

Abstract
Tartary buckwheat (Fagopyrum tataricum) has been established globally as a nutritionally important food item, particularly owing to high levels of bioactive compounds such as rutin. This study investigated the effect of tartary buckwheat extracts (TBEs) on adipogenesis and inflammatory response in 3T3-L1 cells. TBEs inhibited lipid accumulation, triglyceride content, and glycerol-3-phosphate dehydrogenase (GPDH) activity during adipocyte differentiation of 3T3 L1 cells. The mRNA levels of genes involved in fatty acid synthesis, such as peroxisome proliferator-activated receptor-γ (PPAR-γ), CCAAT/enhancer binding protein-α (CEBP-α), adipocyte protein 2 (aP2), acetyl-CoA carboxylase (ACC), fatty acid synthase (FAS), and stearoylcoenzyme A desaturase-1 (SCD-1), were suppressed by TBEs. They also reduced the mRNA levels of inflammatory mediators such as tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), monocyte chemoattractant protein 1 (MCP-1), and inducible nitric oxide synthase (iNOS). In addition, TBEs were decreased nitric oxide (NO) production. These results suggest that TBEs may inhibit adipogenesis and inflammatory response; therefore, they seem to be beneficial as a food ingredient to prevent obesity-associated inflammation.
Keywords: tartary buckwheat, adipogenesis, inflammatory response, 3T3-L1 adipocytes
1. Introduction
Adipocytes play a central role in maintaining whole-body energy balance and lipid homeostasis by storing triglycerides (TGs) or releasing free fatty acids [1,2]. However, excessive fat deposition in adipose tissues can lead to obesity [3]. Obesity is notably associated with various metabolic disorders including hypertension, type 2 diabetes, and cardiovascular diseases [4]. Adipose tissue not only stores fat but is also an important endocrine organ that secretes several physiologically active peptides, including inflammatory cytokines [5]. Obesity causes increased expression of inflammatory cytokines in chronic low-grade inflammation, contributing to pathological dysfunction [6]. Therefore, it is important to develop natural products that might inhibit adipogenesis and inflammation in order to prevent obesity.
Buckwheat (Fagopyrum esculentum) is recognized as a functional food and an important source of high quality protein, abundant flavonoids, and well-balanced essential amino acids and minerals [7,8]. Two types of buckwheat, namely common buckwheat (F. esculentum) and tartary buckwheat (F. tataricum), are consumed globally. Common buckwheat is mainly grown in Europe, USA, Canada, Brazil, South Africa, and Asia. Tartary buckwheat is mainly grown in southwest China, northern India, Bhutan, and Nepal [9,10]. In particular, tartary buckwheat is known to have 9–300 times higher flavonoid content, mainly rutin and quercetin, than common buckwheat [11,12]. In recent years, tartary buckwheat has been attracting research interest for the prevention of various diseases [13,14,15]. Tartary buckwheat is reported to have various beneficial effects, such as antioxidant [16], antitumor [17], and hypoglycemic [18] properties. A recent study reported that rutin-rich tartary buckwheat showed potential effects on decreasing body weight, body fat percentage, and oxidative stress in adult subjects [13]. Yoon and others [19] observed that an 80% ethanolic extract of tartary buckwheat suppressed adipogenesis and reactive oxygen species (ROS) production compared to common buckwheat extract. However, the mechanism of tartary buckwheat extract involved in adipogenesis and inflammatory response remains unclear.
Here, we elucidated the effect of tartary buckwheat extracts (TBEs) on adipogenesis and inflammatory responses and identified the molecular mechanisms underlying its regulatory action. In addition, we investigated the effect of 50% (TBE-50) or 70% (TBE-70) ethanolic extracts of tartary buckwheat on adipogenesis and inflammatory response during adipocyte differentiation.
2. Results and Discussion
2.1. Rutin Contents of TBE
Rutin is a flavonol glycoside composed of quercetin and disaccharide rutinose (Figure 1) and is present in many plants, including buckwheat [20]. The tartary buckwheat contains protocatechuic acid, vanilic acid, syringic acid, ferulic acid, sinapic acid, and quercetin as their phenolic compounds [21]. However, many researchers have reported that rutin is the main active compound found in tartary buckwheat extracts, since more than 80% of total flavonoids in tartary buckwheat extracts are revealed to be rutin [21,22], and rutin accounted for about 85–90% of the total anti-oxidative activity in tartary buckwheat [23]. In this study, we analyzed the content of rutin in 50% (TBE-50) or 70% (TBE-70) ethanolic extracts of tartary buckwheat by the high-performance liquid chromatography (HPLC) method. The HPLC chromatogram of the rutin is presented in Figure 2a–c. The rutin concentrations of TBE-50 and TBE-70 were 84.3 ± 0.7 mg/g and 106.0 ± 1.3 mg/g, respectively (Figure 3). Beneficial effects of rutin on obesity [24,25] and inflammation [26,27] have been reported. Thus, we assumed that TBEs might have effects on adipogenesis and inflammation during adipocyte differentiation, and rutin may contribute in part to beneficial effects of TBEs.
Figure 1.

Chemical structure of rutin.
Figure 2.

HPLC chromatogram of rutin in TBEs. Rutin standard (a); TBE-50 (b); TBE-70 (c).
Figure 3.
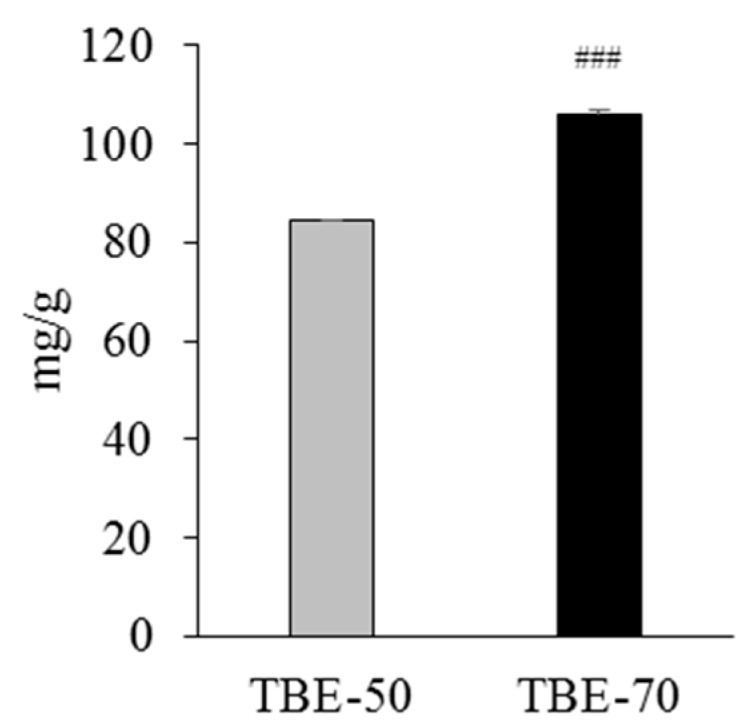
The rutin content of TBE-50 and TBE-70. Rutin content was determined by HPLC. Values are expressed as mean ± SE (n = 3) of three independent experiments. ### p < 0.001 vs. TBE-50.
