

Abstract
Memory for related past experiences can guide current perceptions. However, memory can lead one astray if situational features have changed. Thus, to adaptively use memory to guide perception, one needs to retrieve relevant memories and also to register differences between remembered and current events. Event Memory Retrieval and Comparison Theory proposes that observers associatively activate memories of related previous episodes, and that this guides their ongoing perception. Conflicts between previous and current event features can hurt immediate performance, but if changes are registered and encoded they can lead to highly effective encoding of the prior event, current event, and their relationship. Disruption of these mechanisms could play a role in older adults’ greater susceptibility to event memory interference. Two experiments tested these hypotheses by asking participants to watch movies depicting two fictive days of an actor. Some activities were repeated across days, others were repeated with a changed feature (e.g., waking up to an alarm clock or a phone alarm), and others were performed only on Day 2. One week after watching the Day 2 movie, participants completed a cued recall test. Changes that participants detected but did not remember led to proactive interference in recall, but changes that were successfully detected and remembered led to facilitation. Younger adults detected and remembered more changes than older adults, which partly explained older adults’ differential memory deficit for changed activities. These findings suggest a role for episodic reminding in event perception and a potential source of age differences in event memory.
Keywords: Change Detection, Event Cognition, Interference, Prediction Error, Updating
Many of the activities people experience are not brand new, and not completely novel, but are near repetitions—variations on a theme. For example, imagine that you have a friend who ordered a cheeseburger for dinner the last time that you ate together. After that occasion, your friend discovered a family history of heart disease. When you next have dinner with this friend, you might predict that she would place a similar order as on the last occasion. However, you experience a prediction error when she orders a salad. Registering this change, consciously or not, would help you to better predict her behavior the next time you eat together. The processing associated with registering the change might also affect your encoding of this particular episode. In this paper, we develop a theoretical framework that proposes a mechanism by which people use memory for recent past events to guide their perception of the present, with consequences for subsequent memory. This framework is based on a combination of previous studies of memory for change and previous research on event cognition. It leads to predictions about how memory for changes in events is affected by aging, which we tested in two experiments. Before describing the framework and the present experiments, we describe their origins in studies of event perception, episodic memory, and cognitive aging.
Event Perception and Memory
People perceive naturalistic ongoing activities as discrete events. Theories of event perception hold that everyday activities are represented hierarchically (e.g., Dickman, 1963; Brewer & Dupree, 1983; Zacks & Tversky, 2001), such that smaller units of activity are nested within larger units of activity. For example, the event described above (eating dinner) is comprised of smaller events such as ordering food and eating it, which in turn might be broken down further into events such as picking up a fork, cutting a burger, and taking a drink from a glass of water. Understanding structure in events allows individuals to establish better organized representations of what is happening in their current environment. People spontaneously segment activities during event perception (e.g., Zacks et al., 2001), and this is related to their subsequent memory: Features from event boundaries are often remembered better than event middles (Newtson & Engquist, 1976; Schwan, Garsoffky, & Hesse, 2000; Swallow, Zacks, & Abrams, 2009), individual differences in event segmentation predict individual differences in event memory (Flores, Bailey, Eisenberg, & Zacks, 2017; Sargent et al., 2013; Zacks, Speer, Vettel, & Jacoby, 2006), and interventions to facilitate effective segmentation causally improve memory (Boltz, 1992; Flores et al., 2017; Gold, Zacks, & Flores, 2016; Schwan et al., 2000).
Event Segmentation Theory (EST; Zacks, Speer, Swallow, Braver, & Reynolds, 2007) gives an account of how ongoing activity is segmented into meaningful chunks, with implications for episodic memory formation. According to EST, observers form working memory representations of an ongoing activity, called event models, that represent “what is happening now.” These models serve to guide the comprehension of incoming perceptual information. An effective current event model allows one to make predictions about what will happen in the near future. When there is a spike in prediction error, the current event model is updated. This process of memory updating is experienced by the observer as a boundary between events. Of particular relevance to the present study is the notion that prediction plays a central role in establishing memory representations of discrete units of activity.
The role of prediction in the encoding of event representations has also been discussed in the episodic memory literature. For example, Glenberg (1997) has argued that episodic memories allow individuals to predict physical interactions with their three-dimensional worlds. Related to this, Hintzman (2011) has suggested that involuntary recollections of past events cue potential spatio-temporal regularities and that such remindings can facilitate predicting future events. Finally, work on episodic future thought suggests that mental representations underlying episodic memory also enable thinking predictively about future events (Schacter, Addis, Hassabis, Martin, Spreng, & Szpunar, 2012). In this vein, a recent study of the neurophysiology of event segmentation and memory indicated that prior memories guide anticipatory reinstatement of neural activity in regions associated with segmentation at the level of event models (e.g., Baldassano, Chen, Zadbood, Pillow, Hasson, & Norman, 2017). These findings suggest that episodic memory representations could facilitate detecting change from one instance of an event type to the next; however, no previous studies have investigated this.
Detecting and Remembering Episodic Changes
Episodic changes and associated effects on memory performance have been investigated extensively in the episodic memory literature, in the context of interference effects. It is often the case that episodic changes impair memory when different features are associated with a common cue. For example, in our restaurant example above the two meals are different features associated with a common cue, the friend. Looking back, you might have difficulty remembering which meal your friend ate at the restaurant on the second dinner because you could experience proactive interference from the memory of the first dinner. Indeed, the deleterious effects of such competition have been demonstrated across many paradigms using a variety of stimulus materials (for a review, see Anderson & Neeley, 1996). However, there are some situations in which multiple associations can facilitate remembering. In a classic example, Barnes and Underwood (1959) found retroactive facilitation in paired associate learning when cues were presented with associated responses in separate lists (A-B, A-B’). Consistent with this, Robbins and Bray (1974) found retroactive facilitation in an A-B, A-D paradigm in which participants were told about the relationship between pairs in each list (also see, Bruce & Weaver, 1973). Findings such as these have been taken to suggest that episodic changes can lead to enhanced memory when individuals detect and effectively encode those changes.
Following this proposal, Jacoby, Wahlheim and colleagues (e.g., Jacoby, Wahlheim, & Kelley, 2015; Wahlheim & Jacoby, 2013) proposed the memory-for-change framework to explain when episodic changes should be associated with memory enhancement or impairment. A key component of this framework is the recursive reminding hypothesis (Hintzman, 2011), which states that perceiving a current event can trigger involuntary recollection of past events, and the cognitive operation of reminding can be encoded as part of a configural representation that also includes the constituent events of the reminding (also see, Hintzman, Summers, & Block, 1975; Tzeng & Cotton, 1980; Winograd & Soloway, 1985). This representation is configural because memory for a later event becomes embedded within a representation that includes the operation of reminding and the earlier event. This representational structure can facilitate order memory by allowing individuals to remember which event was the object of reminding and which was the subject of reminding. The recursive process of remembering that a more recent event reminded one of an earlier event provides relative order information.
The empirical findings taken as initial support for the recursive reminding hypothesis were those showing effects of repetitions and item associations on spacing judgments (Hintzman & Block, 1973; Hintzman et al., 1975), effects of spaced repetitions on frequency judgments (Hintzman, 2004) and effects of item associations on relative order judgments (Hintzman, 2010). Jacoby, Wahlheim, and colleagues (e.g., Jacoby et al., 2015; Wahlheim & Jacoby, 2013) extended this hypothesis by proposing a dual-process model that integrates controlled and automatic influences of memory with the mechanics proposed by the recursive remindings hypothesis. According to their model, the retrieval processes involved in detecting a changed event brings the memory of the earlier event into the same context as the current event that contains the changed features. In doing so, people can compare recent and remote event features within working memory, which allows them to detect changes. Such episodic change detection allows features from both events to be encoded into a configural representation that also includes memory for the reminding. Later, when people are able to remember earlier-detected changes, their remembering of the relative order of events will be facilitated by their recollecting that the recent event reminded them of the earlier event. In contrast, failing to recollect earlier-detected changes can impair memory for the more recent event when the strength of the initial competing information is increased by the retrieval process that allowed individuals to initially detect the change. Proactive interference from this strengthening occurs when later retrieval of event features is not based on recollection (also see, Swire, Ecker, & Lewandowsky, 2017).
A clear example of the predicted effects of the memory-for-change framework can be seen in a paired-associate learning experiment from Jacoby et al. (2015, Experiment 3). In this experiment, participants studied two lists of word pairs in which some of the pairs contained one feature that remained the same and another that changed from one presentation to the next (e.g., Presentation 1 = knee-bone, Presentation 2 = knee-bend). Some of these pairs changed between presentations that both appeared within the List 2 context, whereas others changed between presentations that appeared in the List 1 and List 2 contexts. Participants were instructed either to look for all the possible changes in the experiment (List 1 and List 2), or to look only for changes within List 2. The critical question was how participants performed on a final test of memory for the List 2 pairs when items changed from List 1 to List 2. Those who had looked for all possible changes performed much better on these pairs—showing proactive facilitation in memory, and were better able to remember that the pairs had changed between lists. These results provided evidence that retrieving episodic memories in the service of detecting change was associated with facilitated memory for changed events as predicted by the memory-for-change account.
The generalizability of the memory-for-change account has received support from several recent studies showing similar memorial consequences associated with episodic changes. These effects have been shown in memory for the relative order of categorially related pairs of individually presented words (Jacoby & Wahlheim, 2013), list discrimination of word pairs (Jacoby, Wahlheim, & Yonelinas, 2013), memory for positions on controversial issues held by fictional political candidates (Putnam, Wahlheim, & Jacoby, 2014), and misinformation about a fictional crime (Putnam, Sungkhasettee, & Roediger, 2017). The latter two extensions of this framework are especially important for the present investigation because they suggest that episodic change effects can generalize to more naturalistic contexts. The most naturalistic extension was the study by Putnam et al. (2017), which showed that memory for details from a slideshow depicting a fictional crime, that were changed in a subsequently presented narrative (see Okado & Stark, 2005), were better remembered when those changes were detected and remembered. The paradigm used by Putnam et al. has high ecological validity, and the study has both theoretical and practical significance. However, for revealing the mechanisms by which change processing affects memory, the fact that it tested retroactive effects of memory is a limitation. In their paradigm, memory enhancement associated with detecting and remembering change could be the result of retrieval practice of the original information during the subsequent presentation of the narrative, rather than the combination of detecting and remembering change. This suggests the need to examine the association between change processing and proactive effects of memory using naturalistic materials.
Adult Age Differences in Event Perception and Memory for Change
One process that may have profound effects on the ability to use memory for predicting new events is healthy aging. Older adults have well-established deficits in episodic memory tasks, especially those that require self-initiated reinstatement of context from prior events (for reviews, see Balota, Dolan, & Duchek, 2000; Zacks, Hasher, & Li, 2000). Research has shown that age-related episodic memory deficits extend to older adults’ memory for naturalistic activities resulting from poorer encoding of event structure. For example, Zacks, Speer, Vettel, and Jacoby (2006) found that older adults segmented ongoing activities less consistently than younger adults, and this predicted older adults’ poorer memory for event details. In addition, Kurby and Zacks (2011) found that older adults’ segmentation was organized less hierarchically than younger adults, older adults had worse memory than younger adults, and age differences in event segmentation sometimes predicted these memory differences. Given that the ability to segment events depends on one’s sensitivity to changes, these findings suggest that one source of older adults’ impaired event perception and memory may be a diminished sensitivity to changes in ongoing activities. This possibility is consistent with the finding that older adults are less sensitive to changes in context (Balota, Duchek, & Paulin, 1989). Consequently, older adults may form less coherent event representations, which could also impair their ability to detect when event features change from one episode to the next.
In addition to this age-related deficit in detecting ongoing changes, older adults are impaired in their detection of changes that occur between separate episodes. Important for the present study, this deficit could have downstream consequences for remembering change and recall of activity features under conditions of proactive interference. A clear example of this can be seen in a paired associate learning experiment from Wahlheim (2014; Experiment 3). In this experiment, participants studied two lists of word pairs and were instructed to indicate which pairs changed from List 1 to List 2. Older adults were given extra study time in List 2, which allowed them to detect as many changes as younger adults. Despite this, older adults still remembered fewer changes, and this partly explained why older adults showed proactive interference in their recall of List 2 responses, but younger adults did not. The finding that older adults were more susceptible to interference is similar to results shown in other interference paradigms (e.g., Hasher & Zacks, 1988; Jacoby, Debner, & Hay, 2001; Healey, Hasher, & Campbell, 2013). However, the results of Wahlheim (2014) showed that in situations involving episodic change, this deficit is associated with differences in the ability to detect and remember changes. When considered in the context of event perception, these findings suggest that older adults’ impaired ability to form coherent episodic representations of naturalistic activities could impair their ability to detect changes with similar downstream consequences for remembering changes and recall of associated activity features.
Event Memory Retrieval and Comparison Theory
Bringing together the role of prediction in event comprehension proposed by EST (Zacks et al., 2007) with the role of change processing in episodic memory proposed by the memory- for-change framework (Jacoby et al., 2015; Wahlheim & Jacoby, 2013) leads to a novel proposal for the mechanism by which memories guide ongoing processing of the relationship between current perceptions and episodic memories for experienced events, and at the same time update episodic memory representations by encoding event changes. We refer to the account of this mechanism as Event Memory Retrieval and Comparison (EMRC) Theory. In this section, we describe EMRC and explain its implications for the effects of healthy aging on memory for change described above.
