

Socially monogamous songbird couples show off their courtship display in front of other individuals.
Abstract
Social environments can shape animal communication. Although mutual courtship displays are generally thought to function in private communication between a male and a female, we provide experimental evidence that they work in a broader social context than previously thought. We examined the audience effect on mutual courtship in blue-capped cordon-bleus, a socially monogamous songbird. This species is characterized by conspicuous courtship shared between sexes: Both sexes sing songs and sometimes add a unique dance display that looks like human tap dancing. We found that in both sexes, multimodal courtship displays (song accompanied by dance) were promoted in the presence of an audience, especially if it was the opposite sex. In contrast, unimodal displays (song without dance) were suppressed by audiences. Because birds directed the courtship dancing toward their partners (but not the audience), multimodal courtship displays are likely meant to advertise their current mating status to other cordon-bleus.
INTRODUCTION
Although courtship displays are often regarded as dyadic interactions between performers and their potential mates, they can be influenced by other individuals. This phenomenon is called the audience effect (1). Performing courtship display in public provides information such as the mating preference and quality of the performers to bystanding or eavesdropping individuals. It can alter a risk of the prospective mate being usurped or a chance of extra-pair copulation. Thus, animals should adjust their courting behavior depending on who is around. For instance, male Atlantic mollies (Poecilia mexicana) suppress sexual behavior toward preferred females when other males are present to lead potential rivals away from the preferred female (2). Similarly, to avoid sexual conflicts with their mates, male canaries (Serinus canaria) decrease their extra-pair courtships toward unfamiliar females when their own mates are present (3).
Past studies have primarily focused on audience effects on male sexual behavior [for example, insects (4), fish (2, 5), mammals (6), and birds (3, 7)], with few examining how female sexual behavior is affected. Therefore, here, we sought to examine the functions of both male and female courtship within a social environment. In some socially monogamous avian species, both sexes engage in courtship displays, such as song duetting (8), and multimodal courtship displays [for example, magpie-larks (9), albatrosses (10), cranes (11, 12), and grebes (13)]. These types of mutual courtship displays are assumed to play an important role in pair bonding and its maintenance (14, 15), and thus have the potential to convey information related to mating status, which could be useful for mating rivals or individuals who seek opportunities for extra-pair mating. However, few studies have quantitatively examined whether these multimodal courtship displays change depending on the presence of others.
Songbirds in the family Estrildidae are socially monogamous with long-term pair bonding and are characterized by multiple sexual traits (that is, courtship dance and song as well as plumage ornamentation) that are often shared between sexes (16, 17). Among Estrildids, the blue-capped cordon-bleu (Uraeginthus cyanocephalus) is known for especially conspicuous courtship displays in both sexes (16). In addition to song, male and female cordon-bleus “tap dance” by rapidly stamping their feet against their perch several times while hopping (18). This tap dance–like display appears to add visual and vibratory signals to the song (19). We have found that this species is characterized by slight sexual dimorphisms in song traits (20) and dance performance (18). Estrildids are known to usually perform multimodal displays (that is, song with dance) when they are close to potential mating partners, and to perform unimodal displays (that is, song without dance) when they are alone (21, 22). Presumably, multimodal displays convey courtship-target information, as they often include angled-tail posture through which courting individuals “point” at the target individual with their tails (fig. S1 and movie S1), usually throughout the entire courtship display (16). These characteristics led us to predict that cordon-bleus use multimodal courtship to advertise their mating status to others. Testing this idea can give us a clue as to why cordon-bleus evolved to perform these complex multimodal courtship displays in both sexes.
To clarify the function of mutual multimodal courtship displays, we examined how audiences affect cordon-bleu courtship displays. Specifically, we placed paired couples in no-audience and audience conditions and observed their behavior (fig. S2). We expected that audiences could either promote or suppress courtship displays, and the functions of the behavioral changes would vary depending on the sex of the audience (matched or opposite that of a courtship performer; Fig. 1). When multimodal courtship display toward partners is promoted by the presence of an audience (Fig. 1A), it can be understood as an advertisement of mating status. In addition to signaling commitment toward a partner (Fig. 1A, [1a]), courtship displays when the audience is the opposite sex can also function as an implicit appeal for future extra-pair mating with the audience (Fig. 1A, [1b]; discussed later). Courtship displays when the audience is the same sex can function as mate guarding (Fig. 1A, [2]). Conversely, when courtship display is suppressed by the presence of audience (Fig. 1B), it can be seen as an effort to conceal the relationship with a particular individual, which can increase access to future mates [Fig. 1B, [3]; see (3)] or decrease intrasexual competition [Fig. 1B, [4]; see (6)]. We also expected that the direction of the audience effects would differ depending on the sex of the performer because the reproductive benefits of advertising or concealing relationships can differ for male and female birds.
