

Abstract
The mechanism of rapid energy supply to the brain, especially to accommodate the heightened metabolic activity of excited states, is not well-understood. We explored the role of glycogen as a fuel source for neuromodulation using the noradrenergic stimulation of glia in a computational model of the neural-glial-vasculature ensemble (NGV). The detection of norepinephrine (NE) by the astrocyte and the coupled cAMP signal are rapid and largely insensitive to the distance of the locus coeruleus projection release sites from the glia, implying a diminished impact for volume transmission in high affinity receptor transduction systems. Glucosyl-conjugated units liberated from glial glycogen by NE-elicited cAMP second messenger transduction winds sequentially through the glycolytic cascade, generating robust increases in NADH and ATP before pyruvate is finally transformed into lactate. This astrocytic lactate is rapidly exported by monocarboxylate transporters to the associated neuron, demonstrating that the astrocyte-to-neuron lactate shuttle activated by glycogenolysis is a likely fuel source for neuromodulation and enhanced neural activity. Altogether, the energy supply for both astrocytes and neurons can be supplied rapidly by glycogenolysis upon neuromodulatory stimulus.
Author summary
Although efficient compared to computers, the human brain utilizes energy at 10-fold the rate of other organs by mass. How the brain is supplied with sufficient on-demand energy to support its activity in the absence of neuronal storage capacity remains unknown. Neurons are not capable of meeting their own energy requirements, instead energy supply in the brain is managed by an oligocellular cartel composed of neurons, glia and the local vasculature (NGV), wherein glia can provide the ergogenic metabolite lactate to the neuron in a process called the astrocyte-to-neuron shuttle (ANLS). The only means of energy storage in the brain is glycogen, a polymerized form of glucose that is localized largely to astrocytes, but its exact role and conditions of use are not clear. In this computational model we show that neuromodulatory stimulation by norepinephrine induces astrocytes to recover glucosyl subunits from glycogen for use in a glycolytic process that favors the production of lactate. The ATP and NADH produced support metabolism in the astrocyte while the lactate is exported to feed the neuron. Thus, rapid energy demands by both neurons and glia in a stimulated brain can be met by glycogen mobilization.
Introduction
The management of energy in the brain is organized by an oligocellular cooperative called the neural-glial-vasculature ensemble (NGV). Each component is assigned distinct tasks during the chain of events that extract reducing equivalents from glucose to support every brain function. While the continuous supply of energy to the brain is critical for basal functions, rapid boosts in energy demand during higher states of alertness, often in response to neuromodulatory signals, must also be met. There is much controversy about how this kind of brain activity is supported energetically. What is agreed upon is that glucose, glycogen and lactate are the lead actors, with a cadre of support from intermediate metabolites [1–10]. The plot is complicated by dynamic changes in the relative contributions and timing of their roles; sorting all this out requires the insights provided by computational models.
The relationship among the NGV components is still being revealed with increasing interest in the role of glycogen—a form of polymerized glucose that constrains the energy storage capacity in the brain [2,9–15]. It has long been observed that brain glycogen resides almost exclusively in astrocytes [16–18], although its conservative presence in neurons has been noted and associated with hypoxia resistance [19]. Recent studies have more precisely located glycogen granules to the astrocytic lamelliform processes that ensheath synapses[20–23]. In fact, among the first indications of the complexity of coupling between neurons and astrocytes were the observations that synaptic and neuromodulatory activity promote glycogen hydrolysis in the mouse cerebral cortex [5,24]. Brain glycogen is the largest repository of energy in the brain, retaining more glucose equivalents than the amount dissolved in the cytosol, and can supplement the brain for more than an hour under conditions of hypoglycaemia [10].
The concept of the role of glycogen has evolved from a mere glucose storage depot for crisis management [25] to being part and parcel of the dynamic energy milieu [15,26–29]. The on-going turnover of glycogen involves the so-called glycogen shunt in which some of the blood-borne glucose imported into the astrocyte is stored as glycogen before becoming available for glycolysis via glycogenolysis [9,15,30,31].
Glycogenolysis not only contributes to commonplace energy supply [2,5,6,8,15,32–40], but also to handling special requests including stability maintenance during hypoglycemia [41], responding to rapid and high-demand needs signaled by neuromodulatory factors such as norepinephrine (NE) [4], higher local energy demand due to regional stimulation [42–45], memory formation and consolidation [35,46–51] drug addiction [52], as well as sleep and development [29,53,54].
The locus coeruleus (LC) in the brainstem sends far-reaching projections throughout numerous brain regions. In the cortex, these inputs effect neuromodulatory control of arousal, attention and memory via the LC-norepinephrine (LC-NE) arousal circuit [55–57]. The NE is released from axonal varicosities from which it diffuses to find adrenergic receptors on neurons [58] and astrocytes [59]. The activation of β2-adrenergic receptors (β2R) on astrocytes by the volume transmitted NE [60] is thought to mediate the neuromodulatory stimulus-demanding energy supply and consumption in the NGV, with glycogen implicated as a key supplier of lactate [61–71].
Turnover of glycogen in astrocytes is triggered by NE from LC inputs and involves signal transduction mediated by adenyl cyclase and the second messenger cAMP [68,72,73]. Glycogen and β-adrenergic dysregulation are associated with neurodegeneration [46,74] and astrocytic β2 receptors mediate hippocampal long-term memory consolidation and stress response management through training-dependent lactate production [47]. Neuromodulatory stimuli can mobilize more than half of stored glycogen; such glucose dumping could provide rapid and large energy injections into the NGV system [75]. In the cortex, NE containing varicosities are found near glia throughout development and adulthood concomitant with the expression of glycogen, suggesting a persistent role for this pathway, [6,48,66,76–79], and NE release from the LC modulates glycogenolysis and memory consolidation via β2-adrenergic receptors [77,80]. The consumption of glycogen upon circuit activity in cortex [81,82] and its activation and mobilization appear to be rapid [35].
Of particular importance to brain energy supply is the lactate derived from glycolysis in the astrocyte and which is required to support higher metabolic brain activities, including during intense exercise [83], in response to neuromodulation [61,68,71,84] and in support of memory formation [47,50,85]. The production of lactate by whatever means is followed by its export to neighboring neurons through monocarboxylate transporters (MCTs) in a process called the astrocyte-to-neuron lactate shuttle (ANLS) [7,86–89].
This computational model tests the feasibility that glycogenolysis within the NGV ensemble can respond rapidly and sufficiently to provide energy for both astroctyes and neurons in response to neuromodulatory signals [90]. We built on our previous computational model of ANLS to explore the dynamics of glycogen mobilization by NE release from LC terminals and test whether existing knowledge of the enzymatic cascades supports the role of glycogen as a source of energy both to astrocytes and neurons. We observed a rapid degradation of glycogen, expected enzymatic cascades, the production of NADH and ATP and lactate for the neuron via ANLS [7,8,87]. In addition, volume transmission resulting from differences in release distances between the LC terminals and the astrocyte is unlikely to influence outcome, at least in a high ligand affinity second messenger transduction pathway. These results support the idea that glycogenolytic energy supports the enhanced metabolic demand of neuromodulation.
Results
Overview
After using 3D electron microscopy (EM) to determine the locations of glycogen granules in the somatosensory cortex, we employed a computational approach to elucidate the role of glycogen in supporting neuromodulation by building upon our previous NGV model [87]. New model features include a complex, multi-step glycogenolysis pathway, neuromodulation via the LC-NE system in the cortex, and second messenger transduction (cAMP) [91]. We simulated astrocytic stimulation by LC noradrenergic inputs with a focus on the contribution of glycogenolysis to the local and exported energy supplies including the role of lactate shuttling from the astrocyte to the neighboring neuron (ANLS) [7,89].
3D electron microscopy of murine somatosensory cortex
While it has been established that glycogen is located in astrocytes, we further explored the subcellular distribution of glycogen granules within six astrocytic processes from layer I mice somatosensory cortex[92,93] (Fig 1A). We measured the number of granules apparent over a period of 4 (n = 3) and 24 (n = 3) months in 3D reconstruction from EM stacks of 125 cubic micrometers volumes of neuropil. In order to obtain the density of glycogen granules, we divided the total number of granules per each of the reconstructed volumes (3038, 3738 and 11809 in 4 months old, and 6588, 7758 and 4287 in 24 months old) per the volume of the reconstructed astrocyte (10.7 μm3, 10.8 μm3, 17.9 μm3 and 10.6 μm3, 12.3 μm3, 6.5 μm3 for 4 and 24 months old, respectively) and found a stable distribution between the two populations (Fig 1B).
Fig 1. Visualization of glycogen granules in a 3D reconstructed astrocyte.

A) Rendering of one astrocytic process (grey), in semi-transparency to highlight the presence within its cytosol of the glycogen granules. B) Top and bottom panels, rendering of two reconstructed astrocytic processes (green, adult, red, aged), semi-transparent to show the intracellular content of glycogen granules (grey). Whisker plot of the density of glycogen granules per astrocytic process in adult (4 months old 442.3 ± 112.2 granules / μ3, n = 3) and aged (24 months old; 526.3 ±98.6 granules / μ3, n = 3).
Modeling glycogenolysis stimulated by LC-NE volume transmission to astrocytes
Model diagram
We integrated selected features of our previous NGV model [87] with two new computational modules: one for NE neurotransmission and cAMP second messenger transduction and one for glycogen metabolism (Fig 2A illustrates the compartmental scheme). The parameters for the neuromodulation and glycogen modules can be found in S3 Table.
Fig 2. Noradrenergic modulation in glia.

