

Abstract
Atopic dermatitis (AD) is a chronic inflammatory skin disease with the hallmark characteristics of pruritus, psychological stress, and sleep disturbance, all possibly associated with an increased risk of attention-deficit/hyperactivity disorder (ADHD). However, the etiology of the possible association between AD and ADHD is still not well understood. 2,4-dinitrochlorobenzene or corticosterone was used to evaluate the atopic symptom and its psychologic stress in the atopic mice model. Melatonin, corticotropin-releasing hormone, corticotropin-releasing hormone receptor, urocortin, proopiomelanocortin, adrenocorticotropic hormone, corticosterone, cAMP, cAMP response element-binding protein, dopamine and noradrenaline were analyzed spectrophotometrically, and the expression of dopamine beta-hydroxylase and tyrosine hydroxylase were measured by Western blotting or immunohistochemistry. AD-related psychological stress caused an increase in the levels of dopamine beta-hydroxylase and tyrosine hydroxylase, degradation of melatonin, hyper-activity of the hypothalamic-pituitary-adrenal axis, and dysregulation of dopamine and noradrenaline levels (ADHD phenomena) in the locus coeruleus, prefrontal cortex, and striatum of the AD mouse brain. Notably, melatonin administration inhibited the development of ADHD phenomena and their-related response in the mouse model. This study demonstrated that AD-related psychological stress increased catecholamine dysfunction and accelerated the development of psychiatric comorbidities, such as ADHD.
Introduction
ADHD is a common childhood disorder affecting around 7% of all children, and often persists into adulthood1,2. According to the Diagnostic and Statistical Manual of Mental Disorders, ADHD includes 3 subtypes: inattentive, hyperactive-impulsive, and combined. To define these subtypes, 2 symptom dimensions have been used: inattention and hyperactivity-impulsivity3,4. These symptoms appear with varying severity and can lead to adverse consequences, including anxiety, depression, delinquency, sleep disorders, and circadian rhythm defects2,3,5. Despite these consequences, the complex molecular mechanisms underlying ADHD are still understudied and unclear. The main hypothesis, centered on monoamine neurotransmission, supposes that in ADHD, complex interactions between the dopamine, noradrenaline, and serotonin systems are deregulated; human studies have associated ADHD with genes encoding monoamine receptors and transporters6,7. Most treatments available today include the administration of psycho stimulants that increase levels of available dopamine and are believed to restore monoaminergic balance, which is altered during ADHD development8–10.
Atopic dermatitis (AD) is a type of chronic inflammation of the skin characterized by eczematous skin lesions, papules, severe pruritus, and excoriations11,12. AD affects 10–20% of children worldwide and persists into adulthood in a minority of cases (approx. 2–3% of adults), with an increased prevalence in urbanized societies13. Importantly, AD is associated with other non-allergic diseases, including psychiatric and behavioral disorders14–17. The prevalence of depression, stress-related disorders, and anxiety are significantly higher in the AD population18,19. Recent data indicate that chronic AD might directly or indirectly lead to an increased risk of psychiatric comorbidities, such as ADHD20. Epidemiological data suggests that the global prevalence of AD and ADHD has risen parallelly, and several cross-sectional studies have indicated co-occurrence20–22. Furthermore, recent cohort studies have reported a temporal association between AD and ADHD development20,22. Children with AD have a 1.5-fold higher risk for ADHD, and the ascribed population risk for ADHD explained by AD is roughly 9%15,20. According to many clinicians, it is possible that the psycho-endocrine and psycho-neuroimmunological effects of AD are caused by elevated inflammatory cytokine levels, continuous sensory stimuli, disturbed sleep, and increased stress15,20. Currently, the etiology of the association between AD and ADHD is still not well understood.
Many studies have focused on the role of stress as a relevant trigger of AD symptoms. Patients with AD demonstrate blunted hypothalamic-pituitary-adrenal (HPA) axis responsiveness and over-reactivity of the sympathetic-adrenal-medullary system to psychosocial stress23,24. Considering immune-regulatory roles of the HPA axis and the sympathetic-adrenal-medullary systems, especially under stress, an aberrant responsiveness of these systems may increase susceptibility to allergic inflammation and may be a psycho-biological mechanism of stress-related aggravation of AD24,25. These ideas are supported by a study showing raised glucocorticoid and adrenocorticotropic hormone (ACTH) levels under stress in adults with AD, which indicate a dysfunctional HPA axis. Interestingly, response dysfunction of the HPA axis during stress exposure is accompanied by atopy-related immunological changes in response to stressors such as increased immunoglobulin E levels, altered IL4 and interferons concentrations, and aggravation of AD symptomatology25–27. Additionally, recent papers report that continuous high levels of corticotropin-releasing factor-related to psychological stress can trigger AD-like skin lesions25,28,29. Surprisingly, according to Corminas et al., the cortisol response to stress in adults with ADHD was not significantly different from that in the healthy controls3. Nevertheless, clear differences can be observed between the combined and inattentive subtypes3,30. Compared with the mainly inattentive adults, patients with combined ADHD exhibited a blunted cortisol response to stress3. In contrast, the inattentive ADHD subtype was characterized by a trend towards a higher cortisol response than that observed in the controls3,30,31. These reports suggest the significance of distinguishing the inattentive and combined subtypes in the diagnosis of ADHD. Further, these findings implicate dysregulation of the HPA-axis, which is involved in several physiological systems and is central to stress regulation, in the development of ADHD.
The action of glucocorticoids on many physiological systems is to mediate stress-responses, but their long-term chronic elevation impairs catecholamine systems32–35. Specifically, ACTH and the sympathetic nervous system stimulate the synthesis of adrenaline precursors by enhancing the activity of tyrosine hydroxylase (TH) and dopamine beta-hydroxylase (DβH), two enzymes involved in catecholamine synthesis36–38. ACTH also stimulates the adrenal cortex to release corticosterone (CORT), which enhances adrenaline synthesis38,39. Moreover, patients with AD show elevated norepinephrine concentrations, similar to the results observed for patients with ADHD40. Thus, several clinical studies have found associations between AD, ADHD, and attention deficits.
Thus, we hypothesized that psychological stress caused by AD can increase DβH levels, which increases overall dysregulation of dopamine and noradrenaline, creating a state of chronic psychiatric comorbidities, such as ADHD. Our previous studies indicated that atopic stress significantly accelerates neuronal injury and increases neuro-inflammation, primarily via exacerbating a chronic state of blunted glucocorticoid activity and suppressing melatonin-mediated feedback25. In the present study, we employed an AD model to test the aforementioned hypothesis, which included analyzing dopamine and noradrenaline imbalance.
Materials and Methods
Chemical
Chemical analysis was performed as previously described25. Additionally, rabbit and goat anti-TH and rabbit anti-DβH antibodies were purchased from EMD Millipore (Billerica, MA, USA). A mouse DβH enzyme-linked immunosorbent assay (ELISA) kit was purchased from Aviva Systems Biology (San Diego, CA). A norepinephrine and dopamine 2-CAT ELISA Kit were purchased from Rocky Mountain Diagnostic, Inc. (Colorado Springs, CO, USA). All other reagents used were of guaranteed or analytical grade.
In vitro and in vivo models
Immortalized human SH-SY5Y cell culture and the 2,4-dinitrochlorobenzene (DNCB) or CORT treatment animal models were established according to previously published methods25,41,42. The institutional animal care committee of the Korea Institute of Oriental Medicine (KIOM) approved the experimental protocols KIOM-16-015 and KIOM-16-112. The experiments were performed according to the guidelines of the Animal Care and Use Committee at KIOM25. The animals were sacrificed between 11:00 a.m. and 14:30 p.m., 7 weeks after sensitization with DNCB or CORT (Fig. 1 shows the study timeline).