Fig. 1. The hypothesized effects that audience could have on courtship display and what they would signify.
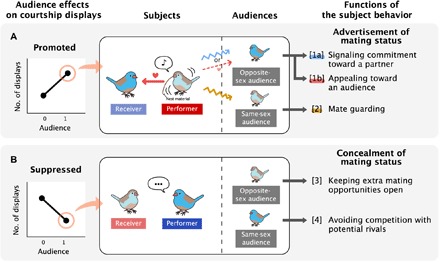
(A) If the cordon-bleu multimodal display is promoted by the presence of audiences, then the display would function as an advertisement for the pair’s relationship. Multimodal displays when the audience is the opposite sex of the performer could indicate commitment toward partners [1a] or an appeal toward the audience for future extra-pair mating [1b]. When the audience is the same sex as the performer, the display could function to guard mates from potential rivals [2]. (B) If the cordon-bleu multimodal display is suppressed by audiences, then the lack of display would act to conceal the mating status and help keep future mating opportunities with potential mates [3] and avoid competition with potential rivals [4].
RESULTS
Multimodal/unimodal displays under no-audience and audience conditions
The presence of an audience significantly affected the frequency of both multimodal and unimodal displays, but in opposite directions (Fig. 2 and Table 1). Compared with pre- and post-experimental controls (fig. S2), both males and females showed more multimodal displays but fewer unimodal displays under audience conditions (Fig. 2 and Table 1). Subject bird sex affected the number of multimodal displays but not unimodal displays (Fig. 2 and Table 1); males produced more multimodal displays than females, which was consistent with the following results in Table 2.
Fig. 2. Effects of audience presence and sex on the number of courtship displays.
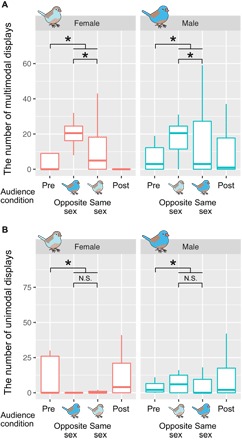
Audience had influences on both (A) multimodal and (B) unimodal displays. Only the data of subjects that performed courtship displays at least once throughout the experiment are plotted (see also Results). All box plots show medians and quartiles. *P < 0.05. N.S., not significant (P > 0.05).
Table 1. Effects of audience presence and subject sex on the number of courtship displays (GLMM, Poisson).
|
Response variable |
Independent variable |
Comparison between pre-experimental control and audience conditions |
Comparison between post-experimental control and audience conditions |
||||||
| Coefficient | SE | z | P | Coefficient | SE | z | P | ||
| The number of multimodal displays |
The presence of audience |
0.733 | 0.096 | 7.673 | <0.001 | 0.945 | 0.104 | 9.069 | <0.001 |
| Subject sex (male or female)* |
4.469 | 1.494 | 2.992 | 0.003 | 4.488 | 1.477 | 3.038 | 0.002 | |
| The number of unimodal displays |
The presence of audience |
−0.440 | 0.093 | −4.729 | <0.001 | −0.716 | 0.087 | −8.269 | <0.001 |
| Subject sex (male or female)* |
1.628 | 1.761 | 0.924 | 0.355 | 0.326 | 1.447 | 0.226 | 0.822 | |
*Estimated values for effects that contain a “subject sex” term are for males.