Figure 1 provides a schematic depiction of the proposed processes involved in the encoding of event changes according to EMRC. The account proposes that as people observe everyday events, they form expectations about the features of upcoming events that will occur in the near future. These expectations (hereafter referred to as predictions) are usually covert, but can also be accessible to conscious awareness in cases where top-down attention is allocated to the relationship between episodic memories and ongoing perception. During ongoing perception, observers encounter activity features that cue recollection of episodic representations of related events. These recollections can occur involuntarily as a consequence of feature overlap between current and earlier events (Berntsen, 1996; Hintzman, 2011) or through self-initiated elaboration of event features as retrieval cues (Jacoby, 1974; Jacoby & Wahlheim, 2013). No matter how it is accomplished, recollecting an earlier event should affect an observer’s predictions about what will happen in the new situation. Retrieval of episodic memories should usually improve prediction, because natural activity often contains consistently recurring feature combinations. In these cases, predictions of repeated activities would be confirmed, leading to the maintenance of stable event models. However, when features change from one event to another related event, this can lead to a prediction error that upregulates attention to unexpected features, leading observers to detect changed features and update their event model.
Figure 1.
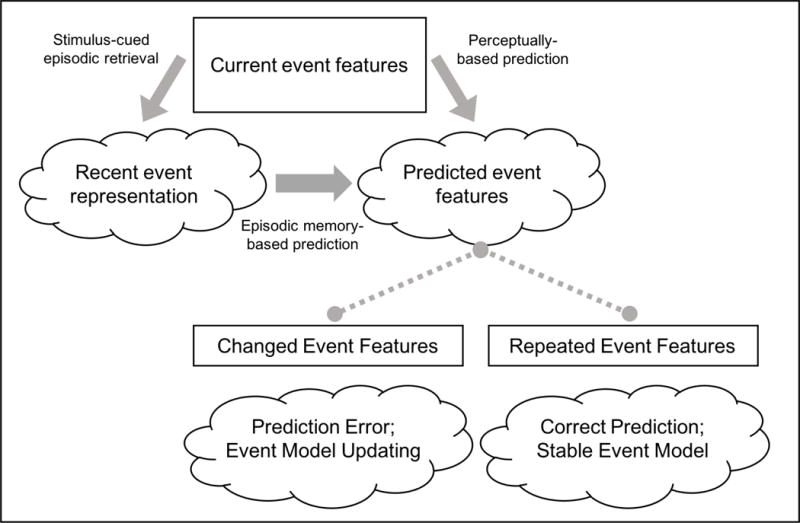
Schematic depiction of the processing chain leading to episodic change detection according to Event Memory Retrieval and Comparison (EMRC) Theory. Boxes represent perceived features, and clouds represent cognitive representations. Arrows indicate the flow of information from perceptual inputs and memory representations to predictions. Dashed lines indicate the influence of predictions on the perception of upcoming event features. EMRC proposes that current event features cue retrieval of recent related event representations. Both those representations and ongoing perceptual information inform predictions about upcoming event features. Changed features in upcoming events lead to prediction errors and event model updating, whereas repeated features tend to lead to maintenance of stable event models.
In the short term, episode-based prediction errors are likely to incur a processing cost and to interfere with ongoing processing. However, these very costs may have long-term benefits for episodic memory updating, if they allow the observer to encode a representation that includes: (a) the original prediction of a repeated event, (b) the fact of the prediction error, and (c) the unexpected features. The formation of configural representations that comprise changed features and the cognitive operations associated with detecting change will allow the observer to subsequently remember what happened during the original event and the new event, and the temporal context associated with each event. This will produce proactive facilitation of the original event for retrieval of the new event when the relationship between the events can be accessed via remembering the earlier-detected change. In contrast, when detected changes cannot be remembered, proactive interference should occur. This would result from the retrieval of the original event that lead to change detection increasing the strength of its competitive features without improving memory for the temporal context associated with those features. However, proactive interference from the original features should not occur when change is detected but the original features cannot be recalled at that time, as the inaccessibility of the earlier features would render them non-competitive with memory for changed features.
An important aspect of EMRC is that the memorial benefits associated with detecting and remembering change depend on the cognitive system registering (consciously or not) the prediction error. However, there will also be instances when prediction errors are not registered and changes are undetected. In these instances, proactive effects of memory will partly depend on the reason why change goes undetected. When change goes undetected because the original features were not encoded effectively, recall of the changed Day 2 features should be similar to recall of once-presented features. In this situation, changed Day 2 features would in effect be perceived as once-presented. In contrast, when change goes undetected because the original features were encoded but inaccessible during Day 2, proactive interference from response competition should only occur when the original features can be recovered at test. In addition, proactive effects of memory associated with undetected changes should partly depend on the extent to which episodes from similar event models compete during retrieval (Radvansky, 2005, 2012). Two related events belonging to distinct event models should not compete, resulting in similar memory for changed activities and once-presented activities. In contrast, two related activities belonging to similar models should create proactive interference to the extent that the models are similar. These hypothetical effects of event segregation are reminiscent of interference reduction due to context differentiation (for reviews, see Abra, 1972; Smith & Vela, 2001). Importantly, proactive facilitation could never be obtained for undetected changes because similar events could at best be represented independently.
In addition to providing a mechanistic account of the processes involved in the comprehension of event changes, EMRC also has implications for understanding age-related differences in the processing of such changes. Older adults experience deficits in attention and episodic memory (for reviews, see Balota et al., 2000; Zacks et al., 2000) that affect their ability to bind event features (e.g., Naveh-Benjamin, 2000) and perceive structure in ongoing activities (Kurby & Zacks, 2011; Zacks et al., 2006). According to EMRC, these deficits should lead to poorer encoding of discrete event representations. The diminished coherence of event representations would reduce the frequency of stimulus-cued retrievals of prior events due to fewer perceived overlapping features with current events and diminished support for self- initiated cue-elaboration. Consequently, older adults should experience fewer episode-based prediction errors than younger adults, leading older adults to detect and remember fewer event changes. When older adults do detect changes, their diminished ability to later remember those changes (Wahlheim, 2014) should lead them to access fewer configural representations, resulting in a greater susceptibility to interference among competing activities features. If this proposal is correct, specific impairments in aspects of this processing chain may be targeted for remediation.
The Present Experiments
The primary purpose of the present investigation was to provide an initial test of some of the aspects of the processing chain proposed by EMRC. To do this, we developed the everyday changes paradigm, which combines procedures from the event perception and episodic memory literatures. In this paradigm, participants watch movies depicting an actor performing everyday activities across the course of two fictive days in her life. The relationship of stimulus features between these episodes are varied such that some activities recur with a critical feature being repeated (e.g., ordering a cheeseburger on both days), other activities have features that change across the days (cheeseburger/salad), and control activities only appear on Day 2 (salad only). In some variations of this paradigm, participants are asked to overtly classify the relationship between activities while watching the Day 2 movie, which provides a measure of the extent to which participants initially detect the relationship between activity features on each day. After watching the movies, participants take a cued-recall test that asks them to: 1) recall Day 2 activity features, 2) indicate whether they remember features of the activity changing between days, and 3) recall Day 1 features for activities that they remember changing.
The general design of the everyday changes paradigm parallels variations of the verbal learning A-B, A-D paradigm, including those used by Jacoby, Wahlheim, and colleagues (Jacoby et al., 2015; Wahlheim & Jacoby, 2013). However, as mentioned above, natural activities differ from verbal paired associates in an important way: Natural activities have meaningful temporal structure. For example, the association between entering a restaurant and ordering a meal is a reliable regularity that is learned over many experiences with restaurant meals. The everyday changes paradigm captures this important feature of natural activities.
We derived our hypotheses from earlier findings showing that detecting and remembering change counteracts interference, presumably via the encoding and retrieval of configural representations comprised of multiple event features and the cognitive operations that associate those features (e.g., Jacoby et al., 2015; Wahlheim & Jacoby, 2013). We hypothesized that cued recall of Day 2 features for changed activities would show proactive facilitation when participants detected changed features while watching the Day 2 movie and remembered that those features changed when taking a later memory test. In contrast, we hypothesized that Day 2 recall would show proactive interference when participants initially detected changed features, but could not remember that those features changed on a later test. Conversely, we expected that detecting but not remembering changed features would lead to more intrusions of Day 1 features, due to the boost in strength from retrieval practice of Day 1 features, whereas detecting and remembering changes would reduce Day 1 intrusions due to the memorial benefits associated with remembering change. Critically, we propose that the initial detection of change necessary for encoding configural representations results from episode-based prediction errors.
We further hypothesized that older adults would be more susceptible to interference, and that this would reflect their impaired processing of changes relative to younger adults. As described above, we derived this hypothesis from the fact that older adults have well-established deficits in attention and memory (for reviews see, Balota et al., 2000; Zacks et al., 2000) that extend to the formation of associations among perceived stimulus features (e.g., Naveh-Benjamin, 2000) and to perception and memory for structure in ongoing events (Kurby & Zacks, 2011; Zacks et al., 2006). These deficits should lead to impaired encoding of event features, which would reduce older adults’ ability to detect episodic changes. This would result from impoverished event representations decreasing the feature overlap necessary for current events to cue recall of recent related events.
If older adults are less likely to retrieve representations of related previous events, then they should be less likely to predict upcoming activity features on the basis of those recent events and more likely to predict activity features based on current perceptual information. However, the extent to which older adults will be able to make effective perceptual predictions will depend on how well they can comprehend ongoing activities. Older adults show more inter-individual variability in their comprehension of naturalistic activities relative to younger adults, and this age difference is associated with older adults’ lower memory for features of those activities (e.g., Zacks et al., 2006). Consequently, older adults should be impaired overall in their ability to engage in predictive processing relative to younger adults. Further, even when older adults do recall activities that lead to memory-based prediction errors, their binding deficit could diminish their ability to encode multiple features together into configural representations, rendering them less likely to represent together the retrieval event, the fact of the prediction error, and the temporal contexts associated with events. This could result in older adults being less likely to remember the source of the features from related events when they correctly remember that an activity changed. We describe specific hypotheses for two experiments in more detail below.
Experiment 1
The primary purpose of Experiment 1 was to examine the effects of episodic changes on memory for features of naturalistic activities in older and younger adults. During a first session, participants watched two movies of an actor performing everyday activities on two fictive days in her life. The activities were either repeated, changed, or new (control) on Day 2. Approximately one week later, participants returned for a second session and took a cued recall test that asked about activity features from Day 2 and also whether each activity had changed from Day 1 to Day 2—a judgment of remembering change. We assumed that this measure tapped into participants’ ability to remember an earlier established relationship between changed activity features. In Experiment 1, we did not measure change detection directly, to preserve the naturalistic viewing experience of watching an actor perform an uninterrupted sequence of everyday activities. Thus, failures of remembering change could reflect failures of initial detection, or forgetting of the experience of the detection. In Experiment 2, we compromised the naturalness of the viewing situation and directly measured change detection during Day 2 viewing. To foreshadow, the results of the two procedures converged closely.
Following EMRC, we hypothesized that instances of remembering change would often indicate that during Day 2 viewing, participants experienced a prediction error that triggered change detection and memory updating. This updating would be accomplished by features from each day being encoded along with their temporal contexts and the cognitive operations that established their relationship. Therefore, we hypothesized that remembering change would be associated with higher rates of correct recall of Day 2 features and lower rates of intrusions of Day 1 features. Regarding age-related differences, we expected older adults to experience fewer memory-based prediction errors to changed Day 2 activities due to their deficits in encoding and remembering Day 1 activities. We also expected these deficits to result in older adults remembering fewer changes. Finding that older adults remember fewer changes and show impaired memory for activity features and their source would be consistent with findings showing that older adults are more susceptible to interference effects partly because they are impaired in their ability to remember changes (Wahlheim, 2014).
Method
The full stimulus sets for the materials used in the present experiments (i.e., movies of the actor performing everyday activities and cued recall questions), anonymized data files, data with coded responses, and analysis scripts can be downloaded from the following URL: https://osf.io/t2qjk/. The research reported here was approved by the Institutional Review Boards of Washington University in St. Louis (Experiments 1 and 2) and The University of North Carolina at Greensboro (Experiment 2). We report how we determined our sample size, all data exclusions, all manipulations, and all measures in this experiment (Simmons, Nelson, & Simonsohn, 2011) below.
Participants
The participants were 36 younger adults (26 females; MAge = 20.19, SD = 1.43, range: 18-23) from Washington University in St. Louis and 36 older adults (27 females; MAge = 77.19, SD = 8.07, range: 61-94) from the St. Louis community. Three older adults were excluded from analyses because they scored lower than 25 on the Mini-Mental State Exam (MMSE; Folstein, Folstein, & McHugh, 1975). The final sample of 33 older adults (25 females; MAge = 77.00, SD = 7.79; range: 61-90) all had MMSE scores of 25 or above (M = 28.61, SD = 1.60, range: 25-30). We based our selection of these sample sizes on an earlier study of proactive effects of memory and associated effects of remembering change in older and younger adults (Wahlheim, 2014). The effect sizes for significant main effects of age on cued recall performance were large. We converted the smallest of those effect sizes (ηp2 = .20) to Cohen’s f = 0.5 and conducted a power analysis using G*Power Version 3.1.9.2. The analysis indicated that at least 17 participants per group were needed to detect comparable effects with power = .80 and α = .05. We chose a larger sample size here because we were unsure how the current materials would behave.