A) Schematic compartmental diagram of the NGV model with noradrenergic locus coeruleus (LC) inputs, astrocyte, extracellular and neuronal compartments. The vasculature blood flow has been clamped for these simulations for simplicity. B) Distance of NE release site from astrocyte was simulated as differences in rise time constant (four NE waveforms with τrise = 1, 10, 100 and 1000 sec). C) Corresponding NE receptor (β2R) activation levels show maximum receptor activation to each NE waveform. Inset: time domain zoom. Astrocytic β2R receptor activation is largely invariant except within initial 50 ms from neurotransmitter release when source of NE release is varied over 4 orders of magnitude. D) Dose-response relationship for NE and β2R activation (Kd = 300 nM). E) cAMP production levels in response to 1 second pulses of NE and τrise = 10 (representing a constant, relatively close proximity of the LC input, see S1 Text) at 4 different adenylate cyclase amplification factors selected in order to empirically produce a wide dynamic range of cAMP. Inset: time domain zoom. NE duration indicated by gray shaded areas.
Norepinephrine—β2-adrenergic receptor dynamics
We simulated the release of NE from LC varicosities by creating simple waveforms of NE with single rise and decay time constants. Volume transmission of NE at four distances from the astrocyte was simulated by varying the rise time constant (τrise) of the NE wave front as it encountered the astrocytic β2R; this would clearly impact the amount of NE reaching the astrocytic receptors. These waveforms were 10 seconds in duration at τrise = 10, 100, 1000, or 10000 ms, (Fig 2B). The activation of the β2R to each of these release patterns demonstrated that the high affinity of the receptor (Kd = 300 nM) makes for an almost all or nothing response to NE no matter what the waveform or corresponding concentration might be (Fig 2C and inset). A dose-response relationship for NE and normalized β2R activity demonstrated the functional concentration range for our ligand-receptor system (Fig 2D).
Based on the results from simulations of NE release (see S1 Text), we chose τrise = 10ms for the remainder of the simulations reported in this study. We then chose 4 adenylate cyclase amplification factors so as to yield a wide dynamic range of cAMP production in the astrocyte in response to the NE stimulus (Fig 2E and inset zoom). The duration of NE application is indicated by the gray shaded area in all relevant figures henceforth.
Enzyme cascade resulting from cAMP formation
The expected sequence of enzyme activations in response to NE-elicited cAMP was observed including protein kinase A (PKA), glycogen phosphorylase a (GPa), hexokinase/phosphofructokinase combined (HKPFK), phosphoglycerate kinase (PGK), pyruvate kinase (PK) and lactate dehydrogenase (LDH) (Fig 3A, real values; Fig 3B normalized, zoomed insets in both panels A and B focus on rise trajectories showing the slower development of LDH). Although the responses begin in less than 1 sec, it takes about 6 seconds for the group of enzymes to reach their (1-1/e) fold levels. The expected inverse activation relationships between protein phosphatase 1 (PP1) and PP1 bound to GPa (PP1-GPa), as well as the between GPa and GSa, were accurately simulated (Fig 3C).
Fig 3. Activation of glycolytic enzyme cascade by cAMP in the astrocytic compartment.
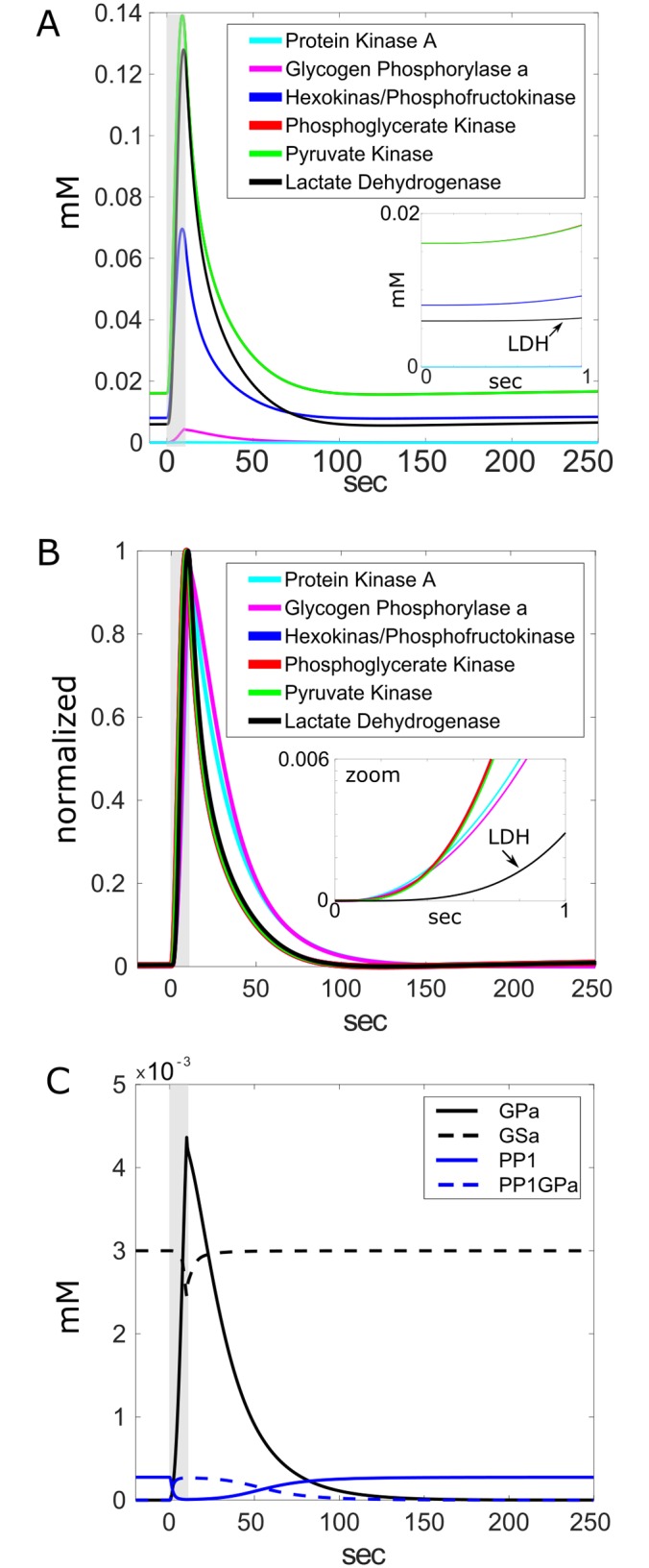
A) The sequence of glycolytic enzyme cascade includes: protein kinase A (PKA), glycogen phosphorylase a (GPa), hexokinase/phosphofructokinase combined (HKPFK), phosphoglycerate kinase (PGK), pyruvate kinase (PK) and lactate dehydrogenase (LDH). B) Responses are normalized to emphasize temporal relationship. Insets in A and B: Zoom-in showing later activation of LDH. C) Separated from other enzymes for clarity, reciprocal enzyme relationships shown for protein phosphatase 1 (PP1) and when complexed with glycogen phosphorylase (PP1-GPa), as well as GPa and glycogen synthase (GSa). NE duration indicated by gray shaded areas.
Metabolites and byproducts of glycolysis
The cascade of metabolites produced by the sequential activation of the battery of glycogenolytic and glycolytic enzymes was observed, including glucose-6-phosphate (G6P), glyceraldehyde-3-phosphate (GAP), phosphoenolpyruvate (PEP), pyruvate (PYR) and finally lactate (LAC) (percent increase featured in Fig 4A1, the glucose shown in panel 4A1 is only normalized ordinate in 4A2 to show smaller responses). Plotting the normalized responses reveals an extra-slow and long LAC response, as well as an undershoot of PYR and GAP (Fig 4A2). The glucose originating only from glycogen and is shown in panel 4A1 to illustrate the rapid conversion to G6P. The liberation of scores of mM equivalents of glucose that are quickly converted to G6P upon activation of cAMP pathways is not surprising considering the calculations in S2 Text that suggest an astrocyte might store hundreds of mM equivalents of glucose. The cytosolic glucose concentration, as well as that of other metabolites from panel 1, are shown in panel 3 of Fig 4A.
Fig 4. Production of metabolites by cAMP-dependent, NE-stimulated glycogen mobilization.

A1) Percent increase of sequence of metabolites triggered by cAMP including: glucose-6-phosphate (G6P), glyceraldehyde-3-phosphate (GAP), phosphoenolpyruvate (PEP), pyruvate (PYR) and lactate (LAC). Inset: zoom that better shows relative rises of smaller responses from phoshoenolpyruvate to lactate. A2) same metabolites as in A1 but normalized to emphasize longer response development and duration of lactate (LAC). A3) same metabolites as in A1, shown as concentrations. B) Production of ergogenic byproducts ATP and NADH in response to cAMP. B1) Percent increase showing relative magnitude. B2) Normalized traces showing relative time of activation. NE duration indicated by gray shaded areas. B3) same metabolites as in B1, shown as concentrations.
The robust production of the ergogenic byproducts ATP and NADH in response to cAMP was also observed. The relative magnitudes by percent increases indicate a larger cytosolic NADH response (Fig 4B1; >500% increase in NADH and 100% increase in ATP) and the normalized responses showing relative time course show a slower ATP response and an undershoot of mitochondrial NADH prior to stabilization (Fig 4B2). The concentrations of these metabolites are also shown in panel 4B3.
Glycogen mobilization and cellular energy status
NE-induced cAMP production in the astrocyte resulted in the degradation of glycogen that scaled with the dose of cAMP in the astrocyte (Fig 5A). For all doses significant degradation of glycogen appears in less than 5 seconds, with a time constant of decay at the largest dose of 29 seconds. Indicators of cellular energy status NAD+/NADH ratio (oxidative state, astrocytic cytosol) as well as energy charge ((ATP + 0.5ADP)/(ATP+ADP+AMP)) changed in response to the cAMP-dependent glycogenolysis within expected ranges (Fig 5B).
Fig 5. Glycogen and cellular energy status.