Table 2. Factors that influence the number of courtship displays under audience conditions (GLMM, Poisson).
| Response variable | Independent variable | Coefficient | SE | z | P |
| The number of multimodal displays |
Audience sex (same or opposite sex)* |
−0.413 | 0.088 | −4.704 | <0.001 |
| The number of audience unimodal displays |
−0.004 | 0.002 | −2.737 | 0.006 | |
| Present order of audience (first or second) |
−0.160 | 0.090 | −1.778 | 0.075 | |
| Subject sex (male or female)† |
4.100 | 1.422 | 2.884 | 0.004 | |
| The number of unimodal displays |
Audience sex (same or opposite sex)* |
−0.178 | 0.158 | −1.124 | 0.261 |
| The number of audience unimodal displays |
−0.009 | 0.002 | −5.000 | <0.001 | |
| Present order of audience (first or second) |
−0.766 | 0.165 | −4.628 | <0.001 | |
| Subject sex (male or female)† |
1.156 | 1.780 | 0.649 | 0.516 |
*Estimated values for effects that contained an “audience sex” term are for same-sex audience conditions.
†Estimated values for effects that contain a “subject sex” term are for males.
Two pairs did not show any multimodal display in any of the four conditions. In the other 18 pairs, at least one bird within a pair performed at least one multimodal display across the four experimental conditions (fig. S3). In any given pair, we usually observed either male-to-female or female-to-male courtship display. Only one pair performed duet dance–like behavior that was determined to be a simultaneous expression of dance display [see (23)]. In two other pairs, both males and females performed multimodal displays, but at different times (fig. S3). We did not observe any song-duetting behavior during the course of the experiment.
Multimodal/unimodal displays depended on audience sex
When the sex of an individual subject did not match the sex of the audience, it performed significantly more multimodal displays but not unimodal displays (Fig. 2 and Table 2). In addition, we found that audience singing suppressed both multimodal and unimodal displays (Table 2). The order of audience conditions (that is, first or second audience conditions; fig. S1) also negatively affected the number of multimodal and unimodal displays (Table 2). This indicates that courting rates were affected by habituation (Table 2).
Who are the multimodal displays for? Dance-bout duration and tail angling
Birds danced significantly longer when their partners stayed on the same perch (movie S1), and courtship performers angled their tails toward their partners (table S1, fig. S1, and movie S1). This was not affected by audience position (Table 3). Thus, despite the presence of an audience, the displays themselves were always directed toward the partner, not the audience. We found no sex difference in dance-bout duration (Table 3).
Table 3. Effects of partner and audience position on dance-bout duration (GLMM, Gaussian).
Positions of paired birds and audiences were classified as binomial variables (see Materials and Methods).
| Response variable | Independent variable | Coefficient | SE | t | P |
| Dance-bout duration | Partner position | 5.558 | 2.023 | 2.747 | 0.006 |
| Audience position | 0.536 | 1.495 | 0.358 | 0.720 | |
| Subject sex (male or female)* |
−1.370 | 4.139 | −0.331 | 0.744 |
*Estimated values for effects that contain a “subject sex” term are for males.
DISCUSSION
Although courtship displays are generally assumed to represent private communication between a performer and their partner, this study revealed that both male and female courtship displays are adjusted depending on broader social environment. In both males and females, multimodal displays were promoted in same- and opposite-sex audience conditions, while unimodal displays were suppressed (Fig. 2). These results suggest that multimodal and unimodal displays have different roles.
Increased multimodal display under audience conditions indicates that cordon-bleus perform these displays to advertise their closed mating status to surrounding individuals (Fig. 1A). Multimodal courtship displays generally serve as more appealing signal than unimodal displays (24). Considering that birds increased the number of multimodal displays when the audience was the opposite sex (Fig. 2A), the multimodal display likely functions as signaling commitment toward partners (Fig. 1A). We observed that performers adjusted dance duration in response to partners but not the audience (Table 3). In addition, gestures such as angled tails during multimodal display can provide more information about who is being courted than can song alone (fig. S1, table S1, and movie S1). These results suggest that the courtship was mostly targeted toward partners. The partners also often angled their tails toward courtship performers (fig. S1 and movie S1). The multimodal courtship display in audience conditions can be interpreted as a way of showing off within-pair communication. In contrast, the results from past studies of audience effects indicate that males generally tried to conceal mating status to avoid sexual conflicts and maximize their future mating success (3), or show false preference so that rival males did not try to court their preferred females (2).
Although the multimodal displays are gesturally directed toward the paired partner but not to audience, the possibility remains that they also function to self-advertise one’s attractiveness toward opposite-sex audiences (Fig. 1A, [1b]) and contribute to future extra-pair mating. Particularly when mate-choice copying exists, advertising mating status is favorable because it provides males with more chances of extra-pair paternity and females with more options for genetic fathers of their offspring. Extra-pair mating is a common phenomenon in socially monogamous birds (25). As far as we know, there is no information about cordon-bleu extra-pair mating, but it likely occurs similar to other Estrildid finches (26). The present short-term experiments using randomly paired birds simulated the situation of intersexual communication during the pair-formation phase. However, we did not look at long-term consequences of courtship display, such as pair-bonding or extra-pair mating, which should be addressed in future research.
