

Abstract
On the 150th anniversary of Korbinian Brodmann’s birth, and the 100th anniversary of his death, Zilles celebrates his pioneering role in brain mapping. With the aid of hitherto unpublished documents and figures, he explains the concepts behind Brodmann’s cytoarchitectonic maps and considers their impact on current neuroimaging approaches.
Introduction
The year 2018 is the 150th anniversary of Korbinian Brodmann’s birth (17 November 1868) as well as the 100th anniversary of his death (22 August 1918). Given the historical, scientific and enduring importance of his work for the field of brain mapping, the dual anniversary is the right occasion to remember the life and scientific work of this pioneer (Fig. 1).
Figure 1.

Korbinian Brodmann (17 November 1868–22 August 1918). With permission of the C. & O. Vogt Archive, Institute of Brain Research, University Düsseldorf.
A cited reference search in the Web of Science carried out in July 2018 resulted in over 170 000 citations of Brodmann’s work, mainly of his monography (Brodmann, 1909) (for a comprehensive list of all major publications by Brodmann and a short description of their content, see Supplementary Table 1). His publications on the cytoarchitectonic parcellation of the entire human cerebral cortex made him a founder of the field of anatomical brain mapping. The number of publications with references to different versions of his maps (Brodmann, 1908a, 1909, 1910, 1912, 1914) dramatically increased since the advent of neuroimaging using PET and MRI, and is still increasing (Fig. 2B). The maps have become particularly popular in recent times for localization of activations using functional MRI and for meta-analyses of structural and functional relationships. The maps are also the fundament for freely available mapping tools (e.g. www.fmriconsulting.com/brodmann/Introduction.html), and frequently used atlases (e.g. Talairach and Tournoux, 1988), in which his 2D schematic drawing of the cytoarchitectonic subdivisions of the cerebral cortex has been tentatively registered as a 3D representation. Since two-thirds of the cortical surface are hidden in the sulci, and Brodmann, as well as the other authors, did not show the precise position of intrasulcal boundaries of the cortical areas in the maps, and only occasionally described their positions in the text or figures (Zilles and Amunts, 2010), all such transformations from the 2D drawings to a 3D reference brain will remain questionable and are based more on subjective assumptions than real evidence from Brodmann’s work.
Figure 2.

Influence of Brodmann's monography and map on modern day neuroimaging studies. (A) Title page of the famous monography (Brodmann, 1909) and the final version of the map of the entire human cerebral cortex (Brodmann, 1910). (B) Development of the citations of Brodmann’s work up to the year 2017. Source: Web of Science.
Nowadays, Brodmann’s maps dominate his legacy, showing 48 cortical areas of the human cerebral cortex (Fig. 2A), and some further subdivisions in the later modifications of this map (Brodmann, 1910, 1914). For detailed comparisons of the different versions see Judas et al. (2012). His theoretical concepts of the organizational principles and evolution of the cortex, however, as well as their potential functional implications are largely ignored, although they are at least equally important parts of his legacy. Despite an English translation of the monography by Laurence Garey (1994), the theoretical concepts described therein are often not recognized and thus ‘rediscovered’ in modern publications. Only the schematic maps of the human and non-human primate brains are widely used. His early original publications (Brodmann, 1903a, b, 1905a, b, 1906, 1908a, b) contain even more detailed and important aspects for anatomical brain mapping than the reduction to schematic maps.
The present publication aims to remember Brodmann’s life and work, his influence on, and importance for, actual research concepts in various fields of modern brain mapping. It contains among others hitherto unpublished illustrations (photos and drafts of drawings with handwritten inscriptions by him) generously provided by the Archive of the Cécile and and Oskar Vogt Institute of Brain Research, University of Düsseldorf, Germany and the Korbinian-Brodmann-Museum, Hohenfels-Liggersdorf, Germany.
The early years of Brodmann
Korbinian Brodmann’s life was full of tragedies, difficult social and career problems, but also unexpected lucky coincidences. His life and work seem to be a converging lens that clearly focuses the historical situation in neuroscience and the appearance of general theories on the organization of the cerebral cortex at the beginning of the 20th century.
The difficulties started with the social conditions of his childhood. He was born in Liggersdorf, a small village in Southern Germany and part of the principality of Hohenzollern-Sigmaringen. He was the illegitimate son of Sophie Benkler (Fig. 3A), the daughter of a very poor farmer (Fig. 3D), who had to work as a maidservant in the house (Fig. 3C) of the wealthy Brodmann family. His father, Josef Brodmann (Fig. 3B), did not marry Sophie until 1886, when Korbinian was 18 years old. From this time on, his family name was Brodmann. His Christian name Korbinian was given to him by the priest of the village—a usual custom at this time used for illegitimate children.
Figure 3.

Photographs portraying Brodmann's life as a child and a young adult. Korbinian Brodmann’s mother Sophie Benkler (A) and his father Josef Brodmann (B). Hofgut of the Brodmann family (C) and his birthplace, the farming house of the Benkler family (D). With permission of the Brodmann Museum, Hohenfels-Liggersdorf. Oskar Vogt (E) at the age of 33 years. Painting by W. Döring, 1905. With permission of the C. & O. Vogt Archive, Institute of Brain Research, University Düsseldorf. Alexandersbad (F) around the time of the first encounter of Vogt and Brodmann. With permission of the Brodmann Museum, Hohenfels-Liggersdorf.
After 6 years in basic school at his birthplace Liggersdorf, he went to a secondary school in Überlingen for 2 years, to the gymnasium in Sigmaringen for 1 year, and finally to the gymnasium in Konstanz for 6 years. He finished his higher school education at the gymnasium in Konstanz, although his school teacher wrote in the class book of the basic school in Liggersdorf: ‘although he is weakly capable, he went on to the high school’.
After gymnasium, he studied medicine between 1889 and 1890 in Munich, then in Würzburg (1890–91), where he attended a lecture on medical physics delivered by Wilhelm Conrad Röntgen (1845–1923), the discoverer of the X-rays and first recipient of the Nobel Prize for Physics. Berlin (1891–92) was the next station of the student, where he attended—besides his lectures and courses in medicine—lectures on Immanuel Kant’s philosophy, which led to a lifelong interest in his work. Finally, he studied in Freiburg from 1892 to 1895, where he passed the final exam and received permission to work as a physician (Approbation; 21 February 1895). Immediately after approbation he worked for some weeks as a general practitioner in Wehr in the Black Forest. He then moved to Lausanne for further education in medicine by attending clinical lectures, and then to Munich as a physician in the so-called Reisnerianum, a hospital for sick children. Brodmann also attended the lectures of Hubert von Grashey (1839–1914) in psychiatry during the winter semester of 1895/1896.
A fateful encounter
The year 1896 brought a fateful encounter for Brodmann’s scientific future. He had planned to recover from a diphtheria infection in Alexandersbad, a spa in the Spruce Mountains (Fig. 3F). Here he met Oskar Vogt (Fig. 3E). The young physician Vogt had just returned from his studies in neurology and hypnosis with the famous French neurologist Joseph Jules Dejerine (1849–1917) (Supplementary Fig. 1) in Paris. Oskar Vogt (1870–1959) offered him a position as a physician in the psychiatric unit of Alexandersbad. Vogt led this unit during the summer season. The flamboyant personality of Vogt and his ideas about the future of brain research apparently fascinated the 2-year older Brodmann, who was described as ‘an intense and earnest man, reserved to the point of timidity, but could flare, on occasion, into a temper’ (Rose, 1979). Vogt finally convinced Brodmann to accept the position in Alexandersbad and to specialize in psychiatry and hypnosis.
Brodmann spent the winter semester of 1896/97 in Berlin to improve his knowledge in hypnosis, neuropathology and neuroanatomy. He then moved to Leipzig, met the psychologist Wilhelm Wundt (1832–1920) (Supplementary Fig. 1B), who is recognized as the father of experimental psychology, and the psychiatrist Paul Flechsig (1847–1929) (Supplementary Fig. 1C), who analysed myelogenesis, i.e. the appearance of myelinated fibre tracts during ontogeny. In Leipzig, Brodmann wrote a letter to Vogt reporting on his skills in hypnosis:
‘Miss [NN] is slowly but steadily improving. Her anxiety, from which she has suffered for already two years and which could not be successfully treated by Flechsig … and others, disappeared after five months of hypnosis by me, and she is definitely cured from her anxiety. However, the patient is not completely healthy because she suffers from depression and many somatic problems.’ (letter without signature in the Cécile and Oskar Vogt Archive, C. & O. Vogt Institute for Brain Research, University Düsseldorf).
During the period in Leipzig, Brodmann finished his MD thesis on ‘Chronic Sclerosis of Ependyma’ (Brodmann, 1898a).
