

Cancer-related anemia (CRA) is part of the broader group of anemias caused by inflammation, in which macrophages and inflammatory cytokines, in particular interleukin (IL)-6 and hepcidin, play a major role.1,2 Typically, CRA is characterized by low levels of serum iron, but high levels of ferritin.2,3 The macrophages are actually the cells that use denial of iron from the microenvironment as an important strategy to accomplish their key role in innate immunity and host defenses.4 In response to different stimuli activated macrophages may polarize into either M1 or M2 phenotype,5 each defined by a specific iron metabolism profile.6 The exact role exerted by polarized tumor-associated macrophages (TAM) and their related cytokines in influencing the incidence and severity of CRA has not yet been demonstrated and it is an aim of the present study. In this regard, we believed that the microenvironment of the peritoneal cavity of patients with ovarian cancer could be the ideal system to evaluate these mechanisms. In the abdominal cancer milieu, macrophages live and interact with cancer cells and they are the most represented non-neoplastic cellular component.
In the present study ascitic fluid from 85 patients suffering from primary epithelial ovarian/fallopian tube cancer (Table 1) was used to assess TAM polarization, associated changes of inflammatory and iron metabolism parameters, and its relationship with hemoglobin. Moreover, we evaluated the ability of in vitro TAM to release IL-6 and hepcidin. Hypothesizing a role for ascitic IL-6 in mediating macrophage polarization, we tested the ability of ascites to polarize peripheral blood macrophages, which are typically resting, and induce hepcidin synthesis, and then the effect of an anti-IL-6 antibody on such activities. Patients were evaluated at diagnosis before any anti-neoplastic treatment and ascitic fluid was collected at the time of laparoscopic surgery that, in our surgical practice, precedes any additional surgical time. The mean hemoglobin level was 10.7±1.0 g/dL (Table 1) and was inversely correlated with high serum levels of IL-6, C-reactive protein, hepcidin and ferritin, thus identifying the typical picture of CRA (Table 1, Online Supplementary Table S1).
Table 1.
Patients’ clinical and laboratory characteristics.
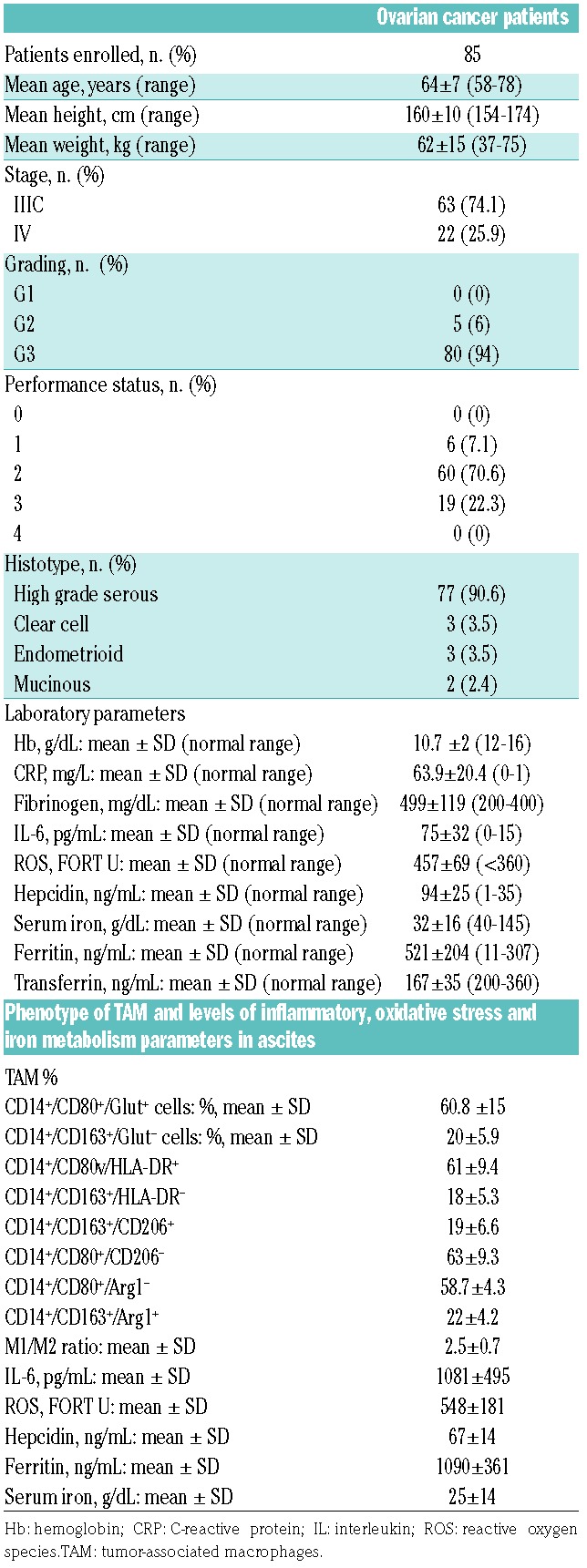
To evaluate the potential link between CRA and TAM polarization, we used flow cytometry to analyze the phenotype of TAM isolated by double density gradient (Ficoll-Hypaque 75/100%) from the ascitic fluid: CD14+ cells were the most represented population in the window corresponding to monocytes (86.8%), with <20% contamination by non-lympho-monocytes and more than 90% of viable cells. For the analysis of surface markers linked to macrophage polarization we chose CD80 and HLA-DR, which are linked to M1 polarization, and CD163 and CD206, which are established markers for M2 polarization;7 additionally, we studied Glut-1 expression, typically associated with increased glycolytic activity of M1, and Arg1 expression, mostly linked to the metabolic profile of M2.8
