

Abstract
Ionizing radiation (IR) therapy is a major cancer treatment modality and an indispensable auxiliary treatment for primary and metastatic cancers, but invariably results in debilitating organ dysfunctions. IR-induced depletion of neural stem/progenitor cells in the subgranular zone of the dentate gyrus in the hippocampus where neurogenesis occurs is considered largely responsible for deficiencies such as learning, memory, and spatial information processing in patients subjected to cranial irradiation. Similarly, IR therapy-induced intestinal injuries such as diarrhea and malabsorption are common side effects in patients with gastrointestinal tumors and are believed to be caused by intestinal stem cell drop out. Hematopoietic stem cell transplantation is currently used to reinstate blood production in leukemia patients and pre-clinical treatments show promising results in other organs such as the skin and kidney, but ethical issues and logistic problems make this route difficult to follow. An alternative way to restore the injured tissue is to preserve the stem cell pool located in that specific tissue/organ niche, but stem cell response to ionizing radiation is inadequately understood at the molecular mechanistic level. Although embryonic and fetal hypersensity to IR has been very well known for many decades, research on embryonic stem cell models in culture concerning molecular mechanisms have been largely inconclusive and often in contradiction of the in vivo observations. This review will summarize the latest discoveries on stem cell radiosensitivity, highlighting the possible molecular and epigenetic mechanism(s) involved in DNA damage response and programmed cell death after ionizing radiation therapy specific to normal stem cells. Finally, we will analyze the possible contribution of stem cell-specific chromatin’s epigenetic constitution in promoting normal stem cell radiosensitivity.
Facts
Ionizing radiation is a common cancer treatment, but it is often accompanied by side effects which cause normal tissue injuries and a decline in the quality of life.
Radioprotective drugs have been proven effective in vitro but fail to replicate their effect in vivo; the only FDA-approved drug available, Amifostine, is currently used to reduce xerostomia but it has also been proven to offer protection against several chemotherapeutic agents.
The loss of the stem cell pool is believed to be the cause of the normal tissue injuries and stem cells have been proven to be highly radiosensitive compared to differentiated cells.
Stem cell radiosensitivity is regulated by pluralistic mechanisms that involve both epigenetic and molecular signaling. Improved understanding of the regulatory pathways that make stem cells radiosensitive would lead to innovative radioprotective drug development and novel therapies to eradicate cancer while preserving the stem/progenitor cells.
Open questions
Do stem and non-stem cells respond differently to DNA breaks?
Are stem cells epigenetically programmed to favor cell death instead of repair and survival after radiation exposure?
What are the molecular mechanisms involved in the stem cell radiosensitivity?
Introduction
Following induction of DNA damage, cells respond in different ways and this DNA damage response (DDR) depends on several variables, such as cell cycle, post-translational modifications of the signaling cascade, and chromatin configuational changes1–3. When the DNA strand break is not severe or irreparable, cells respond by activating DNA repair pathways. Double-strand break repair is achieved by two major DNA repair pathways: homologous recombinational repair pathway (HR) which operates only in the post-replicative S or G2/M phases of cell division cycle and requires a homologous sister chromatid and non-homologous end joining (NHEJ) which operates mostly in the pre-replicative G1 phase of the cell cycle and is the most prominent form of DNA repair mechanism in terminally differentiated cells. When the damage is irreparable, cells respond with cell cycle arrest, apoptosis, senescence, or several other cell mechanisms4,5.
Ionizing radiation (IR) therapy is commonly used to treat cancers with the aim of inducing DNA double-strand breaks (DSBs) in cancer cells. The use of radiation therapy to kill cancer cells also causes DNA damage in the surrounding normal tissue and patients who undergo IR exposure experience treatment-related symptoms during therapy, months or even years after. Early side effects include erythema, dry desquamation, intestinal malabsorption, hyperpigmentation, and hair loss6–8. Late effects include skin atrophy, dryness, telangiectasia, dyschromia, dyspigmentation, fibrosis, ulcers, and neurocognitive decline9–12. Many decades ago it was perceived that a single stem cell was able to partially replenish the physiology of IR-damaged tissues13,14 and lack of this cell pool can lead to different side effects, such as accelerated aging, cognitive impairment, and poor learning and memory, especially in pediatric brain cancer patients. Stem cells at the pluripotent stage are capable of self-renewal and can produce all undifferentiated cell types of the tissue of origin, serving as an internal repair system by dividing and replenishing/replacing the dead cell populations. Because of their ability to restore damaged tissue, research has been driven towards the pathway of stem cell transplantation: although the only approved stem cell transplant in clinical practice is the bone marrow transplant used for cancers affecting the blood or immune system such as leukemia, lymphoma, or multiple myeloma, many other fields have been explored and recent findings suggest stem cell transplantation may provide a useful intervention strategy for minimizing the adverse effects of several pathologies such as Parkinson’s disease, amyotrophic later sclerosis, diabetes mellitus, heart disease, and much more15–20. However, application of stem cell transplant therapy to alleviate cognitive deficits in CNS malignancy treatment regimen has enormous practical limitations. Therefore, in loco stem cells could restore the normal physiology for tissues that have been injured by IR, given that the radiation did not cause any DNA damage in those cells21,22. Current studies in the literature are, however, inconclusive regarding the capability of DNA repair in stem cells after IR exposure, with several studies suggesting stem cells are more prone to activate apoptotic response pathway instead of DDR23–30.
This review will give an overview of normal tissue injury after radiation exposure and normal stem cell radiosensitivity to elucidate the developments in known mechanisms underlying radiation response in order to find a suitable pharmacological approach to protect stem cells from radiation-induced cell death.
Tissue injury after irradiation and known mitigation/radioprotection agents
The response of normal tissues to radiotherapy differs from one another after a chemical, thermal, or mechanical stress: radiation therapy damages DNA, proteins, and lipids in the cell through direct ionization or by production of free radicals, such as reactive oxygen species31. It also increases apoptosis, chromosomal aberrations, and mutation frequency in cells32. Fluorescence in situ hybridization probes show significant increases in chromosomal aberrations in murine splenocytes after total body irradiation, with a high number of translocations33, while in human lymphocytes a radiation dose-dependent increase of micronuclei formation has been observed, with multi-chromosome material detected above 2 Gy34.
These injuries can initiate several signaling pathways, including the inflammatory response, eventually leading to cell death, senescence, or disruption of tissue physiological functions35,36. Radiation injury-directed therapies can be divided into three groups: prevention of radiation damage by intervention before treatment, injection of a radioprotective agent during or immediately after radiation therapy in order to minimize the development of clinical injury, and treatment of radiation injury after therapy to prevent the progression to clinical impact37. Some commonly available treatments have shown promise in prevention or mitigation of normal tissue injury after irradiation, making drug-based approaches important for normal tissue radioprotection. Sulfhydryl (SH) compounds are known for their ability to act as radioprotectors; both cysteine and cysteamine have been proven to protect animals from the effects of total body radiation if they are injected or ingested before the radiation exposure. Sh-mediated cytoprotection has mainly two components: these radioprotectors are free-radical scavenging which are able to withdraw oxygen-based free-radical generation; moreover, their ability to donate hydrogen atoms has the potential of improving DNA repair. The only radioprotective drug approved by the U.S. Food and Drug Administration for use in radiation therapy is amifostine, a non-reactive phosphorothioate which is converted to its active form by the enzyme alkaline phosphatase. This drug has been proven to reduce side effects such as dry mouth and difficulties in eating or speaking in patients with head and neck cancer treated with radiotherapy, but it also showed significant protection against several chemotherapeutic agents. Amifostine, in pratice, is used only for the reduction of xerostomia38.
Another treatment of radiation injury after therapy to prevent the progression to clinical impact is stem cell transplantation. It is known that the stem cell pool has the ability to restore normal tissue structure/function: in fact, in 1967 an experiment by Withers elegantly showed that even one stem cell was sufficient to regrow nodules on a confined annulus of irradiated mouse skin39. Later on, the technique was perfected to obtain the survival characteristics of crypt cells of the mouse jejunum40, testicular stem cells41, and kidney tubules42. It has also been shown that stem cells are able to produce colonies when transplanted from a donor animal to a different site in a recipient mouse43,44. Due to their ability to repopulate an injured tissue, adult stem cells have been proposed for use in the treatment of radiation-induced tissue damage. Hematopoietic stem cell transplantation is currently used to reinstate blood production in leukemia patients, but several pre-clinical treatments with stem cells have shown promising results in different organs (i.e. skin, eye, lung, kidney) and might potentially translate into tissue radioprotection strategies45. However, there are many issues regarding use of stem cell therapy, including the lack of definitive markers for stem cells in different tissues, the rarity of the population for a successful transplant, together with major ethical and expense concerns covering the conventional radiotherapy. Therefore, a possible way to avoid complications related to transplant is to find new ways to protect the stem cell pool residing in the irradiated tissue. It is in fact suggested that stem cells are highly sensitive to IR; thus their ability to repopulate an injured tissue could be compromised after radiotheraphy. The mechanisms responsible for stem cell radiosensitivity have not been completely elucidated yet.
