

Summary
The neuronal microtubule-associated protein tau, MAPT, is central to the pathogenesis of many dementias. Autosomal-dominant mutations in MAPT cause inherited frontotemporal dementia (FTD), but the underlying pathogenic mechanisms are unclear. Using human stem cell models of FTD due to MAPT mutations, we find that tau becomes hyperphosphorylated and mislocalizes to cell bodies and dendrites in cortical neurons, recapitulating a key early event in FTD. Mislocalized tau in the cell body leads to abnormal microtubule movements in FTD-MAPT neurons that grossly deform the nuclear membrane. This results in defective nucleocytoplasmic transport, which is corrected by microtubule depolymerization. Neurons in the post-mortem human FTD-MAPT cortex have a high incidence of nuclear invaginations, indicating that tau-mediated nuclear membrane dysfunction is an important pathogenic process in FTD. Defects in nucleocytoplasmic transport in FTD point to important commonalities in the pathogenic mechanisms of tau-mediated dementias and ALS-FTD due to TDP-43 and C9orf72 mutations.
Keywords: Alzheimer’s disease, frontotemporal dementia, Tau, MAPT, iPSC, human neurons, nuclear membrane, microtubule dynamics, nucleocytoplasmic transport
Graphical Abstract
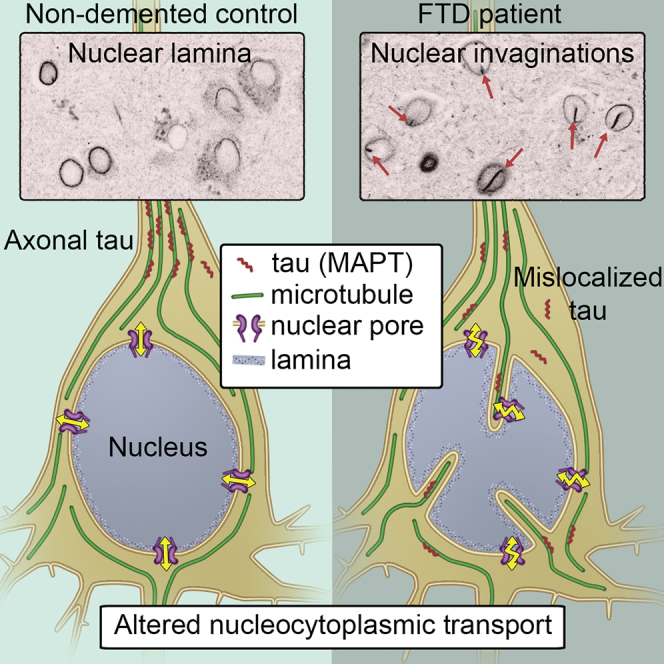
Highlights
-
•
Tau mutations cause microtubule-mediated deformation of the nucleus in dementia
-
•
Nuclear deformation results in defective nucleocytoplasmic transport
-
•
Neuronal nuclei are deformed in the post-mortem frontotemporal dementia brain
-
•
Disrupted nucleocytoplasmic transport is shared in multiple dementias
Paonessa et al. show that mutations in the gene encoding the microtubule-associated protein tau that cause frontotemporal dementia result in microtubule-mediated deformation of the neuronal nucleus and disrupted nucleocytoplasmic transport. These data indicate that perturbed nucleocytoplasmic transport is a pathogenic mechanism in multiple forms of neurodegenerative disease.
Introduction
The microtubule-associated protein tau (MAPT; tau) is involved in the pathogenesis of several different forms of dementia, including Alzheimer’s disease (AD), progressive supranuclear palsy, Pick’s disease, corticobasal degeneration, and frontotemporal dementia (FTD) (Lee et al., 2001, Spillantini and Goedert, 2013). FTD is the third most common cause of dementia, after AD and vascular dementia (Rossor et al., 2010). Autosomal-dominant missense and splicing mutations in MAPT are causes of inherited or familial FTD (FTD-MAPT) (D’Souza et al., 1999, Goedert et al., 2012, Hutton et al., 1998). However, although it is well established that these mutations lead to hyperphosphorylation and aggregation of tau protein in vivo (Ballatore et al., 2007, Goedert et al., 2012), the cell biology of neuronal dysfunction and progressive neurodegeneration in this condition are currently not fully understood.
In healthy neurons, tau protein is almost exclusively localized to the axon, and several mechanisms have been suggested for its highly polarized cellular localization, including selective mRNA and protein transport, local translation, and local degradation (Wang and Mandelkow, 2016). Mislocalization and aggregation of tau in neuronal cell bodies are common features of tau-mediated dementias, including FTD and AD (Fu et al., 2016, Thies and Mandelkow, 2007, Zempel and Mandelkow, 2015). Protein aggregation is widely considered as inherently pathogenic in neurodegeneration (Fitzpatrick et al., 2017, Hernández-Vega et al., 2017), altering many cellular functions, most notably autophagy and proteostasis (Bence et al., 2001, Caballero et al., 2018, Lim and Yue, 2015). However, how MAPT mutations lead to tau hyperphosphorylation and mislocalization, the effects of this mislocalization on neuronal cell biology, and how this contributes to neuronal dysfunction and neurodegeneration all remain poorly understood.
As a typical microtubule-binding protein, tau has several roles in regulating microtubule function and intracellular transport (Wang and Mandelkow, 2016). Tau binds both alpha and beta tubulin subunits of microtubules and has been demonstrated to both stabilize and promote microtubule growth (Kadavath et al., 2015, Witman et al., 1976). The presence of tau on microtubules can alter directions and rates of axonal transport (Dixit et al., 2008, Trinczek et al., 1999). Tau is a natively disordered protein and has recently been found to undergo fluid phase transitions at higher concentrations, nucleating microtubules when it does so (Hernández-Vega et al., 2017). Therefore, it is likely that the changes in tau levels, post-translational modifications, and cellular localization that occur in dementia lead to alterations in microtubule biology, particularly in the neuronal cell body.
To address the question of how MAPT mutations lead to neuronal dysfunction and neurodegeneration, we investigated the effects of two different classes of MAPT mutations on the cell biology of human iPSC-derived cortical neurons. We find that both missense and splicing MAPT mutations cause mislocalization of tau to the cell bodies of neurons and marked changes in microtubule dynamics. Microtubules in the cell bodies of FTD-MAPT neurons actively deform the nuclear membrane, disrupting nucleocytoplasmic transport. Defects in nuclear envelope function, including nucleocytoplasmic transport, are an important pathological process in ALS-FTD because of repeat expansions in C9orf72 and TDP-43 mutations (Chou et al., 2018, Zhang et al., 2015, Zhang et al., 2016, Zhang et al., 2018). Our findings demonstrate that dysfunction of the nuclear membrane due to altered microtubule dynamics is a pathogenic process in dementias involving tau, expanding the group of neurodegenerative diseases that involve disrupted nucleocytoplasmic transport and suggesting common mechanisms of neuronal dysfunctional in these heterogeneous conditions.
Results
Increased Phosphorylation and Altered Cellular Localization of Tau in FTD-MAPT Neurons
To study the effects on neuronal cell biology of FTD-MAPT mutations, we generated excitatory cortical neurons (Shi et al., 2012b) from induced pluripotent stem cells (iPSCs) derived from individuals with different autosomal-dominant mutations in MAPT that are causal for early-onset FTD (Figures 1 and S1). We studied two different types of mutations: the MAPT IVS10+16 autosomal-dominant mutation, which increases inclusion of exon 10, encoding the second microtubule-binding repeat and thus altering the ratio of three (3R) and four (4R) tau isoforms (Hutton et al., 1998, Sposito et al., 2015), and the autosomal-dominant MAPT P301L missense mutation that produces an aggregation-prone form of tau (Wang and Mandelkow, 2016) (Figures 1 and S1). The cortical identity of the neurons generated and the reproducibility of the culture compositions among genotypes was confirmed by assessing the expression of a set of classifier genes that define different neuronal cell types (Figure S1).
Figure 1.

Increased Phosphorylation and Altered Cellular Localization of Tau in FTD-MAPT Neurons
(A) Phosphorylated tau (pS404; AT8 [pS202/pT205]) is increased as a fraction of total tau (epitope 243–441) in frontotemporal dementia (FTD)-MAPT neurons (MAPT IVS10+16-A/B and MAPT P301L) compared with non-demented and MAPT P301 isogenic control neurons (induced pluripotent stem cell [iPSC]-derived neurons at 120 days in vitro [DIV]; three biological replicates). β-actin and β3-tubulin were used as controls. Molecular weight (kDa) is indicated.
(B) Tau isoforms with three (3R; red) or four (4R; inclusion of region 2; green) microtubule-binding regions were detected by western blot analysis of dephosphorylated protein extracts from iPSC-derived control and FTD-MAPT cortical neurons (120 DIV) and from post-mortem human cerebral cortex (non-demented individual). Tau isoforms were identified relative to a commercial tau ladder (Sigma). Molecular weight (kDa) is indicated.
(C) Peptide sequences identified by tau immunoprecipitation (IP)/mass spectrometry from iPSC-derived cortical neurons, confirming the inclusion of repeat 2 (corresponding to exon 10) of 4R tau. In MAPT P301L neurons, both proline and leucine were identified at position 301 (highlighted red). See also Figure S1.
(D) Tau protein is mislocalized to MAP2-positive cell bodies and dendrites in iPSC-derived FTD-MAPT neurons. Confocal images of iPSC-derived control and FTD-MAPT neurons (120 DIV; tau, green; MAP2, red; DAPI, blue). Hyperphosphorylated, AT8-positive tau (AT8; green) is found in cell bodies of FTD-MAPT neurons (arrows) but not in controls (β3-tubulin, red; DAPI, blue). Scale bars, 20 μm.
(E) Increased co-localization of tau and MAP2 protein in FTD-MAPT neurons, compared with non-demented control neurons, analyzed by Pearson’s R correlation (control lines, gray bars; FTD-MAPT lines, black).