Figure 1.
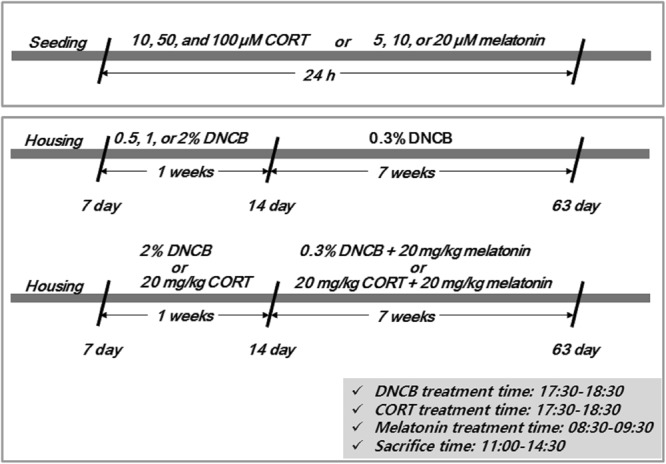
Summary of the experimental design.
Brain tissue preparation and immunofluorescence analysis
Brain tissue preparation was performed as previously described25. Briefly, the mice were decapitated; the skull was then removed and the brain was dissected around selected regions including the locus coeruleus (LC), prefrontal cortex (PC), and striatum (ST) using a brain matrix for kit-based analyses. All tissues were frozen in situ by immediate direct immersion in liquid nitrogen in order to prevent decomposition. Immunofluorescence analysis was then performed as described in a previous study25.
Measurement of stress-related factors
Melatonin, urocortin (UCN), DβH, proopiomelanocortin (POMC), ACTH, CORT, corticotropin-releasing hormone (CRH), corticotropin-releasing hormone receptor 1 (CRHR1), cyclic adenosine monophosphate (cAMP), CREB phosphorylation (pCREB), norepinephrine, and dopamine were quantified using commercially available kits according to the instruction manuals or previously published methods25.
Statistical analysis
All statistical parameters were calculated using Graphpad Prism 5.0 software (Graphpad Software, San Diego, CA, USA). Values are expressed as means ± standard error of the mean (S.E.M.). Statistical comparisons between the different treatments were performed using a one-way ANOVA with Tukey’s multiple comparison post hoc test. p values of <0.05 were considered to be statistically significant.
Results
Effects of CORT and melatonin on DβH levels in SH-SY5Y cells
We measured DβH levels using ELISA kits after exposing SH-SY5Y cells to CORT or melatonin. CORT exposure significantly increased DβH levels (Fig. 2A and Supplementary Table 1) and melatonin exposure significantly decreased DβH levels (Fig. 2B and Supplementary Table 1).
Figure 2.

Effects of CORT on expression of DβH in SH-SY5Y cells. The levels of DβH were quantified using ELISA kits (A). Effects of melatonin on expression of DβH in SH-SY5Y cells. The levels of DβH were quantified using ELISA kits (B). Values are shown as means ± standard error of the mean. *P < 0.05, **P < 0.01, and ***P < 0.001 compared with the control group. CORT, corticosterone; DβH, dopamine beta-hydroxylase.
Effects of DNCB on melatonin in the LC, PC, and ST
We measured melatonin levels using an ELISA kit. The 1–2% DNCB groups demonstrated significantly decreased melatonin levels in the LC (Fig. 3A and Supplementary Table 2) and PC (Fig. 3B and Supplementary Table 2). However, they showed only weak decreases in the ST (Fig. 3C and Supplementary Table 2).
Figure 3.
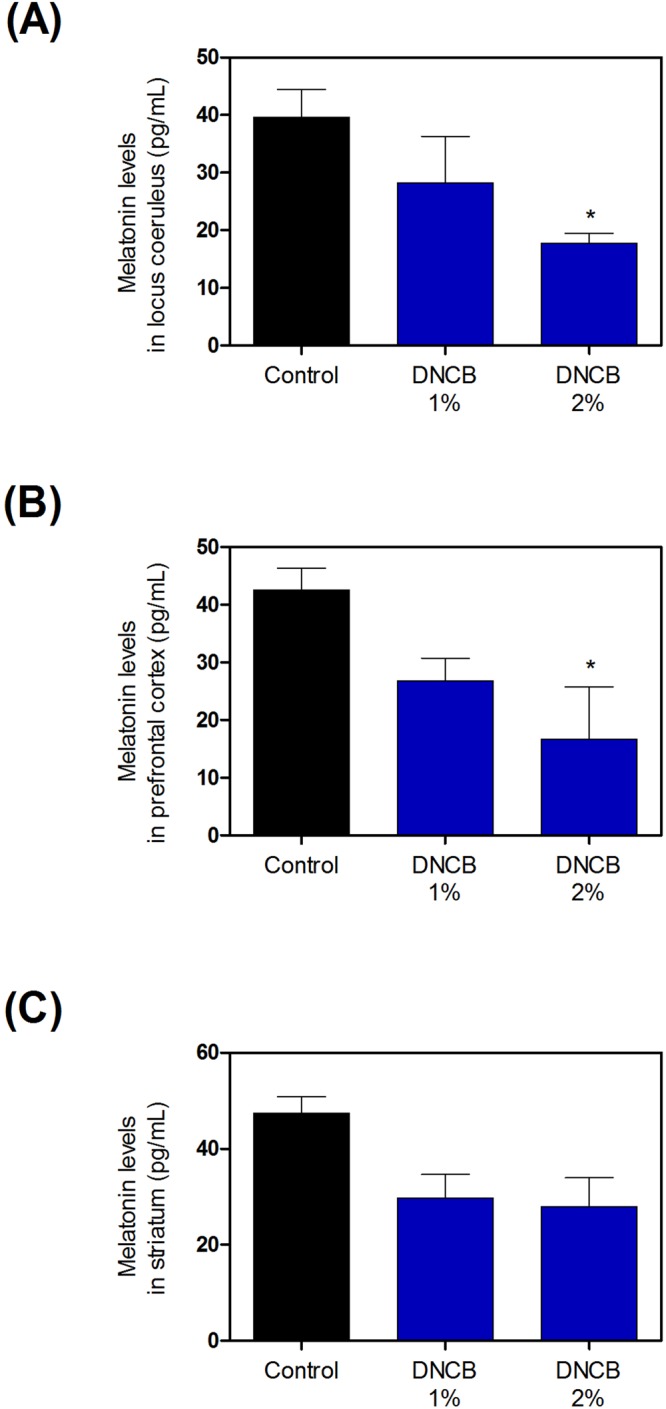
Effects of DNCB on expression of melatonin in brain. The levels of melatonin in locus coeruleus (A), prefrontal cortex (B), and striatum (C) of were quantified using ELISA kits. Values are presented as means ± standard error of the mean. *P < 0.05 compared with the control group. DNCB, dinitrochlorobenzene; ELISA, enzyme-linked immunosorbent assay.
Effects of melatonin on DNCB-induced HPA axis activity biomarkers
To determine whether melatonin affected stress hormone responses, we measured CRH and CRHR levels. Treatment with DNCB significantly increased CRH levels (Fig. 4A–C, and Supplementary Table 3), while treatment with 20 mg/kg melatonin reduced the DNCB-induced CRH level increase (Fig. 4A–C, and Supplementary Table 3). Also, treatment with DNCB significantly increased CRHR levels (Fig. 3D–F, and Supplementary Table 3), while treatment with 20 mg/kg melatonin reduced the DNCB-induced CRHR increase (Fig. 4D–F, and Supplementary Table 3).