As compensation, younger adults received either $10 per hour or partial course credit, and older adults received $10 per hour. The two sessions of the experiment lasted approximately 3 hours total. Vocabulary scores on the Shipley Institute of Living Scale (Shipley, 1986) were significantly higher for the 33 older adults in the final sample (M = 36.33, SD = 2.70) than for the 30 younger adults who took the vocabulary test (M = 34.67, SD = 2.26), t(61) = 2.64, p = .01, d = 0.67 (six younger adults did not take the vocabulary test due to experimenter error). The average years of education did not differ between older (M = 14.79, SD = 2.64) and younger (M = 14.14, SD = 1.29) adults, t(67) = 1.31, p = .19, d = 0.31. All participants were tested individually.
Design and Materials
A 2 × 3 mixed factorial design was used. Age (younger vs. older) was a between-subjects variable, and Activity Type (repeated, control, or changed) was manipulated within-subjects. The materials were videos of a female actor performing daily activities on two fictional days in her life. The activities took place in or around the actor’s home. There were two versions of each activity that differed on a thematically central feature (see Figure 2). For some of the activities, the criterial feature that differed between versions was an object that the actor interacted with (e.g., the towel she hung in the bathroom; Figure 2, top panels), whereas for other activities, the criterial feature was the action itself (e.g., the type of exercise she performed on a yoga mat; Figure 2, bottom panels).
Figure 2.

Example still frames from changed activities. The left column shows examples of activities from a Day 1 movie, and the right column shows examples of changed versions of those activities that appeared in the Day 2 movie. The top row shows an example of an object change and the bottom row shows an example of an action change.
The material set consisted of 62 total activities (51 critical; 11 fillers). For the critical activities, 45 included object changes and 6 included action changes. For the filler activities, 10 included object changes and 1 included an action change. Filler activities were interspersed among the critical activities to improve the coherence of action sequences. All filler activities repeated across movies. To counterbalance critical activities across conditions, the 51 critical activities were divided into 3 groups of 17. Each activity appeared once in each condition across three experimental formats. To create the activity type conditions, we varied the versions of activities in the Day 1 movie across three formats, leaving out control activities that only appeared in Day 2, and held constant the version of activities in the Day 2 movie.
Each format of the Day 1 movie depicted the actor performing 45 activities (34 critical; 11 fillers). The durations of the Day 1 movies were: 29 min and 15 s (Format 1), 30 min and 14 s (Format 2), and 28 min and 37 s (Format 3). The Day 2 movie depicted the actor performing 62 activities (51 critical; 11 untested fillers); its duration was 41 min and 24 s. The activity types appeared in fixed-random orders such that no more than three subsequent critical activities were from the same condition in either movie. The average serial position of critical activities in each movie was equated across conditions to control for serial position effects.
Sixty-two test cues appeared on the final cued recall test. Each cue probed participants’ memory for the criterial feature of the relevant activity that could have changed (e.g., “What type of towel did the actor hang in the bathroom?”).
Procedure
Participants completed tasks in two sessions separated by approximately one week. The average delay between sessions (M = 7.10 days, SD = 0.63, range: 6 - 12) did not differ between older and younger adults. All movies were presented on a computer monitor at a 1280 × 720 aspect ratio. The instructions, movies, and memory test described below were all presented using E-Prime 2 software (Psychology Software Tools, Pittsburgh, PA, USA).
During Session 1, participants first read an instruction screen informing them that they would be presented with two movies depicting an actor perform everyday activities on two fictive days in her life. Participants were told that their task would be to watch the actor and to pay careful attention to her actions. After discussing the task with the experimenter, participants watched the entire Day 1 movie without interruption. Immediately after the Day 1 movie ended, participants were presented with another instruction screen. The instructions told participants that their next task would be to watch the same actor perform another set of actions that took place during a day later in her week and that they should pay attention to her actions. After reading these instructions, participants watched the Day 2 movie without interruption.
During Session 2, participants first read an instruction screen that described the cued recall test that they were about to perform. They were told that their task would be to recall features from the activities that they watched in Session 1 and to identify which of those activities changed between the Day 1 and Day 2 movies. To illustrate the type of changes that participants should identify, two example activities that did not appear in the experiment were shown prior to the test. The first example showed the actor cleaning the kitchen counter with a washcloth, and the second example showed the actor cleaning the same counter with a paper towel. Test cues for each critical activity (e.g., “What type of towel did the actor hang in the bathroom?”) appeared individually in the same order as they appeared in the Day 2 movie. This order was selected to disambiguate activities with similar features using relative temporal information. For each activity, participants were first prompted to recall the Day 2 feature (e.g., “maroon bath towel”) by typing their response onto the screen. Next, participants indicated if the activity had changed between movies by pressing the “1” key to indicate “yes” and the “2” key to indicate “no.” When participants indicated that an activity had changed, they were prompted to recall the feature from the Day 1 movie (e.g., “white hand towel”) by typing their response. Most participants entered their own responses, but some older adults preferred to have the experimenter enter them. The number of older adults who preferred this was not recorded.
After the cued recall test, all participants completed a computerized version of the Shipley vocabulary test. Older adults completed the MMSE last.
Results and Discussion
The effects of the experimental manipulations were modeled using linear and logistic mixed effects models, fit with the lme4 package in R (Bates, Maechler, Bolker, & Walker, 2015); hypothesis tests were performed using the Anova function of the car package (Fox & Weisberg, 2011), and post hoc comparisons using the Tukey method were conducted using the lsmeans package (Lenth, 2016). The logistic models operate on log-likelihoods; for data presentation we have converted estimates of cell means and confidence intervals back to probabilities. The level for significance was set at α = .05.
Recall Performance
Participants’ responses when attempting to recall Day 2 features were classified into four types. As an illustration of these response types, consider an activity in which the actor hung a white hand towel on Day 1 and a maroon bath towel on Day 2 during the corresponding activity (see Figure 2). Day 2 recall responses were correct descriptions of the criterial activity features in the Day 2 movie (e.g., “she hung a maroon towel”). Day 1 intrusions for changed activities were responses that included the criterial feature from Day 1 (e.g., “she hung a small white towel”) instead of the Day 2 feature. Baseline rates for Day 1 intrusions were estimated for repeated and control activities from instances when responses included features that would have appeared in the Day 1 movie had the activity changed. Ambiguous responses were correct action descriptions that did not distinguish between days (e.g., “she hung a towel”); and Other Error responses included both commission errors that did not include criterial features from either day and omitted responses. Note that commission errors sometimes included criterial features from non-target activities and other times included features that were not associated with an earlier- viewed activity. EMRC does not make any clear predictions regarding Ambiguous and Other Error responses at this point, so we do not report analyses of those responses here. Two raters independently coded responses. Cohen’s κ for the initial ratings (κ = .87, p < .001) showed almost perfect agreement between raters (Landis & Koch, 1977). Discrepancies were resolved through discussion.
To examine how age and watching Day 1 activities affected memory for Day 2 activities, we computed response probabilities for Day 2 recall and Day 1 intrusions for each activity type separately for older and younger adults. We compared these probabilities for repeated and changed activities with control activities using separate logistic mixed effects models, with Age and Activity Type as fixed effects, and subjects and activities as random effects. Modeling the random effect of activities was especially important when examining conditional probabilities in recall due to the potential for item selection artifacts.
The analysis of correct recall of Day 2 activities (Figure 3, top panels) revealed that younger adults recalled significantly more activities than older adults, χ2(1) = 24.01, p < .001. In addition, repeated activities were remembered best and control activities were remembered worst, leading to a significant main effect of Activity Type, χ2(2) = 124.31, p < .001. Post hoc tests showed that, for both age groups, all three activity types differed from each other, smallest z ratio = 2.64, p = .02, with one exception: For older adults, the changed and control activities were not significantly different, z ratio = 0.66, p = .79. Despite this, the Age × Activity Type interaction was not significant, χ2(2) = 1.69, p = .43. These results suggested that older adults’ had a differentially greater memory deficit for changed activities.
Figure 3.
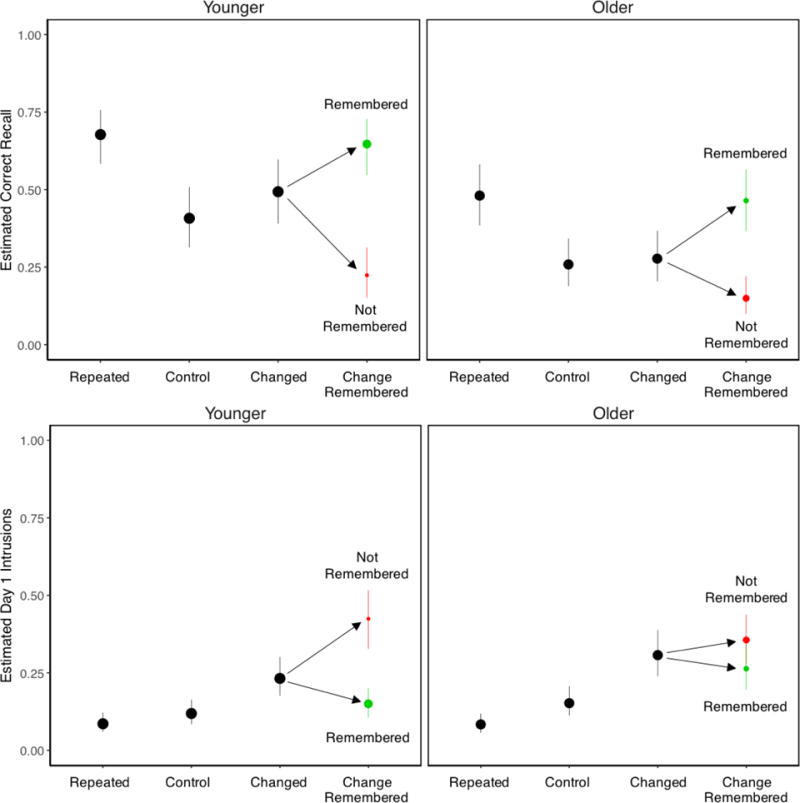
Probabilities of Day 2 correct recall (top panels) and Day 1 intrusions (bottom panels). The areas of conditional points (in green and red) indicate probabilities of remembering change (correct classification of changed activities). Green points indicate that change was remembered (changed activities were correctly classified) and red points indicate that change was not remembered (changed activities were incorrectly classified). The heights of conditional points indicate probabilities of Day 2 recall and Day 1 intrusions. Error bars are bootstrap 95% confidence intervals.
The analyses of Day 1 intrusions (Figure 3, bottom panels) revealed that older adults produced more Day 1 intrusions than younger adults, leading to a significant main effect of Age, χ2(1) = 4.34, p = .037. In addition, both age groups produced Day 1 intrusions most often for changed activities and least often for repeated activities, leading to a significant main effect of Activity Type, χ2(2) = 144.38, p < .001. The Age × Activity Type interaction was not significant, χ2(2) = 3.09, p = .21. Post hoc tests showed that, for younger adults, all three activity types differed significantly from each other, smallest z ratio = 3.76, p < .001, and for older adults, changed activities differed significantly from control and repeated activities, smallest z ratio = 5.23, p < .001, and control activities differed marginally from repeated activities, z ratio = 2.06, p = .098. These results showed that older adults’ memory deficit extended to their incorrect recall of non-target activity features from the Day 1 movie.
Remembering Change
We assumed that correct classifications of changed activities on the cued recall test indicated instances for which change was remembered. It is important to note that remembering change subsequent to viewing both Days, as measured here, is different than detecting change during viewing of the changed clip. Here, we assumed that differences in remembering change resulting from differences reflect earlier differences in detecting change that we did not directly measure in Experiment 1. (In Experiment 2, we measured both concurrent change detection and subsequent change remembering.)
Table 1 displays the probabilities of participants’ classification of activities as having features that changed between the Day 1 and Day 2 movies. The probability of remembering change, as indicated by correct classification of changed activities, was greater than the probabilities of incorrectly classifying repeated and control activities as changed, leading to a significant main effect of Activity Type, χ2(2) = 462.94, p < .001. Younger adults classified more activities as changed than older adults, leading to a significant main effect of Age, χ2(1) = 10.91, p < .001. A significant Age × Activity Type interaction indicated that this difference was greatest for changed activities, χ2(2) = 21.60, p < .001. These results showed that younger adults were better able to correctly classify changed activities than older adults, indicating that younger adults remembered more activity changes.
Table 1.
