

Abstract
The plant oncogene rolD stimulates the reproductive phase transition in plants. We define here the function of its gene product. We show that the RolD protein bears sequence homology with ornithine cyclodeaminase, an uncommon enzyme of specialized-niche eubacteria and archaea that catalyzes the unusual NAD+-dependent conversion of ornithine to proline. To confirm the prediction of the bioinformatic analysis, the RolD protein was expressed in Escherichia coli and purified. An ornithine-dependent NAD+ reduction that can be ascribed only to ornithine cyclodeaminase (OCD) activity was detected both in bacterial extracts containing RolD and in assays on the purified RolD protein. Furthermore, OCD activity was observed in soluble extracts from plants overexpressing rolD. The role of rolD in plant pathogenesis and its effect on plant reproductive development are discussed in light of the newly demonstrated enzymatic activity of its gene product.
The neoplastic disease hairy
root of dicotyledonous plants, an abundant ectopic growth of
adventitious roots, is because of transfer of transferred DNA
(T-DNA) from the Ri plasmid of the infecting Agrobacterium
rhizogenes to plant cells (1–4). Of the 18 ORF localized in the
Ri T-DNA (5), four coincide with genetic loci (rolA,
B, C, D) identified by transposon
mutagenesis from the effects of their inactivation on the virulence
properties of A. rhizogenes (6). When individually inserted
in plants, the rol oncogenes affect plant growth and
development, each in its characteristic and distinctive way. Transgenic
plants containing rolA show wrinkled leaves and reduced
internodal distance (7, 8). Plants expressing rolB show
flower heterostyly and abundant adventitious rooting (7, 9, 10).
rolC results in reduced apical dominance, altered leaf
morphology, and reduced seed production (7). More recently, the effects
of rolD have been shown to consist primarily in a strong
acceleration and stimulation of flowering both in tobacco plants and
cultured tissues (11) and Arabidopsis (M. L. Mauro and
M. M. Altamura, personal communication). So far, nothing is
known on the biochemical function of the RolD protein. Of the other Rol
proteins, RolA has been suggested to be involved in the metabolism of
gibberellins (12) and RolC in the deconjugation of cytokinins (13).
RolB increases the sensitivity of transformed cells to auxin (14, 15)
by perturbing the hormone's signal transduction, possibly through its
protein tyrosine phosphatase activity (16). Where determined, the
intracellular localization of the proteins encoded by the
rol genes is consistent with and supportive of their
biochemical functions. The RolB oncoprotein, apparently involved in
signal transduction, has been localized in the plasma membrane fraction
of transformed cells (16), whereas RolC, a β-glucosidase, is a
cytoplasmic protein (13). In this paper, we demonstrate that
rolD encodes a functional OCD converting ornithine into
proline and NH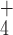 , localized in the cytoplasm of
transformed plant cells. We discuss the origin and effects of the
oncogene in light of this enzymatic activity.
, localized in the cytoplasm of
transformed plant cells. We discuss the origin and effects of the
oncogene in light of this enzymatic activity.
Materials and Methods
Plasmid Constructs.
The 35S∷rolD construct was derived from PBin-rolD (11) by amplifying the rolD coding region with suitable primers. The resulting fragment was cloned in pBI121 (CLONTECH). Gene fusions 6×His-rolD and 6×His-dhfr-rolD for expression in Escherichia coli M15(Rep4) were prepared by cloning the rolD coding region in the vectors pQE30 and, respectively, pQE13 (Qiagen, Chatsworth, CA).
Purification of RolD and Antibody Production.
Purification of denatured 6×His-DHFR-RolD (DHFR, dihydrofolate reductase) and 6×His-RolD proteins was performed on a Ni2+-chelate column according to Qiagen specifications. Rabbit polyclonal antibodies were raised against 6×His-DHFR-RolD and purified. For Western blot analysis, proteins were separated on 12% SDS/PAGE and electroblotted into Hybond C-plus membrane (Amersham Pharmacia). Anti-RolD antibodies were used at 1:1,000 dilution; commercial anti-6×His monoclonals (Amersham Pharmacia) were used according to the supplier's specifications. Nitro-blue tetrazolium/5-bromo-4-chloro-indolyl phosphate was used for visualization. Refolding of the denatured 6×His-RolD protein was performed on a PD10 gel filtration column (Amersham Pharmacia). Elution was performed in a refolding buffer containing 100 mM Tris⋅HCl (pH 7.5), 10% (vol/vol) glycerol, 10 mM KCl, 5 mM MgCl2, 20 mM βmercaptoethanol, and 0.1 mM NAD+.