Figure 4.

Effects of melatonin on DNCB-induced expression of CRH and CRHR in locus coeruleus, prefrontal cortex, and striatum of the brain. The levels of CRH (A–C) and CRHR (D–F) in of were quantified using ELISA kits. Representative photomicrographs are shown (G). Values are shown as means ± standard error of the mean. *P < 0.05 and **P < 0.01 compared with the control group; #P < 0.05 and ##P < 0.01 compared with the 2% DNCB-alone group. DNCB, dinitrochlorobenzene; CRH, corticotropin releasing hormone; CRHR, corticotropin releasing hormone receptor; ELISA, enzyme-linked immunosorbent assay.
Further, to determine whether melatonin affected the stress cascade, we measured UCN, POMC, ACTH, and CORT levels. Treatment with DNCB significantly increased the levels of UCN (Fig. 5A–C, and Supplementary Table 4), while treatment with 20 mg/kg melatonin reduced the DNCB-induced UCN increase (Fig. 5A–C, and Supplementary Table 4). Treatment with DNCB significantly increased the levels of POMC (Fig. 5D–F, and Supplementary Table 4), while treatment with 20 mg/kg melatonin reduced this DNCB-induced POMC increase (Fig. 5D–F, and Supplementary Table 4). Treatment with DNCB significantly increased the levels of ACTH (Fig. 5G–I, and Supplementary Table 4), while treatment with 20 mg/kg melatonin reduced this DNCB-induced ACTH increase (Fig. 5G–I, and Supplementary Table 4). Treatment with DNCB significantly increased the levels of CORT (Fig. 5J–L, and Supplementary Table 4), while treatment with 20 mg/kg melatonin reduced this DNCB-induced CORT increase (Fig. 5J–L, and Supplementary Table 4).
Figure 5.

Effects of melatonin on DNCB-induced expression of the CRH-related HPA axis cascade. The levels of UCN (A–C), POMC (D–F), ACTH (G–I), and CORT (J–L) were measured by ELISA kit. Values are means ± standard error of the mean. *P < 0.05, **P < 0.01, **P < 0.01 and ***P < 0.001 compared with the control group; #P < 0.05, ##P < 0.01, and ###P < 0.001 compared with the 2% DNCB-alone group. DNCB, dinitrochlorobenzene; CRH, corticotropin releasing hormone; HPA, hypothalamic-pituitary-adrenal; UCN; POMC, pro-opiomelanocortin; ACTH, adrenocorticotropic hormone; CORT, corticosterone; ELISA, enzyme-linked immunosorbent assay.
Effects of melatonin on DNCB-induced stress response cAMP-pCREB in the LC
We assessed cAMP-pCREB pathway activity in the LC. Exposure of skin to 1–2% DNCB significantly increased cAMP (Fig. 6A and Supplementary Table 5) and pCREB (Fig. 6B and Supplementary Table 5) levels in the LC (Fig. 6B,C, and Supplementary Table 5). However, treatment with 20 mg/kg melatonin reduced this DNCB-induced cAMP increase (Fig. 6A and Supplementary Table 5) and pCREB (Fig. 6B and Supplementary Table 5).
Figure 6.
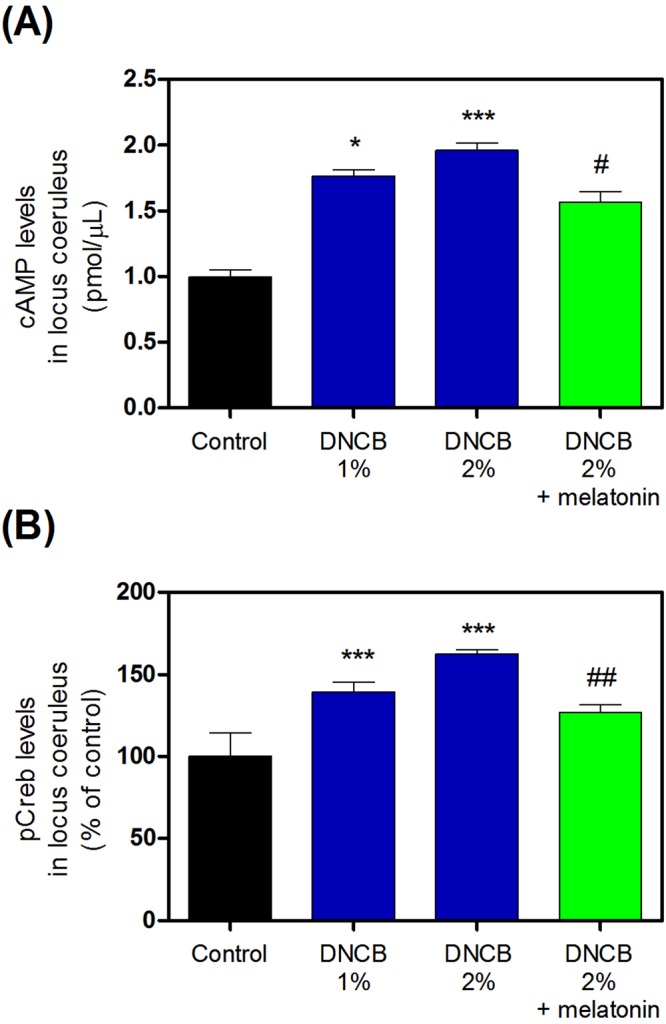
Effects of melatonin on DNCB-induced expression of the stress response cAMP-pCREB signaling in the locus coeruleus. The expression of cAMP (A) and pCREB (B) were measured by ELISA kit or immunofluorescence. Values are means ± standard error of the mean. *P < 0.05 and ***P < 0.001 compared with the control group. DNCB, dinitrochlorobenzene; cAMP, cyclic adenosine monophosphate; pCREB, phosphorylated C-element response binding protein; ELISA, enzyme-linked immunosorbent assay.
Effects of melatonin on DNCB-induced dopamine metabolic enzyme levels
To determine whether melatonin affects dopamine responses, we measured TH and DβH levels. Treatment with DNCB and DNCB + melatonin did not change the levels of TH compared with the control group (Fig. 7A–C, and Supplementary Table 6). However, treatment with DNCB significantly increased the levels of DβH (Fig. 7E–G, and Supplementary Table 6), while treatment with 20 mg/kg melatonin reduced the DNCB-induced DβH increase (Fig. 7D–F, and Supplementary Table 6).
Figure 7.

Effects of melatonin on DNCB-induced expression of the dopamine-related proteins in the locus coeruleus, prefrontal cortex, and striatum. The expression of tyrosine hydroxylase (A–C) and dopamine beta-hydroxylase (DβH; D–F) were measured by immunofluorescence. Representative photomicrographs are shown (G). Values are means ± standard error of the mean. *P < 0.05 and ***P < 0.001 compared with the control group.
Effects of melatonin on DNCB-induced dopamine and norepinephrine levels
We measured dopamine and norepinephrine levels after treatment with DNCB. Treatment with DNCB significantly increased the levels of dopamine (Fig. 8A–C, and Supplementary Table 7), while treatment with 20 mg/kg melatonin reduced this DNCB-induced dopamine increase (Fig. 8A–C, and Supplementary Table 7). Furthermore, treatment with DNCB significantly increased the levels of norepinephrine (Fig. 8D–F, and Supplementary Table 7), while treatment with 20 mg/kg melatonin reduced this DNCB-induced norepinephrine increase (Fig. 8D–F, and Supplementary Table 7).
Figure 8.