Significance was determined for three-sample comparison of non-demented control and two MAPT IVS10+16 lines using one-way ANOVA followed by Tukey’s test (∗p < 0.05 and ∗∗∗p < 0.001). Pairwise comparison of the MAPT P301L line and its isogenic control was carried out using Student’s t test (∗p < 0.05); error bar represents SEM; n = 3 independent experiments.
See also Figure S1.
Total tau content was similar in neurons of each genotype, collectively referred to here as FTD-MAPT neurons (Figure 1A). Notably, tau phosphorylation was increased in FTD-MAPT neurons compared with controls (Figure 1A), including at Ser404 and Ser202/Thr205 (AT8), epitopes typically hyperphosphorylated in tau-mediated dementias (Alonso et al., 2004, Wang et al., 2013). As both mutations are dependent on expression of exon 10 of MAPT, we confirmed translation of exon 10 in neurons generated from all iPSC lines by western blotting and mass spectrometry (Figures 1B, 1C, and S1). No significant difference in insoluble, aggregated tau was detected by sarkosyl extraction between neurons of each genotype (Figure S1).
Mislocalization of tau from axons to neuronal cell bodies and dendrites is an early event in FTD in vivo (Götz et al., 1995, Hoover et al., 2010, Kowall and Kosik, 1987). As expected, control neurons showed a predominantly axonal distribution of tau, with tau largely absent from MAP2-positive neuronal cell bodies and dendrites (Figure 1D). In contrast, tau was commonly present in MAP2-positive cell bodies and dendrites in both MAPT IVS10+16 and P301L neurons (Figures 1D and 1E). Furthermore, tau within cell bodies and dendrites of FTD-MAPT neurons was hyperphosphorylated, as detected by AT8-immunoreactivity (phospho-S202/T205) (Figure 1D).
Microtubules Invade the Nucleus in FTD-MAPT Neurons
Given the mislocalization of tau to the cell bodies of FTD-MAPT neurons, we studied neuronal microtubule dynamics in control and FTD-MAPT neurons (Figures 2 and S2). Actively extending microtubules were live-imaged in iPSC-derived neurons of each genotype by expression of GFP-tagged EB3 (Figure 2; Videos S1 and S2), the microtubule plus-end binding protein (+TIP) (Akhmanova and Steinmetz, 2008). Total microtubule movements were not different between non-demented control and FTD-MAPT neurons, with similar rates of extensions and retractions measured among the various genotypes (Table S1).
Figure 2.

Microtubules Invade the Nucleus in FTD-MAPT Neurons
(A) Total microtubule trajectories (cumulative over a 200 s interval) overlaid on stills from GFP-EB3 live imaging (gray) demonstrate multiple microtubule extensions into the nuclei of FTD-MAPT neurons (MAPT IVS10+16-B and MAPT P301L; 120 DIV), compared with non-demented control neurons; white dotted line indicates the nuclear membrane.
(B) GFP-EB3 growth tracks from representative neurons (cumulative over a 200 s interval) from FTD-MAPT neurons (MAPT IVS10+16-A/B and MAPT P301L) compared with non-demented and MAPT P301 isogenic control neurons (iPSC-derived neurons at 120 DIV; three biological replicates).
Red arrows indicate examples of trajectories within the nucleus. Bars indicate the number of sampled neurons with (black), and without (white), nuclear EB3 growth tracks; n = number of imaged neurons. Scale bars, 10 μm.
Videos obtained from live GFP-EB3 imaging of control (non-demented control and MAPT P301-isogenic) and familial FTD-MAPT (MAPT IVS10+16-A, MAPT IVS10+16-B and MAPT P301L) iPSCs-derived neurons, (100 DIV). Timing is indicated on the top right.
Videos obtained from live GFP-EB3 imaging of familial FTD-MAPT (MAPT IVS10+16-A) iPSCs-derived neurons, (100 DIV) before and after nocodazole (33μM) for two hours. Scale bar = 10 μm.
However, microtubule dynamics were qualitatively different in the cell bodies of control neurons compared with FTD-MAPT neurons. Non-demented control neurons typically had many actively growing microtubules within the cell body that extended around a smooth, oval nucleus (Figures 2A and 2B; Video S1). In contrast, many FTD-MAPT neurons had microtubules with plus ends projecting into the nucleus (15 of 26 FTD-MAPT neurons) (Figure 2B), an event that was infrequently detected in both groups of control neurons (2 of 19) (Figure 2B). Notably, those microtubules that abnormally projected into the nucleus in FTD-MAPT neurons frequently originated from a pronounced focus that resembled a microtubule organizing center (Figure 2A; Video S1). These pronounced foci were not detected in either of the control neuronal lines. We confirmed that the EB3+ microtubules that project into the neuronal nucleus are dynamically growing microtubules, as the mobility of EB3+ comets was greatly reduced following acute microtubule depolymerization (with nocodazole) (Figures S2Band S2C; Video S2).
Microtubules Deform the Nuclear Envelope in FTD-MAPT Neurons
Microtubules couple to the nuclear membrane through the LINC complex (Crisp et al., 2006). This physical association results in transmission of mechanical forces that influence nuclear shape and integrity (Chang et al., 2015), affecting the function of the nuclear envelope (Webster et al., 2009). Given the abnormal projection of microtubules into the nucleus in FTD-MAPT neurons, we studied the shape of the nuclear envelope in iPSC-derived neurons. Marked differences were present in nuclear shape between non-demented controls and FTD-MAPT neurons, as demonstrated by large folds, or invaginations, of the laminB1-positive inner nuclear lamina within the nucleus (Figure 3). The neuronal identity of invaginated cells was confirmed by the co-staining with the cortical deep layer transcription factor CTIP2 and the pan-neuronal protein β3-tubulin (Figure 3).
Figure 3.

Microtubules Deform the Nuclear Envelope in FTD-MAPT Neurons
(A) Marked abnormalities of nuclear lamina shape in FTD-MAPT neurons. Confocal images of the nuclear lamina (laminB1, green) in FTD-MAPT neurons (MAPT IVS10+16-A/B and MAPT P301L; neuronal transcription factor CTIP2, red) compared with non-demented and MAPT P301 isogenic control neurons (120 DIV). White arrows indicate examples of nuclei with pronounced deformation of the nuclear lamina.
(B) FTD-MAPT neurons have increased numbers of cells with deformed nuclear membranes, as defined by the shape of the inner nuclear lamina. Schematic of image analysis method used to quantify nuclear invaginations, as a measure of distortion of the nuclear membrane: nuclear area was established using DAPI (blue), and nuclear lamina (laminB1, green) was assigned as either nuclear boundary or invaginated (i.e., within the nucleus). The fraction of total laminB1 that was invaginated was used empirically to define a threshold for defining neurons as having nuclear membrane invaginations (see Figure S3 for details). Between 25% (MAPT P301L) and 40% (MAPT IVS10+16) of FTD-MAPT neurons have nuclear invaginations, compared with fewer than 10% of control neurons. Significance was determined for non-demented control and two MAPT IVS10+16 lines using one-way ANOVA followed by Tukey’s test (∗p < 0.05); pairwise comparison of the MAPT P301L line and its isogenic control was carried out using Student’s t test (∗p < 0.05); error bar represents SEM; n = 3 independent experiments.
(C) Acute depolymerization of microtubules reverses nuclear lamina invaginations and restores rounded nuclear shapes. Confocal images of control and FTD-MAPT neurons (using genotypes described in A; 120 DIV), treated with DMSO (vehicle) or 10 μM nocodazole for 3 h (laminB1, green; β3-tubulin, red; DAPI, blue). Scale bars, 10 μm.
(D) The proportion of FTD-MAPT neurons with nuclear lamina invaginations is significantly reduced by nocodazole treatment.
Quantification of neurons with abnormalities of the nuclear lamina was carried out as in B. n = 3 independent experiments; error bars represent SEM. Significance was determined using one-way ANOVA followed by Tukey’s test (∗p < 0.05); error bar represents SEM; n = 3 independent experiments.
See also Figure S3.
Quantification of the proportions of neurons with deformation of the nuclear membrane, as defined by the presence of laminB1-positive regions within the nucleus, (Figures 3B and S3), demonstrated that deep nuclear invaginations were present in approximately 25% of MAPT P301L and 40% of MAPT IVS10+16 neurons, compared with fewer than 10% of control neurons (Figure 3B). To confirm that microtubules actively deformed the nucleus in FTD-MAPT neurons, we acutely depolymerized microtubule with the small molecule nocodazole. This significantly reduced the proportion of neurons with nuclear invaginations and restored round nuclear morphology (Figures 3C and 3D). We conclude that the pronounced deformations of the neuronal nuclear membrane in FTD-MAPT neurons are actively mediated by microtubules.
Super-resolution Imaging Demonstrates Close Apposition of Tau and Tubulin within Nuclear Lamina Invaginations
To further study the spatial relationships between tau, microtubules, and the nuclear envelope, we conducted a detailed analysis of the neuronal nucleus in iPSC-derived neurons using three-dimensional (3D) stimulated emission depletion (STED) super-resolution imaging. Three-dimensional STED imaging demonstrated that invaginations of the nuclear lamina present in FTD-MAPT neurons commonly extended deeply into the nucleus, in some cases traversing the entire length of the nucleus, forming pronounced folds (Figure 4A). In comparison, nuclei from non-demented controls had a regular, smooth morphology, with few examples of nuclear lamina invaginations (Figure 4A).
Figure 4.

Super-resolution Imaging Demonstrates Close Apposition of Tau and Tubulin within Nuclear Lamina Invaginations
(A) Three-dimensional reconstructions using STED imaging of the nuclear lamina (laminB1, green) in FTD-MAPT iPSC-derived neurons (MAPT IVS10+16 and MAPT P301L) reveal pronounced nuclear invaginations compared with non-demented and MAPT P301-isogenic control neurons (120 DIV). Scale bar, 10 μm.