2.2. Effect of TBEs on 3T3-L1 Cell Viability
TBEs were tested for the potential cytotoxic effects that might exert on 3T3-L1 cells. Cells were treated with TBE-50 or TBE-70 at various concentrations (0 (control), 0.1, 1, 10, 50, 100, or 500 μg/mL), and incubated for 1, 2, 5, or 7 days. Cytotoxicity was unaffected by 0, 0.1, 1, 10, 50, and 100 μg/mL of TBE-50 and TBE-70 after 7 days of incubation (Figure 4). However, 7 days of incubation with high doses (500 μg/mL) of TBE-50 and TBE-70 significantly decreased cell viability by 13.2% and 10.3%, respectively, compared to the control. The cells did not show any toxicity upon treatment with TBE-50 and TBE-70 at concentrations of 0.1–100 µg/mL. Therefore, this in vitro study was performed at a nontoxic range of concentrations below 100 μg/mL with both extracts.
Figure 4.

Effects of TBEs on cell viability in 3T3-L1 cells. Cells were treated with 0 (control), 0.1, 1, 10, 50, 100, or 500 µg/mL of TBE-50 (a) or TBE-70 (b), and incubated for 1, 2, 5, or 7 days. Cell viability was determined using the WST-8 assay. Values are expressed as mean ± SE (n = 3) of three independent experiments. * p < 0.05 vs. control.
2.3. Effects of TBEs on Lipid Accumulation and TG Content during Adipocyte Differentiation
Adipogenesis is the process by which preadipocytes become mature adipocytes, when exposed to appropriate environmental condition and gene expression. Mature adipocytes enlarge in size while accumulating lipids that finally fill the cells. In this study, the effects of TBEs on intracellular lipid accumulation and triglyceride (TG) content during 3T3-L1 adipocyte differentiation were measured. Cells were treated with 100 μg/mL of TBE-50 or TBE-70 for 2, 5, or 7 days (d2 to d9) as shown in Figure 5a. The intracellular lipid content was measured by Oil Red O staining. On Day 7 (d9), change of adipocyte differentiation was presented with Oil Red O staining (Figure 5b). After 7 days of incubation, TBE-70 inhibited the intracellular lipid content by 15.2% compared to the control cells, whereas TBE-50 did not result in significant change (Figure 5c). To determine the intracellular TG content, 3T3-L1 adipocytes were treated with 0 (control), 1, 50, and 100 μg/mL of TBE-50 and TBE-70 for 7 days. The 100 μg/mL of TBE-70 significantly decreased the TG content by 22.3% compared to the control cells. Further, the intracellular TG content of the TBE-70 was found to be lower than that of TBE-50 by 15.3% (Figure 5d). In a previous study, supplementation of buckwheat leaf and flower mixture resulted in reduced weight gain and plasma lipid concentrations in rats fed a high-fat diet [28]. In addition, a study in humans revealed that the rutin-rich tartary buckwheat showed potential effects on decreasing the body fat percentage and body weight in adults [13]. Meanwhile, the anti-adipogenic effects of phenolic acids present in tartary buckwheat such as rutin [29], ferulic acid [30], and quercetin [31,32,33] have been reported. Our results showed that TBEs reduced the intracellular lipid and TG concentrations during adipocyte differentiation. These results imply that TBEs would be beneficial at suppressing fat accumulation, and these effects seem to be a result of complex action of phenolic acids and rutin in TBEs.
Figure 5.

Effects of TBEs on intracellular lipid accumulation and TG content during adipocyte differentiation. Scheme of the 3T3-L1 preadipocyte differentiation experiment (a). 3T3-L1 cells were treated with 100 µg/mL of TBE-50 or TBE-70, and incubated for 2, 5, or 7 days (d2 to d9). On Day 7 (d9), change of adipocyte differentiation was presented with Oil Red O staining (b). Intracellular lipid content (c) was stained with oil-red O dye, and dissolved the stained oil droplets with isopropanol and quantified by spectrophotometric analysis. Representative cell images were captured at 200× magnification. 3T3-L1 cells were treated with 0 (MDI treated control), 1, 50, and 100 µg/mL of TBE-50 and TBE-70, and incubated for 7 days. Intracellular TG content (d) was determined using enzymatic colorimetric methods. MDI, medium containing 3-isobutyl-1-methylxanthine, dexamethasone and insulin. Values are expressed as mean ± SE (n = 3) of three independent experiments. * p < 0.05, ** p < 0.01 and *** p < 0.01 vs. MDI-treated control. # p < 0.05 vs. TBE-50.
2.4. Effect of TBEs on GPDH Activity in Adipocytes
One of the possible methods of inhibiting lipid accumulation in adipocytes is blocking lipid synthetic pathways in their cells. The enzyme glycerol-3-phosphate dehydrogenase (GPDH) plays a major role in the TG synthesis pathway and is linked to characteristic changes that occur during adipose conversion [34]. To elucidate the mechanism by which TBEs inhibits lipid accumulation, GPDH activity was measured in 3T3-L1 adipocytes. The 3T3-L1 adipocytes were incubated with 0 (control), 1, 50, and 100 μg/mL of TBE-50 and TBE-70, and incubated for 7 days. GPDH activity was significantly decreased by 10.7% and 25.1%, respectively, in the presence of 50 and 100 μg/mL of TBE-70, compared to the untreated control (Figure 6). However, no significant differences in GPDH activity were observed in the presence of TBE-50. Yoon and others [19] have reported that an 80% ethanolic extract of tartary buckwheat showed relatively higher inhibition of GPDH activity and lipid accumulation than the common buckwheat extract in 3T3-L1 adipocytes. In addition, Hsu and Yen have reported that rutin inhibits intracellular TG accumulation and GPDH activity in adipocytes during differentiation [29]. Similarly, in our result, GPDH activity was dose-dependently reduced by the TBE-70 in 3T3-L1 adipocytes. Thus, it can be speculated that TBEs may have a suppressive effect on lipid synthesis partially via reduction of GPDH activity during adipocyte differentiation.
Figure 6.

Effects of TBEs on GPDH activity in adipocytes. GPDH activity was determined using a GPDH assay kit. 3T3-L1 adipocytes were treated with 0 (MDI treated control), 1, 50, and 100 µg/mL of TBE-50 and TBE-70, and incubated for 7 days. MDI, medium containing 3-isobutyl-1-methylxanthine, dexamethasone, and insulin. Values are expressed as mean ± SE (n = 3) of three independent experiments. * p < 0.05, ** p < 0.01 and *** p < 0.001 vs. MDI-treated control.