Suppression of unimodal displays in audience conditions might help to avoid conflicts and unnecessary competition, as birdsong often serves as an aggressive signal (27). This is supported by the fact that audience unimodal display suppressed subject courtship displays (Table 2).
Furthermore, our results suggest that female cordon-bleu courtship displays are as sensitive to social environments as male displays, and that the displays have similar functions for males and females. Although males produced more multimodal displays than females throughout the experiment, the multimodal displays were increased similarly in male and female birds in the audience condition, thus suggesting that multimodal displays in cordon-bleus evolved through mutual sexual selection. This was actually rather surprising because song function is not necessarily shared between sexes in songbirds (28, 29). Although previous songbird studies have shown that the presence of conspecific individuals alters female behaviors such as mate choice (30) and contact behavior with her partner (31), as far as we know, ours is the first experimental study to show audience effects on female multimodal courtship display and to compare them with the effects that audiences have on male displays.
As our analysis of male and female courtship display in cordon-bleus shows, in addition to sexual selection, the complexity and diversity of courtship communication in animals can be subject to social factors. To date, most courtship-display research has been conducted under the premise that complex multimodal courtship displays have evolved in males through female mate choice (32) [for example, spiders (33), frogs (34), fishes (35), and birds (36, 37)]. Therefore, much less is known about the factors responsible for the evolution of complex female displays. Apparently, adding visual information to songs evolved in conjunction with pair communication in the social life of cordon-bleus (16). We propose that pair bonding and group living are two main factors that promoted the evolution of both male and female multimodal display. This idea might be applicable to courtship displays in other socially monogamous birds, for example, the mutual courtship dance of Laysan albatrosses (Diomedea immutabilis) in colony (10) and the group courtship dance of male and female greater flamingos (Phoenicopterus roseus) (38).
MATERIALS AND METHODS
Experimental design
We arbitrarily matched 20 pairs of males and females using 18 male and 11 female adult blue-capped cordon-bleus (>6 months old). Because of the limited number of subjects, two males and five females were used as subjects twice, and two females were used as subjects three times in the experiment. Pseudoreplication due to multiple use of the same subjects was statistically dealt with using random-effect models (see below). Subjects were also used as audiences because the number of birds was limited.
Paired birds experienced four conditions: pre-experimental control (no-audience 1), male audience, female audience, and post-experimental control (no-audience 2; fig. S2). Each pair was introduced into an experimental cage (60 cm × 48 cm × 48 cm) 1 day before pre-experimental control conditions for habituation and then observed during the pre-experimental control. The next day, we introduced the audience bird (male or female) into a cage (60 cm × 48 cm × 48 cm) adjacent to the subject pair’s cage. Next, we repeated the audience condition with the opposite sex. The two cages were separated by a fine wire mesh partition; therefore, they could interact visually and auditorily, but not physically (fig. S2 and movie S1). The order of audience sex was randomized for each pair. For the first audience condition, 12 pairs were presented with male audiences and 8 with female audiences. On the last day of the experiment, pairs were observed without an audience for a post-experimental control. Fifteen pairs experienced the four conditions on four different days, and five pairs experienced male and female audience conditions on the same day. We confirmed that the results did not change even after eliminating the data from these five pairs (table S2).
We recorded the behavior of subject pairs and audience for 2 hours under each condition with a normal-speed camera (Q3HD, Zoom; National Television System Committee format, 30 frames/s). The camera was placed close to the wall of the audience cage (see movie S1). The two cages for the subject pair and audience were placed in a chamber (170 cm × 60 cm × 58 cm, lined with soundproof foam) to minimize disturbance. Before and after experiencing all four conditions, subject birds were kept in same-sex home cages. Similarly, audience birds were housed together in a soundproof chamber except for when they took part in the experiment. Two nests and abundant nesting materials were always available in the subject cages. To minimize the effect of courting activity of the audience, we did not provide nest materials in the audience cage since cordon-bleus generally started courtship by picking up nest materials (16, 18). Birds were provided with a finch seed mixture, cuttlebone, water, and cucumber ad libitum. All individuals were kept on a 14-hour light/10-hour dark schedule (lights on 07:00 to 21:00) at approximately 22°C and 50% humidity. Procedures were in accordance with national laws and approved by Institutional Animal Care and Use Committee guidelines and the government of Upper Bavaria (approval number 311.5-5682.1/1-2014-021).