Preparation of Plant Extracts.
SR1 (17) transformed tobacco plants were obtained and grown as previously described (11). Eight PCR-positive 35S∷rolD transformants (T1) were propagated in the growth chamber and transferred to the greenhouse with a comparable number of controls. T1 plants were selfed and the T2 progeny used for further analysis.
Leaves from greenhouse-grown T2 35S∷rolD and control plants (untransformed SR1 plants and plants transformed with the vector alone) were frozen in liquid nitrogen and homogenized in 100 mM Tris⋅HCl, pH 7.5/10% (vol/vol) glycerol/10 mM KCl/5 mM MgCl2/100 mM β-mercaptoethanol/400 mM sucrose/1 mM phenylmethylsulfonyl fluoride (extraction buffer). Crude homogenates were filtered through Miracloth (Calbiochem) and centrifuged at 10,000 × g for 10 min. The supernatant was centrifuged at 100,000 × g for 60 min and the resulting soluble fraction collected and dialyzed against several changes of extraction buffer in the presence of 10 μM NAD+. All purification steps were carried out at 4°C.
Enzymatic Assays.
Enzymatic activity was assayed on soluble protein extracts (typically 50–100 μg protein) from E. coli and from plant leaves and on purified RolD protein (50–100 ng) in 10 mM Hepes, pH 8.2/1 mM EDTA/2 mM NAD+/10 mM DTT/1 mM ornithine (reaction solution). The accumulation of NADH as a reaction product was measured by recording the increase of the absorbance at 340 nm as a function of time, with a Shimadzu UV-160A spectrophotometer. For proline detection, 2 mg of dialyzed soluble extract was dialyzed overnight at room temperature against 10 ml of reaction buffer. After dialysis, the 10-ml reaction buffer was loaded onto a DOWEX 2 column (Sigma), washed with 0.1 M ammonium bicarbonate (pH 8.2), and eluted with 30 ml 0.1 M HCl. The eluate was lyophilized and amino acid analysis performed monitoring the ninhydrin derivative at 440 nm by using a Beckman model 168 diode array detector.
Results
The RolD Protein Has Strong Sequence Homology with OCDs.
By means of the Smith–Waterman algorithm as implemented by the ssearch interface (http://sss.berkeley.edu/), a significant similarity (25% amino acid identity over 90% of the protein; E value = 0.0014) was found between the protein RolD and the OCD from Archeoglobus fulgidus. To confirm this indication, a similarity profile was built by first identifying OCD homologues by means of the blastp program (http://www.ncbi.nlm.nih.gov) and subsequently aligning them to RolD with clustalw (http://www.ebi.ac.uk/). As can be seen in Fig. 1, there is strong similarity between RolD and a number of different OCDs. All proteins share similar length ranging between 313 and 362 amino acids, and conservative amino acid substitutions as well as strictly conserved amino acid residues are present throughout the multialignment. Furthermore, a similarity profile between RolD and different OCDs was confirmed by using the iterative program psi-blast. The extremely low E values of the matches in the similarity profile, ranging between 1 × 10−32 and 1 × 1−94, warrant the high significance of these results.
Figure 1.