2.5. Effect of TBEs on mRNA Expression of Genes Involved in Fatty Acid Synthesis in Adipocytes
During adipogenesis, enhanced expression of c-fos, c-jun, junB, c-myc, and CCAAT/enhancer binding proteins (C/EBP)-β and -δ is observed [35]. Activated C/EBP-β and -δ mediate the expression of peroxisome proliferator-activated receptor γ (PPARγ) and C/EBP-α [35]. To understand the mechanism underlying the anti-adipogenic effect of TBEs in adipocyte, the mRNA levels of genes involved in fatty acid synthesis were measured. At the molecular level, adipogenesis is driven by a complex signaling cascade that involves key transcription factors, such as proliferator-activated receptor-γ (PPAR-γ) and CCAAT/enhancer binding protein-α (CEBP-α) [36]. When PPAR-γ expression is stimulated, lipid biosynthesis pathways are activated through the expression of target genes, such as CEBP-α and aP2 [37,38]. The adipogenic marker gene adipocyte protein 2 (aP2) is highly expressed as a result of adipocyte differentiation [39]. Acetyl-CoA carboxylase (ACC) is the rate-limiting enzyme of fatty acid synthesis that catalyzes the carboxylation of acetyl-CoA to produce malonyl-CoA [40]. Fatty acids are synthesized from malonyl-CoA through processes catalyzed by fatty acid synthase (FAS) and stearoylcoenzyme A desaturase-1 (SCD-1) [41]. Suppression of genes involved in fatty acid synthesis, such as ACC, FAS, and SCD-1, leads to reduction of adipocyte triglyceride synthesis and accumulation [42]. In this study, we investigated the mRNA levels of genes involved in fatty acid synthesis, such as PPAR-γ, CEBP-α, aP2, ACC, FAS, and SCD-1 in 3T3-L1 adipocytes to investigate the mechanisms involved in the anti-adipogenic effect of TBEs. The 3T3-L1 adipocytes were treated with 100 µg/mL of TBE-50 and TBE-70 and incubated for 7 days. The mRNA levels of PPAR-γ, CEBP-α, aP2, ACC, FAS, and SCD-1 were significantly reduced by 39.3%, 52.0%, 31.3%, 44.0%, 34.7%, and 35.0%, respectively, in TBE-70 compared to untreated control cells (Table 1). TBE-50 significantly decreased the mRNA level of CEBP-α by 23% compared to the untreated control. Additionally, the mRNA level of CEBP-α, ACC, FAS, and SCD-1 was significantly lower (by 37.3%, 36.6%, 31.9%, and 32.9%, respectively) in the TBE-70 than TBE-50. In a previous study, it has been reported that oral administration of germinated buckwheat diminished fatty liver by suppressing the expression of key adipogenic transcriptional factors, such as PPAR-γ and CEBP-α in hepatocytes [43]. In addition, treatment of phenolic acids including rutin downregulates mRNA expression of PPAR-γ, C/EBP-α, and leptin, as well as upregulates adiponectin mRNA expression in differentiating adipocytes [29]. Our results indicate that the mRNA levels of PPAR-γ, CEBP-α, aP2, ACC, FAS, and SCD-1 decreased in cells treated with TBE-70 more so than those in cells treated with TBE-50. In contrast, TBE-50 only showed downregulation of CEBP-α expression. Therefore, the results indicate that TBEs may have anti-adipogenic effects in adipocytes, and the inhibitory effect could be in part explained by suppression of adipocyte-specific gene expression including PPAR-γ, CEBP-α, aP2, ACC, FAS, and SCD-1.
Table 1.
Effects of TBEs on mRNA expression of adipocytes-specific genes in adipocytes.
| Genes | Control | TBE-50 | TBE-70 |
|---|---|---|---|
| PPAR-γ | 1.00 ± 0.06 | 0.85 ± 0.13 | 0.61 ± 0.06 * |
| CEBP-α | 1.00 ± 0.07 | 0.77 ± 0.03 * | 0.48 ± 0.04 **,# |
| aP2 | 1.00 ± 0.02 | 0.86 ± 0.10 | 0.69 ± 0.01 * |
| ACC | 1.00 ± 0.12 | 0.88 ± 0.09 | 0.56 ± 0.03 *,# |
| FAS | 1.00 ± 0.12 | 0.88 ± 0.09 | 0.56 ± 0.03 *,# |
| SCD-1 | 1.00 ± 0.03 | 0.97 ± 0.10 | 0.65 ± 0.04 *,# |
mRNA level was measured using real-time quantitative polymerase chain reaction (qPCR). Values represent fold changes compared to the control. Data are expressed as mean ± SE of at least three independent experiments, each performed in triplicate (n = 3). * p < 0.05 and ** p < 0.01 vs. control. # p < 0.05 vs. TBE-50.
2.6. Effect of TBEs on mRNA Expression of Inflammatory Mediators and NO Production in Adipocytes
The tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), monocyte chemoattractant protein 1 (MCP-1), and inducible nitric oxide synthase (iNOS) are involved in the derivation and maintenance of chronic inflammatory responses in obesity [44,45]. In our study, to elucidate the inflammatory response of TBE, we analyzed the mRNA expression of inflammatory mediators such as TNF-α, IL-6, MCP-1, and iNOS, and nitric oxide (NO) production in adipocytes. As the lipid content in the adipose tissue increases, adipocytes synthesize TNF-α and IL-6, thus directly causing the inflammatory response [46]. Toll-like receptors (TLRs) are a family of molecules that are involved in the innate immunity. In obese adipocytes, the expression of several TLRs is elevated and activation of these receptors is thought to produce chemotactic signals, such as MCP-1, which provoke macrophage infiltration [47,48]. Activated macrophages in adipose tissue secrete pro-inflammatory cytokines, which leads to chronic low-grade inflammation in obesity [47]. It has been reported that treatment of citrus flavonoid naringenin during adipocyte differentiation inhibits TLR2 expression [48]. In addition, naringenin treatment suppresses NO production and TNF-α secretion from RAW 264.7 macrophages [49]. Moreover, coculture of 3T3-L1 adipocytes and RAW 264.7 macrophages enhances the production of TNF-α, MCP-1, and NO compared with the control cultures, while the treatment with naringenin chalcone dose-dependently inhibits the production of these pro-inflammatory mediators [50].