Let us briefly return to Alexandersbad because of its importance in the fate of Brodmann and Vogt. Alexandersbad was the most fashionable spa for upper class people with real or imagined diseases. Vogt, as head of the psychiatric unit at this spa, successfully treated members of the Krupp family (Supplementary Fig. 1D–G). From this time on, he became the personal physician of the Krupps, who owned the world’s largest industrial complex producing steel, warships, tanks, and cannons. Friedrich Alfred Krupp, the so-called ‘cannon king’, had a great influence on Emperor Wilhelm II, and was instrumental in arming the German troops before World War I. Vogt remained the personal physician of Gustav Krupp von Bohlen and Halbach (1870–1950) and his wife Bertha (1886–1957), who followed Friedrich Alfred Krupp not only as the head of the Krupp company, but also with enormous political influence during the Third Reich. Vogt’s relationship with the Krupp family was extremely important for Brodmann’s and Vogt’s scientific future, because Brodmann went on to receive a postdoctoral position in the so-called ‘Neurobiological Central Station’ in Berlin (Fig. 4A). It had been founded by Vogt in 1901, was located in the rented first to third floors of a private house, and consisted of a medical practice for hypnosis on the first floor, a research laboratory on the second floor, and animal and photographic facilities on the third floor. The ‘Neurobiological Central Station’ was a completely private institution financially supported by the Krupp family and Vogt’s private income as a physician. The ‘Station’ was founded against the resistance of most of the academic establishment at the University of Berlin. Here, Brodmann started his pioneering work on the cytoarchitecture of mammalian brains. However, before he joined Vogt in Berlin, he spent another 2 years in Jena and Frankfurt.
Figure 4.

Photographs portraying Brodmann's time in Berlin. (A) ‘Neurobiological Central Station’, Berlin, Magdeburgerstr. 16. (B–D) Korbinian Brodmann during his time in Vogt’s ‘Neurobiologischem Laboratorium’ in Berlin. With permission of the C. & O. Vogt Archive, Institute of Brain Research, University Düsseldorf.
Evolutionary theories and neuropathology: the theoretical and practical basis of Brodmann’s work
Brodmann planned to improve his experience in psychiatry and neuroanatomy during this period (1898 to 1900). More important for Brodmann’s scientific development than his activity as a physician in the psychiatric hospital of the University in Jena, however, was the fact that Jena was the home of Ernst Haeckel (1834–1919) (Supplementary Fig. 2A). He was a physician, marine biologist, philosopher and artist and the most prominent fighter in Germany for Darwin’s theory. His wonderful paintings of the radiolarians, calcareous, medusas and siphonophores were admired by the scientific world (Supplementary Fig. 2B) and had a great influence on the art around the turn of the 19–20th centuries, i.e. the art nouveau (‘Jugendstil’). His influence on the entire spectrum of life science and its promotion in Germany cannot be overestimated. Thus, Brodmann was immersed in this world of Darwinian ideas. His efforts in comparative neuroanatomy and embryology to understand the organization of the structure in the adult brain reflect Haeckel’s influence on his work during the following 20 years. One of Haeckel’s most famous statements is that ontogeny recapitulates phylogeny. Brodmann’s search for homologous cortical layers and areas in the brains of various mammals and his studies of embryonic and foetal stages of brain development are reflections of his time in Jena, and served him as a signpost in the jungle of different structures in the adult brain.
During 1900–01, Brodmann worked as a physician at the Municipal Mental Asylum in Frankfurt, where he met Ludwig Edinger (1855–1918) (Supplementary Fig. 2C), one of the founders of comparative neuroanatomy. Based on his studies of ‘old’ and ‘newly acquired’ parts of the human brain, Edinger coined the terms ‘Palaeencephalon’ and ‘Neencephalon’ for those parts of the brain that appear early or late, respectively, during brain evolution. Brodmann (1909) and Vogt and Vogt (1919) adapted this distinction in their cyto- and myeloarchitectonic studies, and introduced the terms ‘neocortex’ equivalent to isocortex, and ‘palaeocortex’ equivalent to the olfactory part of the allocortex. How much Brodmann appreciated the work of Edinger is highlighted by the fact that Brodmann asked Edinger for a critical review and agreement before he would publish his ontology of major cortical subdivisions.
The most important contact for Brodmann’s further career was the chance to meet the neuropathologist Alois Alzheimer (1864–1915) (Supplementary Fig. 2D) in Frankfurt. Alzheimer explicitly supported Vogt’s former advice: Brodmann should seek his future as a neuroanatomist in psychiatry. He apparently convinced him, because Brodmann accepted the offer by Vogt to join in the newly founded ‘Neurobiological Central Station’ in Berlin (Fig. 4B–D). When he later applied for a new position after his time in Berlin and Tübingen, Brodmann described this most important decision for his scientific work in his austere and prosaic style: ‘From August 1901 onwards I was assistant at the neurobiological laboratory of the University of Berlin, where I served under Doctor O. Vogt until my entry into my current position as assistant physician of the Royal University Clinic for Mental and Nervous Conditions on October 1st 1910.’ (translation of a hand-written curriculum vitae of Brodmann provided by the Brodmann Museum).
Localization in the cerebral cortex: the years with Oskar and Cécile Vogt in Berlin
In 1902, the ‘Neurobiological Central Station’ changed its name into ‘Neurobiological Laboratory of the University of Berlin’ in a process initiated by letters of Vogt and Friedrich Alfred Krupp. It was not a simple renaming, but a revolution in Berlin’s academic structure, because Vogt had started with a private laboratory completely outside the university structure. Now, this unusual and suspected ‘Central Station’ was to be transformed into an official unit and independent department of the University of Berlin. Vogt immediately faced the furious resistance of the majority of the professors in Berlin, amongst them the anatomist Wilhem von Waldeyer, who were encouraged by the well-recognized scientist and director of the University Hospital for Psychiatry in Leipzig, Paul Flechsig. In the past, Flechsig had supervised both Vogt and Brodmann. During Vogt’s time in Flechsig’s department in Leipzig, in 1894, Vogt accused Flechsig of making data, from Vogt’s then unpublished dissertation, public. The young physician insulted the mighty Professor Flechsig by calling him ‘a great scoundrel’ (Letter of Oskar Vogt to Auguste Forel. cited by Peiffer, 2013, p. 229). This was the beginning of a life-long enmity. Thus, Flechsig alarmed his friends at the University in Berlin when the private ‘Central Station’ was to be transformed into an official department of the university. Flechsig wrote in an official derogatory letter, which was forwarded to the Ministry of Culture, that Oskar Vogt was ‘a pathological personality, a degenerate … in the field of brain research a nearly ridiculous figure, immensely conceited, without any deeper knowledge … a ridiculous tattler and chatterbox’ [own translation of a document in Geheimes Staatsarchiv, Preußischer Kulturbesitz (GStA PK) Berlin: Rep. I,76 Va, Sekt. 2, Tit X, Nr. 11, Bd. XII, Adhib. I. betr. Neuro-Biologisches Laboratorium]. Despite all protest of the university, the imperial ministry decided to install the ‘Neurobiological Laboratory of the University of Berlin’, under the leadership of Oskar Vogt. The decision was made following intervention by Friedrich Alfred Krupp, the friend of Emperor Wilhelm II, and Vogt’s reliable supporter since the days of Alexandersbad. Vogt won the battle but acquired new enemies. I have recounted this episode, because this long-lasting animosity is the reason behind the later rejection of Brodmann’s habilitation thesis. The foundation of the new department had, however, a positive effect on Brodmann because, for the first time, he received a position financially secured by the state.
The years between 1901 and 1910 were the period during which Brodmann’s work on cytoarchitecture and localization was created. His work was not exclusively focused on cytoarchitecture. Between 1897 and 1907, he published 14 papers on hypnosis (Brodmann, 1897, 1898b, 1902), astrocytes (Brodmann, 1899), neuropathology (Brodmann, 1900; Bielschowsky and Brodmann, 1905), psychopathology (Brodmann, 1902, 1902–1903a, 1904), polarization microscopy of myelinated nerve fibres (Brodmann, 1901), brain activity and blood flow (Brodmann, 1902–1903b), fibrillogeny and myelogenesis (Brodmann, 1907), and hundreds of printed reviews of actual publications of other authors. He was a very critical reviewer, and his reviews increased the number of his opponents. However, his most influential studies are those on cytoarchitecture and the organization of the cerebral cortex. In a series of eight papers (Brodmann, 1903a, b, 1905a, b, 1906, 1908a, b, c), he founded—together with his contemporary, Campbell—not only the field of cytoarchitectonic mapping of the entire cortex, but also provided fundamental insights into the structural organization of the cerebral cortex, its composition by different cell types, the differences between iso- and allocortex, phylogenetic aspects in brains of different species, and, finally, created the concept of structural homologies of cortical regions in mammals.