TAM presented a mixed phenotype with a prevalence of CD14+/CD80+/Glut+ (60.8±15%) and CD14+/CD80+/HLA-DR+ (61±9.4%) cells, resembling a M1 phenotype, and low percentages of CD14+/CD163+/Glut− (20±5.9%), CD14+/CD163+/CD206+ (19±6.6%) and CD14+/CD163+/Arg1+ (22±4.2%) cells, indicative of a M2 phenotype, with a mean M1/M2 ratio of 2.5±0.7 (Table 1). Consistently, the glucose uptake, evaluated by a fluorescently labeled deoxyglucose analog and represented by the mean fluorescence intensity, was greater in CD14+/CD80+ TAM than in CD14+/CD163+ TAM (Online Supplementary Figure S1). Such M1 polarization of TAM was associated with CRA and significantly inversely correlated with hemoglobin levels (Table 2). These results indicate that M1 TAM in the tumor microenvironment may play an important role not only in the etiopathogenesis but also in the severity of anemia. The Glut-1 expression and glucose uptake of TAM, as well as their high percentage in the S phase of the cell cycle (Online Supplementary Figure S2) at the time of collection demonstrated that these cells were functionally activated. Indeed, the functional activity of macrophages is cell cycle-dependent and strictly related to their ability to enter the cell cycle and start replication.9 The TAM assessed by us could, therefore, be chronically activated by tissue damage induced by the neoplastic process; in turn, activated macrophages result in the synthesis of cytokines and other factors also on the basis of their polarization.7
Table 2.
Correlation of TAM polarization phenotype and functional status with hemoglobin, synthesis of IL-6 and hepcidin, and ascitic levels of IL-6, ROS and iron metabolism-related factors.

In the light of the above results we evaluated the ability of cultured in vitro TAM to release two of the main mediators involved in the pathogenesis of CRA, i.e., IL-6 and hepcidin. Firstly, we demonstrated that, after 24 h, unstimulated TAM released significantly higher levels of IL-6 and hepcidin in culture medium in comparison to cultured unstimulated CD14+ macrophages isolated from peripheral blood of the same patient. We, therefore, verified whether these TAM could be further activated in culture by assessing the percentage of cells progressing into the S phase after the addition of the mitogen phytohemagglutinin (PHA). The percentage of TAM in the S phase increased significantly, demonstrating that TAM could be activated by PHA (Online Supplementary Figure S2). The IL-6 and hepcidin released by PHA-stimulated TAM increased significantly compared to that released by non-stimulated cells (P=0.0005 for IL-6, and P=0.014 for hepcidin) (Online Supplementary Figure S3A,B). The synthesis of IL-6 and hepcidin by cultured TAM was compared to that of CD14+ macrophages isolated from peripheral blood. The mean increases in IL-6 and hepcidin in culture medium of PHA-stimulated peripheral blood CD14+ macrophages were significantly lower than those found in TAM cultures (P<0.001 for both). These results support the hypothesis that the ability to produce IL-6 and hepcidin is peculiar to activated TAM from the cancer microenvironment. Notably, the extent of the increase in IL-6 and hepcidin in the culture medium by PHA-stimulated TAM was positively correlated with M1 polarization and the M1/M2 ratio; additionally, it was negatively correlated with hemoglobin concentration (Table 2).