Probabilities of Classifying Activities as Changed on the Cued Recall Test as a Function of Age and Activity Type: Experiment 1
| Age | Activity Type
|
||
|---|---|---|---|
| Repeated | Control | Changed | |
| Younger | .12 [.08, .15] | .27 [.21, .33] | .67 [.61, .73] |
| Older | .12 [.09, .16] | .19 [.14, .24] | .45 [.37, .52] |
Note: Probabilities for Changed activities are correct classifications that index remembering change, whereas probabilities for Repeated and Control activities are incorrect classifications that are akin to false alarms. Bootstrap 95% confidence intervals are displayed in brackets.
Day 1 Recall When Change Was Remembered
When participants classified activities as changed on the cued recall test, they were also prompted to recall the original Day 1 activity features. This measure was included to enable inferences about the extent to which participants earlier encoded changed features from both movies together during Day 2 viewing as part of a configural representation. These responses were coded in the same manner as for Day 2 recall. Two independent raters showed almost perfect agreement in their ratings (κ = .85, p < .001). Discrepancies were resolved through discussion. Consistent with our predictions, correct recall of Day 1 activity features for correctly classified changed activities was significantly lower for older (M = .55, CI = [.44, .66]) than younger (M = .73, CI = [.65, .81]) adults, χ2(1) = 7.68, p < .001. These results suggested that older adults had earlier encoded Day 1 and Day 2 activity features together less effectively than younger adults. However, these results might also suggest that older and younger adults used different criteria for classifying an activity as changed on some occasions, with older adults potentially being more liberal in their use of that classification.
Recall of Changed Activities Conditionalized on Remembering Change
As described in the Introduction, we expected that correct classification of changed activities (i.e., remembering change) would be associated with enhanced memory for Day 2 activity features. This prediction is based on the assumption of EMRC that remembering change should allow participants to access activity features from both days and the temporal relationship between the activities. We tested this by analyzing correct recall of Day 2 activity features conditionalized on whether changed activities were correctly classified (red and green points in Figure 3). Separate logistic mixed effects models were fit to correct Day 2 recall and Day 1 intrusions, with Age and Remembering Change (correct vs. incorrect classifications of changed activities) as fixed effects, and subjects and activities as random effects. Redundant main effects of Age are not reported. We examined associations between remembering change and memory for activity features separately for Day 2 recall and Day 1 intrusions by conditionalizing on whether changed activities were correctly classified.
EMRC predicts that the memorial benefits associated with correct classifications of changed activities (i.e., remembering change) should manifest in better recall of Day 2 features and fewer intrusions of Day 1 features. The analysis of Day 2 recall revealed higher recall of changed activity features when those activities were classified as changed as compared to when they were not, leading to a significant main effect of Remembering Change, χ2(1) = 111.11, p < .001. There was no significant Age × Remembering Change interaction, χ2(1) = 0.65, p = .42. Post hoc tests comparing Day 2 recall for changed activities conditionalized on whether they were classified as changed with Day 2 recall for control activities were conducted next to examine proactive effects of memory. Day 2 recall was significantly higher for correctly classified changed activities than control activities, smallest z ratio = 4.33, p < .001, whereas recall was significantly lower for incorrectly classified changed activities than control activities, smallest z ratio = 6.34, p < .001. These results replicate earlier findings showing proactive facilitation when change was remembered and proactive interference when change was not remembered (e.g., Wahlheim & Jacoby, 2013).
EMRC also predicts that the memorial differences in Day 2 recall associated with remembering change depend on whether Day 1 features are correctly recalled at test. Correct recall of Day 1 features should lead to proactive facilitation of Day 2 recall by enhancing cue elaboration at test. In contrast, failing to correctly recall Day 1 features should lead to proactive interference either because those features are recalled without source information or because their unconscious evocation exerts interference, perhaps via blocking. The magnitude of such interference should be similar to when change is not remembered. However, this will depend on how often changes are initially detected during encoding and how often change classifications on the cued recall test reflect guessing. Proactive interference effects should be greater when more changes are detected during encoding but not remembered at test due to more competing responses being strengthened by study phase retrievals during change detection. These effects should also be greater when more classifications at test are based on guessing because those instances should reflect retrieval of competing responses without sufficient diagnostic information about their original source.
To examine the memorial effects associated with remembering change and recall of Day 1 features, Day 2 recall was conditionalized on three forms of remembering change (Table 2, top rows): correct classification of changed activities (change remembered) and correct recall of Day 1 features, correct classification of changed activities (changed remembered) without recall of Day 1 features, and incorrect classification of change activities (change not remembered). These data were fit with a logistic mixed effects model that included Age and Remembering Change (correctly classified and Day 1 recalled, correctly classified and Day 1 not recalled, and incorrectly classified) as fixed effects, and subjects and activities as random effects. When changed activities were correctly classified, Day 2 recall was higher when Day 1 features were recalled than when Day 1 features were not recalled, leading to a significant main effect of Remembering Change, χ2(2) = 196.33, p < .001. There was no significant Age × Remembering Change interaction, χ2(2) = 4.25, p = .12. Post hoc tests revealed no significant differences in Day 2 recall when changed activities were correctly classified without recall of Day 1 features and when changed activities were incorrectly classified, largest z ratio = 1.78, p = .18. These results show that enhanced Day 2 recall associated with remembering change depended on correct recall of Day 1 features. This suggests that the benefits associated with remembering change depend on participants being able to access multiple elements of configural representations formed during Day 2 viewing.
Table 2.
Probabilities of Day 2 Recall for Changed Activities conditionalized on Activity Type Classification during the Cued Recall Test and on Day 1 Recall as a Function of Age: Experiments 1 and 2
| Experiment | Age | Correctly Classified
|
Incorrectly Classified | |
|---|---|---|---|---|
| Day 1 Recalled | Day 1 Not Recalled | |||
| Experiment 1 | Younger | .84 [.77, .91] | .20 [.10, .29] | .20 [.12, .28] |
| Older | .71 [.59, .83] | .22 [.12, .32] | .14 [.08, .20] | |
|
| ||||
| Experiment 2 | Younger | .86 [.80, .93] | .33 [.19, .47] | .40 [.29, .51] |
| Older | .64 [.51, .77] | .30 [.17, .43] | .26 [.17, .34] | |
Note: We assume that correct classifications of changed activities (Correctly Classified) reflect remembering that activity features changed from Day 1 to Day 2, and incorrect classifications of changed activities (Incorrectly Classified) reflect a lack of remembering that activity features changed. Bootstrap 95% confidence intervals are displayed in brackets.
The analysis of Day 1 intrusions (Figure 3) revealed fewer intrusions when changed activities were correctly classified as compared to when they were incorrectly classified, leading to a significant main effect of Remembering Change, χ2(1) = 36.86, p < .001. The reduction in Day 1 intrusions associated with remembering change was greater for younger than older adults, resulting in a significant Age × Remembering Change interaction, χ2(1) = 11.73, p < .001. The smaller benefits obtained for older adults likely reflected a combination of their less effective formation of configural representations during Day 2 viewing and greater tendency to classify activities as changed based on non-diagnostic information.
Summary
The Day 2 recall results showed that repeated viewing of the same event increased performance relative to watching an event once, and that changing a central event feature was associated with proactive facilitation when the change was remembered and proactive interference when the change was not remembered. In contrast, remembering change was associated with lower Day 1 intrusion rates. The proactive facilitation in Day 2 recall reflected instances when both change was remembered and Day 1 features were recalled. EMRC predicts this, because Day 2 recall should be facilitated by the elaborative encoding that occurs when configural representations are formed as a result of encoding detected changes. In addition, later recall of Day 2 features should also benefit from the additional cue elaboration afforded by remembering change and recall of Day 1 features. According to EMRC, a crucial stage leading to a configural representation being available during final recall is experiencing a prediction error due to the changed feature on Day 2 being different from the Day 1 feature that is expected on the basis of episodic retrieval. However, this experiment did not include a direct measure of change detection, only remembering change. This precluded an examination of the role of change detection in forming configural representations. Consequently, we conducted Experiment 2 to obtain a direct measure of change detection and to attempt a replication of the findings in Experiment 1.
Experiment 2
Experiment 1 showed proactive facilitation in memory for changed activity features for both age groups when change was remembered and the earlier features could be recalled. This finding is consistent with earlier studies (e.g., Jacoby et al., 2015; Wahlheim, 2014) as well as our a priori predictions. However, in order to preserve uninterrupted viewing of the Day 2 movie in Experiment 1, we did not distinguish the effects of detecting a change from the effects of later remembering that a change had occurred. In Experiment 2, we did so by adding a direct measure of change detection after the presentation of each individual Day 2 activity. We also included an approximately one-week delay between viewing of the Day 1 and Day 2 movies, because pilot testing showed that participants detected nearly every change when the two movies were shown consecutively in the same session. Thus, there were three sessions, each separated by approximately one week: Day 1 viewing, Day 2 viewing with a change detection task, and cued recall with a remembering change task.
The measure of remembering change that we included on the final cued recall test was modified slightly to more precisely examine participants’ ability to classify activity types. Rather than just asking whether the feature in question had changed from Day 1 to Day 2, we asked participants to classify each item as repeated, changed, or new. We included this three-alternative judgment for both Session 2 (change detection judgments) and Session 3 (change remembering judgments). Finally, we examined participants’ meta-awareness of their classification accuracy by asking them to make confidence judgments for those classifications. Older adults are typically impaired in their metacognitive monitoring of recollective information (for a review, see Dodson, 2017), and the inclusion of confidence judgments allowed us to assess whether this deficit extends to activity type classifications, which are likely to often be recollective experiences.
Consistent with predictions of EMRC, we expected that the retrieval processes that enable change detection during Day 2 viewing would allow participants to encode Day 1 and 2 features within configural representations. Consequently, later accessing those representations via remembering change should be associated with enhanced Day 2 recall. However, the retrieval of Day 1 features during Day 2 viewing that enables change detection should increase the accessibility of Day 1 features. This would make Day 1 features more competitive on a later test when recollective processes cannot be deployed to oppose the increased strength of those features. This leads to the prediction that Day 2 recall will be enhanced when change is detected and remembered, but impaired when change is detected but not remembered. Conversely, this hypothetical improvement in memory for temporal relationships should result in Day 1 intrusion rates being lower when change is detected and remembered, and higher when change is detected but not remembered. This trade-off should determine overall levels of recall performance, and also determine the degree of older adults’ differential deficit for changed activity features.
Based on the fact that older adults have impaired encoding of event structure (Kurby & Zacks, 2011; Zacks et al., 2006), we expected that older adults would detect relationships between activities during Day 2 viewing less accurately than younger adults. This should result in older adults detecting and remembering fewer changes and recalling Day 1 features less often when correctly classifying changed activities. These age-related deficits should then have detrimental consequences for Day 2 recall, which would partly contribute to a differential deficit in recall of changed activity features. Finally, this constellation of age-related deficits associated with poorer memory accuracy might also have consequences for metacognitive awareness of the accuracy of activity classifications. The classification of the relationships between activities on separate episodes should mostly rely on recollection-based retrieval, as these classifications will be informed by memory for event features and their origins. In addition, older adults often show poorer metacognitive accuracy when metacognitive judgments are primarily based on recollective processes (for a review, see Dodson, 2017). This leads to the prediction that older adults should show poorer metacognitive accuracy in their confidence judgments to the extent that accurate activity type classifications require recollection of earlier event features.
Method
Participants
We initially planned to collect data from at least 36 participants per age group to match the sample sizes used in Experiment 1. We scheduled appointments for more than 36 younger adults anticipating that some would drop out before completing all three sessions. The entire sample of younger adults included 44 participants from Washington University in St. Louis. However, data from six younger adults were excluded from analyses because five participants did not complete all three sessions, and the computer malfunctioned for one participant. The final younger adult sample included 38 participants (23 females; MAge = 19.84, SD = 1.26, range: 18-23). We recruited a total of 37 older adult participants from the Greensboro, NC community. Data from one older adult were excluded because the computer malfunctioned. The final older adult sample included 36 participants (20 females; MAge = 70.22, SD = 2.88, range: 65-75). All the older adults had MMSE scores of 25 or above (M = 28.44, SD = 1.25, range: 25-30).
As compensation, younger adults received partial course credit and older adults received $10 per hour. The experiment lasted approximately 3 hours total. Vocabulary scores were significantly higher for older (M = 34.97, SD = 3.44) than younger (M = 33.50, SD = 2.24) adults, t(72) = 2.19, p = .03, d = 0.51. In addition, the average years of education were significantly higher for older (M = 15.97, SD = 2.10) than younger (M = 13.50, SD = 1.23) adults, t(72) = 6.22, p < .001, d = 1.43. All participants were tested individually.
Design and Materials
The design and materials were identical to Experiment 1, except that the Day 2 movies appeared as individual clips so that participants could classify the type of activity after each. The duration of each activity varied widely (M = 38.23 s, SD = 25.86 s, range: 4-109 s).
Procedure
The procedure maintained most of the key elements of Experiment 1, but Experiment 2 added an overt measure of change detection during Day 2 viewing and an additional session. The delays between each of the three sessions were approximately one week (M = 7.16 days, SD = 1.17, range: 6-17) and did not differ between age groups.
During Session 1, participants read instructions describing the Day 1 movie that they would be watching. Participants were instructed to pay attention to the actions performed by the actor. Prior to watching the Day 1 movie, participants watched an example activity of the sort the actor would perform. In this activity, the actor cleaned the kitchen counter with a washcloth. Participants then watched the Day 1 movie uninterrupted.