The commercially available technical equipment necessary for the ambitious plans of Brodmann were to a large degree inadequate when he started his collaboration with Vogt. A large microtome had to be designed and manufactured by themselves, embedding and staining procedures had to be improved, and novel photographic techniques created, in order to prepare complete series of sections (each about 20–30 μm thick) of entire human hemispheres (up to several thousand sections per hemisphere). The histological slides produced were of superior quality (for cell body staining, Nissl’s original staining procedure with methylene blue was later replaced by cresyl violet) were greatly admired by Campbell (1905). How perfectly they developed the laboratory work can still be seen on the original micrographs (Figs 5 and 6).
Figure 5.
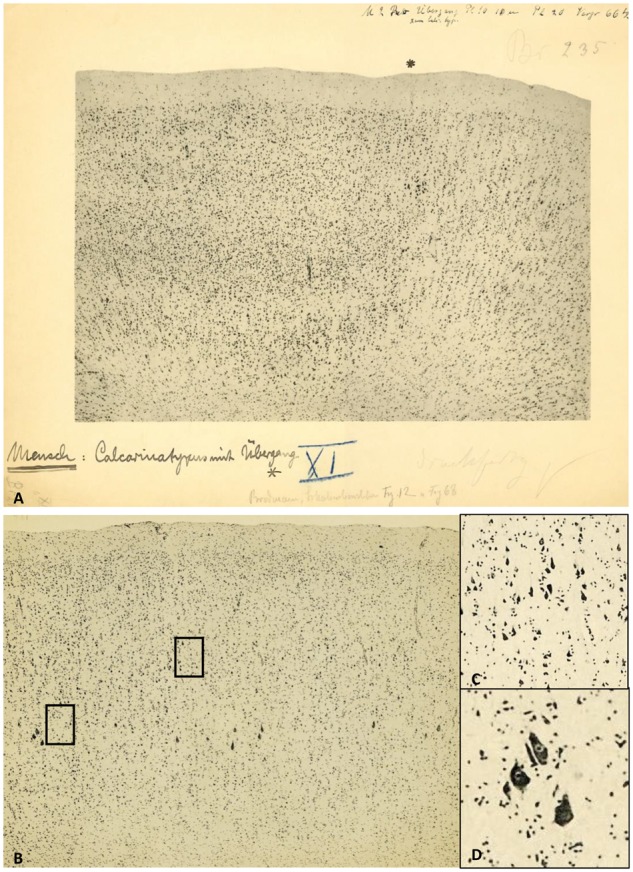
Micrographs of the human visual cortex with annotations by Brodmann. (A) Border (asterisk) between the primary (BA17, ‘Calcarinatypus’, right) and secondary (BA18, left) visual cortex of a human brain. Original micrograph with hand-written notes by K. Brodmann. (B) Brodmann’s area 4 (primary motor cortex) with Betz giant pyramidal cells (left). Note the excellent quality of the photographic technique in the magnified details (inserts C and D on the right; their positions are indicated as rectangles in the original micrograph). With permission of the C. & O. Vogt Archive, Institute of Brain Research, University Düsseldorf.
Figure 6.

Original micrographs of human cortical areas by K. Brodmann (specimen M15). Transcriptions of Brodmann’s hand writing: (A) Area occipitalis BA18, secondary visual cortex at the transition to BA17. (B) Praeoccipitalis BA19 (higher visual cortex). (C) Area parietalis sup. (precuneus ant) BA7. (D) Area parietalis inf. ant = supramarg BA40, inferior parietal cortex. (E) Area pariet. post. inf = angularis BA39, temporo-occipital cortex. (F) Transition from BA4 to BA3, (primary motor to primary somatosensory cortex). (G) Area postcentralis oralis BA3, primary somatosensory cortex. (H) Area frontalis agranularis BA6, premotor cortex. Bottom: Higher magnifications of the original micrographs (A–H) demonstrate the larger (or at least equally large) pyramidal cells in layer III compared with those cells in layer V. This cytoarchitectonic feature is called ‘externopyramidization’ and typical for higher unimodal (BA18 as an example in the visual system) and multimodal (examples BA7 and BA39) areas. In contrast, the primary motor (BA4), somatosensory (BA3), and visual (BA17) cortices display larger pyramidal cells in layer V than III. Note the wider layer IV in primary sensory (BA3 and BA17) than multimodal (BA7 and BA39) areas, and the lack of a clearly visible layer IV in the primary motor area (BA4). With permission of the C. & O. Vogt Archive, Institute of Brain Research, University Düsseldorf.
Cytoarchitectonics and the universal feature six layers
Brodmann’s monograph from 1909 is a summary of his concept of cytoarchitecture. However, his so often copied and schematic 2D drawing of the human cortical map dominates the knowledge of Brodmann’s work to such a degree, that the text of the monograph and the original publications are almost forgotten, although an English translation of the monograph is available (Garey, 1994). Brodmann describes in its first part, the principles of his comparative neuroanatomical approach, which led to the definition of homologue cortical layers and areas in various mammalian species. Furthermore, the studies of foetal brains enabled the principal distinction between hetero- and homogenetic, i.e. allo- and neocortical, regions (developmental approach). In the second part, the parcellation of the entire cerebral cortex into various distinct areas (Brodmann initially used the word ‘typus’ for an area) (Brodmann’s hand-written remarks in Figs 5–7) is comprehensively displayed by micrographs and maps of the human brain and those of various mammalian species. In the third part, he critically discusses the problem of assignment of functions to histologically defined areas.
Brodmann criticizes previous concepts to explain the functional meaning of cortical organization by focusing on the occurrence of single cell types. He states:
‘Not only is it not proven, but it is highly unlikely on general biological considerations, that a special sensory function is related to a cell type of particular structure. The essential for the elaboration of any cortical function, even the most primitive sensory perception, is not the individual cell type but cell groupings.’ (Brodmann, 1909, p. 6; this and all following citations of pages in Brodmann’s monograph are from Garey, 1994).
He clarifies this position by conceding that certain cell types may have certain physiological properties, but these functional characteristics could not be studied during his lifetime because the necessary single cell recording technique was not available.
Brodmann repeatedly expressed his respect for the work of Ramón y Cajal (1900–1906) of analysing single cell types in various cortical regions, but critically discussed Cajal’s statement that there is a simplification of the layering in lissencephalic mammals by a decrease in the number of layers, and by the loss of the inner granular layer in these species, thus forming a four- or five-layered cortex in mouse and rabbit, whereas six, seven, or up to nine layers are found in man. Brodmann wrote in the monograph:
‘In contrast to Cajal … I have for years supported the idea that … this six-layered pattern is visible in all orders, either permanently or at least as a temporary ontogenetic stage … even in those cortical zones where it disappears in the mature brain. … I maintain now as before that the original pattern for the whole mammalian order is the … six-layered type and that all variations in cortical structure are derived from this … The only exceptions are certain “rudimentary” cortical zones. In man these include on the one hand a relatively small part of the rhinencephalon [this points to the paleocortex, K.Z.], that undergoes considerable development in macrosmatic animals … and on the other hand the more or less extensive cortical regions of the cingulate gyrus adjacent to the corpus callosum, mainly around its anterior half and at the splenium [this points to the periarchicortex, K.Z.]. … these zones have an atypical laminar pattern, or at least … not … a six-layered ontogenetic transitional stage’ (Brodmann, 1909, pp. 17–18).
Brodmann’s six-layers concept is universally accepted in modern studies and textbooks. He is convinced that the analysis of cell types does not enable the parcellation of the entire cortex. Only the Betz cells represent an exemption, as they occur only in the primary motor cortex [Brodmann area (BA)4]. With his reservation regarding Cajal’s concept, he is in agreement with Campbell, who states in his monumental monograph of 1905:
‘Ramón y Cajal’s work shows blemishes … for although he describes the histology of these parts in a degree of detail which is almost bewildering, he either gives no boundaries at all, or only draws vague lines round the areas which he has examined with such extraordinary minuteness.’ (Campbell, 1905, p. 8).
However, it was not the primary goal of Cajal to define a cortical map. He was interested in the detection of single cell types, and thus, paved the way for modern cellular neuroanatomy and physiology.
Brodmann also denies the concept of layers as structural units with a unique function, the so-called stratigraphic approach represented by Ariens Kappers (1877–1947). Brodmann stated: ‘Certainly there is at first sight something attractive … in taking the characteristic and striking cortical layers … to parcellate the cortex… However, for the moment we know nothing definite about the significance of the individual layers’ (Brodmann, 1909, p. 7). His opinion that a single cortical layer cannot represent complex neuropsychological functions such as perception, cognition, motor or emotional activities still holds true, but the roles of supragranular, granular and infragranular layers are presently studied in the context of connectivity between areas and their hierarchical position. Types of connectivity are now correlated with the width of layers, the relation of the size of pyramidal cells in layers III and V, and the width and visibility of layer IV, e.g. pyramidal cells in layer III of higher sensory and multimodal areas increase in size over those in layer V (Fig. 6, also see Amunts and Zilles, 2015 and Zilles and Amunts, 2015; ‘externopyramidization’ of Goulas et al., 2018). It has been proposed that the laminar origin of long intrahemispheric connections is associated with those cytoarchitectonic features from less to more differentiated cortical areas (Sanides, 1970; Barbas, 1986, 2015; Markov et al., 2014; Goulas et al., 2018), but the methods to directly demonstrate feedforward or feedback connections of single layers were not available in Brodmann’s time.