Embryonic and fetal DDR and sensitivity to IR
Radiation exposure of embryos and fetuses is of great concern for radiological diagnostics, radioprotection, and human health. Typical fetal doses from diagnostic radiology are usually low but some radio-diagnostic-specific treatments like cancer radiotherapy can expose the fetus to higher and potentially unsafe radiation doses46. The deleterious effect of IR to the embryo or fetus is strongly dependent on the stage of development, the absorbed dose, and fractionation47–49. According to extensive studies performed on animal models, a dose of 0.05–0.5Gy at the first and second week of pregnancy will slightly increase the incidence of implantation failure, but surviving embryos will probably have no significant health effects. During early organogenesis (third to sixth week postconception) the embryo is very vulnerable to growth retardation, teratogenic, and lethal effects of high-dose irradiation, while during early fetal stage the fetus develops a higher tolerance to radiation exposure, although the central nervous system can be seriously affected by high levels of IR exposure. At the end of pregnancy, the fetuses generally do not show any malformation after exposure to low amounts of radiation50. Moreover, some effects of radiation can only be observed many years post-exposure, such as behavioral disorders, infertility, neoplasia, and shorter life span51. Therefore, a definitive trend of hypersensitivity of embryonic cells has been observed in vivo over the past many decades in clinics and in experimental models as well.
However, research on embryonic stem cell in “culture” models have been inconclusive and often contradict the in vivo observations most likely due to late-passage spontaneous transformation effects in culture. It has been reported that DSBs caused by IR exposure in human embryonic stem cells (hESC) lead to the activation of the ataxia telangiectasia mutated (ATM) signaling pathway52,53, activating several proteins such as p53, Chk2, and Nbs1. Additionally, the number of γ-H2AX IR-induced foci (IRIF) increases dramatically within minutes following IR and returns almost to control levels in unexposed hESCs cultures within 24 h54. Besides the role of ATM in H2AX phosphorylation, Rad3-related (ATR) kinase was also proposed to play a role in the process53,55. However, our studies on multiple stem cell tissue niches in vivo and early-passage primary stem and isogenic differentiated non-stem cells have shown that murine embryonic stem cells are unable to activate ATM after IR-induced DNA DSB24,30 and that the γ-H2AX IRIF are significantly reduced compared to differentiated counterparts23. Similar results were observed by Suchorska and colleagues comparing human embryonic SCs, human induced pluripotent SCs, and primary human dermal fibroblasts56. Human embryonic stem cells show a robust apoptotic response after low IR exposure25,57 and a dramatic decrease in cell viability26,56 in a cell-cycle-dependent fashion58,59. Interestingly, the surviving hESCs continued to express common pluripotency markers and embryonic transcription factors, such as Oct4, Sox2, and Nanog25,52,53,56,60. The protection of hESCs from apoptosis following different exposures has been investigated and many biological pathways are shown to be involved61–67, but there are insufficient details regarding the response of hESCs after IR exposure. DNA synthesis and cell proliferation were partially inhibited68–71 but high expressions of anti-apoptotic protein Bcl-2 were found after irradiation together with low levels of pro-apoptotic proteins, indicating that the apoptotic response was not induced after IR exposure68,72–75. In contrast, early-passage primary murine embryonic stem cells showed high apoptotic response after 6 Gy exposure compared to their differentiated isogenic progeny23,24 together with high levels of pro-apoptotic factor Bax and low levels of anti-apoptotic protein Bcl-2 (ref. 30). hESCs show a higher number of aberrant mitotics after 2 Gy dose of irradiation compared with negative control and an arrest of the cell cycle in G(2)/M52. The influence of IR exposure on hESCs transcriptional response has been analyzed by Sokolov and colleagues: they found that hESCs have a clear pro-apoptotic transcriptional response 2 h after IR exposure, with upregulation of several genes such as BTG2, CDKN1A, SESN1, IER5, GADD45A which are genes often associated with temporal cell cycle arrest. At 16 h post IR the transcription patterns change, showing a strong expression of genes involved in pro-survival pathways and general metabolic signaling54. We observed IR-induced changes in the murine embryonic stem cell transcriptome which were associated with affecting biosynthesis, such as ribosome expression, while the non-stem cell transcriptome promoted activation of survival signaling pathways as well as protein digestion and absorption30. This aspect of the IR influence on hESCs has not been scrutinized deeply yet; thus, further studies are required. We therefore believe there is a need of serious evaluation of appropriate stem cell radiosensitivity and DDR research models to establish a common platform for inferring and proposing mitigating and/or preventive measures for radioprotection.
Adult stem cells sensitivity to IR
Adult stem cells have the tendency to respond differently from embryonic stem cells after IR-induced DNA damage. While hESC preferentially undergo apoptosis after IR treatment, adult stem cells exhibit a broad variety of responses and studies of stem cell radiosensitivity/radioresistance in literature are generally inconclusive (reviewed in refs. 76,77). A possible explanation is that stem cells in different tissues reside in niches that have tissue-specific environmental factors needed for stemness maintenance; moreover, the cell cycle status seems likely to play a role. Most importantly, the stem cell models used for studying the phenomenon or the mechanism is crucial in drawing inferences. It is in fact known that some adult stem cells are in the G0 phase of the cell cycle and are therefore quiescent. Many factors intervene in the maintenance of this status, which is believed to be important to preserve long-term proliferative potential and genomic integrity78,79. However, as quiescent cells, adult stem cells lack DNA damage checkpoints and are unable to activate cell cycle-dependent repair pathways, thus genomic integrity is not maintained80–83.
DDR activation
Human mesenchymal stem cells (hMSCs) are believed to be relatively resistant to IR exposure84–87. The proposed explanation for hMSCs is that their niche in vivo is hypoxic, so hMSCs may already be able to hinder the IR effects by having signaling machinery capable of responding to several insults at normoxic conditions88. DDR, like ATM protein phosphorylation, cell-cycle checkpoint activation, and DSB repair are some of the cellular mechanisms proposed to explain hMSCs radioresistance72,85,89,90. Some studies observed high levels of phosphorylated histone H2AX in MSC after IR exposure as a marker of DNA DSB repair process74,85–87,91,92, while other studies found overexpression 3 days post IR indicating a possible execution of cell senescence program68. Recently, it has been proposed that human MSCs exposed to prolonged X-irradiation accumulated γ-H2AX and 53BP1 foci differently compared to acute X-irradiation93; a careful distinction between the roles of apoptosis-related pan-nuclear γ-H2AX versus DDR-related γ-H2AX IRIF specific to the DSBs is required23.
After 4 Gy of IR treatment, human hematopoietic stem cells (hHSCs) did not show any activation of the cell cycle checkpoints29, but high levels of γ-H2AX have been reported in hHSC subpopulations when compared to more mature progenitors at 12 h post 3 Gy of IR, indicative of persistent DNA DSBs in these cells94. These results are in contrast to those presented after non-IR-related oxidative stress induction in hHSCs. It has been shown that hHSCs respond to oxidative damage with a strong activation of ATM, p53, 53BP1, CHK2, and FOXO3a and a senescence-like cell cycle arrest95,96. Recently, Biechonski and colleagues97 found that hematopoietic stem and early progenitor cells exhibit reduced NHEJ activities in comparison to non-lineage committed progenitors, along with more persistent 53BP1 foci. Moreover, the DNA repair process in hHSCs needs the presence of thrombopoietin, which supports important microenvironmental factors in the regulation of hHSCs IR exposure response98. Keratinocyte and melanocyte stem cells have reportedly shown high levels of γ-H2AX IRIF after 5 Gy irradiation with reduced colony-forming activity in culture and delayed hair cycle in vivo99 while lower dose of IR did not exert any effect on cell survival100.
Limited studies on DNA DSB repair in human neural stem cell (hNSC) cultures have reported the presence of γ-H2AX foci after irradiation, which reaches the IRIF level of non-IR treated cells 3 h after IR exposure27. However, careful in vivo experimentations by our group that has been published in recent years are in contrast with these findings where we demonstrate that instead of foci formation, pan-nuclear H2AX-S139 phosphorylation is observed and that murine neural stem cells selectively undergo IR-induced apoptosis compared to the non-stem differentiated cells in the hippocampal tissue niche (Fig. 1a). Stem cells in intestine (Fig. 1b) and testis (Fig. 1c) showed the same response, with superficial tissues like testis showing massive depletion of spermatogenesis in response to irradiation (Fig. 1d). As mentioned before, careful observation of DSB IRIFs versus pan-nuclear apoptotic γ-H2AX in multiple tissue niches and primary culture models, noγ-H2AX IRIFs were detected in stem cells (Fig. 2), indicating that stem cells are deficient in enabling DDR activation and DNA repair after radiation exposure23.
Fig. 1. Stem cells are more radiosensitive than their differentiated progeny.

a TUNEL staining (yellow-green signals) was performed in the dentate gyrus of the hippocampus obtained from brains of C57BL/6 mice after 6 Gy whole-body IR. b TUNEL staining was performed in tissue sections of adult murine intestine after 6 Gy IR. c TUNEL staining was performed in tissue sections of adult murine testis after 6 Gy IR. d TUNEL staining was performed in tissue sections of adult murine testis after 12 Gy IR. Scale bar = 10 µm. Original picture included for illustrative purposes, referring to ref. 23
Fig. 2. Stem cells (marked with red signals) fail to form γ-H2AX foci (green signals) after ionizing radiation treatment.

γ-H2AX was detected in adult murine testis stem cells by immunostaining after 0Gy (A) and 6 Gy IR (B). Scale bar = 10 µm. Original picture included for illustrative purposes, referring to ref. 23
More detailed studies are required to determine whether the varying DDRs observed in several stem cell populations is due to dissimilar cell cycle status, since stem cells from diverse niches proliferate at different rates or may be in a quiescent or proliferative stage.