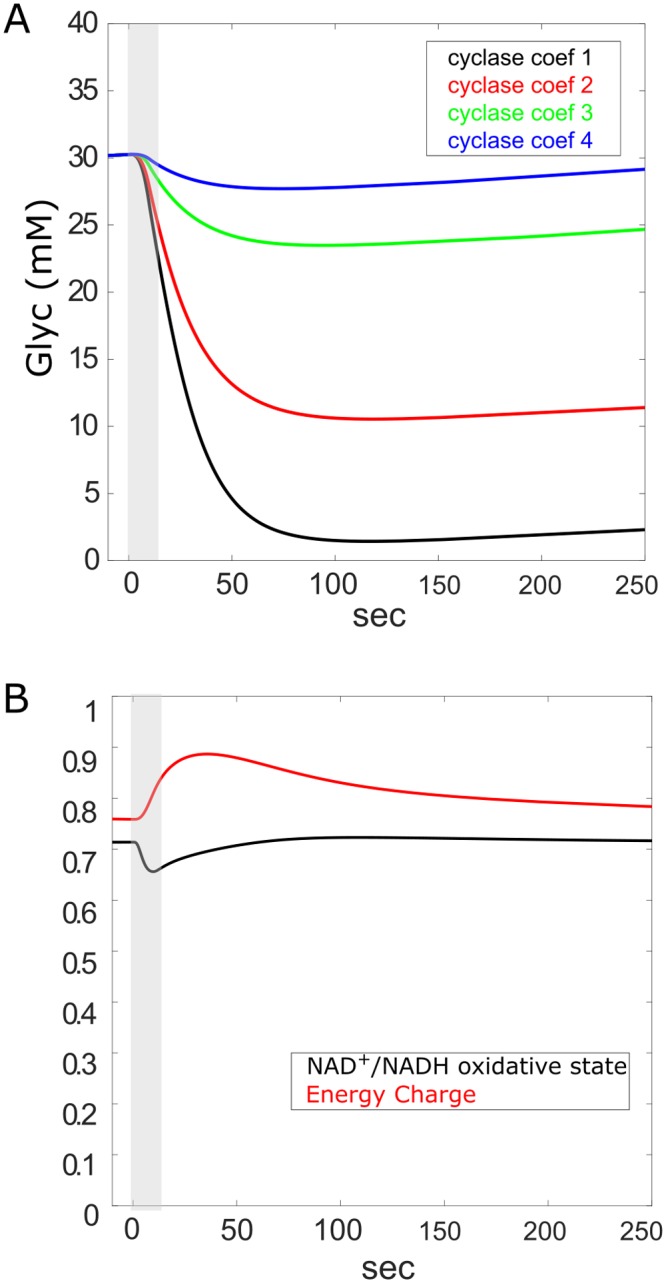
A) mobilization of glycogen in response to NE-triggered cAMP at each of 4 cyclase amplification coefficients (cyclase coef. in panels). B) Indicators of cellular energy status: astrocytic, cytosolic NAD+/NADH ratio (oxidative state) as well as energy charge ((ATP + 0.5ADP)/(ATP+ADP+AMP)) in response to cAMP. NE duration indicated by gray shaded areas.
Astrocyte-to-neuron lactate shuttle ANLS
Of particular interest to our current study was the production and fate of lactate from glycogenolysis and whether it can plausibly participate in the astrocyte-to-neuron lactate shuttle [7,87]. While the production of lactate in the astrocyte was demonstrated (Fig 4), we further examined to what degree the lactate could be exported and found robust and rapid transport of lactate to the extracellular space from where it was imported into the adjacent neuronal compartment (Fig 6A). When the lactate in the neuron, the extracellular space and the neuron were plotted together, evident was the similarity in the lactate transients, shifted only slightly in time as the wave of lactate passed from one compartment to the other. The rise time constant of the lactate response was 13 sec. The direction and timing of lactate flow in the NE- stimulated and cAMP-dependent ANLS is better seen by magnifying the traces (zoom in 6B).
Fig 6. Glycogen derived lactate shuttle.

A) Lactate (LAC) transients from 3 compartments in response to NE-dependent cAMP signaling. Responses from astrocyte, extracellular space and neuron all show same kinetics and are nearly overlapping, but slightly shifted in time, reflecting the transport time between compartments. B) Zoom-in of a region of almost overlapping LAC traces from 3 compartments that demonstrates the flow of LAC from astrocyte to extracellular space to neighboring neuron. Arrows indicate direction of LAC wave flow. C) ANLS with a characteristic lactate oxidative dip (upper panel, arrow) produced by synaptic excitation instead of NE stimulation (lower panel, V from neuron). The lactate dip is absent from the ANLS produced by glycogenolysis (panel A). NE duration indicated by gray shaded areas.
Our previous NGV model demonstrated the production of lactate from synaptic transmission activity that was characterized by an initial dip (corresponding to the use of lactate for energy) with a nadir around 20 seconds post stimulus (Fig 6C). In contrast, in this new study, the lactate signal resulting from NE-stimulated glycogenolysis lacked the initial dip, even in the neuron, and rose continuously with stimulus duration and decayed immediately upon stimulus cessation (Fig 6A). These results suggest that the astrocyte-to-neuron lactate shuttle (ANLS) activated by glycogenolysis, while lacking the initial oxidative dip, is at least as robust as that induced by synaptic activity.
Discussion
The role of glycogen in neuromodulation
The value of the role of glycogen in balancing the energy budget of the brain should not be discounted given its abundance in astrocytes in the vicinity of synapses [21,23] and experimental evidence for its involvement in supporting brain activity [15,35,40,48,49,65,68,82,94]. What is not clear is the feasibility of glycogen being able to respond rapidly and sufficiently enough to neuromodulators that regulate neuronal circuit activity and to what degree the ANLS is involved [47,51,83,85,89,95–97]. Since glycogenolysis has been suggested to provide energy to both neurons and astrocytes during learning, the involvement of lactate would be a likely candidate in this mechanism [49]. Accordingly, we have investigated the role of astrocytic glycogen in fueling and mediating neuromodulation in a computational model of glycogenolytic and noradrenergic transduction pathways along with elements of our previous NGV model [87].
Localization of glycogen
Anatomical evidence from 3D EM for the proximity of glycogen granules to synaptic regions in the somatosensory cortex demonstrates that glycogen is well-placed for a major role in the energetic support of brain activity (Fig 1). Although lower than muscle glycogen levels, brain glycogen is thought to store more glucosyl energy than soluble glucose [10] and our calculations support this view (S2 Text). One benefit of warehousing energy in the form of glycogen would be the buffering of glucose supplies locally without contributing to the osmotic tension associated with free glucose [20,21,23,23,98]. An additional advantage might be conveyed by reducing advanced glycation end products (AGEs) that are associated with age-related neurodegenerative disorders (e.g., [99]).
The EM results place glycogen near synapses, but to what extent is this source of energy destined for local astrocytic needs versus export for neuronal consumption? A summary of experimental evidence suggests both. Glycogen is degraded by neuronal stimulation [82], can sustain gray and white matter survival in the absence of glucose [100,101] and is required to provide fast local ATP to astrocytic SERCA pumps [6]. Many studies have shown that glycogen contributes to the constitutive requirements of active neurons and not simply for rapid energy needs [1,8,62,102]. In co-cultures of cerebellar neurons and astrocytes, energy from glycogen is required both to support astrocytic demands as well as for neurotransmitter release in the accompanying neuron [103]. ATP production in astrocytes depends on glycogenolysis [40] and glucose deprivation in cultured astrocytes leads to glycogen depletion and export of lactate [104].
Other research proposes that glycogen is mobilized to produce rapid energy during intense neuronal activity [31,105] since glycogenolysis can be initiated by neurotransmitters (e.g., NE and VIP) via a cAMP dependent mechanism [5,66,68,106,107] and this mechanism forms a component of the coupling mechanism between astroglial and neuronal energy metabolism within the NGV [108]. Glycogenolysis activated by NE inputs from the LC has been implicated in memory consolidation, even perhaps factoring into the etiology of Alzheimer’s disease [77,80], chronic stress-induced atrophy and depression [109], as well as diabetic neuropathy [110].
Lactate from glycogen
A preponderance of evidence thus far implicates the glycolytic metabolite lactate as the major energy vehicle for the astrocytic support of neuronal activity and cognitive functions [8,88,95,96,111–114]. Much as in muscle, lactate derived from glycogen can serve as an energy supply buffer between fast and slow energy requirements [115]. During intense neural activity, lactate derived from glycogen provides the necessary energy to sustain synaptic activity in the CNS [68,116,117].
The fate of lactate specifically produced from glycogen in astrocytes remains controversial. Lactate may be used in astrocytes where it can support local energy demands or be exported to neurons or parts unknown [49,118,119]. Other ergogenic molecules are derived from glycogen phosphorylation in the astrocyte such as NADH and ATP and remain there (Figs 3 and 4). Our model tested the viability of utilizing glycogen as a source of energy locally in the astrocyte or by the neuron, or both. Our simulation results reported here support the view that glycogen can feasibly support both roles when the astrocyte is stimulated by neuromodulatory signals. Mobilization of glycogen by NE-stimulated cAMP signaling rapidly degrades glycogen with a time constant of 29 seconds (Fig 5), resulting in the production of ATP and NADH for astrocytic use (Fig 4) and lactate that is produced with a time constant of 13 seconds and entirely shuttled to the neuron (Fig 6).
The fact that we observe a small increase in lactate compared to the very large amount G6P produced suggests that lactate production from glycogen may require concomitant kinetic control of rate-limiting glycolytic enzymes or priming reactions [120]. Glycogen degradation, therefore, may exert a leveraging effect on glycolysis in conjunction with other glycolytic signals. If this were to be the case, one would expect a much lower or more compartmentalized effect of cAMP on glycogenolysis in vivo. In either case, a much more detailed model in terms of reaction steps, regulation and spatial constraints should follow these results.
