippon Foundation Nereus Program. We thank the GFW partnership for their technical support and for providing the global fishing effort estimates that we used in this study. They were an indispensable asset to this study. We also thank Dalhousie University, in cooperation with B.W., D.P.T., and K.B., for hosting a workshop in May 2017 that helped shape some of the ideas and analyses presented here. Funding: G.O.C., D.C.D., and P.N.H. received financial support from the Nippon Foundation Nereus Program to lead this research. K.B., D.P.T., and B.W. acknowledge support from the Google Earth Engine Research Award for the funding opportunity to convene a workshop that was instrumental for structuring the study. Author contributions: All authors were engaged in the intellectual design of the project, and all participated in editing the manuscript. G.O.C., D.C.D., and P.N.H. coordinated the research effort. G.R. and K.B. helped with the analysis and pre-processing of the GFW data. Competing interests: The authors declare that they have no competing interests. Data and materials availability: All data needed to evaluate the conclusions in the paper are present in the paper and/or the Supplementary Materials. Additional data and code related to this paper may be requested from the authors or the GFW portal (www.globalfishingwatch.org).
SUPPLEMENTARY MATERIALS
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/4/8/eaat3681/DC1
Supplementary Materials and Methods
Fig. S1. The proportion of 2015 and 2016 fishing effort (hours) in ABNJ by gear.
Fig. S2. The proportion of pelagic longline fishing effort attributed to the main fishing States or territories.
Fig. S3. Accuracy values obtained for the 2015 and 2016 monthly boosted regression tree models after applying an ROC threshold.
Fig. S4. Accuracy values obtained for the 2015 and 2016 monthly boosted regression tree models after applying an MPD threshold.
Fig. S5. The predictive accuracy of the monthly BRTs after projecting them onto future environments.
Fig. S6. Distribution of predicted and observed fishing effort in January and July of 2015 using different thresholds: ROC and MPD.
Fig. S7. The SST partial dependence plots from the monthly 2015 models.
Fig. S8. The temperature at 400-m partial dependence plots from the monthly 2015 models.
Fig. S9. The DCS partial dependence plots from the monthly 2015 models.
Fig. S10. The oxygen at 400-m partial dependence plots from the monthly 2016 models.
Fig. S11. The SST partial dependence plots from the monthly 2015 models.
Fig. S12. The distribution of fishing effort intensity as a function of the Euclidean distance (kilometers) to the continental shelf.
Fig. S13. Monthly variable importance scores for boosted regression trees using background pseudoabsence points from the entire high seas areas for 2015 and 2016.
Table S1. Various model performance indices of the monthly BRTs for 2015 and 2016.
Table S2. Various model performance indices of the monthly BRTs for 2015 and 2016.
Table S3. Various model performance indices of the temporally averaged BRT model.
Table S4. Various model performance indices of the temporally averaged BRT model.
Table S5. Results from the Wilcoxon signed-rank test comparing the performance of monthly models to the temporally averaged model.
Table S6. Amount of fundamental niche occupied by pelagic longliners.
Table S7. The 2015 VI scores.
Table S8. The 2016 VI scores.
Table S9. Average 2016 model performance metrics using different environmental variables.
Table S10. Description of the variable type and source for each of the 14 biophysical and physiographic predictors.
Table S11. The number of presence and pseudoabsence points in 2015.
Table S12. The number of presence and pseudoabsence points in 2016.
REFERENCES AND NOTES
- 1.Swartz W., Sala E., Tracey S., Watson R., Pauly D., The spatial expansion and ecological footprint of fisheries (1950 to present). PLOS ONE 5, e15143 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Sea Around Us Concepts, Design and Data, D. Pauly, D. Zeller, Eds. (2015); www.seaaroundus.org.