In this study, we investigated the mRNA expression of inflammatory mediators, such as TNF-α, IL-6, MCP-1, and iNOS, and NO production on TBE-50 and TBE-70 in 3T3-L1 cells. The cells were treated with 100 µg/mL of TBE-50 and TBE-70 and incubated for 7 days. TBE-70 decreased the mRNA levels of inflammatory mediators such as TNF-α, IL-6, MCP-1, and iNOS by 52.8%, 48.7%, 36.3% and 59.7%, respectively, compared with those in the untreated control (Table 2). TBE-50 decreased the mRNA level of iNOS by 35.7% compared to the untreated control. Further, the mRNA level of IL-6 and iNOS was significantly lower (by 32.5% and 37.3%, respectively) in the TBE-70 than TBE-50. NO production was significantly inhibited by 27.7% in TBE-70 compared to the control, and the TBE-70 was decreased more than TBE-50 (by 15.5%) (Figure 7). It has been reported that iNOS is a key mediator in obesity-induced inflammation, and an enzyme involved in the production of NO [51]. In a previous study, tartary buckwheat fractions and rutin effectively inhibited the production of ROS, NO, and IL-6, and downregulated the mRNA expression levels of pro-inflammatory factors including nuclear factor kappa B, cyclooxygenase-2, and iNOS in lipopolysaccharide (LPS)- and interferon-γ-stimulated RAW 264.7 cells [15]. In particular, rutin exhibits anti-inflammatory properties by inhibiting the release of TNF-α from monocytes [52] and human peripheral blood neutrophils [53]. Moreover, rutin exerts protective effects on inflammatory diseases such as acute pancreatitis [54], diabetic cardiomyopathy [55], neuroinflammation [56] in vivo. Meanwhile, phenolic acid present in tartary buckwheat-such as protocatechuic acid [57,58], syringic acid [59], ferulic acid [58], sinapic acid [60], quercetin [61], and rutin [62]-have shown beneficial effects on LPS-induced inflammation in RAW 264.7 cells have been reported, indicating that rutin and other phenolic acids present in TBEs might be multiply associated with anti-inflammatory effects of TBEs in adipocytes. In the present study, we first found that TBEs decreased the mRNA levels of inflammatory mediators, such as TNF-α, IL-6, MCP-1, and iNOS, and NO production during adipocyte differentiation, and these results suggest that TBEs would be beneficial in suppressing obesity-related inflammation.
Table 2.
Effects of TBEs on mRNA expression of inflammation mediators in adipocytes.
| Genes | Control | TBE-50 | TBE-70 |
|---|---|---|---|
| TNF-α | 1.00 ± 0.03 | 0.71 ± 0.06 | 0.47 ± 0.11 ** |
| IL-6 | 1.00 ± 0.09 | 0.76 ± 0.08 | 0.51 ± 0.04 **,# |
| MCP-1 | 1.00 ± 0.06 | 0.80 ± 0.06 | 0.64 ± 0.06 * |
| iNOS | 1.00 ± 0.10 | 0.64 ± 0.04 * | 0.40 ± 0.03 **,# |
mRNA level was measured using qPCR. Values represent fold changes compared to the control. Data are expressed as mean ± SE of at least three independent experiments, each performed in triplicate (n = 3). * p < 0.05 and ** p < 0.01 vs. control. # p < 0.05 vs. TBE-50.
Figure 7.

Effects of TBEs on NO production in adipocytes. 3T3-L1 adipocytes were treated with 100 µg/mL of TBE-50 or TBE-70, and incubated for 7 days. The production of NO was measured using Griess reagent. Values are expressed as mean ± SE (n = 3) of three independent experiments. * p < 0.05 vs. control. # p < 0.05 vs. TBE-50.
3. Materials and Methods
3.1. Materials
The 3T3-L1 cells were obtained from American Type Culture Collection (Manassas, VA, USA). Dulbecco’s modified Eagle’s medium (DMEM), glutamine, penicillin-streptomycin, fetal bovine serum (FBS), and TRIzol reagent were obtained from Invitrogen (Carlsbad, CA, USA). The cell count kit-8 (CCK-8) was purchased from Dojindo Laboratories (Kumamoto, Japan). An assay kit for TG was obtained from Asan Pharmaceutical Co. (Seoul, Korea). The GPDH activity assay kit was from Takara (Kyoto, Japan). The bicinchoninic acid (BCA) protein assay kit was obtained from Thermo Scientific (Pittsburgh, PA, USA). Universal SYBR® Green PCR Master Mix was purchased from Qiagen (Chatsworth, CA, USA). M-MLV reverse transcriptase was purchased from Promega (Madison, WI, USA). The NO assay kit was purchased from Thermo Scientific (Pittsburgh, PA, USA).
3.2. Preparation of Tartary Buckwheat Extracts
The extracts of tartary buckwheat were kindly supplied by the SKBioland Co. (Ansan, Gyeonggi, Korea). Tartary buckwheat was purchased from buckwheat associative corporation in Jeju, Korea. The extraction was performed as follows: 100 g of tartary buckwheat was subjected to extraction with 50% or 70% ethanol (1:15 (w/v)) at 80 °C for 3 h, and was then filtered. The filtrate was vacuum evaporated and the concentrated liquid was spray-dried (Dongjin ENG Inc., Siheung, Korea) and used as TBE-50 and TBE-70, respectively.
3.3. HPLC Analysis
The rutin content of TBE-50 and TBE-70 was determined by HPLC. Briefly, each sample was dissolved in methanol in 1 mg/mL concentration and was filtered through a 0.45 μm PVDF syringe filter. The Agilent technologies 1260 infinity (Agilent Technologies, Santa Clara, CA, USA) with a ZORBAX Eclipse XDB-C18 column (250 mm × 4 mm, 5 μm pore size, Agilent technologies) was used for analysis. The mobile phases consisted of 0.1% formic acid in water (Solvent A) and 0.1% formic acid in acetonitrile (Solvent B). The gradient elution used was as follows: 0–5 min, 5% B, 5–30 min, 10% B; 30–40 min, 40% B; 40–45 min, 90% B; 45–55 min, 5% B. The flow rate was 0.3 mL/min and the volume of sample injection was 20 μL. The column temperature was constant at 30 °C.
3.4. Cell Culture
The culture of 3T3-L1 cells was initially conducted in DMEM containing 10% (v/v) FBS, 100 units/mL penicillin, 100 μg/mL streptomycin, and 2 mM glutamine under conditions of 37 °C and 5% CO2. For the induction of adipocyte differentiation, the preadipocytes were cultured to confluence (Day 0, d0) and exposed to a differentiation medium containing 0.5 mM isobutylmethylxanthine, 1 μM dexamethasone, and 5 μg/mL insulin (MDI) for 2 days (d2). The cells were cultured with 5 μg/mL insulin for 2 days (d4) and then subsequently cultured in DMEM containing 10% FBS for 5 days (d9). TBE-50 and TBE-70 were treated with the medium for 7 days (from d2 to d9). Cells incubated without treatment served as a control. All assays were carried out in triplicate.
3.5. Cell Viability Assay
The cytotoxicity of TBE-50 and TBE-70 in adipocytes was evaluated by the WST-8 [2-(2-methoxy-4-nitropheyl)-3-(4-nitrophenyl)-5-(2,4-dinitrophenyl)-2H-tetrazolium, monosodium salt method, using a commercial CCK-8 kit as described previously [63]. Differentiated 3T3-L1 adipocytes were incubated for 1, 2, 5, or 7 days with TBE-50 or TBE-70 (at a dose of 0, 0.1, 1, 10, 50, 100, or 500 µg/mL). The absorbance was read using a Varioskan plate reader (Thermo Electron, Waltham, MA, USA) at 450 nm, and the results are expressed as the percentage of the control. All assays were carried out in triplicate.
