

Abstract
Background
Bacterial wilt caused by Ralstonia solanacearum is one of the most important diseases in pepper worldwide, however, the molecular mechanism underlying pepper resistance to bacterial wilt remains poorly understood.
Results
Herein, a novel RD leucine-rich repeat receptor-like kinase, CaLRR-RLK1, was functionally characterized in immunity against R. solanacearum. CaLRR-RLK1 was targeted exclusively to plasma membrane and was up-regulated by R. solanacearum inoculation (RSI) as well as by the exogenous application of salicylic acid (SA), methyl jasmonate (MeJA) or ethephon (ETH). The silencing of CaLRR-RLK1 led to enhanced susceptibility of pepper plants to RSI, accompanied by down-regulation of immunity-related genes including CaACO1, CaHIR1, CaPR4 and CaPO2. In contrast, transient overexpression of CaLRR-RLK1 triggered hypersensitive response (HR)-like cell death and H2O2 accumulation in pepper leaves, manifested by darker trypan blue and DAB staining respectively. In addition, the ectopic overexpression of CaLRR-RLK1 in tobacco plants enhanced resistance R. solanacearum, accompanied with the immunity associated marker genes including NtPR2, NtPR2, NtHSR203 and NtHSR515. Furthermore, it was found that CaHDZ27, a positive regulator in pepper response to RSI in our previous study, transcriptionally activated CaLRR-RLK1 by direct targeting its promoter probably in a CAATTATTG dependent manner.
Conclusion
The study revealed that CaLRR-RLK1 confers pepper resistance to R. solanacearum as the direct targeting of CaHDZ27.
Electronic supplementary material
The online version of this article (10.1186/s12870-018-1609-6) contains supplementary material, which is available to authorized users.
Keywords: CaLRR-RLK1, CaHDZ27, Pepper, Ralstonia solanacearum, Plant immunity
Background
Bacterial wilt, caused by Ralstonia solanacearum [1], is a highly destructive vascular disease in pepper (Capsicum annuum), a solanaceous vegetable of great agricultural importance. It is believed that the most efficient way to cope with such kind of disease is to develop crop variety with high level of disease resistance, a better understanding of molecular basis of pepper resistance to R. solanacearum will benefit its genetic improvement.
Upon challenge by pathogens, plant employ two interconnected layers of immunity including pathogen-associated molecular pattern (PAMP)-triggered immunity (PTI) and effector-triggered immunity (ETI) to protect them [2, 3]. Plant perceive exogenous PAMPs derived from microbial pathogens by cell surface-localized proteins, known as pattern recognition receptors (PRRs), which results in PTI. In addition, PRRs can also activate immune response by recognizing the endogenous damage-associated molecular patterns (DAMPs), which are produced as a consequence of pathogen infection and perception [4]. The adapted pathogen develop effectors, which are secreted and injected into host cells where they repress PTI by targeting specific signaling components, leading effectors triggered susceptivity (ETS) [5, 6]. PTI, also called basal defense, is required but not sufficient for disease resistance. Under the selection pressure of ETS, plant might evolve specific receptors, typically resistance (R) proteins, to detect the effectors and result in ETI, which is often accompanied with hypersensitive reaction (HR) and systematic acquired resistance (SAR). Although ETI is more robust, intensive and more directly associated with transcriptional regulation of defense gene expression than PTI, overlapping downstream molecular events have been found between PTI and ETI [7].
Leucine-rich repeat receptor-like protein kinases (LRR-RLKs) are typical plant cell surface-localized PRRs, they contain an extracellular domain with up to 30 leucine-rich repeat (LRRs) to perceive ligands including small molecules, peptides, and entire proteins [8, 9], a TMD (trans-membrane domain) for its cell membrane targeting and an intracellular serine/threonine (Ser/Thr) kinase domain to phosphorylate specific substrates for the signal transduction. Approximately 223 LRR-RLKs in Arabidopsis [10, 11], 226 in Zea mays [12], 234 in tomato [13], 379 in populous [14], 303 in Brassica rapa [15], 188 in pepper [16] have been identified. Some LRR-RLKs have been implicated to regulate plant innate immunity [17–19]. For example, a well-studied PRR LAGELLIN SENSITIVE 2 (FLS2) recognizes and binds a 22-amino acid epitope of bacterial flagellin called flg22 [20, 21]. Upon perception of flg22, FLS2 interacts with another LRR-RLK, BRI1-ASSOCIATED RECEPTOR KINASE 1 (BAK1) [22, 23], and phosphorylates the NADPH oxidase RbohD by association with a receptor-like cytoplasmic kinase (RLCK), BOTRYTIS-INDUCED KINASE 1 (BIK1) [24, 25], leading to a calcium burst and reactive oxygen species (ROS) production, and thereby mounts PAMP-triggered immunity (PTI). By sensing and binding Ax21 derived from pathogen, XA21 confers broad-spectrum resistance to the bacterial disease caused by the pathogen of Xanthomonas oryzae pv. oryzae (Xoo) [26, 27]. In addition, LRR-RLKs may participate in other biological processes such as plant growth, development and response to abiotic stresses [28]. For example, CLAVATA1 (CLV1) determines shoot and floral meristem size in Arabidopsis [29]. OsGIRL1-overexpressing transgenic Arabidopsis plants were hypersensitive in response to salt stress and heat stress, or hyposensitive in response to gamma-ray treatment and osmotic stress [30]. However, the majority members within LRR-RLKs family in different plant species remain functionally unidentified. In particular, so far, no LRR-RLK has been functionally characterized during pepper response to R. solanacearum.
A key step in both PTI and ETI is the massive transcriptional reprogramming dictated by various transcription factors. Through integrating the signal initiation and along the signaling pathways transmitting them into appropriate transcription of many defense associated genes including PRRs, R proteins and other signaling components [31–33], transcription factors play important roles in the regulation of plant immunity [34–38]. The HD-Zip transcription factors, characterized by two conserved domains, the DNA-binding homeodomain (HD) and an adjacent leucine zipper (LZ) motif, are unique to plants [39]. HD-Zip proteins are classified into four subfamilies (I–IV) based on their conserved HD-Zip domain, gene structure and additional conserved motifs [40]. HD-Zip I members were reported to play essential roles in response to environmental conditions [41, 42]. A HD-Zip I protein in pepper, CaHDZ27, was previously found to function as a positive regulator in plant immunity toward R. solanacearum by forming homodimers [42]. However, how CaHDZ27 linked with the upstream signaling components remains to be elucidated.
In the present study, a pepper putative LRR-RLK gene CaLRR-RLK1 which is up-regulated by the inoculation of R. solanacearum was functionally characterized in immunity, and its transcriptional regulation by CaHDZ27 was identified. The results showed that CaLRR-RLK1 act as a positive regulator in pepper response to RSI, the promoter of CaLRR-RLK1 is targeted by CaHDZ27 in a pseudopalindromic DNA (CAATTATTG) dependent manner, and it is up-regulated by CaHDZ27 during pepper response to RSI.
Results
Isolation and sequence analysis of CaLRR-RLK1
A putative LRR-RLK gene in pepper was originally identified as up-regulation in pepper plants challenged by R. solanacearum in our previous gene differential expression display by cDNA-AFLP (cDNA-amplified fragment length polymorphism). For cDNA-AFLP analysis, 64 primer pairs were used for selective PCR amplification and total 400 transcript derived fragments (TDFs) obtained. Among them, 114 displayed altered expression patterns after inoculation of R. solanacearum, of which 35 up-regulated and 79 down-regulated. The corresponding gene sequence including the promoter, 5’UTR, 3’UTR and the CDS were identified using pepper genome sequence (http://peppersequence.genomics.cn/page/species/index.jsp). The gene product was designated CaLRR-RLK1 (GenBank accession number: XP_016560889) as it represents the first report of a LRR-RLK from pepper. The full-length complementary DNA (cDNA) sequence of CaLRR-RLK1 was cloned by PCR with a specific pair of primers. By sequence analysis, the cDNA contains an open reading frame (ORF) of 2955 bps in length, its deduced amino acid sequence are 984 aa in length, containing a predicted extracellular region with a N-terminal signal peptide (SP, residues 1–25), a LRR_NT2 (a leucine-rich repeat N-terminal domain (residues 24–66), 21 extracellular leucine-rich repeats (LRRs, residues 96–596), a trans-membrane domain (TMD, residues 626–648) and a kinase domain (residues 684–946) (Fig. 1a).