(B) Tau is in close proximity to the nuclear lamina within nuclear invaginations of FTD-MAPT neurons. STED imaging of control and FTD-MAPT iPSC-derived neurons (using genotypes described in A; 120 DIV; tau, green; β3-tubulin, red; DNA, Yo-Pro, blue). Detail from white boxes in upper panels, showing both merge of all channels, and single channel images of tau (green) or β3-tubulin (red). Dashed lines indicate the boundary between the nucleus (Nu) and cytoplasm (Cy). Arrows indicate invaginations into the nucleus.
(C) Nuclear invaginations are lined with nuclear lamina and contain tau. STED imaging of MAPT IVS10+16-A neurons (120 DIV; tau, green; laminB1, red). Arrow indicates tau within a nuclear invagination, in close proximity to the laminB1-positive inner nuclear lamina.
As observed by confocal microscopy, tau protein was found to be abundant in the cell bodies of FTD-MAPT neurons by STED imaging and in those neurons was closely apposed to the outer nuclear membrane (Figure 4B). Both tau protein and neuronal tubulin were found within nuclear lamina invaginations in FTD-MAPT neurons (Figure 4B). STED imaging demonstrated that tau within nuclear membrane invaginations is within hundreds of nanometers of the nuclear lamina (Figure 4C). Given that laminB1 filaments line the inner surface of the nuclear envelope, which is typically of the order of 15–60 nm in width (Burke and Stewart, 2013, Gerace and Huber, 2012), we conclude that tau is in close proximity to proteins in the outer membrane of the nuclear envelope in FTD-MAPT neurons.
Tau-Containing Nuclear Lamina Invaginations in Neurons of the Post-mortem FTD-MAPT Cerebral Cortex
Having identified nuclear lamina defects in iPSC-derived FTD-MAPT neurons, we asked whether alterations of the nuclear lamina are also a feature of FTD-MAPT in vivo. To do so, we studied the incidence of invaginations of the nuclear lamina in the frontal and temporal cortex from two separate cohorts from independent brain banks, both containing individuals diagnosed with FTD due to MAPT IVS10+16 mutations and compared with age-matched non-demented controls. These cohorts were analyzed separately using different methods for detecting laminB1. We quantified the fraction of all nuclei with invaginations within each brain region (Figure 5). Similar results were obtained in both cohorts: the frequency of nuclear lamina invaginations was higher in the deep cortical layers in post-mortem cerebral cortices from individuals with FTD due to the MAPT IVS10+16 mutation, compared with non-demented control individuals (Figures 5 and S4). This was the case in both frontal and temporal cortex (Figures 5B, 5E, and 5F).
Figure 5.

Tau-Containing Nuclear Lamina Invaginations in Neurons of the Post-mortem FTD-MAPT Cerebral Cortex
(A) Increased incidence of laminB1-positive nuclear invaginations in vivo, in post-mortem FTD-MAPT IVS10+16 cortex compared with age-matched controls (cohort 1). Immunohistochemistry of laminB1 in (top) frontal and (bottom) temporal cortex from individuals with FTD due to the MAPT IVS10+16 mutation or age-matched controls (non-demented). Red arrows indicate nuclei exhibiting nuclear invaginations.
(B) Percentage of invaginated nuclei in deep layers of frontal and temporal cortex of two control (green) and two MAPT IVS10+16 individuals (red), calculated from 20 individual imaging fields (cohort 1). Nuclei were scored by three observers, blinded to the identity of the post-mortem samples, and the averages of the three measurements are shown. Error represents SEM.
(C) Hyperphosphorylated, AT8+ tau within nuclear lamina invaginations in neurons of the frontal (top) and temporal (bottom) cortex of an individual carrying MAPT IVS10+16 mutation. Representative neurons, showing an extensive nuclear invagination (laminB1, green; DAPI, gray) containing hyperphosphorylated tau (AT8, red). White arrows indicate nuclear invaginations. Scale bars, 10 μm. See also Figure S4.
(D) Representative confocal images from post-mortem FTD-MAPT IVS10+16 cortex compared with age-matched controls (cohort 2) showing laminB1 (red) and DAPI (blue); white arrows indicate nuclear invaginations. Scale bar, 10 μm.
(E and F) Left: percentage of invaginated nuclei in deep layers of frontal (E) and temporal (F) cortex from six aged-matched controls and six FTD-MAPT IVS10+16 individuals (cohort 2) quantified using the analysis method described in Figures 3 and S3. Points indicate quantifications from individual fields. Right: pairwise comparison of the control and FTD-MATP groups show increased percentage of invaginated nuclei in pathology (points indicate individuals; cohort 2) carried out using Student’s t test (∗∗∗p < 0.001); error bar represents SEM.
Data from iPSC-derived FTD-MAPT neurons suggested that laminB1-positive nuclear invaginations would be associated with the presence of phosphorylated tau within the neuronal cell body. Consistent with this, we found that the nuclear lamina was grossly disrupted in neurons that had high levels of hyperphosphorylated (AT8+) tau and neurofibrillary tangles in the post-mortem FTD-MAPT IVS10+16 cerebral cortex (Figures 5C and S4B), and those neurons frequently contained pronounced nuclear lamina invaginations (Figures 5C and S4B). Furthermore, nuclear lamina invaginations in such neurons also commonly contained AT8-positive hyperphosphorylated tau (Figure 5C).
Disrupted Nucleocytoplasmic Transport in FTD-MAPT Neurons
Abnormalities of the nuclear lamina are also found in aging diseases, such as Hutchinson-Gilford progeria syndrome (Broers et al., 2006). Nuclear membrane distortion in response to mechanical forces leads to deleterious effects on many aspects of nuclear function, disrupting nucleocytoplasmic transport (Kelley et al., 2011). We confirmed that the nuclear lamina/membrane invaginations present in iPSC-derived FTD-MAPT neurons also contained nuclear pores within these membrane folds, with nuclear pores (labeled by NUP98) co-localizing with laminB1-positive invaginations (Figure S5A).
To assess whether alterations in the nuclear membrane in FTD-MAPT neurons result in defects in nucleocytoplasmic transport, we expressed NES:GFP and NLS:RFP from a single construct in iPSC-derived neurons (Mertens et al., 2015). This assay enables measurement of the integrity of both nuclear localization and accumulation and cytoplasmic retention and nuclear exclusion within individual neurons (Figures 6A and S5B). Control iPSC-derived neurons had discrete cellular distributions of each protein, with prominent nuclear RFP and cytosolic GFP (Figure 6B). In contrast, localization of NLS:RFP was altered in FTD-MAPT neurons such that there was a marked decrease in the nuclear/cytoplasmic RFP ratio (Figure 6C). Conversely, nuclear exclusion of NES:GFP was reduced in FTD-MAPT neurons, with an increase of GFP within the nucleus (Figure 6C). Together, these data demonstrate defects in the selective permeability of the nuclear envelope in FTD-MAPT neurons, indicating a general failure of nucleocytoplasmic transport within FTD-MAPT neurons.
Figure 6.

Disrupted Nucleocytoplasmic Transport in FTD-MAPT Neurons
(A) Functional assay demonstrates disrupted nucleocytoplasmic transport in human iPSC-derived FTD-MAPT neurons. Schematic illustrates lentiviral vector for co-expression of NES:GFP (nuclear export signal fused to GFP) and NLS:RFP (nuclear localization signal fused to RFP) within human neurons and relative distributions of both proteins in healthy neurons and in cells with defective nucleocytoplasmic transport.
(B) Representative confocal images of control and FTD-MAPT neurons (MAPT IVS10+16-A/B and MAPT P301L; all 120 DIV) expressing GFP:NES and RFP:NLS (GFP, green; RFP, red; β3-tubulin). Grayscale images of NES-GFP and NLS-RFP localization in representative cells of each genotype are shown: FTD-MAPT neurons show an increase of GFP within the nucleus and a reduction in nuclear localization of NLS-RFP. Scale bars, 20 μm.
(C) Quantification of the nuclear/cytoplasmic ratio for both NES:GFP and NLS:RFP demonstrates altered nuclear transport in FTD-MAPT genotypes relative to controls: NES:GFP is increased in the nuclei of FTD-MAPT neurons, whereas NLS:RFP is decreased (points indicate quantifications from individual fields, different shapes indicate measurements from different experiments). Significance was determined for non-demented control and two MAPT IVS10+16 lines using one-way ANOVA followed by Dunnett’s test (∗p < 0.05, ∗∗p < 0.01, and ∗∗∗p < 0.001). Pairwise comparison of the MAPT P301L line and its isogenic control was performed using Student’s t test (∗p < 0.05); error bar represents SEM; n = 4 independent experiments.
(D) Effect of nocodazole (Noc) on the nuclear/cytoplasmic ratio for both NES:GFP and NLS:RFP. Nuclear/cytoplasmic ratio is restored in the presence of nocodazole (+) in FTD-MATP neurons compared with vehicle (−) treated neurons (points indicate quantifications from individual fields, different shapes indicate measurements from different experiments).
Significance was determined using one-way ANOVA followed by Dunnett’s test (∗p < 0.05, ∗∗p < 0.01, and ∗∗∗p < 0.001); error bar represents SEM; n = 3 independent experiments.
See also Figure S5.
Defective nucleocytoplasmic transport may be a cumulative phenomenon due to protracted nuclear membrane damage, or an acute process due to microtubule-mediated nuclear membrane deformation. To distinguish between these mechanisms, we acutely depolymerized microtubules with nocodazole in control and FTD-MAPT neurons and quantified nucleocytoplasmic transport using the NES:GFP/NLS:RFP assay. This treatment corrected the distribution of both NES:GFP and NLS:RFP in FTD-MAPT neurons, restoring the nucleocytoplasmic ratios to the level of healthy control neuron (Figure 6D). We conclude that defective nucleocytoplasmic transport in FTD-MAPT neurons is an ongoing process due to microtubule-mediated deformation of the nuclear membrane.