Behavioral measurements
As measures of courting activity, we quantified the number of multimodal and unimodal displays under each condition. Unimodal displays comprised only singing behavior, and multimodal displays comprised singing behavior that was accompanied by dance. For each of the four conditions, we also recorded whether the dance and song displays were solos or duets. Dance/song duetting was defined as simultaneous dancing/singing by both members of a pair (8, 23).
To assess the effects of any audience courting activity on subject-pair behavior, we counted the number of unimodal displays produced by the audiences. We did not take into account multimodal displays from the audiences because they were not expected to dance without nest materials (see subjects and experimental procedure). However, two male audience birds did dance during the experiment. Although these unexpected audience displays could have affected the number of subject displays, we confirmed that the results remained unchanged even after removing these data (table S3).
To detect for whom multimodal displays were performed, we measured subject dance-bout duration under audience conditions and checked the positions of the paired birds and audience at that time. Positions of paired birds were scored as 1 if they were on the same perch as the dancer and 0 if they were on another perch. Audience position was scored as 1 if they were on the partition or the perch nearest to the partition (see movie S1) without singing and sleeping and 0 if they were on another perch or the floor. We also assessed whether the subjects of a pair angled their tails toward their partners during courtship when on the same perch (fig. S1).
Statistical analysis
To test whether audience presence affected courtship displays, we compared the number of multimodal and unimodal displays between no-audience and audience conditions, and between subject males and subject females. We did not pool first and second no-audience conditions to avoid erroneously detecting significant differences between audience and no-audience conditions. This was because we found that the number of multimodal displays in the second no-audience condition was substantially lower than in the first no-audience condition (see Fig. 2), which was presumably because of habituation. Thus, we did not combine the two conditions because doing so would underestimate the real number of displays when no audience was present.
For the audience conditions, we recorded the following data for analysis to determine the factors responsible for courtship activity: For each focal bird, we noted (i) whether the audience was the same or opposite sex, (ii) how often the audience sang unimodal displays, (iii) whether the audience was the first or second audience condition, and (iv) the sex of the focal bird. In addition, we tested whether the position of partners and audiences affected dance-bout duration.
For these analyses, we used a generalized linear mixed-effect model (GLMM). The numbers of multimodal and unimodal displays were analyzed with a Poisson distribution. Dance-bout duration was analyzed with a Gaussian distribution. In all analyses, we considered pair ID nested within bird ID as random effects to control for non-independence of data. All statistical analyses were performed using R 3.3.0 (39).
Supplementary Material
Acknowledgments
We thank the Max Planck Institute for Ornithology staff for providing the experimental equipment and maintaining the birds, especially A. Leitão for help with our experiments. We also thank A. Phillips from Edanz Group (www.edanzediting.com/ac) for editing a draft of this manuscript. Funding: We appreciate the financial support from the Japan Society for Promotion of Science (JSPS). Our study was supported by JSPS Grants-in-Aid for Young Scientists (grant numbers 22800002 and 23680027) to M.S. and a Grant-in-Aid for JSPS Fellows (grant number 15J02516) to N.O. Author contributions: All authors designed the experiments. N.O. carried out the experiments. N.O. and M.S. analyzed the data. All authors discussed the results and contributed to the writing of the manuscript. Competing interests: The authors declare that they have no competing interests. Data and materials availability: All data needed to evaluate the conclusions in the paper are present in the paper and/or the Supplementary Materials. Additional data related to this paper may be requested from N.O.
SUPPLEMENTARY MATERIALS
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/4/10/eaat4779/DC1
Fig. S1. Examples of tail direction during a courtship display in which a male is courting a female.
Fig. S2. Protocol design and schematic views of the apparatus used in this experiment.
Fig. S3. Cumulative number of pairs that performed duet or solo multimodal displays in the four experimental conditions.
Table S1. The list of subject birds that performed multimodal displays when their partner stayed nearby and the percentage of the dance bouts with tail angling.