During Session 2, participants read instructions describing the relationship between the activities they watched in the Day 1 movie and the activities that they would be watching in the Day 2 movie. They were also instructed that their task would be to indicate how Day 2 activities related to Day 1 activities (repeated, changed, or new). To clarify the activities that should be classified as changed, participants were shown the example activity from Session 1 followed by another version of that activity showing the actor cleaning the kitchen counter with a paper towel (instead of a washcloth). The Day 2 activities then appeared as individual clips. After each clip, participants classified the type of activity. Participants classified activities as repeated by pressing the “1” key, as changed by pressing the “2” key, and as new by pressing the “3” key. Participants also rated their confidence in their classifications on a scale from 1 (not confident) - 5 (very confident) by pressing the corresponding key. They were encouraged to use the full range of the scale. Finally, after classifying an activity as changed, participants were prompted to recall the original Day 1 feature by typing their responses on to the screen.
During Session 3, participants completed a cued recall test of Day 2 activity features similar to Experiment 1. However, as in Session 2: activity type classifications included three alternatives, participants made confidence judgments for those classifications, and participants recalled Day 1 features after each activity that they classified as changed. After the cued recall test, all participants completed a computerized version of the Shipley vocabulary test. Older adults completed the MMSE last.
Results and Discussion
Recall Performance
Recall responses were coded in the same manner as Experiment 1. Two raters initially showed substantial agreement in their classifications, κ = .67, p < .001. Disagreements were resolved by discussion. Day 2 recall and Day 1 intrusions were compared for each activity type for both age groups (Figure 4) using the same logistic mixed effects models as in Experiment 1.
Figure 4.
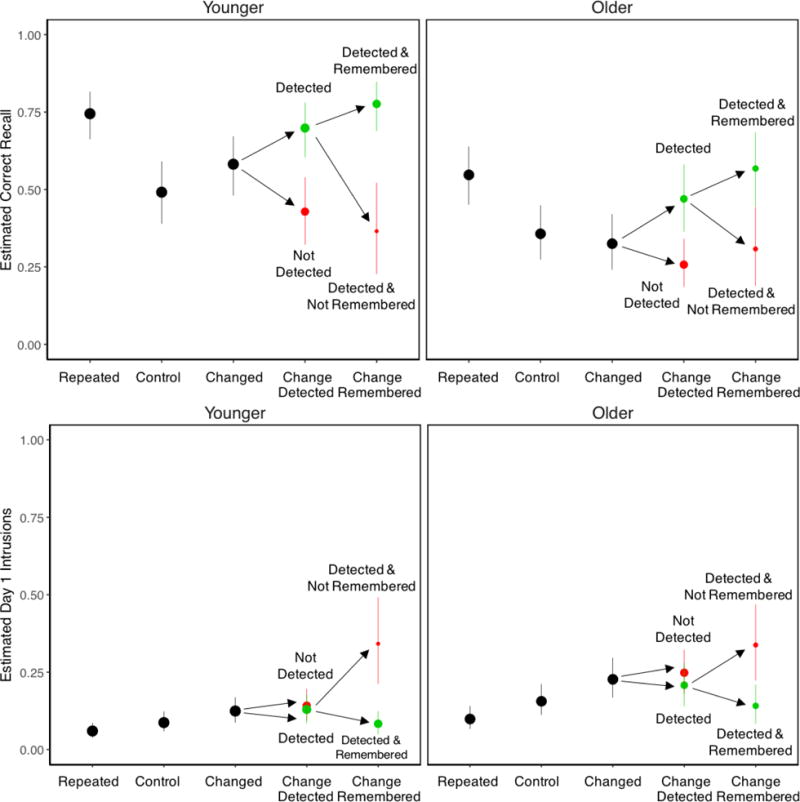
Probabilities of Day 2 correct recall (top panels) and Day 1 intrusions (bottom panels). The areas of conditional points (in green and red) for changed activities refer to probabilities of change detection (correct classification of changed activities during Day 2 viewing) and remembering change (correct classification of changed activities during the cued recall test). Green points correspond to detected or remembered changes (correctly classified changed activities), whereas red points correspond to changes that were not detected or remembered (incorrectly classified changed activities). The heights of conditional points indicate probabilities of Day 2 recall and Day 1 intrusions. Error bars are bootstrap 95% confidence intervals.
Day 2 recall performance was higher for younger than older adults, leading to a significant main effect of Age, χ2(1) = 42.05, p < .001. Recall was highest for repeated activities for both age groups, whereas recall of control activities was the lowest for younger but not older adults. This led to a significant main effect of Activity Type, χ2(2) = 115.45, p < .001, that was qualified by a significant Age × Activity Type interaction, χ2(2) = 7.70, p = .02. Post hoc tests indicated that recall differed across all three activity types, smallest z ratio = 2.86, p = .01, with one exception: For older adults, recall was not significantly different between changed and control activities, z ratio = 1.06, p = .54. These results show that older adults experienced a differential deficit in memory for changed activity features, replicating results from Experiment 1. However, the statistical evidence is stronger in this case due to the significant interaction.
As in Experiment 1, Day 1 intrusions for changed activities reflected activity features that appeared during Day 1 being incorrectly recalled as appearing on Day 2. In addition, Day 1 intrusions for repeated and control activities were baseline estimates of the extent to which features that could have appeared on Day 1 were recalled. Older adults produced more Day 1 intrusions than younger adults, leading to a significant main effect of Age, χ2(1) = 24.50, p < .001. These intrusions were produced most often for changed activities and least often for repeated activities, leading to a significant main effect of Activity Type, χ2(2) = 58.97, p < .001. The Age × Activity Type interaction was not significant, χ2(2) = 0.61, p = .74. Post hoc tests showed significantly more intrusions for changed than for repeated activities, smallest z ratio = 3.83, p < .001, whereas the difference between all other pairwise comparisons was marginally significant, smallest z ratio = 2.06, p = .099. The failure to replicate the significantly higher intrusion rate for changed than control activities in Experiment 1 was likely due to the longer interval between movies in Experiment 2 creating more differentiation between sources, which would reduce the interference caused by Day 1 activity features.
Activity Type Classifications (Detecting and Remembering Change)
Four older adults were excluded from the following analyses because inspection of their data suggested that they did not understand the activity type classification instructions. One participant appeared to have mis-mapped the response keys, confusing changed and control activities, whereas the other three participants never used the “new” classification. Table 3 displays the probabilities of correct classifications for activity types during the Detection Phase in Session 2 (top rows) and the Cued Recall Test in Session 3 (bottom rows). Note that these data are displayed differently from the comparable analyses in Experiment 1 because including all three response options in Experiment 2 allowed us to assess classification accuracy for each activity type. The data from each session were fit with separate logistic mixed effects models with Age and Activity Type as fixed effects, and subjects and activities as random effects.
Table 3.
Probabilities of Correct Activity Type Classification as a function of Phase, Age, and Activity Type: Experiment 2
| Phase | Age | Activity Type
|
||
|---|---|---|---|---|
| Repeated | Control | Changed | ||
| Detection Phase (Session 2) | Younger | .82 [.79, .86] | .86 [.83, .89] | .58 [.52, .63] |
| Older | .74 [.69, .79] | .63 [.58, .69] | .43 [.38, .49] | |
|
| ||||
| Cued Recall Phase (Session 3) | Younger | .81 [.76, .85] | .59 [.53, .65] | .54 [.48, .60] |
| Older | .67 [.61, .73] | .35 [.29, .42] | .40 [.34, .47] | |
Note: Bootstrap 95% confidence intervals are displayed in brackets.
The analyses of classification accuracy during Session 2 showed that younger adults classified activity types more accurately than older adults, leading to a significant main effect of Age, χ2(1) = 51.62, p < .001. There was also a significant effect of Activity Type, χ2(2) = 225.86, p < .001, and a significant Age × Activity Type interaction, χ2(2) = 18.35, p < .001. Post hoc tests showed that for younger adults, correct classification of repeated and control activities was not significantly different, z ratio = 1.82, p = .16, whereas correct classification was significantly higher for repeated and control than changed activities, smallest z ratio = 9.36, p < .001. For older adults, correct classification was significantly higher for repeated than control and for control than changed activities, smallest z ratio = 3.71, p < .001.
The analyses of classification accuracy during Session 3 also showed that younger adults classified activity types more accurately than older adults, leading to a significant effect of Age, χ2(1) = 43.80, p < .001. There was also a significant effect of Activity Type, χ2(2) = 206.19, p < .001, showing that classification accuracy was significantly higher for repeated than control and changed activities, smallest z ratio = 12.98, p < .001, and not significantly different between control and change activities, z ratio = 0.23, p = .97. The Age × Activity Type interaction was marginally significant, χ2(2) = 5.46, p = .06.
Together, the analyses from Sessions 2 and 3 showed that older adults were less accurate at detecting and remembering changed activities relative to younger adults. These results also showed that older adults were generally impaired in their ability to classify activity types. EMRC does not make any predictions about whether (or how) these age-related deficits should differ across activity types, so we do not have a clear interpretation for the interactions obtained above.
Day 1 Recall for Activities Remembered as Changed (Session 3)
When participants indicated that an activity changed between days on the recall test, they were also prompted to recall the original Day 1 features. Similar to Experiment 1, this measure was used to make inferences about the extent to which participants encoded changed features from both movies as part of configural representations. Responses were coded in the same manner as Experiment 1, and raters showed almost perfect agreement in their initial ratings (κ = .81, p < .001). Discrepancies were resolved through discussion. Replicating Experiment 1, older adults’ episodic memory deficit resulted in their recalling fewer Day 1 features after classifying activities as changed on the cued recall test. Correct Day 1 recall for changed activities classified as such was significantly lower for older (M = .51, [CI = .41, .62]) than younger (M = .75, [CI = .68, .83]) adults, χ2(1) = 18.38, p < .001. These results provide further evidence that the quality of configural representations was poorer for older than younger adults, and also suggested that both age groups differed somewhat in the bases for their classifications, with older adults presumably relying on non-diagnostic information more often than younger adults.
Recall of Changed Activities Conditionalized on Detecting and Remembering Change
As described above, EMRC predicts that correct classification of changed activity features on both Session 2 (detecting change) and Session 3 (remembering change) will be associated with better recall of Day 2 activity features and lower rates of Day 1 intrusions. This was based on two assumptions: First, detecting change should enable participants to elaborate their encoding by integrating activity features from both movies together as part of configural representations. Second, remembering change should elaborate retrieval cues to improve access to the contents of those representations. Thus, successful detection and remembering of change should result in proactive facilitation in Day 2 recall and fewer intrusions of Day 1 features. When change cannot not be remembered, earlier detection of change enabled by retrieving Day 1 features during Day 2 viewing should result in more competition from Day 1 features on the cued recall test because the source of those features will less often be recollected. This should result in proactive interference effects that reduce Day 2 recall accuracy and increase rates of intrusions from Day 1 features. To test this prediction, we conditionalized recall performance on whether changed activities were correctly classified in Sessions 2 and 3 (red and green points in Figure 4). We fitted logistic mixed effects models to Day 2 recalls and Day 1 intrusions with Detecting Change, Remembering Change, and Age as fixed effects, and subjects and activities as random effects. We do not report main effects of Age that are redundant with earlier analyses.
The analyses of Day 2 recall (Figure 4, top panels) revealed significant main effects of Detecting Change, χ2(1) = 10.61, p = .001, and Remembering Change, χ2(1) = 25.02, p < .001, as well as a significant Detecting Change × Remembering Change interaction, χ2(1) = 10.65, p = .001. The interaction showed that recall performance associated with correct classification of changed activities during Day 2 viewing (change detection) depended on whether participants correctly classified changed activities during the cued recall test (remembering change). Specifically, Day 2 recall was higher when changed activities were correctly classified during Day 2 viewing and during the cued recall test relative to when changed activities were only correctly classified during Day 2 viewing, z ratio = 6.26, p < .001. In contrast, when changed activities were incorrectly classified during Day 2 viewing, Day 2 recall did not differ depending on the accuracy of activity type classification on the cued recall test, z ratio = 1.01, p = .31. Consequently, Day 2 recall for incorrectly classified changed activities is not further conditionalized on classification accuracy on the cued recall test in Figure 4 for ease of visualization. These effects were comparable for both age groups, as the Age variable did not enter into any significant interactions, largest χ2(1) = 1.32, p = .25.
Post hoc tests comparing Day 2 recall of control activities with changed activities that were correctly classified during Day 2 viewing conditionalized on classification accuracy during the cued recall test were conducted next to examine potential differences in proactive effects of memory. For both age groups, Day 2 recall was significantly higher for changed activities correctly classified at test than for control activities, smallest z ratio = 3.36, p = .002, whereas recall was significantly lower for changed activities incorrectly classified at test than for control activities for older adults, z ratio = 2.59, p = .026, but not younger adults, z ratio = 1.76, p = .183. These results were consistent with earlier findings showing proactive facilitation associated with remembering change for both age groups and proactive interference associated with remembering change for older adults. The finding that younger adults did not show proactive interference when they incorrectly classified changed activities during the cued recall test may have resulted from too few observations to detect differences in cued recall performance. However, the increased temporal separation between movies in this experiment relative to Experiment 1 (i.e., approximately one week vs. a few minutes) could have reduced the competition between event representations here relative to Experiment 1. We return to this issue in the General Discussion.