The RolD protein bears strong similarity to OCDs from different organisms. clustalw multiple alignment of the amino acid sequence of RolD and mu-crystallins from mouse (Mus, sp:054983), human (homo, sp: Q14894), and Kangaroo (Macropus, pir: S27950); OCD from Archeoglobus fulgidus (Archeoglobus, gi:2648890); OCD from Corynebacterium glutamicum (Corynebacterium, emb: CAA07634); OCD from Methanobacterium thermoautotrophicum (Methanobacterium, gi:2622612); OCD from nopaline A. tumefaciens plasmid pTic58 (Agrobacterium, sp: P09773); OCD from Sinorhizobium melitoti (Sinorhizobium, X64613); OCD from octopine A. tumefaciens plasmid R10 (octopine, gi:1066073); OCD from octopine A. tumefaciens plasmid AGRT5 (AGRT5, gi:95109); OCD from Rhodobacter capsulatus (Rhodobacter, gi:3128286); OCD from Brucella abortus (Brucella, sp:059175). The numbers identify protein sequences in the SwissProt (sp), Pir (pir), European Molecular Biology Laboratory (emb), and GenBank (gi) databases. The amino acid residues are colored according to their physicochemical properties. Aliphatic/hydrophobic (ILVAMWFC), blue; hydrophilic (STNQ), green; positively charged (RK), red; negatively charged (ED), magenta; tyrosine and histidine (YH), light blue; proline (P), yellow; glycine (G), orange.
RolD Expressed in E. coli Has OCD Activity.
To verify the OCD activity predicted by the bioinformatic analysis, a histidine-tagged RolD protein was expressed in E. coli, as shown in Fig. 2A. The 6×His-RolD protein was detected in bacterial protein extracts with polyclonal anti-RolD antibodies raised from a purified 6×His-dihydrofolate reductase-RolD fusion protein, and with commercial monoclonal anti-6×His antibodies (Fig. 2 B and C). Because OCD converts ornithine to proline and specifically requires NAD+ as cofactor, bacterial protein extracts were incubated in the presence of ornithine and NAD+. The formation of NADH as reaction product was monitored through the kinetics of increase of absorbance at 340 nm (the absorbance maximum of NADH). As shown in the spectrophotometric tracings reported in Fig. 3A, NAD+ is indeed reduced to NADH in an ornithine-dependent fashion, because in the absence of ornithine, no activity is observed. No activity was detected in control extracts from E. coli expressing the vector alone (not shown).
Figure 2.

Expression of the RolD protein in E. coli. (A) PAGE of total protein extracts from uninduced (lane1) and IPTG-induced (lane 2) E. coli harboring the 6×His-rolD construct, and of purified denatured (lane 3) and refolded (lane 4) 6×His-RolD protein. (B and C) Immunoblot detection of the 6×His-RolD protein in extracts from E. coli expressing the 6×His-rolD construct with polyclonal anti-RolD antibodies (B) and monoclonal anti-6×His antibodies (C).
Figure 3.

The RolD protein expressed in E. coli has OCD activity. The absorbance at 340 nm of the reaction mixture was spectrophotometrically monitored in continuum and converted in NADH produced per milligram of protein. (A) Activity of total protein extracts from E. coli expressing the 6×His-rolD construct (Upper) and the same in the absence of ornithine (Lower). (B) Activity of the purified refolded 6×His-RolD protein (Upper) and the same in the absence of ornithine (Lower).
The presence of the N-terminal histidine tag allowed purification of 6×His-RolD by affinity chromatography. Most of the 6×His-RolD protein accumulates in the bacteria in the insoluble form. We thus purified the denatured recombinant protein (Fig. 2A, lane 3) and subsequently recovered a small amount of it in the native soluble form through a refolding protocol (Fig. 2A, lane 4). As shown in Fig. 3B, the refolded RolD protein has ornithine-dependent OCD activity.
Extracts from Plants Overexpressing rolD Have OCD Activity.
To achieve a high level of expression of the oncogene in plants, SR1 tobacco plants (17) were transformed with a construct carrying rolD under the control of the strong constitutive CaMV35S promoter (35S∷rolD plants). Protein blot experiments with anti-RolD antibodies revealed the presence of the RolD protein in extracts from leaves of the 35S∷rolD plants, as shown in Fig. 4A, lane 2. No signal was detected in leaves from untransformed SR1 plants (Fig. 4A, lane 1), plants transformed with the empty vector (not shown), or in leaves presented with preimmune antiserum (not shown). By means of differential centrifugation, we localized RolD in the cytosolic fraction of cells from leaves of the 35S∷rolD plants, as shown in Fig. 4A, lane 4.
Figure 4.