Effects of melatonin on DNCB-induced expression of the dopamine and noradrenaline contents in the locus coeruleus, prefrontal cortex, and striatum of the brain. The expression of dopamine (A–C) and noradrenaline (D–F) were measured using ELISA kits. Values are means ± standard error of the mean. *P < 0.05 and ***P < 0.001 compared with the control group. DNCB, dinitrochlorobenzene; ELISA, enzyme-linked immunosorbent assay.
Effects of melatonin on CORT-induced dopamine and norepinephrine levels
We determined dopamine and norepinephrine levels after treatment with CORT. Treatment with CORT significantly increased the levels of dopamine (Fig. 9A–C, and Supplementary Table 8), while treatment with 20 mg/kg melatonin reduced this CORT-induced dopamine increase (Fig. 9A–C, and Supplementary Table 8). Furthermore, treatment with CORT also significantly increased the levels of norepinephrine (Fig. 9D–F, and Supplementary Table 8), while treatment with 20 mg/kg melatonin reduced this CORT-induced norepinephrine increase (Fig. 9D–F, and Supplementary Table 8).
Figure 9.

Effects of melatonin on CORT-induced expression of the dopamine and noradrenaline contents in the locus coeruleus, prefrontal cortex, and striatum of the brain. The expression of dopamine (A–C) and noradrenaline (D–F) were measured by ELISA kits. Values are means ± standard error of the mean. *P < 0.05 and ***P < 0.001 compared with the control group. CORT, corticosterone; ELISA, enzyme-linked immunosorbent assay.
Discussion
The data presented herein demonstrate that the atopy-induced stress response significantly increased the presence of signaling molecules involved in ADHD. This response is due to factors such as dopamine and noradrenalin imbalances (via the up-regulation of DβH), which exacerbate HPA dysfunction and suppress the melatonin feedback system. Atopic stress led to: (1) induction of CRH-related and suppressing melatonin signaling in the LC and ST; (2) increased levels of DβH that did not alter TH levels; and (3) decreased dopamine levels and increased noradrenalin levels. Melatonin reversed these effects on dopamine and noradrenalin via stimulation of the normal HPA axis. To the best of our knowledge, in the first study to demonstrate that AD-caused psychologic stress increases catecholamine dysfunction and accelerated the development of psychiatric comorbidities, such as ADHD via dysregulation of the HPA/melatonin signaling pathways.
ADHD, characterized by inattention, hyperactivity/impulsivity, or both, is one of the most common psychiatric disorders of childhood43. Approximately one third of medication-free children with ADHD experience chronic sleep-onset insomnia43–45. Although the safety and efficacy of melatonin treatment for sleep-onset insomnia in children without ADHD have been well-documented, melatonin efficacy has not been studied in medication-free children with ADHD and sleep-onset insomnia; this patient group is of special interest for several reasons46,47. First, medication-free children with both disorders exhibit a delayed evening increase in endogenous melatonin levels, and this phase delay predicts strengthening of the sleep phase, which normalizes the effect of exogenous melatonin in children without ADHD46. Second, because treating sleep-related disorders other than insomnia improves daytime function in children with ADHD, treating insomnia may have important consequences for ADHD treatment strategies48–50. The night time circadian rise in melatonin levels correlates with a nighttime circadian drop in glucocorticoids51,52. Any chronic, late-night stressor (e.g., shift-work53) can result in excessively high nighttime cortisol levels, which may impair the normal morning circadian increase in corticosteroid levels54. It is known that circadian rhythms are highly related to the HPA axis, which is a key neuroendocrine mediator of physiological responses to psychological stressors55–57. In a previous study, we found that hyper-activation of HPA axis induced by atopic chronic psychological stress was associated with neurotoxicity and cognitive impairment, and it also blunted neuroendocrine responses to stress25. In this and our previous paper, we focused on the idea that neuroendocrine contribution to the responses in skin to stress is promoted, in part, by local synthesis of all elements of the HPA axis. Skin has the ability to synthesize glucocorticoids from cholesterol or steroid intermediates of systemic origin58,59. By interacting with glucocorticoid receptors, they regulate skin immune functions as well as functions and phenotype of the epidermal, dermal and adnexal compartments58,59. Most of the biochemical (enzyme and transporter activities) and regulatory principles of cutaneous glucocorticosteroidogenesis (neuropeptides mediated activation of cAMP and protein kinase A dependent pathways) are similar to those operating in classical steroidogenic organs58–61.
According to a recent report, stimulation of cutaneous corticosteroidogenesis can occur via this skin homologue of the HPA axis which is dependent on the functional activating CRHR and processing POMC60,61. Specifically, the stimulating cortisol synthesis by IL1 is intriguing because IL1 serves as a local signal of skin inflammatory injury60,61. It is possible that IL1 may stimulate corticosteroidogenesis indirectly, through up-regulating CRH or POMC peptides or by itself, which has been reported to occur in the adrenals60,61. CRH, in addition to indirect stimulation may directly stimulate local corticosteroidogenesis because it increases cAMP62,63. A similar direct action of CRH on adrenocortical region has been reported62,63. An additional regulating-mechanism involves the activation or inactivation of glucocorticosteroids by locally expressed cortisone reductase, such as 11β-hydroxysteroid dehydrogenase type1 and 262–64.
In this and our previous paper, we confirmed that melatonin control anti-inflammation by regulating the HPA axis via the skin. Melatonin plays an important role in the regulating circadian timing and neuro-immunologic function54,65. Recent studies have linked decreases in melatonin output to insomnia in aged patients and sleep disturbances in patients with Alzheimer’s disease25,66–69. In AD, a severity score ≥48.7 predicts elevated levels of immunoglobulin, sensitive allergen, and poor sleep efficiency, possibly because of reduced nocturnal melatonin secretion, pruritus, and associated scratching25,69. One study reported disrupted melatonin secretion in eczema, possibly due to partial action of the sympathetic nervous reduction that regulates secretion of melatonin17,25. Recent studies have shown that melatonin is synthesized in numerous extrapineal sites54,65,70. Additionally, melatonin is regulated by rapid metabolism in the liver and peripheral organs including the skin54. Several researchers have proposed that melatonin and its metabolites affect skin functions and structures through actions mediated by intracutaneously expressed cell-surface and putative nuclear receptors54,70. Melatonin exerts both receptor-dependent and receptor-independent protective effects against oxidative stress and can attenuate environmental skin stressor–induced damage71. The effects of the common environmental skin stressors are modulated by melatonin via a complex intra-cutaneous melatonergic anti-oxidative system, with ultraviolet radiation-enhanced melatonin metabolism generating bioactive melatonin metabolites such as acetyl-N-formyl-5-methoxykynurenamine70,71. These properties suggest that melatonin is an important endogenous effector of intracutaneous stress responses. In fact, there is a shortage of information concerning the regulation of HPA axis-melatonin-circadian rhythms on the glucocorticosteroidogenesis signaling system in skin61–63. However, previously, we found that NC/Nga mice exposed to atopic stress exhibited substantial reductions in hypothalamic melatonin membrane receptor expression and in hypothalamic and intracutaneous melatonin expression25. Additionally, historically, oriental medicine has suggested a similar connection with the thought that biological rhythm is an inherent connotation of “harmony between human and nature”72. Our previous study did not show whether intracutaneous melatonin affects the brain, but many other studies have suggested that it has an important effect. Further research is certainly needed, but our observation of decreased melatonin levels in the LC, PC, and ST in DNCB-exposed NC/Nga mice supports this hypothesis.
