

Abstract
Jellyfish populations apparently have increased in some places around the world and human problems with them also have increased. However, effects of jellyfish outbreaks in the ecosystems remain poorly understood and little or no information is available on their dietary preferences - in relation to the seasonal shifts of prey abundance - and on the potential variability of their impact on marine food webs. The mauve stinger Pelagia noctiluca (Forsskål, 1775) is by far the most common outbreak-forming scyphozoan jellyfish in the Western Mediterranean. By use of a combination of stomach contents, stable isotope (SI) and fatty acid (FA) analyses, we tested the hypothesis that changes in the seasonal dietary sources of P. noctiluca parallel changes in the FA and SI composition. Stomach content and biomarker analyses suggested that P. noctiluca is not a selective predator, cyclically shifting between carnivory and omnivory depending on the seasonality of accessible prey. The combination of SI and FA analyses highlighted the importance of microzooplankton as prey. Specific FA biomarkers showed that the diet of P. noctiluca changed seasonally depending on the availability of living plankton or suspended detritus. This study also revealed significant biochemical differences between jellyfish somatic and gonadal tissues, with total fatty acid concentration in the gonads up to ten times higher than in the somatic tissues.
Introduction
Studies of trophic relationships help to unravel mechanisms of marine ecosystem functioning by illuminating foraging patterns of key species and related flows of energy and matter. Due to the inherent spatial and temporal variability of pelagic communities, tracing trophic relationships may be not as straightforward as in benthic habitats, leaving large gaps in the mechanistic interpretation of unpredicted biological events such as massive and recurrent outbreaks of gelatinous zooplankton communities in several coastal waters worldwide1. For instance, carnivorous jellyfish are mainly subject to bottom-up controls from their forage base rather than to top-down controls from predators2. This suggests that knowing what, how much, and where they eat would be key information to establish ecophysiological optima and trophic links supporting regime shifts in marine ecosystems coupled to local or temporary dominance of jellyfish. This knowledge is of particular importance for predicting the potential impact of jellyfish on marine ecosystem functioning and services, and eventually on human activities in coastal areas3,4.
A range of methods has been used to study trophic relationships in aquatic organisms, each with its own strengths and drawbacks. Stomach content analysis is the most direct approach5,6. This methodology may be useful to describe the diet and provide quantitative estimates of ingested prey. However, digestibility of prey differs significantly and identifiable items in stomach contents may often represent the most recently ingested food or prey requiring long digestion times7. Microzooplankton do not leave easily detectable remains and are therefore usually neglected, with the loss of key information7,8. An alternative approach to stomach content analysis could be experimental feeding incubations, which may give insights on food selectivity and ingestion rates, but suffer from artifact and over-simplification of complex natural food assemblages in the laboratory9. Indeed, the feeding rate of gelatinous species has often been underestimated in grazing experiments performed in artificial containers6. The use of identifiable molecular biomarkers, which pass from food sources to the consumer, is a complementary approach to detect small, soft-bodied prey, like ciliates and flagellates10. Such tracers may convey and integrate information on a consumer’s diet over days to months10–12, but separation of food sources is not straightforward. Each method is sensitive to different timescales and aspects of the diet, often leading to conflicting results on trophic relationships13. These biomarkers are complementary to the stomach content approach, but not sufficient in some cases when complex trophic interactions have to be described7.
Overall fatty acid composition and specific fatty acids (FAs) used as biomarkers can help to elucidate trophic relationships in food webs. Indeed, some FAs can only be synthesized de novo by certain taxa, especially by algae and bacteria, and then are conservatively transferred to the next trophic level unchanged or modified in an expected, detectable way7,10,14. For example, calanoid copepods are thought to be the only source of C20:1 (n-11 and n-9) and C22:1 (n-11 and n-9) monounsaturated fatty acids15; the identification of these FAs in the tissues of jellyfish suggests direct predation on copepods or other crustacean zooplankton16. However, knowledge of the FA metabolic pathways in many invertebrate taxa is in its infancy. FAs may be used successfully as qualitative trophic tracers in marine food webs to verify or identify predator–prey relationships, if the assumption that tracking FAs from source to consumers can be reliably met. To understand how food source contributions may vary seasonally or through the life span, stable isotope (SI) analysis becomes a powerful tool in the overall approach17. In fact, the complexity of food webs and the rarity of unique biomarkers impose uncertainties in the interpretation of FA data from field studies that must be resolved with complementary information such as stomach contents or stable isotopic data7.
The proportions of SIs of carbon (δ13C) and nitrogen (δ15N) may vary depending on the nutrient source, the physiology of the primary producers, and the trophic level of the consumers. SI analysis has successfully been used to elucidate food source partitioning18,19 and food web dynamics20–22. Its application in gelatinous zooplankton trophic ecology, however, is still infrequent23–27, but may help to identify trophic guilds and their potential prey sources in combination with various complementary methodologies17. So far, studies on the foraging ecology of the scyphozoan Pelagia noctiluca (Forsskål, 1775), the predominant gelatinous predator in the central and western Mediterranean Sea28, have relied solely on the analysis of stomach contents27,29,30, with a single exception31.
Pelagia noctiluca was first recorded in 1785 in the Strait of Messina32, at the confluence of the Southern Tyrrhenian and Ionian seas, where it now regularly undergoes massive population outbreaks with important effects on the structure and organization of the overall planktonic community29. Comparison of stomach contents and zooplankton composition indicate that this jellyfish is a generalist predator, with a clear seasonal variation in its diet27,29. The seasonal changes in diet may lead to seasonal changes in the composition of FAs and, as for Aurelia aurita in the North Sea, to the temporal variability in jellyfish δ15N and δ13C SI signatures33. Other factors like physiology, changes in behavior, investment in reproductive output also could affect FAs and SIs34. Thus, study of the seasonal variation of biochemical markers could help us to better understand their periodic reproduction events, because the biochemical composition of specific tissues, such as the gonads, may affect the quality of the breeding stock35. The simultaneous application of the three methodologies (e.g., stomach contents, FA and SI analyses) to clarify the trophic position of P. noctiluca and its trophic linkages (by predation or competition) with other taxa has not been attempted previously.
In this study, our objective was to clarify the predatory trophic impact of P. noctiluca medusae on the coastal marine food web by elucidating potential jellyfish dietary preferences over the year in relation to the seasonal shifts of prey availabilities. We used a combination of stomach contents, SI and FA analyses to test the hypothesis that changes in the seasonal dietary sources of P. noctiluca are paralleled by changes in the FA and SI composition of somatic and gonadic tissues, potentially accounting for the seasonal variations of reproductive and outbreak-forming potential of jellyfish.
Results
Stomach contents
The abundances of zooplankton taxa in the stomach contents of P. noctiluca and in the surface water layers were comparable (Table 1, Supplementary Information 1). Copepods were the most abundant group all year, followed by fish eggs and pteropods (Table 1).
Table 1.
Monthly prey composition of zooplankton taxa in the stomach contents of Pelagia noctiluca.
| Nov (23) | Dec (17) | Feb (3) | Apr (12) | May (20) | Jun (27) | Jul (13) | Sep (13) | ||
|---|---|---|---|---|---|---|---|---|---|
| Prey | Siphonophores | 0.04 ± 0.04 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0.08 ± 0.0 |
| Pteropods | 0.70 ± 0.21+ | 7.71 ± 3.65 | 0 ± 0 | 0.22 ± 0.22 | 0.45 ± 0.35 | 0.26 ± 0.11 | 0 ± 0 | 0 ± 0 | |
| Ostracods | 0.04 ± 0.04 | 0 ± 0 | 0 ± 0 | 0.11 ± 0.11 | 0.1 ± 0.06 | 0 ± 0 | 0.08 ± 0.07 | 0 ± 0 | |
| Furcilia | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 − | 0.05 ± 0.05 | 0 ± 0 | 0 ± 0 | 0 ± 0 | |
| Fish larvae | 0.04 ± 0.04 | 0.06 ± 0.05 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0.04 ± 0.03 | 0.08 ± 0.07 | 0 ± 0 | |
| Fish eggs | 0.52 ± 0.16+ | 0.41 ± 0.12+ | 0 ± 0 | 1.33 ± 0.57 | 0.80 ± 0.22+ | 0.52 ± 0.16+ | 0.08 ± 0.07 | 0 ± 0 | |
| Copepods | 12.34 ± 3.45+ | 6.65 ± 2.62 | 0.33 ± 0.33 | 36.66 ± 8.17 | 21.95 ± 5.79+ | 10.37 ± 1.99+ | 15.23 ± 9.95 | 1.38 ± 0.46 | |
| Chaetognaths | 0.43 ± 0.19 | 0.60 ± 0.32+ | 0 ± 0 | 0 ± 0 | 1.35 ± 0.70 | 0.22 ± 0.12 | 0 ± 0 | 0 ± 0 | |
| Appendicularians | 0.04 ± 0.04 | 0 ± 0 | 0 ± 0 | 0.11 ± 0.11 | 0.20 ± 0.15 | 0.07 ± 0.05 | 0 ± 0 | 0 ± 0 | |
The numbers of jellyfish analyzed are in brackets beside each month. Values are expressed as mean number of prey medusa−1 ± standard error. Bold highlighted values indicate significant prey selection: “−” Significant negative selection; “+”positive significant selection. Significances were calculated using Chi-square test.