The results demonstrate the rapid production and export of lactate into the extracellular space and the neighboring neuron as a result of NE-stimulated cAMP production. The lactate exported to the neuron via MCTs stimulated the production of neuronal NADH similarly to the ANLS triggered by synaptic activity in our previous model (Fig 6A). The glycogen-derived NADH signal (Fig 4) mimics the experimental observation of [121] that related ANLS to increases in neuronal NADH. That glycogen can produce so much lactate to support neuronal functions, as well as NADH and ATP to support astocytic energy demands, is consistent with its observed role in preventing spreading depression through a mechanism that involves lactate [122].
The lack of an initial dip in lactate concentration (Fig 6B; as reported by [87]), which has been attributed to an initial oxidative consumption of lactate in the neuron in response to synaptic activity prior to eventual increases in production [123], suggests that the cAMP-dependent mobilization of large amounts of glucose from glycogen stores is anaerobic and that the presence or absence of the dip could be a signature of aerobic or anaerobic lactate signaling, respectively. The dumping of glucose observed during glygogenolysis is consistent with the large amounts of glucose stored in glycogen (S2 Text) and supports the idea of a compartmentalization of energy resources [103,124,125]. If so much glucose were not stored in glycogen and rapidly metabolized to downstream products it would present a potentially lethal challenge to the astrocytes osmotic balance, especially in the small volumes where glycogen is found [20]. Subsequent iterations and improvements of this model will implement a separate compartment for the fine astrocytic processes surrounding synapses that contain glycogen.
Thus, to the already familiar ANLS described experimentally [7,87,97,102,126] and computationally from neuronal glutamatergic and electrical activity [86,87] we confidently add the plausibility of ANLS stimulated by glycogenolysis triggered by neuromodulation. Given the persistent lactate gradient from astrocytes to neurons [127], it is not surprising that lactate derived from any source would rapidly be transported by the array of MCTs and even high capacity ion channels in the astrocyte and neuron [88,113,128].
The LC-NE network
The simulation results also lend credence to the idea that β2Rs participate in long-term hippocampal learning via a mechanism involving lactate export to neurons [47,129], and validate the involvement of a lactate rescue of cocaine-induced conditioned memory when glycogenolysis is impaired [130]. The apparent irrelevance of the distance of NE release from the glia (Fig 2) suggests a system fine-tuned to detect and respond to neuromodulatory signals; the mechanism of using high-affinity receptors in a volume transmission scenario could effectively approximate wired transmission in a volume transmission setting [131,132].
β2R activation triggers astrocytic glycogenolysis and dysregulation of these mechanisms is associated with neurodegenerative diseases [133]. Impairment of β2R adrenergic expression on astrocytes has been associated with the etiology of multiple sclerosis with a mechanism possibly involving the dysregulation of glycogenolysis [74]. It is tempting, therefore, to speculate that the involvement of neuromodulation via astrocytes in neuropsychiatric diseases might be related to their role in energy supply [134–138].
Conclusions and predictions
The results of our 3D electron microscopy and computational modeling study supports the plausibility that glycogenolysis plays a major mechanistic role in fueling and transducing the neuromodulatory signals mediated by cAMP. Significantly, we conclude that 1) glycogen granule density in layer 1 of somatosensory cortex is stable between 4–24 months, the type of reliable expression that would be consistent with expectations for a fuel source responsible for support of on-demand activity; 2) the distance of NE release from the astrocyte is not critically important, implying that volume transmission effects can be mitigated by high-affinity receptor or rapid transduction systems; 3) glycogenolysis evoked by cAMP elevations generate energy in the form of ATP, NADH and lactate production, thus supplying energy to both the astrocyte and the neuron; and 4) astrocytic lactate derived from glycogen is shuttled rapidly and preferentially to the neuron (ANLS). 5) Altogether, our model supports observations of the involvement of glycogen and lactate in supplying energy to both astrocytes and neurons during learning events related to neuromodulatory inputs, as well as their involvement in related disease states [35,45,47–49,51,52,97,108,109,139]. 6) The success of the model validates our bottom-up modeling approach as a tool to complement and guide basic and disease-related experimental studies.
Methods
3D EM reconstruction
We reconstructed astrocytic processes and the glycogen granules within the astrocytic profiles of six volumes of 5x5x5 μm3 from FIBSEM image stacks (courtesy of Graham Knott, BioEM, EPFL, Switzerland). Original samples were acquired from layer I somatosensory cortex of wild type mice aged 4 and 24 months (N = 3 per sample). Astrocytes were reconstructed using the carving, semi-automated algorithm [140] of the ilastik 1.2 software (www.ilastik.org). Glycogen granules were reconstructed using the trakEM2 software, by placing a sphere in the center of each granule and adjusting its diameter to the size of the granule (Fig 1)
Modeling
Our modeling approach was to adapt our previous NGV model [87] by adding new modules without changing the previous equations or parameters except where required for integration of the new modules into the original model. We provide all the equations in this manuscript for ease of reference.
Simulation environment
All simulations were carried-out in NEURON [141], using a fixed time step of 3 μs with Euler integration, and was run either on a Ubuntu 14.04 LTS workstation with a 3.6 GHz Intel Core i7-4790 CPU and 15.6 GB RAM, or on the Blue Gene/Q in Lugano, Switzerland. Matlab was used for data analysis. We found that the model was highly sensitive to the fixed time-step required for integration into larger models due to the rapid and wide-range of biochemical reactions. Other researchers wishing to adapt our model to their purposes should consider a variable time-step.
Neurotransmitter diffusion
To quantify the effects of diffusion on the waveform of NE, we computed the summed concentration from a point release source at various lateral distances from the point of release of norephinephrine (NE) from the locus coeruleus (LC) varicosities to the astrocytes as a function of time (t) and lateral distance (xdist) according the procedures and equations in S1 Text. From these calculations, we chose a 10 ms rise time constant (τrise) for the majority of the simulations in this study and lengthened this value 3 additional orders of magnitude in order to simulate progressively distant terminals for a dose-response effect. Due to the saturation of the β2-adrenergic receptors (β2Rs) on the astrocytes by the NE from the LC, a scaling factor was introduced for the cAMP production by adenylate cyclase in order to produce a wide NE-cAMP dose-response relationship (Fig 1).
Glucose storage capacity of glycogen
We have made calculations of the glucose storage capacity of glycogen in astrocytes and the effect of glycogenic glucosyl liberation on intracellular glucose concentration (S2 Text) and found that glycogen is capable of storing hundreds of mM equivalents of glucose in one astrocyte. These calculations were made to support simulation results suggesting the release of scores of mM equivalents of glucose upon stimulation.
Glycogen module
We built our glycogen shunt module with components from our previous multi-scale NGV metabolic model [87], without re-optimizing or recalibration of the original model, such that each voxel in the circuit contains a unicompartmental point model of the system of differential equations. Most of the equations for the glycogen module were adapted from [91]. Additional mechanisms or rate constants were taken from [142] (cAMP kinase rate constants), [143] (cAMP decay time constant), [144] (Kd for NE). The use of the type of glycogen phosphorylase from [91] is supported by experimental results suggesting that glycogen in astrocytes is mobilized by the muscle form of the enzyme glycogen phosphorylase [145]. We chose the muscle pattern of regulation of glycogen phosphorylase over the liver because muscle and brain isozymes share greater identity with regard to nucleotide and deduced amino acid sequences and their role in responding to physiological activity is similar [146]. Our model incorporated the feature of a dynamic Kd in order to account for the interactions between GSa and GPa wherein GPa has an inhibitory effect on the activation of GS [91,147]. Model equations and rate constants appear in S1 and S2 Tables, respectively, while parameters can be found in S3 Table.
Neuromodulation-free simulations
In order to demonstrate the ANLS produced by neuronal excitation by glutamate in the absence of neuromodulation and glycogenolysis, the original NGV model [87] was used (Fig 6C) in the absence of the neuromodulation and glycogenolysis modules.