3.6. Oil-Red O Staining
Lipid content was measured as previously described [63]. Briefly, 3T3-L1 adipocytes were treated with 100 μg/mL of TBE-50 or TBE-70. On Days 2, 5, and 7, the cells were rinsed in phosphate-buffered saline (PBS) (pH 7.4) and then fixed in 10% (v/v) formalin in PBS. To quantify accumulated lipid levels, cells were stained for 15 min with oil-red O dye (6 parts of saturated oil-red O in isopropanol and 4 parts of water). Oil droplets stained with oil-red O were dissolved in 4% (v/v) Nonidet P-40 in isopropanol and quantified by measuring absorbance at 520 nm [64]. The results are expressed as the percentage of the control.
3.7. TG Assay
For the measurement of intracellular TG, a colorimetric TG Assay kit was used according to the method as described previously [63]. 3T3-L1 adipocytes were lysed using a buffer containing 1% Triton X-100 in PBS, and the level of TG was determined using a commercial TG assay kit. Cellular TG content was then normalized to the protein concentration measured by a BCA protein assay kit.
3.8. GPDH Activity
GPDH activity was analyzed as described previously [65] using a commercial kit. In brief, 3T3-L1 adipocytes were treated with 100 μg/mL of TBE-50 or TBE-70. The cells were then disrupted by homogenization and centrifuged at 4 °C for 10 min. The supernatant was assayed for the GPDH activity by monitoring the decrease of NADH in the presence of dihydroxyacetone phosphate and measuring absorbance at 340 nm. Cellular protein level was determined using a BCA protein assay kit. The results are expressed as the percentage of the control.
3.9. Real-Time qPCR
Total RNA was extracted from 3T3-L1 adipocytes by using TRIzol Reagent. The cDNAs were synthesized from 4 μg RNA by using M-MLV reverse transcriptase. Quantitative real-time PCR was then carried out in 25 μL of Universal SYBR® Green PCR Master Mix using a fluorometric thermal cycler (Rotor-GeneTM 2000; Corbett Research, Mortlake, NSW, Australia). Primer3 software (Version 2.3.6, MA, USA) was used for the primer design [66]. The sequences of the primers used are presented in Table 3. For relative quantification, the delta–delta Ct method was used [67], and β-actin was used as an endogenous control. Values presented represent fold changes compared to the control.
Table 3.
Primers used for qPCR.
| Name | GeneBank No. | Primer Sequence (5′-3′) |
|---|---|---|
| ACC | AY451393 | F: CAAGTGCTCAAGTTTGGCGC |
| R: CAAGAACCACCCCGAAGCTC | ||
| aP2 | NM_024406 | F: CGACAGGAAGGTGAAGAGCA |
| R: ATTCCACCACCAGCTTGTCA | ||
| β-actin | NM_007393 | F: GGACCTGACAGACTACCTCA |
| R: GTTGCCAATAGTGATGACCT | ||
| CEBP-α | NM_007678 | F: ATAGACATCAGCGCCTACAT |
| R: TCCCGGGTAGTCAAAGTCAC | ||
| FAS | AF127033 | F: CTGGCATTCGTGATGGAGTC |
| R: TGTTTCCCCTGAGCCATGTA | ||
| IL-6 | NM_031168 | F: CCTTCCTACCCCAATTTCCA |
| R: TAACGCACTAGGTTTGCCGA | ||
| iNOS | BC062378.1 | F: GCTACTGGGTCAAAGACAAG |
| R: GCTGAACTTCCAGTCATTGT | ||
| MCP-1 | NM_019812 | F: TGCTGACCCCAAGAAGGAAT |
| R: TGAGGTGGTTGTGGAAAAGG | ||
| PPAR-γ | NM_011146 | F: TTGATTTCTCCAGCATTTCT |
| R: TGTTGTAGAGCTGGGTCTTT | ||
| SCD-1 | AF509567 | F: ATGGATATCGCCCCTACGAC |
| R: GATGTGCCAGCGGTACTCAC | ||
| TNF-α | NM_013693 | F: AGCACAGAAAGCATGATCCG |
| R: CCACAAGCAGGAATGAGAA |
3.10. NO Production
The production of NO was measured as nitrite using a commercial kit according to the manufacturer’s instructions. 3T3-L1 adipocytes were treated with 100 μg/mL of TBE-50 or TBE-70 for 7 days. The NO released into the medium was reacted with Griess reagent (1% sulfanilamide +0.1% naphthylendiamine dihydrochloride, 1:1) for 30 min at room temperature, and the absorbance was measured at 548 nm. The nitrite concentration was determined using sodium nitrite as a standard. The cellular protein concentration was determined using a BCA protein assay kit. The NO concentrations were normalized to the cellular protein content. Values are presented as percentage of untreated control.
3.11. Statistical Analysis
Values are expressed as mean ± standard error of the mean (SEM). Statistical analyses were performed using SPSS software (version 23; IBM Corporation, Armonk, NY, USA). The significance of differences between two groups of TBE-50 and TBE-70 with the same concentration was determined with a Student’s t-test (two-tailed). Significant differences among different concentrations of treatment group were analyzed using a one-way analysis of variance (ANOVA), followed by Tukey’s multiple comparison tests. p < 0.05 indicated a significant difference.
4. Conclusions
Taken together, our results demonstrate that TBEs may inhibit adipogenesis and inflammatory response during adipocyte differentiation of 3T3-L1 cells. These effects of TBEs were partially mediated by reducing GPDH activity and NO production, and by modulating the expression of genes involved in fatty acid synthesis and inflammatory mediators. Thus, TBEs may be useful as a potential food ingredient to prevent obesity-associated inflammation.
Acknowledgments
This study was supported by SKbioland, the Ministry of Agriculture, Food and Rural Affairs of the Korean Government, and the National Research Foundation of Korea (NRF) funded by the Korean Government (No. 2016R1A2B4011021).
Author Contributions
Y.K. and M.-S.L. conceived and designed the experiments. M.-S.L. carried out experiments and data analysis. Y.S. wrote the manuscript. S.J. and S.-Y.K. participated in several experiments. Y.-H.J. and C.-T.K. analyzed the TBE-50 and TBE-70. M.-K.Y., S.-J.L., J.S., and H.-J.Y. provided the TBE-50 and TBE-70. All authors read and approved the final manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Footnotes
Sample Availability: Not available.