The total abundance of zooplankton prey in the stomach contents of P. noctiluca differed significantly over the year (Table 2). The highest values were in April and May, with 39 ± 8 (mean ± SE) and 25 ± 1 prey medusa−1, respectively. Copepods were consumed throughout the period of investigation, mostly in spring (April and May, 37 ± 8 and 22 ± 6 copepods medusa−1) and early summer (June and July, 10 ± 2 and 15 ± 9 copepods medusa−1), whereas few were found at the end of summer (September, 1 ± 0.5 copepods medusa−1) or in winter (February, 0.3 ± 0.3 copepods medusa−1) (Table 1). Pteropods were particularly abundant prey in December and May, while copepods and fish eggs were numerous in spring.
Table 2.
One-way and 3-way PERMANOVA comparing (A) mean total prey (Total, prey medusa−1) and prey composition from stomach content analyses of Pelagia noctiluca; (B) mean δ15N and δ13C values from stable isotope analysis, and (C) total amount of fatty acids (ANOVA: Total), type of fatty acids (PERMANOVA: Saturated Fatty Acids, Monounsaturated Fatty Acids, and Polyunsaturated Fatty Acids: SFA-MUFA-PUFA), fatty acid composition for jellyfish and for plankton (FA composition).
| df | MS | F | p | |
|---|---|---|---|---|
| A. Stomach content | ||||
| Total | ||||
| Month | 8 | 1432 | 3.26 | ** |
| Residuals | 128 | 438 | ||
| Prey composition | ||||
| Month | 9 | 12.95 | 7.33 | *** |
| Residuals | 139 | 1.76 | ||
| B. Stable isotopes | ||||
| δ15N | ||||
| Month | 8 | 1.22 | 12.21 | *** |
| Residuals | 36 | 0.11 | ||
| δ13C | ||||
| Month | 8 | 2.92 | 27.93 | *** |
| Residuals | 36 | 0.1 | ||
| C. Fatty acids | ||||
| Total | ||||
| Month | 8 | 22.23 | 0.83 | 0.51 |
| Sex | 1 | 32.86 | 1.23 | 0.25 |
| Bp | 1 | 714 | 26.79 | *** |
| Month*Sex | 8 | 7.14 | 0.26 | 0.94 |
| Month*Bp | 8 | 26.19 | 0.98 | 0.41 |
| Sex*Bp | 1 | 3.57 | 0.13 | 0.73 |
| Month*Sex*Bp | 6 | 7.58 | 0.28 | 0.9 |
| Residuals | 50 | 26.65 | ||
| SFA-MUFA-PUFA | ||||
| Month | 8 | 714 | 0.93 | 0.47 |
| Sex | 1 | 1262 | 1.64 | 0.2 |
| Bp | 1 | 110 | 0.14 | 0.76 |
| Month*Sex | 8 | 1343 | 1.75 | 0.09 |
| Month*Bp | 8 | 1608 | 2.1 | * |
| Sex*Bp | 1 | 6153 | 8.04 | ** |
| Month*Sex*Bp | 6 | 376 | 0.49 | 0.83 |
| Residuals | 50 | 764 | ||
| FA composition (medusae) | ||||
| Month | 8 | 1308 | 1.65 | * |
| Sex | 1 | 2251 | 2.83 | * |
| Bp | 1 | 6793 | 8.56 | *** |
| Month*Sex | 8 | 608 | 0.76 | 0.76 |
| Month*Bp | 8 | 1164 | 1.46 | 0.08 |
| Sex*Bp | 1 | 5476 | 6.9 | *** |
| Month*Sex*Bp | 6 | 606 | 0.76 | 0.74 |
| Residuals | 50 | 792 | ||
| FA composition (plankton) | ||||
| Season | 2 | 2487 | 3.16 | ** |
| Plankton Size | 2 | 1143 | 1.45 | 0.19 |
| Season*Plankton. Size | 4 | 1037 | 1.32 | 0.24 |
| Residuals | 10 | 785 | ||
Significant p-values (≤0.05, ≤0.01, ≤0.001) are indicated by one, two, or three asterisks, respectively. Sex = Medusa sex; Bp = Body part; df = degree of freedom; MS = mean of square; F = statistic F test.
Monthly prey selection calculated by Pearre’s selection index for all prey of P. noctiluca showed significant positive selection for copepods (November, May, and June), pteropods (November) and fish eggs (November, December, May, and June), and negative selection for ostracods (December) and furcilia (April) (Table 1). For other all prey types and months, P. noctiluca showed a non-selective foraging strategy.
Stable Isotopes
Both δ13C and δ15N of P. noctiluca differed significantly over the year (Table 2). δ13C ranged from −20.5 ± 0.1‰ in April to −18.2 ± 0.1‰ in February (Fig. 1). In general, autumn and winter isotopic signatures had higher values than those in spring months (Fig. 1). δ15N ranged from 3.7 ± 0.1‰ in December to 5.0 ± 0.2‰ in July, with lower values in autumn and winter months (i.e., December and February) than in spring and summer (June, July, and August; Fig. 1).
Figure 1.
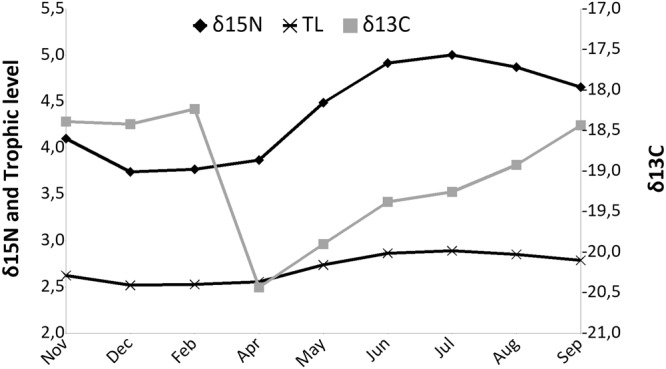
Monthly variations (mean ± standard error) of δ13C and δ15N (‰) values and trophic level (TL) in Pelagia noctiluca tissues.
Analysis of trophic level (TL) values (Fig. 1) showed the diet of P. noctiluca ranged from carnivorous in late spring-summer, with the highest TL value in July (2.8 ± 0.05), to omnivorous in late autumn-winter, with the lowest TL value in December (2.5 ± 0.02).
Comparison of δ15N and δ13C isotopic compositions showed that P. noctiluca jellyfish and macrozooplankton (≥1000 µm) had similar δ15N values, highlighting a shared, high trophic level for these two categories (Fig. 2). 15N-depleted values of mesozooplankton (200–1000 µm) and microplankton (<200 µm) revealed a lower trophic level than jellyfish and macrozooplankton. Medusa samples from different periods of the year showed clear separation (Fig. 2) in three main clusters represented by jellyfish sampled in spring (April-May), spring-summer (June-July-August-September), or in autumn and winter months (November-December-February).
Figure 2.

Bi-plot of stable isotope composition for Pelagia noctiluca in different season and size class of plankton preys.
The results of the SIAR mixing model showed that P. noctiluca primarily consumed microplankton (95% credibility interval: 38–53%) and mesozooplankton (95% credibility interval: 33–51%) and, to a lesser extent, macrozooplankton (95% credibility interval: 7–17%) (Fig. 3).
Figure 3.
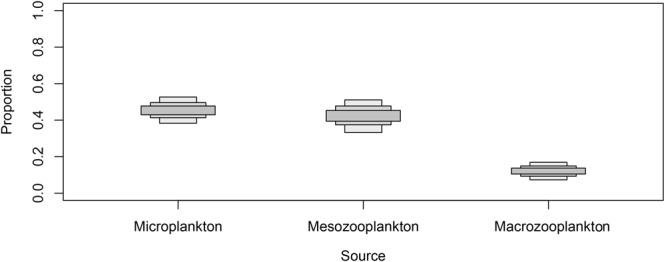
Modeled proportion of prey in the diet of Pelagia noctiluca medusae obtained using a stable isotope mixing model.
Fatty acids (FAs)
The FA concentration (µg of total FAs per mg of dry tissue) was higher and more variable in the gonads than in somatic tissue for both male and female jellyfish (Fig. 4). Female gonads accumulated FAs during three periods: spring (April: 10 ± 0.3, May: 8 ± 0.8 and June: 11 ± 2 µg fatty acid *mg−1 dry tissue), late summer (September: 7 ± 0.9 µg fatty acid * mg−1 dry tissue) and late autumn (December: 11 ± 1 µg fatty acid * mg−1 dry tissue), while male gonads had only two peaks (May: 7 ± 1 and December: 12 ± 2 µg fatty acid * mg−1 dry tissue) (Fig. 4).