What is a cytoarchitectonic area?
Central to Brodmann’s efforts of parcellation and localization in the cerebral cortex is the question: What is a meaningful concept of a cytoarchitectonic area? He described his approach as follows:
‘The most important … finding of our comparative topography of the cerebral cortex is that its … organization manifests a common architectonic plan in all mammals, a standardised pattern of layers. First, we were able to distinguish two types of such architectonic cortical formations, derived from (presumably) different primitive cytoarchitectures, the heterogenetic and the homogenetic types. The essential features of each of these structural types is consistently demonstrable throughout all species, with greater or lesser modifications. Further, we ascertained that in all mammals the homogenetic cortex again manifests a regular series of variations (homologous types), that themselves all stem from a common basic histological form, the original six-layered primitive tectogenetic type. Finally, we were able to show that, thanks to these homologous structural formations, it is possible to demonstrate an essentially standardised organisation of the surface of the hemispheres throughout the whole mammalian class into spatially delineated zones, or secondary cortical organs, that we denominate regions and fields (or areas).’ (Brodmann, 1909, pp. 219–20).
Supplementary Fig. 3 shows a comparison between Brodmann’s maps of four different species. The heterogenetic type is the allocortex, divided into palaeo- and archicortex, which comprise the olfactory, hippocampal and parahippocampal cortices, as well as the cingulate and retrosplenial cortices. The homogenetic type is the iso- or neocortex, subdivided into numerous areas. What supports Brodmann’s position that his statements hold true for all mammals? It is his work on not less than 64 different vertebrate species ranging from prototherians to metatherians, and many eutherian species including a large variety of non-human primates (Fig. 7A–F). He additionally collected quantitative data on the motor, prefrontal, visual and olfactory cortices during the following years (Brodmann, 1910, 1912, 1913). This comparative anatomical approach enabled Brodmann to propose and develop a combined evolutionary/cytoarchitectonic concept for the definition of areas, e.g. the unified motor-somatosensory region of insectivores (Supplementary Fig. 3D) differentiates into numerous motor, sensory and association areas in the human brain, which demonstrates a developmental gradient from insectivores to primates.
‘A topical separation into a giant pyramidal area [BA4] and an agranular frontal area [BA6] is not possible; the two spatially completely separate areas of higher species have not yet differentiated here [in the hedgehog], but form a common zone. … Rather, in our brain map caudal to the precentral region there follows a homogeneously structured zone [parietal cortex of the present Supplementary Fig. 3D] that cannot be further divided into individual areas and must wholly represent these two regions; thus in it lie the combined areas 1 to 3, 5 and 7 of other mammals [see present Supplementary Fig. 3A–C], undifferentiated.’ (Brodmann, 1909, p. 170).
‘The parietal lobe is divisible into the four areas 5, 7, 39 and 40 [see present Supplementary Fig. 3A] that correspond to only two … in most other brains, or often to only one [see present Supplementary Fig. 3D] … there undoubtedly emerges an increase in the specifically differentiated cortical mass that is manifested … by a larger number of differentiated areas; in other words, there are new additions to the cortex. That such localisational transformations of particular cortical zones should also take place in lower, less well organised species, is a proof that we are dealing here with real progressive differentiation’ (Brodmann, 1909, p. 198).
Figure 7.

Hand drawings of Brodmann (with permission of the Brodmann Museum, Hohenfels-Liggersdorf, Germany). (A) Lateral view of the left hemisphere of a platypus (Ornithorhynchus paradoxus, Monotremata). (B) Lateral view of the right hemisphere of a spiny anteater (Echidna, Monotremata). (C) Mesial surface of the right hemisphere of a common brushtail possum (Trichosurus vulpecula). ‘Am’ in the yellow circle depicts the Corpus amygdaloideum. Large black dot commissura anterior. The commissura anterior is far smaller in the human than in the marsupial brain, since it preferentially connects allocortical regions, which are large in marsupials but belong to the relatively small (relative to the isocortex) human allocortex (archi- and palaeocortex). (D) Mesial surface of the right hemisphere of the Kinkajou (Cercoleptes caudivolvolus). Fine dots label the primary visual cortex (BA17), other symbols mark the olfactory and entorhinal cortex. (E) Mesial surface of the right hemisphere of a prosimian (Lemur niger). Large black pyramids mark the area praeparietalis (BA5), filled circles the primary motor cortex (BA4), open circles the premotor cortex (BA6), dots the primary visual cortex (BA17), and the large and small crosses the cingulate cortex (par of BA24 and the whole BA23, respectively). This sketch is an early stage of Fig. 99 in the monography (Brodmann, 1909). (F) Lateral view of the left hemisphere of a kangaroo (Macropus pennicillatus). Blue contour primary motor cortex (BA4), red primary visual cortex (BA17), yellow olfactory cortex. (G) Brodmann together with Cécile and Oskar Vogt in the ‘Neurobiological Laboratory’ of the University of Berlin. From left to right Korbinian Brodmann, Cécile and Oskar Vogt, the technician Louise Bosse, and the scientific collaborators Max Lewandowski and Max Borchert. Photo taken around 1905. With permission of the C. & O. Vogt Archive, Institute of Brain Research, University Düsseldorf, Photograph No. 272.
In conclusion, a cortical area has an evolutionary history. The identification of a cytoarchitectonical area requires the demonstration of homologue structures and the modification of its laminar pattern in comparative neuroanatomical studies. Brodmann’s parcellation work and the definition of a cytoarchitectonic area is, therefore, founded on the theory of brain evolution. The aim of his studies was not a demonstration of as many subdivisions of the cortex as technically possible.
Function and cytoarchitectonics
The structural-functional relationships were a central topic in the most influential contemporary publications on the anatomy of the cerebral cortex (Meynert, 1866, 1868; Wernicke, 1874; Lichtheim, 1885; Flechsig, 1894, 1896a, b, 1901; Campbell, 1904, 1905).
Meynert’s studies are based on the analyses of fibres and their putative termination sites in the cerebral cortex, and distinguished two different types of nerve fibres: projection and association fibres (Meynert, 1866, 1868). The former convey sensory stimuli to cortical termination sites, i.e. the ‘sensorial sphere’ localized in the posterior part of the hemisphere behind the central sulcus. The association fibres exclusively connect cortical sites, and enable the linkage between sensory input, imagination, and motoric activity. The cortex was, therefore, parcellated into two functional and structural parts: a subordinate centre represented by the terminal sites of sensory subcortical afferents, and the higher centre represented by terminal sites of association fibres. Wernicke (1874) and Lichtheim (1885) further developed the ideas of Meynert in their studies of aphasic syndromes: ‘I do not consider the function to be localized in one spot of the brain, but rather to result from the combined action of the whole sensorial sphere’ (Lichtheim, 1885, p. 477).
Flechsig, the adversary of Brodmann and Vogt since their stay in Leipzig, overstrained Meynerts concept. He formulated the ‘myelogenetic basic law’ (Flechsig, 1894, 1896a, b) based on studies of myelogenesis in foetuses, newborns and children. He correctly stated that projection fibres mature early, and association fibres mature late, but he added that cortical centres are the origin and target of simultaneously maturing nerve fibres of functionally equal implication, either projection or association fibres. Thus, he introduced a sharp distinction between both types of fibres and their associated centres. ‘Since these sensorial spheres [the termination fields of the projection fibres, K.Z.] … represent only one and the smaller part of the cerebral cortex, large parts remain, in which projection systems can never be detectable.’ (Flechsig, 1927, p. 84). He described 35 centres in three large association regions of the cortex (Supplementary Figs 4 and 5). The association centres are ‘a kind of battle ground, where the lower desires fight with the higher sentiments and ideas for the reign – at least in nobler individuals’ (Flechsig, 1896b, p. 4) (translation K.Z.). This is clearly a social-darwinistic perspective of brain organization (Hagner, 1999) reflecting the general ideological, non-scientific narrative of Flechsig’s functional concept. The psychologists Theodor Lipps (1851–1914), Carl Stumpf (1848–1936), and Hermann Ebbinghaus (1850–1909), however, stated that it remains unclear what Flechsig really means with the term ‘function’; functions like those mentioned by Flechsig are probably not explainable by morphological findings. Flechsig’s separation of association centres and their fibres from projection centres was denied by Joseph Jule Dejerine and Augusta Dejerine-Klumpke (1885) based on their pioneering anatomical and pathological studies. They demonstrated the occurrence of normal or degenerated projection fibres in nearly all cortical regions including Flechsig’s association centres. Vogt (1887) also emphasized that association and projection fibres are not separated by specific brain regions, a fundamental assumption of Flechsig’s original association theory. ‘Psychological considerations do not require special association centers regarding the physiological aspect. … the identification of an anatomical center does not explain the physiological processes in this center.’ (Vogt, 1887, p. 358–60). Flechsig’s anatomical foundation of his functional concept of association centres was not accepted by Brodmann and Vogt, although they highly regarded his myelogenetic findings as an important step forward. Brodmann also critically assessed Flechsig’s method to subdivide the cortex into structural-functional units:
‘If myelogenetic studies do not reveal at all such a marked difference and, more so, if judging by the temporal sequence of myelinisation the two central gyri represent a homogeneous and unseparable entity, it is evidence … that myelinisation can only be considered an indicator of functional localisation with considerable reservations and with critical prudence.’ (Brodmann, 1909, p. 255), and later he continues: ‘The delineation of myelogenetic centers, particulary of the late myelinating association centers is mainly subjected to the arbitrariness of the observer regarding the number, extension and special localization of distinct areas.’ (Brodmann, 1914, p. 118) (translation K.Z.).