Discussion
The cellular and molecular biology of the pathogenesis of FTD due to MAPT mutations is not well understood. Currently, it is thought that MAPT mutations all lead to tau protein aggregation and that protein aggregation is the primary driver of neurodegeneration (Ballatore et al., 2007, Spillantini and Goedert, 2013). However, protein aggregation may represent only the late stage of the disease, and the processes preceding and leading to neurofibrillary tangle formation and cellular dysfunction remain to be elucidated. Here we report the use of human stem cell systems to study the effects of those mutations on neuronal cell biology, finding that tau-mediated dementias are accompanied by defective neuronal nucleocytoplasmic transport.
Focusing on two different types of MAPT mutations causal for FTD, we have found that both IVS10+16 and P301L mutations lead to marked defects in nucleocytoplasmic transport in human neurons. We find that both missense and splicing mutations in MAPT alter tau protein localization and phosphorylation within iPSC-derived neurons within 4 months in cell culture, recapitulating a well-described aspect of early FTD pathology in vivo (Götz et al., 1995, Hoover et al., 2010, Kowall and Kosik, 1987), without detectable tau aggregation. Mislocalization of tau in the cell bodies of FTD-MAPT neurons in culture leads to marked changes in microtubule dynamics, causing deformation of the nuclear membrane both in cell culture and in the human FTD-MAPT cortex in vivo. Disruption of the nuclear lamina is commonly associated with dysfunction of the nuclear envelope, and we find marked disruption of nucleocytoplasmic transport in FTD-MAPT neurons. Disrupted nucleocytoplasmic transport is due to ongoing microtubule-mediated nuclear membrane deformation, as it is corrected by acute microtubule depolymerization. Together, these data indicate that perturbation of the function of the nuclear membrane and disruption of nucleocytoplasmic transport is an important pathological process in FTD due to MAPT mutations.
An early event in FTD is the mislocalization of tau from axons to cell bodies and dendrites, and this key stage in disease progression is also an early event in iPSC-derived models of FTD-MAPT. In vivo, mislocalization of tau is typically associated with tau hyperphosphorylation (Götz et al., 1995, Spillantini and Goedert, 2013). We find this also occurs in iPSC-derived FTD-MAPT neurons, where we detected increased tau phosphorylation at serine 202/threonine 205 (the AT8 epitope) and also at serine 404. The sequence in which mislocalization and hyperphosphorylation take place in FTD in vivo, and in iPSC-derived FTD-MAPT neurons in culture, is not currently clear, nor are the mechanisms by which these processes occur. The appearance of tau within cell bodies and dendrites indicates a breakdown of the cellular polarity mechanisms that maintain the axonal enrichment of tau protein and its exclusion from the somatodendritic compartment, mechanisms that are poorly understood.
The two heterozygous, dominant MAPT mutations studied here have different effects on tau protein in neurons. The MAPT P301L missense mutation, like many missense mutations in the microtubule-binding region domain of MAPT, increases the tendency of tau to aggregate in cell-free assays and in transgenic mouse models (von Bergen et al., 2005, Lewis et al., 2000, Shammas et al., 2015). In contrast, the IVS10+16 mutation is not a coding mutation but rather is an intronic single base change that favors the inclusion of exon 10 in the MAPT mRNA, increasing the amount of tau containing four microtubule-binding repeats (4R), relative to the three-repeat (3R) form (Hutton et al., 1998). However, despite these differences, the changes in the forms of tau in both MAPT P301L and IVS10+16 neurons both lead to mislocalization and increased phosphorylation of tau. This finding suggests that either the presence of a pool of P301L tau, or a shift in the 3R/4R tau ratio, alter a common pathway that regulates tau distribution within neurons, tau phosphorylation, or both.
In both MAPT IVS 10+16 and P301L mutant neurons, the appearance of tau in cell bodies is accompanied by marked qualitative changes in neuronal microtubule dynamics. Microtubules in FTD-MAPT neurons actively deform the nuclear envelope, which we find can be reversed by depolymerization of microtubules. Tau has multiple roles in stabilizing microtubules (Wang and Mandelkow, 2016), and microtubules are coupled to the nuclear membrane through the LINC complex (Chang et al., 2015, Crisp et al., 2006, Luo et al., 2016). Therefore, it is likely that the overall effect of the presence of tau in the cell body is to promote microtubule stability, leading to increased pushing forces on the nuclear membrane and the formation of invaginations in the nuclear membrane. As tau has recently also been found to promote microtubule nucleation when undergoing phase transitions at high concentration (Hernández-Vega et al., 2017), accumulation of tau in the cell body may also lead to increased pushing forces on the nucleus by facilitating microtubule nucleation.
Alterations in nuclear shape and nuclear membrane function are a common feature of cellular aging, including in the nervous system, and are associated with multiple deleterious changes in nuclear biology, including chromatin changes and disrupted nucleocytoplasmic transport (Frost, 2016, Oberdoerffer and Sinclair, 2007). Drosophila models of FTD, with neuronal expression of human MAPT R406W, have nuclear shape abnormalities and chromatin changes (Frost et al., 2016). Recently, nuclear membrane disruption has been reported in a MAPT P301L transgenic mouse model of FTD (Eftekharzadeh et al., 2018). Perturbations of the nuclear lamina have been described in the post-mortem AD brain (Frost et al., 2016), including the juxtaposition of neurofibrillary tangles of tau with the nuclear membrane (Sheffield et al., 2006). We also find here an increase in nuclear invaginations in neurons of the human post-mortem MAPT IVS10+16 cortex, and the presence of hyperphosphorylated tau within nuclear invaginations in tangle-bearing neurons. Together, these different studies are consistent with a pathological effect of tau within the neuronal cell body in FTD and AD, whereby the presence of tau alters microtubule biology, resulting in pronounced abnormalities of the neuronal nucleus and defective nucleocytoplasmic transport. Recent findings have shown a direct interaction of aggregated tau with the nuclear pore complex (Eftekharzadeh et al., 2018), suggesting that the microtubule disruption of nuclear pore function reported here may be mediated by tau protein at the nuclear membrane.
Microtubule deformation of the nucleus is a phenotype also seen in classic laminopathies such as the accelerated aging disorder Hutchinson-Gilford progeria syndrome (HGPS), in which the primary defect is due to mutant lamin A/C protein (Capell and Collins, 2006). In that case, microtubules also contribute to nuclear deformations, leading to defects in nucleocytoplasmic transport (Kelley et al., 2011, Larrieu et al., 2014, Snow et al., 2013). Changes in nuclear envelope function in other neurodegenerative diseases, including ALS-FTD due to repeat expansions in C9orf72, Huntington’s disease, and AD, have recently been reported (Freibaum et al., 2015, Grima et al., 2017, Jovičić et al., 2015, Zhang et al., 2015).
Our finding here of disruption of the neuronal nuclear membrane as a consequence of MAPT mutations in FTD extends this pathogenic mechanism to dementias in which protein aggregation has been thought to be the primary driver of neurodegeneration. These data suggest that dysfunction of the nuclear membrane may be a common pathogenic process in diverse neurodegenerative diseases, which could be targeted therapeutically with agents that regulate microtubule functions, nucleocytoplasmic transport, and/or associated processes.
STAR★Methods
Key Resources Table
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Antibodies | ||
| 4R-tau; rabbit | Cosmo Bio | Cat#CAC-TIP-4RT-P01; RRID:N/A |
| Alexa Fluor 488 donkey anti-rabbit | Thermo Fisher Scientific | Cat#A21206; RRID:AB_2535792 |
| Alexa Fluor 488 goat anti-chicken | Thermo Fisher Scientific | Cat#A11039; RRID:AB_142924 |
| Alexa Fluor 594 donkey anti-mouse | Thermo Fisher Scientific | Cat#A21203; RRID:AB_141633 |
| Alexa Fluor 647 donkey anti-rabbit | Thermo Fisher Scientific | Cat#A31573; RRID:AB_2536183 |
| Alexa Fluor 647 goat anti-rat | Thermo Fisher Scientific | Cat#A21247; RRID:AB_141778 |
| CTIP2; rat monoclonal | Abcam | Cat#ab18465; RRID:AB_10015215 |
| GFP; chicken polyclonal | Abcam | Cat#ab13970; RRID:AB_300798 |
| IRDye 680 RD donkey anti-mouse | LI-COR bioscience | Cat#925-68072; RRID:N/A |
| IRDye 800 CW donkey anti-rabbit | LI-COR bioscience | Cat#926-32213; RRID:AB_621848 |
| LaminB1 [119D5-F1]; mouse monoclonal | Abcam | Cat#ab8982; RRID:AB_1640627 |
| LaminB1; rabbit polyclonal | Abcam | Cat#ab16048; RRID:AB_443298 |
| MAP2; chicken polyclonal | Abcam | Cat#ab5392; RRID:AB_2138153 |
| NUP98; rat monoclonal | Abcam | Cat#ab50610; RRID:AB_881769 |
| Phospho-tau pS202/T205 - AT8; mouse monoclonal | Thermo Fisher Scientific | Cat#MN1020; RRID:AB_223647 |
| Phospho-tau S404; rabbit monoclonal | Abcam | Cat#ab92676; RRID:AB_10561457 |
| tau RD3; mouse monoclonal | Millipore | Cat#05-803; RRID:AB_310013 |
| Tbr1; rabbit polyclonal | Abcam | Cat#ab31940; RRID:AB_2200219 |