- 3.Sala E., Mayorga J., Costello C., Kroodsma D., Palomares M. L. D., Pauly D., Sumaila U. R., Zeller D., The economics of fishing the high seas. Sci. Adv. 4, eaat2504 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.United Nations Food and Agriculture Organization (FAO), The State of World Fisheries and Aquaculture 2014 (FAO, 2014). [Google Scholar]
- 5.Juan-Jordá M. J., Mosqueira I., Cooper A. B., Freire J., Dulvy N. K., Global population trajectories of tunas and their relatives. Proc. Natl. Acad. Sci. U.S.A. 108, 20650–20655 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Clark M. R., Althaus F., Schlacher T. A., Williams A., Bowden D. A., Rowden A. A., The impacts of deep-sea fisheries on benthic communities: A review. ICES J. Mar. Sci. 73, i51–i69 (2015). [Google Scholar]
- 7.Druel E., Gjerde K. M., Sustaining marine life beyond boundaries: Options for an implementing agreement for marine biodiversity beyond national jurisdiction under the United Nations Convention on the Law of the Sea. Mar. Policy 49, 90–97 (2014). [Google Scholar]
- 8.Cullis-Suzuki S., Pauly D., Failing the high seas: A global evaluation of regional fisheries management organizations. Mar. Policy 34, 1036–1042 (2010). [Google Scholar]
- 9.Gilman E., Passfield K., Nakamura K., Performance of regional fisheries management organizations: Ecosystem-based governance of bycatch and discards. Fish Fish. 15, 327–351 (2014). [Google Scholar]
- 10.Worm B., Tittensor D. P., Range contraction in large pelagic predators. Proc. Natl. Acad. Sci. U.S.A. 108, 11942–11947 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Allain V., Nicol S., Polovina J., Coll M., Olson R., Griffiths S., Dambacher J., Young J., Molina J. J., Hoyle S., Lawson T., International workshop on opportunities for ecosystem approaches to fisheries management in the Pacific Ocean tuna fisheries. Rev. Fish Biol. Fish. 22, 29–33 (2012). [Google Scholar]
- 12.de Souza E. N., Boerder K., Matwin S., Worm B., Improving fishing pattern detection from satellite AIS using data mining and machine learning. PLOS ONE 11, e0158248 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.McCauley D. J., Woods P., Sullivan B., Bergman B., Jablonicky C., Roan A., Hirshfield M., Boerder K., Worm B., Ending hide and seek at sea. Science 351, 1148–1150 (2016). [DOI] [PubMed] [Google Scholar]
- 14.Natale F., Gibin M., Alessandrini A., Vespe M., Paulrud A., Mapping fishing effort through AIS data. PLOS ONE 10, e0130746 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.W. Merten, A. Reyer, J. Savitz, J. Amos, P. Woods, B. Sullivan, Global Fishing Watch: Bringing transparency to global commercial fisheries. https://arxiv.org/abs/1609.08756 (2016).
- 16.Queiroz N., Humphries N. E., Mucientes G., Hammerschlag N., Lima F. P., Scales K. L., Miller P. I., Sousa L. L., Seabra R., Sims D. W., Ocean-wide tracking of pelagic sharks reveals extent of overlap with longline fishing hotspots. Proc. Natl. Acad. Sci. U.S.A. 113, 1582–1587 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cuthbert R., Hilton G., Ryan P., Tuck G. N., At-sea distribution of breeding Tristan albatrosses Diomedea dabbenena and potential interactions with pelagic longline fishing in the South Atlantic Ocean. Biol. Conserv. 121, 345–355 (2005). [Google Scholar]
- 18.Hazen E. L., Jorgensen S., Rykaczewski R. R., Bograd S. J., Foley D. G., Jonsen I. D., Shaffer S. A., Dunne J. P., Costa D. P., Crowder L. B., Block B. A., Predicted habitat shifts of Pacific top predators in a changing climate. Nat. Clim. Chang. 3, 234–238 (2013). [Google Scholar]