Table S2. Results of the same analysis as in Table 2 after eliminating the data from the five pairs whose two audience conditions were conducted on the same day.
Table S3. Results of the same analysis as in Table 2 after eliminating the data from two pairs in which male audience birds danced during the experiment.
Movie S1. Male blue-capped cordon-bleu courtship display directed to its partner in the presence of a female audience.
REFERENCES AND NOTES
- 1.Zuberbühler K., Audience effects. Curr. Biol. 18, R189–R190 (2008). [DOI] [PubMed] [Google Scholar]
- 2.Plath M., Richter S., Tiedemann R., Schlupp I., Male fish deceive competitors about mating preferences. Curr. Biol. 18, 1138–1141 (2008). [DOI] [PubMed] [Google Scholar]
- 3.Ung D., Amy M., Leboucher G., Heaven it’s my wife! Male canaries conceal extra-pair courtships but increase aggressions when their mate watches. PLOS ONE 6, e22686 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Fitzsimmons L. P., Bertram S. M., Playing to an audience: The social environment influences aggression and victory displays. Biol. Lett. 9, 20130449 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Auld H. L., Godin J.-G. J., Sexual voyeurs and copiers: Social copying and the audience effect on male mate choice in the guppy. Behav. Ecol. Sociobiol. 69, 1795–1807 (2015). [Google Scholar]
- 6.Overduin-De Vries A. M., Massen J. J. M., Spruijt B. M., Sterck E. H. M., Sneaky monkeys: An audience effect of male rhesus macaques (Macaca mulatta) on sexual behavior. Am. J. Primatol. 74, 217–228 (2012). [DOI] [PubMed] [Google Scholar]
- 7.Vignal C., Mathevon N., Mottin S., Audience drives male songbird response to partner’s voice. Nature 430, 448–451 (2004). [DOI] [PubMed] [Google Scholar]
- 8.Hall M. L., A review of hypotheses for the functions of avian duetting. Behav. Ecol. Sociobiol. 55, 415–430 (2004). [Google Scholar]
- 9.Ręk P., Magrath R. D., Multimodal duetting in magpie-larks: How do vocal and visual components contribute to a cooperative signal’s function? Anim. Behav. 117, 35–42 (2016). [Google Scholar]
- 10.Meseth E. H., The dance of the Laysan albatross, Diomedea immutabilis. Behaviour 54, 217–257 (1975). [Google Scholar]
- 11.Takeda K. F., Kutsukake N., Complexity of mutual communication in animals exemplified by pair dances in the red-crowned crane. Japanese J. Anim. Psychol. 68, 25–37 (2018). [Google Scholar]
- 12.Masatomi H., Some observations on mating behaviour of several cranes in captivity. J. Ethol. 1, 62–69 (1983). [Google Scholar]
- 13.Nuechterlein G. L., Storer R. W., The pair-formation displays of the Western Grebe. Condor 84, 351–369 (1982). [Google Scholar]
- 14.Malacarne G., Cucco M., Camanni S., Coordinated visual displays and vocal duetting in different ecological situations among western palearctic non-passerine birds. Ethol. Ecol. Evol. 3, 207–219 (1991). [Google Scholar]
- 15.Wachtmeister C. A., Display in monogamous pairs: A review of empirical data and evolutionary explanations. Anim. Behav. 61, 861–868 (2001). [Google Scholar]
- 16.D. Goodwin, Estrildid Finches of the World (Cornell Univ. Press, 1982). [Google Scholar]
- 17.Soma M., Garamszegi L. Z., Evolution of courtship display in Estrildid finches: Dance in relation to female song and plumage ornamentation. Front. Ecol. Evol. 3, 1–3 (2015). [Google Scholar]
- 18.Ota N., Gahr M., Soma M., Tap dancing birds: The multimodal mutual courtship display of males and females in a socially monogamous songbird. Sci. Rep. 5, 16614 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ota N., Gahr M., Soma M., Songbird tap dancing produces non-vocal sounds. Bioacoustics 26, 161–168 (2017). [Google Scholar]
- 20.Geberzahn N., Gahr M., Undirected (solitary) birdsong in female and male blue-capped cordon-bleus (Uraeginthus cyanocephalus) and its endocrine correlates. PLOS ONE 6, e26485 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.R. Zann, The Zebra Finch: A Synthesis of Field and Laboratory Studies (Oxford Univ. Press, 1996). [Google Scholar]
- 22.Kunkel P., Zum Verhalten einiger Prachtfinken (Estrildinae). Z. Tierpsychol. 16, 302–350 (1959). [Google Scholar]