Apoptotic response activation
hHSCs respond with massive apoptotic activation following low dose IR, showing both dose and time dependency28,29,101,102 and an implication of Bcl-2, p53, and ASPP1 in the process94. The same response has been observed in murine HSCs after 2 Gy treatment30. The clonogenic potential in 2 Gy-irradiated hHSCs decreased to ∼50–60% compared with the non-irradiated control103 while Wang et al.104,105 found that 6.5 Gy total body irradiation drove HSC to senescence. On the contrary, Chang and collaborators106 found that in vivo 1 Gy exposure was sufficient to drive HSC to quiescence, without any apoptotic effect observed in stem cells. No significant alterations in cellular senescence and apoptosis were detected in HSCs after exposure to low dose of radiation107.
hNSCs were previously shown to undergo programmed cell death after low, modest, and high doses of IR24,27,57,108 in a dose-dependent way27, with the surviving populations mostly starting a senescence process109, although hNSC radiosensitivity seems to be subpopulation-dependent110. Additionally, it has been recently observed that 10 Gy treatment affected both the cell survival and degree of differentiation in NSC through a bystander effect and activation of pro-apoptotic factors111. The apoptosis-susceptible nature of the irradiated hNSCs has been associated with a TRAIL-R2-mediated signaling cascade with activation of caspase-3 (ref. 112) and with prolonged upregulation of phosphorylated p53 (ref. 113). Activation of caspase-3 was found in murine spermatogonial stem cells after 6 Gy IR exposure (Fig. 3) and has been published before23,24,30.
Fig. 3. Stem cells undergo apoptosis after ionizing radiation treatment.
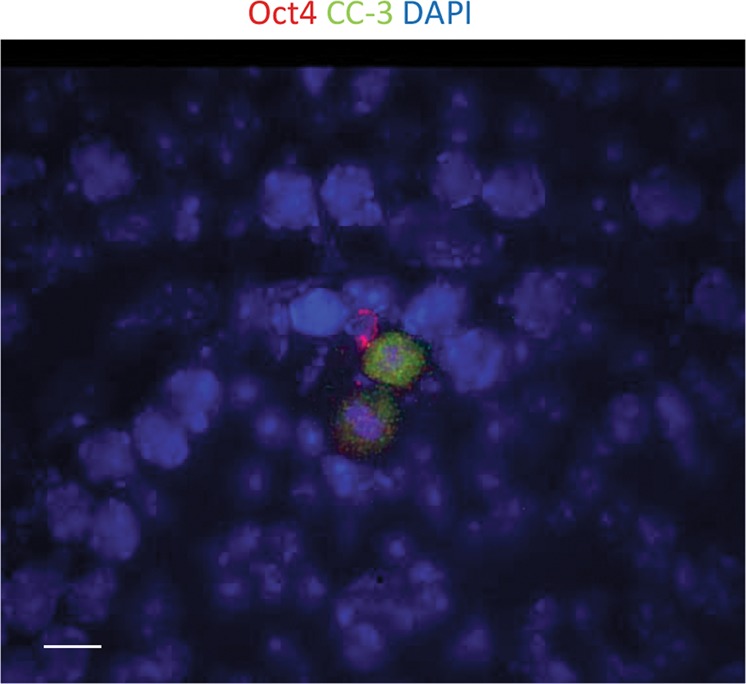
Cleaved caspase-3 (CC-3) was detected in adult murine testis stem cells by immunostaining after 6 Gy IR. Scale bar = 10 µm. Original picture included for illustrative purposes, referring to ref. 23
P53-dependent apoptosis has also been observed in Lgr5+ intestinal stem cells after irradiation in culture and mice114. In vivo experiments on mice show that 10% of intestinal stem cells initiate apoptosis after low doses, although cell death had no appreciable effect on tissue architecture115,116, while 15 Gy treatment caused more widespread apoptosis of Lgr5+ intestinal stem cells, resulting in a failure to restore viability of the small intestines117. We found Lgr5+ intestinal stem cells in ex vivo organoid cultures undergo massive apoptosis after 6 Gy irradiation while proliferative cells marked with Ki67 positivity largely comprised of SSEA1-negative non-stem cells in the same organoids showed greater radioresistance30.
The effect of IR on mesenchymal stem cells has been widely scrutinized in the past: minimal induction of cellular apoptosis was observed after treatment up to 20 Gy, with MSCs showing high expression levels of anti-apoptotic proteins BCL-2 and BCL-XL and low levels of pro-apoptotic proteins such as Puma (reviewed in ref. 73).
Although different stem cell populations respond with a diverse degree of programmed cell death, the dose utilized seems to play a fundamental role in the process.
Cell cycle arrest
hMSCs show a high and constant level of p53 after high doses of gamma-irradiation68, with ATM implicated in the post -translational modifications of p53 on ser 15, Chk1 on ser 345, and Chk2 on thr 68 (ref. 118). On the contrary, Kurpinski et al.119 found a modest gene expression change 5 h post 1 Gy of IR in hMSCs, although the downregulation of cyclin E2 (CCNE2) caused cell cycle arrest at this time point, while Jin et al.120 observed a complex response in the transcriptomic analysis of hMSCs after IR exposures, with early-response genes expressed at low doses and late-response gene at the highest doses. These conflicting results can be explained in the elegant work of Wu and colleagues: using early- and late-passage mesenchymal stem cells, they found IR exposure resulted in a decreasing trend in arresting both early- and late-passage MSCs in the G0/G1 phase up to 72 h post IR, and a substantial accumulation of early-passage MSCs in the G2/M phase. These results indicate that early-passage MSCs possess more effective cell cycle checkpoints in G2/M following IR exposure. It is likely that DNA damage is repaired through error-free HR in early-passage MSCs. Given that more late-passage MSCs were in the G0/G1 phase, the error-prone NHEJ can be the major DNA repair mechanism for late-passage MSCs, which may result in more genomic alterations87.
Intestinal stem cells undergo massive apoptosis after 20 Gy irradiation and show G2/M arrest induced by radiation to prevent mitotic catastrophe in a p53-independent manner121. In contrast, lower doses induced p53-mediated cell cycle arrest and protect the stem cell niche after DNA damage122. Lgr5+ intestinal stem cell radiosensitivity has been suggested to be CDK 4/6-dependent123, although a more recent study suggested that their radiosensitivity is related to DNA damage-dependent activation of Wnt signaling124. Moreover, although small and large intestines possess seemingly similar Lgr5+ stem cells, colonic epithelial stem cells (CESC) have been shown to be markedly more radioresistant in vivo than small intestinal crypt base columnar stem cells. Lgr5+ CESCs displayed delayed checkpoint recovery at 48 h post-19 Gy, which correlated with complete DSB repair and regeneration of colonic mucosa125.
It seems therefore that the cell cycle status might play an important role in stem cell radiosensitivity, dictating the activation of error-prone or error-free DNA repair mechanism after IR.
The pluralistic regulatory mechanisms of stem cell sensitivity to IR
Stem cell radiosensitivity is due to several signaling pathways that are activated/deactivated in response to irradiation. We believe that pluralistic interaction of molecular and epigenetic mechanisms collectively regulate and impart IR hypersensitive phenotype to the normal stem cells. Recently, we have found that stem cells constitutively express PP2A, which antagonizes and impairs DDR activation and promotes apoptosis at 0 Gy (Fig 4a, b) and after 6 Gy treatment (Fig. 4c), as has been published before30, confirming the hypothesis of Chowdhury et al.126.
Fig. 4. Stem cells display constitutively elevated IR-induced PP2A levels.

a PP2A detected in adult murine intestine by immunostaining at 0 Gy. b PP2A detected in adult murine testis by immunostaining at 0 Gy. c PP2A detected in adult murine testis by immunostaining at 30 min after 6 Gy. Scale bar = 10 µm. Original picture included for illustrative purposes, referring to ref. 30
The method by which gene expression is regulated without altering the genomic sequence, known as epigenetics, is another key factor involved in this multifaceted mechanism. Epigenetic changes include DNA methylation, histone acetylation, and miRNA-regulated gene expression127,128. Epigenetic alterations have been found to contribute to the pathogenesis of radiation-induced carcinogenesis129 by the reactivation of oncogenes and the silencing of tumor suppressor genes130. These events can result in genomic instability and consequent carcinogenesis in many models131–134.
Many epigenetic studies on ESC and induced pluripotent SC maintenance and differentiation have been extensively reviewed135–140, while little is known about epigenetic modifications after IR exposure.
Recently, the relationship between histone modifications and stem cell radiation responses has led to new insights: acetylation and methylation on different residues of H3 histone has been shown to play an important role in the radiosensitivity of stem cells. H3K9 modifications have been previously analyzed in ES cells141–143. Local downregulation of H3K9ac has been reported in ES cells, accompanying recruitment of the transcription factor OCT4 to DNA lesions144. In contrast, we found that ES cells are unable to deacetylate H3K9 at the DNA damage site compared to differentiated cells (Fig 5a, b), thereby limiting trimethylation on the same amino acid residue and consequent DNA damage repair as has been published before24 (Fig. 5c, d). In fact, H3K9 trimethylation is fundamental in the regulation of ATM activation after DNA DSB145,146. Deacetylation of H3K9 and consequent trimethylation has been proven to increase stem cells radioresistance, decrease IR-induced apoptotic response, and induce DDR activation24. Moreover, constitutively elevated histone-3 lysine-56 acetylation (H3K56ac) in stem cells results in a repressive chromatin environment that interferes with DDR activation and promotes radiosensitivity. Recruitment of histone deacetylases and deacetylation of H3K56ac is required at DSB for adequate DDR activation in non-stem cells147,148. We have observed higher H3K56 acetylation levels exclusively in neural stem cells of the dentate gyrus displaying elevated H3K56ac23 (Fig 6a, b). Knockdown of H3K56 acetyltransferase p300 reduced H3K56ac and significantly decreased radiosensitivity, restored DDR function, and increased stem cell survival23.