In the present study, we tried to investigate effects of stress in the LC. Some anti-depressants as well as the ADHD medication atomoxetine, are believed to act on LC neurons73,74. The LC is responsible for mediating several sympathetic effects of stress; it is activated by stress and responds by increasing norepinephrine secretion, which in turn alters cognitive function (through the PC), increases motivation (through nucleus accumbens), and activates the HPA axis75. After HPA axis stimulation, norepinephrine stimulates the secretion of CRH from the hypothalamus, which induces ACTH release from the anterior pituitary and subsequent cortisol synthesis in the adrenal glands55,76,77. Norepinephrine released from the LC will inhibit its own production, and CRH will inhibit its own production while causing the LC to increase norepinephrine production73. Thus, to determine whether mechanisms of the dopamine and noradrenaline imbalance caused by atopic stress were regulated by melatonin, we measured ADHD signaling patterns in an NC/Nga atopic mouse model. We showed that DNCB significantly increased levels of HPA axis-related response substances such as CRH, CRHR, UCN, POMC, ACTH, and CORT, while treatment with melatonin significantly reduced these levels in DNCB-treated mice. Further, corticosteroid-mediated psychological stress responses utilize various signal transduction systems78–80. Studies have shown that in hippocampal neurons or slices, ERK1/2 respond to stressful stimuli through the transcription factor CREB (cyclic response element binding protein), which activates c-Fos via CRE sites in promoter regions79. Additionally, stress increases phosphorylating CREB80. Moreover, the PI3K-cAMP-CREB pathway activity is elevated in the noradrenergic neurons of the LC, which is the main source of noradrenaline in the brain37. PI3Kc gene KO mice show increased CREB activation via elevation of cAMP levels in the LC and alters the dopamine/noradrenaline balance in the PC and ST7,37. These changes facilitate the development of core ADHD-related phenotypes, including hyperactivity and attention deficit, as well as secondary features such as memory and social impairments7,37. Overexpression of CREB in the LC of normal animals produces similar behavioral changes, and down-regulation of CREB activity in the LC of mutant mice reverses the phenotype7,37. In the present study, we observed significantly increased cAMP and pCREB expression in the LC; subchronic melatonin reduced cAMP and pCREB expression in DNCB-treated mice.
Further, we examined DβH and TH levels. DβH is anenzyme that converts DA into NE and is coreleased with catecholamines81. TH is involved in the conversion of phenylalanine to dopamine81. As the rate-limiting enzyme in the synthesis of catecholamines, TH plays a key role in the physiology of adrenergic neurons; TH is regularly used as a marker for dopaminergic neurons81–83. DβH is a genetic marker and may reflect individual susceptibility to developing psychosis in the context of exposure to traumatic events84. Generally, adrenal catecholamines are known to mediate many of the physiological consequences of the “fight or flight” response to stress84,85. However, the mechanisms by which the long-term responses to repeated stress exposure are mediated are not well understood. According to McMahon et al., both TH and DβH levels are elevated by single and repeated exposure to immobilization stress84,85. Surprisingly, we observed no or slight change in TH expression in the LC. However, DβH levels were significantly upregulated in the LC, PC, and ST of DNCB-treated mice. Further, subchronic melatonin reduced DβH expression. We also observed significantly decreased dopamine and increased noradrenaline levels in the LC, PC, and ST of DNCB-treated mice; subchronic melatonin treatment reduced these levels. This pattern demonstrated similar results in CORT injection model. In this study, we suggested that the imbalance resulting from increased release of dopamine and noradrenaline for potential modulation of physiological or immunological responses is the consequence of the upregulated DβH expression and the unchanged TH expression in DNCB exposed to atopic stress. However, future research will be needed to analyze the specific mechanisms system. Thus, AD-related neuropsychological stress caused the relationship of normal glucocorticoid/melatonin disruption and accelerated dopamine dysregulation.
This study has some limitations which have to be pointed out. We did not assess whether atopic stress induced ADHD behavioral pattern of chronic stress, because the scratching behavior and memory impairments interfered with the rodent behavioral tasks. Several pilot experiments were conducted, but they were unsuccessful. Additionally, melatonin is correlated with the circadian rhythm phase shift of the circadian clock; hence, a change in sleep-wake time is expected. However, the experiments were unsuccessful as itching interfered with the sleep-wake time and the sleep-wake time criterion was unclear. Thus, future studies will need to analyze the wake-sleep states, specific response of stress, and their potential behavioral outcomes.
Conclusions
In conclusion, we found that atopic stress accelerated HPA-axis dysfunction, increased norepinephrine, and decreased dopamine (Fig. 10). Atopic stress induced HPA axis-related response dysfunction and ERK-CREB signaling pathway, which reduced melatonin levels in the LC, PC, and ST. Moreover, atopy-induced stress accelerated DβH level increases and dopamine consumption. Further, we discovered that melatonin inhibited reduced dopamine consumption by the inhibition of DβH via regulating HPA in AD models. Therefore, our findings suggest that the CORT-melatonin disequilibrium might contribute to dysfunction of dopamine by causing or enhancing neurodegeneration, which could lead to disorders such as ADHD.
Figure 10.

Schematic of the mechanism proposed for the effects of melatonin on the hypothalamic-pituitary-adrenal (HPA) axis and attention deficit hyperactivity disorder (ADHD) pathogenesis.
Electronic supplementary material
Acknowledgements
We thank Prof. Kim, Prof. Park, and Dr. Kim for providing the ELISA kits. We also thank Prof. Chung for help in the Department Institute of Human-Environment Interface Biology, Medical Research Center, Seoul National University Hospital (SNUH) laboratory and office. This work was supported by a National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIP) (NRF-2015R1C1A1A01054802).
Author Contributions
Dr. Park and Prof. Kim conceived the study and designed the experiments. Dr. Park, Prof. Jung, and Prof. Yang conducted the experiments and data analysis and assisted in data interpretation and manuscript preparation. Dr. Park, Prof. Kim, Prof. Jung, Prof. Park, and Prof. Yang wrote, corrected, and revised the entire manuscript. All authors provided suggestions for and approved the manuscript.
Competing Interests
The authors declare no competing interests.
Footnotes
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Gunhyuk Park, Email: parkgunhyuk@gmail.com.
Yong-ung Kim, Email: ykim@dhu.ac.kr.
Electronic supplementary material
Supplementary information accompanies this paper at 10.1038/s41598-018-33317-x.