Supporting information
(DOCX)
(DOCX)
(DOCX)
(DOCX)
(DOCX)
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
Supported by a CRG grant from King Abdullah University of Science and Technology "KAUST-EPFL Alliance for Neuro-inspired High-Performance Computing", as well as ETH Board funding to the Blue Brain Project. Support by the NCCR Synapsy and the Prefargier Foundation to PJM are also acknowledged. Calculations were performed on the EPFL Blue Brain IV BlueGene/Q hosted at the Swiss National Supercomputing Center (CSCS) in Lugano. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Dienel GA. The metabolic trinity, glucose-glycogen-lactate, links astrocytes and neurons in brain energetics, signaling, memory, and gene expression. Neurosci Lett. 2015;637: 18–25. 10.1016/j.neulet.2015.02.052 [DOI] [PubMed] [Google Scholar]
- 2.Hertz L, Peng L, Dienel GA. Energy metabolism in astrocytes: high rate of oxidative metabolism and spatiotemporal dependence on glycolysis/glycogenolysis. J Cereb Blood Flow Metab Off J Int Soc Cereb Blood Flow Metab. 2007;27: 219–249. 10.1038/sj.jcbfm.9600343 [DOI] [PubMed] [Google Scholar]
- 3.Hui S, Ghergurovich JM, Morscher RJ, Jang C, Teng X, Lu W, et al. Glucose feeds the TCA cycle via circulating lactate. Nature. 2017;551: 115–118. 10.1038/nature24057 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Magistretti PJ, Morrison JH. Noradrenaline- and vasoactive intestinal peptide-containing neuronal systems in neocortex: functional convergence with contrasting morphology. Neuroscience. 1988;24: 367–378. [DOI] [PubMed] [Google Scholar]
- 5.Magistretti PJ, Morrison JH, Shoemaker WJ, Sapin V, Bloom FE. Vasoactive intestinal polypeptide induces glycogenolysis in mouse cortical slices: a possible regulatory mechanism for the local control of energy metabolism. Proc Natl Acad Sci U S A. 1981;78: 6535–6539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Muller MS. Functional impact of glycogen degradation on astrocytic signalling. Biochem Soc Trans. 2014;42: 1311–1315. 10.1042/BST20140157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Pellerin L, Magistretti PJ. Glutamate uptake into astrocytes stimulates aerobic glycolysis: a mechanism coupling neuronal activity to glucose utilization. Proc Natl Acad Sci U S A. 1994;91: 10625–10629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Pellerin L, Magistretti PJ. Sweet sixteen for ANLS. J Cereb Blood Flow Metab Off J Int Soc Cereb Blood Flow Metab. 2012;32: 1152–1166. 10.1038/jcbfm.2011.149 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Walls AB, Heimbürger CM, Bouman SD, Schousboe A, Waagepetersen HS. Robust glycogen shunt activity in astrocytes: Effects of glutamatergic and adrenergic agents. Neuroscience. 2009;158: 284–292. 10.1016/j.neuroscience.2008.09.058 [DOI] [PubMed] [Google Scholar]
- 10.Gruetter R. Glycogen: the forgotten cerebral energy store. J Neurosci Res. 2003;74: 179–183. 10.1002/jnr.10785 [DOI] [PubMed] [Google Scholar]
- 11.Falkowska A, Gutowska I, Goschorska M, Nowacki P, Chlubek D, Baranowska-Bosiacka I. Energy Metabolism of the Brain, Including the Cooperation between Astrocytes and Neurons, Especially in the Context of Glycogen Metabolism. Int J Mol Sci. 2015;16: 25959–25981. 10.3390/ijms161125939 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Magistretti PJ, Allaman I. Glycogen: a Trojan horse for neurons. Nat Neurosci. 2007;10: 1341–1342. 10.1038/nn1107-1341 [DOI] [PubMed] [Google Scholar]
- 13.Magistretti PJ, Allaman I. A Cellular Perspective on Brain Energy Metabolism and Functional Imaging. Neuron. 2015;86: 883–901. 10.1016/j.neuron.2015.03.035 [DOI] [PubMed] [Google Scholar]
- 14.Magistretti PJ, Pellerin L, Rothman DL, Shulman RG. Energy on demand. Science. 1999;283: 496–497. [DOI] [PubMed] [Google Scholar]
- 15.Obel LF, Müller MS, Walls AB, Sickmann HM, Bak LK, Waagepetersen HS, et al. Brain glycogen-new perspectives on its metabolic function and regulation at the subcellular level. Front Neuroenergetics. 2012;4: 3 10.3389/fnene.2012.00003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ibrahim MZ. Glycogen and its related enzymes of metabolism in the central nervous system. Adv Anat Embryol Cell Biol. 1975;52: 3–89. [DOI] [PubMed] [Google Scholar]
- 17.Phelps CH. Barbiturate-induced glycogen accumulation in brain. An electron microscopic study. Brain Res. 1972;39: 225–234. [DOI] [PubMed] [Google Scholar]
- 18.Watanabe H, Passonneau JV. Factors affecting the turnover of cerebral glycogen and limit dextrin in vivo. J Neurochem. 1973;20: 1543–1554. [DOI] [PubMed] [Google Scholar]
- 19.Duran J, Guinovart JJ. Brain glycogen in health and disease. Mol Aspects Med. 2015;46: 70–77. 10.1016/j.mam.2015.08.007 [DOI] [PubMed] [Google Scholar]
- 20.Cali C, Baghabra J, Boges DJ, Holst GR, Kreshuk A, Hamprecht FA, et al. Three-dimensional immersive virtual reality for studying cellular compartments in 3D models from EM preparations of neural tissues. J Comp Neurol. 2016;524: 23–38. 10.1002/cne.23852 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Cali C, Kare K, Boges DJ, Agus M, Magistretti PJ. Sparse reconstruction of neurons and glial cells of layer VI somatosensory cortex of a juvenile rat. 2017.
- 22.Pfeiffer-Guglielmi B, Fleckenstein B, Jung G, Hamprecht B. Immunocytochemical localization of glycogen phosphorylase isozymes in rat nervous tissues by using isozyme-specific antibodies. J Neurochem. 2003;85: 73–81. [DOI] [PubMed] [Google Scholar]
- 23.Oe Y, Baba O, Ashida H, Nakamura KC, Hirase H. Glycogen distribution in the microwave-fixed mouse brain reveals heterogeneous astrocytic patterns. Glia. 2016;64: 1532–1545. 10.1002/glia.23020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hof PR, Pascale E, Magistretti PJ. K+ at concentrations reached in the extracellular space during neuronal activity promotes a Ca2+-dependent glycogen hydrolysis in mouse cerebral cortex. J Neurosci Off J Soc Neurosci. 1988;8: 1922–1928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Siesjö BK. Brain energy metabolism and catecholaminergic activity in hypoxia, hypercapnia and ischemia. J Neural Transm Suppl. 1978; 17–22. [PubMed] [Google Scholar]
- 26.Brown AM, Baltan Tekkök S, Ransom BR. Energy transfer from astrocytes to axons: the role of CNS glycogen. Neurochem Int. 2004;45: 529–536. 10.1016/j.neuint.2003.11.005 [DOI] [PubMed] [Google Scholar]
- 27.Chambers TW, Daly TP, Hockley A, Brown AM. Contribution of glycogen in supporting axon conduction in the peripheral and central nervous systems: the role of lactate. Front Neurosci. 2014;8: 378 10.3389/fnins.2014.00378 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lavoie S, Allaman I, Petit J-M, Do KQ, Magistretti PJ. Altered glycogen metabolism in cultured astrocytes from mice with chronic glutathione deficit; relevance for neuroenergetics in schizophrenia. PloS One. 2011;6: e22875 10.1371/journal.pone.0022875 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Petit J-M, Magistretti PJ. Regulation of neuron-astrocyte metabolic coupling across the sleep-wake cycle. Neuroscience. 2015;323: 135–56. 10.1016/j.neuroscience.2015.12.007 [DOI] [PubMed] [Google Scholar]
- 30.Magistretti PJ, Allaman I. Brain Energy Metabolism In: Pfaff DW, editor. Neuroscience in the 21st Century: From Basic to Clinical. New York, NY: Springer New York; 2013. pp. 1591–1620. 10.1007/978-1-4614-1997-6_56 [DOI] [Google Scholar]
- 31.Shulman RG, Hyder F, Rothman DL. Cerebral energetics and the glycogen shunt: neurochemical basis of functional imaging. Proc Natl Acad Sci U S A. 2001;98: 6417–6422. 10.1073/pnas.101129298 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Brown AM. Brain glycogen re-awakened. J Neurochem. 2004;89: 537–552. 10.1111/j.1471-4159.2004.02421.x [DOI] [PubMed] [Google Scholar]
- 33.Choi HB, Gordon GRJ, Zhou N, Tai C, Rungta RL, Martinez J, et al. Metabolic communication between astrocytes and neurons via bicarbonate-responsive soluble adenylyl cyclase. Neuron. 2012;75: 1094–1104. 10.1016/j.neuron.2012.08.032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Dinuzzo M, Mangia S, Maraviglia B, Giove F. The role of astrocytic glycogen in supporting the energetics of neuronal activity. Neurochem Res. 2012;37: 2432–2438. 10.1007/s11064-012-0802-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Gibbs ME, Hutchinson DS. Rapid turnover of glycogen in memory formation. Neurochem Res. 2012;37: 2456–2463. 10.1007/s11064-012-0805-2 [DOI] [PubMed] [Google Scholar]
- 36.Ransom CB, Ransom BR, Sontheimer H. Activity-dependent extracellular K+ accumulation in rat optic nerve: the role of glial and axonal Na+ pumps. J Physiol. 2000;522 Pt 3: 427–442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Schousboe A, Sickmann HM, Walls AB, Bak LK, Waagepetersen HS. Functional importance of the astrocytic glycogen-shunt and glycolysis for maintenance of an intact intra/extracellular glutamate gradient. Neurotox Res. 2010;18: 94–99. 10.1007/s12640-010-9171-5 [DOI] [PubMed] [Google Scholar]