References
- 1.Frühbeck G., Gómez-Ambrosi J., Muruzábal F.J., Burrell M.A. The adipocyte: A model for integration of endocrine and metabolic signaling in energy metabolism regulation. Am. J. Physiol. Endocr. Metab. 2001;280:E827–E847. doi: 10.1152/ajpendo.2001.280.6.E827. [DOI] [PubMed] [Google Scholar]
- 2.Tang Q.Q., Jiang M.S., Lane M.D. Possible role of SpI as repressor of the C/EBPa gene in the adipocyte differentiation program. Mol. Cell Biol. 1999;19:4855–4865. doi: 10.1128/MCB.19.7.4855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hill J.O., Peters J.C. Environmental contributions to the obesity epidemic. Science. 1998;280:1371–1374. doi: 10.1126/science.280.5368.1371. [DOI] [PubMed] [Google Scholar]
- 4.Alessi M.C., Lijnen H.R., Bastelica D., Juhan-Vague I. Adipose tissue and atherothrombosis. Pathophysiol. Haemost. Thromb. 2003;33:290–297. doi: 10.1159/000083816. [DOI] [PubMed] [Google Scholar]
- 5.Ahima R.S., Flier J.S. Adipose tissue as an endocrine organ. Trends Endocr. Metab. 2000;11:327–332. doi: 10.1016/S1043-2760(00)00301-5. [DOI] [PubMed] [Google Scholar]
- 6.Ouchi N., Parker J.L., Lugus J.J., Walsh K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011;11:85–97. doi: 10.1038/nri2921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wijngaard H.H., Arendt E.K. Buckwheat. Cereal Chem. 2006;83:391–401. doi: 10.1094/CC-83-0391. [DOI] [Google Scholar]
- 8.Eggum B.O., Kreft I., Javornik B. Chemical composition and protein quality of buckwheat (Fagopyrum esculentum Moench) Plant Foods Hum. Nutr. 1980;30:175–179. doi: 10.1007/BF01094020. [DOI] [Google Scholar]
- 9.Choi B.H. Status of buckwheat genetic resources in East Asia-1991. Korean J. Breed. Sci. 1992;24:293–301. [Google Scholar]
- 10.Campbell C.G. Buckwheat: Fagopyrum esculentum Moench. Volume 19. Bioversity International; Manitoba, MB, Canada: 1997. Major and minor production areas; p. 52. [Google Scholar]
- 11.Fabjan N., Rode J., Košir I.J., Wang Z., Zhang Z., Kreft I. Tartary buckwheat (Fagopyrum tataricum Gaertn.) as a source of dietary rutin and quercitrin. J. Agric. Food Chem. 2003;51:6452–6455. doi: 10.1021/jf034543e. [DOI] [PubMed] [Google Scholar]
- 12.Minghe Z., Fukang Q. Tartary flavonoids characteristics and its applications; Proceedings of the 7th International Symposium on Buckwheat; Winnipeg, MB, Canada. August 1998; pp. 40–45. [Google Scholar]
- 13.Nishimura M., Ohkawara T., Sato Y., Satoh H., Suzuki T., Ishiguro K., Nodac T., Morishitac T., Nishihiraa J. Effectiveness of rutin-rich Tartary buckwheat (Fagopyrum tataricum Gaertn.) ‘Manten-Kirari’ in body weight reduction related to its antioxidant properties: A randomised, double-blind, placebo-controlled study. J. Funct. Foods. 2016;26:460–469. doi: 10.1016/j.jff.2016.08.004. [DOI] [Google Scholar]
- 14.Hu Y., Hou Z., Liu D., Yang X. Tartary buckwheat flavonoids protect hepatic cells against high glucose-induced oxidative stress and insulin resistance via MAPK signaling pathways. Food Funct. 2016;7:1523–1536. doi: 10.1039/C5FO01467K. [DOI] [PubMed] [Google Scholar]
- 15.Choi S.Y., Choi J.Y., Lee J.M., Lee S., Cho E.J. Tartary buckwheat on nitric oxide-induced inflammation in RAW264. 7 macrophage cells. Food Funct. 2015;6:2664–2670. doi: 10.1039/C5FO00639B. [DOI] [PubMed] [Google Scholar]
- 16.Liu C.L., Chen Y.S., Yang J.H., Chiang B.H. Antioxidant activity of tartary (Fagopyrum tataricum (L.) Gaertn.) and common (Fagopyrum esculentum Moench) buckwheat sprouts. J. Agric. Food Chem. 2008;56:173–178. doi: 10.1021/jf072347s. [DOI] [PubMed] [Google Scholar]
- 17.Guo X., Zhu K., Zhang H., Yao H. Purification and characterization of the antitumor protein from Chinese tartary buckwheat (Fagopyrum tataricum Gaertn.) water-soluble extracts. J. Agric. Food Chem. 2007;55:6958–6961. doi: 10.1021/jf071032+. [DOI] [PubMed] [Google Scholar]
- 18.Yao Y., Shan F., Bian J., Chen F., Wang M., Ren G. d-chiro-inositol-enriched tartary buckwheat bran extract lowers the blood glucose level in KK-Ay mice. J. Agric. Food Chem. 2008;56:10027–10031. doi: 10.1021/jf801879m. [DOI] [PubMed] [Google Scholar]
- 19.Yoon B.R., Cho B.J., Lee H.K., Kim D.J., Rhee S.K., Hong H.D., Kim K.T., Cho C.W., Choi H.S., Lee B.Y., et al. Antioxidant and anti-adipogenic effects of ethanolic extracts from tartary and common buckwheats. Korean J. Food Pres. 2012;19:123–130. doi: 10.11002/kjfp.2012.19.1.123. [DOI] [Google Scholar]
- 20.Kurisawa M., Chung J.E., Uyama H., Kobayashi S. Enzymatic synthesis and antioxidant properties of poly (rutin) Biomacromolecules. 2003;4:1394–1399. doi: 10.1021/bm034136b. [DOI] [PubMed] [Google Scholar]
- 21.Sedej I., Sakač M., Mandić A., Mišan A., Tumbas V., Čanadanović-Brunet J. Buckwheat (Fagopyrum esculentum Moench) grain and fractions: Antioxidant compounds and activities. J. Food Sci. 2012;77:C954–C959. doi: 10.1111/j.1750-3841.2012.02867.x. [DOI] [PubMed] [Google Scholar]
- 22.Park B.J., Kwon S.M., Park J.I., Chan K.J., Park C.H. Phenolic compounds in common and tartary buchweat. Korean J. Crop Sci. 2005;50:175–180. [Google Scholar]
- 23.Morishita T., Yamaguchi H., Degi K. The contribution of polyphenols to antioxidative activity in common buckwheat and Tartary buckwheat grain. Plant Prod. Sci. 2007;10:99–104. doi: 10.1626/pps.10.99. [DOI] [Google Scholar]
- 24.Choi I., Park Y., Choi H., Lee E.H. Anti-adipogenic activity of rutin in 3T3-L1 cells and mice fed with high-fat diet. Biofactors. 2006;26:273–281. doi: 10.1002/biof.5520260405. [DOI] [PubMed] [Google Scholar]
- 25.Hsu C.L., Wu C.H., Huang S.L., Yen G.C. Phenolic compounds rutin and o-coumaric acid ameliorate obesity induced by high-fat diet in rats. J. Agric. Food Chem. 2009;57:425–431. doi: 10.1021/jf802715t. [DOI] [PubMed] [Google Scholar]