Fig. 1.

The deduced amino acid sequence analysis of CaLRR-RLK1. a Schematic diagram of CaLRR-RLK1 protein domain architecture. b Comparison of the protein kinase subdomain of CaLRR-RLK1with that of serine/threonine protein kinases from Ipomoea nil (XP_019195246), Manihot esculenta (XP_021632827), Punica granatum (OWM66138), Sesamum indicum (XP_011073656), Helianthus annuus (XP_021998473), Solanum lycopersicum (XP_004231961), Solanum tuberosum (XP_006357742), Solanum pennellii (XP_015067035), Nicotiana sylvestris (XP_009804629), Nicotiana tabacum (XP_016449272), Nicotiana tomentosiformis (XP_009618480) and Coffea canephora (CDP02659)
The kinase domain shares high sequence homology with that of other RLK proteins (Fig. 1b). The conserved active site signature motif (VHRDVKSNNILLD residues) and the G-T/S-XX-Y/F-X-APE motif [43] indicate that CaLRR-RLK1 is a Ser/Thr kinase rather than a Tyr kinase. In addition, CaLRR-RLK1 appear to be a RD type protein kinase since it contains a conserved arginine-aspartic acid motif in its kinase subdomain [4].
Phylogenetic tree analysis was performed based on the amino acid sequence of CaLRR-RLK1 and its close homologs in other plant species. The result showed that CaLRR-RLK1 falls into a clade with Solanum tuberosum LRR-RLK (XP_006357742), Solanum lycopersicum LRR-RLK (XP_004231961) and Solanum pennellii LRR-RLK (XP_015067035) (Additional file 1: Figure S1).
Expression of CaLRR-RLK1 induced by R. solanacearum or exogenous hormones
To confirm the inducible expression of CaLRR-RLK1 in pepper plants against R. solanacearum inoculation, we detected its transcript levels in pepper leaves by qRT-PCR analysis. The result showed that the transcript levels of CaLRR-RLK1 were increased at 12 to 48 h post inoculation of R. solanacearum compared to the mock treatment (Fig. 2a).
Fig. 2.

The transcription of CaLRR-RLK1 in pepper leaves inoculated with R. solanacearum as well as exogenous application of methyl jasmonate (MeJA), ethephon (ETH) or salicylic acid (SA). a qRT–PCR analysis of transcript levels ofCaLRR-RLK1 in pepper leaves challenged with R. solanacearum inoculation compared to the mock treatment. b-d Relative transcript levels of CaLRR-RLK1 gene in leaves of pepper plants after treated with SA, MeJA or ETH. Values are the mean ± standard error (n = 6).Asterisks (P < 0.05) indicate significant differences as analyzed by the Student’s t-test
SA, JA and ET are signaling molecules that participate in the regulation of plant immunity [44–47]. To confirm the result that CaLRR-RLK1was induced by RSI, the transcript levels of CaLRR-RLK1 in pepper plants treated with exogenous applications of SA, MeJA or ETH were measured by qRT-PCR. The result showed that the transcript levels of CaLRR-RLK1 were up-regulated dramatically and peaked at 6 h post application (hpa) of exogenous application of MeJA (Fig. 2b), followed by a gradual decrease during 12–48 hpa. Upon application of ETH, the CaLRR-RLK1 transcript abundance was induced as early as 3 hpa (Fig. 2c). Similarly, the maximum transcripts occurred at 3 hpa of SA and then decreased to basal levels at 12 hpa (Fig. 2d). These results imply that CaLRR-RLK1 is involved in pepper defense against R. solanacearum regulated by a signaling mediated by SA, JA and ET.
Subcellular localization of CaLRR-RLK1
To study the subcellular localization of CaLRR-RLK1, we fused the full length ORF of CaLRR-RLK1 without the termination codon to 5′ terminal of the green fluorescent protein (GFP), resulting 35S:CaLRR-RLK1-GFP construct, using 35S:GFP and 35S:CBL1n-RFP (specifically targeting to the plasma membrane) [48] as controls. By agro-infiltration, CaLRR-RLK1-GFP and CBL1n-RFP or GFP and CBL1n-RFP were transiently co-overexpressed in Nicotiana benthamiana leaves. The results showed that green fluorescent signal of CaLRR-RLK1-GFP completely overlapped with the red fluorescent signal in the plasma membrane, while the control GFP was ubiquitously distributed throughout the cell (Fig. 3), indicating the localization of CaLRR-RLK1 in the plasma membrane.
Fig. 3.
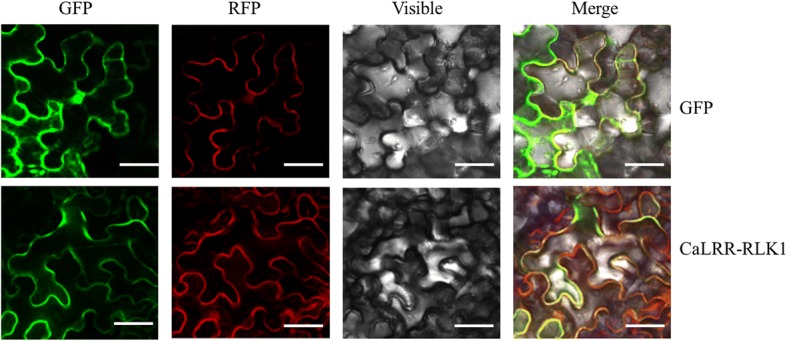
Subcellular localization of CaLRR-RLK1 by agro-infiltration in mesophyll cells of Nicotiana benthamiana leaves. Transient co-overexpression of CBL1n-RFP andCaLRR-RLK1-GFP or GFP in N. benthamiana leaves. Fluorescencewas detected using a confocal microscope. GFP, green fluorescent protein; RFP, red fluorescent protein. Bars = 50 μm
CaLRR-RLK1 silencing in pepper results in enhanced susceptibility to R. solanacearum
To further analyze the possible role of CaLRR-RLK1 in pepper response to RSI, virus-induced gene silencing (VIGS) approach was used to investigate the effect of silencing of CaLRR-RLK1 on pepper resistance to RSI. A fragment 182 bps in length within the 3’-UTR region of CaLRR-RLK1 was cloned into pTRV2 vector resulting in TRV:CaLRR-RLK1 (using TRV:00 as a control). The Agrobacterium GV3101 cells harboring TRV:CaLRR-RLK1 were infiltrated onto the cotyledon of the 12–14 days old pepper plants, and the TRV:CaPDS pepper plants were used to monitor the gene silencing process. When the seedlings were grown for about 25 days, plants infiltrated with TRV:CaPDS exhibited bleaching phenotype in their newly emerging leaves, the TRV:00 and TRV:CaLRR-RLK1 pepper plants were inoculated with R. solanacearum, and their transcript levels of CaLRR-RLK1 were measured by qRT-PCR using randomly selected 6 TRV:CaLRR-RLK1 pepper plants to detect the silencing efficiency of CaLRR-RLK1. The result showed that transcript levels of TRV:CaLRR-RLK1 in R. solanacearum inoculated or mock treated plants were about 20–30% of that in the control (TRV:00) plants (Fig. 4a), indicating the success of CaLRR-RLK1 silencing in pepper plants by VIGS.
Fig. 4.