| total tau HT7; mouse monoclonal | Thermo Fisher Scientific | Cat#MN1000; RRID:AB_2314654 |
| total tau; rabbit polyclonal | Dako Cytomation | Cat#A0024; RRID:AB_10013724 |
| β-actin; mouse monoclonal | Sigma | Cat#A2228; RRID:AB_476697 |
| β3-tubulin; mouse monoclonal | BioLegends | Cat#MMS-435P; RRID:AB_2313773 |
| Biological Samples | ||
| Human brain FFPE sections from frontal and temporal cortex (cohort 1) | Queen’s Square Brain Bank, Institute of Neurology, University College London | Tissue request MTA ID 20170112_UCL |
| Human brain FFPE sections from frontal and temporal cortex (cohort 2) | London Neurodegenerative Diseases Brain Bank and Brains for Dementia Research | Tissue request No 1827 |
| Chemicals, Peptides, and Recombinant Proteins | ||
| 2-Mercaptoethanol | Thermo Fisher Scientific | Cat#21985-023 |
| Accutase | Innovative Cell Technologies | Cat#AT104 |
| B-27 supplement | Thermo Fisher Scientific | Cat#17504-044 |
| Bovine Serum Albumin (BSA) | Sigma | Cat#A2153 |
| cOmplete, Mini, EDTA-free Protease Inhibitor Cocktail Tablets | Sigma | Cat#04693159001 |
| Colloidal Blue Staining Kit | Thermo Fisher Scientific | Cat#LC6025 |
| DAB Peroxidase (HRP) Substrate Kit | Vector Laboratories | Cat#SK-4100 |
| DAPI | Sigma | Cat#D9542 |
| Dispase | Thermo Fisher Scientific | Cat#17105 |
| DL-Dithiothreitol solution | Sigma | Cat#646563 |
| DMEM/F-12, GlutaMAX | Thermo Fisher Scientific | Cat#31331-028 |
| DNase I | New England BioLabs | Cat#M0303S |
| Dimethyl sulfoxide (DMSO) | Sigma | Cat#D2650 |
| Donkey serum | Abcam | Cat#ab7475 |
| Dorsomorphin dihydrochloride | Tocris | Cat#3093 |
| Essential 8 medium | Thermo Fisher Scientific | Cat#A1517001 |
| Fibroblast growth factor 2 (FGF2) | PeproTech | Cat#100-18B |
| Halt Phosphatase Inhibitor Cocktail | Thermo Fisher Scientific | Cat#78420 |
| High Capacity cDNA Reverse Transcription kit | Thermo Fisher Scientific | Cat#4368814 |
| Insulin | Sigma | Cat#19278 |
| L-Glutamine | Thermo Fisher Scientific | Cat#25030-024 |
| Laminin | Sigma | Cat#L2020 |
| N-2 supplement | Thermo Fisher Scientific | Cat#17502-048 |
| N-Lauroylsarcosine sodium salt solution | Sigma | Cat#61747 |
| Neurobasal | Thermo Fisher Scientific | Cat#12348-017 |
| Nocodazole | Tocris | Cat#1228 |
| Non-essential amino acid solution | Thermo Fisher Scientific | Cat#11140-050 |
| NuPAGE LDS Sample Buffer | Thermo Fisher Scientific | Cat#NP0007 |
| Paraformaldehyde (PFA) | Sigma | Cat#158127 |
| PBS | N/A | N/A |
| Penicillin-streptomycin | Thermo Fisher Scientific | Cat#15140-122 |
| Phenylmethanesulfonyl fluoride (PMSF) solution | Sigma | Cat#93482 |
| ProLong Gold Antifade Mountant | Thermo Fisher Scientific | Cat#P36930 |
| RIPA Buffer | Sigma | Cat#R0278 |
| SB431542 | Tocris Bioscience | Cat#1614 |
| Sodium pyruvate | Sigma | Cat#S8636 |
| Sudan Black B | Sigma | Cat# 199664 |
| SYBR Green JumpStart Taq Ready Mix | Sigma | Cat#S4438 |
| Triton X-100 | Sigma | Cat#T8787 |
| TRIzol | Thermo Fisher Scientific | Cat#15596026 |
| Tween 20 | Sigma | Cat#P9416 |
| Trisodium citrate dihydrate | Sigma | Cat#S1804 |
| Critical Commercial Assays | ||
| Precision Red Advanced Protein Assay | Cytoskeleton, Inc. | Cat# ADV02-A |
| Experimental Models: Cell Lines | ||
| Human: non-demented control (NDC) iPSC line | Israel et al., 2012 | N/A |
| Human: MAPT IVS10+16-A and MAPT IVS10+16-B iPSC lines | Sposito et al., 2015 | N/A |
| Human: MAPT P301L and MAPT P301L-isogenic iPSC lines | Janssen Pharmaceutica | IMI STEMBANCC project agreement ICD 483960 |
| Oligonucleotides | ||
| 4RMAPT For 5′ – AAGATCGGCTCCACTGAGAA – 3′ | This paper | N/A |
| 4RMAPT Rev 5′ – CACACTTGGACTGGACGTTG – 3′ | This paper | N/A |
| GAPDH For 5′ – AATGAAGGGGTCATTGATGG – 3′ | This paper | N/A |
| GAPDH Rev 5′ – AAGGTGAAGGTCGGAGTCAA – 3′ | This paper | N/A |
| RPS9 For 5′ – CAGCTTCATCTTGCCCTCAT – 3′ | This paper | N/A |
| RPS9 Rev 5′ – CTGCTGACGCTTGATGAGAA – 3′ | This paper | N/A |
| Recombinant DNA | ||
| dsEGFP-EB3-7 | Michael Davidson | Addgene plasmid # 56474; RRID:Addgene_56474 |
| pLVX-EF1alpha-2xGFP:NES-IRES-2xRFP:NLS | Mertens et al., 2015 | Addgene plasmid #71396; RRID:Addgene_71396 |
| Software and Algorithms | ||
| ABI StepOnePlus software | Thermo Fisher Scientific | N/A; RRID:N/A |
| Fiji | Schindelin et al., 2012 | RRID:SCR_002285 |
| Image Studio Lite | LI-COR | RRID:SCR_013715 |
| Mascot | MATRIX Science | RRID:SCR_014322 |
| plusTipTracker | Applegate et al., 2011 | N/A; RRID:N/A |
| Prism 6 | GraphPad | RRID:SCR_002798 |
Contact for Reagent and Resource Sharing
Further information and requests for resources and reagents should be directed to and will be fulfilled by the Lead Contact, Rick Livesey (r.livesey@ucl.ac.uk).
Experimental Model and Subject Details
Human iPSC lines
MAPT IVS10+16-A and MAPT IVS10+16-B mutant iPSCs were as reported in (Sposito et al., 2015). MAPT P301L was generated from Janssen Pharmaceutica by TALEN editing the line MAPT P301-isogenic, under the IMI STEMBANCC project agreement ICD 483960. The non-demented control line was previously reported (Israel et al., 2012). iPSC cells were growth and expanded in feeder-free conditions using Essential 8 Medium (Thermo Fisher Scientific), at 37°C with 5% CO2. Essential 8 Medium was replaced daily.
Human post-mortem brain sections
Human brain sections were obtained from the Queen’s Square Brain Bank, Institute of Neurology, University College London (cohort one) and from the London Neurodegenerative Diseases Brain Bank and Brains for Dementia Research (cohort two). For cohort one, control brains included one male (age 71) and one female (age 56). FTD-MAPT IVS10+16 brains were from two males (age 52 and 66). For cohort two, control brains were from five males (age 63-77) and one female (age 43). FTD-MAPT IVS10+16 brains were from four males (age 48-71) and two females (age 58 and 63). The use of human post-mortem tissues for this study has been approved with Research Ethics Committee reference ID 08/H0718/54+5 for cohort one and reference ID 08/MRE09/38+5 for cohort two.
Method Details
Generation of iPSC-derived cortical neurons and drug treatments
Differentiation of iPSCs to cortical neurons was carried out as described, with minor modifications (Shi et al., 2012b, Shi et al., 2012a). Briefly, dissociated iPSCs were plated on Geltrex (Thermo Fisher Scientific)-coated plates to reach full confluence. Neural induction was initiated the next day (Day 0) by changing the culture medium to a 1:1 mixture of DMEM/N-2 (DMEM/F-12 GlutaMAX; 1 × N-2; 5 μg ml−1 insulin; 1 mM L-glutamine; 100 μm non-essential amino acids; 100 μM β-mercaptoethanol; 50 U ml−1 penicillin and 50 mg ml−1 streptomycin) and Neurobasal/B-27 (Neurobasal; 1 × B-27; 200 mM L-glutamine; 50 U ml−1 penicillin and 50 mg ml−1 streptomycin) media (hereafter referred as N2B27) supplemented with 1 μM dorsomorphin and 10 μM SB431542 to inhibit TGFβ signaling and support neuronal differentiation and neurogenesis, media was replaced every 24 hours. At day 12 neuroepithelial sheet was harvested and dissociated using the enzyme Dispase and replated on laminin-coated plates. The day after, media was replaced with N2B27 containing 20 ng/mL FGF2. N2B27+FGF2 was added freshly daily for 4 days to promote the maturation of neural rosettes. After 4 days FGF2 was withdrawn and neural rosettes were maintained in N2B27 refreshing medium every other day. At day 30 neural rosettes were dissociated using Accutase and neural progenitor cells were plated on laminin-coated plates at 150,000 cells/mm2. Plated neurons were maintained for up to 120 days with a medium change every other day. To establish identity and quality of cortical neuronal inductions, gene expression profiling was performed on a custom gene expression panel. RNA was isolated from induced cortical neurons using TRIzol (Thermo Fisher Scientific), according to the manufacturer’s instructions. Expression levels of mRNAs enriched in deep and upper layer cortical neurons were assessed using a nanoString (nanoString Technologies) gene expression panel of approximately 250 genes. After subtracting the maximum negative control probe counts, gene counts were normalized using the geometric mean of 6 positive control probes and of 7 housekeeping genes (CLTC, GAPDH, GUSB, PPIA, RPLP1, RPS15A, RPS9). For nocodazole (Tocris) treatment, neurons were grown for 120 days in vitro (DIV) and compound was added at 33 μM for 2 hours before imaging. DMSO was used as vehicle.
Protein extraction and western blot analysis
Total cell protein was extracted using RIPA buffer (Sigma) supplemented with protease inhibitors (Sigma) and Halt phosphatase inhibitors (Thermo Fisher Scientific). Protein quantification was performed using Precision Red Advanced Protein Assay buffer (Cytoskeleton, Inc.). For each sample, 30 μg of protein were mixed with 1X NuPAGE LDS Sample Buffer (Thermo Fisher Scientific) + 1 μM Dithiothreitol. Samples were heated at 100°C for 10 minutes and loaded on NuPAGE 4%–12% Bis-Tris gel (Thermo Fisher Scientific). Afterward, proteins were transferred on PVDF membrane (Millipore) for 1 h at 100 V. Membranes were blocked for another 60 min in 5% BSA in PBST (PBS containing 0.05% Tween 20). All primary antibodies were incubated overnight in 5% milk in PBST at 4°C. Next day, membranes were incubated for at least 1 h in secondary antibody and washed gently in PBST buffer for further 30-60 min. Immunoblots were detected using LI-COR Odyssey CLx Infrared Imaging System and processed with the Image Studio Software (LI-COR).