- 26.Gao M., Ma Y., Liu D. Rutin suppresses palmitic acids-triggered inflammation in macrophages and blocks high fat diet-induced obesity and fatty liver in mice. Pharm. Res. 2013;30:2940–2950. doi: 10.1007/s11095-013-1125-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Nafees S., Rashid S., Ali N., Hasan S.K., Sultana S. Rutin ameliorates cyclophosphamide induced oxidative stress and inflammation in Wistar rats: Role of NFκB/MAPK pathway. Chem. Biol. Interact. 2015;231:98–107. doi: 10.1016/j.cbi.2015.02.021. [DOI] [PubMed] [Google Scholar]
- 28.Yuandani, Jantan I., Husain K. 4,5,4′-Trihydroxychalcone, 8,8′-(ethene-1,2-diyl)-dinaphtalene-1,4,5-triol and rutin from Gynura segetum inhibit phagocytosis, lymphocyte proliferation, cytokine release and nitric oxide production from phagocytic cells. BMC Complement. Altern. Med. 2017;17:211. doi: 10.1186/s12906-017-1726-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Nikfarjam B.A., Adineh M., Hajiali F., Nassiri-Asl M. Treatment with Rutin-A Therapeutic Strategy for Neutrophil-Mediated Inflammatory and Autoimmune Diseases: Anti-inflammatory Effects of Rutin on Neutrophils. J. Pharmacopunct. 2017;20:52–56. doi: 10.3831/KPI.2017.20.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Abreu F.F., Souza A.C., Teixeira S.A., Soares A.G., Teixeira D.F., Soares R.C., Santana M.T., Lauton Santos S., Costa S.K., Muscará M.N., et al. Elucidating the role of oxidative stress in the therapeutic effect of rutin on experimental acute pancreatitis. Free Radic. Res. 2016;50:1350–1360. doi: 10.1080/10715762.2016.1247494. [DOI] [PubMed] [Google Scholar]
- 31.Saklani R., Gupta S.K., Mohanty I.R., Kumar B., Srivastava S., Mathur R. Cardioprotective effects of rutin via alteration in TNF-α, CRP, and BNP levels coupled with antioxidant effect in STZ-induced diabetic rats. Mol. Cell. Biochem. 2016;420:65–72. doi: 10.1007/s11010-016-2767-1. [DOI] [PubMed] [Google Scholar]
- 32.Hao G., Dong Y., Huo R., Wen K., Zhang Y., Liang G. Rutin inhibits neuroinflammation and provides neuroprotection in an experimental rat model of subarachnoid hemorrhage, possibly through suppressing the RAGE-NF-κB inflammatory signaling pathway. Neurochem. Res. 2016;41:1496–1504. doi: 10.1007/s11064-016-1863-7. [DOI] [PubMed] [Google Scholar]
- 33.Ðurendić-Brenese M., Popović T., Pilija V., Arsić A., Milić M., Kojić D., Jojić N., Milić N. Hypolipidemic and antioxidant effects of buckwheat leaf and flower mixture in hyperlipidemic rats. Bosnina J. Basic Med. Sci. 2013;13:100–108. doi: 10.17305/bjbms.2013.2389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Hsu C.L., Yen G.C. Effects of flavonoids and pehnolic acids on the inhibition of adipogenesis in 3T3-L1 adipocytes. J. Agric. Food Chem. 2007;55:8404–8410. doi: 10.1021/jf071695r. [DOI] [PubMed] [Google Scholar]
- 35.Koh E.J., Kim K.J., Seo Y.J., Choi J., Lee B.Y. Modulation of HO-1 by ferulic acid attenuates adipocyte differentiation in 3T3-L1 cells. Molecules. 2017;22:E745. doi: 10.3390/molecules22050745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Seo M.J., Lee Y.J., Hwang J.H., Kim K.J., Lee B.Y. The inhibitory effects of quercetin on obesity and obesity-induced inflammation by regulation of MAPK signaling. J. Nutr. Biochem. 2015;26:1308–1316. doi: 10.1016/j.jnutbio.2015.06.005. [DOI] [PubMed] [Google Scholar]
- 37.Eseberri I., Miranda J., Lasa A., Churruca I., Portillo M.P. Doses of quercetin in the range of serum concentrations exert delipidating effects in 3T3-L1 preadipocytes by acting on different stages of adipogenesis, but not in mature adipocytes. Oxid. Med. Cell. Longev. 2015;2015:480943. doi: 10.1155/2015/480943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bae C.R., Park Y.K., Cha Y.S. Quercetin-rich onion peel extract suppresses adipogenesis by downregulating adipogenic transcription factors and gene expression in 3T3-L1 adipocytes. J. Sci. Food Agric. 2014;94:2655–2660. doi: 10.1002/jsfa.6604. [DOI] [PubMed] [Google Scholar]
- 39.Spiegelman B.M., Frank M., Green H. Molecular cloning of mRNA from 3T3 adipocytes. Regulation of mRNA content for glycerophosphate dehydrogenase and other differentiation-dependent proteins during adipocyte development. J. Biol. Chem. 1983;258:10083–10089. [PubMed] [Google Scholar]
- 40.Ntambi J.M., Young-Cheul K. Adipocyte differentiation and gene expression. J. Nutr. 2000;130:3122S–3126S. doi: 10.1093/jn/130.12.3122S. [DOI] [PubMed] [Google Scholar]
- 41.Kang S.W., Kang S.I., Shin H.S., Yoon S.A., Kim J.H., Ko H.C., Kim S.J. Sasa quelpaertensis Nakai extract and its constituent p-coumaric acid inhibit adipogenesis in 3T3-L1 cells through activation of the AMPK pathway. Food Chem. Toxicol. 2013;59:380–385. doi: 10.1016/j.fct.2013.06.033. [DOI] [PubMed] [Google Scholar]
- 42.Prusty D., Park B.H., Davis K.E., Farmer S.R. Activation of MEK/ERK signaling promotes adipogenesis by enhancing peroxisome proliferator-activated receptor γ (PPARγ) and C/EBPα gene expression during the differentiation of 3T3-L1 preadipocytes. J. Biol. Chem. 2002;277:46226–46232. doi: 10.1074/jbc.M207776200. [DOI] [PubMed] [Google Scholar]
- 43.Thompson G.M., Trainor D., Biswas C., LaCerte C., Berger J.P., Kelly L.J. A high-capacity assay for PPARγ ligand regulation of endogenous aP2 expression in 3T3-L1 cells. Anal. Biochem. 2004;330:21–28. doi: 10.1016/j.ab.2004.03.061. [DOI] [PubMed] [Google Scholar]
- 44.Duplus E., Forest C. Is there a single mechanism for fatty acid regulation of gene transcription? Biochem. Pharmacol. 2002;64:893–901. doi: 10.1016/S0006-2952(02)01157-7. [DOI] [PubMed] [Google Scholar]
- 45.Thampy K.G., Koshy A.G. Purification, characterization, and ontogeny of acetyl-CoA carboxylase isozyme of chick embryo brain. J. Lipid Res. 1991;32:1667–1673. [PubMed] [Google Scholar]