- 38.Tesfaye N, Seaquist ER, Oz G. Noninvasive measurement of brain glycogen by nuclear magnetic resonance spectroscopy and its application to the study of brain metabolism. J Neurosci Res. 2011;89: 1905–1912. 10.1002/jnr.22703 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Walls AB, Sickmann HM, Brown A, Bouman SD, Ransom B, Schousboe A, et al. Characterization of 1,4-dideoxy-1,4-imino-d-arabinitol (DAB) as an inhibitor of brain glycogen shunt activity. J Neurochem. 2008;105: 1462–1470. 10.1111/j.1471-4159.2008.05250.x [DOI] [PubMed] [Google Scholar]
- 40.Xu J, Song D, Bai Q, Cai L, Hertz L, Peng L. Basic mechanism leading to stimulation of glycogenolysis by isoproterenol, EGF, elevated extracellular K+ concentrations, or GABA. Neurochem Res. 2014;39: 661–667. 10.1007/s11064-014-1244-z [DOI] [PubMed] [Google Scholar]
- 41.Swanson RA, Choi DW. Glial glycogen stores affect neuronal survival during glucose deprivation in vitro. J Cereb Blood Flow Metab Off J Int Soc Cereb Blood Flow Metab. 1993;13: 162–169. 10.1038/jcbfm.1993.19 [DOI] [PubMed] [Google Scholar]
- 42.Cloutier M, Bolger FB, Lowry JP, Wellstead P. An integrative dynamic model of brain energy metabolism using in vivo neurochemical measurements. J Comput Neurosci. 2009;27: 391–414. 10.1007/s10827-009-0152-8 [DOI] [PubMed] [Google Scholar]
- 43.Koivisto H, Leinonen H, Puurula M, Hafez HS, Barrera GA, Stridh MH, et al. Chronic Pyruvate Supplementation Increases Exploratory Activity and Brain Energy Reserves in Young and Middle-Aged Mice. Front Aging Neurosci. 2016;8: 41 10.3389/fnagi.2016.00041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Ransom BR. Glial modulation of neural excitability mediated by extracellular pH: a hypothesis revisited. Prog Brain Res. 2000;125: 217–228. 10.1016/S0079-6123(00)25012-7 [DOI] [PubMed] [Google Scholar]
- 45.Xu J, Song D, Xue Z, Gu L, Hertz L, Peng L. Requirement of glycogenolysis for uptake of increased extracellular K+ in astrocytes: potential implications for K+ homeostasis and glycogen usage in brain. Neurochem Res. 2013;38: 472–485. 10.1007/s11064-012-0938-3 [DOI] [PubMed] [Google Scholar]
- 46.Duran J, Saez I, Gruart A, Guinovart JJ, Delgado-García JM. Impairment in long-term memory formation and learning-dependent synaptic plasticity in mice lacking glycogen synthase in the brain. J Cereb Blood Flow Metab Off J Int Soc Cereb Blood Flow Metab. 2013;33: 550–556. 10.1038/jcbfm.2012.200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Gao V, Suzuki A, Magistretti PJ, Lengacher S, Pollonini G, Steinman MQ, et al. Astrocytic β2-adrenergic receptors mediate hippocampal long-term memory consolidation. Proc Natl Acad Sci U S A. 2016;113: 8526–8531. 10.1073/pnas.1605063113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Gibbs ME, Hutchinson D, Hertz L. Astrocytic involvement in learning and memory consolidation. Neurosci Biobehav Rev. 2008;32: 927–944. 10.1016/j.neubiorev.2008.02.001 [DOI] [PubMed] [Google Scholar]
- 49.Hertz L, Chen Y. Glycogenolysis, an astrocyte-specific reaction, is essential for both astrocytic and neuronal activities involved in learning. Neuroscience. 2017;370: 27–36. 10.1016/j.neuroscience.2017.06.025 [DOI] [PubMed] [Google Scholar]
- 50.Steinman MQ, Gao V, Alberini CM. The Role of Lactate-Mediated Metabolic Coupling between Astrocytes and Neurons in Long-Term Memory Formation. Front Integr Neurosci. 2016;10: 10 10.3389/fnint.2016.00010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Suzuki A, Stern SA, Bozdagi O, Huntley GW, Walker RH, Magistretti PJ, et al. Astrocyte-neuron lactate transport is required for long-term memory formation. Cell. 2011;144: 810–823. 10.1016/j.cell.2011.02.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Boury-Jamot B, Carrard A, Martin JL, Halfon O, Magistretti PJ, Boutrel B. Disrupting astrocyte-neuron lactate transfer persistently reduces conditioned responses to cocaine. Mol Psychiatry. 2015; 10.1038/mp.2015.157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Baud MO, Parafita J, Nguyen A, Magistretti PJ, Petit J-M. Sleep fragmentation alters brain energy metabolism without modifying hippocampal electrophysiological response to novelty exposure. J Sleep Res. 2016;25: 583–590. 10.1111/jsr.12419 [DOI] [PubMed] [Google Scholar]
- 54.Brunet JF, Allaman I, Magistretti PJ, Pellerin L. Glycogen metabolism as a marker of astrocyte differentiation. J Cereb Blood Flow Metab Off J Int Soc Cereb Blood Flow Metab. 2010;30: 51–55. 10.1038/jcbfm.2009.207 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Aston-Jones G, Rajkowski J, Kubiak P, Alexinsky T. Locus coeruleus neurons in monkey are selectively activated by attended cues in a vigilance task. J Neurosci Off J Soc Neurosci. 1994;14: 4467–4480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Song AH, Kucyi A, Napadow V, Brown EN, Loggia ML, Akeju O. Pharmacological Modulation of Noradrenergic Arousal Circuitry Disrupts Functional Connectivity of the Locus Ceruleus in Humans. J Neurosci Off J Soc Neurosci. 2017;37: 6938–6945. 10.1523/JNEUROSCI.0446-17.2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Aston-Jones G, Waterhouse B. Locus coeruleus: From global projection system to adaptive regulation of behavior. Brain Res. 2016;1645: 75–78. 10.1016/j.brainres.2016.03.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Lecas J-C. Locus coeruleus activation shortens synaptic drive while decreasing spike latency and jitter in sensorimotor cortex. Implications for neuronal integration. Eur J Neurosci. 2004;19: 2519–2530. 10.1111/j.0953-816X.2004.03341.x [DOI] [PubMed] [Google Scholar]
- 59.Atzori M, Cuevas-Olguin R, Esquivel-Rendon E, Garcia-Oscos F, Salgado-Delgado RC, Saderi N, et al. Locus Ceruleus Norepinephrine Release: A Central Regulator of CNS Spatio-Temporal Activation? Front Synaptic Neurosci. 2016;8: 25 10.3389/fnsyn.2016.00025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Fuxe K, Borroto-Escuela DO, Romero-Fernandez W, Diaz-Cabiale Z, Rivera A, Ferraro L, et al. Extrasynaptic neurotransmission in the modulation of brain function. Focus on the striatal neuronal-glial networks. Front Physiol. 2012;3: 136 10.3389/fphys.2012.00136 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Allaman I, Pellerin L, Magistretti PJ. Protein targeting to glycogen mRNA expression is stimulated by noradrenaline in mouse cortical astrocytes. Glia. 2000;30: 382–391. [PubMed] [Google Scholar]
- 62.Allaman I, Bélanger M, Magistretti PJ. Astrocyte-neuron metabolic relationships: for better and for worse. Trends Neurosci. 2011;34: 76–87. 10.1016/j.tins.2010.12.001 [DOI] [PubMed] [Google Scholar]
- 63.De Pitta M, Brunel N, Volterra A. Astrocytes: Orchestrating synaptic plasticity? Neuroscience. 2016;323: 43–61. 10.1016/j.neuroscience.2015.04.001 [DOI] [PubMed] [Google Scholar]
- 64.Hertz L, Xu J, Song D, Yan E, Gu L, Peng L. Astrocytic and neuronal accumulation of elevated extracellular K(+) with a 2/3 K(+)/Na(+) flux ratio-consequences for energy metabolism, osmolarity and higher brain function. Front Comput Neurosci. 2013;7: 114 10.3389/fncom.2013.00114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Magistretti PJ. Regulation of glycogenolysis by neurotransmitters in the central nervous system. Diabète Métabolisme. 1988;14: 237–246. [PubMed] [Google Scholar]
- 66.Quach TT, Rose C, Schwartz JC. [3H]Glycogen hydrolysis in brain slices: responses to neurotransmitters and modulation of noradrenaline receptors. J Neurochem. 1978;30: 1335–1341. [DOI] [PubMed] [Google Scholar]
- 67.Schorderet M, Hof P, Magistretti PJ. The effects of VIP on cyclic AMP and glycogen levels in vertebrate retina. Peptides. 1984;5: 295–298. [DOI] [PubMed] [Google Scholar]
- 68.Sorg O, Magistretti PJ. Characterization of the glycogenolysis elicited by vasoactive intestinal peptide, noradrenaline and adenosine in primary cultures of mouse cerebral cortical astrocytes. Brain Res. 1991;563: 227–233. [DOI] [PubMed] [Google Scholar]
- 69.Walls AB, Schousboe A. Brain glycogen: emergency fuel and dynamic function in neurotransmission. Metab Brain Dis. 2015;30: 249 10.1007/s11011-014-9619-z [DOI] [PubMed] [Google Scholar]
- 70.Xue X, Wang LR, Sato Y, Jiang Y, Berg M, Yang DS, et al. Single-walled carbon nanotubes alleviate autophagic/lysosomal defects in primary glia from a mouse model of Alzheimer’s disease. Nano Lett. 2014;14: 5110–7. 10.1021/nl501839q [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Sorg O, Magistretti PJ. Vasoactive intestinal peptide and noradrenaline exert long-term control on glycogen levels in astrocytes: blockade by protein synthesis inhibition. J Neurosci Off J Soc Neurosci. 1992;12: 4923–4931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Edwards C, Nahorski SR, Rogers KJ. In vivo changes of cerebral cyclic adenosine 3’,5’-monophosphate induced by biogenic amines: association with phosphorylase activation. J Neurochem. 1974;22: 565–572. [DOI] [PubMed] [Google Scholar]
- 73.Murphy S, Pearce B. Functional receptors for neurotransmitters on astroglial cells. Neuroscience. 1987;22: 381–394. [DOI] [PubMed] [Google Scholar]