Figure 4.
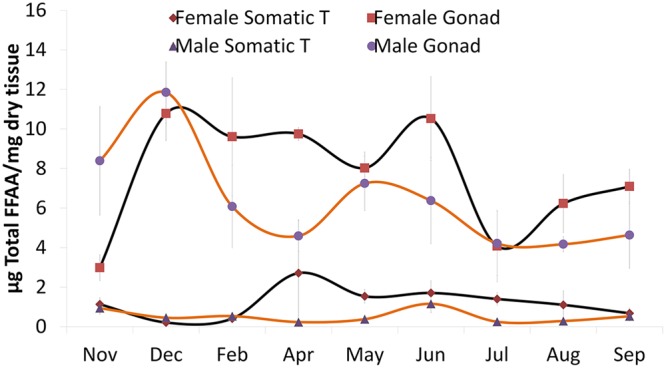
Monthly total fatty acid concentration (mean ± standard error) in different Pelagia noctiluca medusa tissues (somatic, gonad) and sex (male, female) from the Strait of Messina.
The FAs encountered in the present work are presented in Supplementary Information 2. In general, polyunsaturated fatty acids (PUFAs) were the most abundant compounds, representing from 49.5% (November) to 68% (September) of total FAs. Major PUFAs were C20:4n-6 (Arachidonic acid; AA), C22:6n-3 (Docosahexaenoic acid; DHA), and C20:5n-3 (Eicosapentaenoic acid; EPA), which accounted for 37.5 to 41.6% of total FAs. Monounsaturated fatty acids (MUFAs) accounted for 2.3 to 26.3% of total FAs and consisted mostly of C16:1n-7 and C18:1(n-9). Saturated fatty acids (SFAs) were the next most abundant group, consisting mainly of C16:0, C17:0, and C18:0, which accounted for 24.8 to 28.1% of total FAs.
A 3-way PERMANOVA analysis on Log +1 transformed FA data suggested significant dietary shifts of P. noctiluca over the year (Table 2) and that FA composition of jellyfish differed significantly for the three factors considered: Month, Sex, and Body Part (Table 2).
SIMPER analysis showed that the FAs C16:0, C20:5n-6, C20:4n-3, and C22:6n-3 contributed most to the Bray-Curtis dissimilarity between Body Parts, between Sex and, in most cases, among months.
FA composition in zooplankton differed significantly among months (Table 2), although it did not differ among the zooplankton fractions. SIMPER analysis, conducted on a Bray Curtis similarity matrix, indicated homogeneity in FA composition among different spring zooplankton size classes (Similarity: 74.6%). C22:6n-3 contributed most to differentiate spring samples from other seasons, representing 29.42% of total FAs.
The highest ratios of n-3/n-6 (Omega 3/Omega 6 FAs) were recorded in somatic tissue in July, followed by December and February and in the gonads in February-April, September, and December (Fig. 5a). PUFA/SFA (Polyunsaturated/Saturated fatty acid) were higher in June, August, and February in the somatic tissue and in April and August-September in gonads (Fig. 5b). The lowest EPA/DHA (Eicosapentaenoic/Docosahexaenoic acid) ratios were found in May and July in somatic tissues and in May and August in gonads (Fig. 5c).
Figure 5.
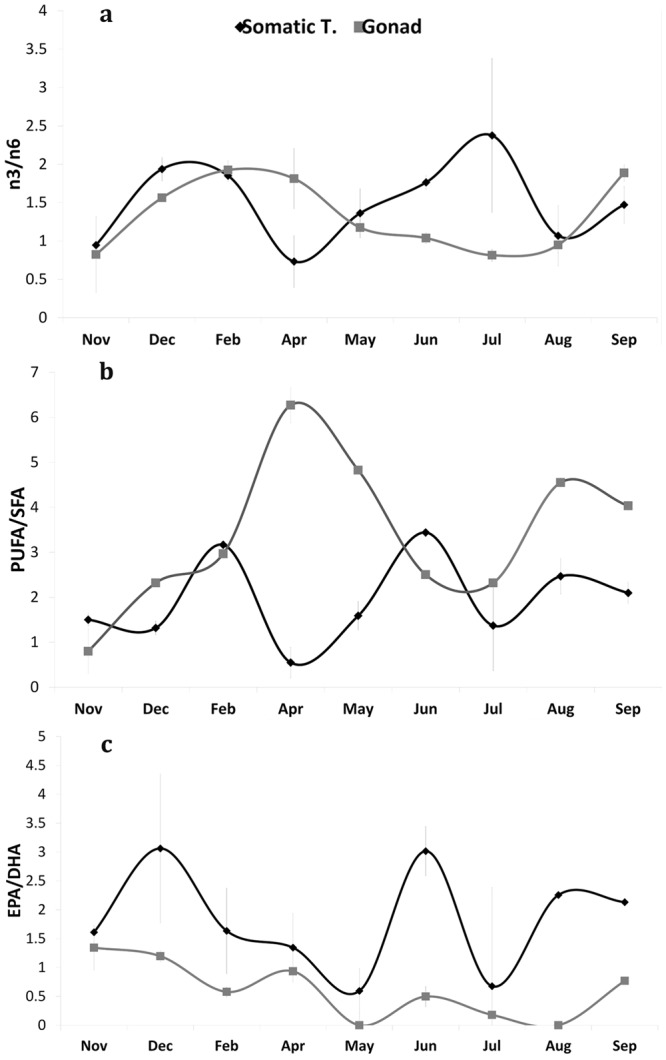
Monthly fatty acid variation (mean ± standard error) of (a) Omega 3/Omega 6 (n3/n6) ratio; (b) polyunsaturated fatty acid/saturated fatty acid (PUFA/SFA) ratio and (c) eicosapentaenoic acid/ docosahexaenoic acid (EPA/DHA) ratio in gonads and somatic tissue of Pelagia noctiluca medusae from the Strait of Messina.
Discussion
The most available prey in the Strait of Messina, e.g., copepods, pteropods, fish eggs, and chaetognaths (Supplementary Information 1), were the main food sources for Pelagia noctiluca medusae in the present study. This jellyfish species has been described as a non-selective predator27,29,36,37, feeding on almost all types of zooplankton and ichthyoplankton27,30,31,38. The stomach contents of P. noctiluca showed a great variety of ingested prey, such as cladocerans, appendicularians, copepods, hydromedusae, siphonophores, and fish eggs. This result is in agreement with most of the above mentioned previous findings27,29,39.
Pearre’s selection index and data from the NW Mediterranean Sea27 concur that P. noctiluca in the Strait of Messina can be considered an unselective predator for most zooplankton taxa, capturing prey in proportion to their availability (Table 1). Apparently, euphausiid furcilia larvae and ostracods were negatively selected by P. noctiluca. However, this result may be biased by the diel vertical migration of those taxa, which are more commonly recorded in shallow water at night40,41, whereas during daytime when medusae were sampled, euphausiid larvae and ostracods move into deeper waters, possibly escaping from predation by jellyfish we collected. Also, the neutral selection of fish larvae as prey for P. noctiluca can be explained by an active anti-predator behavior, which can allow perception of slow predators and their avoidance42,43.
By contrast, fish eggs and copepods were positively selected as preferred prey, confirming the potential impact of P. noctiluca on fish recruitment either directly, by predation on fish eggs44,45, or indirectly by competition with fish larvae for the same food source (copepods)46. Such positive selection was most evident during the periods of sexual reproduction of P. noctiluca35, suggesting a correlation between high quality food and spawning periods. The high occurrence of fish eggs in the stomach of P. noctiluca might reflect the long digestion time for this prey type, as already known for P. noctiluca ephyrae47. Overall, these data corroborate the lack of strong prey selectivity in P. noctiluca medusae. Generalist predation may provide the simplest mechanistic explanation for the significant differences in P. noctiluca prey composition over the year, reflecting the seasonal availability of different prey and a clear opportunistic behavior.