- 46.Ntambi J.M., Miyazaki M. Recent insights into stearoyl-CoA desaturase-1. Curr. Opin. Lipidol. 2003;14:255–261. doi: 10.1097/00041433-200306000-00005. [DOI] [PubMed] [Google Scholar]
- 47.Wong C.P., Kaneda T., Morita H. Plant natural products as an anti-lipid droplets accumulation agent. J. Nat. Med. 2014;68:253–266. doi: 10.1007/s11418-014-0822-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Choi I., Seog H., Park Y., Kim Y., Choi H. Suppressive effects of germinated buckwheat on development of fatty liver in mice fed with high-fat diet. Phytomedicine. 2007;14:563–567. doi: 10.1016/j.phymed.2007.05.002. [DOI] [PubMed] [Google Scholar]
- 49.Carvalho Filho M.A., Ueno M., Carvalheira J.B., Velloso L.A., Saad M.J. Targeted disruption of iNOS prevents LPS induced S nitrosation of IRbeta/IRS 1 and Akt and insulin resistance in muscle of mice. Am. J. Physiol. Endocr. Metab. 2006;291:E476–E482. doi: 10.1152/ajpendo.00422.2005. [DOI] [PubMed] [Google Scholar]
- 50.Hsu C.L., Lin Y.J., Ho C.T., Yen G.C. The inhibitory effect of pterostilbene on inflammatory responses during the interaction of 3T3 L1 adipocytes and RAW 264.7 macrophages. J. Agric. Food Chem. 2013;61:602–610. doi: 10.1021/jf304487v. [DOI] [PubMed] [Google Scholar]
- 51.Hotamisligil G.S., Shargill N.S., Spiegelman B.M. Adipose expression of tumor necrosis factor-alpha: Direct role in obesity-linked insulin resistance. Science. 1993;259:87–91. doi: 10.1126/science.7678183. [DOI] [PubMed] [Google Scholar]
- 52.Kanda H., Tateya S., Tamori Y., Kotani K., Hiasa K.I., Kitazawa R., Kitazawa S., Miyachi H., Maeda S., Egashira K., et al. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J. Clin. Investig. 2006;116:1494–1505. doi: 10.1172/JCI26498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Yoshida H., Watanabe W., Oomagari H., Tsuruta E., Shida M., Kurokawa M. Citrus flavonoid naringenin inhibits TLR2 expression in adipocytes. J. Nutr. Biochem. 2013;24:1276–1284. doi: 10.1016/j.jnutbio.2012.10.003. [DOI] [PubMed] [Google Scholar]
- 54.Lyu S.Y., Park W.B. Production of cytokine and NO by RAW 264.7 macrophages and PBMC in vitro incubation with flavonoids. Arch. Pharm. Res. 2005;28:573–581. doi: 10.1007/BF02977761. [DOI] [PubMed] [Google Scholar]
- 55.Hirai S., Kim Y.I., Goto T., Kang M.S., Yoshimura M., Obata A., Yu R., Kawada T. Inhibitory effect of naringenin chalcone on inflammatory changes in the interaction between adipocytes and macrophages. Life Sci. 2007;81:1272–1279. doi: 10.1016/j.lfs.2007.09.001. [DOI] [PubMed] [Google Scholar]
- 56.Fujimoto M., Shimizu N., Kunii K., Martyn J.A., Ueki K., Kaneki M. A role for iNOS in fasting hyperglycemia and impaired insulin signaling in the liver of obese diabetic mice. Diabetes. 2005;54:1340–1348. doi: 10.2337/diabetes.54.5.1340. [DOI] [PubMed] [Google Scholar]
- 57.Lee J., Ha S.J., Lee H.J., Kim M.J., Kim J.H., Kim Y.T., Song K.M., Kim Y.J., Kim H.K., Jung S.K. Protective effect of Tremella fuciformis Berk extract on LPS-induced acute inflammation via inhibition of the NF-κB and MAPK pathways. Food Funct. 2016;7:3263–3272. doi: 10.1039/C6FO00540C. [DOI] [PubMed] [Google Scholar]
- 58.Yoo S.R., Seo C.S., Lee N.R., Shin H.K., Jeong S.J. Phytochemical analysis on quantification and the inhibitory effects on inflammatory responses from the fruit of xanthii fructus. Pharmacogn. Mag. 2015;11:S585–S591. doi: 10.4103/0973-1296.172966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Yoo S.R., Jeong S.J., Lee N.R., Shin H.K., Seo C.S. Simultaneous determination and anti-inflammatory effects of four phenolic compounds in Dendrobii Herba. Nat. Prod. Res. 2017:10:1–10:4. doi: 10.1080/14786419.2017.1300798. [DOI] [PubMed] [Google Scholar]
- 60.Yun K.J., Koh D.J., Kim S.H., Park S.J., Ryu J.H., Kim D.G., Lee J.Y., Lee K.T. Anti-inflammatory effects of sinapic acid through the suppression of inducible nitric oxide synthase, cyclooxygase-2, and proinflammatory cytokines expressions via nuclear factor-kappaB inactivation. J. Agric. Food Chem. 2008;56:10265–10272. doi: 10.1021/jf802095g. [DOI] [PubMed] [Google Scholar]
- 61.Endale M., Park S.C., Kim S., Kim S.H., Yang Y., Cho J.Y., Rhee M.H. Quercetin disrupts tyrosine-phosphorylated phosphatidylinositol 3-kinase and myeloid differentiation factor-88 association, and inhibits MAPK/AP-1 and IKK/NF-κB-induced inflammatory mediators production in RAW 264.7 cells. Immunobiology. 2013;218:1452–1467. doi: 10.1016/j.imbio.2013.04.019. [DOI] [PubMed] [Google Scholar]
- 62.Agbo M.O., Nworu C.S., Okoye F.B., Osadebe P.O. Isolation and structure elucidaton of polyphenols from Loranthus micranthus Linn. parasitic on Hevea brasiliensis with antiinflammatory property. EXCLI. J. 2014;13:859–868. [PMC free article] [PubMed] [Google Scholar]
- 63.Lee M.S., Kim C.T., Kim I.H., Kim Y. Effects of Capsaicin on Lipid Catabolism in 3T3-L1 Adipocytes. Phytother. Res. 2011;25:935–939. doi: 10.1002/ptr.3339. [DOI] [PubMed] [Google Scholar]
- 64.Moreno-Aliaga M.J., Matsumura F. Endrin inhibits adipocyte differentiation by selectively altering expression pattern of CCAAT/enhancer binding protein-α in 3T3-L1 cells. Mol. Pharmacol. 1999;56:91–101. doi: 10.1124/mol.56.1.91. [DOI] [PubMed] [Google Scholar]
- 65.Lee M.S., Jung S., Oh S., Shin Y., Kim C.T., Kim I.H., Kim Y. Effect of high hydrostatic pressure extract of fresh ginseng on adipogenesis in 3T3-L1 adipocytes. J. Sci. Food Agric. 2015;95:2409–2415. doi: 10.1002/jsfa.6961. [DOI] [PubMed] [Google Scholar]
- 66.Rozen S., Skaletsky H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 1999;132:365–386. doi: 10.1385/9259:365. [DOI] [PubMed] [Google Scholar]
- 67.Livak K.J., Schmittgen T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]