This evidence was indirectly confirmed by the finding that the levels of ascitic IL-6 and hepcidin were positively correlated with TAM M1 polarization and negatively with hemoglobin (Table 2). According to the evidence that M1 polarization is associated with a peculiar shift in iron metabolism-related pathways,6 we found that ascitic ferritin levels were high and positively correlated with M1 TAM and the M1/M2 ratio, while they were inversely correlated with hemoglobin. Vice versa, ascitic free iron levels were inversely correlated with M1 TAM and the M1/M2 ratio and positively correlated with hemoglobin (Tables 1 and 2). According to the hypothesis that M1 polarization and related iron retention may enhance the production of oxidative stress metabolites,10 we found that the levels of reactive oxygen species in ascites were high and positively correlated with the amount of M1 TAM and the M1/M2 ratio, while they were inversely correlated with hemoglobin (Tables 1 and 2). At multivariate regression analysis, values of ascitic IL-6 (β-coefficient -0.359, P=0.007), hepcidin (β-coefficient -0.737, P<0.001), and the M1/M2 ratio (β-coefficient -0.413, P=0.005) were independent predictors of hemoglobin concentration.
Thereafter, we assessed whether ascitic fluid was able to modulate the polarization of peripheral blood macrophages obtained from 20 patients with primary ovarian cancer and the consequent in vitro hepcidin production. The phenotypic assessment of peripheral blood macrophages immediately after their isolation showed that these were resting, non-polarized cells; when cultured with the addition of ascites, they polarized into M1 cells with a significant increase of the M1/M2 ratio (P=0.01) in comparison to cells of the control well (peripheral blood CD14+ macrophages cultured without ascites) (Table 3, Online Supplementary Figure S3C). The increase of M1 macrophages in each experiment was proportional to the concentration of IL-6 in ascites supernatants of the corresponding sample. Thus, hypothesizing that IL-6 could be the main soluble factor responsible for such activation, we added the anti-IL-6 monoclonal antibody tocilizumab11 to peripheral CD14+ macrophages cultured in the presence of ascites: control experiments with an equal dose of the isotype control (IgG1) for the anti-IL-6 antibody were also performed. Since it had already been demonstrated in preclinical animal cancer models that anti-IL-6 treatment was associated with an improvement of anemia,12 we focused our study only on human samples obtained from our patients. After 24 h, the hepcidin in culture medium fell to undetectable levels (Table 3). Furthermore, after 72 h we observed a significant drop in the M1 percentage to a degree proportional to the concentration of monoclonal antibody used, reaching a mean value of 5.7%, which is superimposable to the value observed in the culture of peripheral CD14+ macrophages without ascites. Vice versa, the percentage of M2 was unchanged (Table 3, Online Supplementary Figure S3C). The control experiments with the addition of only the isotype control antibody did not produce any significant changes. This appears to confirm a central role of IL-6 in mediating macrophage M1 polarization and its direct link with hepcidin synthesis in the tumor microenvironment, thus contributing to inducing the functional iron deficiency typical of CRA. However, ascitic fluid contains many other factors besides IL-6, although this is one of the most significant, so the inhibition of the above activities by neutralizing IL-6 does not rule out the potential involvement of other factors in combination.
Table 3.
Phenotype and hepcidin synthesis by CD14+ macrophages isolated from peripheral blood of patients with primary ovarian cancer at baseline and after culture without or with the addition of ascites ± anti-IL-6 monoclonal antibody at increasing doses (1-1000 mg/mL).