Silencing of CaLRR-RLK1enhanced susceptibility of pepper plants to RSI. a The efficiency of CaLRR-RLK1 by VIGS was detected by qRT-PCR in TRV:CaLRR-RLK1 and TRV:00 plants challenged by R.solanacearum. b Disease symptoms of pepper plants at 10 dpi with R. solanacearum. c Disease index of R. solanacearum were evaluated daily for 14 days using a scale of 0 to 4. At least 50 plants were inoculated with R. solanacearum by root irrigation for TRV:CaLRR-RLK1 or TRV:00 plants. Results shown are the means of three independent experiments, each with 10 plants for TRV:CaLRR-RLK1 orTRV:00,bars indicate standard error of the mean. d Growth of R. solanacearum in stems of TRV:CaLRR-RLK1 and TRV:00 plants at 9 days post inoculation of R. solanacearum by root irrigation. e Growth of R. solanacearum in leaves of TRV:CaLRR-RLK1 and TRV:00 plants at 3 days post leaf-inoculation of R. solanacearum. f The relative transcript levels of the defense related genes in leaves of TRV:CaLRR-RLK1andTRV:00 plants. In a, d, e and f, values were the mean ± standard error (n = 6). Asterisks (P < 0.05) indicate significant differences as analyzed by the Student’s t-test
With the CaLRR-RLK1 silencing pepper plants, which were confirmed by qRT-PCR individually, the effect of CaLRR-RLK1 silencing on pepper immunity against RSI was assayed. All of CaLRR-RLK1 silenced pepper plants did not exhibit any morphological changes compared to the control plants. Inoculated with R. solanacearum by root irrigation, TRV:CaLRR-RLK1 plants exhibited significantly enhanced susceptibility to RSI compared to the control plants (Fig. 4b). After 6 days post inoculation (dpi), TRV:CaLRR-RLK1 plants displayed a higher disease index than the control. TRV:CaLRR-RLK1 plants began to display the bacterial wilt symptom at 6 dpi and complete wilt phenotype at 12 dpi, while the control plants did not begin to display the bacterial wilt symptom until 7 dpi and complete wilt until 14 dpi (Fig. 4c). Consistently, a higher population density of R. solanacearum was detected in stems of TRV:CaLRR-RLK1 plants at 9 dpi by root irrigation (Fig. 4d) and also in R. solanacearum inoculated leaves at 3 dpi (Fig. 4e), compared to that in the control plants.
To further investigate whether silencing of CaLRR-RLK1 affect the expression of immunity-related genes in pepper, qRT–PCR analysis was performed. The results showed that expressions of HR related CaHIR1 [49], ROS detoxification-associated CaPO2 [50], ET-responsive ACC oxidase gene CaACO1 [51], JA/ET signaling associated CaPR4 [52] were significantly down-regulated in CaLRR-RLK1 silenced leaves, compared with that in the control plants challenged with RSI. However, there was no significant difference for the transcript levels of SA signaling associated CaPR1 [53] and CaNPR1 [54] (Fig. 4f). These results indicate that silencing of CaLRR-RLK1 increased susceptibility of pepper plants to RSI.
Transit expression of CaLRR-RLK1 triggered HR cell death and induced the transcription of immunity associated genes
To confirm the results in the gene silencing by VIGS, CaLRR-RLK1 was transiently overexpressed via agro-infiltration. GV3101 cells containing 35S:HA-CaLRR-RLK1 or 35S:HA were infiltrated into pepper leaves, the success of the transient overexpression was confirmed by qRT-PCR (Fig. 5a) or immunoblot with antibodies of HA (Additional file 2: Figure S2). Darker staining of trypan blue and DAB were detected in CaLRR-RLK1 transiently overexpressing pepper leaves around the infiltration sites, while no such darker staining was detected in the control plant leaves (35S:HA), indicating that HR cell death and H2O2 accumulation were triggered by transient overexpression of CaLRR-RLK1 (Fig. 5b). Consistently, a significant enhanced electrolyte leakage was also detected in CaLRR-RLK1 transiently overexpressing pepper leaves at 48 and 72 h post infiltration (hpi) compared with that of the control (Fig. 5c).
Fig. 5.

Transient overexpression of CaLRR-RLK1triggered HR mimic cell death in pepper leaves. a The transcript levels ofCaLRR-RLK1 in pepper leaves detected by qRT-PCR. b Trypan blue and DAB staining in leaves transiently expressing CaLRR-RLK1 at 48 h post infiltration. c Electrolyte leakage from leaf discs of CaLRR-RLK1 transiently overexpressed pepper leaves. d Transcript levels of immunity associated genes were detected in CaLRR-RLK1 transient overexpression pepper leaves by qRT-PCR. In a, c and d, values were the mean ± standard error (n = 6). Asterisks (P < 0.05) indicate significant differences as analyzed by the Student’s t-test
Quantitative RT-PCR analysis was used to investigate whether transient expression of CaLRR-RLK1 affect defense associated genes including CaACO1, CaHIR1, CaPR4, CaPO2, CaNPR1 and CaPR1 in pepper leaves. The results showed that CaACO1, CaHIR1, CaPR4 and CaPO2 exhibited enhanced transcript levels in CaLRR-RLK1 transiently overexpressing pepper leaves at 48 hpi compared to that in the control, transcript levels of CaNPR1 and CaPR1 did not alter (Fig. 5d).
Ectopic overexpression of CaLRR-RLK1 enhanced resistance of tobacco plants to RSI
To further confirm that result that CaLRR-RLK1 act as positive regulator in plant immunity, we investigated the effect of its ectopic overexpression on the resistance of tobacco plants to RSI by developing the transgenic tobacco lines via Agrobacterium mediated method. Six transgenic lines (T3) each with at least 10 seedlings were confirmed by kanamycin resistance analysis. Two lines were employed to assay the transcriptional expression of CaLRR-RLK1, the seedlings were inoculated with R. solanacearum cells. RT-PCR result showed that CaLRR-RLK1 constitutively expressed in leaves of the two lines (#2 and #8) (Additional file 3: Figure S3).
Eight-week-old tobacco plants were inoculated with R. solanacearum by root irrigation, the development of bacterial wilt symptoms was monitored. At 7 dpi, wilting and contagion symptoms occurred on the stems of the wild-type plants, while no significant symptom was observed in plants of CaLRR-RLK1 overexpressing lines (Fig. 6a). At 21 dpi, wild-type plants exhibited extremely severe wilting symptoms, but only slight wilt symptoms were observed in plants of CaLRR-RLK1 overexpressing lines (Fig. 6b). At 28 dpi, a significant higher death rate (76.1%) was found in the wild-type plants than that in plants of the two lines, which was 36.5 and 51.1%, respectively (Fig. 6c).
Fig. 6.

Ectopic overexpression of CaLRR-RLK1enhanced resistance of tobacco plants to RSI. a Comparison of the symptom on stems between wild-type and CaLRR-RLK1 overexpressing lines at 7 dpi with R. solanacearum. b Comparison of wilt symptoms in seedlings of CaLRR-RLK1 overexpressing lines and the wild-type at 21 dpi. c Percentage of dead plants at 28 dpi with R. solanacearum by root irrigation. Data were from at least 10 plants for each line. Three independent experiments were performed. Bars represent standard error of the mean. d Relative transcript levels of immunity associated genes in plants of CaLRR-RLK1 overexpressing lines and wild-type at 48 h post inoculated with R. solanacearum. Values were the mean ± standard error (n = 3). Asterisks (P < 0.05) indicate significant differences as analyzed by the Student’s t-test
In addition, the transcript levels of immunity associated genes, including SA-induced pathogenesis-related NtPR2 [55] and NtPR3 [56], HR-related NtHSR201 and NtHSR515 [57], were measured by qRT-PCR in tobacco plants of CaLRR-RLK1 overexpressing lines and the wild type, the results showed that the transcript levels of NtPR2, NtPR3, NtHSR201 and NtHSR515 increased in transgenic plants compared to that in wild-type plants in response to R. solanacearum infection (Fig. 6d).
The promoter of CaLRR-RLK1 was directly bound and transcriptionally modulated by CaHDZ27
Silencing of CaLRR-RLK1 impairs resistance of pepper to RSI, while its overexpression enhances the resistance of pepper or tobacco to RSI. Therefore, its inducible expression under challenge by RSI is crucial for its role as positive regulator. To study the molecular mechanism underlying this inducible expression, the cis-elements in the promoter region of CaLRR-RLK1 of 1574 bps in length was analyzed (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/). Noticeably, a 9 bp pseudopalindromic DNA sequence (CAATTATTG), which is potentially bound by HD-Zip transcription factors [42, 58], was found to be present in the promoter (Fig. 7a). Since our previous study found that CaHDZ27, a HD-Zip I transcription factor, acts a positive regulator in pepper response to RSI, we speculated that CaHDZ27 might act as an upstream regulator of CaLRR-RLK1 by binding the motif CAATTATTG in its promoter. To test if it is the case, we employed chromatin immunoprecipitation (ChIP) assay via HA-CaHDZ27 transient overexpression in pepper leaves, the chromatins were isolated from HA-CaHDZ27 transiently overexpressing pepper leaves after crosslinking with 1% formaldehyde, sheared into fragments of 300-500 bp in length, the DNA fragments bound to CaHDZ27 were immunoprecipitated by antibodies of HA, the de-crosslinked and purified DNA fragments were used as template for qRT- PCR with specific primer pairs based on the sequence flanking the motif (CAATTATTG) in the promoter of CaLRR-RLK1, the result showed that the promoter of CaLRR-RLK1 was significantly enriched by CaHDZ27, and in response to the challenge of R. solanacearum, CaHDZ27 binding enrichment was markedly increased (Fig. 7b), indicating that CaHDZ27 binds to the promoter of CaLRR-RLK1 probably via the motif CAATTATTG.