References
- 1.Kirov R, Brand S. Sleep problems and their effect in ADHD. Expert review of neurotherapeutics. 2014;14:287–299. doi: 10.1586/14737175.2014.885382. [DOI] [PubMed] [Google Scholar]
- 2.Zhou RY, et al. Attention deficit hyperactivity disorder may be a highly inflammation and immune-associated disease. Molecular Medicine Reports. 2017;16:5071–5077. doi: 10.3892/mmr.2017.7228. [DOI] [PubMed] [Google Scholar]
- 3.Corominas-Roso M, et al. Cortisol response to stress in adults with attention deficit hyperactivity disorder. International Journal of Neuropsychopharmacology. 2015;18:pyv027. doi: 10.1093/ijnp/pyv027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lee SJ, Park S, Jeong A, Chae H, Oh K. Temperamental predictors of developmental trajectories of inattention and hyperactivity–impulsivity problems in schoolchildren. Integr Med Res. 2017;6:33–40. doi: 10.1016/j.imr.2016.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.der Heijden KBV, Smits MG, Van Someren EJ, Ridderinkhof KR, Gunning WB. Effect of melatonin on sleep, behavior, and cognition in ADHD and chronic sleep-onset insomnia. Journal of the American Academy of Child & Adolescent Psychiatry. 2007;46:233–241. doi: 10.1097/01.chi.0000246055.76167.0d. [DOI] [PubMed] [Google Scholar]
- 6.del Campo N, Chamberlain SR, Sahakian BJ, Robbins TW. The roles of dopamine and noradrenaline in the pathophysiology and treatment of attention-deficit/hyperactivity disorder. Biological psychiatry. 2011;69:e145–e157. doi: 10.1016/j.biopsych.2011.02.036. [DOI] [PubMed] [Google Scholar]
- 7.D’Andrea, I. et al. Lack of kinase‐independent activity of PI3Kγ in locus coeruleus induces ADHD symptoms through increased CREB signaling. EMBO molecular medicine, e201404697 (2015). [DOI] [PMC free article] [PubMed]
- 8.Castle L, Aubert RE, Verbrugge RR, Khalid M, Epstein RS. Trends in medication treatment for ADHD. Journal of attention disorders. 2007;10:335–342. doi: 10.1177/1087054707299597. [DOI] [PubMed] [Google Scholar]
- 9.Volkow ND, et al. Brain dopamine transporter levels in treatment and drug naive adults with ADHD. Neuroimage. 2007;34:1182–1190. doi: 10.1016/j.neuroimage.2006.10.014. [DOI] [PubMed] [Google Scholar]
- 10.Gilbert DL, et al. Dopamine transporter genotype influences the physiological response to medication in ADHD. Brain. 2006;129:2038–2046. doi: 10.1093/brain/awl147. [DOI] [PubMed] [Google Scholar]
- 11.Spergel JM, Paller AS. Atopic dermatitis and the atopic march. Journal of Allergy and Clinical Immunology. 2003;112:S118–S127. doi: 10.1016/j.jaci.2003.09.033. [DOI] [PubMed] [Google Scholar]
- 12.Saeki H, et al. Guidelines for management of atopic dermatitis. The Journal of dermatology. 2009;36:563–577. doi: 10.1111/j.1346-8138.2009.00706.x. [DOI] [PubMed] [Google Scholar]
- 13.Leung DY. Atopic dermatitis: the skin as a window into the pathogenesis of chronic allergic diseases. Journal of Allergy and Clinical Immunology. 1995;96:302–319. doi: 10.1016/S0091-6749(95)70049-8. [DOI] [PubMed] [Google Scholar]
- 14.Chida Y, Hamer M, Steptoe A. A bidirectional relationship between psychosocial factors and atopic disorders: a systematic review and meta-analysis. Psychosomatic Medicine. 2008;70:102–116. doi: 10.1097/PSY.0b013e31815c1b71. [DOI] [PubMed] [Google Scholar]
- 15.Schmitt J, Buske‐Kirschbaum A, Roessner V. Is atopic disease a risk factor for attention‐deficit/hyperactivity disorder? A systematic review. Allergy. 2010;65:1506–1524. doi: 10.1111/j.1398-9995.2010.02449.x. [DOI] [PubMed] [Google Scholar]
- 16.Buske-Kirschbaum A, et al. Psychoendocrine and psychoneuroimmunological mechanisms in the comorbidity of atopic eczema and attention deficit/hyperactivity disorder. Psychoneuroendocrinology. 2013;38:12–23. doi: 10.1016/j.psyneuen.2012.09.017. [DOI] [PubMed] [Google Scholar]
- 17.Chang Y-S, Chiang B-L. Mechanism of sleep disturbance in children with atopic dermatitis and the role of the circadian rhythm and melatonin. International journal of molecular sciences. 2016;17:462. doi: 10.3390/ijms17040462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Slattery MJ, Essex MJ. Specificity in the association of anxiety, depression, and atopic disorders in a community sample of adolescents. Journal of psychiatric research. 2011;45:788–795. doi: 10.1016/j.jpsychires.2010.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gupta MA, Gupta AK. Psychiatric and psychological co-morbidity in patients with dermatologic disorders. American journal of clinical dermatology. 2003;4:833–842. doi: 10.2165/00128071-200304120-00003. [DOI] [PubMed] [Google Scholar]
- 20.Nygaard U, Riis JL, Deleuran M, Vestergaard C. Attention-Deficit/Hyperactivity Disorder in Atopic Dermatitis: An Appraisal of the Current Literature. Pediatric Allergy, Immunology, and Pulmonology. 2016;29:181–188. doi: 10.1089/ped.2016.0705. [DOI] [PubMed] [Google Scholar]
- 21.Yaghmaie P, Koudelka CW, Simpson EL. Mental health comorbidity in patients with atopic dermatitis. Journal of Allergy and Clinical Immunology. 2013;131:428–433. doi: 10.1016/j.jaci.2012.10.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Strom M, Fishbein A, Paller A, Silverberg J. Association between atopic dermatitis and attention deficit hyperactivity disorder in US children and adults. British Journal of Dermatology. 2016;175:920–929. doi: 10.1111/bjd.14697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hall Jessica M. F., Cruser desAnges, Podawiltz Alan, Mummert Diana I., Jones Harlan, Mummert Mark E. Psychological Stress and the Cutaneous Immune Response: Roles of the HPA Axis and the Sympathetic Nervous System in Atopic Dermatitis and Psoriasis. Dermatology Research and Practice. 2012;2012:1–11. doi: 10.1155/2012/403908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Konakchieva R, Mitev Y, Almeida OF, Patchev VK. Chronic melatonin treatment counteracts glucocorticoid-induced dysregulation of the hypothalamic-pituitary-adrenal axis in the rat. Neuroendocrinology. 1998;67:171–180. doi: 10.1159/000054312. [DOI] [PubMed] [Google Scholar]
- 25.Park, G., Lee, S. H., Oh, D. S. & Kim, Y. U. Melatonin inhibits neuronal dysfunction‐associated with neuroinflammation by atopic psychological stress in NC/Nga atopic‐like mouse models. Journal of pineal research (2017). [DOI] [PubMed]
- 26.Buske-Kirschbaum A, Fischbach S, Rauh W, Hanker J, Hellhammer D. Increased responsiveness of the hypothalamus–pituitary–adrenal (HPA) axis to stress in newborns with atopic disposition. Psychoneuroendocrinology. 2004;29:705–711. doi: 10.1016/S0306-4530(03)00100-8. [DOI] [PubMed] [Google Scholar]
- 27.Buske-Kirschbaum A, Ebrecht M, Hellhammer D. Blunted HPA axis responsiveness to stress in atopic patients is associated with the acuity and severeness of allergic inflammation. Brain, behavior, and immunity. 2010;24:1347–1353. doi: 10.1016/j.bbi.2010.06.013. [DOI] [PubMed] [Google Scholar]
- 28.Amano H, Negishi I, Akiyama H, Ishikawa O. Psychological stress can trigger atopic dermatitis in NC/Nga mice: an inhibitory effect of corticotropin-releasing factor. Neuropsychopharmacology. 2008;33:566–573. doi: 10.1038/sj.npp.1301435. [DOI] [PubMed] [Google Scholar]