Extracts from plants overexpressing rolD have OCD activity. (A) Immunoblot detection of the RolD protein in soluble extracts from leaves of SR1 (lane 1) and 35S∷rolD plants (lane 2), and from subcellular 10,000 × g pellet (lane 3), 100,000 × g supernatant (lane 4) and 100,000 × g pellet (lane 5) from leaves of 35S∷rolD plants. All samples are equivalent to 50 mg of leaves (fresh weight). (B) Activity (see legend to Fig. 3) of protein extracts from leaves of 35S∷rolD plants (Upper) and the same in the absence of ornithine (Lower). (C) Amino acid analyzer tracings showing the production of more abundant proline (arrows) in the extracts from leaves of 35S∷rolD plants than of SR1 plants. The effluents are monitored at 440 nm, and amino acids are identified by coelution with an internal standard (not shown).
Soluble extracts from leaves of 35S∷rolD and of control plants were extensively dialyzed (see Materials and Methods) and then tested for enzymatic activity. The results obtained with extracts from different 35S∷rolD plants were qualitatively similar, and a typical assay on one of them is reported in Fig. 4B. As shown, NAD+ is reduced to NADH and in the absence of ornithine, no activity is observed. The NAD+-reducing activity is proportional to enzyme concentration and is subjected to product inhibition as described for the OCD of Agrobacterium tumefaciens (18). The product responsible for inhibition is NADH, as addition of proline does not affect NAD+ reduction, whereas addition of 1 mM NADH results in total inhibition (not shown). NAD+ is specifically required and neither NADP+ nor FAD can substitute for it (not shown). The OCD of extracts from 35S∷rolD plants is active only under reducing conditions, as also reported for other OCDs (19). None of the extracts from control plants showed any comparable activity.
To assess the production of proline as a result of OCD activity, the enzymatic analysis was performed on a bulk amount of dialyzed soluble extracts (about 2 mg of protein) from 35S∷rolD plants and from SR1 controls. To allow accumulation of proline as reaction product but relieve product inhibition by NADH, the enzymatic assay was performed by dialyzing overnight the extracts against a 10-fold excess of the reaction solution. After a subsequent ion-exchange purification, significantly higher amounts of proline were detected in extracts from the 35S∷roD plant than in SR1 control extracts, as shown in Fig. 4. No differences in other amino acid acids were observed (not shown).
Discussion
The RolD Protein Is a Functional OCD.
In this work, we show by way of biochemical assays based on a bioinformatic prediction that the plant oncogene rolD from A. rhizogenes encodes a functional OCD. The similarity profile between RolD and different OCDs built by means of a combination of iterative and noniterative methods provided a high statistical significance of the alignment and a high level of confidence in the similarity proposed. The prediction on the biochemical function of the RolD protein was experimentally confirmed by enzymatic assays on RolD as expressed in, and purified from, E. coli, and on soluble extracts from plants overexpressing the oncogene. The enzymatic assays revealed a specific ornithine-dependent NAD+ reduction with production of proline that can only be accounted for by OCD activity.
No OCD has been described in plants, where ornithine can be converted to proline only via pyridoxal phosphate-dependent reactions. OCD has been characterized best in A. tumefaciens where this enzymatic activity is involved in the catabolism of the arginine-derived opines that are produced by tumor cells and used as carbon and nitrogen source by the bacterium. In A. tumefaciens, OCD is encoded by genes localized in the non-T-DNA region of the Ti plasmid, i.e., the part that is not inserted in infected plant cells (18, 20, 21). Interestingly, in the closely related A. rhizogenes, OCD is not used by the bacterium for opine catabolism and the encoding gene has become part of the T-DNA, has acquired eukaryotic cis regulatory elements, and is expressed only in plant cells. OCD is also present in the legume bacterial symbiont Sinorhizobium meliloti, where its activity increases the efficiency of root nodulation (22, 23). The presence of OCD in only a limited number of other very particular prokaryotes such as light harvesting (Rhodobacterium) or parasite (Brucella) bacteria or in extremophyle archaea (Archeoglobus, Methanobacterium) further testifies to the specialized-niche character of this unusual enzyme. In animals, the only OCD-related proteins are mu-crystallins (24). This is not surprising, because often eye lens proteins have been recruited among metabolic enzymes (25).