Fig. 7.

CaHDZ27 can bind the motif CAATTATTG in the promoter ofCaLRR-RLK1 andactivateCaLRR-RLK1 transcription. a Scheme of the CaLRR-RLK1 promoter region, primers used in ChIP-qPCR assay were marked by arrows. b ChIP-qPCR assays were performed using the specific primers corresponding to the region containing the motif (CAATTATTG). c The transcript levels of CaLRR-RLK1 in HA-CaHDZ27-SRDX or HA-CaHDZ27 transiently overexpressed pepper leaves. d The transcript levels of CaLRR-RLK1 in CaHDZ27-silenced leaves challenged by R. solanacearum. e GUS activities driven by the promoter of CaLRR-RLK1 were assayed when transiently overexpressing HA-CaHDZ27-SRDX or HA-CaHDZ27 in pepper leaves. f Activities of GUS driven by the pCaLRR-RLK1 or pCaLRR-RLK1mut,in which CAATAATTG was replaced by CAGGGGTTG, when co-transiently overexpressed withHA-CaHDZ27 in pepper leaves. In b, c and e, the values were the mean ± standard error (n = 3), different letters indicate significant differences, as determined by LSD test (P < 0.05). In d and f, asterisks indicate significant difference according to Student’s t test at P < 0.05
To assay if CaLRR-RLK1 is transcriptionally modulated by CaHDZ27, we carried out transient overexpression of HA-CaHDZ27 in pepper leaves by infiltration with GV3101 cells containing 35:HA-CaHDZ27 or 35S:HA, 2 days after agro-infiltration, the transcript levels of CaLRR-RLK1 were checked by qRT-PCR. The result showed that CaLRR-RLK1 was up-regulated in leaves infiltrated with 35S:HA-CaHDZ27, and not down-regulated in leaves infiltrated with 35S:HA-CaHDZ27-SRDX (a repressor version of HA-CaHDZ27), compared with 35S:HA (Fig. 7c). Moreover, we silenced CaHDZ27 in pepper plants by VIGS and checked the transcript level of CaLRR-RLK1 in CaHDZ27-silenced plants by qRT-PCR, the result showed that the transcript level of CaLRR-RLK1 decreased in TRV:CaHDZ27 pepper leaves upon R. solanacearum inoculation compared to that in the control (TRV:00) plants (Fig. 7d). To provide additional evidence that the transcript of CaLRR-RLK1 was activated by CaHDZ27, the promoter of CaLRR-RLK1 was cloned and inserted into pMDC163 [59] to get construct pCaLRR-RLK1:GUS. The GV3101 cells containing pCaLRR-RLK1:GUS were mixed with 35S:HA-CaHDZ27, 35S:HA-CaHDZ27-SRDX or 35S:HA at 1:1 ratio, and infiltrated into the pepper leaves. GUS activity was measured at 2 dpi, the result showed that GUS activities were enhanced by HA-CaHDZ27 transient overexpression, while not decreased by HA-CaHDZ27-SRDX compared to that in the control leaves (Fig. 7e), consistently with the qRT-PCR results. These results indicated that CaLRR-RLK1 is transcriptionally activated by CaHDZ27.
To further determine whether the transcriptional activation of CaLRR-RLK1 by CaHDZ27 is via the motif CAATTATTG within the promoter of CaHDZ27, we mutated the cis-acting element (CAGGGGTTG) in the promoter region of CaLRR-RLK1, and resulted in construct pCaLRR-RLK1mut:GUS. GV3101 cells containing pCaLRR-RLK1:GUS or pCaLRR-RLK1mut:GUS were mixed with 35S:HA-CaHDZ27 and were co-infiltrated into the leaves of N. benthamiana. 48 or 72 h after infiltration, the GUS activity was measured. The result showed that GUS activity was much higher in the leaves co-expressing pCaLRR-RLK1:GUS and 35S:HA-CaHDZ27 than that in pCaLRR-RLK1mut:GUS and 35S:HA-CaHDZ27 co-expressed pepper leaves (Fig. 7f). Taken together, these data support the idea that CaHDZ27 activates the transcription of CaLRR-RLK1 by direct binding to the CAATTATTG motif.
Discussion
CaLRR-RLK1 plays a positive role in pepper immunity
Bacterial wilt incited by R. solanacearum causes heavy losses in pepper, a vegetable of great agricultural importance worldwide. In the present study, our data indicate that CaLRR-RLK1 act as a positive regulator in pepper immunity toward R. solanacearum and it is transcriptionally modulated directly by CaHDZ27.
Based on sequence motifs in the extracellular domain, typical RLKs can be divided into more than 21 subfamilies [11]. So far, three RLKs including CaRLK1 [60–62], CaPIK11 [63–66] and CaLecRK-S.5 [67] in pepper have been functionally characterized. Like CaLRR-RLK1 in the present study, CaRLK1 contains cytoplasmic kinase domain, a transmembrane span, and an extracellular domain which shows low homology to other known plant RLKs. It functions in cell death [62], hypoxia resistance [61] and cell division [60]. CaPIK1 functions as a positive regulator of defense responses and cell death in an SA-dependent manner [65, 66]. L-type lectin receptor kinase CaLecRK-S.5 positively mediated broad-spectrum resistance in pepper [67]. However, no LRR-RLK in pepper has been functionally characterized in plant immunity to date.
CaLRR-RLK1 contains a predicted extracellular region with an N-terminal signal peptide, 21 extracellular leucine-rich repeats, a trans-membrane domain and an intracellular kinase domain. CaLRR-RLK1 appears to be RD Ser/Thr kinase since it contains a conserved arginine-aspartic acid motif in its kinase subdomain. Consistent with the trans-membrane domain, CaLRR-RLK1 exclusively localizes to the plasma membrane. The direct evidence that CaLRR-RLK1 act as a positive regulator came from data that CaLRR-RLK1 silencing by VIGS significantly impaired resistance of pepper plant to RSI and down-regulated the tested immunity associated maker genes. By contrast, the transient overexpression of CaLRR-RLK1 significantly enhanced the HR cell death which is frequently found to occur in ETI and occasionally in PTI [68], the accumulation of H2O2 which is manifested by DAB staining and closely related to HR cell death [69], and the induction of the tested immunity associated marker genes. These data from pepper plants indicate a role of CaLRR-RLK1 as a positive regulator in pepper immunity against RSI, and it was further supported by the data from ectopic expression of CaLRR-RLK1 in tobacco plants. The overexpression of CaLRR-RLK1 significantly enhanced the resistance of tobacco plants to RSI, accompanied with significant up-regulations of the tested immunity associated marker genes including NtPR2 [55], NtPR3 [56], NtHSR201 and NtHSR515 [57].
RLKs were categorized into RD kinases and non-RD kinases according to the RD motif in their kinase domains [70]. By far, well-studied LRR-RLK PRRs, such as FLS2 or Xa21 [71, 72], that recognize the exogenous PAMPs in disease resistance were non-RD kinases [70]. Characterized plant DAMP receptors all contained RD kinase domains [4]. For example, PEPR1/2 and RLK7 have been implicated in activation of PTI against pathogens through perception of peptides AtPEP1 [73, 74] and PIP1 [75] in Arabidopsis, respectively. As a RD kinase domain containing RLK, we speculate that CaLRR-RLK1 might act as a receptor of unidentified DAMPs in pepper defense response to R. solanacearum.
The expression of CaLRR-RLK1 was activated by CaHDZ27 direct binding the CAATTATTG motif in its promoter
Our data indicate an inducible expression of CaLRR-RLK1 upon challenge of R. solanacearum, this enhanced expression might be crucial for profound amplification of the defense signaling to avoid the negative effect of the constitutive immune reaction. Some PRRs such as FLS2 [76] and R genes such as Xa1 [77] were reported to exhibit inducible expression upon pathogen attack. To dissect the mechanism underlying up-regulation of CaLRR-RLK1 upon RSI, its upstream transcription factor is identified. Our data showed that CaHDZ27, a HD-Zip protein that acts as positive regulator in pepper resistance against R. solanacearum [42], directly targets the promoter of CaLRR-RLK1, and activate the expression of CaLRR-RLK1 in a motif CAATAATTG dependent manner. Similar to CaHDZ27 [42], the transcription of CaLRR-RLK1 was also up-regulated by exogenously applied SA, MeJA and ETH, indicating that CaHDZ27 and CaLRR-RLK1 act as a positive regulators in plant immunity during pepper response to RSI. CaLRR-RLK1 contains 21 LRRs in the extracellular domain. Although the ligand of LRRs is not known, CaLRR-RLK1 may play a role in the pepper defense signaling pathway through these LRR domains binding with ligand, and the enhanced expression caused by the transcript factor CaHDZ27 is beneficial to pepper resistance against R. solanacearum.