- 23.Soma M., Iwama M., Mating success follows duet dancing in the Java sparrow. PLOS ONE 12, e0172655 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.M. Stevens, Sensory Ecology, Behaviour, and Evolution (Oxford Univ. Press, 2013). [Google Scholar]
- 25.Griffith S. C., Owens I. P. F., Thuman K. A., Extra pair paternity in birds: A review of interspecific variation and adaptive function. Mol. Ecol., 11, 2195–2212 (2002). [DOI] [PubMed] [Google Scholar]
- 26.Bolton P. E., Rollins L. A., Brazill-Boast J., Kim K.-W., Burke T., Griffith S. C., The colour of paternity: Extra-pair paternity in the wild Gouldian finch does not appear to be driven by genetic incompatibility between morphs. J. Evol. Biol. 30, 174–190 (2017). [DOI] [PubMed] [Google Scholar]
- 27.Searcy W. A., Beecher M. D., Song as an aggressive signal in songbirds. Anim. Behav. 78, 1281–1292 (2009). [Google Scholar]
- 28.Ritchison G., The singing behavior of female Northern cardinals. Condor 88, 156–159 (1986). [Google Scholar]
- 29.Levin R. N., Song behaviour and reproductive strategies in a duetting wren, Thryothorus nigricapillus: II. Playback experiments. Anim. Behav. 52, 1107–1117 (1996). [Google Scholar]
- 30.Kniel N., Bender S., Witte K., Sex-specific audience effect in the context of mate choice in zebra finches. PLOS ONE 11, e0147130 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hoi H., Griggio M., Bearded reedlings adjust their pair-bond behaviour in relation to the sex and attractiveness of unpaired conspecifics. PLOS ONE 7, e32806 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.M. Andersson, Sexual Selection (Princeton Univ. Press, 1994). [Google Scholar]
- 33.Girard M. B., Elias D. O., Kasumovic M. M., Female preference for multi-modal courtship: Multiple signals are important for male mating success in peacock spiders. Proc. Biol. Sci. 282, 20152222 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.de Luna A. G., Hödl W., Amézquita A., Colour, size and movement as visual subcomponents in multimodal communication by the frog Allobates femoralis. Anim. Behav. 79, 739–745 (2010). [Google Scholar]
- 35.Amorim M. C. P., Simões J. M., Fonseca P. J., Turner G. F., Species differences in courtship acoustic signals among five lake Malawi cichlid species (Pseudotropheus spp.). J. Fish Biol. 72, 1355–1368 (2008). [Google Scholar]
- 36.Cooper B. G., Goller F., Multimodal signals: Enhancement and constraint of song motor patterns by visual display. Science 303, 544–546 (2004). [DOI] [PubMed] [Google Scholar]
- 37.Dalziell A. H., Peters R. A., Cockburn A., Dorland A. D., Maisey A. C., Magrath R. D., Dance choreography is coordinated with song repertoire in a complex avian display. Curr. Biol. 23, 1132–1135 (2013). [DOI] [PubMed] [Google Scholar]
- 38.Perrot C., Béchet A., Hanzen C., Arnaud A., Pradel R., Cézilly F., Sexual display complexity varies non-linearly with age and predicts breeding status in greater flamingos. Sci. Rep. 6, 36242 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.R Core Team. R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, 2016); https://www.R-project.org/.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/4/10/eaat4779/DC1
Fig. S1. Examples of tail direction during a courtship display in which a male is courting a female.
Fig. S2. Protocol design and schematic views of the apparatus used in this experiment.
Fig. S3. Cumulative number of pairs that performed duet or solo multimodal displays in the four experimental conditions.
Table S1. The list of subject birds that performed multimodal displays when their partner stayed nearby and the percentage of the dance bouts with tail angling.
Table S2. Results of the same analysis as in Table 2 after eliminating the data from the five pairs whose two audience conditions were conducted on the same day.
Table S3. Results of the same analysis as in Table 2 after eliminating the data from two pairs in which male audience birds danced during the experiment.
Movie S1. Male blue-capped cordon-bleu courtship display directed to its partner in the presence of a female audience.