Similar to Experiment 1, we also examined whether the associations among detecting change, remembering change, and Day 2 recall depended on Day 1 features being correctly recalled when change was remembered. As described earlier, the idea is that correct recall of Day 2 features should be heightened because retrieval of both the fact of the change and the Day 1 features along with their temporal context should effectively cue retrieval of Day 2 features. To examine this, we analyzed Day 2 recall of changed activities that were correctly classified during Day 2 viewing (changes detected) conditionalized on classification accuracy during the cued recall test and Day 1 recall (Table 2, bottom rows). These data were fit with a mixed effects model that included Age and Remembering Change (correctly classified and Day 1 recalled, correctly classified and Day 1 not recalled, incorrectly classified) as fixed effects, and subjects and activities as random effects.
This analysis revealed a significant effect of Remembering Change, χ2(2) = 113.35, p < .001, and a significant Age × Remembering Change interaction, χ2(2) = 6.30, p = .04. These results showed that the memory enhancement in Day 2 recall associated with remembering change was only obtained when Day 1 features were correctly recalled. This advantage was greater for younger than older adults. This age difference could reflect younger adults forming higher quality configural representations when detecting change, due to older adults’ well established deficits in encoding and associative binding. For both age groups, correct Day 2 recall did not differ when changed activities were correctly classified during cued recall and Day 1 features were not recalled compared to when changed activities were incorrectly classified during cued recall, largest z ratio = 1.09, p = .52. Consistent with Experiment 1, these results suggest that the memorial benefits associated with remembering change depended on Day 1 features being correctly recalled. As compared with Experiment 1, these results provide more direct evidence that proactive facilitation in memory for changed activity features is associated with correctly detecting and remembering that those activities changed.
The analyses of Day 1 intrusions (Figure 4, bottom panels) revealed no significant effect of Detecting Change, χ2(1) = 1.97, p = .16, a significant effect of Remembering Change, χ2(1) = 23.68, p < .001, and a significant Detecting Change × Remembering Change interaction, χ2(1) = 9.74, p = .002. The interaction showed that the rate of Day 1 intrusions on changed activities that were correctly classified during Day 2 viewing also depended on whether changes were also correctly classified during the cued recall test. When changed activities were correctly classified during Day 2 viewing, the rate of Day 1 intrusions was significantly lower for those activities than when they were correctly rather than incorrectly classified during the cued recall test, z ratio = 4.68, p < .001. In contrast, when changed activities were incorrectly classified during Day 2 viewing, the rate of Day 1 intrusions did not differ depending on whether those activities were correctly classified during the cued recall test, z ratio = 0.83, p = .41. Consequently, rates of Day 1 intrusions when changed activities were incorrectly classified during Day 2 viewing are not conditionalized on classification accuracy during the cued recall test in Figure 4 for ease of visualization. These effects were comparable for both age groups, as the Age variable did not enter into any significant interactions, largest χ2(1) = 1.98, p = .15. These results show that remembering change facilitated memory for the source of Day 1 features.
Confidence Judgments for Activity Type Classifications
Figure 5 displays the mean confidence judgments for activity type classifications in Sessions 2 and 3. The primary interest was whether participants’ confidence judgments discriminated between correct and incorrect classifications of activity types (i.e., monitoring resolution). Each participant’s confidence judgments were first z-scored because participants could use the confidence scales differently. Then, confidence z-scores were submitted to multilevel linear models with Age, Activity Type, and Classification Accuracy as fixed effects, and activity as a random intercept, and a random slope of accuracy by subject (to model the fact that monitoring resolution might vary by subject). In this analysis, better monitoring resolution is reflected in a larger effect of classification accuracy on confidence. That is, to the extent that one is aware of one’s classification accuracy, confidence should be higher for correct than incorrect classifications. Thus, one critical test is the main effect of accuracy on z-scored confidence. The other critical test is the interaction of accuracy with age, which indicates whether monitoring resolution differed between older and younger adults. These two tests are reported for classifications made in Sessions 2 and 3.
Figure 5.
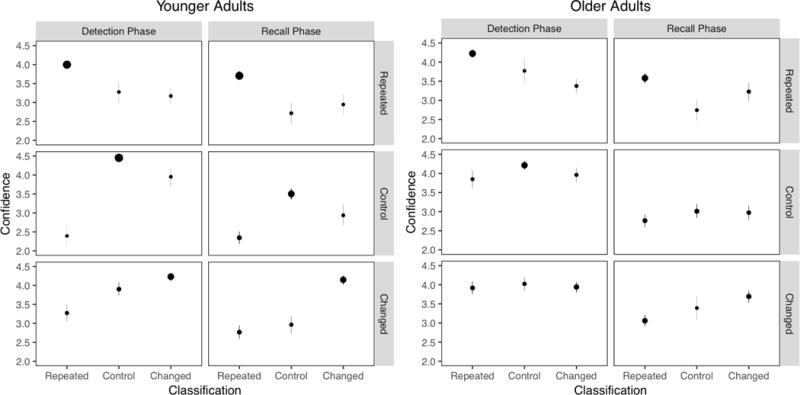
Mean confidence judgments as a function of age, activity type, and activity classification during Session 2 (detection phase) and Session 3 (cued recall phase) of Experiment 2. The point areas indicate the proportion of activities that were classified as each activity type. Error bars are bootstrap 95% confidence intervals.
The mean confidence ratings shown in Figure 5 indicate that younger adults had good monitoring resolution, which was better than that of older adults. For classifications given during Session 2, this led to a large effect of accuracy on z-scored confidence, χ2(1) = 137.1, p < .001, and a significant Age × Accuracy interaction, χ2(1) = 12.9, p < .001. Both patterns held true for judgments made during Session 3: a significant main effect of Accuracy, χ2(1) = 164.2, p < .001, and a significant Age × Accuracy interaction, χ2(1) = 25.0, p < .001. These results are consistent with findings showing age-related deficits in monitoring accuracy in tasks involving recollective processes (for a review, see Dodson, 2017).
Summary
Experiment 2 extended the results of Experiment 1 by showing proactive facilitation in recall of Day 2 features when change was detected and remembered, and proactive interference when change was detected but not remembered. Conversely, Day 1 intrusion rates were higher when change was detected but not remembered relative to when change was detected and remembered. These results confirm the prediction of EMRC that the effects of detecting change should depend on whether change is later remembered, as remembering change enables access to representations that include diagnostic information regarding the temporal relationship between changed activity features. Experiment 2 also replicated the finding in Experiment 1 that older adults’ experienced a differential deficit in impaired recall of changed activities. According to EMRC, this deficit was partly due to older adults detecting and remembering fewer changes. Finally, both age groups could monitor the accuracy of their activity type classifications reasonably well, but older adults showed impaired resolution. These results are consistent with a larger literature showing age-related deficits in monitoring of recollective information.
General Discussion
The present experiments examined adult age differences in event memory when features from everyday activities changed from one episode to the next. A major focus of the present experiments was establishing the extent to which memory for activity features was associated with detecting and remembering episodic changes. The present findings suggest that episodic memory guided the detection of episodic changes in naturalistic actions and that remembering such changes was associated with proactive facilitation of event memory. Older adults showed a differential deficit in memory for changed activities that was associated with their impaired ability to detect and remember event changes. Older adults also showed poorer metacognitive monitoring when classifying the relationship between activities, both when viewing the more recent activities and when recalling the activities on a later test. This deficit was likely the result of activity type classifications being primarily based on recollective processes. Overall, the patterns of results observed here are consistent with the predictions of our theoretical framework (EMRC). We interpret these results and discuss directions for theory development below.
Detecting and Remembering Everyday Changes
The primary goal of the present study was to investigate episodic memory updating and associated effects on source memory in the context of perceived changes in the everyday activities of an actor. To accomplish this, we combined perspectives from the event perception and episodic memory literatures to create a controlled yet naturalistic procedure for examining these everyday cognitive phenomena. Our basic assumption was that effective parsing of perceptual inputs enables encoding and remembering of everyday activity features, which includes relationships between events and regularities in their occurrence. Experiment 1 showed that when activity changes were remembered, source memory for activity features was enhanced in the form of higher correct recall of Day 2 features and fewer intrusions of Day 1 features. Experiment 2 replicated and extended these findings by showing that the memory effects associated with remembering change depended on change being initially detected. Both experiments showed that remembering change was associated with proactive facilitation effects in Day 2 recall, whereas the failure to remember change was associated with proactive interference effects. These findings are consistent with an account of memory for episodic changes proposing that remembering activity features in the context of episodic changes is facilitated when participants can access configural representations the include activity features from multiple episodes and the cognitive operations that inform their temporal relationship (Jacoby et al., 2015; Wahlheim & Jacoby, 2013).
The present results also provided a more precise specification of the conditions required for remembering change to be associated with proactive facilitation. Specifically, such effects were only obtained in both experiments when participants remembered change and could correctly recall Day 1 features when asked following activity type classifications on the cued recall test. Our preferred interpretation is that there is variability is the extent to which activity features from distinct episodes can be effectively integrated into configural representations. This variability is likely to differ across individuals and activities due to differences in the extent to which earlier activity features can be recalled when participants detect that features in an ongoing activity have changed. Further, differences in memory for earlier activity features in the context of naturalistic event perception should depend on differences in perceived event structure, with normative perception being associated with more effective encoding of event features (Zacks et al., 2007). EMRC assumes that configural representations including features from both episodes are formed when earlier features are recollected in the context of perceiving changed features. However, EMRC also assumes that configural representations can be formed even when earlier activity features cannot be recollected, but the quality of the representations will be less elaborate and potentially less informative about the source of activity features. This proposed difference in the structure of configural representations points to the critical role of differences in retrieval of earlier event features that enables detection of episodic changes.
What determines differences in the retrieval of features from prior events when people perceive a new episode? Do these differences reflect variations in the strength of prior episodic representations, or are there qualitatively different routes by which episodic memory biases ongoing processing of event changes? Dual process theories posit that both consciously controlled (recollection) and automatic influences of memory contribute to retrieval across tasks (e.g., Jacoby, 1991; Yonelinas, 2002). Following this assumption, the retrieval of earlier events that enables detection of changes in current events may sometimes be accomplished through recollection or automatic influences. EMRC proposes that detecting change that is accomplished by comparing currently perceived features with recollections of Day 1 features should lead to both features being encoded as part of a configural representation. This type of representation should include information about Day 1 features, Day 2 features, and the temporal relationship between them, resulting in proactive facilitation effects. However, detecting change accomplished by automatic retrieval of Day 1 features would likely not be associated with proactive facilitation effects because configural representations would lack the Day 1 features necessary to elaborate and later cue episodic memory representations for Day 2 features.
The idea that retrieval quality during encoding determines how related episodes will affect source memory is consistent with the finding that hippocampal activity predicts the extent to which encoding of new memories is impaired by reactivating prior memories (e.g., Kuhl, Shah, DuBrow, & Wagner, 2010). Based on this, we suggest that the proactive facilitation observed in the present experiment when change was remembered and Day 1 features were recalled mostly reflected instances when Day 1 features were recollected to enable change detection. In contrast, proactive interference observed when change was remembered but Day 1 features were not recalled likely reflected some instances when change was detected on the basis of automatic influences. These instances may have contributed to such proactive interference effects by impairing encoding of changed activity features. Of course, the proactive interference associated with failure to recall Day 1 features when remembering change could have also resulted from correct classifications of change being based on guessing. Future studies should aim to isolate these contributions to classifications of changed activities.
Consistent with the idea that proactive effects of memory may vary depending on differences in representational structure, the present experiments also showed that the magnitude of proactive interference effects obtained when participants did not correctly classify changed activities during the cued recall test was smaller in Experiment 2 than Experiment 1. These findings are consistent with earlier studies showing that interference effects observed when participants did not correctly classify changed activities at test were greater when the accessibility of earlier memories was enhanced by repetitions (Wahlheim, 2014) and retrieval practice with feedback (Wahlheim, 2015). The present investigation showed that proactive interference effects were greater when each movie was shown in the same session (Experiment 1) rather than different sessions (Experiment 2). This greater distinction between temporal contexts associated with each movie across in Experiment 2 than Experiment 1 may have resulted in event memories competing less with each other. However, there were relatively few instances of activities for which changes were detected but not remembered in Experiment 2, compared to the larger number of instances for which change was not remembered in Experiment 1. It would therefore be valuable to verify these results using a direct within- experiment comparison. If this across-experiment difference is generalizable, it parallels findings showing that context differentiation reduces interference (for reviews, see Abra, 1972; Smith & Vela, 2001). One consequence of this differentiation may be that features associated with distinct but related event models compete less when they are retrieved as separate representations rather than as part of a configural representation.