Conclusions
Our data in the present study indicate that expression of CaLRR-RLK1 was induced by R. solanacearum inoculation or exogenous application of SA, MeJA or ETH. Inhibition of its expression in pepper disrupted the defense ability of plants against R. solanacearum infection. Transit expression of this gene in pepper leaves can lead to cell death, and overexpression of this gene rendered the transgenic tobacco plants with increased disease tolerance. Furthermore, the expression of CaLRR-RLK1 is activated by CaHDZ27 in a pseudopalindromic DNA (CAATTATTG) dependent manner. The experimental evidence collectively demonstrates that CaLRR-RLK1 act as a positive regulator in pepper response to R. solanacearum, and it is transcriptionally activated by CaHDZ27.
Methods
Plant materials and conditions of growth
Pepper (Capsicum annuum cultivar L. cv. Fj8), with medium-resistance to R. solanacearum, was obtained from the pepper breeding group at Fujian Agriculture and Forestry University. Tobacco (Nicotiana tabacum L. cv. Honghuadajinyuan) was kindly supplied by the tobacco breeding group in Fujian Agriculture and Forestry University. Plants were grown under the conditions of 26 °C temperature, 60% relative humidity, 60–70 μmol photons m− 2 s− 1 and 16 h light/8 h dark.
Pathogen inoculation
Virulent strains FJC100301 and FJ1003 strain of R. solanacearum were from pepper and tobacco, respectively. The pathogen strains were amplified according to the method described previously [78]. The R. solanacearum strain was grown in PSA medium (200 g/L of potato, 20 g/L of sucrose, 3 g/L of beef extract, 5 g/L of tryptone). Bacteria were harvested with sterile ddH2O and re-suspended to 1.0 × 108 CFU/ mL. Seedlings were root inoculated by pouring either 30 mL bacterial inoculum or sterile ddH2O as the mock into each pot. Before inoculation, the roots were damaged by making holes in the soil of each pot. For leaf inoculation, the third leaf from the top of each pepper plant at the eight-leaf stage were infiltrated with 10 μL of the R. solanacearum suspension using a syringe without a needle, and the mock treatment was performed with sterile ddH2O.
Exogenous hormones treatments
Six leaf-stage pepper plants were used to investigate gene expression. For salicylic acid (SA), methyl jasmonate (MeJA) and ethephon (ETH) treatments, 1 mM SA, 100 μM MeJA or 100 μM ETH were sprayed onto pepper leaves. In parallel experiments, corresponding solvent or sterile water was sprayed as a control. Pepper leaves were sampled at various time points, frozen and stored at − 80 °C for RNA isolation.
Subcellular localization
The open reading frame (ORF) of CaLRR-RLK1 without the stop codon was PCR amplified from plant cDNA and cloned into pMDC83 to yield 35S:CaLRR-RLK1-GFP. Agrobacterium tumefaciens strain GV3101 containing the constructs 35S:CaLRR-RLK1-GFP or 35S:GFP (used as a control) was re-suspended (OD600 = 0.8) in the induction medium (10 mM MES, 10 mM MgCl2, 200 μM acetosyringone, pH 5.6) and infiltrated into leaves of Nicotiana benthamiana. After 2 days post infiltration, GFP fluorescence was imaged using a Laser Scanning Confocal Microscope (TCS SP8, Leica, Solms, Germany) with an excitation wavelength of 488 nm and a 505–530 nm band-pass emission filter.
VIGS in pepper plants
TRV (tobacco rattle virus)-based VIGS analysis was performed as described previously [42]. A 182-bp gene specific 3′ untranslated region of CaLRR-RLK1 was inserted into the TRV2 vector to yield pTRV2:CaLRR-RLK1 by gateway cloning technique. TRV1 and TRV2:00 (control) or pTRV2:CaLRR-RLK1 in A. tumefaciens strain GV3101 were co-infiltrated into the fully expanded cotyledons of approximately 12–14 days old pepper plants. Then, the seedlings were incubated at 16 °C for 56 h, and grown at 26 °C.
Agrobacterium-mediated transient expression in pepper leaves
The full length cDNA of CaLRR-RLK1 or CaHDZ27 was cloned into pEarleyGate201 to yield 35S:HA-CaLRR-RLK1 construct or 35S:HA-CaHDZ27 by gateway cloning technique. The CaHDZ27 dominant repression construct (35S:HA-CaHDZ27-SRDX) was created by fusing the CaHDZ27 cDNA in frame with the dominant EAR repression sequence [79], which was ligated downstream of the 35S promoter into the vector pEarleyGate201.
A. tumefaciens strain GV3101 harboring the above vector was infiltrated into the pepper leaves at the eight-leaf stage using a syringe without a needle. After 48 or 72 h post infiltration, injected leaves were harvested for further use.
Histochemical staining and ion conductivity measurement
Diaminobenzidine (DAB) and trypan blue staining, as well as ion conductivity measurement assays were performed according to methods described previously [51, 78]. Bacteria-infiltrated leaves were harvested and incubated in 1 mg mL− 1 DAB in the dark overnight and destained with lacticacid/glycerol/ethanol (1:1:3). To visualize the cell death, the infiltrated leaves were stained with trypan blue and destained with chloral hydrate solution. Cell death was quantified by ion conductivity measurement, eight discs (11 mm in diameter) were excised and washed in sterile ddH2O. After incubation in 20 mL of double distilled water with gentle shaking for 3 h, ion conductivity was measured using a Mettler Toledo 326 (Mettler, Zurich, Switzerland).
Quantitative real-time RT-PCR
For quantitative real-time PCR analysis, total RNA from plant leaves was extracted using a TRIzol Reagent (Invitrogen, Carlsbad, CA, USA). First-strand cDNA was synthesized using One Step PrimeScript TM cDNA Synthesis Kit (TaKaRa, Shigo, Japan). Real-time PCR experiments were performed according to the manufacturer’s instructions for the BIO-RAD Real-time PCR system (Foster City, CA, USA) and the SYBR Premix Ex Taq II system (TaKaRa). To normalize the transcript levels, Pepper CaActin (GQ339766) or 18S ribosomal RNA (EF564281) and Tobacco NtEF1α (D63396) or NtActin (U60489) expression was monitored as reference genes in each reaction. The gene-specific primers used for the quantitative real-time RT–PCR analysis are listed in Additional file 4: Table S1 and Table S2. Each gene expression was calculated from three experimental replicates to ensure reproducibility of the results.
ChIP-qPCR analysis
ChIP-qPCR assays were performed essentially as described previously [51]. Briefly, the chromatins were isolated from the pepper leaves infiltrated with 35S:HA-CaHDZ27. The isolated chromatins were cross-linked with 1% formaldehyde, sheared, and precipitated using anti-HA. Enrichment of DNA samples was analyzed by quantitative real-time PCR. Each ChIP value was normalized to its respective input DNA value.
β-Glucuronidase (GUS) enzymatic assay
The CaLRR-RLK1 promoter or its mutation was cloned into pMDC163 to yield pCaLRR-RLK1:GUS or pCaLRR-RLK1mut:GUS construct. The frozen plant material collected from leaves of transiently transformed pepper, was ground with a pestle in the extraction buffer (50 mM phosphate buffer, pH 7.0, 10 mM EDTA, 0.1% Triton X-100, 0.1% sodium lauryl sarcosine, and 10 mM β-mercaptoethanol) and centrifuged at 11000 g for 10 min at 4 °C. Soluble crude protein in the supernatant was quantified using the Bradford method [80]. Quantitative analysis of GUS activity was measured with 4-methylumbelliferyl-D-glucuronide (4-MUG) as the substrate [54].