In contrast to Meynert and Flechsig, Brodmann shifted the scientific focus from white (fibres) to grey matter to define cortical units by cytoarchitecture. He simultaneously emphasized the importance of connectivity by mentioning Flechsig’s studies:
‘The variety and the gradations of form and degree of higher intellectual activity are thus merely the expression of the infinite variability of functional combinations of individual cortical organs. The possibility of such variable and diverse complexity is supported by the evidence that I have given that the cortical surface is composed of numerous such specific morphological organs. … As is well known, Flechsig … believes that his anatomical division, based on the asynchronous myelinisation of different sections of the cortex, also points to a differentiation of organs for individual ‘spiritual powers in the older psychological sense’, without, of course, giving further detailed explanations as to the localisation of such powers.’ (Brodmann, 1909, pp. 250–51).
Between 1900 and 1904 Campbell undertook a combined cyto- and myeloarchitectonic parcellation of the human brain (Supplementary Figs 6 and 7), and provided a functional interpretation of his structural findings (Campbell, 1904, 1905). Campbell described his pioneering work in a presentation read during the Annual General Meeting of the Medico-Psychological Association in London on 22 July 1904:
‘I endeavoured to demonstrate that a comprehensive study of the form and arrangement of these constituents [here he means the myelinated fibres of the cortex, K.Z.] in the normal adult condition afforded a useful guide to precise localisation of function. … Viewed collectively, the human brain harbours two varieties of centres, controlling what we may call “primary” and “higher evolutionary” functions respectively; the former are those common to all animals and essential to survival, viz. centres for movement and common and special sensation; the latter are those complex psychic functions in the possession of which man rises superior to all other beings.’ (Campbell, 1904, p. 651–52).
Here, he apparently refers to projection and association fibres and centres in the cortex following Meynert’s and Flechsig’s concepts, but further developed their concepts by introducing a comparative anatomical approach: ‘In carnivora and other lower animals, cortex can be recognised almost identical in structure with the motor cortex of man and the man-like ape’ (Campbell, 1904, p. 653). He not only introduced the argument of homology, but replaces the speculative ‘control’ concept of Meynert (see above) and the ‘hierarchical’ concept of Flechsig (‘lower desires’ are reigned by ‘higher ideas’, see above) by data-based arguments. This becomes evident when he speaks about the prefrontal and the parietal cortex: ‘For my part I think that histology brings proof of what was previously only ‘surmise’ that it is the very last pallium to appear in the progress of phylogenesis. I would submit that it is a part with a future, but that at present its evolution is incomplete.’ (Campbell, 1904, pp. 655–56). The postcentral gyrus is the main terminal for common sensory impressions for him.
‘I would submit that the “post-central area” proper may be a primary centre, one serving for the recognition of … common sensation … while parts behind, including the “intermediate post-central area” may constitute a higher centre. … Therefore, whether our parietal lobes share with our frontal convolutions the conduct of high psychic processes or not, there is no doubt that both undergo equal expansion in the progress of phylogenic development.’ (Campbell, 1904, pp. 656–58).
Although he admired the perfection of microphotography in Brodmann’s study, his drawings of the laminar architecture (Supplementary Fig. 8) and normal as well as pathologically altered nerve cells (Supplementary Fig. 9) are stunning by their realism and clarity. While agreeing with Campbell in many points, Brodmann stated that Campbell missed the necessary subdivision of the inferior frontal gyrus into three areas (BA44, BA45, and BA47) as separate architectonic entities (Fig. 2A), as well as subdivisions in the temporal and cingulate cortices.
‘in particular, according to all that can be concluded from anatomical localizational data, classic theory, and that at least the anterior sections of the inferior frontal gyrus, and perhaps even part of the actual orbital surface, must be included in it (thus, apart from area 44, also areas 45 and 47 of the brain map…).’ (Brodmann, 1909, p. 259–60).
Elliot Smith (1907) described a more detailed cortical parcellation of the human cortex (Supplementary Figs 3C and 7C) than Campbell (1905). He subdivided the cortex into over 30 cortical areas after observations of more than 200 hemispheres from different ages and ethnic origin. Comparable areas can also be found Brodmann’s cytoarchitectonic map (Supplementary Fig. 3B), on which he worked during the same period as Elliot Smith, who emphasizes ‘that as a rule sulci do not develop with mathematical precision at the exact boundary lines of adjoining areas’ (Elliot Smith, 1907, p. 237). This statement is in complete accordance with Brodmann’s findings. He also emphasized the existence of sharp borders between areas in accordance with Brodmann. ‘The changes in structure occur with the utmost abruptness, so that it is possible to determine with absolute precision the exact boundaries of each area.’ (Elliot Smith, 1907, p. 240). In contrast to previous studies, Elliot Smith avoided any functional or histological interpretation, although his intracortical ‘bands’ apparently match the Baillarger stripes of the primary visual cortex. ‘It is not pretended or in any way assumed that such contrasts in appearance necessarily imply physiological differences or even essential distinctions in histological constitution.’ (Elliot Smith, 1907, p. 238).
Thus, the answer to the question ‘What is the function of an area?’ remained. Brodmann wrote in his monograph: ‘Although my studies of localization are based on purely anatomical considerations … my ultimate goal was the achievement of a theory of function and its pathological deviations.’ (Brodmann, 1909, p. 239). He proceeds by saying: ‘Only … functions of the most elementary kind … can be associated with histological elements, but never complex psychic images’ (Brodmann, 1909, p. 243). And then he proposes a concept of cortical areas, which he sometimes called ‘organs’ or ‘centres’ or ‘types’ of the cerebral cortex:
‘we have now been able to determine that the cerebral cortex consists of a number of individual histologically highly differentiated organs, each of which has a clearly determined position … We have also seen, that such histological “centres” can be delimited in all other mammals as well as in man … and that individual homologous structural zones adopt the same, clearly determined positions on the cortical surface in all mammalian brains’ (Brodmann, 1909, p. 249).
The use of the term ‘organs’ was very unfortunate, because it suggests that Brodmann may be a neophrenologist following Gall’s ideas of circumscribed organs in the cortex subserving complex mental faculties (Gall, 1835). However, Brodmann clearly stated:
‘We must therefore reject as a quite impossible psychological concept the idea that an intellectual faculty or a mental event or a spatial or temporal quality or any other complex, higher psychic function should be represented in a single circumscribed cortical zone, whether one calls this an “association centre” or “thought organ” or anything similar. … one should rather envisage the situation that in each particular case supposed “elementary functional loci” are active in different numbers, in different degrees and in differing combinations. … Such activities are, however, always the result (and not merely the sum) of the function of a large number of sub-organs distributed more or less widely over the cortical surface; they can never be the product of a morphologically or physiologically independent “centre”.’ (Brodmann, 1909, p. 251).
Thus, he was neither a naïve localizationist, nor a neophrenologist. Rather, he stimulated a novel research concept, which neither followed the attempts to assign complex functions to single cortical units nor negated any functional localization by holistic interpretation of functional representation. Thus, his search for functions of cytoarchitectonically defined areas differs from that of his contemporaries, and led to a ‘network’ concept by stating that single loci are active in differing combinations when realizing a complex function. Thus, he paved the way for the functional connectivity analysis of modern neuroimaging using functional MRI or PET.