Fig. 5. Stem cells fail to deacetylate H3K9 after ionizing radiation treatment and trimethylate the same residue.

a H3K9 acetylation was detected in hippocampal stem cells obtained from brains of WT C57BL/6 mice by immunostaining after 6 Gy IR. b H3K9 acetylation was detected in adult murine testis stem cells by immunostaining after 6 Gy IR. c H3K9 trimethylation was detected in hippocampal stem cells obtained from brains of WT C57BL/6 mice by immunostaining after 6 Gy IR. d H3K9 trimethylation was detected in adult murine testis stem cells by immunostaining after 6 Gy IR. Scale bar = 10 µm. Original picture included for illustrative purposes, referring to ref. 24
Fig. 6. Stem cells display higher levels of H3K56ac (red signals) and pan-nuclear apoptotic H2AX-S139 phosphorylation 6 h after irradiation.

H3K56 acetylation and H2AX phosphorylation was detected in hippocampal stem cells obtained from brains of C57BL/6 mice by immunostaining a 3 h and b 6 h after 6 Gy IR. Scale bar = 10 µm. Original picture included for illustrative purposes, referring to ref. 23
A constitutively elevated level of H2AX-Y142 phosphorylation has been found in stem cells compared to differentiated counterparts23. The relationship between γ-H2AX IRIF, pan-γ-H2AX, and H2AX-pY142 has been previously elucidated149, suggesting that the close proximity of persistent H2AX-pY142 sterically hinders access to the S139 site in stem cells, thereby inhibiting DDR signaling and promoting IR-induced apoptosis.
Alterations in methylation levels after 3 Gy of IR have been reported in vivo150–152 and a correlation between global DNA hypomethylation and increased radiosensitivity has been observed in somatic cell lines127,153–155. On the contrary, exposure to 3 Gy X-irradiation does not lead to changes in DNA methylation in murine ES cells and radiosensitivity is independent of DNA methylation levels. However, de novo methyltransferases DNMT3A and DNMT3B may play a role in modulating sensitivity to X-rays in mESCs as their absence seems to have a modest radioprotective effect156. Moreover, DNA methylation status of 50 Long Interspersed Nuclear Element 1 (LINE-1) did not differ significantly in HSCs, hematopoietic progenitor cells, and mononuclear cells in C57BL/6J compared to radiosensitive CBA/J mice immediately after IR. In contrast, a significant decrease in LINE-1 DNA methylation in HSCs was observed in CBA/J 2 months post-treatment, suggesting that epigenetic alterations may potentially serve as driving forces of radiation-induced carcinogenesis157.
5-Hydroxylmethylcytosine (5hmC) is a potential indicator of active DNA demethylation and its distribution patterns provide a global view of gene activation. Murine pluripotent cells have been reported to have constitutively high levels of 5hmC158,159 together with elevated γ-H2AX foci160 while only a minor subset of γ-H2AX foci colocalized with 5hmC in human embryonic stem cells161,162. There is no correlation between 5hmC and radiation response in stem cells. While the role of epigenetics in stem cell pluripotency maintenance and self-renewal as well as differentiation has been largely investigated163, further investigations are needed to elucidate its role in stem-cell-specific IR response (Fig. 7).
Fig. 7. Graphical abstract summarizing the concept of this review.
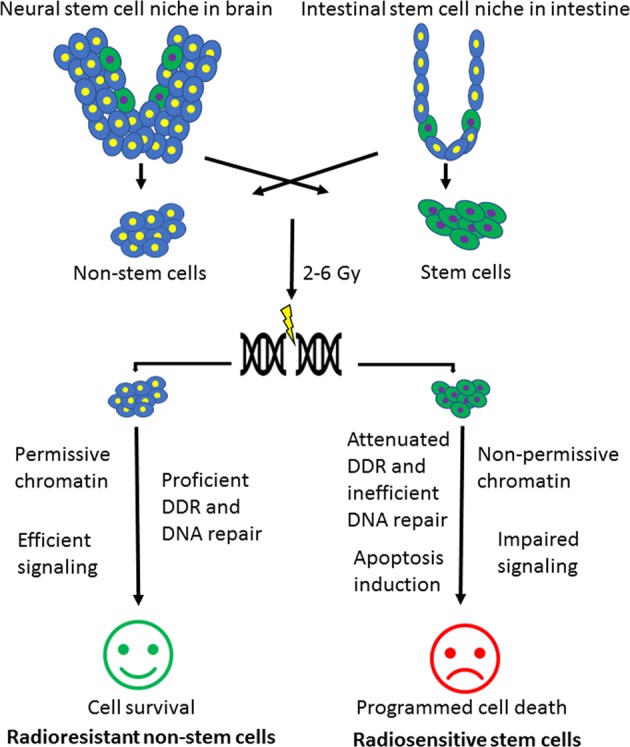
Stem cells reside inside tissue niches together with differentiated non-stem cells. After radiation therapy, non-stem cells respond with chromatin modifications which allows the damage site to become accessible to DDR and repairosome complex to repair the DNA damage. On the contrary, stem cells chromatin configuration remains non-permissive and DNA repair factors are not recruited on damage site. This attenuated DDR, impaired signaling, and induction of apoptosis leads to radiation hypersensitivity of stem cells
Conclusions
Radiation therapy treats many types of cancer effectively, but like other treatments, it can cause complications. Tissue injuries are observed in many cancer patients undergoing IR treatment and the preservation of healthy cells has become an important medical concern, especially in pediatric oncology. Stem cell transplants, although effective in hematopoietic cancer patients, have enormous practical limitations and therefore the protection of in loco radiosensitive stem cells seems to be a more feasible strategy. Our present knowledge on mechanisms of normal stem cell radiosensitivity is still evolving in a piecemeal manner, but clearly the stem cell-specific responses to IR exposure involve molecular genetic, signaling, and epigenetic regulations. Discovering the pluralistically interacting mechanisms and regulatory molecular targets involved in normal stem cell radiosensitivity could lead to innovative therapies that can eradicate cancer while preserving the stem/progenitor cells.
Acknowledgements
We would like to apologize to all colleagues whose work could not be cited due to space limitations. We acknowledge work of all previous members of the our laboratory. Funding was provided by NIH R01CA174966 and Department of Radiation Oncology of Washington University School of Medicine.
Conflict of interest
The authors declare that they have no conflict of interest.
Footnotes
Edited by I. Lavrik
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Orthwein A, et al. Mitosis inhibits DNA double-strand break repair to guard against telomere fusions. Science. 2014;344:189–193. doi: 10.1126/science.1248024. [DOI] [PubMed] [Google Scholar]
- 2.Biehs R, et al. DNA double-strand break resection occurs during non-homologous end joining in G1 but is distinct from resection during homologous recombination. Mol. Cell. 2017;65:671–684 e675. doi: 10.1016/j.molcel.2016.12.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Tsouroula K, et al. Temporal and spatial uncoupling of DNA double strand break repair pathways within mammalian heterochromatin. Mol. Cell. 2016;63:293–305. doi: 10.1016/j.molcel.2016.06.002. [DOI] [PubMed] [Google Scholar]
- 4.Campisi J, d'Adda di Fagagna F. Cellular senescence: when bad things happen to good cells. Nat. Rev. Mol. Cell Biol. 2007;8:729–740. doi: 10.1038/nrm2233. [DOI] [PubMed] [Google Scholar]
- 5.Edinger A, Thompson C. Death by design: apoptosis, necrosis and autophagy. Curr. Opin. Cell Biol. 2004;16:663–669. doi: 10.1016/j.ceb.2004.09.011. [DOI] [PubMed] [Google Scholar]
- 6.Heydarirad G, Rezaeizadeh H, Choopani R, Mosavat SH, Ameri A. Efficacy of a traditional Persian medicine preparation for radiation-induced xerostomia: a randomized, open-label, active-controlled trial. J. Integr. Med. 2017;15:201–208. doi: 10.1016/S2095-4964(17)60333-9. [DOI] [PubMed] [Google Scholar]
- 7.Ryan JL. Ionizing radiation: the good, the bad, and the ugly. J. Invest. Dermatol. 2012;132:985–993. doi: 10.1038/jid.2011.411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.McQuestion M. Evidence-based skin care management in radiation therapy. Semin. Oncol. Nurs. 2006;22:163–173. doi: 10.1016/j.soncn.2006.04.004. [DOI] [PubMed] [Google Scholar]