Possible Role of RolD/OCD in Hairy Root Induction and in Flowering.
In light of the enzymatic activity demonstrated for the RolD protein, its developmental effects in hairy root induction and in floral transition of transformed plants could be because of either the accumulation of its reaction product (proline) or the depletion of its substrate (ornithine).
Transposon inactivation of rolD in the Ri plasmid T-DNA was shown to impair the virulence of agropine-type A. rhizogenes. On infection with a rolD-mutated strain, root initiation still occurs but subsequent root growth is impaired (6), pointing to a role of rolD in root elongation. The proline produced by the OCD enzymatic activity of RolD may prompt an explanation for the role of this protein in hairy root growth. A substantial increase of proline concentration in the growing region of maize primary roots at low water potential has in fact been reported, a finding that suggests a role for proline biosynthesis in sustaining root growth under these conditions (26). An increase in the amount of proline could affect the rate of biosynthesis of hydroxyproline-rich glycoproteins (HRGPs, extensins, and arabinogalactan proteins). These proteins are structural constituents of the plant cell wall and are thought to play a key role in the regulation of cell division, cell wall self assembly, and cell extension (27). Alternatively, stimulation of root growth by rolD could be related to the depletion of ornithine and consequent possible alterations of the polyamine pool, of which ornithine is a precursor. Overexpression of arginine decarboxylase–arginine is another precursor of polyamines, resulting in an increased level of putrescine, was shown to impair root growth in tobacco plants (28).
When individually inserted in plants, the oncogene rolD has a remarkable effect on development. Tobacco plants expressing rolD under the control of its promoter set earlier and more abundant flowers than untransformed controls (11). More recently, we observed stimulation of flowering also in rolD-Arabidopsis plants (M. L. Mauro and M. M. Altamura, personal communication). This intriguing effect of the oncogene on plant reproductive transition may also be related to the production of proline because of the OCD activity. In addition to its role as stress-related osmoprotectant, evidence is accumulating that may point to a role of proline in flower transition. Analysis of the free amino acid composition in tomato revealed that proline content in flowers is 60 times higher than in any other organ. It has been suggested that proline is imported via a specific transporter from vegetative tissues to the flower, where increased proline levels were correlated with flower formation (29). Support for this hypothesis may also come from work on the Arabidopsis gene encoding 1-pyrroline-5-carboxylate syntethase, a key enzyme in proline biosynthesis. Arabidopsis plants expressing an antisense to P5CS show a defective inflorescence elongation and a role for proline in morphogenesis has been suggested (30). Accordingly, Arabidopsis plants overexpressing P5CS produce much more abundant flowers than controls under saline stress (31). In addition, the same P5CS proline biosynthetic gene has been identified among the targets of CONSTANS, a transcriptional activator that controls flowering time in Arabidopsis (32).
As discussed above for root growth, also the effects of overproduction of proline on reproductive transition may be because of stimulation of hydroxyproline-rich glycoproteins biosynthesis, as extensins are developmentally regulated and their occurrence correlates with changes in cell morphology and with the formation of anatomical patterns (33, 34). Accumulation of proline in specific developmental stages may, instead, act as a signal in analogy with the recognized role of amino acids as regulators in yeast and animals (35, 36).
Alternatively, depletion of the ornithine pool may underlie the effect of RolD in flower transition. As mentioned, ornithine is a precursor of polyamines, and there is evidence suggesting an involvement of both free and conjugated polyamines in flowering (37–39).
The promoter of the rolD oncogene is developmentally regulated and is very active in elongating and differentiating tissues and in organs with determinate growth—cotyledons, leaves, and flowers—rolD expression parallels the age gradient of the tissues (40). It will be interesting to assess whether and how the accumulation of proline and/or the depletion of ornithine in specific developmental stages could affect the reproductive phase transition in plants. The finding reported in this work provides a powerful tool to investigate in this direction.