Brodmann’s work is a paradigmatic shift from functional considerations in the succession of phrenology as used by Gall, Meynert, and Flechsig to a new research concept, which is guided by the principle of evolution of brain structures. The importance of Brodmann’s cytoarchitectonic maps is not primarily reasoned by the higher number of areas. Von Economo and Koskinas (1925) will present a map with many more areas only a few years later. Brodmann’s maps fundamentally differ from all previous attempts by the detection and specification of the ‘common root of cortical layering’ (Brodmann, 1909, p. 6), i.e. the six-layered basic type of cortex in all mammals. An area is identified as being homologue if it shows the same local modification of the characteristic features of its basic cytoarchitectonic type, i.e. koniocortical, granular, dys/agranular, number of sublayers, cell types and sizes. Thus, the interspecies similarity in cytoarchitectonic features justifies the comparability, and not primarily the topological relation between cortical areas. This microscopically founded concept proved to be the breakthrough for any comparative mapping and is widely accepted today (Pandya et al., 2015; Palomero-Gallagher and Zilles, 2017; Zilles and Palomero-Gallagher, 2017). Brodmann’s map is not the result of a simple race for more areas, but the consequent application of morphological arguments to the entire cerebral cortex.
Brodmann also compared his map with results of electrical stimulations reported in the contemporary literature. The studies of Sherrington and Grünbaum (1902) and Vogt and Vogt (1907) were particularly important for his position. Electrical stimulation of the central and precentral region supported the idea that cortical motor activity can be elicited also outside BA4, which contains the giant pyramidal cells.
‘The Area gigantopyramidalis is found within the electrically excitable zone of the cerebral cortex, but does not cover the whole extension of this zone. We could definitely demonstrate in many monkeys that the region of electrically excitable foci of the cortex is larger in rostral direction than the Area gigantopyramidalis. The physiological and histological borders and also do not match in dorso-medial direction.’ (Brodmann, 1906, p. 395) (translated by K.Z.).
It became evident that the excitomotor zone, identified by electrical stimulation, extends anteriorly well beyond BA4—the primary motor cortex—in all animals (Vogt and Vogt, 1907). Thus, BA4 is not the only cortical motor centre.
‘Thus we arrive at the conclusion that, in very different animals, there exists a considerable correspondence between a physiologically very important cortical zone, the electrically excitable motor area, and an anatomically defined zone. … They could firstly be due to deficiencies in technique and observation, but secondly it is also possible that the more extensive anatomical region might represent a higher element within which the smaller “excitatory zone” represents only a partial function, as it is absolutely not necessary to think that the electrically excitable zone must be identical with the “motor region” in the strictest sense, that is the centre for voluntary movement.’ (Brodmann, 1909, p. 256–57).
The comparison of BA8 with electrical stimulation studies in lemur and cercopithecus brains (Vogt and Vogt, 1907, 1926) is a further example for the subdivision of the more posterior part of the frontal lobe into primary motor, premotor, and putative frontal eye field regions (Supplementary Fig. 10). Furthermore, the somatotopic map in BA4 (Supplementary Fig. 10) reminds of Penfield's homunculus long before Penfield and Boldrey (1937).
Brodmann’s maps are not the last word in cytoarchitectonics
Brodmann revolutionized research on the microstructure of the human brain by establishing the common six-layered laminar ‘bauplan’ and its regional modifications for the entire neocortex of mammals. His studies of brain evolution (Brodmann, 1906, 1912) in numerous mammalian species, which he selected under an evolutionary aspect, and the discovery of the modifications of the laminar structure during ontogeny (Brodmann, 1909, 1914) were the imperative conditions.
Since his map is mainly used nowadays for the localization of activation foci in neuroimaging studies, one limitation of his maps becomes evident. He did not register the interindividual variability (with the exception of the thickness of layers; Brodmann, 1908c) of the localization and size of the areas (Lashley and Clark, 1946). To comply with this variability, cortical maps must be probabilistic, as borders of cytoarchitectonic areas as well as those of functional foci are highly variable between subjects (Zilles and Amunts, 2010). Such probabilistic cytoarchitectonic maps are now available (Eickhoff et al., 2005; http://www.fz-juelich.de/SharedDocs/Downloads/INM/INM-7/DE/SPM_Toolbox/Toolbox_22c.html).
A further limitation of Brodmann’s cytoarchitectonic studies is the observer-dependent definition of borders of cortical areas. This drawback is shared by all classical cytoarchitectonic studies including the monumental work of von Economo and Koskinas (1925). It led to severe attacks against cytoarchitectonic studies in general. Bailey and von Bonin came to doubt after having accepted this type of research in their earlier work. Their ‘evaluation’ of Brodmann’s work has been described recently (Zilles and Amunts, 2010). They argued that the cerebral cortex was over-parcellated by Brodmann and followers ‘the efforts of all these authors were meticulous to the point of hair splitting.’ (Bailey and von Bonin, 1951). They highlighted the lack of observer independency. Advances in computerized image analysis (Schleicher et al., 1999) have overcome this limitation.
Finally, Brodmann’s delineations must be corrected in the regions of higher multimodal (visual) and association cortices (parietal, temporal and prefrontal). In most cases, necessary subdivisions are lacking in his maps, as shown in recent publications based on computerized image analysis and multimodal brain mapping (e.g. Amunts and Zilles, 2006; Toga et al., 2006; Weiner et al., 2014; Gomez et al., 2017). These methods were not available at the time.
A sad end to the years in Berlin and the move to Tübingen
In 1908, Brodmann submitted his habilitation thesis on ‘Cytoarchitectonic parcellation of the cerebral cortex in prosimians’—a requirement for obtaining a professorship position. The thesis has been officially rejected by the University of Berlin because of not having published clinical studies (letter with signature 208 in the Cécile and Oskar Vogt Archive, C. and O. Vogt Institute for Brain Research, University Düsseldorf)—which is apparently incorrect, since Brodmann had published studies on the effects and methods of hypnosis in the therapy of psychiatric diseases (Brodmann, 1897, 1898a, b, 1900, 1902, 1902–1903a, b). The real cause behind the curtain was Brodmann’s severe and published criticism of the work of the influential psychiatrist Theodor Ziehen in Berlin and the enduring intrigues of Paul Flechsig against Oskar Vogt and his ambitious new Neurobiological Laboratory, as well as against all of Vogt’s collaborators including Brodmann (Fig. 7).
As a consequence of this dirty battle, the atmosphere between Vogt and Brodmann worsened, because Brodmann suspected that Vogt did not support him sufficiently, although Vogt addressed letters to Berlin University and many colleagues outside fighting for Brodmann (letter with signature 208 in the Cécile and Oskar Vogt Archive, C. and O. Vogt Institute for Brain Research, University Düsseldorf). The quarrel culminated in an exchange of letters, in which Vogt threatened Brodmann with a summary dismissal (letter with signature 208 in the Cécile and Oskar Vogt Archive, C. and O. Vogt Institute for Brain Research, University Düsseldorf). Finally, Brodmann cancelled his position in Berlin, and moved to the Clinic for Psychiatry and Neurology of the University Tübingen, headed by Robert Gaupp. Brodmann was appointed as head of the anatomy laboratory in 1910. His habilitation was accepted by the University of Tübingen on 7 February 1911, and he became an appointed professor. However, the beginning of World War I stopped all scientific plans. Between 1914 and 1916, he had to serve as a physician in a field hospital and took care of soldiers with brain injuries (Fig. 8A).
Figure 8.

Photographs portraying Brodmann's life as an established MD. (A) Brodmann as a physician in a hospital for soldiers. Photo taken between 1914 and 1916. With permission of the Brodmann Museum, Hohenfels-Liggersdorf. (B) Korbinian Brodmann and his wife Margarete Francke in the year 1917. With permission of the C. & O. Vogt Archive, Institute of Brain Research, University Düsseldorf.
In 1913, Brodmann gave the main lecture during the congress of Nature Researches and Physicians in Vienna on the comparative anatomy of the cerebral cortex and anthropological questions (Brodmann, 1913). Two major questions were discussed in this lecture: ‘Generally speaking, does the analysis of the localization in the cerebral cortex provide arguments for statements on the higher or lower organization of the brain and thus the higher or lower mental abilities of a mammal, particularly the human species? and Can localizations be found in the brains of different human races, which can be interpreted as indicators of a more primitive condition, and as such eventually for the problem of human races?’ (translated by K.Z.). Although he denied possible correlations between a larger brain or a greater volume of the cortex with better intellectual abilities, he followed the widely accepted research direction of the 19th and the beginning of the 20th century by replacing measurements of the entire brain by those of the size of single cortical areas (Brodmann, 1912). The cerebral cortex had already played a central role in such discussions; it was seen as the central control level, like the capital of a country (Meynert, 1891). This narrative of hierarchical relationships between higher and lower brain regions used pairs of terms such as conscious/unconsciousness, expedient/automatic, healthy/unhealthy, rational/irrational, civilization/savagery. Brain research had attained a position where it outreached to an explanation of sociocultural, political and ethical issues.