Adult Age Differences in Change Processing
The results discussed thus far suggest that differences in proactive effects of memory can result from variations in the memorability of individual activities and the extent to which encoding contexts create response competition. In addition to these sources of variability, individual participants are also likely to differ in their ability to encode features from both days together as part of configural representations. Here, we examined adult age differences in change detection and downstream effects on remembering change and associated recall performance. Older adults showed a differential deficit in Day 2 recall for changed activities that was associated with poorer encoding of and later access to configural representations. Evidence for this was shown here as older adults correctly recalled fewer Day 1 features than younger adults when changed activities were correctly classified during the cued recall test.
This age-related deficit may have reflected that older adults detected change during Day 2 viewing on the basis of automatic influences of memory for Day 1 activities more often than younger adults. This would be consistent with the well-established finding that older adults have a selective recollection deficit (e.g., Jennings & Jacoby, 1993). Earlier evidence for this possibility was shown in a study by Wahlheim (2014) using an A-B, A-D paradigm, as older adults were impaired in their recall of List 1 responses when they detected changed responses in List 2. This impairment led older adults to incorrectly classify changed activities at test more often than younger adults, which was associated with older adults’ greater susceptibility to proactive interference. These findings demonstrate how the negative effects of older adults’ impaired encoding of everyday activities (Kurby & Zacks, 2011; Zacks et al., 2006) lead to deficits in detecting changes, which had negative downstream effects on their ability to remember changes and recall activity features. These age-related differences may not be surprising given the constellation of processing deficits suffered by older adults. Indeed, many of the well-established processes responsible for such impairment were likely to have contributed to the age differences reported here. Despite the fact that we cannot isolate the many processes that led to these deficits, these results do implicate a few key aspects of a processing chain that may explain several critical processes involved in older adults’ impoverished event comprehension.
Older adults’ impoverished event comprehension may have also contributed to the impaired metacognitive monitoring shown in their activity type classifications. In Experiment 2, confidence judgments discriminated more poorly between correct and incorrect classifications of activities for older than younger adults. This monitoring deficit may have been partly due to lower fidelity representations being insufficiently diagnostic as bases for classifications. This is consistent with our suggestion that older adults are less likely recollect prior events when viewing ongoing activities. This deficit is also noteworthy because Wahlheim (2014, Experiment 2) showed that older and younger adults’ confidence judgments were comparably sensitive to recall performance differences associated with remembering change. This was despite the fact that the retrieval task placed high demands on recollective processes. Together, these findings indicate that although older adults might show impaired metacognitive accuracy in their judgments regarding the relationships between events, they are largely aware of the memory advantages associated with remembering change. More generally, this discrepancy is similar to findings in the cognitive aging literature showing that older adults are more prone to false remembering (for a review, see Dodson, 2017), but are also acutely aware of their memory deficits, especially when predicting later memory performance (e.g., Hertzog & Dunlosky, 2011). The discrepancy in older and younger adults’ metacognitive accuracy regarding classification of changed activities highlights the need for future studies to illuminate which variables moderate this relationship.
From Memory-for-Change to Event Memory Retrieval and Comparison Theory
The present results replicate earlier findings that can be explained by the memory-for- change framework. But can the memory-for-change framework sufficiently explain the present results? We argue that the memory-for change framework cannot sufficiently explain the present results because it does not account for the predictive processing involved in the perception of temporally structured events. As described in the Introduction, we propose that a merger between the memory-for-change framework and EST can provide a more comprehensive account (EMRC) of how dynamic event changes are encoded and represented, along with the memorial consequences associated with detecting and remembering those changes.
EMRC assumes that the feature overlap between everyday events and prior event representations can cue retrieval of earlier events. We propose that such retrievals can serve to guide predictions of upcoming events, and that the perception of unexpected event features leads to prediction errors. These prediction errors should upregulate attention to unexpected features and trigger event model updating consistent with predictions from EST (Zacks et al., 2007). Current event features should then be encoded with retrieved features and associated cognitive operations into a configural representation, consistent with the memory-for-change framework (Jacoby et al., 2015; Wahlheim & Jacoby, 2013). Importantly, the formation of these representations requires that features of prior events be recollected when changes are detected during their initial encoding. The benefits of such integrative encoding should then be expressed when configural representations are accessed via remembering change. However, retrieval of recent related events when detecting change should increase the strength of earlier-event representations, making them a more robust source of interference when recollective processes cannot be engaged during later retrieval.
Although the findings reported here are consistent with EMRC, the current experiments do not directly demonstrate a role for memory-based predictive processing. However, research from the discourse comprehension literature provides support for the proposal that predictive processing plays a role in detecting episodic changes during event perception. Findings from the discourse comprehension literature are relevant to investigations of event perception as both tasks may be governed by similar underlying processes. Evidence for this has been shown by situation models and event segmentation having similar effects on text and movie comprehension (Baggett, 1979; Lichtenstein & Brewer, 1980; Zacks, Speer, & Reynolds, 2009). In the discourse comprehension literature, evidence for the role of prediction has been shown using eye tracking methods. For example, Altmann and Kamide (1999) examined whether knowledge of semantic relationships would enable prediction of action plans when listening to sentences and viewing objects that corresponded to those sentences. They found that participants looked towards objects that were consistent with spoken verbs prior to those verbs being mentioned in the sentences. These findings were taken as evidence for memory-based anticipation of upcoming features (also see, Kamide, Altmann, & Haywood, 2003).
The discourse comprehension literature might also inform hypotheses about age differences in the use of predictive processing in detecting episodic changes. For example, Federmeier, McLennan, DeOchoa, and Kutas (2002) used event related potentials to examine differences in older and younger adults’ ability to predict word features based on sentence context. Their results showed that sentence context facilitated processing of within-category violations that were related to expected words, for younger but not older adults. This was shown by N400 amplitudes being facilitated for within-category violations less effectively for older adults. These results suggest that older adults may be less able to construct representations that allow for the anticipation of upcoming features. Similar mechanisms may operate in event perception as older adults form less coherent event models that impair episodic retrieval of prior activity features, which diminishes their ability to anticipate upcoming activity features.
EMRC asserts that episodic retrieval influences predictive processing, which in turn affects current encoding. A key challenge for future research is to separate the mechanisms of episodic retrieval and predictive processing in online comprehension. For example, one might directly assay memory retrieval using pattern-based functional MRI (Rissman & Wagner, 2012). In addition, to directly assay predictive processing, a promising approach is to measure anticipatory eye-movements prior to episodic changes. Individuals fixate to objects in naturalistic activities prior to when an actor makes contact with the object (e.g., Hayhoe, Shrivastava, Mruczek, & Pelz, 2003; Land & McLeod, 2000). EMRC predicts that this anticipatory behavior should be heightened by retrieval of related prior events.
In addition to the challenge of directly measuring predictive processing, EMRC will face two other key challenges. The first challenge will be to derive a direct test of the assumptions of EMRC that does not depend on conditionalizing on correct classification of changed activities both when participants just viewed those changes (change detection) and when participants attempt to retrieve whether those changes occurred (remembering change). This conditionalization can introduce selection artifacts due to differences across subjects, activities, and fluctuations in the encoding of activities across the experimental session. Jacoby, Wahlheim, and colleagues have largely addressed the issue of selection artifacts due to subjects and items in earlier studies using hierarchical regression to show that the memorial consequences associated with detecting and remembering change remain when the models account for differences in the memory abilities of subjects and memorability of items (e.g., Jacoby et al., 2013; Wahlheim & Jacoby, 2013). The linear mixed effect models used in the present study improve upon that approach by simultaneously accounting for variability due to subjects and items while assessing differences in memory associated with detecting and remembering change. The present results agree with earlier findings showing memory differences associated with change processing when accounting for correlations due to subjects and items. However, another source of variance, natural fluctuations in attention, is not accounted for by either approach. We are currently planning experiments that directly examine change processing without the need for conditionalization and that allow us to model variance due to attentional fluctuation.
The second challenge will be to establish the generalizability of our findings to event perception and memory in contexts outside of the laboratory. The material set developed for the present experiments represented a compromise between the experimental control offered by a well-characterized stimulus set and the ecological validity offered by naturalistic activities. The current materials hold the most promise for testing whether the effects of change processing will generalize to everyday contexts, as the actions were scripted to be common everyday activities. However, the two version of each activity were not perfect analogs to episodic changes that are encountered in everyday life. Here, the peripheral features of changed events were held constant to highlight the changed features, but peripheral as well as central features will change across events perceived in everyday life. Another direction for future research will be to develop an even more naturalistic set of materials that allows peripheral features to vary in a more naturalistic way to further explore the generalizability of the present findings.
Conclusion
The present study represents the first attempt to integrate perspectives from the event perception, episodic memory, and cognitive aging literatures to examine the memorial consequences associated with detecting and remembering episodic changes in naturalistic activities. These findings open a new avenue for research that cuts across fundamental areas of inquiry in cognitive psychology and neuroscience. These results also demonstrate the viability of this approach for investigating the perception of and memory for changes in naturalistic activities in older and younger adults. Future directions involving a combination of cognitive and neuroscientific methods will allow us to further develop a comprehensive theoretical framework that attempts to explain how individuals update their memory to adapt to changes in the actions of others and how such updating might contribute to their own everyday activities.
Context of the Research.
This research grew from the intersection of research by Wahlheim and Jacoby on the memory- for-change framework with work by Zacks and colleagues on event representations in episodic memory. For us, a key theoretical insight was that the temporal dynamics of change processing are of major theoretical importance. EMRC proposes that, during ongoing comprehension, recent episodic memory representations are retrieved, that the contents of these episodic memories inform one’s working models of the current activity, and that this informs predictions about what will happen in the near future. When such memory-guided predictions lead to errors, this induces a processing cascade that alters new memory encoding. This processing chain illustrates that retrieval and encoding are cyclically linked.
Acknowledgments
This project was funded by NIA grants T32AG000030 and R21AG05231401, and by internal funding from the University of North Carolina at Greensboro. We thank Larry Jacoby for consultation on the experimental designs, Sara Estle for volunteering to be filmed while performing everyday activities, Shaney Flores for his assistance filming, and Jodi Backalar, Heather Bartels, Kathleen Bettencourt, Jemila Collier, Madeleine Fish, Shelly Gupta, Keshav Jayaraman, Jack Kuhns, and Zach Pugh for their assistance with data collection.
Footnotes
The data reported here were presented at the 57th and 58th annual meetings of the Psychonomic Society and the 2017 North Carolina Cognition Conference. The data and stimulus materials are publicly available through the following URL: https://osf.io/t2qjk/.
Author Contributions
Both authors developed the study concept and contributed equally to the study design. C. N. Wahlheim programmed the experiments and supervised data collection and coding. J. M. Zacks performed analyses with input from C N. Wahlheim. C. N. Wahlheim drafted the manuscript, and both authors made revisions. Both authors approved the final version for submission.