δ13C and δ15N values of P. noctiluca jellyfish were compared with those of three different plankton size classes. Marine animals are 0–2‰ enriched in carbon isotopes31,48 and 3.2‰ enriched in nitrogen isotopes49 relative to those in their diets through the food web. Average δ13C values for jellyfish were about 1.8‰ higher than the values in mesozooplankton and fish eggs (200–1000 µm) and 2.4‰ higher than microplankton (<200 μm size class). Average δ15N values for jellyfishes were about 2.8‰ higher than the values in zooplankton 200–1000 μm size class and 3.6‰ higher than in microplankton <200 μm size class, indicating the enrichment of the heavy isotopes in the P. noctiluca tissues. Trophic fractionation of carbon isotope ratios between predators and their prey was considered to be ~2‰ for P. noctiluca31; thus, our results indicate that the main diet of P. noctiluca is composed of a mix of zooplankton size classes, which is consistent with the stomach content results. The same results come from our analysis of the nitrogen SI in P. noctiluca, with a rate of fractionation of 3.2‰. The isotopic fingerprint of microplankton detected in the medusa tissues can be the result of direct ingestion of bio-seston or indirect assimilation from the stomachs of mesozooplankton grazers that were consumed by P. noctiluca, like herbivorous copepods or appendicularians50. Grazing on microplankton is known for many cnidarians, such as Aurelia moon jellyfish from the eastern Atlantic, which ingested various microplankton taxa (e.g., particularly non-loricate ciliates, rotifers, flagellates, and others)51. Moreover, microplankton may represent an important food source for this jellyfish, particularly when mesozooplankton taxa are at low concentration23,52.
Analysis of seasonal variability of food preferences is uncommon for jellyfish. Our results suggest that δ13C in jellyfish varied from a high in autumn-winter months (~−18‰) to its lowest value in April (~−20‰). This variation may be related to the Strait of Messina having an alternating tidal regime driven by strong upwelling currents, with a seasonal peak from late spring to late summer53. Upwelled waters are characterized by a high micro-phytoplankton fraction54,55, with decreased δ13C and δ15N values at the base of the food web56. The seasonal upwelling peak may be followed by an opportunistic increase of direct consumption/indirect assimilation of micro-phytoplankton by P. noctiluca that is mirrored by the seasonal reduction of δ13C values in the jellyfish tissues. The SI results also agree with jellyfish stomach content data, in which most of the identifiable prey consisted of copepods and other herbivorous crustaceans. This can be interpreted as additional evidence of the lack of selection in the diet of P. noctiluca and the plasticity of its foraging and assimilation ability, because detritus is more abundant in autumn-early winter and diatom blooms are more abundant in late winter-spring months55.
High δ15N values and high trophic levels were found in summer in accordance with the increase in mesozooplankton abundance. Other studies on temporal changes in SI composition of dietary sources in food webs agree with the seasonal variation in C and N values in P. noctiluca tissues from the upwelling system in the Strait of Messina. Fukuda and Naganuma57 found that the diet of Aurelia medusae from the Sea of Japan shifted seasonally from a diatom-based food chain in spring to a detritus-based food chain in autumn. Recently, Javidpour et al.33 also suggested a temporal dietary shift from mesozooplankton to microplankton and re-suspended organic matter from the benthos for A. aurita from the Kiel Fjord (Baltic Sea). Other studies demonstrate feeding on microplankton58–60, but so far, only one study59 has demonstrated the ability of cnidarians to assimilate phytoplankton. Fresh material from algal blooms or reworked seston from resuspension or low quality material have different signals and are key to understand the trophic guilds of aquatic species61. Pelagia noctiluca can assimilate all zooplankton according to their abundance in the environment and smaller foods being digested in the stomach of its prey, utilizing an omnivorous assimilation strategy most of the year and an opportunistic carnivorous assimilation strategy during summer months.
The fatty acids also yielded results in accordance with all of the previous findings. Trophic biomarkers may be useful to understand the relationships of organisms in the food web and also to trace seasonal and ontogenetic changes in their physiological states14,17. Most of the previous research on lipid storage and FA metabolism in marine organisms dealt with taxa other than cnidarian jellyfish62,63. In addition, most studies on jellyfish FAs analyzed extracts from whole specimens without considering possible differences in FAs in different body parts, as appropriate to their respective physiological roles64. The study of trophic interactions through compositional analysis of fatty acids in different jellyfish tissues was carried out only for the colonial siphonophore Nanomia cara16, separated into two different body parts: the nectosome (specialized for swimming) and the siphosome (specialized for feeding and reproduction), with the siphosome yielding more fatty acids than the nectosome. Here we show for the first time the relationship between FAs and the diet of P. noctiluca medusae over a year, as well as the importance of considering FA extracts from somatic and gonadal tissues separately. The FA extracts from P. noctiluca soma and gonads highlighted their differences in terms of FA composition and storage capacity, respectively. The somatic tissue had low concentrations of FAs throughout the year, while the gonadal tissue had total FA concentrations that at times during the annual cycle exceeded ten times that in the somatic tissue of medusae, in agreement with previous findings based on other methods, e.g. on total biochemical composition, ash-free dry mass, or C:N ratios65–67.
Interestingly, EPA and AA values in the zooplankton fractions (9 ± 2 µg*mg−1dry tissue and 2 ± 3 µg*mg−1dry tissue, respectively) were lower than in P. noctiluca medusae (17 ± 2 µg*mg−1 dry tissue and 11 ± 1 µg*mg−1 dry tissue, respectively). This may be due to selective FA accumulation by jellyfish through indirect assimilation or direct predation of primary producers or microzooplankton (as shown by the gut contents). A comparable accumulation mechanism is known in other groups (e.g., copepods and cladocerans)10,63; however, molecular studies are now providing progress towards understanding the mechanisms of long-chain PUFA biosynthesis in marine invertebrates. Candidate genes for LC-PUFA biosynthetic enzymes (e.g., desaturases and elongases) have been confirmed from crustaceans and molluscs63. Thus, the possibility that part of the EPA and AA was generated through biosynthetic pathways in the medusa tissues, rather than representing assimilation from prey, needs to be investigated.
Palmitic Acid and DHA values were high in zooplankton samples, therefore jellyfish can accumulate these two FAs from the direct feeding on meso- and macrozooplankton14,57. The transfer of FAs from prey to P. noctiluca was not surprising. Siphonophores are known to assimilate over 90% of the carbon and nitrogen from their prey68. Therefore, the ability of selected FA assimilation and storage by P. noctiluca may be a crucial reproductive strategy, taking into consideration that part of FA energy will be used for the gonadal investment and the development of future ephyra stages.
FA biomarkers may provide complementary information about seasonal alternation between carnivory and omnivory. Their composition in P. noctiluca tissues varied for all factors considered in this study: SIMPER analysis highlighted the high contributions of C16:0 (Palmitic Acid), C20:4n-3 (Arachidonic Acid, AA), C20:5n-6 (Eicosapentanoic Acid, EPA), and C22:6n-3 (Docosahexaenoic Acid, DHA) in Bray-Curtis Dissimilarity between sex, body parts, and among months. EPA and DHA were most abundant in the gonads. These are essential FAs in fish broodstock diets, having critical functions as the main components of phospholipids of cell membranes69. The EPA/DHA ratio potentially may be used to determine the degree of carnivory, because DHA is highly conserved through the food web, being preferentially incorporated into polar lipids70. Consequently, the EPA/DHA ratio should decrease towards higher trophic levels14. Further, Cripps and Atkinson71 proposed that the ratio of polyunsaturated to saturated fatty acids (PUFA/SFA) in krill may be a useful indicator for carnivorous (high PUFA/SFA) versus herbivorous (low PUFA/SFA) feeding. In the gonads of P. noctiluca, both EPA/DHA and PUFA/SFA ratios suggest medium to high trophic levels (i.e., from secondary to tertiary consumer levels) for P. noctiluca medusae, in agreement with the results obtained from the SI analysis. Overall, the complementary information presented here reinforces the idea of a medium to high trophic position of P. noctiluca medusa, depending on the available food sources in the water column.
The reproductive tissues showed a wider seasonal oscillation of total FAs than did the somatic ones, related to the seasonal, biphasic cycle of spawning events observed in P. noctiluca35. The great accumulation capacity of FAs in the gonads may provide long-term signals of the quality and quantity of the diet, but the biochemical composition of gonads may be largely variable according to the reproductive cycle and the available food for an opportunistic species. Seasonal FA biomarker changes in jellyfish has been correlated with the population fitness throughout the year17. A strong seasonal pattern of FA biomarkers has been found in Aurelia moon jellyfish57, with lower quality FA composition in autumn (due to a prominent detritus-based nutrition) than in spring (due to a paramount diatom-based food source). This biochemical difference might be critical for the activation of sexual reproduction and eventually for increased larval supply and recruitment. In Aurelia and other scyphozoan with a benthic polyp stage, any increment of polyp populations will facilitate jellyfish outbursts by means of temperature-dependent strobilation timers72. Data on FAs stored in P. noctiluca medusae from the Strait of Messina demonstrated that energy flux was higher in spring-early summer and linked to the phytoplankton blooms, while the autumn decrease of n-3 FAs and increased abundance in SFAs (November-December, coincident with the refractory organic matter period10) was consistent with a prevalent detritus-based diet. The spring-early summer peak of copepods in the environment54 and in the stomach contents was also reflected in the amount of copepod-specific FA biomarkers. The abundance of copepods and fish eggs in the stomach contents is also reflected in the total FA concentration in the gonads, with a high energy investment for sexual reproduction.