‘The human being develops more and more into a brain-animal. Our brain will play an ever-growing role in our development. This development will also provide larger risks for our health. A successful future of our species depends to a large degree from the development of a sort of brain hygiene.’ (Vogt, 1912) (translated by K.Z.).
Brain research should ‘analyse the abilities and activities of each individual … by applying neuropsychological measures to each individual … to detect the genetic factors of the mental personality … this way providing the long awaited scientific basis for a voluntary selection, and ethnic hygiene of the future.’ (Vogt, 1912) (translated by K.Z.). Along these concepts, Brodmann measured the surface of the cortex, and found that the cortical surface hidden in the sulci is absolutely and relatively larger in brains of ‘Europeans’ than in those of indigenous populations (‘Naturmenschen’), and even larger than in mentally handicapped people. He also measured the brains of various non-human primates, carnivora, ungulata, cetaceans, pinnipedia, rodentia, insectivores, edentata, marsupialia, and monotremata. He stated that the size of the entire cortex depends on the phylogenetic position, body and brain size, and does not enable any conclusion on correlations between these parameters and intelligence. He further states that the degree of cortical folding is not a measure of intelligence, nor a characteristic of race. He finally concluded that macromorphological measures are not reliable measures of psychological properties, because such features (e.g. sulci and gyri) seldom coincide with the borders of cortical areas, as determined by cytoarchitecture. Thus, he compared four brain regions, whose borders are sharp and clearly homologue in an interspecies comparison, i.e. the motor cortex, the total prefrontal cortex, the primary visual cortex, and the olfactory part of the allocortex. In the prefrontal and motor cortices, he found considerable inter-species differences in absolute and relative size (relative to the total cortical surface). The primary visual cortex is larger in primates than in most other species, and despite a considerable inter-subject difference, larger in Europeans than in members of the African indigenous population. However, the relative size of BA17 is smallest in humans, because their parietal lobe is disproportionally developed. The smallest relative size was reached by the Europeans, followed by the somewhat larger sizes in the members of the African indigenous populations, and even larger sizes in mentally handicapped or microcephalic Europeans. The largest relative sizes are found in non-human primates. The olfactory cortex shows the smallest relative size of its surface in humans but much larger sizes in all other mammals. Because of the small size of the human brain sample, Brodmann mentioned the preliminary character of his study, but despite this restriction, he emphasized the ethnic differences in the relative sizes of the prefrontal, motor and visual regions between Europeans and non-Europeans. He also emphasized the regional sizes of handicapped people. His discussion of the occurrence of the lunate sulcus sharpens his conclusion by stating that a lunate sulcus is found in non-human primates and brains of indigenous Africans, but not in European brains. Although he did not explicitly speculate on the functional superiority based on the size of regions, Brodmann followed with these statements the internationally predominating opinion of anthropology at his time. He arrived in a dangerous neighbourhood with rising racism and its ‘scientific’ supporters.
In 1916, Brodmann moved as prosector to the Nietleben mental asylum near Halle in the Eastern part of Germany, where he stayed until 1918. In this position he was assured of reasonable financial security. He met the technician Margarete Francke, who became his wife on 3 April 1917 (Fig. 8B). In 1918 their daughter Ilse was born. During the Nietleben period, Brodmann made a further step forward regarding his brain map: after many autopsies, he stated that the primary visual cortex can show a considerable inter-subject variability in size and extent, as already mentioned in his 1913 publication (Brodmann, 1913). In two talks in Würzburg and Halle, he discussed individual variations of the visual sphere and the clinical importance of shot wounds to the back of the head. He would never finish this important supplement to his brain map. In 1914 he published a summary of his previous work together with remarks on the physiology of the brain in a large handbook chapter (Brodmann, 1914). This was Brodmann’s last publication.
Climax of Brodmann’s personal career and sudden death
In 1918, Brodmann reached the climax of his career. He was appointed as head of the Department for Topographical Anatomy in the famous Research Institute for Neurology in Munich. This institute was founded by the eminent psychiatrist Emil Kraepelin. It was the world’s first privately financed, university-independent and interdisciplinary brain research institute. The neuropathologists Alois Alzheimer and Walther Spielmeyer, and the anatomist Franz Nissl were most renowned heads of other departments. Brodmann started a collaboration with Nissl. However, Brodmann’s fate annihilated all great plans. Lawrence Garey (2002) wrote:
‘On the 17th of August 1918, he developed what seemed to be a simple influenza, but after a few days signs of septicaemia appeared. It is thought that an old infection that he had contracted during an autopsy some time earlier had flared up. Brodmann was normally very strong and healthy, and even saw his illness as a way of catching up a backlog of work. He seemed not to suspect that this was not to be. One day he was seen to be making writing motions on his bed with his finger before sinking back, dead’.
He died suddenly before he could move to this new promising position in Munich.
Brodmann’s work remains a seminal landmark for structural localization of cortical areas in neuroimaging research. Thousands of studies still refer to Brodmann’s map of the human cerebral cortex. During the first decade of the 20th century, he paved the way for brain mapping. He is the tragic figure in our field of research on whose shoulders we stand. Vogt wrote in his biography of Brodmann:
‘Just at the moment when he had begun to live a very happy family life and when, after years of interruption because of war work, he was able to take up his research activities again in independent and distinguished circumstances, just at the moment when his friends were looking forward to a new era of successful research from him, a devastating infection snatched him away after a short illness, on August 22nd 1918’ (Vogt, 1952).
Supplementary Material
Acknowledgements
The author wishes to thank the Brodmann Museum, Hohenfels-Liggersdorf for the permission to use photos related to Brodmann biography and hand drawings of brain maps by himself. My special thanks goes to Ursula Grell, Berlin. Thousands of documents and rare photos now in the Vogt Archive of the C. and O. Vogt Institute for Brain Research of the University of Düsseldorf would not be professionally archived and available for researchers without her tremendous work and enduring enthusiasm. I also have to thank Dr Markus Fix for extremely careful research on the bibliography of Brodmann, which is part of his dissertation thesis (Fix, 1994), and Dr Nicola Palomero-Gallagher for helpful discussions during the work on this manuscript.
Funding
No funding was received towards this work.
References
- Amunts K, Zilles K. A multimodal analysis of structure and function in Broca’s region. In: Grodzinski Y, Amunts K, editors, Broca’s Region. New York, NY: Oxford University Press; 2006. p. 17–30. [Google Scholar]
- Amunts K, Zilles K. Architectonic mapping of the human brain beyond Brodmann. Neuron 2015; 88: 1086–107. [DOI] [PubMed] [Google Scholar]
- Barbas H. Pattern in the laminar origin of corticocortical connections. J Comp Neurol 1986; 1252: 415–22. [DOI] [PubMed] [Google Scholar]
- Barbas H. General cortical and special prefrontal connections: principles from structure to function. Annu Rev Neurosci 2015; 38: 269–89. [DOI] [PubMed] [Google Scholar]
- Bailey P, von Bonin G. The isocortex of man. Urbana, IL: University of Illinois Press; 1951. [Google Scholar]
- Bielschowsky M, Brodmann K. Zur feineren Histologie und Histopathologie der Großhirnrinde mit besondere Berücksichtigung der Dementia paralytika, Dementia senilis und Idiotie. J Psychol 1905; 5: 173–199. [Google Scholar]
- Campbell AW. Histological studies on the localisation of cerebral function. J Mental Sci 1904; 50: 651–9. [Google Scholar]