Plant transformation
The full length cDNA of CaLRR-RLK1 was cloned into plant binary vector pK7WG2 to yield 35S:CaLRR-RLK1 construct. Agrobacterium-mediated transformation of tobacco (Nicotiana tabacum L. cv. Honghuadajinyuan) was performed. Transformants were selected on Murashige and Skoog (MS) agar plates containing 50 μg mL− 1 kanamycin. Successful transformation was confirmed by RT-PCR using the CaLRR-RLK1 gene-specific primers. Among the selected transgenic plants, T2 lines #2 and #8 were used in this study.
Additional files
Figure S1. The phylogenetic relationship of CaLRR-RLK1 protein with other plant LRR-RLKs. An unrooted neighbor-joining tree was built by MEGA 4.0. (TIF 30 kb)
Figure S2. Transient overexpression of HA-CaLRR-RLK1 in pepper leaves detected by immunoblot. (TIF 36 kb)
Figure S3. The expression of CaLRR-RLK1 in representative T3 transgenic tobacco plants was checked by RT-PCR. Wild-type (WT) tobacco plants as the blank control, and NtEF1α served as an endogenous control. (TIF 21 kb)
Table S1. Pepper primers used for qPCR in this study. Table S2. Tobacco primers used for qPCR in this study. Table S3. Primers used in these experiments. (DOCX 21 kb)
Acknowledgements
We thank Mark D. Curtis for kindly providing the Gateway destination vectors and Dr. S. P. Dinesh-Kumar (Yale University) for the pTRV1 and pTRV2 vectors.
Funding
This work was supported by grants from the National Natural Science Foundation of China (31301254, 31372061, 31601761, 31401890, 31401312 and 31260482) and Natural Science Foundation of Fujian Province (2018 J01616). The funders were not involved in the design of the study, the data collection and analysis, and writing the manuscript.
Availability of data and materials
All data generated or analysed during this study are included in this published article (and its Additional file 4).
Abbreviations
- ChIP
Chromatin immunoprecipitation
- ETI
Effector-triggered immunity
- ETS
Effectors triggered susceptivity
- GFP
Green fluorescent protein
- hpa
H post application
- LRR-RLK
Leucine-rich repeat receptor-like kinase
- non-RD
Non-arginine-aspartate
- ORF
Open reading frame
- PAMP
Pathogen-associated molecular pattern
- PTI
PAMP-triggered immunity
- RSI
Ralstonia solanacearum inoculation
- SAR
Systematic acquired resistance
- TMD
Trans-membrane domain
- VIGS
Virus-induced gene silencing
Authors’ contributions
SM and SH conceived and designed the research. SM, FG, LS, SY and WH performed the experiment. WC performed data analysis. YW contributed to the acquisition of transgenic tobacco lines. SH and YW made key comments on the design of the trail, the article writing and the revisions. SM and SH wrote the article. All authors read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Shaoliang Mou, Email: moushaoliang@163.com.
Feng Gao, Email: 2423544334@qq.com.
Lei Shen, Email: shorttubelycoris07@163.com.
Sheng Yang, Email: 419239918@qq.com.
Weihong He, Email: 1574480429@qq.com.
Wei Cheng, Email: chengwei8503@163.com.
Yang Wu, Email: 40770249@qq.com.
Shuilin He, Email: shlhe201304@aliyun.com.
References
- 1.Du HS, Chen B, Zhang XF, Zhang FL, Miller SA, Rajashekara G, Xu XL, Geng SS. Evaluation of Ralstonia solanacearum infection dynamics in resistant and susceptible pepper lines using bioluminescence imaging. Plant Dis. 2017;101(2):272–278. doi: 10.1094/PDIS-05-16-0714-RE. [DOI] [PubMed] [Google Scholar]
- 2.Hein I, Gilroy EM, Armstrong MR, Birch PR. The zig-zag-zig in oomycete-plant interactions. Mol Plant Pathol. 2009;10(4):547–562. doi: 10.1111/j.1364-3703.2009.00547.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Jones JD, Dangl JL. The plant immune system. Nature. 2006;444(7117):323–329. doi: 10.1038/nature05286. [DOI] [PubMed] [Google Scholar]
- 4.Dardick C, Schwessinger B, Ronald P. Non-arginine-aspartate (non-RD) kinases are associated with innate immune receptors that recognize conserved microbial signatures. Curr Opin Plant Biol. 2012;15(4):358–366. doi: 10.1016/j.pbi.2012.05.002. [DOI] [PubMed] [Google Scholar]
- 5.Liu ZH, Faris JD, Oliver RP, Tan KC, Solomon PS, McDonald MC, McDonald BA, Nunez A, Lu SW, Rasmussen JB, et al. SnTox3 Acts in Effector Triggered Susceptibility to Induce Disease on Wheat Carrying the Snn3 Gene. PLoS Pathog. 2009;5(9). [DOI] [PMC free article] [PubMed]
- 6.Deller S, Hammond-Kosack KE, Rudd JJ. The complex interactions between host immunity and non-biotrophic fungal pathogens of wheat leaves. J Plant Physiol. 2011;168(1):63–71. doi: 10.1016/j.jplph.2010.05.024. [DOI] [PubMed] [Google Scholar]
- 7.Peng Y, van Wersch R, Zhang Y. Convergent and divergent signaling in PAMP-triggered immunity and effector-triggered immunity. Mol Plant-Microbe Interact. 2018;31(4):403–409. doi: 10.1094/MPMI-06-17-0145-CR. [DOI] [PubMed] [Google Scholar]
- 8.Fischer I, Dievart A, Droc G, Dufayard JF, Chantret N. Evolutionary dynamics of the Leucine-rich repeat receptor-like kinase (LRR-RLK) subfamily in angiosperms. Plant Physiol. 2016;170(3):1595–1610. doi: 10.1104/pp.15.01470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Shiu SH, Bleecker AB. Plant receptor-like kinase gene family: diversity, function, and signaling. Sci STKE. 2001(113):re22. [DOI] [PubMed]
- 10.Gou X, He K, Yang H, Yuan T, Lin H, Clouse SD, Li J. Genome-wide cloning and sequence analysis of leucine-rich repeat receptor-like protein kinase genes in Arabidopsis thaliana. BMC Genomics. 2010;11:19. doi: 10.1186/1471-2164-11-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wu Y, Xun Q, Guo Y, Zhang J, Cheng K, Shi T, He K, Hou S, Gou X, Li J. Genome-wide expression pattern analyses of the Arabidopsis Leucine-rich repeat receptor-like kinases. Mol Plant. 2016;9(2):289–300. doi: 10.1016/j.molp.2015.12.011. [DOI] [PubMed] [Google Scholar]
- 12.Song W, Wang B, Li X, Wei J, Chen L, Zhang D, Zhang W, Li R. Identification of Immune Related LRR-Containing Genes in Maize (Zea mays L.) by Genome-Wide Sequence Analysis. Int J Genomics. 2015;2015:231358. doi: 10.1155/2015/231358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wei Z, Wang J, Yang S, Song Y. Identification and expression analysis of the LRR-RLK gene family in tomato (Solanum lycopersicum) Heinz 1706. Genome. 2015;58(4):121–134. doi: 10.1139/gen-2015-0035. [DOI] [PubMed] [Google Scholar]
- 14.Zan Y, Ji Y, Zhang Y, Yang S, Song Y, Wang J. Genome-wide identification, characterization and expression analysis of populus leucine-rich repeat receptor-like protein kinase genes. BMC Genomics. 2013;14:318. doi: 10.1186/1471-2164-14-318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rameneni JJ, Lee Y, Dhandapani V, Yu X, Choi SR, Oh MH, Lim YP. Genomic and post-translational modification analysis of Leucine-rich-repeat receptor-like kinases in Brassica rapa. PLoS One. 2015;10(11):e0142255. doi: 10.1371/journal.pone.0142255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kim S, Park M, Yeom SI, Kim YM, Lee JM, Lee HA, Seo E, Choi J, Cheong K, Kim KT, et al. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat Genet. 2014;46(3):270–278. doi: 10.1038/ng.2877. [DOI] [PubMed] [Google Scholar]