Acknowledgments
This work is dedicated to the memory of Franco Tatò. We thank Prof. Donatella Barra and Dr. Giusi Pitari for helpful discussions and Dr. Manuela Citterich for suggestions on bioinformatic analysis. This work was partially supported by grants from Ministero dell'Università e della Ricerca Scientifica e Technologica, Consiglio Nazionale delle Ricerche (Italy) (PF Biotecnologie), and the European Union (FP5 contract REGIA) to P.C.
Abbreviation
- OCD
ornithine cyclodeaminase
- T-DNA
transferred DNA
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
References
- 1.Chilton M-D, Tepfer D A, Petit A, Casse-Delbart F, Tempé J. Nature (London) 1982;295:432–434. [Google Scholar]
- 2.Spanò L, Pomponi M, Costantino P, Van Slogteren G M S, Tempé J. Plant Mol Biol. 1982;1:291–300. doi: 10.1007/BF00027560. [DOI] [PubMed] [Google Scholar]
- 3.White F F, Ghidossi G, Gordon M P, Nester E W. Proc Natl Acad Sci USA. 1982;79:3193–3197. doi: 10.1073/pnas.79.10.3193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Willmitzer L, Sanchez-Serrano J, Buschfeld E, Schell J. Mol Gen Genet. 1982;186:16–22. [Google Scholar]
- 5.Slightom J L, Durand-Tardif M, Jouanin L, Tepfer D. J Biol Chem. 1986;261:108–121. [PubMed] [Google Scholar]
- 6.White F F, Taylor B H, Huffmman G A, Gordon M P, Nester E W. J Bacteriol. 1985;164:33–44. doi: 10.1128/jb.164.1.33-44.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Schmülling T, Schell J, Spena A. EMBO J. 1988;7:2621–2629. doi: 10.1002/j.1460-2075.1988.tb03114.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sinkar V P, Pythoud F, White F F, Nester E W. Genes Dev. 1988;2:688–697. doi: 10.1101/gad.2.6.688. [DOI] [PubMed] [Google Scholar]
- 9.Cardarelli M, Mariotti D, Pomponi M, Spanò L, Capone I, Costantino P. Mol Gen Genet. 1987;209:475–480. doi: 10.1007/BF00331152. [DOI] [PubMed] [Google Scholar]
- 10.Spena A, Schmülling T, Koncz C, Schell J. EMBO J. 1987;6:3891–3899. doi: 10.1002/j.1460-2075.1987.tb02729.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mauro M L, Trovato M, De Paolis A, Gallelli A, Costantino P, Altamura M M. Dev Biol. 1996;180:693–700. doi: 10.1006/dbio.1996.0338. [DOI] [PubMed] [Google Scholar]
- 12.Dehio C, Grossmann K, Schell J, Schmülling T. Plant Mol Biol. 1993;23:1199–1210. doi: 10.1007/BF00042353. [DOI] [PubMed] [Google Scholar]
- 13.Estruch J J, Chriqui D, Grossmann K, Schell J, Spena A. EMBO J. 1991;10:2889–2895. doi: 10.1002/j.1460-2075.1991.tb07838.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Spanò L, Mariotti D, Cardarelli M, Branca C, Costantino P. Plant Physiol. 1988;87:479–483. doi: 10.1104/pp.87.2.479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Maurel C, Barbier-Brygoo H, Spena A, Tempé J, Guern J. Plant Physiol. 1991;97:212–216. doi: 10.1104/pp.97.1.212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Filippini F, Rossi V, Marin O, Trovato M, Costantino P, Downey P M, Lo Schiavo F, Terzi M. Nature (London) 1996;379:499–500. doi: 10.1038/379499a0. [DOI] [PubMed] [Google Scholar]
- 17.Maliga P A, Sz-Breznovits A, Marton L. Nat New Biol. 1973;244:29–30. doi: 10.1038/newbio244029a0. [DOI] [PubMed] [Google Scholar]
- 18.Sans N, Schindler U, Schröder J. Eur J Biochem. 1988;173:123–130. doi: 10.1111/j.1432-1033.1988.tb13975.x. [DOI] [PubMed] [Google Scholar]