- 29.Jiang J, et al. Oral administration of Yokukansan inhibits the development of atopic dermatitis-like lesions in isolated NC/Nga mice. Journal of dermatological science. 2009;56:37–42. doi: 10.1016/j.jdermsci.2009.07.003. [DOI] [PubMed] [Google Scholar]
- 30.Hastings PD, Fortier I, Utendale WT, Simard LR, Robaey P. Adrenocortical functioning in boys with attention-deficit/hyperactivity disorder: examining subtypes of ADHD and associated comorbid conditions. Journal of abnormal child psychology. 2009;37:565. doi: 10.1007/s10802-008-9292-y. [DOI] [PubMed] [Google Scholar]
- 31.Abelson JL, Curtis GC, Cameron OG. Hypothalamic-pituitary-adrenal axis activity in panic disorder: effects of alprazolam on 24 h secretion of adrenocorticotropin and cortisol. Journal of psychiatric research. 1996;30:79–93. doi: 10.1016/0022-3956(95)00035-6. [DOI] [PubMed] [Google Scholar]
- 32.Eto K, Mazilu-Brown JK, Henderson-MacLennan N, Dipple KM, McCabe ER. Development of catecholamine and cortisol stress responses in zebrafish. Molecular genetics and metabolism reports. 2014;1:373–377. doi: 10.1016/j.ymgmr.2014.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Feenstra MG, Kalsbeek A, Van Galen H. Neonatal lesions of the ventral tegmental area affect monoaminergic responses to stress in the medial prefrontal cortex and other dopamine projection areas in adulthood. Brain research. 1992;596:169–182. doi: 10.1016/0006-8993(92)91545-P. [DOI] [PubMed] [Google Scholar]
- 34.Henry C, et al. Prenatal stress in rats facilitates amphetamine-induced sensitization and induces long-lasting changes in dopamine receptors in the nucleus accumbens. Brain research. 1995;685:179–186. doi: 10.1016/0006-8993(95)00430-X. [DOI] [PubMed] [Google Scholar]
- 35.Pani L, Porcella A, Gessa G. The role of stress in the pathophysiology of the dopaminergic system. Molecular psychiatry. 2000;5:14. doi: 10.1038/sj.mp.4000589. [DOI] [PubMed] [Google Scholar]
- 36.Daubner SC, Le T, Wang S. Tyrosine hydroxylase and regulation of dopamine synthesis. Archives of biochemistry and biophysics. 2011;508:1–12. doi: 10.1016/j.abb.2010.12.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Darcq E, Kieffer BL. PI3K signaling in the locus coeruleus: a new molecular pathway for ADHD research. EMBO molecular medicine. 2015;7:859–861. doi: 10.15252/emmm.201505266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Boesgaard S, Hagen C, Andersen AN, Fenger M, Eldrup E. Effect of dopamine, dopamine D-1 and D-2 receptor modulation on ACTH and cortisol levels in normal men and women. Acta endocrinologica. 1990;122:29–36. doi: 10.1530/acta.0.1220029. [DOI] [PubMed] [Google Scholar]
- 39.Petrossians P, Thonnard A-S, Beckers A. Medical treatment in Cushing’s syndrome: dopamine agonists and cabergoline. Neuroendocrinology. 2010;92:116–119. doi: 10.1159/000317716. [DOI] [PubMed] [Google Scholar]
- 40.Buske-Kirschbaum A, Geiben A, Höllig H, Morschhäuser E, Hellhammer D. Altered responsiveness of the hypothalamus-pituitary-adrenal axis and the sympathetic adrenomedullary system to stress in patients with atopic dermatitis. The Journal of Clinical Endocrinology & Metabolism. 2002;87:4245–4251. doi: 10.1210/jc.2001-010872. [DOI] [PubMed] [Google Scholar]
- 41.Park G, Oh D-S, Lee MG, Lee CE, Kim Y-u. 6-Shogaol, an active compound of ginger, alleviates allergic dermatitis-like skin lesions via cytokine inhibition by activating the Nrf2 pathway. Toxicology and applied pharmacology. 2016;310:51–59. doi: 10.1016/j.taap.2016.08.019. [DOI] [PubMed] [Google Scholar]
- 42.Park G, et al. Coriander alleviates 2, 4-dinitrochlorobenzene-induced contact dermatitis-like skin lesions in mice. Journal of medicinal food. 2014;17:862–868. doi: 10.1089/jmf.2013.2910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Neto, F. K., Noschang, R. & Nunes, M. L. The relationship between epilepsy, sleep disorders, and attention deficit hyperactivity disorder (ADHD) in children: A review of the literature. Sleep Science (2016). [DOI] [PMC free article] [PubMed]
- 44.Heijden KB, Smits MG, Gunning WB. Sleep hygiene and actigraphically evaluated sleep characteristics in children with ADHD and chronic sleep onset insomnia. Journal of sleep research. 2006;15:55–62. doi: 10.1111/j.1365-2869.2006.00491.x. [DOI] [PubMed] [Google Scholar]
- 45.Corkum P, Tannock R, Moldofsky H. Sleep Disturbances in Children With Attention‐Deficit/Hyperactivity Disorder. Journal of the American Academy of Child & Adolescent Psychiatry. 1998;37:637–646. doi: 10.1097/00004583-199806000-00014. [DOI] [PubMed] [Google Scholar]
- 46.Van der Heijden KB, Smits MG, Someren EJV, Boudewijn Gunning W. Idiopathic Chronic Sleep Onset Insomnia in Attention‐Deficit/Hyperactivity Disorder: A Circadian Rhythm Sleep Disorder. Chronobiology international. 2005;22:559–570. doi: 10.1081/CBI-200062410. [DOI] [PubMed] [Google Scholar]
- 47.Konofal E, Lecendreux M, Cortese S. Sleep and ADHD. Sleep medicine. 2010;11:652–658. doi: 10.1016/j.sleep.2010.02.012. [DOI] [PubMed] [Google Scholar]
- 48.Ali N, Pitson D, Stradling J. Sleep disordered breathing: effects of adenotonsillectomy on behaviour and psychological functioning. European journal of pediatrics. 1996;155:56–62. doi: 10.1007/BF02309859. [DOI] [PubMed] [Google Scholar]
- 49.Walters AS, et al. Dopaminergic therapy in children with restless legs/periodic limb movements in sleep and ADHD. Pediatric neurology. 2000;22:182–186. doi: 10.1016/S0887-8994(99)00152-6. [DOI] [PubMed] [Google Scholar]
- 50.van Geijlswijk IM, Korzilius HP, Smits MG. The use of exogenous melatonin in delayed sleep phase disorder: a meta-analysis. Sleep. 2010;33:1605–1614. doi: 10.1093/sleep/33.12.1605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Fatima G, Sharma V, Verma N. Circadian variations in melatonin and cortisol in patients with cervical spinal cord injury. Spinal cord. 2016;54:364–367. doi: 10.1038/sc.2015.176. [DOI] [PubMed] [Google Scholar]
- 52.Duffy JF, et al. Peak of circadian melatonin rhythm occurs later within the sleep of older subjects. American Journal of Physiology-Endocrinology And Metabolism. 2002;282:E297–E303. doi: 10.1152/ajpendo.00268.2001. [DOI] [PubMed] [Google Scholar]
- 53.Lewy AJ, Ahmed S, Jackson JML, Sack RL. Melatonin shifts human orcadian rhythms according to a phase-response curve. Chronobiology international. 1992;9:380–392. doi: 10.3109/07420529209064550. [DOI] [PubMed] [Google Scholar]
- 54.Slominski AT, et al. Metabolism of melatonin in the skin: Why is it important? Experimental dermatology. 2017;26:563–568. doi: 10.1111/exd.13208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Smith SM, Vale WW. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. Dialogues in clinical neuroscience. 2006;8:383. doi: 10.31887/DCNS.2006.8.4/ssmith. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Bornstein SR, Engeland WC, Ehrhart-Bornstein M, Herman JP. Dissociation of ACTH and glucocorticoids. Trends in Endocrinology & Metabolism. 2008;19:175–180. doi: 10.1016/j.tem.2008.01.009. [DOI] [PubMed] [Google Scholar]