- 17.Hu H, Wang J, Shi C, Yuan C, Peng C, Yin J, Li W, He M, Ma B, Wang Y, et al. A receptor like kinase gene with expressional responsiveness on Xanthomonas oryzae pv. oryzae is essential for Xa21-mediated disease resistance. Rice (N Y) 2015;8(1):34. doi: 10.1186/s12284-015-0069-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Greeff C, Roux M, Mundy J, Petersen M. Receptor-like kinase complexes in plant innate immunity. Front Plant Sci. 2012;3:209. doi: 10.3389/fpls.2012.00209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Chaparro-Garcia A, Wilkinson RC, Gimenez-Ibanez S, Findlay K, Coffey MD, Zipfel C, Rathjen JP, Kamoun S, Schornack S. The receptor-like kinase SERK3/BAK1 is required for basal resistance against the late blight pathogen phytophthora infestans in Nicotiana benthamiana. PLoS One. 2011;6(1):e16608. doi: 10.1371/journal.pone.0016608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Gomez-Gomez L, Boller T. FLS2: an LRR receptor-like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis. Mol Cell. 2000;5(6):1003–1011. doi: 10.1016/S1097-2765(00)80265-8. [DOI] [PubMed] [Google Scholar]
- 21.Salomon S, Robatzek S. Induced endocytosis of the receptor kinase FLS2. Plant Signal Behav. 2006;1(6):293–295. doi: 10.4161/psb.1.6.3594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Heese A, Hann DR, Gimenez-Ibanez S, Jones AME, He K, Li J, Schroeder JI, Peck SC, Rathjen JP. The receptor-like kinase SERK3/BAK1 is a central regulator of innate immunity in plants. P Natl Acad Sci USA. 2007;104(29):12217–12222. doi: 10.1073/pnas.0705306104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Roux M, Schwessinger B, Albrecht C, Chinchilla D, Jones A, Holton N, Malinovsky FG, Tor M, de Vries S, Zipfel C. The Arabidopsis Leucine-rich repeat receptor-like kinases BAK1/SERK3 and BKK1/SERK4 are required for innate immunity to Hemibiotrophic and biotrophic pathogens. Plant Cell. 2011;23(6):2440–2455. doi: 10.1105/tpc.111.084301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Li L, Li M, Yu L, Zhou Z, Liang X, Liu Z, Cai G, Gao L, Zhang X, Wang Y, et al. The FLS2-associated kinase BIK1 directly phosphorylates the NADPH oxidase RbohD to control plant immunity. Cell Host Microbe. 2014;15(3):329–338. doi: 10.1016/j.chom.2014.02.009. [DOI] [PubMed] [Google Scholar]
- 25.Zhang M, Chiang YH, Toruno TY, Lee D, Ma M, Liang X, Lal NK, Lemos M, Lu YJ, Ma S, et al. The MAP4 kinase SIK1 ensures robust extracellular ROS burst and antibacterial immunity in plants. Cell Host Microbe. 2018;24(3):379–391. doi: 10.1016/j.chom.2018.08.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Park CJ, Han SW, Chen X, Ronald PC. Elucidation of XA21-mediated innate immunity. Cell Microbiol. 2010;12(8):1017–1025. doi: 10.1111/j.1462-5822.2010.01489.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Song WY, Wang GL, Chen LL, Kim HS, Pi LY, Holsten T, Gardner J, Wang B, Zhai WX, Zhu LH, et al. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science. 1995;270(5243):1804–1806. doi: 10.1126/science.270.5243.1804. [DOI] [PubMed] [Google Scholar]
- 28.Li J, Tax FE. Receptor-like kinases: key regulators of plant development and defense. J Integr Plant Biol. 2013;55(12):1184–1187. doi: 10.1111/jipb.12129. [DOI] [PubMed] [Google Scholar]
- 29.Clark SE, Williams RW, Meyerowitz EM. The CLAVATA1 gene encodes a putative receptor kinase that controls shoot and floral meristem size in Arabidopsis. Cell. 1997;89(4):575–585. doi: 10.1016/S0092-8674(00)80239-1. [DOI] [PubMed] [Google Scholar]
- 30.Park S, Moon JC, Park YC, Kim JH, Kim DS, Jang CS. Molecular dissection of the response of a rice leucine-rich repeat receptor-like kinase (LRR-RLK) gene to abiotic stresses. J Plant Physiol. 2014;171(17):1645–1653. doi: 10.1016/j.jplph.2014.08.002. [DOI] [PubMed] [Google Scholar]
- 31.Hok S, Danchin EG, Allasia V, Panabieres F, Attard A, Keller H. An Arabidopsis (malectin-like) leucine-rich repeat receptor-like kinase contributes to downy mildew disease. Plant Cell Environ. 2011;34(11):1944–1957. doi: 10.1111/j.1365-3040.2011.02390.x. [DOI] [PubMed] [Google Scholar]
- 32.Sanabria NM, van Heerden H, Dubery IA. Molecular characterisation and regulation of a Nicotiana tabacum S-domain receptor-like kinase gene induced during an early rapid response to lipopolysaccharides. Gene. 2012;501(1):39–48. doi: 10.1016/j.gene.2012.03.073. [DOI] [PubMed] [Google Scholar]
- 33.Yoodee S, Kobayashi Y, Songnuan W, Boonchird C, Thitamadee S, Kobayashi I, Narangajavana J. Phytohormone priming elevates the accumulation of defense-related gene transcripts and enhances bacterial blight disease resistance in cassava. Plant Physiol Biochem. 2018;122:65–77. doi: 10.1016/j.plaphy.2017.11.016. [DOI] [PubMed] [Google Scholar]
- 34.Birkenbihl RP, Liu S, Somssich IE. Transcriptional events defining plant immune responses. Curr Opin Plant Biol. 2017;38:1–9. doi: 10.1016/j.pbi.2017.04.004. [DOI] [PubMed] [Google Scholar]
- 35.Moore JW, Loake GJ, Spoel SH. Transcription dynamics in plant immunity. Plant Cell. 2011;23(8):2809–2820. doi: 10.1105/tpc.111.087346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Bhattacharjee S, Garner CM, Gassmann W. New clues in the nucleus: transcriptional reprogramming in effector-triggered immunity. Front Plant Sci. 2013;4:364. doi: 10.3389/fpls.2013.00364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Buscaill P, Rivas S. Transcriptional control of plant defence responses. Curr Opin Plant Biol. 2014;20:35–46. doi: 10.1016/j.pbi.2014.04.004. [DOI] [PubMed] [Google Scholar]
- 38.Tsuda K, Somssich IE. Transcriptional networks in plant immunity. New Phytol. 2015;206(3):932–947. doi: 10.1111/nph.13286. [DOI] [PubMed] [Google Scholar]
- 39.Roodbarkelari F, Groot EP. Regulatory function of homeodomain-leucine zipper (HD-ZIP) family proteins during embryogenesis. New Phytol. 2017;213(1):95–104. doi: 10.1111/nph.14132. [DOI] [PubMed] [Google Scholar]
- 40.Ariel FD, Manavella PA, Dezar CA, Chan RL. The true story of the HD-zip family. Trends Plant Sci. 2007;12(9):419–426. doi: 10.1016/j.tplants.2007.08.003. [DOI] [PubMed] [Google Scholar]
- 41.Gao D, Appiano M, Huibers RP, Chen X, Loonen AE, Visser RG, Wolters AM, Bai Y. Activation tagging of ATHB13 in Arabidopsis thaliana confers broad-spectrum disease resistance. Plant Mol Biol. 2014;86(6):641–653. doi: 10.1007/s11103-014-0253-2. [DOI] [PubMed] [Google Scholar]
- 42.Mou S, Liu Z, Gao F, Yang S, Su M, Shen L, Wu Y, He S. CaHDZ27, a Homeodomain-Leucine zipper I protein, positively regulates the resistance to Ralstonia solanacearum infection in pepper. Mol Plant-Microbe Interact. 2017;30(12):960–973. doi: 10.1094/MPMI-06-17-0130-R. [DOI] [PubMed] [Google Scholar]
- 43.Krupa A, Preethi G, Srinivasan N. Structural modes of stabilization of permissive phosphorylation sites in protein kinases: distinct strategies in Ser/Thr and Tyr kinases. J Mol Biol. 2004;339(5):1025–1039. doi: 10.1016/j.jmb.2004.04.043. [DOI] [PubMed] [Google Scholar]
- 44.Kunkel BN, Brooks DM. Cross talk between signaling pathways in pathogen defense. Curr Opin Plant Biol. 2002;5(4):325–331. doi: 10.1016/S1369-5266(02)00275-3. [DOI] [PubMed] [Google Scholar]
- 45.Zhao S, Qi X. Signaling in plant disease resistance and symbiosis. J Integr Plant Biol. 2008;50(7):799–807. doi: 10.1111/j.1744-7909.2008.00702.x. [DOI] [PubMed] [Google Scholar]