- 19.Muth W L, Costilow R N. J Biol Chem. 1974;249:7457–7462. [PubMed] [Google Scholar]
- 20.Sans N, Schröder G, Schröder J. Eur J Biochem. 1987;167:81–87. doi: 10.1111/j.1432-1033.1987.tb13306.x. [DOI] [PubMed] [Google Scholar]
- 21.Schindler U, Sans N, Schröder J. J Bacteriol. 1989;171:847–854. doi: 10.1128/jb.171.2.847-854.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Soto M J, Zorzano A, Garcia-Rodriguez F M, Mercado-Blanco J, Lopez-Lara I M, Olivares J, Toro N. Mol Plant–Microbe Interact. 1994;7:703–707. doi: 10.1094/mpmi-7-0703. [DOI] [PubMed] [Google Scholar]
- 23.Jimenez-Zurdo J I, van Dillewijn P, Soto M J, de Felipe M R, Olivares J, Toro N. Mol Plant–Microbe Interact. 1995;8:492–498. doi: 10.1094/mpmi-8-0492. [DOI] [PubMed] [Google Scholar]
- 24.Kim R Y, Gasser R, Wistow G J. Proc Natl Acad Sci USA. 1992;89:9292–9296. doi: 10.1073/pnas.89.19.9292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Piatigorsky J, Kantorow M, Gopal-Srivastava R, Tomarev S I. EXS. 1994;71:241–250. doi: 10.1007/978-3-0348-7330-7_24. [DOI] [PubMed] [Google Scholar]
- 26.Verslues P E, Sharp R E. Plant Physiol. 1999;119:1349–1360. doi: 10.1104/pp.119.4.1349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Varner J E, Lin L-S. Cell. 1989;56:231–239. doi: 10.1016/0092-8674(89)90896-9. [DOI] [PubMed] [Google Scholar]
- 28.Masgrau C, Altabella T, Farräs R, Flores D, Thompson A J, Besford R T, Tiburcio A F. Plant J. 1997;11:465–473. doi: 10.1046/j.1365-313x.1997.11030465.x. [DOI] [PubMed] [Google Scholar]
- 29.Schwacke R, Grallath S, Breitkreuz K E, Stransky E, Stransky H, Frommer W B, Rentsch D. Plant Cell. 1999;11:377–392. doi: 10.1105/tpc.11.3.377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Nanjo T, Kobayashi N T, Yoshiba Y, Sanada Y, Wada K, Tsukaya H, Kabkubari Y, Yamaguchi-Shinozaki K. Plant J. 1999;18:185–193. doi: 10.1046/j.1365-313x.1999.00438.x. [DOI] [PubMed] [Google Scholar]
- 31.Kavi Kishor P B, Hong Z, Miao G-H, Hu C-A A, Verma D P. Plant Physiol. 1995;108:1387–1394. doi: 10.1104/pp.108.4.1387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Samach A, Onouchi H, Gold S E, Ditta G S, Schwarz-Sommer S, Yanofsky M F, Coupland G. Science. 2000;288:1613–1616. doi: 10.1126/science.288.5471.1613. [DOI] [PubMed] [Google Scholar]
- 33.Knox J P. FASEB J. 1995;9:1004–1012. doi: 10.1096/fasebj.9.11.7544308. [DOI] [PubMed] [Google Scholar]
- 34.Cooper J B, Heuser J E, Varner J E. Plant Physiol. 1994;104:747–752. doi: 10.1104/pp.104.2.747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Fafournoux P, Bruhat A, Jousse C. Biochem J. 2000;351:1–12. doi: 10.1042/0264-6021:3510001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Daphne A, van Sluijters P, Dubbelhuis P F, Blommaart F C, Meijer J. Biochem J. 2000;351:545–550. doi: 10.1042/0264-6021:3510545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Evans P T, Malmberg R L. Annu Rev Plant Physiol. 1989;40:235–269. [Google Scholar]
- 38.Galston A W, Sawhney R K. Plant Physiol. 1990;94:406–410. doi: 10.1104/pp.94.2.406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Martin-Tanguy J. Physiol Plant. 1997;100:675–688. [Google Scholar]
- 40.Trovato M, Mauro M L, Costantino P, Altamura M M. Protoplasma. 1997;197:111–120. [Google Scholar]