In conclusion, our study reveals a strong association between the severity of CRA and the amount of M1 TAM as well as hepcidin and IL-6 released by these cells. Notably, we found that IL-6 is responsible for macrophage polarization into the M1 phenotype. IL-6 blockade with a specific monoclonal antibody inhibited this polarization and the consequent synthesis of hepcidin. Indeed, starting from the hypothesis that the regulation of the functional iron deficiency induced by M1 macrophages could be a key element of this form of anemia, in a randomized clinical trial in patients with advanced cancer we showed the efficacy of lactoferrin, a modulator of iron trafficking and storage, in the treatment of CRA.13 In this regard, a very recent study14 showed that lactoferrin is able to downregulate M1 macrophage polarization and the related changes of iron metabolism, thus providing a strong biological basis for the role that we have hypothesized here could be played by M1 polarization and the consequent synthesis of IL-6 in the pathogenesis of CRA. These findings provide insights into the use of anti-IL-6 monoclonal antibody as a treatment for CRA based on the regulation of macrophage polarization through the inhibition of IL-6 activity.15
Supplementary Material
Acknowledgments
The authors would like to thank Ivan Collu, Martina Trudu, Andreea Voicu, and Federica Concas for their technical assistance.
Footnotes
Funding: this work was supported by the “Associazione Sarda per la ricerca in Oncologia Ginecologica-ONLUS” with funding from the “Banco di Sardegna” Foundation (grant n. 5335, 2014).
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Weiss G, Goodnough LT. Anemia of chronic disease. N Engl J Med. 2005;352(10):1011–1023. [DOI] [PubMed] [Google Scholar]
- 2.Macciò A, Madeddu C, Gramignano G, et al. The role of inflammation, iron, and nutritional status in cancer-related anemia: results of a large, prospective, observational study. Haematologica. 2015; 100(1):124–132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Davis SL, Littlewood TJ. The investigation and treatment of secondary anaemia. Blood Rev. 2012;26(2):65–71. [DOI] [PubMed] [Google Scholar]
- 4.Soares MP. Macrophages and iron metabolism. Immunity. 2016; 44(3):492–504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Biswas SK, Mantovani A. Macrophage plasticity and interaction with lymphocyte subsets: cancer as a paradigm. Nat Immunol. 2010;11(10):889–896. [DOI] [PubMed] [Google Scholar]
- 6.Cairo G, Recalcati S, Mantovani A, Locati M. Iron trafficking and metabolism in macrophages: contribution to the polarized phenotype. Trends Immunol. 2011;32(6):241–247. [DOI] [PubMed] [Google Scholar]
- 7.Murray PJ, Allen JE, Biswas SK, et al. Macrophage activation and polarization: nomenclature and experimental guidelines. Immunity. 2014;41(1):14–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Freemerman AJ, Johnson AR, Sacks GN, et al. Metabolic reprogramming of macrophages: glucose transporter 1 (GLUT1)-mediated glucose metabolism drives a proinflammatory phenotype. J Biol Chem. 2014;289(11):7884–7896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Luo Y, Tucker SC, Casadevall A. Fc- and complement-receptor activation stimulates cell cycle progression of macrophage cells from G1 to S. J Immunol. 2005;174(11):7226–7233. [DOI] [PubMed] [Google Scholar]
- 10.Biswas SK, Mantovani A. Orchestration of metabolism by macrophages. Cell Metabolism. 2012;15(4):432–437. [DOI] [PubMed] [Google Scholar]
- 11.Obeng JA, Amoruso A, Camaschella GL, Sola D, Brunelleschi S, Fresu LG. Modulation of human monocyte/macrophage activity by tocilizumab, abatacept and etanercept: an in vitro study. Eur J Pharmacol. 2016;80:33–37. [DOI] [PubMed] [Google Scholar]
- 12.Noguchi-Sasaki M, Sasaki Y, Shimonaka Y, Mori K, Fujimoto-Ouchi K. Treatment with anti-IL-6 receptor antibody prevented increase in serum hepcidin levels and improved anemia in mice inoculated with IL-6-producing lung carcinoma cells. BMC Cancer. 2016;16:270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Macciò A, Madeddu C, Gramignano G, Mulas C, Sanna E, Mantovani G. Efficacy and safety of oral lactoferrin supplementation in combination with rHuEPO-beta for the treatment of anemia in advanced cancer patients undergoing chemotherapy: open-label, randomized controlled study. Oncologist. 2010;15(8):894–902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cutone A, Rosa L, Lepanto MS, et al. Lactoferrin efficiently counteracts the inflammation-induced changes of the iron homeostasis system in macrophages. Front Immunol. 2017;8:705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rochette L, Gudjoncik A, Guenancia C, Zeller M, Cottin Y, Vergely C. The iron-regulatory hormone hepcidin: a possible therapeutic target¿ Pharmacol Ther. 2015;146:35–52. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.