In conclusion, stomach contents and biomarker analysis showed that P. noctiluca in the study area is not a selective predator, cyclically shifting its foraging strategy between carnivory and omnivory, depending on the seasonal availability of food items. Such cyclical changes seem to be tuned with the phenology of sexual reproduction of jellyfish, and to their abundance-size spectra. In turn, the high abundance of P. noctiluca may exert a top-down control on the zooplankton populations and the pelagic food webs.
Stomach contents and the selectivity index do not support for P. noctiluca in the Strait of Messina a role as main predator of fish larvae, as elsewhere hypothesized30,37. However, an impact on fish populations cannot be ruled out, as it may take place through three alternative, non-mutually exclusive ways: (1) direct top-down interaction by predation on fish eggs; (2) indirect competitive interaction with fish larvae, by dietary overlap for mesozooplankton as a food source; (3) indirect, bottom-up control, by grazing on microplankton so decreasing the available energy for upper trophic levels. The combination of SI and, potentially, FA analyses highlighted for the first time the importance of microzooplankton as prey for P. noctiluca, a neglected or underestimated role by direct stomach content analysis, showing that multiple mechanisms may coincide during jellyfish outbreaks to reduce food availability to higher trophic levels. The recognition of the opportunistic feeding behavior in a dominant, outbreak-forming species, such as P. noctiluca, corroborates the hypothesis that the variability of jellyfish trophic roles in plankton communities may be key both to their short-term ecological dominance and long-term evolutionary success73.
Methods
Ethics statement
No specific permits were required for the described field studies in the Strait of Messina. The species collected is the most common native jellyfish in the Mediterranean Sea and is not protected throughout its range. Sampling points did not include any protected or private lands.
Study Area
The study was carried out in the Strait of Messina (Sicily, Italy). This site is strongly influenced by the peculiar hydrodynamic regime of the Strait, characterized by a six-hour alternation of northward (from the Ionian to Tyrrhenian seas) and southward (from the Tyrrhenian to Ionian seas) tidal currents, with upwelling and downwelling water masses reaching up to 200 cm s−1 speed29. The hydrodynamic complexity of the Strait ecosystem has a major influence on the spatial and vertical distribution of the organisms, especially on zooplankton communities53. The regular alternation of water masses prevents stratification of the water column and the formation of a summer thermocline. For this reason, the Strait is characterized by a higher productivity than other Mediterranean coastal areas.
In this study, surface water temperature was measured monthly using a bucket thermometer, and our data were implemented with daily time-series sea surface temperature downloaded from www.mareografico.it (ISPRA).
Sampling
Live P. noctiluca specimens were collected two times per month from November 2011 to September 2012 with a 1-cm mesh hand net dipped from the boat. Due to adverse weather conditions and the absence of jellyfish from surface waters, we were unable to collect P. noctiluca in October 2011, January and March 2012. On board, jellyfish diameters were measured using a rule to the nearest mm. The specimens for stomach content analysis were fixed immediately after capture in 4% formaldehyde/water solution and stored individually in a container, to search all possible prey also in the formaldehyde/water solution, due to an event of stomach egestion during preservation.
Specimens for biomarker analysis were placed in a 50-l tank, with a continuous flow of seawater. Immediately after sampling (~2 hours), jellyfish were brought to the laboratory to proceed with morphometric and morphological analysis and sex identification74.
Three male and three female living jellyfish were selected from each sampling (i.e., six males and six females each month). The gonadal tissue was dissected from the somatic tissue for each specimen and each separately frozen in liquid nitrogen for subsequent trophic biomarker analyses. To obtain particulate organic matter for SI analysis, 3 liters of 200-µm-filtered sea water were re-filtered under vacuum through pre-ashed (4 h at 500 °C) Whatman GF/F glass fiber filters.
To provide additional information on P. noctiluca food availability and dietary preferences, two zooplankton samples, one each for taxonomic and biochemical analyses, were taken monthly in the same zone where jellyfish were sampled, using a WP2 net (40-cm mouth opening; 200 µm mesh size) with 10-minute surface horizontal hauls (1–2 m depth) at an average speed of 1 knot, in the same time and space of medusa samples. Zooplankton samples for taxonomic identification were fixed in 4% formaldehyde/water solution. Zooplankton samples for biochemical analyses were divided according into three size classes: <200 µm, 200–1000 µm and >1000 µm, and each fraction was filtered in pre-weighted GF-A filter.
Stomach contents and stable isotope analysis
The recognizable prey taxa within the gastric pouches, manubrium, oral arms and mucus of 149 specimens of homogeneous size (see Table 1, including the number of samples per month) were identified to the finest possible taxonomic level with magnification of a stereomicroscope. Values of Pearre’s electivity index (C)75 were calculated for the most common zooplankton taxa by the numbers of prey m−3 and the average numbers of prey medusa−1 for each sample, using an ordinary chi-square test to assess their statistical significance.
Jellyfish somatic tissue (5 medusae for each month), zooplankton samples and filters with particulate organic matter were oven-dried at 60 °C and homogenized using mortar and pestle. To remove carbonate structures, one sub-sample of each sample for carbon isotope analysis in zooplankton and particulate organic matter was acidified by adding 1 M HCl drop-by-drop76. The samples were re-dried at 60 °C for 24 h.
All samples (medusa, zooplankton, and microplankton) were weighed in tin capsules, combusted and the resultant gases were analyzed in a Thermo Electron Isotope Ratio Mass Spectrometer for SI abundance. Internal laboratory standards were used after every five samples to calibrate the system. Isotopic values were expressed in δ notation as parts per thousand (‰) differences from international standards (Vienna Pee Dee Belemnite and atmospheric N2 for carbon and nitrogen respectively):
| 1 |
where X is equal to 15N or 13C and R is the corresponding ratio 13C/12C or 15N/14N. Based on replicates of laboratory standards, analytical precision was ±0.2 and ± 0.1‰ for δ15N and δ13C respectively. Trophic levels (TL) of P. noctiluca were estimated assuming an average increase in δ15N values of 3.2‰ between successive trophic levels49,77, and an average increase in δ13C values of 2‰ between successive trophic levels31. In agreement with23, we did not use the trophic enrichment factors value for nitrogen proposed by D’ Ambra et al.78, because the trophic level of P. noctiluca would result in unrealistically high trophic positions.
TL were estimated according to the following formula:
| 2 |
where δ15Nf and δ15Nref are the nitrogen isotopic signature of the medusa and a baseline reference organism respectively; 3.2 is the δ15N expected isotopic fractionation per trophic level; TFref is the trophic level of the baseline organism. In this study the baseline organisms were the mesozooplanktonic calanoid copepods (TFref = 2).
Fatty acid extraction
Approximately 100 mg dry weight (±0.01) of medusa tissue (Gonad or Somatic tissue) and 50 mg dry weight (±0.01) of different zooplankton size fractions were extracted by microwave-assisted extraction (5 min at 70 °C) with 10 mL of 3:1 dichloromethane-methanol, and using 2-octyldodecanoic acid and 5β-cholanic acid as internal standards. The extract was taken to near dryness in a centrifugal vacuum concentrator at a constant temperature and fractionated by solid phase extraction according to Ruiz et al.79. The sample was re-dissolved on 0.5 mL of chloroform and eluted through a 500 mg amino-propyl column previously conditioned with 4 mL of n-hexane. The first fraction was eluted in 3 mL of chloroform: 2-propanol (2:1), and the fatty acids recovered in 8.5 mL of diethyl ether: acetic acid (98:2). The FA fraction was methylated using a solution of 20% methanol/BF3 heated at 90 °C for 1 h. The reaction was quenched with 4 mL of water saturated with NaCl. The methyl esters of FFA were recovered by extracting twice in 3 mL of n-hexane. The combined extracts were taken to near dryness, re-dissolved in 1.5 mL of chloroform, eluted through a glass column filled with Na2SO4 to remove residual water, taken to dryness under a gentle nitrogen flux, and stored at −20 °C until analysis. The samples were re-dissolved in 40 μl (for medusa tissue) or 100 μl (for zooplankton size fraction) of isooctane. Gas chromatographic analysis was performed with a Thermo Finnigan Trace Gas Chromatography Ultra instrument equipped with a flame ionization detector, a splitless injector and a DB–5 Agilent column (30 m length, 0.25 mm internal diameter and 0.25 μm phase thickness). Helium was used as a carrier gas at 33 cm s−1. The oven temperature was programmed to increase from 50 °C to 320 °C at 10 °C−min. The injector and detector temperatures were 300 °C and 320 °C, respectively. The methyl esters of fatty acids were identified by comparing their retention times with those of FA standards (37 FAME compounds, Supelco® Mix C4-C24). Fatty acids were quantified by integrating areas under the peaks in the gas chromatograph traces (Chromquest 4.1 software) with calibrations derived from internal standards80. The Mix C4-C24 satisfied almost all the fatty acid identification; only in few cases GC-MS has been used to identify unknown peaks.