- 9.Hymes SR, Strom EA, Fife C. Radiation dermatitis: clinical presentation, pathophysiology, and treatment 2006. J. Am. Acad. Dermatol. 2006;54:28–46. doi: 10.1016/j.jaad.2005.08.054. [DOI] [PubMed] [Google Scholar]
- 10.Chang DW, te Marvelde L, Chua BH. Prospective study of local control and late radiation toxicity after intraoperative radiation therapy boost for early breast cancer. Int. J. Radiat. Oncol. Biol. Phys. 2014;88:73–79. doi: 10.1016/j.ijrobp.2013.09.049. [DOI] [PubMed] [Google Scholar]
- 11.Hoeller U, Bonacker M, Bajrovic A, Alberti W, Adam G. Radiation-induced plexopathy and fibrosis. Is magnetic resonance imaging the adequate diagnostic tool? Strahlenther. Onkol. 2004;180:650–654. doi: 10.1007/s00066-004-1240-3. [DOI] [PubMed] [Google Scholar]
- 12.Dorr W, Hendry JH. Consequential late effects in normal tissues. Radiother. Oncol. 2001;61:223–231. doi: 10.1016/S0167-8140(01)00429-7. [DOI] [PubMed] [Google Scholar]
- 13.Hellman S, Botnick L. Stem cell depletion: an explanation of the late effects of cytotoxins. Int. J. Radiat. Oncol. Biol. Phys. 1977;2:181–184. doi: 10.1016/0360-3016(77)90028-1. [DOI] [PubMed] [Google Scholar]
- 14.Thotala DK, Geng L, Dickey AK, Hallahan DE, Yazlovitskaya EM. A new class of molecular targeted radioprotectors: GSK-3beta inhibitors. Int. J. Radiat. Oncol. Biol. Phys. 2010;76:557–565. doi: 10.1016/j.ijrobp.2009.09.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kim HW, et al. Dual effects of human placenta-derived neural cells on neuroprotection and the inhibition of neuroinflammation in a rodent model of Parkinson's disease. Cell Transplant. 2018;27:814–830. doi: 10.1177/0963689718766324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Eve DJ, et al. Reduction of microhemorrhages in the spinal cord of symptomatic ALS mice after intravenous human bone marrow stem cell transplantation accompanies repair of the blood-spinal cord barrier. Oncotarget. 2018;9:10621–10634. doi: 10.18632/oncotarget.24360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Thomsen, G. M. et al. Transplantation of neural progenitor cells expressing glial cell line-derived neurotrophic factor into the motor cortex as a strategy to treat amyotrophic lateral sclerosis. Stem Cells10.1002/stem.2825 (2018). [DOI] [PubMed]
- 18.Yin Y, et al. The homing of human umbilical cord-derived mesenchymal stem cells and the subsequent modulation of macrophage polarization in type 2 diabetic mice. Int. Immunopharmacol. 2018;60:235–245. doi: 10.1016/j.intimp.2018.04.051. [DOI] [PubMed] [Google Scholar]
- 19.Mucci, A.et al. iPSC-derived macrophages effectively treat pulmonary alveolar proteinosis in Csf2rb-deficient mice. Stem Cell Rep. 10.1016/j.stemcr.2018.07.006 (2018). [DOI] [PMC free article] [PubMed]
- 20.Zhang, D. Y. et al. Sirtuin3 protects aged human mesenchymal stem cells against oxidative stress and enhances efficacy of cell therapy for ischaemic heart diseases. J. Cell Mol. Med.10.1111/jcmm.13821 (2018). [DOI] [PMC free article] [PubMed]
- 21.Acharya MM, Rosi S, Jopson T, Limoli CL. Human neural stem cell transplantation provides long-term restoration of neuronal plasticity in the irradiated hippocampus. Cell Transplant. 2015;24:691–702. doi: 10.3727/096368914X684600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Baulch JE, et al. Cranial grafting of stem cell-derived microvesicles improves cognition and reduces neuropathology in the irradiated brain. Proc. Natl Acad. Sci. USA. 2016;113:4836–4841. doi: 10.1073/pnas.1521668113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jacobs KM, et al. Unique epigenetic influence of H2AX phosphorylation and H3K56 acetylation on normal stem cell radioresponses. Mol. Biol. Cell. 2016;27:1332–1345. doi: 10.1091/mbc.E16-01-0017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Meyer B, et al. Histone H3 lysine 9 acetylation obstructs ATM activation and promotes ionizing radiation sensitivity in normal stem cells. Stem Cell Rep. 2016;7:1013–1022. doi: 10.1016/j.stemcr.2016.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Solokov, M. & Neumman, R. Human embryonic stem cell responses to ionizing radiation exposures: current state of knowledge and future challenges. Stem Cells Int. 2012, 579104 (2012). [DOI] [PMC free article] [PubMed]
- 26.Filion T, et al. Survival responses of human embryonic stem cells to DNA damage. J. Cell. Physiol. 2009;220:586–592. doi: 10.1002/jcp.21735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Acharya M, et al. Consequences of ionizing radiation-induced damage in human neural stem cells. Free Radic. Biol. Med. 2010;49:1846–1855. doi: 10.1016/j.freeradbiomed.2010.08.021. [DOI] [PubMed] [Google Scholar]
- 28.Katoh O, Tauchi H, Kawaishi K, Kimura A, Satow Y. Expression of the vascular endothelial growth factor (VEGF) receptor gene, KDR, in hematopoietic cells and inhibitory effect of VEGF on apoptotic cell death caused by ionizing radiation. Cancer Res. 1995;55:5687–5692. [PubMed] [Google Scholar]
- 29.Becker D, et al. Response of human hematopoietic stem and progenitor cells to energetic carbon ions. Int. J. Radiat. Biol. 2009;85:1051–1059. doi: 10.3109/09553000903232850. [DOI] [PubMed] [Google Scholar]
- 30.Fabbrizi MR, et al. Transient PP2A inhibition alleviates normal tissue stem cell susceptibility to cell death during radiotherapy. Cell Death Dis. 2018;9:492. doi: 10.1038/s41419-018-0559-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lomax ME, Folkes LK, O'Neill P. Biological consequences of radiation-induced DNA damage: relevance to radiotherapy. Clin. Oncol. (R. Coll. Radiol.) 2013;25:578–585. doi: 10.1016/j.clon.2013.06.007. [DOI] [PubMed] [Google Scholar]
- 32.Close DM, Nelson WH, Bernhard WA. DNA damage by the direct effect of ionizing radiation: products produced by two sequential one-electron oxidations. J. Phys. Chem. A. 2013;117:12608–12615. doi: 10.1021/jp4084844. [DOI] [PubMed] [Google Scholar]
- 33.Katsube T, et al. Effects of chronic restraint-induced stress on radiation-induced chromosomal aberrations in mouse splenocytes. Mutat. Res. 2017;813:18–26. doi: 10.1016/j.mrgentox.2016.11.005. [DOI] [PubMed] [Google Scholar]
- 34.Balajee AS, Bertucci A, Taveras M, Brenner DJ. Multicolour FISH analysis of ionising radiation induced micronucleus formation in human lymphocytes. Mutagenesis. 2014;29:447–455. doi: 10.1093/mutage/geu041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Fajardo LF. The pathology of ionizing radiation as defined by morphologic patterns. Acta Oncol. 2005;44:13–22. doi: 10.1080/02841860510007440. [DOI] [PubMed] [Google Scholar]
- 36.Denham JW, Hauer-Jensen M. The radiotherapeutic injury—a complex 'wound'. Radiother. Oncol. 2002;63:129–145. doi: 10.1016/S0167-8140(02)00060-9. [DOI] [PubMed] [Google Scholar]
- 37.Kalman NS, Zhao SS, Anscher MS, Urdaneta AI. Current status of targeted radioprotection and radiation injury mitigation and treatment agents: a critical review of the literature. Int. J. Radiat. Oncol. Biol. Phys. 2017;98:662–682. doi: 10.1016/j.ijrobp.2017.02.211. [DOI] [PubMed] [Google Scholar]
- 38.Hall, E. J. & Giaccia, A. J. Radiobiology for the Radiologist. 6th edn (Lippincott Williams & Wilkins, 2006).