References
- Abra JC. List differentiation and forgetting. In: Duncan CP, Sechrest L, Melton AW, editors. Human memory: Festschrift in honor of Benton J Underwood. New York, NY: Appleton-Century Crofts; 1972. pp. 25–57. [Google Scholar]
- Altmann GT, Kamide Y. Incremental interpretation at verbs: Restricting the domain of subsequent reference. Cognition. 1999;73:247–264. doi: 10.1016/s0010-0277(99)00059-1. [DOI] [PubMed] [Google Scholar]
- Anderson MC, Neely JH. Interference and inhibition in memory retrieval. In: Bjork EL, Bjork RA, editors. Handbook of perception and cognition (2nd ed): Memory. San Diego, CA: Academic Press; 1996. pp. 237–313. [Google Scholar]
- Baggett P. Structurally equivalent stories in movie and text and the effect of the medium on recall. Journal of Verbal Learning & Verbal Behavior. 1979;18:333–356. [Google Scholar]
- Baldassano C, Chen J, Zadbood A, Pillow JW, Hasson U, Norman KA. Discovering event structure in continuous narrative perception and memory. Neuron. 2017;95:709–721. doi: 10.1016/j.neuron.2017.06.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balota DA, Dolan PO, Duchek JM. Memory changes in healthy older adults. In: Tulving E, Craik FIM, editors. The Oxford handbook of memory. New York: Oxford University Press; 2000. pp. 395–409. [Google Scholar]
- Balota DA, Duchek JM, Paullin R. Age-related differences in the impact of spacing, lag, and retention interval. Psychology & Aging. 1989;4:3–9. doi: 10.1037//0882-7974.4.1.3. [DOI] [PubMed] [Google Scholar]
- Barnes JM, Underwood BJ. “Fate” of first-list associations in transfer theory. Journal of Experimental Psychology. 1959;58:97–105. doi: 10.1037/h0047507. [DOI] [PubMed] [Google Scholar]
- Bates D, Maechler M, Bolker B, Walker S. Fitting Linear Mixed-Effects Models Using lme4. Journal of Statistical Software. 2015;67:1–48. [Google Scholar]
- Berntsen D. Involuntary autobiographical memory. Applied Cognitive Psychology. 1996;10:435–454. [Google Scholar]
- Boltz M. Temporal accent structure and the remembering of filmed narratives. Journal of Experimental Psychology: Human Perception and Performance. 1992;18:90–105. doi: 10.1037//0096-1523.18.1.90. [DOI] [PubMed] [Google Scholar]
- Brewer WF, Dupree DA. Use of plan schemata in the recall and recognition of goal-directed actions. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1983;9:117–129. [Google Scholar]
- Bruce D, Weaver GE. Retroactive facilitation in short-term retention of minimally learned paired associates. Journal of Experimental Psychology. 1973;100:9–17. [Google Scholar]
- Dickman HR. The perception of behavioral units. In: Barker RG, editor. The stream of behavior. New York: Appleton-Century-Crofts; 1963. pp. 23–41. [Google Scholar]
- Dodson CS. Aging and Memory. In: Byrne JH, editor. Learning and Memory: A Comprehensive Reference 2E, Volume: Cognitive Psychology of Memory. Elsevier; 2017. [Google Scholar]
- Federmeier KD, McLennan DB, De Ochoa E, Kutas M. The impact of semantic memory organization and sentence context information on spoken language processing by younger and older adults: An ERP study. Psychophysiology. 2002;39:133–146. doi: 10.1017/S0048577202001373. [DOI] [PubMed] [Google Scholar]
- Flores S, Bailey HR, Eisenberg ML, Zacks JM. Event segmentation improves event memory up to one month later. Journal of Experimental Psychology: Learning, Memory, & Cognition. 2017;43:1183–1202. doi: 10.1037/xlm0000367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Folstein MF, Folstein SE, McHugh PR. “Mini-mental state”. A practical method for grading the cognitive state of patients for the clinician. Journal of Psychiatric Research. 1975;12:189–198. doi: 10.1016/0022-3956(75)90026-6. [DOI] [PubMed] [Google Scholar]
- Fox J, Weisberg S. An {R} Companion to Applied Regression. Second. Thousand Oaks CA: Sage; 2011. [Google Scholar]
- Glenberg AM. What memory is for. Behavioral and Brain Sciences. 1997;20:1–18. doi: 10.1017/s0140525x97000010. [DOI] [PubMed] [Google Scholar]
- Gold DA, Zacks JM, Flores S. Effects of cues to event segmentation on subsequent memory. Cognitive Research: Principles and Implications. 2017;2:1. doi: 10.1186/s41235-016-0043-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hasher L, Zacks RT. Working memory, comprehension, and aging: A review and a new view. Psychology of Learning and Motivation. 1988;22:193–225. [Google Scholar]
- Hayhoe MM, Shrivastava A, Mruczek R, Pelz JB. Visual memory and motor planning in a natural task. Journal of Vision. 2003;3:49–63. doi: 10.1167/3.1.6. [DOI] [PubMed] [Google Scholar]
- Healey MK, Hasher L, Campbell KL. The role of suppression in resolving interference: Evidence for an age-related deficit. Psychology & Aging. 2013;28:721–728. doi: 10.1037/a0033003. [DOI] [PubMed] [Google Scholar]
- Hertzog C, Dunlosky J. Metacognition in later adulthood: Spared monitoring can benefit older adults’ self-regulation. Current Directions in Psychological Science. 2011;20:167–173. doi: 10.1177/0963721411409026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hintzman DL. Research strategy in the study of memory: Fads, fallacies, and the search for the “coordinates of truth. Perspectives on Psychological Science. 2011;6:253–271. doi: 10.1177/1745691611406924. [DOI] [PubMed] [Google Scholar]
- Hintzman DL, Block RA. Memory for the spacing of repetitions. Journal of Experimental Psychology. 1973;99:70–74. [Google Scholar]
- Hintzman DL. Judgment of frequency vs. recognition confidence: Repetition and recursive reminding. Memory & Cognition. 2004;32:336–350. doi: 10.3758/bf03196863. [DOI] [PubMed] [Google Scholar]
- Hintzman DL. How does repetition affect memory? Evidence from judgments of recency. Memory & Cognition. 2010;38:102–115. doi: 10.3758/MC.38.1.102. [DOI] [PubMed] [Google Scholar]
- Hintzman DL, Summers JJ, Block RA. Spacing judgments as an index of study-phase retrieval. Journal of Experimental Psychology: Human Learning and Memory. 1975;1:31–40. [Google Scholar]
- Jacoby LL. The role of mental contiguity in memory: Registration and retrieval effects. Journal of Verbal Learning and Verbal Behavior. 1974;13:483–496. [Google Scholar]
- Jacoby LL. A process dissociation framework: Separating automatic from intentional uses of memory. Journal of Memory and Language. 1991;30:513–541. [Google Scholar]
- Jacoby LL, Debner JA, Hay JF. Proactive interference, accessibility bias, and process dissociations: Valid subjective reports of memory. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2001;27:686–700. [PubMed] [Google Scholar]
- Jacoby LL, Wahlheim CN. On the importance of looking back: The role of recursive remindings in recency judgments and cued recall. Memory & Cognition. 2013;41:625–637. doi: 10.3758/s13421-013-0298-5. [DOI] [PubMed] [Google Scholar]
- Jacoby LL, Wahlheim CN, Kelley CM. Memory consequences of looking back to notice change: Retroactive and proactive facilitation. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2015;41:1282–1297. doi: 10.1037/xlm0000123. [DOI] [PubMed] [Google Scholar]
- Jacoby LL, Wahlheim CN, Yonelinas AP. The role of detection and recollection of change in list discrimination. Memory & Cognition. 2013;41:638–649. doi: 10.3758/s13421-013-0313-x. [DOI] [PubMed] [Google Scholar]
- Jennings JM, Jacoby LL. Automatic versus intentional uses of memory: Aging, attention, and control. Psychology and Aging. 1993;8:283–293. doi: 10.1037//0882-7974.8.2.283. [DOI] [PubMed] [Google Scholar]
- Kamide Y, Altmann GTM, Haywood SL. The timecourse of prediction in incremental sentence processing: Evidence from anticipatory eye movements. Journal of Memory and Language. 2003;49:133–156. [Google Scholar]
- Kuhl BA, Shah AT, DuBrow S, Wagner AD. Resistance to forgetting associated with hippocampus-mediated reactivation during new learning. Nature Neuroscience. 2010;13:501–506. doi: 10.1038/nn.2498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurby CA, Zacks JM. Age differences in the perception of hierarchical structure in events. Memory & Cognition. 2011;39:75–91. doi: 10.3758/s13421-010-0027-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Land MF, McLeod P. From eye movements to actions: How batsmen hit the ball. Nature Neuroscience. 2000;3:1340–1345. doi: 10.1038/81887. [DOI] [PubMed] [Google Scholar]
- Landis JR, Koch GG. The measurement of observer agreement for categorical data. Biometrics. 1977;33:159–174. [PubMed] [Google Scholar]
- Lenth RV. Least-Squares Means: The R Package lsmeans. Journal of Statistical Software. 2016;69:1–33. [Google Scholar]
- Lichtenstein ED, Brewer WF. Memory for goal-directed events. Cognitive Psychology. 1980;12:412–445. [Google Scholar]
- Naveh-Benjamin M. Adult age differences in memory performance: Tests of an associative deficit hypothesis. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2000;26:1170–1187. doi: 10.1037//0278-7393.26.5.1170. [DOI] [PubMed] [Google Scholar]
- Newtson D, Engquist G. The perceptual organization of ongoing behavior. Journal of Experimental Social Psychology. 1976;12:436–450. [Google Scholar]
- Okado Y, Stark CE. Neural activity during encoding predicts false memories created by misinformation. Learning & Memory. 2005;12:3–11. doi: 10.1101/lm.87605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Putnam AL, Sungkhasettee VW, Roediger HL. When misinformation improves memory: The effects of recollecting change. Psychological Science. 2017;28:36–46. doi: 10.1177/0956797616672268. [DOI] [PubMed] [Google Scholar]
- Putnam AL, Wahlheim CN, Jacoby LL. Memory for flip-flopping: Detection and recollection of political contradictions. Memory & Cognition. 2014;42:1198–1210. doi: 10.3758/s13421-014-0419-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radvansky GA. Situation models, propositions, and the fan effect. Psychonomic Bulletin & Review. 2005;12:478–483. doi: 10.3758/bf03193791. [DOI] [PubMed] [Google Scholar]
- Radvansky GA. Across the event horizon. Current Directions in Psychological Science. 2012;21:269–272. [Google Scholar]
- Rissman J, Wagner AD. Distributed Representations in Memory: Insights from Functional Brain Imaging. Annual Review of Psychology. 2012;63:101–128. doi: 10.1146/annurev-psych-120710-100344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robbins D, Bray JF. The spacing effect and the A-B, A-C paradigm: Evidence for retroactive facilitation. Journal of Experimental Psychology. 1974;103:420–425. [Google Scholar]
- Sargent JQ, Zacks JM, Hambrick DZ, Zacks RT, Kurby CA, Bailey HR, et al. Event segmentation ability uniquely predicts event memory. Cognition. 2013;129:241–255. doi: 10.1016/j.cognition.2013.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schacter DL, Addis DR, Hassabis D, Martin VC, Spreng RN, Szpunar KK. The future of memory: Remembering, imagining, and the brain. Neuron. 2012;76:677–694. doi: 10.1016/j.neuron.2012.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider W, Eschman A, Zuccolotto A. E-Prime: User’s guide. Psychology Software Incorporated; 2002. [Google Scholar]
- Schwan S, Garsoffky B, Hesse FW. Do film cuts facilitate the perceptual and cognitive organization of activity sequences? Memory & Cognition. 2000;28:214–223. doi: 10.3758/bf03213801. [DOI] [PubMed] [Google Scholar]
- Shipley WC. Shipley Institute of Living Scale. Los Angeles: Western Psychological Services; 1986. [Google Scholar]
- Simmons JP, Nelson LD, Simonsohn U. False-positive psychology: Undisclosed flexibility in data collection and analysis allows presenting anything as significant. Psychological Science. 2011;22:1359–1366. doi: 10.1177/0956797611417632. [DOI] [PubMed] [Google Scholar]
- Smith SM, Vela E. Environmental context-dependent memory: A review and meta-analysis. Psychonomic Bulletin & Review. 2001;8:203–220. doi: 10.3758/bf03196157. [DOI] [PubMed] [Google Scholar]
- Swallow KM, Zacks JM, Abrams RA. Event boundaries in perception affect memory encoding and updating. Journal of Experimental Psychology: General. 2009;138:236–257. doi: 10.1037/a0015631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swire B, Ecker UKH, Lewandowsky S. The role of familiarity in correcting inaccurate information. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2017;43:1948–1961. doi: 10.1037/xlm0000422. [DOI] [PubMed] [Google Scholar]
- Tzeng OJL, Cotton B. A study-phase retrieval model of temporal coding. Journal of Experimental Psychology: Human Learning and Memory. 1980;6:705–716. [Google Scholar]
- Wahlheim CN. Proactive effects of memory in young and older adults: The role of change recollection. Memory & Cognition. 2014;42:950–964. doi: 10.3758/s13421-014-0411-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wahlheim CN. Testing can counteract interference by integrating competing information. Memory & Cognition. 2015;43:27–38. doi: 10.3758/s13421-014-0455-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wahlheim CN, Jacoby LL. Remembering change: The critical role of recursive remindings in proactive effects of memory. Memory & Cognition. 2013;41:1–15. doi: 10.3758/s13421-012-0246-9. [DOI] [PubMed] [Google Scholar]
- Wahlheim CN, Zacks JM. Memory guides the comprehension of event changes for older and younger adults. 2017 Oct 4; doi: 10.1037/xge0000458. Retrieved from osf.io/t2qjk. [DOI] [PMC free article] [PubMed]
- Winograd E, Soloway RM. Reminding as a basis for temporal judgments. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1985;11:262–271. doi: 10.1037//0278-7393.11.2.262. [DOI] [PubMed] [Google Scholar]
- Yonelinas AP. The nature of recollection and familiarity: A review of 30 years of research. Journal of Memory and Language. 2002;46:441–517. [Google Scholar]
- Zacks JM, Braver TS, Sheridan MA, Donaldson DI, Snyder AZ, Ollinger JM, et al. Human brain activity time-locked to perceptual event boundaries. Nature Neuroscience. 2001;4:651–655. doi: 10.1038/88486. [DOI] [PubMed] [Google Scholar]
- Zacks JM, Speer NK, Reynolds JR. Segmentation in reading and film comprehension. Journal of Experimental Psychology: General. 2009;138:307–327. doi: 10.1037/a0015305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zacks JM, Speer NK, Swallow KM, Braver TS, Reynolds JR. Event perception: A mind-brain perspective. Psychological Bulletin. 2007;133:273–293. doi: 10.1037/0033-2909.133.2.273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zacks JM, Speer NK, Vettel JM, Jacoby LL. Event understanding and memory in healthy aging and dementia of the Alzheimer type. Psychology and Aging. 2006;21:466–482. doi: 10.1037/0882-7974.21.3.466. [DOI] [PubMed] [Google Scholar]
- Zacks JM, Tversky B. Event structure in perception and conception. Psychological Bulletin. 2001;127:3–21. doi: 10.1037/0033-2909.127.1.3. [DOI] [PubMed] [Google Scholar]
- Zacks RT, Hasher L, Li KZH. Human memory. In: Craik FIM, Salthouse TA, editors. The handbook of aging and cognition. 2nd. Hillsdale, NJ: Erlbaum; 2000. pp. 293–357. [Google Scholar]