- 19.Druon J.-N., Fromentin J.-M., Hanke A. R., Arrizabalaga H., Damalas D., Tičina V., Quílez-Badia G., Ramirez K., Arregui I., Tserpes G., Reglero P., Deflorio M., Oray I., Karakulak F. S., Megalofonou P., Ceyhan T., Grubišić L., MacKenzie B. R., Lamkin J., Afonso P., Addis P., Habitat suitability of the Atlantic bluefin tuna by size class: An ecological niche approach. Prog. Oceanogr. 142, 30–46 (2016). [Google Scholar]
- 20.Roberts J. J., Best B. D., Mannocci L., Fujioka E., Halpin P. N., Palka D. L., Garrison L. P., Mullin K. D., Cole T. V. N., Khan C. B., McLellan W. A., Pabst D. A., Lockhart G. G., Habitat-based cetacean density models for the U.S. Atlantic and Gulf of Mexico. Sci. Rep. 6, 22615 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Soykan C. U., Eguchi T., Kohin S., Dewar H., Prediction of fishing effort distributions using boosted regression trees. Ecol. Appl. 24, 71–83 (2014). [DOI] [PubMed] [Google Scholar]
- 22.Howell E. A., Kobayashi D. R., Parker D. M., Balazs G. H., Polovina J. J., TurtleWatch: A tool to aid in the bycatch reduction of loggerhead turtles Caretta caretta in the Hawaii-based pelagic longline fishery. Endanger. Species Res. 5, 267–278 (2008). [Google Scholar]
- 23.J. Raskie, “Geospatial analysis of fisheries to improve federal enforcement,” thesis, Nicholas School of the Environment, Duke University (2013). [Google Scholar]
- 24.Hobday A. J., Hartog J. R., Spillman C. M., Alves O., Seasonal forecasting of tuna habitat for dynamic spatial management. Can. J. Fish. Aquat. Sci. 68, 898–911 (2011). [Google Scholar]
- 25.Pinsky M. L., Fogarty M., Lagged social-ecological responses to climate and range shifts in fisheries. Clim. Chang. 115, 883–891 (2012). [Google Scholar]
- 26.Kroodsma D. A., Mayorga J., Hochberg T., Miller N. A., Boerder K., Ferretti F., Wilson A., Bergman B., White T. D., Block B. A., Woods P., Sullivan B., Costello C., Worm B., Tracking the global footprint of fisheries. Science 359, 904–908 (2018). [DOI] [PubMed] [Google Scholar]
- 27.Crespo G. O., Dunn D. C., A review of the impacts of fisheries on open-ocean ecosystems. ICES J. Mar. Sci. 74, 2283–2297 (2017). [Google Scholar]
- 28.Bertrand S., Bertrand A., Guevara-Carrasco R., Gerlotto F., Scale-invariant movements of fishermen: The same foraging strategy as natural predators. Ecol. Appl. 17, 331–337 (2007). [DOI] [PubMed] [Google Scholar]
- 29.Reygondeau G., Maury O., Beaugrand G., Marc Fromentin J., Fonteneau A., Cury P., Biogeography of tuna and billfish communities. J. Biogeogr. 39, 114–129 (2012). [Google Scholar]
- 30.Arrizabalaga H., Dufour F., Kell L., Merino G., Ibaibarriaga L., Chust G., Irigoien X., Santiago J., Murua H., Fraile I., Chifflet M., Goikoetxea N., Sagarminaga Y., Aumont O., Bopp L., Herrera M., Fromentin J. M., Bonhomeau S., Global habitat preferences of commercially valuable tuna. Deep-Sea Res. Pt. II Top. Stud. Oceanogr. 113, 102–112 (2015). [Google Scholar]
- 31.Evans K., Langley A., Clear N. P., Williams P., Patterson T., Sibert J., Hampton J., Gunn J. S., Behaviour and habitat preferences of bigeye tuna (Thunnus obesus) and their influence on longline fishery catches in the western Coral Sea. Can. J. Fish. Aquat. Sci. 65, 2427–2443 (2008). [Google Scholar]
- 32.Brill R. W., Bigelow K. A., Musyl M. K., Fritsches K. A., Warrant E. J., Bigeye tuna (Thunnus obesus) behavior and physiology and their relevance to stock assessments and fishery biology. Col. Vol. Sci. Pap. ICCAT 57, 142–161 (2005). [Google Scholar]
- 33.Neilson J. D., Loefer J., Prince E. D., Royer F., Calmettes B., Gaspar P., Lopez R., Andrushchenko I., Seasonal distributions and migrations of Northwest Atlantic swordfish: Inferences from integration of pop-up satellite archival tagging studies. PLOS ONE 9, e112736 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Walli A., Teo S. L. H., Boustany A., Farwell C. J., Williams T., Dewar H., Prince E., Block B. A., Seasonal movements, aggregations and diving behavior of Atlantic bluefin tuna (Thunnus thynnus) revealed with archival tags. PLOS ONE 4, e6151 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ardron J., Gjerde K., Pullen S., Tilot V., Marine spatial planning in the high seas. Mar. Policy 32, 832–839 (2008). [Google Scholar]