- 39.Withers HR. The dose-survival relationship for irradiation of epithelial cells of mouse skin. Br. J. Radiol. 1967;40:187–194. doi: 10.1259/0007-1285-40-471-187. [DOI] [PubMed] [Google Scholar]
- 40.Withers HR. Regeneration of intestinal mucosa after irradiation. Cancer. 1971;28:75–81. doi: 10.1002/1097-0142(197107)28:1<75::AID-CNCR2820280115>3.0.CO;2-8. [DOI] [PubMed] [Google Scholar]
- 41.Withers HR, Hunter N, Barkley HT, Jr, Reid BO. Radiation survival and regeneration characteristics of spermatogenic stem cells of mouse testis. Radiat. Res. 1974;57:88–103. doi: 10.2307/3573759. [DOI] [PubMed] [Google Scholar]
- 42.Withers HR, Mason KA, Thames HD., Jr. Late radiation response of kidney assayed by tubule-cell survival. Br. J. Radiol. 1986;59:587–595. doi: 10.1259/0007-1285-59-702-587. [DOI] [PubMed] [Google Scholar]
- 43.McCulloch EA, Till JE. The sensitivity of cells from normal mouse bone marrow to gamma radiation in vitro and in vivo. Radiat. Res. 1962;16:822–832. doi: 10.2307/3571281. [DOI] [PubMed] [Google Scholar]
- 44.Gould MN, Biel WF, Clifton KH. Morphological and quantitative studies of gland formation from inocula of monodispersed rat mammary cells. Exp. Cell Res. 1977;107:405–416. doi: 10.1016/0014-4827(77)90362-7. [DOI] [PubMed] [Google Scholar]
- 45.Coppes RP, van der Goot A, Lombaert IM. Stem cell therapy to reduce radiation-induced normal tissue damage. Semin. Radiat. Oncol. 2009;19:112–121. doi: 10.1016/j.semradonc.2008.11.005. [DOI] [PubMed] [Google Scholar]
- 46.Donnelly EH, Smith JM, Farfan EB, Ozcan I. Prenatal radiation exposure: background material for counseling pregnant patients following exposure to radiation. Disaster Med. Public Health Prep. 2011;5:62–68. doi: 10.1001/dmp.2011.2. [DOI] [PubMed] [Google Scholar]
- 47.De Santis M, et al. Radiation effects on development. Birth Defects Res. C Embryo Today. 2007;81:177–182. doi: 10.1002/bdrc.20099. [DOI] [PubMed] [Google Scholar]
- 48.Brent RL. Protection of the gametes embryo/fetus from prenatal radiation exposure. Health Phys. 2015;108:242–274. doi: 10.1097/HP.0000000000000235. [DOI] [PubMed] [Google Scholar]
- 49.Shaw P, Duncan A, Vouyouka A, Ozsvath K. Radiation exposure and pregnancy. J. Vasc. Surg. 2011;53:28S–34S. doi: 10.1016/j.jvs.2010.05.140. [DOI] [PubMed] [Google Scholar]
- 50.Brent RL. Clinical teratology counseling and consultation case report: exposure to diagnostic radiation early in pregnancy. Teratology. 1992;46:31–33. doi: 10.1002/tera.1420460106. [DOI] [PubMed] [Google Scholar]
- 51.Wakeford R, Little MP. Risk coefficients for childhood cancer after intrauterine irradiation: a review. Int. J. Radiat. Biol. 2003;79:293–309. doi: 10.1080/0955300031000114729. [DOI] [PubMed] [Google Scholar]
- 52.Momcilovic O, et al. Ionizing radiation induces ataxia telangiectasia mutated-dependent checkpoint signaling and G2 but not G1 cell cycle arrest in pluripotent human embryonic stem cells. Stem Cells. 2009;27:1822–1835. doi: 10.1002/stem.123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Suvorova II, Kozhukharova IV, Nikol'skii NN, Pospelov VA. [ATM/ATR signaling pathway activation in human embryonic stem cells after DNA damage] Tsitologiia. 2013;55:841–851. [PubMed] [Google Scholar]
- 54.Sokolov M, Panyutin I, Panyutin I, Neumann R. Dynamics of the transcriptome response of cultured human embryonic stem cells to ionizing radiation exposure. Mutat. Res. 2011;709-710:40–48. doi: 10.1016/j.mrfmmm.2011.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Adams B, Golding S, Rao R, Valerie K. Dynamic dependence on ATR and ATM for double-strand break repair in human embryonic stem cells and neural descendants. PLoS ONE. 2010;5:e10001. doi: 10.1371/journal.pone.0010001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Suchorska WM, Augustyniak E, Lukjanow M. Comparison of the early response of human embryonic stem cells and human induced pluripotent stem cells to ionizing radiation. Mol. Med. Rep. 2017;15:1952–1962. doi: 10.3892/mmr.2017.6270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Barazzuol L, Jeggo PA. In vivo sensitivity of the embryonic and adult neural stem cell compartments to low-dose radiation. J. Radiat. Res. 2016;57(Suppl 1):i2–i10. doi: 10.1093/jrr/rrw013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Dumitru R, et al. Human embryonic stem cells have constitutively active Bax at the Golgi and are primed to undergo rapid apoptosis. Mol. Cell. 2012;46:573–583. doi: 10.1016/j.molcel.2012.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Mujoo K, et al. Differentiation of human induced pluripotent or embryonic stem cells decreases the DNA damage repair by homologous recombination. Stem Cell Rep. 2017;9:1660–1674. doi: 10.1016/j.stemcr.2017.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Wilson K, et al. Effects of ionizing radiation on self-renewal and pluripotency of human embryonic stem cells. Cancer Res. 2010;70:5539–5548. doi: 10.1158/0008-5472.CAN-09-4238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Yang C, et al. Opposing putative roles for canonical and noncanonical NFkappaB signaling on the survival, proliferation, and differentiation potential of human embryonic stem cells. Stem Cells. 2010;28:1970–1980. doi: 10.1002/stem.528. [DOI] [PubMed] [Google Scholar]
- 62.Bai H, et al. Bcl-xL enhances single-cell survival and expansion of human embryonic stem cells without affecting self-renewal. Stem Cell Res. 2012;8:26–37. doi: 10.1016/j.scr.2011.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Conklin J, Baker J, Sage J. The RB family is required for the self-renewal and survival of human embryonic stem cells. Nat. Commun. 2012;3:1244. doi: 10.1038/ncomms2254. [DOI] [PubMed] [Google Scholar]
- 64.Ardehali R, et al. Overexpression of BCL2 enhances survival of human embryonic stem cells during stress and obviates the requirement for serum factors. Proc. Natl Acad. Sci. USA. 2011;108:3282–3287. doi: 10.1073/pnas.1019047108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Edel M, et al. Rem2 GTPase maintains survival of human embryonic stem cells as well as enhancing reprogramming by regulating p53 and cyclin D1. Genes Dev. 2010;24:561–573. doi: 10.1101/gad.1876710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Vinarsky V, et al. Human embryonic and induced pluripotent stem cells express TRAIL receptors and can be sensitized to TRAIL-induced apoptosis. Stem. Cells Dev. 2013;22:2964–2974. doi: 10.1089/scd.2013.0057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Zhang L, et al. Activated mitochondrial apoptosis in hESCs after dissociation involving the PKA/p-p53/Bax signaling pathway. Exp. Cell Res. 2018;369:226–233. doi: 10.1016/j.yexcr.2018.05.024. [DOI] [PubMed] [Google Scholar]
- 68.Cmielova J, et al. Gamma radiation induces senescence in human adult mesenchymal stem cells from bone marrow and periodontal ligaments. Int. J. Radiat. Biol. 2012;88:393–404. doi: 10.3109/09553002.2012.666001. [DOI] [PubMed] [Google Scholar]
- 69.Ko E, Lee K, Hwang D. Human umbilical cord blood-derived mesenchymal stem cells undergo cellular senescence in response to oxidative stress. Stem. Cells Dev. 2012;21:1877–1886. doi: 10.1089/scd.2011.0284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Havelek R, et al. Ionizing radiation induces senescence and differentiation of human dental pulp stem cells. Folia Biol. (Krakow) 2013;59:188–197. doi: 10.14712/fb2013059050188. [DOI] [PubMed] [Google Scholar]
- 71.Sergeeva VA, et al. Low-dose ionizing radiation affects mesenchymal stem cells via extracellular oxidized cell-free DNA: a possible mediator of bystander effect and adaptive response. Oxid. Med. Cell Longev. 2017;2017:9515809. doi: 10.1155/2017/9515809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Nicolay N, et al. Mesenchymal stem cells retain their defining stem cell characteristics after exposure to ionizing radiation. Int. J. Radiat. Oncol. Biol. Phys. 2013;87:1171–1178. doi: 10.1016/j.ijrobp.2013.09.003. [DOI] [PubMed] [Google Scholar]
- 73.Nicolay N, et al. Mesenchymal stem cells are resistant to carbon ion radiotherapy. Oncotarget. 2015;6:2076–2087. doi: 10.18632/oncotarget.2857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Alessio N, et al. Low dose radiation induced senescence of human mesenchymal stromal cells and impaired the autophagy process. Oncotarget. 2015;6:8155–8166. doi: 10.18632/oncotarget.2692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Li XH, Ha CT, Xiao M. MicroRNA-30 inhibits antiapoptotic factor Mcl-1 in mouse and human hematopoietic cells after radiation exposure. Apoptosis. 2016;21:708–720. doi: 10.1007/s10495-016-1238-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Prise KM, Saran A. Concise review: stem cell effects in radiation risk. Stem Cells. 2011;29:1315–1321. doi: 10.1002/stem.690. [DOI] [PubMed] [Google Scholar]
- 77.Squillaro T, Galano G, De Rosa R, Peluso G, Galderisi U. Concise review: the effect of low-dose ionizing radiation on stem cell biology: a contribution to radiation risk. Stem Cells. 2018 doi: 10.1002/stem.2836. [DOI] [PubMed] [Google Scholar]
- 78.Fuchs E. The tortoise and the hair: slow-cycling cells in the stem cell race. Cell. 2009;137:811–819. doi: 10.1016/j.cell.2009.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Cheung TH, Rando TA. Molecular regulation of stem cell quiescence. Nat. Rev. Mol. Cell Biol. 2013;14:329–340. doi: 10.1038/nrm3591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Kim JS, et al. Independent and sequential recruitment of NHEJ and HR factors to DNA damage sites in mammalian cells. J. Cell Biol. 2005;170:341–347. doi: 10.1083/jcb.200411083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Delacote F, Lopez BS. Importance of the cell cycle phase for the choice of the appropriate DSB repair pathway, for genome stability maintenance: the trans-S double-strand break repair model. Cell Cycle. 2008;7:33–38. doi: 10.4161/cc.7.1.5149. [DOI] [PubMed] [Google Scholar]