Statistical analysis
Differences in the total numbers of prey in the stomach contents of each jellyfish (number of prey medusae−1) among months were analyzed by one-way analysis of variance (ANOVA), after testing homogeneity of variance by Cochran’s test, while differences of monthly diet composition were analyzed by one-way Permutational Multivariate ANOVA (PERMANOVA81) performed on a triangular matrix based on Euclidean distances.
To define the diet of P. noctiluca based on δ13C and δ15N of medusae and their potential prey, we computed a mixing model using the package SIAR82 (version 4.1) downloaded with R (“R: A language and environment for statistical computing” 2013; version 2.13.1) from the Comprehensive Archive Network site (CRAN, http://cran.r-project.org/). The mixing model was based on prey composition in the predator’s diet inferred from all feasible prey. The proportional diet composition of medusae was estimated using the following fractionation values: δ13C 2 + 0.3‰31, and δ15N 3.2 + 0.1‰49. Moreover, differences in δ13C and δ15N values among different sampled months, were investigated separately using one-way ANOVA. The total amount of FAs was investigated using 3-way PERMANOVA. In this case the independent variables used were: Month (fixed with 8 levels), Sex (fixed and orthogonal with two levels), and Body part (fixed and orthogonal with two levels: gonadic and somatic tissue). Differences in FA composition of medusae among Months, Sex, and Body parts were tested using 3-way PERMANOVA based on Bray-Curtis similarities matrix of log +1 transformed data. In this case we preferred the use of Bray-Curtis similarities matrix because of the large number of variables considered (FAs) and the presence of zero, following the recommendation of Legendre & Legendre83. The SIMPER routine was used to identify the dissimilarity among months, as well as the FAs responsible for the observed differences. FA composition of zooplankton sampling were analyzed using 2-way PERMANOVA, in order to highlight differences among different months and zooplankton size and their interaction.
All analyses were performed using PRIMER6 & PERMANOVA+84,85 and R (version 2.13.1; CRAN, http://cran.r-project.org/).
Electronic supplementary material
Acknowledgements
This work has received funding from the European Union’s projects MED-JELLYRISK (grant n. I-A/1.3/098 - ENPI CBCMED programme), VECTORS (Vectors of Change in Oceans and Seas Marine Life, Impact on Economic Sectors, grant n. 266445, FP7th programme) and CERES (Climate Change and European Aquatic Resources, grant n. 678193, Horizon 2020 programme).
Author Contributions
G.M., V.L.F., S.P. and S.R. conceived the ideas and designed the methodology; G.M. collected the data; G.M., U.T. and S.R. performed the biochemical analyses; G.M. analyzed the data; G.M., S.V., J.E.P., S.R. and S.P. led the writing of the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
Competing Interests
The authors declare no competing interests.
Footnotes
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Giacomo Milisenda, Email: giacomo.milisenda@gmail.com.
Stefano Piraino, Email: stefano.piraino@unisalento.it.
Electronic supplementary material
Supplementary information accompanies this paper at 10.1038/s41598-018-30474-x.
References
- 1.Purcell JE, Uye S, Lo W. Anthropogenic causes of jellyfish blooms and their direct consequences for humans: a review. Mar. Ecol. Prog. Ser. 2007;350:153–174. doi: 10.3354/meps07093. [DOI] [Google Scholar]
- 2.Pauly D, Graham W, Libralato S, Morissette L, Deng Palomares ML. Jellyfish in ecosystems, online databases, and ecosystem models. Hydrobiologia. 2008;616:67–85. doi: 10.1007/s10750-008-9583-x. [DOI] [Google Scholar]
- 3.Boero, F. Review of jellyfish blooms in the Mediterranean and Black Sea Available at: http://www.vliz.be/imis/imis.php?module=ref&refid=226251 (2013).
- 4.Boero F, et al. Gelatinous plankton: irregularities rule the world (sometimes) Mar. Ecol. Prog. Ser. 2008;356:299–310. doi: 10.3354/meps07368. [DOI] [Google Scholar]
- 5.Purcell JE. Pelagic cnidarians and ctenophores as predators: selective predation, feeding rates, and effects on prey populations. Ann. Inst. océanogr., Paris. 1997;73:125–137. [Google Scholar]
- 6.Purcell JE. Extension of methods for jellyfish and ctenophore trophic ecology to large-scale research. Hydrobiologia. 2009;616:23–50. doi: 10.1007/s10750-008-9585-8. [DOI] [Google Scholar]
- 7.Pitt KA, Connolly RM, Meziane T. Stable isotope and fatty acid tracers in energy and nutrient studies of jellyfish: a review. Hydrobiologia. 2008;616:119–132. doi: 10.1007/s10750-008-9581-z. [DOI] [Google Scholar]
- 8.Sullivan BK, Garcia JR, Klein-MacPhee G. Prey selection by the scyphomedusan predator Aurelia aurita. Mar. Biol. 1994;121:335–341. doi: 10.1007/BF00346742. [DOI] [Google Scholar]
- 9.Toonen RJ, Chia F-S. Limitations of laboratory assessments of coelenterate predation: Container effects on the prey selection of the Limnomedusa, Proboscidactyla flavicirrata (Brandt) J. Exp. Mar. Biol. Ecol. 1993;167:215–235. doi: 10.1016/0022-0981(93)90032-J. [DOI] [Google Scholar]
- 10.Rossi S, Sabatés A, Latasa M, Reyes E. Lipid biomarkers and trophic linkages between phytoplankton, zooplankton and anchovy (Engraulis encrasicolus) larvae in the NW Mediterranean. J. Plankton Res. 2006;28:551–562. doi: 10.1093/plankt/fbi140. [DOI] [Google Scholar]
- 11.Alonzo F, Virtue P, Nicol S, Nichols P. Lipids as trophic markers in Antarctic krill. III. Temporal changes in digestive gland lipid composition of Euphausia superba in controlled conditions. Mar. Ecol. Prog. Ser. 2005;296:81–91. doi: 10.3354/meps296081. [DOI] [Google Scholar]
- 12.Fry B, Arnold C. Rapid 13C/12C turnover during growth of brown shrimp (Penaeus aztecus) Oecologia. 1982;54:200–204. doi: 10.1007/BF00378393. [DOI] [PubMed] [Google Scholar]
- 13. Båmstedt, U., Gifford, D. J., Irigoien, X., Atkinson, A. & Roman, M. Feeding. In ICES Zooplankton Methodology Manual 1st Edition (eds Harris, R., Wiebe, P., Lenz, J., Skjoldal, H. R. & Huntley, M. B. T.) 297–399 (Academic Press, 2000). 10.1016/B978-012327645-2/50009-8.
- 14.Dalsgaard J, St John M, Kattner G, Müller-Navarra D, Hagen W. Fatty acid trophic markers in the pelagic marine environment. Adv. Mar. Biol. 2003;46:225–340. doi: 10.1016/S0065-2881(03)46005-7. [DOI] [PubMed] [Google Scholar]
- 15.Kattner, G. & Hagen, W. Lipids in marine copepods: latitudinal characteristics and perspective to global warming. In Lipids in Aquatic Ecosystems (eds Kainz, M., Brett, T. M. & Arts, T. M.) 257–280 (Springer New York, 2009). 10.1007/978-0-387-89366-2_11.