Sarkosyl extraction
iPSC-derived neurons (120 DIV) were homogenized in Tris-NaCl buffer (25 mM TrisHCl, 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, 5 mM Na4P2O7, pH 7.6) supplemented with 1 mM PMSF, protease inhibitors (Sigma) and Halt phosphatase inhibitors (Thermo Fisher Scientific). Homogenate was subjected to ultracentrifugation at 150,000 g for 30 min at 4°C. The pellet was re-suspended in an equal volume of 10 mM TrisHCl, 0.8 M NaCl, 10% sucrose, pH 7.6 supplemented with 1 mM PMSF, protease inhibitors and Halt phosphatase inhibitors. The re-suspended pellet was centrifuged at 20,000 g for 30 min at 4°C, the supernatant was incubated with 1% sarkosyl (N-lauroylsarkosine sodium salt; Sigma) for 1 hour at room temperature and ultracentrifuged at 150,000 g for 30 min at 4°C. The resulting pellet (sarkosyl insoluble fraction) was resuspended in 1X NuPAGE LDS Sample Buffer (Thermo Fisher Scientific) + 1 mM Dithiothreitol. Samples were heated at 100°C for 10 minutes and loaded on NuPAGE 4%–12% Bis-Tris gel (Thermo Fisher Scientific).
Immunoprecipitation and mass spectrometric analysis of intracellular tau
Tau was immunoprecipitated from 1 mg of total protein extracted from iPSC-derived neurons (120 DIV) using a polyclonal anti-tau antibody (Dako Cytomation). Immunoprecipitated samples were analyzed by western blot using a monoclonal tau antibody (MN1000; Thermo Fisher Scientific) or stained with colloidal blue (Thermo Fisher Scientific). Bands that corresponded to tau by western blot analysis were excised from the colloidal blue SDS-PAGE. Peptide masses of digested protein samples were determined using a Bruker ultrafleXtreme Maldi mass spectrometer in reflectron mode and ms/ms fragmentation performed in LIFT mode. Data analysis was with FlexAnalysis, BioTools and ProteinScape software (Bruker). Database searches of the combined mass fingerprint-ms/ms data were performed using Mascot (http://www.matrixscience.com).
RNA extraction and qRT-PCR analysis
Total RNA was extracted using TRIzol according to manufacturer protocol (Invitrogen). 1 μg of RNA was treated with DNase I (New England BioLabs) and 500 ng were retrotranscribed using the High Capacity cDNA Reverse Transcription kit (Applied Biosystems). qRT-PCR were performed in a StepOnePlus instrument (Applied Biosystems) using the SYBR Green JumpStart Taq Ready Mix (Sigma) in a final volume of 15 μl, using the following protocol: 95°C for 2 min, 40 cycles at 94°C for 15 s, 60°C for 30 s. MAPT mRNA expression was assessed relative to GAPDH and RPS9 housekeeping genes using the specific primers reported in the Key Resource Table. Results were analyzed using the ABI StepOnePlus software (Thermo Fisher Scientific).
Confocal microscopy and image analysis
For immunofluorescent labeling and imaging, cells were washed 3 times in PBS and then fixed using 4% paraformaldehyde (v/v) in PBS for 15 minutes at RT. After 3 washes in PBS, cells were permeabilised in PBS+0.3% Triton X-100 (Sigma; Tx) for 15 minutes at room temperature (RT). After 3 washes in PBS, cells were blocked using 5% donkey serum in PBS+0.3% Triton X-100 (PBS-Tx+5% DS) for 1 hr at RT. For AT8-tau immunostaining (ThermoFisher; MN1020), 5% BSA (Sigma) (w/v) in PBS+0.3% Triton X-100 (PBS-Tx+5% BSA) was used as a blocking agent. Primary antibodies were diluted in PBS-Tx+5% DS or PBS-Tx+5% BSA as indicated below and incubated overnight at 4°C. Cells were washed 3 times in PBS and incubated 1 hr in the dark at RT with secondary antibodies diluted 1:1000 in PBS-Tx+5% DS or PBS-Tx+5% BSA. After 3 washes in PBS, samples were incubated for 5 minutes at RT with DAPI diluted 1:5000 in PBS and then washed 3 additional times with PBS. Samples were mounted using ProLong Gold antifade (Thermo Fisher Scientific).
Standard confocal images were acquired with an Olympus Inverted FV1000 confocal microscope and processed using Fiji software (Schindelin et al., 2012). STED imaging was performed on a custom built, dual color, beam scanning system with gated detection optically identical to the instrument described in (Bottanelli et al., 2016).
For image analysis of colocalization of tau and MAP2, Pearson’s R correlation was calculated using the Coloc2 plugin for Fiji (https://imagej.net/Coloc2). To quantify nuclear invaginations neurons were co-stained for LaminB1 and DAPI. Nuclear lamina signal was assigned as either nuclear boundary or invaginated (i.e., within the DAPI stained area defining the nucleus) using a custom plugin for the Fiji bioimage analysis software. Nuclei with a proportion of invaginated laminB1 that exceeded 0.3 were considered as invagination positive (see Figure 3B and S3 for details). At least 5 imaging fields from three independent experiments for genotype were analyzed.
Staining of human formalin-fixed paraffin embedded (FFPE) brain sections
FFPE slides were deparaffinised and rehydrated, then boiled for 20 minutes in 10 mM Tri-sodium citrate buffer, pH6 + 0.05% Tween 20, for antigen retrieval. Slides were cooled to RT before staining. 3,3′-Diaminobenzidine (DAB) staining was performed using the DAB Peroxidase (HRP) Substrate Kit (Vector Laboratories) according to manufacturer’s instructions. After DAB staining, whole sections were imaged using an Axio Scan.Z1 microscope (Zeiss). For immunofluorescence, after antigen retrieval slides were blocked using 5% BSA in PBS+0.3% Triton X-100 (PBS-Tx+5% BSA) for 20 minutes at RT. Primary antibodies were diluted in PBS-Tx+5% BSA as indicated below and incubated O/N at 4°C in humidified chamber. Slides were washed 3 times in PBS and incubated for 30 minutes in the dark at RT with secondary antibodies diluted 1:1000 in PBS-Tx+5% BSA. After 3 washes in PBS, samples were incubated 5 minutes with DAPI diluted 1:5000 in PBS. After 3 washes in PBS, slides were incubated for 20 minutes with 0.01% Sudan Black B (Sigma) in 70% ethanol. Samples were mounted using ProLong Gold antifade (Thermo Fisher Scientific). Images were acquired through Olympus Inverted FV1000 confocal microscope and processed using the Fiji software.
For cohort one, nuclear lamina invaginations were quantified after 3,3′-Diaminobenzidine (DAB) staining of LaminB1. Nuclei were scored from 20 randomly acquired imaging fields from each individual. Nuclei were considered positive (folded) if invaginations extended into the nuclear interior for at least 3 μm. The percentage of folded respect to total nuclei was calculated for each imaging field. For cohort two, neurons were co-stained for LaminB1 and DAPI. Nuclear lamina signal was assigned as either nuclear boundary or invaginated (i.e., within the DAPI stained area defining the nucleus) using custom plugin for the Fiji bioimage analysis software. Nuclei with a proportion of invaginated laminB1 that exceeded 0.3 were considered as invagination positive (see Figure 3B and S3 for details).
Live imaging of microtubule dynamics
Neurons were grown to 100 DIV in individual μ-Dish 35 mm dishes (Ibidi) and transfected with a plasmid encoding for GFP-EB3 (gift from Michael Davidson; Addgene plasmid # 56474). 48h after transfection, neurons were subjected to live imaging using a Leica SP5 microscope equipped with a controlled environment chamber (37°C; 5% CO2). Images were acquired at resonant scanning with a 63x objective (1frame/sec). Resulting movies were analyzed using the plusTipTracker software (Applegate et al., 2011).
Nucleocytoplasmic transport assay
Nucleocytoplasmic trafficking was analyzed by infection of 120 DIV human iPSC-derived neurons with the pLVX-EF1alpha-2xGFP:NES-IRES-2xRFP:NLS construct (Addgene plasmid #71396; Mertens et al., 2015). After 6 days, neurons were fixed and immunostained for β3-tubulin and GFP. Only cells positive for neuron-specific β3-tubulin were considered. The nuclear to cytoplasmic ratios of both GFP and RFP (nucRFP:cytRFP and nucGFP:cytGFP) were calculated separately using the integrated density of ROIs drawn within and outside the nucleus (see Fig.S5 for details).
Quantification and statistical analysis
Unless otherwise specified, data are presented as mean values of the number of independently conducted experiments indicated in the legend of each figure. Error bars represent the standard error of mean (SEM). Statistical analysis was performed using the Prism6 analytical software (GraphPad). Unpaired Student’s t test was used to compare differences between two groups, assuming the data were normally distributed. One-way ANOVA followed by Tukey’s or Dunnett’s correction for multiple testing (as indicated in figure legends) was used to analyze the differences between more than two groups. ∗∗∗ p < 0.01, ∗∗p < 0.01, ∗p < 0.05.
Acknowledgments
F.J.L.’s group is supported by a Wellcome Trust Senior Investigator Award (WT101052MA), the Alborada Trust’s funding of the Alzheimer’s Research UK Stem Cell Research Centre, and Dementias Platform UK. S.P.J. is a Wellcome Trust Senior Investigator. S.W. and J.H. received funding from the National Institute for Health Research University College London Hospitals Biomedical Research Centre and S.W. from an ARUK Senior Research Fellowship (ARUK-SRF2016B-2). D.L. is a Wellcome Trust Henry Dale fellow (86609). Research in S.P.J.’s and F.J.L.’s groups benefits from core support to the Gurdon Institute from the Wellcome Trust and Cancer Research UK.