- 82.Mao Z, Bozzella M, Seluanov A, Gorbunova V. Comparison of nonhomologous end joining and homologous recombination in human cells. DNA Repair (Amst.). 2008;7:1765–1771. doi: 10.1016/j.dnarep.2008.06.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Georgoulis, A., Vorgias, C. E., Chrousos, G. P. & Rogakou, E. P. Genome Instability and gammaH2AX. Int. J. Mol. Sci. 18, 10.3390/ijms18091979 (2017). [DOI] [PMC free article] [PubMed]
- 84.Chen M, et al. The sensitivity of human mesenchymal stem cells to ionizing radiation. Int. J. Radiat. Oncol. Biol. Phys. 2006;66:244–253. doi: 10.1016/j.ijrobp.2006.03.062. [DOI] [PubMed] [Google Scholar]
- 85.Prendergast A, Cruet-Hennequart S, Shaw G, Barry F, Carty M. Activation of DNA damage response pathways in human mesenchymal stem cells exposed to cisplatin or gamma-irradiation. Cell Cycle. 2011;10:3768–3777. doi: 10.4161/cc.10.21.17972. [DOI] [PubMed] [Google Scholar]
- 86.Oliver L, et al. Differentiation-related response to DNA breaks in human mesenchymal stem cells. Stem Cells. 2013;31:800–807. doi: 10.1002/stem.1336. [DOI] [PubMed] [Google Scholar]
- 87.Wu PK, et al. Early passage mesenchymal stem cells display decreased radiosensitivity and increased DNA repair activity. Stem Cells Transl. Med. 2017;6:1504–1514. doi: 10.1002/sctm.15-0394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Sokolov M, Neumann R. Lessons learned about human stem cell responses to ionizing radiation exposures: a long road still ahead of us. Int. J. Mol. Sci. 2013;14:15695–15723. doi: 10.3390/ijms140815695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Sugrue T, Lowndes NF, Ceredig R. Hypoxia enhances the radioresistance of mouse mesenchymal stromal cells. Stem Cells. 2014;32:2188–2200. doi: 10.1002/stem.1683. [DOI] [PubMed] [Google Scholar]
- 90.Islam MS, Stemig ME, Takahashi Y, Hui SK. Radiation response of mesenchymal stem cells derived from bone marrow and human pluripotent stem cells. J. Radiat. Res. 2015;56:269–277. doi: 10.1093/jrr/rru098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Sugrue T, Lowndes N, Ceredig R. Mesenchymal stromal cells: radio-resistant members of the bone marrow. Immunol. Cell Biol. 2013;91:5–11. doi: 10.1038/icb.2012.61. [DOI] [PubMed] [Google Scholar]
- 92.Serakinci N, et al. Ectopically hTERT expressing adult human mesenchymal stem cells are less radiosensitive than their telomerase negative counterpart. Exp. Cell Res. 2007;313:1056–1067. doi: 10.1016/j.yexcr.2007.01.002. [DOI] [PubMed] [Google Scholar]
- 93.Tsvetkova A, et al. gammaH2AX, 53BP1 and Rad51 protein foci changes in mesenchymal stem cells during prolonged X-ray irradiation. Oncotarget. 2017;8:64317–64329. doi: 10.18632/oncotarget.19203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Milyavsky M, et al. A distinctive DNA damage response in human hematopoietic stem cells reveals an apoptosis-independent role for p53 in self-renewal. Cell Stem Cell. 2010;7:186–197. doi: 10.1016/j.stem.2010.05.016. [DOI] [PubMed] [Google Scholar]
- 95.Yahata T, et al. Accumulation of oxidative DNA damage restricts the self-renewal capacity of human hematopoietic stem cells. Blood. 2011;118:2941–2950. doi: 10.1182/blood-2011-01-330050. [DOI] [PubMed] [Google Scholar]
- 96.Li X, et al. Binding to WGR domain by salidroside activates PARP1 and protects hematopoietic stem cells from oxidative stress. Antioxid. Redox Signal. 2014;20:1853–1865. doi: 10.1089/ars.2013.5600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Biechonski S, et al. Attenuated DNA damage responses and increased apoptosis characterize human hematopoietic stem cells exposed to irradiation. Sci. Rep. 2018;8:6071. doi: 10.1038/s41598-018-24440-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.de Laval B, et al. Thrombopoietin-increased DNA-PK-dependent DNA repair limits hematopoietic stem and progenitor cell mutagenesis in response to DNA damage. Cell Stem Cell. 2013;12:37–48. doi: 10.1016/j.stem.2012.10.012. [DOI] [PubMed] [Google Scholar]
- 99.Aoki H, Hara A, Motohashi T, Kunisada T. Keratinocyte stem cells but not melanocyte stem cells are the primary target for radiation-induced hair graying. J. Invest. Dermatol. 2013;133:2143–2151. doi: 10.1038/jid.2013.155. [DOI] [PubMed] [Google Scholar]
- 100.Rachidi W, et al. Sensing radiosensitivity of human epidermal stem cells. Radiother. Oncol. 2007;83:267–276. doi: 10.1016/j.radonc.2007.05.007. [DOI] [PubMed] [Google Scholar]
- 101.Shao L, Luo Y, Zhou D. Hematopoietic stem cell injury induced by ionizing radiation. Antioxid. Redox Signal. 2014;20:1447–1462. doi: 10.1089/ars.2013.5635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Moehrle BM, et al. Stem cell-specific mechanisms ensure genomic fidelity within HSCs and upon aging of HSCs. Cell Rep. 2015;13:2412–2424. doi: 10.1016/j.celrep.2015.11.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Ishikawa J, Hayashi N, Yamaguchi M, Monzen S, Kashiwakura I. Characteristics of human CD34+ cells exposed to ionizing radiation under cytokine-free conditions. J. Radiat. Res. 2015;56:678–690. doi: 10.1093/jrr/rrv024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Wang Y, et al. Total body irradiation causes residual bone marrow injury by induction of persistent oxidative stress in murine hematopoietic stem cells. Free Radic. Biol. Med. 2010;48:348–356. doi: 10.1016/j.freeradbiomed.2009.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Wang Y, Kellner J, Liu L, Zhou D. Inhibition of p38 mitogen-activated protein kinase promotes ex vivo hematopoietic stem cell expansion. Stem. Cells Dev. 2011;20:1143–1152. doi: 10.1089/scd.2010.0413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Chang J, et al. Low doses of oxygen ion irradiation cause acute damage to hematopoietic cells in mice. PLoS ONE. 2016;11:e0158097. doi: 10.1371/journal.pone.0158097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Miousse IR, et al. Exposure to low-dose (56)Fe-ion radiation induces long-term epigenetic alterations in mouse bone marrow hematopoietic progenitor and stem cells. Radiat. Res. 2014;182:92–101. doi: 10.1667/RR13580.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Katsura M, et al. Effects of chronic low-dose radiation on human neural progenitor cells. Sci. Rep. 2016;6:20027. doi: 10.1038/srep20027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Zou Y, et al. Responses of human embryonic stem cells and their differentiated progeny to ionizing radiation. Biochem. Biophys. Res. Commun. 2012;426:100–105. doi: 10.1016/j.bbrc.2012.08.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Li, Y. Q., Cheng, Z. & Wong, S. Differential apoptosis radiosensitivity of neural progenitors in adult mouse hippocampus. Int. J. Mol. Sci. 17, 10.3390/ijms17060970 (2016). [DOI] [PMC free article] [PubMed]
- 111.Ivanov VN, Hei TK. Radiation-induced glioblastoma signaling cascade regulates viability, apoptosis and differentiation of neural stem cells (NSC) Apoptosis. 2014;19:1736–1754. doi: 10.1007/s10495-014-1040-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Ivanov VN, Hei TK. A role for TRAIL/TRAIL-R2 in radiation-induced apoptosis and radiation-induced bystander response of human neural stem cells. Apoptosis. 2014;19:399–413. doi: 10.1007/s10495-013-0925-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Isono M, et al. Carbon-ion beams effectively induce growth inhibition and apoptosis in human neural stem cells compared with glioblastoma A172 cells. J. Radiat. Res. 2015;56:856–861. doi: 10.1093/jrr/rrv033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Wang X, et al. Pharmacologically blocking p53-dependent apoptosis protects intestinal stem cells and mice from radiation. Sci. Rep. 2015;5:8566. doi: 10.1038/srep08566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Zhu Y, Huang YF, Kek C, Bulavin DV. Apoptosis differently affects lineage tracing of Lgr5 and Bmi1 intestinal stem cell populations. Cell Stem Cell. 2013;12:298–303. doi: 10.1016/j.stem.2013.01.003. [DOI] [PubMed] [Google Scholar]
- 116.Metcalfe C, Kljavin NM, Ybarra R, de Sauvage FJ. Lgr5+ stem cells are indispensable for radiation-induced intestinal regeneration. Cell Stem Cell. 2014;14:149–159. doi: 10.1016/j.stem.2013.11.008. [DOI] [PubMed] [Google Scholar]
- 117.Hua G, et al. Crypt base columnar stem cells in small intestines of mice are radioresistant. Gastroenterology. 2012;143:1266–1276. doi: 10.1053/j.gastro.2012.07.106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Cmielova J, et al. The effect of ATM kinase inhibition on the initial response of human dental pulp and periodontal ligament mesenchymal stem cells to ionizing radiation. Int. J. Radiat. Biol. 2013;89:501–511. doi: 10.3109/09553002.2013.775530. [DOI] [PubMed] [Google Scholar]
- 119.Kurpinski K, et al. Differential effects of x-rays and high-energy 56Fe ions on human mesenchymal stem cells. Int. J. Radiat. Oncol. Biol. Phys. 2009;73:869–877. doi: 10.1016/j.ijrobp.2008.10.002. [DOI] [PubMed] [Google Scholar]
- 120.Jin Y, et al. Comprehensive analysis of time- and dose-dependent patterns of gene expression in a human mesenchymal stem cell line exposed to low-dose ionizing radiation. Oncol. Rep. 2008;19:135–144. [PubMed] [Google Scholar]