- 57.Pravosudov VV. Long-term moderate elevation of corticosterone facilitates avian food-caching behaviour and enhances spatial memory. Proceedings of the Royal Society of London B: Biological Sciences. 2003;270:2599–2604. doi: 10.1098/rspb.2003.2551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Slominski A, Wortsman J, Tuckey R, Paus R. Differential expression of HPA axis homolog in the skin. Mol Cell Endocrinol. 2007;265–266:143–149. doi: 10.1016/j.mce.2006.12.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Slominski A, Manna P, Tuckey R. On the role of skin in the regulation of local and systemic steroidogenic activities. Steroids. 2015;103:72–88. doi: 10.1016/j.steroids.2015.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Slominski A, et al. Key role of CRF in the skin stress response system. Endocr Rev. 2013;34:827–884. doi: 10.1210/er.2012-1092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Slominski A, et al. A novel pathway for sequential transformation of 7-dehydrocholesterol and expression of the P450scc system in mammalian skin. Eur J Biochem. 2004;271:4178–4188. doi: 10.1111/j.1432-1033.2004.04356.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Slominski A, et al. Steroidogenesis in the skin: implications for local immune functions. J Steroid Biochem Mol Biol. 2013;137:107–123. doi: 10.1016/j.jsbmb.2013.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Slominski A, Manna P, Tuckey R. Cutaneous glucocorticosteroidogenesis: securing local homeostasis and the skin integrity. Exp Dermatol. 2014;23:369–374. doi: 10.1111/exd.12376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Slominski A, Brożyna A, Tuckey R. Cutaneous Glucocorticoidogenesis and Cortisol Signaling Are Defective in Psoriasis. J Invest Dermatol. 2017;137:1609–1611. doi: 10.1016/j.jid.2017.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Slominski, A. T. et al. Melatonin: a cutaneous perspective on its production, metabolism, and functions. Journal of Investigative Dermatology (2018). [DOI] [PMC free article] [PubMed]
- 66.Karasek M. Melatonin, human aging, and age-related diseases. Experimental gerontology. 2004;39:1723–1729. doi: 10.1016/j.exger.2004.04.012. [DOI] [PubMed] [Google Scholar]
- 67.Gubin DG, Gubin GD, Gapon LI, Weinert D. Daily melatonin administration attenuates age-dependent disturbances of cardiovascular rhythms. Current aging science. 2016;9:5–13. doi: 10.2174/1874609809666151130220011. [DOI] [PubMed] [Google Scholar]
- 68.Carretero M, et al. Long‐term melatonin administration protects brain mitochondria from aging. Journal of pineal research. 2009;47:192–200. doi: 10.1111/j.1600-079X.2009.00700.x. [DOI] [PubMed] [Google Scholar]
- 69.Chang Y-S, et al. Atopic dermatitis, melatonin, and sleep disturbance. Pediatrics. 2014;134:e397–e405. doi: 10.1542/peds.2014-0376. [DOI] [PubMed] [Google Scholar]
- 70.Slominski AT, et al. Melatonin, mitochondria, and the skin. Cellular and Molecular Life Sciences. 2017;74:3913–3925. doi: 10.1007/s00018-017-2617-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Slominski A, Tobin DJ, Zmijewski MA, Wortsman J, Paus R. Melatonin in the skin: synthesis, metabolism and functions. Trends in Endocrinology & Metabolism. 2008;19:17–24. doi: 10.1016/j.tem.2007.10.007. [DOI] [PubMed] [Google Scholar]
- 72.Zhang T, Yan L, Ma S, He J. Human biological rhythm in traditional Chinese medicine. Journal of Traditional Chinese Medical Sciences. 2016;3:206–2011. doi: 10.1016/j.jtcms.2016.12.004. [DOI] [Google Scholar]
- 73.Benarroch EE. The locus ceruleus norepinephrine system Functional organization and potential clinical significance. Neurology. 2009;73:1699–1704. doi: 10.1212/WNL.0b013e3181c2937c. [DOI] [PubMed] [Google Scholar]
- 74.Ramos BP, Arnsten AF. Adrenergic pharmacology and cognition: focus on the prefrontal cortex. Pharmacology & therapeutics. 2007;113:523–536. doi: 10.1016/j.pharmthera.2006.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Chiew, K. S. & Braver, T. S. Positive affect versus reward: emotional and motivational influences on cognitive control. Frontiers in psychology2 (2011). [DOI] [PMC free article] [PubMed]
- 76.Pacak K, et al. Effects of various stressors on in vivo norepinephrine release in the hypothalamic paraventricular nucleus and on the pituitary‐adrenocortical axis. Annals of the New York Academy of Sciences. 1995;771:115–130. doi: 10.1111/j.1749-6632.1995.tb44675.x. [DOI] [PubMed] [Google Scholar]
- 77.Mizoguchi K, Shoji H, Ikeda R, Tanaka Y, Tabira T. Persistent depressive state after chronic stress in rats is accompanied by HPA axis dysregulation and reduced prefrontal dopaminergic neurotransmission. Pharmacology Biochemistry and Behavior. 2008;91:170–175. doi: 10.1016/j.pbb.2008.07.002. [DOI] [PubMed] [Google Scholar]
- 78.Kwon M-S, et al. The effect of single or repeated restraint stress on several signal molecules in paraventricular nucleus, arcuate nucleus and locus coeruleus. Neuroscience. 2006;142:1281–1292. doi: 10.1016/j.neuroscience.2006.07.027. [DOI] [PubMed] [Google Scholar]
- 79.Trentani A, Kuipers S, Ter Horst G, Den Boer J. Selective chronic stress‐induced in vivo ERK1/2 hyperphosphorylation in medial prefrontocortical dendrites: Implications for stress‐related cortical pathology? European Journal of Neuroscience. 2002;15:1681–1691. doi: 10.1046/j.1460-9568.2002.02000.x. [DOI] [PubMed] [Google Scholar]
- 80.Qi X, Lin W, Li J, Pan Y, Wang W. The depressive-like behaviors are correlated with decreased phosphorylation of mitogen-activated protein kinases in rat brain following chronic forced swim stress. Behavioural brain research. 2006;175:233–240. doi: 10.1016/j.bbr.2006.08.035. [DOI] [PubMed] [Google Scholar]
- 81.Zeng C, Jose PA. Dopamine Receptors. Hypertension. 2011;57:11–17. doi: 10.1161/HYPERTENSIONAHA.110.157727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Park G, Park Y-J, Yang HO, Oh MS. Ropinirole protects against 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine (MPTP)-induced neurotoxicity in mice via anti-apoptotic mechanism. Pharmacology Biochemistry and Behavior. 2013;104:163–168. doi: 10.1016/j.pbb.2013.01.017. [DOI] [PubMed] [Google Scholar]
- 83.Prince J. Catecholamine dysfunction in attention-deficit/hyperactivity disorder: an update. Journal of clinical psychopharmacology. 2008;28:S39–S45. doi: 10.1097/JCP.0b013e318174f92a. [DOI] [PubMed] [Google Scholar]
- 84.Hamner MB, Gold PB. Plasma dopamine beta-hydroxylase activity in psychotic and non-psychotic post-traumatic stress disorder. Psychiatry research. 1998;77:175–181. doi: 10.1016/S0165-1781(98)00002-X. [DOI] [PubMed] [Google Scholar]
- 85.McMahon A, et al. Regulation of Tyrosine Hydroxylase and Dopamine β‐Hydroxylase mRNA Levels in Rat Adrenals by a Single and Repeated Immobilization Stress. Journal of neurochemistry. 1992;58:2124–2130. doi: 10.1111/j.1471-4159.1992.tb10954.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.