Author Contributions
F.P., L.D.E., D.L., and F.J.L. conceived and designed the experiments. F.P. and L.D.E. collected and analyzed the experimental data. F.P., L.D.E., R.S., and F.J.L. wrote the manuscript. J.H. and S.W. contributed cell lines. All authors edited and approved the final manuscript.
Declaration of Interests
The authors declare no competing interests.
Published: January 15, 2019
Footnotes
Supplemental Information includes five figures, one table, and two videos and can be found with this article online at https://doi.org/10.1016/j.celrep.2018.12.085.
Supplemental Information
Analysis recombinant microtubule tip protein EB3 growth speed, life-time, pause events and shrinking events in iPSC-derived neurons for each genotype in the study (MAPT IVS10+16-A/B, MAPT P301L, non-demented control and MAPT P301-isogenic control). Number of cells (n) analyzed per genotype are indicated in the table.
References
- Akhmanova A., Steinmetz M.O. Tracking the ends: a dynamic protein network controls the fate of microtubule tips. Nat. Rev. Mol. Cell Biol. 2008;9:309–322. doi: 10.1038/nrm2369. [DOI] [PubMed] [Google Scholar]
- Alonso Adel.C., Mederlyova A., Novak M., Grundke-Iqbal I., Iqbal K. Promotion of hyperphosphorylation by frontotemporal dementia tau mutations. J. Biol. Chem. 2004;279:34873–34881. doi: 10.1074/jbc.M405131200. [DOI] [PubMed] [Google Scholar]
- Applegate K.T., Besson S., Matov A., Bagonis M.H., Jaqaman K., Danuser G. plusTipTracker: quantitative image analysis software for the measurement of microtubule dynamics. J. Struct. Biol. 2011;176:168–184. doi: 10.1016/j.jsb.2011.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ballatore C., Lee V.M.-Y., Trojanowski J.Q. Tau-mediated neurodegeneration in Alzheimer’s disease and related disorders. Nat. Rev. Neurosci. 2007;8:663–672. doi: 10.1038/nrn2194. [DOI] [PubMed] [Google Scholar]
- Bence N.F., Sampat R.M., Kopito R.R. Impairment of the ubiquitin-proteasome system by protein aggregation. Science. 2001;292:1552–1555. doi: 10.1126/science.292.5521.1552. [DOI] [PubMed] [Google Scholar]
- Bottanelli F., Kromann E.B., Allgeyer E.S., Erdmann R.S., Wood Baguley S., Sirinakis G., Schepartz A., Baddeley D., Toomre D.K., Rothman J.E., Bewersdorf J. Two-colour live-cell nanoscale imaging of intracellular targets. Nat. Commun. 2016;7:10778. doi: 10.1038/ncomms10778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Broers J.L.V., Ramaekers F.C.S., Bonne G., Yaou R.B., Hutchison C.J. Nuclear lamins: laminopathies and their role in premature ageing. Physiol. Rev. 2006;86:967–1008. doi: 10.1152/physrev.00047.2005. [DOI] [PubMed] [Google Scholar]
- Burke B., Stewart C.L. The nuclear lamins: flexibility in function. Nat. Rev. Mol. Cell Biol. 2013;14:13–24. doi: 10.1038/nrm3488. [DOI] [PubMed] [Google Scholar]
- Caballero B., Wang Y., Diaz A., Tasset I., Juste Y.R., Stiller B., Mandelkow E.-M., Mandelkow E., Cuervo A.M. Interplay of pathogenic forms of human tau with different autophagic pathways. Aging Cell. 2018;17:e12692. doi: 10.1111/acel.12692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Capell B.C., Collins F.S. Human laminopathies: nuclei gone genetically awry. Nat. Rev. Genet. 2006;7:940–952. doi: 10.1038/nrg1906. [DOI] [PubMed] [Google Scholar]
- Chang W., Worman H.J., Gundersen G.G. Accessorizing and anchoring the LINC complex for multifunctionality. J. Cell Biol. 2015;208:11–22. doi: 10.1083/jcb.201409047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou C.-C., Zhang Y., Umoh M.E., Vaughan S.W., Lorenzini I., Liu F., Sayegh M., Donlin-Asp P.G., Chen Y.H., Duong D.M. TDP-43 pathology disrupts nuclear pore complexes and nucleocytoplasmic transport in ALS/FTD. Nat. Neurosci. 2018;21:228–239. doi: 10.1038/s41593-017-0047-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crisp M., Liu Q., Roux K., Rattner J.B., Shanahan C., Burke B., Stahl P.D., Hodzic D. Coupling of the nucleus and cytoplasm: role of the LINC complex. J. Cell Biol. 2006;172:41–53. doi: 10.1083/jcb.200509124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D’Souza I., Poorkaj P., Hong M., Nochlin D., Lee V.M., Bird T.D., Schellenberg G.D. Missense and silent tau gene mutations cause frontotemporal dementia with parkinsonism-chromosome 17 type, by affecting multiple alternative RNA splicing regulatory elements. Proc. Natl. Acad. Sci. U S A. 1999;96:5598–5603. doi: 10.1073/pnas.96.10.5598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dixit R., Ross J.L., Goldman Y.E., Holzbaur E.L.F. Differential regulation of dynein and kinesin motor proteins by tau. Science. 2008;319:1086–1089. doi: 10.1126/science.1152993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eftekharzadeh B., Daigle J.G., Kapinos L.E., Coyne A., Schiantarelli J., Carlomagno Y., Cook C., Miller S.J., Dujardin S., Amaral A.S. Tau protein disrupts nucleocytoplasmic transport in Alzheimer’s disease. Neuron. 2018;99:925–940.e7. doi: 10.1016/j.neuron.2018.07.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fitzpatrick A.W.P., Falcon B., He S., Murzin A.G., Murshudov G., Garringer H.J., Crowther R.A., Ghetti B., Goedert M., Scheres S.H.W. Cryo-EM structures of tau filaments from Alzheimer’s disease. Nature. 2017;547:185–190. doi: 10.1038/nature23002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freibaum B.D., Lu Y., Lopez-Gonzalez R., Kim N.C., Almeida S., Lee K.-H., Badders N., Valentine M., Miller B.L., Wong P.C. GGGGCC repeat expansion in C9orf72 compromises nucleocytoplasmic transport. Nature. 2015;525:129–133. doi: 10.1038/nature14974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frost B. Alzheimer’s disease: An acquired neurodegenerative laminopathy. Nucleus. 2016;7:275–283. doi: 10.1080/19491034.2016.1183859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frost B., Bardai F.H., Feany M.B. Lamin dysfunction mediates neurodegeneration in tauopathies. Curr. Biol. 2016;26:129–136. doi: 10.1016/j.cub.2015.11.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fu H., Hussaini S.A., Wegmann S., Profaci C., Daniels J.D., Herman M., Emrani S., Figueroa H.Y., Hyman B.T., Davies P., Duff K.E. 3D visualization of the temporal and spatial spread of tau pathology reveals extensive sites of tau accumulation associated with neuronal loss and recognition memory deficit in aged tau transgenic mice. PLoS ONE. 2016;11:e0159463. doi: 10.1371/journal.pone.0159463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerace L., Huber M.D. Nuclear lamina at the crossroads of the cytoplasm and nucleus. J. Struct. Biol. 2012;177:24–31. doi: 10.1016/j.jsb.2011.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goedert M., Ghetti B., Spillantini M.G. Frontotemporal dementia: implications for understanding Alzheimer disease. Cold Spring Harb. Perspect. Med. 2012;2:a006254. doi: 10.1101/cshperspect.a006254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Götz J., Probst A., Spillantini M.G., Schäfer T., Jakes R., Bürki K., Goedert M. Somatodendritic localization and hyperphosphorylation of tau protein in transgenic mice expressing the longest human brain tau isoform. EMBO J. 1995;14:1304–1313. doi: 10.1002/j.1460-2075.1995.tb07116.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grima J.C., Daigle J.G., Arbez N., Cunningham K.C., Zhang K., Ochaba J., Geater C., Morozko E., Stocksdale J., Glatzer J.C. Mutant Huntingtin disrupts the nuclear pore complex. Neuron. 2017;94:93–107.e6. doi: 10.1016/j.neuron.2017.03.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hernández-Vega A., Braun M., Scharrel L., Jahnel M., Wegmann S., Hyman B.T., Alberti S., Diez S., Hyman A.A. Local nucleation of microtubule bundles through tubulin concentration into a condensed tau phase. Cell Rep. 2017;20:2304–2312. doi: 10.1016/j.celrep.2017.08.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoover B.R., Reed M.N., Su J., Penrod R.D., Kotilinek L.A., Grant M.K., Pitstick R., Carlson G.A., Lanier L.M., Yuan L.-L. Tau mislocalization to dendritic spines mediates synaptic dysfunction independently of neurodegeneration. Neuron. 2010;68:1067–1081. doi: 10.1016/j.neuron.2010.11.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hutton M., Lendon C.L., Rizzu P., Baker M., Froelich S., Houlden H., Pickering-Brown S., Chakraverty S., Isaacs A., Grover A. Association of missense and 5′-splice-site mutations in tau with the inherited dementia FTDP-17. Nature. 1998;393:702–705. doi: 10.1038/31508. [DOI] [PubMed] [Google Scholar]