- 74.De Keyser J, Zeinstra E, Wilczak N. Astrocytic β2-adrenergic receptors and multiple sclerosis. Neurobiol Dis. 2004;15: 331–339. 10.1016/j.nbd.2003.10.012 [DOI] [PubMed] [Google Scholar]
- 75.Cambray-Deakin M, Pearce B, Morrow C, Murphy S. Effects of extracellular potassium on glycogen stores of astrocytes in vitro. J Neurochem. 1988;51: 1846–1851. [DOI] [PubMed] [Google Scholar]
- 76.Cohen Z, Molinatti G, Hamel E. Astroglial and vascular interactions of noradrenaline terminals in the rat cerebral cortex. J Cereb Blood Flow Metab Off J Int Soc Cereb Blood Flow Metab. 1997;17: 894–904. 10.1097/00004647-199708000-00008 [DOI] [PubMed] [Google Scholar]
- 77.Gibbs ME, Hutchinson DS, Summers RJ. Noradrenaline release in the locus coeruleus modulates memory formation and consolidation; roles for α- and β-adrenergic receptors. Neuroscience. 2010;170: 1209–1222. 10.1016/j.neuroscience.2010.07.052 [DOI] [PubMed] [Google Scholar]
- 78.Hertz L, Lovatt D, Goldman SA, Nedergaard M. Adrenoceptors in brain: cellular gene expression and effects on astrocytic metabolism and [Ca(2+)]i. Neurochem Int. 2010;57: 411–420. 10.1016/j.neuint.2010.03.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.O’Dowd BS, Barrington J, Ng KT, Hertz E, Hertz L. Glycogenolytic response of primary chick and mouse cultures of astrocytes to noradrenaline across development. Brain Res Dev Brain Res. 1995;88: 220–223. [DOI] [PubMed] [Google Scholar]
- 80.Gibbs ME. Role of Glycogenolysis in Memory and Learning: Regulation by Noradrenaline, Serotonin and ATP. Front Integr Neurosci. 2015;9: 70 10.3389/fnint.2015.00070 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Madsen PL, Cruz NF, Sokoloff L, Dienel GA. Cerebral oxygen/glucose ratio is low during sensory stimulation and rises above normal during recovery: excess glucose consumption during stimulation is not accounted for by lactate efflux from or accumulation in brain tissue. J Cereb Blood Flow Metab Off J Int Soc Cereb Blood Flow Metab. 1999;19: 393–400. 10.1097/00004647-199904000-00005 [DOI] [PubMed] [Google Scholar]
- 82.Swanson RA. Physiologic coupling of glial glycogen metabolism to neuronal activity in brain. Can J Physiol Pharmacol. 1992;70 Suppl: S138–144. [DOI] [PubMed] [Google Scholar]
- 83.Matsui T, Omuro H, Liu Y-F, Soya M, Shima T, McEwen BS, et al. Astrocytic glycogen-derived lactate fuels the brain during exhaustive exercise to maintain endurance capacity. Proc Natl Acad Sci U S A. 2017;114: 6358–6363. 10.1073/pnas.1702739114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Pellerin L, Stolz M, Sorg O, Martin JL, Deschepper CF, Magistretti PJ. Regulation of energy metabolism by neurotransmitters in astrocytes in primary culture and in an immortalized cell line. Glia. 1997;21: 74–83. [DOI] [PubMed] [Google Scholar]
- 85.Alberini CM, Cruz E, Descalzi G, Bessières B, Gao V. Astrocyte glycogen and lactate: New insights into learning and memory mechanisms. Glia. 2017;66: 1244–1262. 10.1002/glia.23250 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Genc S, Kurnaz IA, Ozilgen M. Astrocyte-neuron lactate shuttle may boost more ATP supply to the neuron under hypoxic conditions—in silico study supported by in vitro expression data. BMC Syst Biol. 2011;5: 162 10.1186/1752-0509-5-162 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Jolivet R, Coggan JS, Allaman I, Magistretti PJ. Multi-timescale modeling of activity-dependent metabolic coupling in the neuron-glia-vasculature ensemble. PLoS Comput Biol. 2015;11: e1004036 10.1371/journal.pcbi.1004036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Mason S. Lactate Shuttles in Neuroenergetics-Homeostasis, Allostasis and Beyond. Front Neurosci. 2017;11: 43 10.3389/fnins.2017.00043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Pellerin L, Magistretti PJ. Sweet sixteen for ANLS. JCerebBlood Flow Metab. 2012;32: 1152–1166. 10.1038/jcbfm.2011.149 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Nuriya M, Takeuchi M, Yasui M. Background norepinephrine primes astrocytic calcium responses to subsequent norepinephrine stimuli in the cerebral cortex. Biochem Biophys Res Commun. 2017;483: 732–738. 10.1016/j.bbrc.2016.12.073 [DOI] [PubMed] [Google Scholar]
- 91.Xu K, Morgan KT, Todd Gehris A, Elston TC, Gomez SM. A whole-body model for glycogen regulation reveals a critical role for substrate cycling in maintaining blood glucose homeostasis. PLoS Comput Biol. 2011;7: e1002272 10.1371/journal.pcbi.1002272 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Calì C, Wawrzyniak M, Becker C, Maco B, Cantoni M, Jorstad A, et al. The effects of aging on neuropil structure in mouse somatosensory cortex—A 3D electron microscopy analysis of layer 1. PLOS ONE. 2018;13: e0198131 10.1371/journal.pone.0198131 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Cali C, Wawrzyniak M, Becker C, Maco B, Cantoni M, Jorstad A, et al. Data from: The effects of aging on neuropil structure in mouse somatosensory cortex—A 3D electron microscopy analysis of layer 1. 2018; 10.5061/dryad.bh78sn5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Hertz L, Xu J, Song D, Du T, Li B, Yan E, et al. Astrocytic glycogenolysis: mechanisms and functions. Metab Brain Dis. 2015;30: 317–333. 10.1007/s11011-014-9536-1 [DOI] [PubMed] [Google Scholar]
- 95.Barros LF. Metabolic signaling by lactate in the brain. Trends Neurosci. 2013;36: 396–404. 10.1016/j.tins.2013.04.002 [DOI] [PubMed] [Google Scholar]
- 96.Bouzier-Sore A-K, Voisin P, Canioni P, Magistretti PJ, Pellerin L. Lactate is a preferential oxidative energy substrate over glucose for neurons in culture. J Cereb Blood Flow Metab Off J Int Soc Cereb Blood Flow Metab. 2003;23: 1298–1306. 10.1097/01.WCB.0000091761.61714.25 [DOI] [PubMed] [Google Scholar]
- 97.Magistretti PJ, Pellerin L. Regulation by neurotransmitters of glial energy metabolism. Adv Exp Med Biol. 1997;429: 137–143. [DOI] [PubMed] [Google Scholar]
- 98.Cataldo AM, Broadwell RD. Cytochemical identification of cerebral glycogen and glucose-6-phosphatase activity under normal and experimental conditions. II. Choroid plexus and ependymal epithelia, endothelia and pericytes. J Neurocytol. 1986;15: 511–524. [DOI] [PubMed] [Google Scholar]
- 99.Shimizu F, Sano Y, Tominaga O, Maeda T, Abe M, Kanda T. Advanced glycation end-products disrupt the blood-brain barrier by stimulating the release of transforming growth factor-β by pericytes and vascular endothelial growth factor and matrix metalloproteinase-2 by endothelial cells in vitro. Neurobiol Aging. 2013;34: 1902–1912. 10.1016/j.neurobiolaging.2013.01.012 [DOI] [PubMed] [Google Scholar]
- 100.Ransom BR, Fern R. Does astrocytic glycogen benefit axon function and survival in CNS white matter during glucose deprivation? Glia. 1997;21: 134–141. [PubMed] [Google Scholar]
- 101.Wender R, Brown AM, Fern R, Swanson RA, Farrell K, Ransom BR. Astrocytic glycogen influences axon function and survival during glucose deprivation in central white matter. J Neurosci Off J Soc Neurosci. 2000;20: 6804–6810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Magistretti PJ. Neuron-glia metabolic coupling and plasticity. Exp Physiol. 2011;96: 407–410. 10.1113/expphysiol.2010.053157 [DOI] [PubMed] [Google Scholar]
- 103.Sickmann HM, Walls AB, Schousboe A, Bouman SD, Waagepetersen HS. Functional significance of brain glycogen in sustaining glutamatergic neurotransmission. J Neurochem. 2009;109 Suppl 1: 80–86. 10.1111/j.1471-4159.2009.05915.x [DOI] [PubMed] [Google Scholar]
- 104.Dringen R, Hamprecht B. Differences in glycogen metabolism in astroglia-rich primary cultures and sorbitol-selected astroglial cultures derived from mouse brain. Glia. 1993;8: 143–149. 10.1002/glia.440080302 [DOI] [PubMed] [Google Scholar]
- 105.Brown AM, Tekkök SB, Ransom BR. Glycogen regulation and functional role in mouse white matter. J Physiol. 2003;549: 501–512. 10.1113/jphysiol.2003.042416 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Cambray-Deakin M, Pearce B, Morrow C, Murphy S. Effects of neurotransmitters on astrocyte glycogen stores in vitro. J Neurochem. 1988;51: 1852–1857. [DOI] [PubMed] [Google Scholar]
- 107.Magistretti PJ, Hof P, Schorderet M. The increase in cyclic-AMP levels elicited by vasoactive intestinal peptide (VIP) in mouse cerebral cortical slices is potentiated by ergot alkaloids. Neurochem Int. 1984;6: 751–753. [DOI] [PubMed] [Google Scholar]
- 108.Magistretti PJ, Sorg O, Yu N, Martin JL, Pellerin L. Neurotransmitters regulate energy metabolism in astrocytes: implications for the metabolic trafficking between neural cells. Dev Neurosci. 1993;15: 306–312. 10.1159/000111349 [DOI] [PubMed] [Google Scholar]
- 109.Zhao Y, Zhang Q, Shao X, Ouyang L, Wang X, Zhu K, et al. Decreased Glycogen Content Might Contribute to Chronic Stress-Induced Atrophy of Hippocampal Astrocyte volume and Depression-like Behavior in Rats. Sci Rep. 2017;7: 43192 10.1038/srep43192 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Idris I, Thomson GA, Sharma JC. Diabetes mellitus and stroke. Int J Clin Pract. 2006;60: 48–56. 10.1111/j.1368-5031.2006.00682.x [DOI] [PubMed] [Google Scholar]