- 16.Rossi S, Youngbluth MMJ, Jacoby CA, Pagès F, Garrofé X. Fatty acid trophic markers and trophic links among seston, crustacean zooplankton and the siphonophore Nanomia cara in Georges Basin and Oceanographer Canyon. Sci. Mar. 2008;72:403–416. doi: 10.3989/scimar.2008.72n2403. [DOI] [Google Scholar]
- 17.Kelly J, Scheibling R. Fatty acids as dietary tracers in benthic food webs. Mar. Ecol. Prog. Ser. 2012;446:1–22. doi: 10.3354/meps09559. [DOI] [Google Scholar]
- 18.Ben-David M, Flynn RW, Schell DM. Annual and seasonal changes in diets of martens: evidence from stable isotope analysis. Oecologia. 1997;111:280–291. doi: 10.1007/s004420050236. [DOI] [PubMed] [Google Scholar]
- 19.Szepanski MM, Ben-David M, Van Ballenberghe V. Assessment of anadromous salmon resources in the diet of the Alexander Archipelago wolf using stable isotope analysis. Oecologia. 1999;120:327–335. doi: 10.1007/s004420050866. [DOI] [PubMed] [Google Scholar]
- 20.Fry B. Food web structure on Georges Bank from stable C, N, and S isotopic compositions. Limnol. Oceanogr. 1988;33:1182–1190. doi: 10.4319/lo.1988.33.5.1182. [DOI] [Google Scholar]
- 21.Hobson K, Harold E. Determination of the trophic relationships within a high Artic marine food web using δ13C and δ15N analysis. Mar. Ecol. Prog. Ser. 1988;84:9–18. doi: 10.3354/meps084009. [DOI] [Google Scholar]
- 22.Søreide JE, Hop H, Carroll ML, Falk-Petersen S, Hegseth EN. Seasonal food web structures and sympagic–pelagic coupling in the European Arctic revealed by stable isotopes and a two-source food web model. Prog. Oceanogr. 2006;71:59–87. doi: 10.1016/j.pocean.2006.06.001. [DOI] [Google Scholar]
- 23.Fleming NEC, Harrod C, Newton J, Houghton JDR. Not all jellyfish are equal: isotopic evidence for inter- and intraspecific variation in jellyfish trophic ecology. PeerJ. 2015;3(e1110):1–21. doi: 10.7717/peerj.1110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.D’Ambra I, Graham WM, Carmichael RH, Malej A, Onofri V. Predation patterns and prey quality of medusae in a semi-enclosed marine lake: implications for food web energy transfer in coastal marine ecosystems. J. Plankton Res. 2013;35:1305–1312. doi: 10.1093/plankt/fbt065. [DOI] [Google Scholar]
- 25.Ying C, Ying W, Jing Z, Na W. Potential dietary influence on the stable isotopes and fatty acid compositions of jellyfishes in the Yellow Sea. J. Mar. Biol. Assoc. United Kingdom. 2012;92:1325–1333. doi: 10.1017/S0025315412000082. [DOI] [Google Scholar]
- 26.Naman SM, et al. Stable isotope-based trophic structure of pelagic fish and jellyfish across natural and anthropogenic landscape gradients in a fjord estuary. Ecol. Evol. 2016;6:8159–8173. doi: 10.1002/ece3.2450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Tilves U, et al. Natural diet and predation impacts of Pelagia noctiluca on fish eggs and larvae in the NW Mediterranean. J. Plankton Res. 2016;38:1243–1254. doi: 10.1093/plankt/fbw059. [DOI] [Google Scholar]
- 28.Canepa, A. et al. Pelagia noctiluca in the Mediterranean Sea. in Jellyfish Blooms (eds Pitt, K. A. & Lucas, C. H.)237–266 (Springer Netherlands, 2014). 10.1007/978-94-007-7015-7_11.
- 29.Rosa S, Pansera M, Granata A, Guglielmo L. Interannual variability, growth, reproduction and feeding of Pelagia noctiluca (Cnidaria: Scyphozoa) in the Straits of Messina (Central Mediterranean Sea): Linkages with temperature and diet. J. Mar. Syst. 2013;111–112:97–107. doi: 10.1016/j.jmarsys.2012.10.001. [DOI] [Google Scholar]
- 30.Sabatés A, et al. Planktonic cnidarian distribution and feeding of Pelagia noctiluca in the NW Mediterranean Sea. Hydrobiologia. 2010;645:153–165. doi: 10.1007/s10750-010-0221-z. [DOI] [Google Scholar]
- 31.Malej A, Faganelli J, Pezdic J. Stable isotopes and biochemical fractionation in the marine pelagic food chain: the jellyfish Pelagia noctiluca and net zooplankton. Mar. Biol. 1993;116:565–570. doi: 10.1007/BF00355475. [DOI] [Google Scholar]
- 32.Spallanzani L. Méduses phosphoriques observées dans le détroit de Messine. Voyages dans les deux Siciles et dans quelques uarties des Apennins. 1799;1:137–171. [Google Scholar]
- 33.Javidpour J, Cipriano-Maack AN, Mittermayr A, Dierking J. Temporal dietary shift in jellyfish revealed by stable isotope analysis. Mar. Biol. 2016;163:1–9. doi: 10.1007/s00227-016-2892-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Viladrich, N. et al. Variation in lipid and free fatty acid content during spawning in two temperate octocorals with different reproductive strategies: surface versus internal brooder. Coral Reef. 35(3), issn=07224028, 10.1007/s00338-016-1440-1.
- 35.Milisenda G, et al. Reproductive and bloom patterns of Pelagia noctiluca in the Strait of Messina, Italy. Estuar. Coast. Shelf Sci. 2018;201:29–39. doi: 10.1016/j.ecss.2016.01.002. [DOI] [Google Scholar]
- 36.Daly Yahia MN, Goy J, Daly Yahia-Kéfi O. Distribution and ecology of Medusae and Scyphomedusae (Cnidaria) in Tunis Gulf (SW Mediterranean) Oceanol. Acta. 2003;26:645–655. doi: 10.1016/j.oceact.2003.05.002. [DOI] [Google Scholar]
- 37.Larson RJ. A note on the feeding, growth, and reproduction of the epipelagic scyphomedusa Pelagia noctiluca (Forskal) Biol. Oceanogr. 1987;4:447–454. [Google Scholar]
- 38.Giorgi, R., Avian, M., De Olazabal, S. & Rottini-Sandrini, L. Feeding of Pelagia noctiluca in Open Sea. in UNEP workshop, Jellyfish blooms in the Mediterranean Sea, Proceedings of II Workshop on Jellyfish in the Mediterranean Sea. 102–111 (1991).
- 39.Malej A. Behaviour and trophic ecology of the jellyfish Pelagia noctiluca (Forsskal, 1775) J. Exp. Mar. Ecol. 1989;126:259–270. doi: 10.1016/0022-0981(89)90191-3. [DOI] [Google Scholar]
- 40.Chavtur VG. Vertical distribution of pelagic ostracods (Ostracoda, Halocyprinidae) in the Australian - New Zealand sector of the Southern Ocean. Russ. J. Mar. Biol. 2003;29:90–99. doi: 10.1023/A:1023996306419. [DOI] [Google Scholar]
- 41.Pillar SC. Diel variation in the vertical distribution of some common zooplankton species off the west coast of South Africa. South African J. Mar. Sci. 1984;2:71–80. doi: 10.2989/02577618409504360. [DOI] [Google Scholar]
- 42.Fuiman LA, Magurran AE. Development of predator defences in fishes. Rev. Fish Biol. Fish. 1994;4:145–183. doi: 10.1007/BF00044127. [DOI] [Google Scholar]
- 43.Hubbs C, Blaxter JHS. Ninth Larval Fish Conference. Trans. Am. Fish. Soc. 1986;115:98–114. doi: 10.1577/1548-8659(1986)115<98:NLFCDO>2.0.CO;2. [DOI] [Google Scholar]
- 44.Gordoa A, Acuña JL, Farrés R, Bacher K. Burst Feeding of Pelagia noctiluca ephyrae on Atlantic Bluefin Tuna (Thunnus thynnus) Eggs. PLoS One. 2013;8:e74721. doi: 10.1371/journal.pone.0074721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Purcell JE, et al. Digestion and predation rates of zooplankton by the pleustonic hydrozoan Velella velella and widespread blooms in 2013 and 2014. J. Plankton Res. 2014;37:1056–1067. doi: 10.1093/plankt/fbv031. [DOI] [Google Scholar]
- 46.Tilves U, et al. Trophic interactions of the jellyfish Pelagia noctiluca in the NW Mediterranean: Evidence from stable isotope signatures and fatty acid composition. Mar. Ecol. Prog. Ser. 2018;591:101–116. doi: 10.3354/meps12332. [DOI] [Google Scholar]
- 47.Purcell JE, et al. Digestion times and predation potentials of Pelagia noctiluca eating fish larvae and copepods in the NW Mediterranean Sea. Mar. Ecol. Prog. Ser. 2014;510:201–213. doi: 10.3354/meps10790. [DOI] [Google Scholar]
- 48.Michener, R. H. & Schell, D. M. Stable isotopes ratios as tracers in marine aquatic foodwebs. In Stable Isotopes in Ecology and Environmental Sciences. (eds Lajtha, K. & Michener, R. H.) 138–157 (1994).
- 49.Post DM. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology. 2002;83:703–718. doi: 10.1890/0012-9658(2002)083[0703:USITET]2.0.CO;2. [DOI] [Google Scholar]
- 50.Purcell, J. E., Sturdevant, M. V. & Galt, C. P. A review of appendicularians as prey of fish and invertebrate predators. In Response of marine ecosystems to global change: Ecological impact of appendicularians. (eds Gorsky, G., Youngbluth, M. J. & Deibel, D.). Contemporary Publishing International, Paris, pp. 359–435 (2005).