- 36.D. Kacev, R. L. Lewison, Satellite remote sensing in support of fisheries management in global oceans, in Earth Science Satellite Applications (Springer, 2016), pp. 207–222. [Google Scholar]
- 37.M. A. Cervera, A. Ginesi, On the performance analysis of a satellite-based AIS system, 10th International Workshop on Signal Processing for Space Communications (SPSC 2008), Rhodes Island, Greece, 6 to 8 October, 2008. [Google Scholar]
- 38.Lewison R. L., Crowder L. B., Read A. J., Freeman S. A., Understanding impacts of fisheries bycatch on marine megafauna. Trends Ecol. Evol. 19, 598–604 (2004). [Google Scholar]
- 39.Dunn D. C., Kot C. Y., Halpin P. N., A comparison of methods to spatially represent pelagic longline fishing effort in catch and bycatch studies. Fish. Res. 92, 268–276 (2008). [Google Scholar]
- 40.Barbet-Massin M., Jiguet F., Albert C. H., Thuiller W., Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 3, 327–338 (2012). [Google Scholar]
- 41.J. Franklin, Mapping Species Distributions: Spatial Inference and Prediction (Cambridge Univ. Press, 2010). [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/4/8/eaat3681/DC1
Supplementary Materials and Methods
Fig. S1. The proportion of 2015 and 2016 fishing effort (hours) in ABNJ by gear.
Fig. S2. The proportion of pelagic longline fishing effort attributed to the main fishing States or territories.
Fig. S3. Accuracy values obtained for the 2015 and 2016 monthly boosted regression tree models after applying an ROC threshold.
Fig. S4. Accuracy values obtained for the 2015 and 2016 monthly boosted regression tree models after applying an MPD threshold.
Fig. S5. The predictive accuracy of the monthly BRTs after projecting them onto future environments.
Fig. S6. Distribution of predicted and observed fishing effort in January and July of 2015 using different thresholds: ROC and MPD.
Fig. S7. The SST partial dependence plots from the monthly 2015 models.
Fig. S8. The temperature at 400-m partial dependence plots from the monthly 2015 models.
Fig. S9. The DCS partial dependence plots from the monthly 2015 models.
Fig. S10. The oxygen at 400-m partial dependence plots from the monthly 2016 models.
Fig. S11. The SST partial dependence plots from the monthly 2015 models.
Fig. S12. The distribution of fishing effort intensity as a function of the Euclidean distance (kilometers) to the continental shelf.
Fig. S13. Monthly variable importance scores for boosted regression trees using background pseudoabsence points from the entire high seas areas for 2015 and 2016.
Table S1. Various model performance indices of the monthly BRTs for 2015 and 2016.
Table S2. Various model performance indices of the monthly BRTs for 2015 and 2016.
Table S3. Various model performance indices of the temporally averaged BRT model.
Table S4. Various model performance indices of the temporally averaged BRT model.
Table S5. Results from the Wilcoxon signed-rank test comparing the performance of monthly models to the temporally averaged model.
Table S6. Amount of fundamental niche occupied by pelagic longliners.
Table S7. The 2015 VI scores.
Table S8. The 2016 VI scores.
Table S9. Average 2016 model performance metrics using different environmental variables.
Table S10. Description of the variable type and source for each of the 14 biophysical and physiographic predictors.
Table S11. The number of presence and pseudoabsence points in 2015.
Table S12. The number of presence and pseudoabsence points in 2016.