- 111.Barros LF, Deitmer JW. Glucose and lactate supply to the synapse. Brain Res Rev. 2010;63: 149–159. 10.1016/j.brainresrev.2009.10.002 [DOI] [PubMed] [Google Scholar]
- 112.Bélanger M, Allaman I, Magistretti PJ. Brain energy metabolism: focus on astrocyte-neuron metabolic cooperation. Cell Metab. 2011;14: 724–738. 10.1016/j.cmet.2011.08.016 [DOI] [PubMed] [Google Scholar]
- 113.Sotelo-Hitschfeld T, Niemeyer MI, Mächler P, Ruminot I, Lerchundi R, Wyss MT, et al. Channel-mediated lactate release by K+-stimulated astrocytes. J Neurosci Off J Soc Neurosci. 2015;35: 4168–4178. 10.1523/JNEUROSCI.5036-14.2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Tanaka M, Nakamura F, Mizokawa S, Matsumura A, Matsumura K, Murata T, et al. Role of lactate in the brain energy metabolism: revealed by Bioradiography. Neurosci Res. 2004;48: 13–20. [DOI] [PubMed] [Google Scholar]
- 115.Shulman RG, Rothman DL. 13 C NMR of intermediary metabolism: implications for systemic physiology. AnnuRevPhysiol. 2001;63: 15–48. [DOI] [PubMed] [Google Scholar]
- 116.Brown AM, Ransom BR. Astrocyte glycogen and brain energy metabolism. Glia. 2007;55: 1263–1271. 10.1002/glia.20557 [DOI] [PubMed] [Google Scholar]
- 117.Morgenthaler FD, Koski DM, Kraftsik R, Henry P-G, Gruetter R. Biochemical quantification of total brain glycogen concentration in rats under different glycemic states. Neurochem Int. 2006;48: 616–622. 10.1016/j.neuint.2005.12.034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Dringen R, Gebhardt R, Hamprecht B. Glycogen in astrocytes: possible function as lactate supply for neighboring cells. Brain Res. 1993;623: 208–214. [DOI] [PubMed] [Google Scholar]
- 119.Sibson NR, Dhankhar A, Mason GF, Rothman DL, Behar KL, Shulman RG. Stoichiometric coupling of brain glucose metabolism and glutamatergic neuronal activity. Proc Natl Acad Sci U S A. 1998;95: 316–321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Jouaville LS, Pinton P, Bastianutto C, Rutter GA, Rizzuto R. Regulation of mitochondrial ATP synthesis by calcium: Evidence for a long-term metabolic priming. Proc Natl Acad Sci. 1999;96: 13807–13812. 10.1073/pnas.96.24.13807 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Kasischke KA, Vishwasrao HD, Fisher PJ, Zipfel WR, Webb WW. Neural activity triggers neuronal oxidative metabolism followed by astrocytic glycolysis. Science. 2004;305: 99–103. 10.1126/science.1096485 [DOI] [PubMed] [Google Scholar]
- 122.Kilic K, Karatas H, Donmez-Demir B, Eren-Kocak E, Gursoy-Ozdemir Y, Can A, et al. Inadequate Brain Glycogen or Sleep Increases Spreading Depression Susceptibility. Ann Neurol. 2017;83: 61–73. 10.1002/ana.25122 [DOI] [PubMed] [Google Scholar]
- 123.Aubert A, Pellerin L, Magistretti PJ, Costalat R. A coherent neurobiological framework for functional neuroimaging provided by a model integrating compartmentalized energy metabolism. Proc Natl Acad Sci U S A. 2007;104: 4188–4193. 10.1073/pnas.0605864104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Chen Y, Hertz L. Noradrenaline effects on pyruvate decarboxylation: correlation with calcium signaling. J Neurosci Res. 1999;58: 599–606. [DOI] [PubMed] [Google Scholar]
- 125.Jolivet R, Magistretti PJ, Weber B. Deciphering neuron-glia compartmentalization in cortical energy metabolism. Front Neuroenergetics. 2009;1: 4 10.3389/neuro.14.004.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Aubert A, Costalat R, Magistretti PJ, Pellerin L. Brain lactate kinetics: Modeling evidence for neuronal lactate uptake upon activation. Proc Natl Acad Sci U S A. 2005;102: 16448–16453. 10.1073/pnas.0505427102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Machler P, Wyss MT, Elsayed M, Stobart J, Gutierrez R, von Faber-Castell A, et al. In Vivo Evidence for a Lactate Gradient from Astrocytes to Neurons. Cell Metab. 2016;23: 94–102. 10.1016/j.cmet.2015.10.010 [DOI] [PubMed] [Google Scholar]
- 128.Tang F, Lane S, Korsak A, Paton JFR, Gourine AV, Kasparov S, et al. Lactate-mediated glia-neuronal signalling in the mammalian brain. Nat Commun. 2014;5: 3284 10.1038/ncomms4284 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Jensen CJ, Demol F, Bauwens R, Kooijman R, Massie A, Villers A, et al. Astrocytic β2 Adrenergic Receptor Gene Deletion Affects Memory in Aged Mice. PloS One. 2016;11: e0164721 10.1371/journal.pone.0164721 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Boury-Jamot B, Carrard A, Martin JL, Halfon O, Magistretti PJ, Boutrel B. Disrupting astrocyte-neuron lactate transfer persistently reduces conditioned responses to cocaine. Mol Psychiatry. 2016;21: 1070–6. 10.1038/mp.2015.157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Fuxe K, Borroto-Escuela DO, Romero-Fernandez W, Zhang W-B, Agnati LF. Volume transmission and its different forms in the central nervous system. Chin J Integr Med. 2013;19: 323–329. 10.1007/s11655-013-1455-1 [DOI] [PubMed] [Google Scholar]
- 132.Zoli M, Agnati LF. Wiring and volume transmission in the central nervous system: the concept of closed and open synapses. Prog Neurobiol. 1996;49: 363–380. [DOI] [PubMed] [Google Scholar]
- 133.Dong J, Chen X, Cui M, Yu X, Pang Q, Sun J. β2-adrenergic receptor and astrocyte glucose metabolism. J Mol Neurosci MN. 2012;48: 456–463. 10.1007/s12031-012-9742-4 [DOI] [PubMed] [Google Scholar]
- 134.Bender CL, Calfa GD, Molina VA. Astrocyte plasticity induced by emotional stress: A new partner in psychiatric physiopathology? Prog Neuropsychopharmacol Biol Psychiatry. 2015;65: 68–77. 10.1016/j.pnpbp.2015.08.005 [DOI] [PubMed] [Google Scholar]
- 135.Carrard A, Elsayed M, Margineanu M, Boury-Jamot B, Fragnière L, Meylan EM, et al. Peripheral administration of lactate produces antidepressant-like effects. Mol Psychiatry. 2016; 10.1038/mp.2016.179 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Russell VA, Oades RD, Tannock R, Killeen PR, Auerbach JG, Johansen EB, et al. Response variability in Attention-Deficit/Hyperactivity Disorder: a neuronal and glial energetics hypothesis. BehavBrain Funct. 2006;2: 30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Todd RD, Botteron KN. Is attention-deficit/hyperactivity disorder an energy deficiency syndrome? Biol Psychiatry. 2001;50: 151–158. [DOI] [PubMed] [Google Scholar]
- 138.Verkhratsky A, Zorec R, Rodríguez JJ, Parpura V. Astroglia dynamics in ageing and Alzheimer’s disease. Curr Opin Pharmacol. 2016;26: 74–79. 10.1016/j.coph.2015.09.011 [DOI] [PubMed] [Google Scholar]
- 139.Seidel JL, Shuttleworth CW. Contribution of astrocyte glycogen stores to progression of spreading depression and related events in hippocampal slices. Neuroscience. 2011;192: 295–303. 10.1016/j.neuroscience.2011.05.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Straehle CN, Köthe U, Knott G, Hamprecht FA. Carving: scalable interactive segmentation of neural volume electron microscopy images. Med Image Comput Comput-Assist Interv MICCAI Int Conf Med Image Comput Comput-Assist Interv. 2011;14: 653–660. [DOI] [PubMed]
- 141.Hines ML, Carnevale NT. NEURON: a tool for neuroscientists. Neurosci Rev J Bringing Neurobiol Neurol Psychiatry. 2001;7: 123–135. [DOI] [PubMed] [Google Scholar]
- 142.Boras BW, Kornev A, Taylor SS, McCulloch AD. Using Markov state models to develop a mechanistic understanding of protein kinase A regulatory subunit RIα activation in response to cAMP binding. J Biol Chem. 2014;289: 30040–30051. 10.1074/jbc.M114.568907 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Conti M, Mika D, Richter W. Cyclic AMP compartments and signaling specificity: Role of cyclic nucleotide phosphodiesterases. J Gen Physiol. 2014;143: 29–38. 10.1085/jgp.201311083 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Meunier H, Labrie F. Specificity of the beta 2-adrenergic receptor stimulating cyclic AMP accumulation in the intermediate lobe of rat pituitary gland. Eur J Pharmacol. 1982;81: 411–420. [DOI] [PubMed] [Google Scholar]
- 145.Jakobsen E, Bak LK, Walls AB, Reuschlein A-K, Schousboe A, Waagepetersen HS. Glycogen Shunt Activity and Glycolytic Supercompensation in Astrocytes May Be Distinctly Mediated via the Muscle Form of Glycogen Phosphorylase. Neurochem Res. 2017;42: 2490–2494. 10.1007/s11064-017-2267-z [DOI] [PubMed] [Google Scholar]
- 146.Newgard CB, Littman DR, van Genderen C, Smith M, Fletterick RJ. Human brain glycogen phosphorylase. Cloning, sequence analysis, chromosomal mapping, tissue expression, and comparison with the human liver and muscle isozymes. J Biol Chem. 1988;263: 3850–3857. [PubMed] [Google Scholar]
- 147.Mutalik VK, Venkatesh KV. Quantification of the glycogen cascade system: the ultrasensitive responses of liver glycogen synthase and muscle phosphorylase are due to distinctive regulatory designs. Theor Biol Med Model. 2005;2: 19 10.1186/1742-4682-2-19 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
(DOCX)
(DOCX)
(DOCX)
(DOCX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.