- 51.Stoecker DK, Michaels AE, Davis LH. Grazing by the jellyfish, Aurelia aurita, on microzooplankton. J. Plankton Res. 1987;9:901–915. doi: 10.1093/plankt/9.5.901. [DOI] [Google Scholar]
- 52.Smetacek V. The annual cycle of protozooplankton in the Kiel Bight. Mar. Biol. 1981;63:1–11. doi: 10.1007/BF00394657. [DOI] [Google Scholar]
- 53.Azzaro F, Decembrini F, Raffa F, Crisafi E. Seasonal variability of phytoplankton fluorescence in relation to the Straits of Messina (Sicily) tidal upwelling. Ocean Sci. Discuss. 2007;4:415–440. doi: 10.5194/osd-4-415-2007. [DOI] [Google Scholar]
- 54.D’Alcalà MR, et al. Seasonal patterns in plankton communities in a pluriannual time series at a coastal Mediterranean site (Gulf of Naples): an attempt to discern recurrences and trends. Sci. Mar. 2004;68:65–83. doi: 10.3989/scimar.2004.68s165. [DOI] [Google Scholar]
- 55.Rossi S, Gili JM. Composition and temporal variation of the near-bottom seston in a Mediterranean coastal area. Estuar. Coast. Shelf Sci. 2005;65:385–395. doi: 10.1016/j.ecss.2005.05.024. [DOI] [Google Scholar]
- 56.Cifuentes L, Sharp J, Fogel M. Stable carbon and nitrogen isotope biogeochemistry in the Delaware estuary. Limnol. Ocean. 1988;33:1102–1115. doi: 10.4319/lo.1988.33.5.1102. [DOI] [Google Scholar]
- 57.Fukuda Y, Naganuma T. Potential dietary effects on the fatty acid composition of the common jellyfish Aurelia aurita. Mar. Biol. 2001;138:1029–1035. doi: 10.1007/s002270000512. [DOI] [Google Scholar]
- 58.Colin S, Costello J. Omnivory by the small cosmopolitan hydromedusa. Aglaura hemistoma. Limnol. Oceanogr. 2005;50:1264–1268. doi: 10.4319/lo.2005.50.4.1264. [DOI] [Google Scholar]
- 59.Gili JM, Duró A, García-Valero J, Gasol JM, Rossi S. Herbivory in small carnivores: benthic hydroids as an example. J. Mar. Biol. Assoc. United Kingdom. 2008;88:1541–1546. doi: 10.1017/S0025315408003214. [DOI] [Google Scholar]
- 60.Regula C, Colin S, Costello J, Kordula H. Prey selection mechanism of ambush-foraging hydromedusae. Mar. Ecol. Prog. Ser. 2009;374:135–144. doi: 10.3354/meps07756. [DOI] [Google Scholar]
- 61.Elias-Piera F, Rossi S, Gili J, Orejas C. Trophic ecology of seven Antarctic gorgonian species. Mar. Ecol. Prog. Ser. 2013;477:93–106. doi: 10.3354/meps10152. [DOI] [Google Scholar]
- 62.Larson RJ, Harbison GR. Source and fate of lipids in polar gelatinous zooplankton. Arctic. 1989;42:339–346. doi: 10.14430/arctic1675. [DOI] [Google Scholar]
- 63.Monroig Ó, Tocher DR, Navarro JC. Biosynthesis of polyunsaturated fatty acids in marine invertebrates: Recent advances in molecular mechanisms. Mar. Drugs. 2013;11:3998–4018. doi: 10.3390/md11103998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Nelson MM, Phleger CF, Mooney BD, Nichols PD. Lipids of gelatinous Antarctic zooplankton: Cnidaria and Ctenophora. Lipids. 2000;35:551–559. doi: 10.1007/s11745-000-555-5. [DOI] [PubMed] [Google Scholar]
- 65.Lucas CH. Biochemical composition of Aurelia aurita in relation to age and sexual maturity. J. Exp. Mar. Biol. Ecol. 1994;183:179–192. doi: 10.1016/0022-0981(94)90086-8. [DOI] [Google Scholar]
- 66.Doyle TK, Houghton JDR, McDevitt R, Davenport J, Hays JC. The energy density of jellyfish: Estimates from bomb-calorimetry and proximate-composition. J. Exp. Mar. Biol. Ecol. 2007;343(2):239–252. doi: 10.1016/j.jembe.2006.12.010. [DOI] [Google Scholar]
- 67.Lucas CH, Pitt KA, Purcell JE, Lebrato M, Condon RH. What’s in a jellyfish? Proximate and elemental composition and biometric relationships for use in biogeochemical studies. Ecology. 2011;92(8):1704. doi: 10.1890/11-0302.1. [DOI] [Google Scholar]
- 68.Purcell JE. Digestion rates and assimilation efficiencies of siphonophores fed zooplankton prey. Mar. Biol. 1983;73:257–261. doi: 10.1007/BF00392251. [DOI] [Google Scholar]
- 69.Watanabe T. Lipid nutrition in fish. Comp. Biochem. Physiol. 1982;73:3–15. [Google Scholar]
- 70.Scott CL, Kwasniewski S, Falk-Petersen S, Sargent JR. Species differences, origins and functions of fatty alcohols and fatty acids in the wax esters and C. finmarchicus from Arctic waters. Mar. Ecol. Prog. Ser. 2002;235:127–134. doi: 10.3354/meps235127. [DOI] [Google Scholar]
- 71.Cripps GC, Atkinson A. Fatty acid composition as an indicator of carnivory in Antarctic krill. Euphausia superba. Can. J. Fish. Aquat. Sci. 2000;57:31–37. doi: 10.1139/f00-167. [DOI] [Google Scholar]
- 72.Fuchs B, et al. Regulation of polyp-to-jellyfish transition in Aurelia aurita. Curr. Biol. 2014;24:263–273. doi: 10.1016/j.cub.2013.12.003. [DOI] [PubMed] [Google Scholar]
- 73.Piraino S, Fanelli G, Boero F. Variability of species’ roles in marine communities: Change of paradigms for conservation priorities. Mar. Biol. 2002;140:1067–1074. doi: 10.1007/s00227-001-0769-2. [DOI] [Google Scholar]
- 74.Milisenda, G. et al. Jellyfish as prey: Frequency of predation and selective foraging of Boops boops (Vertebrata, Actinopterygii) on the mauve stinger Pelagia noctiluca (Cnidaria, Scyphozoa). PLoS One9 (2014). [DOI] [PMC free article] [PubMed]
- 75.Pearre S., Jr. Estimating prey preference bypPredators: Uses of various indices, and a proposal of another based on χ2. Can. J. Fish. Aquat. Sci. 1982;39:914–923. doi: 10.1139/f82-122. [DOI] [Google Scholar]
- 76.Jacob U, Mintenbeck K, Brey T, Knust R, Beyer K. Stable isotope food web studies: a case for standardized sample treatment. Mar. Ecol. Prog. Ser. 2005;287:251–253. doi: 10.3354/meps287251. [DOI] [Google Scholar]
- 77.Vander Zanden, M. J. & Rasmussen, J. B. Variation in δ15N and δ13C trophic fractionation: implications for aquatic food web studies. Limnol. Oceanogr. 46, (2001).
- 78.D’Ambra I, Carmichael RH, Graham WM. Determination of δ13C and δ15N and trophic fractionation in jellyfish: Implications for food web ecology. Mar. Biol. 2014;161:473–480. doi: 10.1007/s00227-013-2345-y. [DOI] [Google Scholar]
- 79.Ruiz J, Antequera T, Andres AI, Petron MJ. & Muriel, E. Improvement of a solid phase extraction method for analysis of lipid fractions in muscle foods. Anal. Chim. Acta. 2004;520:201–205. doi: 10.1016/j.aca.2004.04.059. [DOI] [Google Scholar]
- 80.Soler-Membrives A, Rossi S, Munilla T. Feeding ecology of NW Mediterranean sea spiders (Pycnogonida): temporal variation in fatty acid composition. Estuar. Coast. Shelf Sci. 2011;92:588–597. doi: 10.1016/j.ecss.2011.02.018. [DOI] [Google Scholar]
- 81.Anderson MJ. Permutation tests for univariate or multivariate analysis of variance and regression. Can. J. Fish. Aquat. Sci. 2001;58:626–639. doi: 10.1139/f01-004. [DOI] [Google Scholar]
- 82.Parnell, A. & Jackson, A. L. SIAR: Stable Isotope Analysis in R (2013). https:// cran. r-project. org/web/packages/ siar/.
- 83.Legendre, P. & Legendre, L. NumericalEcology, 3rd edn, vol. 24 of Developments in Environmental Modelling, pp. 1006. Elsevier, Amsterdam (2012).
- 84.Anderson, M., Gorley, R. & Clarke, K. P. for PRIMER: guide to software and statistical methods. Prim. Plymouth, UK (2008).
- 85.Clarke, K. R. & Warwich, R. Change in marine communities: an approach to statistical analysis statistical analysis and interpretation, 2nd edition. (Plymouth MarineLaboratory) (2001).
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.