- Campbell AW. Histological studies on the localization of cerebral function. Cambridge: Cambridge University Press; 1905 [Google Scholar]
- Dejerine JJ, Dejerine-Klumpke A. Anatomie des centres nerveux. Vol. 1 Paris: Rueff; 1885. [Google Scholar]
- Eickhoff S, Stephan KE, Mohlberg H, Grefkes C, Fink GR, Amunts K, et al. . A new SPM toolbox for combining probabilistic cytoarchitectonic maps and functional imaging data. Neuroimage 2005; 25: 1325–35. [DOI] [PubMed] [Google Scholar]
- Elliot Smith G. A new topographical survey of the human cerebral cortex, being an account of the distribution of the anatomically distinct cortical areas and their relationship to the cerebral sulci. J Anat Physiol 1907; 41: 237–54. [PMC free article] [PubMed] [Google Scholar]
- Fix M. Leben und Werk des Gehirnanatomen Korbinian Brodmann (1868-1918). Inaugural dissertation, Tübingen: Eberhard-Karls-Universität Tübingen; 1994. [Google Scholar]
- Flechsig P. Gehirn und Seele. Rektorrede am 31. Oktober 1894. Leipzig: L. Edelmann; 1894. [Google Scholar]
- Flechsig P. Die Localisation der geistigen Vorgänge insbesondere der Sinnesempfindungen des Menschen. Leipzig: Veit & Comp; 1896a. [Google Scholar]
- Flechsig P. Die Grenzen geistiger Gesundheit und Krankheit. Rede gehalten zur Feier des Geburtstags Sr. Majestät des Königs Albert von Sachsen am 23. April 1896. Leipzig: Veith & Comp; 1896b. [Google Scholar]
- Flechsig P. Developmental (myelogenetic) localisation of the cerebral cortex in the human subject. Lancet 1901; 2: 1027–9. [Google Scholar]
- Flechsig P. Meine myelogenetische Hirnlehre mit biographischer Einleitung. Berlin: Springer; 1927. [Google Scholar]
- Gall FJ. On the functions of the brain and of each of its parts: with observations on the possibility of determining the instincts, propensities, and talents, or the moral and intellectual dispositions of men and animals, by the configuration of the brain and head. 6 Vols., trans. Winslow Lewis, Jr. Boston: Marsh, Capen and Lyon; 1835. [Google Scholar]
- Garey LJ. Brodmann’s “localisation in the cerebral cortex”. Garey LJ, translator/editor. London: Smith-Gordon; 1994. 2nd edn 1999: London: Imperial College Presocietys; 3rd edn.2006: Heidelberg: Springer. [Google Scholar]
- Garey, L. History of Neuroscience: Korbinian Brodmann (1868-1918). IBRO History of Neuroscience, 2002. [http://ibro.org/wp-content/uploads/2018/07/Brodmann-Korbinian.pdf] [Google Scholar]
- Gomez J, Barnett MA, Natu VS, Mezer A, Palomero-Gallagher N, Weiner KS, et al. . Growth of tissue in human cortex is coupled with the development of face processing. Science 2017; 355: 68–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goulas A, Zilles K, Hilgetag CC. Cortical gradients and laminar projections in mammals. Trends Neurosci 2018. doi: 10.1016/j.tins.2018.06.003 [DOI] [PubMed] [Google Scholar]
- Hagner M. Gehirnführung. Zur Anatomie geistiger Funktionen, 1870-1930, In: Hagner M, editor. Ecce cortex. Göttingen: Wallstein Verlag; 1999. pp. 177–205. [Google Scholar]
- Judas M, Cepanec M, Sedmak G. Brodmann’s map of the human cerebral cortex – or Brodmann’s maps. Transl Neurosci 2012; 3: 67–74. [Google Scholar]
- Lashley KS and Clark G. The cytoarchitecture of the cerebral cortex of Ateles: a critical examination of architectonic studies. J Comp Neurol 1946; 85: 223–305. [DOI] [PubMed] [Google Scholar]
- Lichtheim L. On aphasia. Brain 1885; 7: 433–84. [Google Scholar]
- Markov NT, Julien Vezoli J, Chameau P, Falchier A, Quilodran R, Huissoud C, et al. . Anatomy of hierarchy: feedforward and feedback pathways in macaque visual cortex. J Comp Neurol 2014; 522: 225–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meynert T. Ein Fall von Sprachstörung, anatomisch begründet. In Braun C, Duchek A, Schlager L, editors. Zeitschrift der K.u.K, Vol. 22 Gesellschaft der Ärzte in Wien; 1866. p. 152–189. [Google Scholar]
- Meynert T. Der Bau der Gross-Hirnrinde und seine örtliche Verschiedenheiten, nebst einem pathologisch-anatomischen Corollarium. Neuwied, Leipzig: Heuser; 1868 [Google Scholar]
- Meynert T. (1891) Das Zusammenwirken der Gehirnteile. Verhandl. d. 10. Intern Med Kongresses 1: 173–90. Summary by Ziehen T (1892) In: Zeitschrift für Psychologie und Physiologie der Sinnesorgane, 3: 59–61. [Google Scholar]
- Palomero-Gallagher N, Zilles K. Cortical layers: cyto-, myelo-, receptor- and synaptic architecture in human cortical areas. Neuroimage 2017. doi: https://10.1016/j.neuroimage.2017.08.035 [DOI] [PubMed] [Google Scholar]
- Pandya DN, Petrides M, Seltzer B, Cipolloni PB. Cerebral cortex: architecture, connections, and the dual origin concept. Oxford: Oxford University Press; 2015. [Google Scholar]
- Peiffer J. Hirnforschung in Deutschland 1849 bis 1974: Briefe zur Entwicklung von Psychiatrie und Neurowissenschaften sowie zum Einfluss des politischen Umfeldes auf Wissenschaftler. Berlin: Springer; 2013. [Google Scholar]
- Penfield W and Boldrey E. Somatic motor and sensory representation in the cerebral cortex of man as studied by electrical stimulation. Brain 1937; 60: 389–440. [Google Scholar]
- Ramón y, Cajal S. Studien über die Hirnrinde des Menschen. Bresler J, translator. Vol. 1. Die Sehrinde, 1900, Vol. 2. Die Bewegungsrinde, 1900, Vol. 3. Die Hörrinde, 1902, Vol. 4. Die Riechrinde beim Menschen und Säugetier, 1903, Vol. 5. Vergleichende Strukturbeschreibung und Histogenesis der Hirnrinde, 1906. Leipzig, Barth; 1900-1906. [Google Scholar]
- Rose JE. Korbinian Brodmann. In: Haymaker W, Schiller F, editors. The founders of neurology. 2nd edn.Springfield, IL: Charles C Thomas; 1979. [Google Scholar]
- Sanides F. Functional architecture of motor and sensory cortices in primates in the light of a new concept of neocortex evolution. In: Noback CR, Montagna W, editors. The primate brain: advances in primatology. New York, NY: Appleton-Century-Crofts Educational Division/Meredith Corporation; 1970. pp. 137–208. [Google Scholar]
- Schleicher A, Amunts K, Geyerünbaur S, Morosan, P, Zilles K. Observer-independent method for microstructural parcellation of cerebral cortex: a quantitative approach to cytoarchitectonics. Neuroimage 1999; 9: 165–77. [DOI] [PubMed] [Google Scholar]
- Sherrington CS, Grünbaum ASF. A discussion of the motor cortex as exemplified in the Anthropoid Apes. Brit Med J part 1902; 2: 784–5. [Google Scholar]
- Talairach J, Tournoux P. Co-planar stereotaxic atlas of the human brain. Stuttgart: Thieme; 1988. [Google Scholar]
- Toga AW, Thompson PM, Mori S, Amunts K, Zilles K. Towards multimodal atlases of the human brain. Nat Rev Neurosci 2006; 7: 952–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vogt C, Vogt O. Zur Kenntnis der elektrisch erregbaren Hirnrindengebiete bei den Säugetieren. J Psychol Neurol 1907; 8: 277–456. [Google Scholar]
- Vogt C, Vogt O. Allgemeinere ergebnisse unserer hirnforschung. J Psychol Neurol 1919; 25: 279–462. [Google Scholar]
- Vogt C, Vogt O. Die vergleichend-architektonische und die vergleichend-reizphysiologische Felderung der GroBhirnrinde unter besonderer Berücksichtigung der menschlichen. Die Naturwissenschaften 1926; 50: 1190–4. [Google Scholar]
- Vogt O. Flechsig’s Associationscentrenlehre, ihre Anhänger und Gegner. Zeitschrift Hypnotismus 1887; 5: 347–61. [Google Scholar]
- Vogt O. Bedeutung, Ziele und Wege der Hirnforschung. Nord Süd 1912; 36: 309–14. [Google Scholar]
- Vogt O. Korbinian Brodmann (1868-1918). In: Kolle K, editor. Große Nervenärzte. Vol. 2 Stuttgart: Thieme; 1952. p. 40–4. [Google Scholar]
- von Economo C, Koskinas GN. Die Cytoarchitektonik der Hirnrinde des erwachsenen Menschen. Wien, Berlin: Springer; 1925. [Google Scholar]
- Weiner K, Golarai G, Caspers J, Mohlberg H, Zilles K, Amunts K, et al. . The mid-fusiform sulcus: a landmark identifying both cytoarchitectonic and functional divisions of the human fusiform gyrus. Neuroimage 2014; 84: 453–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wernicke C. Der aphasische Symptomenkomplex. Eine psychologische Studie auf anatomischer Basis. Breslau: Cohn und Weigert; 1874. [Google Scholar]
- Zilles K, Amunts K. Centenary of Brodmann’s map conception and fate. Nature Rev Neurosci 2010; 11: 139–45. [DOI] [PubMed] [Google Scholar]
- Zilles K, Amunts K. Anatomical basis for functional specialization. In: Uludag K, Ugurbil K, Berliner L, editors. fMRI: from nuclear spins to brain function. New York, NY: Springer; 2015. p. 27–66. [Google Scholar]
- Zilles K, Palomero-Gallagher N. Multiple transmitter receptors in regions and layers of the human cerebral cortex. Front Neuroanat 2017; 11: 78. doi: 10.3389/fnana.2017.00078 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.