- Israel M.A., Yuan S.H., Bardy C., Reyna S.M., Mu Y., Herrera C., Hefferan M.P., Van Gorp S., Nazor K.L., Boscolo F.S. Probing sporadic and familial Alzheimer’s disease using induced pluripotent stem cells. Nature. 2012;482:216–220. doi: 10.1038/nature10821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jovičić A., Mertens J., Boeynaems S., Bogaert E., Chai N., Yamada S.B., Paul J.W., 3rd, Sun S., Herdy J.R., Bieri G. Modifiers of C9orf72 dipeptide repeat toxicity connect nucleocytoplasmic transport defects to FTD/ALS. Nat. Neurosci. 2015;18:1226–1229. doi: 10.1038/nn.4085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kadavath H., Hofele R.V., Biernat J., Kumar S., Tepper K., Urlaub H., Mandelkow E., Zweckstetter M. Tau stabilizes microtubules by binding at the interface between tubulin heterodimers. Proc. Natl. Acad. Sci. U S A. 2015;112:7501–7506. doi: 10.1073/pnas.1504081112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelley J.B., Datta S., Snow C.J., Chatterjee M., Ni L., Spencer A., Yang C.-S., Cubeñas-Potts C., Matunis M.J., Paschal B.M. The defective nuclear lamina in Hutchinson-Gilford progeria syndrome disrupts the nucleocytoplasmic Ran gradient and inhibits nuclear localization of Ubc9. Mol. Cell. Biol. 2011;31:3378–3395. doi: 10.1128/MCB.05087-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kowall N.W., Kosik K.S. Axonal disruption and aberrant localization of tau protein characterize the neuropil pathology of Alzheimer’s disease. Ann. Neurol. 1987;22:639–643. doi: 10.1002/ana.410220514. [DOI] [PubMed] [Google Scholar]
- Larrieu D., Britton S., Demir M., Rodriguez R., Jackson S.P. Chemical inhibition of NAT10 corrects defects of laminopathic cells. Science. 2014;344:527–532. doi: 10.1126/science.1252651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee V.M., Goedert M., Trojanowski J.Q. Neurodegenerative tauopathies. Annu. Rev. Neurosci. 2001;24:1121–1159. doi: 10.1146/annurev.neuro.24.1.1121. [DOI] [PubMed] [Google Scholar]
- Lewis J., McGowan E., Rockwood J., Melrose H., Nacharaju P., Van Slegtenhorst M., Gwinn-Hardy K., Paul Murphy M., Baker M., Yu X. Neurofibrillary tangles, amyotrophy and progressive motor disturbance in mice expressing mutant (P301L) tau protein. Nat. Genet. 2000;25:402–405. doi: 10.1038/78078. [DOI] [PubMed] [Google Scholar]
- Lim J., Yue Z. Neuronal aggregates: formation, clearance, and spreading. Dev. Cell. 2015;32:491–501. doi: 10.1016/j.devcel.2015.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo Y., Lee I.-W., Jo Y.-J., Namgoong S., Kim N.-H. Depletion of the LINC complex disrupts cytoskeleton dynamics and meiotic resumption in mouse oocytes. Sci. Rep. 2016;6:20408. doi: 10.1038/srep20408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mertens J., Paquola A.C.M., Ku M., Hatch E., Böhnke L., Ladjevardi S., McGrath S., Campbell B., Lee H., Herdy J.R. Directly reprogrammed human neurons retain aging-associated transcriptomic signatures and reveal age-related nucleocytoplasmic defects. Cell Stem Cell. 2015;17:705–718. doi: 10.1016/j.stem.2015.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oberdoerffer P., Sinclair D.A. The role of nuclear architecture in genomic instability and ageing. Nat. Rev. Mol. Cell Biol. 2007;8:692–702. doi: 10.1038/nrm2238. [DOI] [PubMed] [Google Scholar]
- Rossor M.N., Fox N.C., Mummery C.J., Schott J.M., Warren J.D. The diagnosis of young-onset dementia. Lancet Neurol. 2010;9:793–806. doi: 10.1016/S1474-4422(10)70159-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schindelin J., Arganda-Carreras I., Frise E., Kaynig V., Longair M., Pietzsch T., Preibisch S., Rueden C., Saalfeld S., Schmid B. Fiji: an open-source platform for biological-image analysis. Nat. Methods. 2012;9:676–682. doi: 10.1038/nmeth.2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shammas S.L., Garcia G.A., Kumar S., Kjaergaard M., Horrocks M.H., Shivji N., Mandelkow E., Knowles T.P.J., Mandelkow E., Klenerman D. A mechanistic model of tau amyloid aggregation based on direct observation of oligomers. Nat. Commun. 2015;6:7025. doi: 10.1038/ncomms8025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheffield L.G., Miskiewicz H.B., Tannenbaum L.B., Mirra S.S. Nuclear pore complex proteins in Alzheimer disease. J. Neuropathol. Exp. Neurol. 2006;65:45–54. doi: 10.1097/01.jnen.0000195939.40410.08. [DOI] [PubMed] [Google Scholar]
- Shi Y., Kirwan P., Livesey F.J. Directed differentiation of human pluripotent stem cells to cerebral cortex neurons and neural networks. Nat. Protoc. 2012;7:1836–1846. doi: 10.1038/nprot.2012.116. [DOI] [PubMed] [Google Scholar]
- Shi Y., Kirwan P., Smith J., Robinson H.P.C., Livesey F.J. Human cerebral cortex development from pluripotent stem cells to functional excitatory synapses. Nat. Neurosci. 2012;15:477–486. doi: 10.1038/nn.3041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snow C.J., Dar A., Dutta A., Kehlenbach R.H., Paschal B.M. Defective nuclear import of Tpr in Progeria reflects the Ran sensitivity of large cargo transport. J. Cell Biol. 2013;201:541–557. doi: 10.1083/jcb.201212117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spillantini M.G., Goedert M. Tau pathology and neurodegeneration. Lancet Neurol. 2013;12:609–622. doi: 10.1016/S1474-4422(13)70090-5. [DOI] [PubMed] [Google Scholar]
- Sposito T., Preza E., Mahoney C.J., Setó-Salvia N., Ryan N.S., Morris H.R., Arber C., Devine M.J., Houlden H., Warner T.T. Developmental regulation of tau splicing is disrupted in stem cell-derived neurons from frontotemporal dementia patients with the 10 + 16 splice-site mutation in MAPT. Hum. Mol. Genet. 2015;24:5260–5269. doi: 10.1093/hmg/ddv246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thies E., Mandelkow E.-M. Missorting of tau in neurons causes degeneration of synapses that can be rescued by the kinase MARK2/Par-1. J. Neurosci. 2007;27:2896–2907. doi: 10.1523/JNEUROSCI.4674-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trinczek B., Ebneth A., Mandelkow E.M., Mandelkow E. Tau regulates the attachment/detachment but not the speed of motors in microtubule-dependent transport of single vesicles and organelles. J. Cell Sci. 1999;112:2355–2367. doi: 10.1242/jcs.112.14.2355. [DOI] [PubMed] [Google Scholar]
- von Bergen M., Barghorn S., Biernat J., Mandelkow E.-M., Mandelkow E. Tau aggregation is driven by a transition from random coil to beta sheet structure. Biochim. Biophys. Acta. 2005;1739:158–166. doi: 10.1016/j.bbadis.2004.09.010. [DOI] [PubMed] [Google Scholar]
- Wang Y., Mandelkow E. Tau in physiology and pathology. Nat. Rev. Neurosci. 2016;17:5–21. doi: 10.1038/nrn.2015.1. [DOI] [PubMed] [Google Scholar]
- Wang J.-Z., Xia Y.-Y., Grundke-Iqbal I., Iqbal K. Abnormal hyperphosphorylation of tau: sites, regulation, and molecular mechanism of neurofibrillary degeneration. J. Alzheimers Dis. 2013;33(Suppl 1):S123–S139. doi: 10.3233/JAD-2012-129031. [DOI] [PubMed] [Google Scholar]
- Webster M., Witkin K.L., Cohen-Fix O. Sizing up the nucleus: nuclear shape, size and nuclear-envelope assembly. J. Cell Sci. 2009;122:1477–1486. doi: 10.1242/jcs.037333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Witman G.B., Cleveland D.W., Weingarten M.D., Kirschner M.W. Tubulin requires tau for growth onto microtubule initiating sites. Proc. Natl. Acad. Sci. U S A. 1976;73:4070–4074. doi: 10.1073/pnas.73.11.4070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zempel H., Mandelkow E.-M. Tau missorting and spastin-induced microtubule disruption in neurodegeneration: Alzheimer disease and hereditary spastic paraplegia. Mol. Neurodegener. 2015;10:68. doi: 10.1186/s13024-015-0064-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang K., Donnelly C.J., Haeusler A.R., Grima J.C., Machamer J.B., Steinwald P., Daley E.L., Miller S.J., Cunningham K.M., Vidensky S. The C9orf72 repeat expansion disrupts nucleocytoplasmic transport. Nature. 2015;525:56–61. doi: 10.1038/nature14973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y.-J., Gendron T.F., Grima J.C., Sasaguri H., Jansen-West K., Xu Y.-F., Katzman R.B., Gass J., Murray M.E., Shinohara M. C9ORF72 poly(GA) aggregates sequester and impair HR23 and nucleocytoplasmic transport proteins. Nat. Neurosci. 2016;19:668–677. doi: 10.1038/nn.4272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang K., Daigle J.G., Cunningham K.M., Coyne A.N., Ruan K., Grima J.C., Bowen K.E., Wadhwa H., Yang P., Rigo F. Stress granule assembly disrupts nucleocytoplasmic transport. Cell. 2018;173:958–971.e17. doi: 10.1016/j.cell.2018.03.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Videos obtained from live GFP-EB3 imaging of control (non-demented control and MAPT P301-isogenic) and familial FTD-MAPT (MAPT IVS10+16-A, MAPT IVS10+16-B and MAPT P301L) iPSCs-derived neurons, (100 DIV). Timing is indicated on the top right.
Videos obtained from live GFP-EB3 imaging of familial FTD-MAPT (MAPT IVS10+16-A) iPSCs-derived neurons, (100 DIV) before and after nocodazole (33μM) for two hours. Scale bar = 10 μm.
Analysis recombinant microtubule tip protein EB3 growth speed, life-time, pause events and shrinking events in iPSC-derived neurons for each genotype in the study (MAPT IVS10+16-A/B, MAPT P301L, non-demented control and MAPT P301-isogenic control). Number of cells (n) analyzed per genotype are indicated in the table.