- 121.Liao Y, et al. Paradoxical roles of elongation factor-2 kinase in stem cell survival. J. Biol. Chem. 2016;291:19545–19557. doi: 10.1074/jbc.M116.724856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Gurley KE, Ashley AK, Moser RD, Kemp CJ. Synergy between Prkdc and Trp53 regulates stem cell proliferation and GI-ARS after irradiation. Cell Death Differ. 2017;24:1853–1860. doi: 10.1038/cdd.2017.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Wei L, et al. Inhibition of CDK4/6 protects against radiation-induced intestinal injury in mice. J. Clin. Invest. 2016;126:4076–4087. doi: 10.1172/JCI88410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Tao S, et al. Wnt activity and basal niche position sensitize intestinal stem and progenitor cells to DNA damage. EMBO J. 2017;36:2920–2921. doi: 10.15252/embj.201797813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Hua G, et al. Distinct levels of radioresistance in Lgr5+ colonic epithelial stem cells versus Lgr5+ small intestinal stem cells. Cancer Res. 2017;77:2124–2133. doi: 10.1158/0008-5472.CAN-15-2870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Chowdhury D, et al. gamma-H2AX dephosphorylation by protein phosphatase 2A facilitates DNA double-strand break repair. Mol. Cell. 2005;20:801–809. doi: 10.1016/j.molcel.2005.10.003. [DOI] [PubMed] [Google Scholar]
- 127.Aypar U, Morgan WF, Baulch JE. Radiation-induced genomic instability: are epigenetic mechanisms the missing link? Int. J. Radiat. Biol. 2011;87:179–191. doi: 10.3109/09553002.2010.522686. [DOI] [PubMed] [Google Scholar]
- 128.Kovalchuk O, Baulch JE. Epigenetic changes and nontargeted radiation effects—is there a link? Environ. Mol. Mutagen. 2008;49:16–25. doi: 10.1002/em.20361. [DOI] [PubMed] [Google Scholar]
- 129.Li L, et al. Effect of ionizing radiation at low dose on transgenerational carcinogenesis by epigenetic regulation. Lab. Anim. Res. 2017;33:92–97. doi: 10.5625/lar.2017.33.2.92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Song W, et al. Increased p16 DNA methylation in mouse thymic lymphoma induced by irradiation. PLoS ONE. 2014;9:e93850. doi: 10.1371/journal.pone.0093850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Yamazaki J, et al. The epigenome of AML stem and progenitor cells. Epigenetics. 2013;8:92–104. doi: 10.4161/epi.23243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Wakita S, et al. Mutations of the epigenetics-modifying gene (DNMT3a, TET2, IDH1/2) at diagnosis may induce FLT3-ITD at relapse in de novo acute myeloid leukemia. Leukemia. 2013;27:1044–1052. doi: 10.1038/leu.2012.317. [DOI] [PubMed] [Google Scholar]
- 133.Shih A, Abdel-Wahab O, Patel J, Levine R. The role of mutations in epigenetic regulators in myeloid malignancies. Nat. Rev. Cancer. 2012;12:599–612. doi: 10.1038/nrc3343. [DOI] [PubMed] [Google Scholar]
- 134.Holz-Schietinger C, Matje D, Reich N. Mutations in DNA methyltransferase (DNMT3A) observed in acute myeloid leukemia patients disrupt processive methylation. J. Biol. Chem. 2012;287:30941–30951. doi: 10.1074/jbc.M112.366625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Harikumar A, Meshorer E. Chromatin remodeling and bivalent histone modifications in embryonic stem cells. EMBO Rep. 2015;16:1609–1619. doi: 10.15252/embr.201541011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Gładych M, Andrzejewska A, Oleksiewicz U, Estécio M. Epigenetic mechanisms of induced pluripotency. Contemp. Oncol. (Pozn.) 2015;Q19:A30–38. doi: 10.5114/wo.2014.47135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Shiraki N, Ogaki S, Kume S. Profiling of embryonic stem cell differentiation. Rev. Diabet. Stud. 2014;11:102–114. doi: 10.1900/RDS.2014.11.102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Kraushaar D, Zhao K. The epigenomics of embryonic stem cell differentiation. Int. J. Biol. Sci. 2013;9:1134–1144. doi: 10.7150/ijbs.7998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Liang G, Zhang Y. Embryonic stem cell and induced pluripotent stem cell: an epigenetic perspective. Cell Res. 2013;23:49–69. doi: 10.1038/cr.2012.175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Ben-David U, Kopper O, Benvenisty N. Expanding the boundaries of embryonic stem cells. Cell Stem Cell. 2012;10:666–677. doi: 10.1016/j.stem.2012.05.003. [DOI] [PubMed] [Google Scholar]
- 141.Etchegaray JP, et al. The histone deacetylase SIRT6 controls embryonic stem cell fate via TET-mediated production of 5-hydroxymethylcytosine. Nat. Cell Biol. 2015;17:545–557. doi: 10.1038/ncb3147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Qiao Y, Wang R, Yang X, Tang K, Jing N. Dual roles of histone H3 lysine 9 acetylation in human embryonic stem cell pluripotency and neural differentiation. J. Biol. Chem. 2015;290:2508–2520. doi: 10.1074/jbc.M114.603761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Hammoud SS, et al. Chromatin and transcription transitions of mammalian adult germline stem cells and spermatogenesis. Cell Stem Cell. 2014;15:239–253. doi: 10.1016/j.stem.2014.04.006. [DOI] [PubMed] [Google Scholar]
- 144.Bartova E, et al. Recruitment of Oct4 protein to UV-damaged chromatin in embryonic stem cells. PLoS ONE. 2011;6:e27281. doi: 10.1371/journal.pone.0027281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Ayrapetov MK, Gursoy-Yuzugullu O, Xu C, Xu Y, Price BD. DNA double-strand breaks promote methylation of histone H3 on lysine 9 and transient formation of repressive chromatin. Proc. Natl Acad. Sci. USA. 2014;111:9169–9174. doi: 10.1073/pnas.1403565111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Sun Y, et al. Histone H3 methylation links DNA damage detection to activation of the tumour suppressor Tip60. Nat. Cell Biol. 2009;11:1376–1382. doi: 10.1038/ncb1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Zhu Q, et al. Damaged DNA-binding protein down-regulates epigenetic mark H3K56Ac through histone deacetylase 1 and 2. Mutat. Res. 2015;776:16–23. doi: 10.1016/j.mrfmmm.2015.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Miller KM, et al. Human HDAC1 and HDAC2 function in the DNA-damage response to promote DNA nonhomologous end-joining. Nat. Struct. Mol. Biol. 2010;17:1144–1151. doi: 10.1038/nsmb.1899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Solier S, Pommier Y. The apoptotic ring: a novel entity with phosphorylated histones H2AX and H2B and activated DNA damage response kinases. Cell Cycle. 2009;8:1853–1859. doi: 10.4161/cc.8.12.8865. [DOI] [PubMed] [Google Scholar]
- 150.Giotopoulos G, et al. DNA methylation during mouse hemopoietic differentiation and radiation-induced leukemia. Exp. Hematol. 2006;34:1462–1470. doi: 10.1016/j.exphem.2006.06.008. [DOI] [PubMed] [Google Scholar]
- 151.Pogribny I, Raiche J, Slovack M, Kovalchuk O. Dose-dependence, sex- and tissue-specificity, and persistence of radiation-induced genomic DNA methylation changes. Biochem. Biophys. Res. Commun. 2004;320:1253–1261. doi: 10.1016/j.bbrc.2004.06.081. [DOI] [PubMed] [Google Scholar]
- 152.Wang J, et al. Genome-wide screen of DNA methylation changes induced by low dose X-ray radiation in mice. PLoS ONE. 2014;9:e90804. doi: 10.1371/journal.pone.0090804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Dote H, et al. Enhancement of in vitro and in vivo tumor cell radiosensitivity by the DNA methylation inhibitor zebularine. Clin. Cancer Res. 2005;11:4571–4579. doi: 10.1158/1078-0432.CCR-05-0050. [DOI] [PubMed] [Google Scholar]
- 154.Narayan A, Tuck-Muller C, Weissbecker K, Smeets D, Ehrlich M. Hypersensitivity to radiation-induced non-apoptotic and apoptotic death in cell lines from patients with the ICF chromosome instability syndrome. Mutat. Res. 2000;456:1–15. doi: 10.1016/S0027-5107(00)00103-2. [DOI] [PubMed] [Google Scholar]
- 155.Aypar U, Morgan WF, Baulch JE. Radiation-induced epigenetic alterations after low and high LET irradiations. Mutat. Res. 2011;707:24–33. doi: 10.1016/j.mrfmmm.2010.12.003. [DOI] [PubMed] [Google Scholar]
- 156.Armstrong CA, et al. DNMTs are required for delayed genome instability caused by radiation. Epigenetics. 2012;7:892–902. doi: 10.4161/epi.21094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Miousse, I. R. et al. Inter-strain differences in LINE-1 DNA methylation in the mousehematopoietic system in response to exposure to ionizing radiation. Int. J. Mol. Sci.18, 10.3390/ijms18071430 (2017). [DOI] [PMC free article] [PubMed]
- 158.Ruzov A, et al. Lineage-specific distribution of high levels of genomic 5-hydroxymethylcytosine in mammalian development. Cell Res. 2011;21:1332–1342. doi: 10.1038/cr.2011.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Tahiliani M, et al. Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science. 2009;324:930–935. doi: 10.1126/science.1170116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Banath JP, et al. Explanation for excessive DNA single-strand breaks and endogenous repair foci in pluripotent mouse embryonic stem cells. Exp. Cell Res. 2009;315:1505–1520. doi: 10.1016/j.yexcr.2008.12.007. [DOI] [PubMed] [Google Scholar]
- 161.Kafer GR, et al. 5-Hydroxymethylcytosine marks sites of DNA damage and promotes genome stability. Cell Rep. 2016;14:1283–1292. doi: 10.1016/j.celrep.2016.01.035. [DOI] [PubMed] [Google Scholar]
- 162.Takashima Y, et al. Resetting transcription factor control circuitry toward ground-state pluripotency in human. Cell. 2014;158:1254–1269. doi: 10.1016/j.cell.2014.08.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Festuccia N, Gonzalez I, Navarro P. The epigenetic paradox of pluripotent ES cells. J. Mol. Biol. 2017;429:1476–1503. doi: 10.1016/j.jmb.2016.12.009. [DOI] [PubMed] [Google Scholar]