- 46.Tsuda K, Sato M, Stoddard T, Glazebrook J, Katagiri F. Network properties of robust immunity in plants. PLoS Genet. 2009;5(12):e1000772. doi: 10.1371/journal.pgen.1000772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Yang DL, Yang Y, He Z. Roles of plant hormones and their interplay in rice immunity. Mol Plant. 2013;6(3):675–685. doi: 10.1093/mp/sst056. [DOI] [PubMed] [Google Scholar]
- 48.Batistic O, Sorek N, Schultke S, Yalovsky S, Kudla J. Dual fatty acyl modification determines the localization and plasma membrane targeting of CBL/CIPK Ca2+ signaling complexes in Arabidopsis. Plant Cell. 2008;20(5):1346–1362. doi: 10.1105/tpc.108.058123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Choi HW, Kim DS, Kim NH, Jung HW, Ham JH, Hwang BK. Xanthomonas filamentous hemagglutinin-like protein Fha1 interacts with pepper hypersensitive-induced reaction protein CaHIR1 and functions as a virulence factor in host plants. Mol Plant-Microbe Interact. 2013;26(12):1441–1454. doi: 10.1094/MPMI-07-13-0204-R. [DOI] [PubMed] [Google Scholar]
- 50.Choi HW, Kim YJ, Lee SC, Hong JK, Hwang BK. Hydrogen peroxide generation by the pepper extracellular peroxidase CaPO2 activates local and systemic cell death and defense response to bacterial pathogens. Plant Physiol. 2007;145(3):890–904. doi: 10.1104/pp.107.103325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Cai H, Yang S, Yan Y, Xiao Z, Cheng J, Wu J, Qiu A, Lai Y, Mou S, Guan D, et al. CaWRKY6 transcriptionally activates CaWRKY40, regulates Ralstonia solanacearum resistance, and confers high-temperature and high-humidity tolerance in pepper. J Exp Bot. 2015;66(11):3163–74. [DOI] [PubMed]
- 52.Lee B, Park YS, Yi HS, Ryu CM. Systemic induction of the small antibacterial compound in the leaf exudate during Benzothiadiazole-elicited systemic acquired resistance in pepper. Plant Pathol J. 2013;29(3):350–355. doi: 10.5423/PPJ.NT.02.2013.0018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Kim DS, Hwang BK. An important role of the pepper phenylalanine ammonia-lyase gene (PAL1) in salicylic acid-dependent signalling of the defence response to microbial pathogens. J Exp Bot. 2014;65(9):2295–2306. doi: 10.1093/jxb/eru109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Shen L, Liu Z, Yang S, Yang T, Liang J, Wen J, Liu Y, Li J, Shi L, Tang Q, et al. Pepper CabZIP63 acts as a positive regulator during Ralstonia solanacearum or high temperature-high humidity challenge in a positive feedback loop with CaWRKY40. J Exp Bot. 2016;67(8):2439–2451. doi: 10.1093/jxb/erw069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Naoumkina MA, He X, Dixon RA. Elicitor-induced transcription factors for metabolic reprogramming of secondary metabolism in Medicago truncatula. BMC Plant Biol. 2008;8:132. doi: 10.1186/1471-2229-8-132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Yang Y, Zhang Y, Ding P, Johnson K, Li X, Zhang Y. The ankyrin-repeat transmembrane protein BDA1 functions downstream of the receptor-like protein SNC2 to regulate plant immunity. Plant Physiol. 2012;159(4):1857–1865. doi: 10.1104/pp.112.197152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Czernic P, Huang HC, Marco Y. Characterization of hsr201 and hsr515, two tobacco genes preferentially expressed during the hypersensitive reaction provoked by phytopathogenic bacteria. Plant Mol Biol. 1996;31(2):255–265. doi: 10.1007/BF00021788. [DOI] [PubMed] [Google Scholar]
- 58.Palena CM, Gonzalez DH, Chan RL. A monomer-dimer equilibrium modulates the interaction of the sunflower homeodomain leucine-zipper protein Hahb-4 with DNA. Biochem J. 1999;341(Pt 1):81–87. [PMC free article] [PubMed] [Google Scholar]
- 59.Curtis MD, Grossniklaus U. A gateway cloning vector set for high-throughput functional analysis of genes in planta. Plant Physiol. 2003;133(2):462–469. doi: 10.1104/pp.103.027979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Lee DJ, Choi HJ, Moon ME, Chi YT, Ji KY, Choi D. Superoxide serves as a putative signal molecule for plant cell division: overexpression of CaRLK1 promotes the plant cell cycle via accumulation of O2(−) and decrease in H2 O2. Physiol Plant. 2017;159(2):228–243. doi: 10.1111/ppl.12487. [DOI] [PubMed] [Google Scholar]
- 61.Lee DJ, Chi YT, Kim DM, Choi SH, Lee JY, Choi GW. Ectopic expression of CaRLK1 enhances hypoxia tolerance with increasing alanine production in Nicotiana spp. Plant Mol Biol. 2014;86(3):255–270. doi: 10.1007/s11103-014-0227-4. [DOI] [PubMed] [Google Scholar]
- 62.Yi SY, Lee DJ, Yeom SI, Yoon J, Kim YH, Kwon SY, Choi D. A novel pepper (Capsicum annuum) receptor-like kinase functions as a negative regulator of plant cell death via accumulation of superoxide anions. New Phytol. 2010;185(3):701–715. doi: 10.1111/j.1469-8137.2009.03095.x. [DOI] [PubMed] [Google Scholar]
- 63.Han SW, Hwang BK. Molecular functions of Xanthomonas type III effector AvrBsT and its plant interactors in cell death and defense signaling. Planta. 2017;245(2):237–253. doi: 10.1007/s00425-016-2628-x. [DOI] [PubMed] [Google Scholar]
- 64.Kim DS, Kim NH, Hwang BK. The Capsicum annuum class IV chitinase ChitIV interacts with receptor-like cytoplasmic protein kinase PIK1 to accelerate PIK1-triggered cell death and defence responses. J Exp Bot. 2015;66(7):1987–1999. doi: 10.1093/jxb/erv001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Kim DS, Kim NH, Hwang BK. GLYCINE-RICH RNA-BINDING PROTEIN1 interacts with RECEPTOR-LIKE CYTOPLASMIC PROTEIN KINASE1 and suppresses cell death and defense responses in pepper (Capsicum annuum) New Phytol. 2015;205(2):786–800. doi: 10.1111/nph.13105. [DOI] [PubMed] [Google Scholar]
- 66.Kim DS, Hwang BK. The pepper receptor-like cytoplasmic protein kinase CaPIK1 is involved in plant signaling of defense and cell-death responses. Plant J. 2011;66(4):642–655. doi: 10.1111/j.1365-313X.2011.04525.x. [DOI] [PubMed] [Google Scholar]
- 67.Woo JY, Jeong KJ, Kim YJ, Paek KH. CaLecRK-S.5, a pepper L-type lectin receptor kinase gene, confers broad-spectrum resistance by activating priming. J Exp Bot. 2016;67(19):5725–5741. doi: 10.1093/jxb/erw336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Roy S, Nandi AK. Arabidopsis thaliana methionine sulfoxide reductase B8 influences stress-induced cell death and effector-triggered immunity. Plant Mol Biol. 2017;93(1–2):109–120. doi: 10.1007/s11103-016-0550-z. [DOI] [PubMed] [Google Scholar]
- 69.Wang Y, Lin A, Loake GJ, Chu C. H2O2-induced leaf cell death and the crosstalk of reactive nitric/oxygen species. J Integr Plant Biol. 2013;55(3):202–208. doi: 10.1111/jipb.12032. [DOI] [PubMed] [Google Scholar]
- 70.Dardick C, Ronald P. Plant and animal pathogen recognition receptors signal through non-RD kinases. PLoS Pathog. 2006;2(1):14–28. doi: 10.1371/journal.ppat.0020002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Chen X, Chern M, Canlas PE, Jiang C, Ruan D, Cao P, Ronald PC. A conserved threonine residue in the juxtamembrane domain of the XA21 pattern recognition receptor is critical for kinase autophosphorylation and XA21-mediated immunity. J Biol Chem. 2010;285(14):10454–10463. doi: 10.1074/jbc.M109.093427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Parrott DL, Huang L, Fischer AM. Downregulation of a barley (Hordeum vulgare) leucine-rich repeat, non-arginine-aspartate receptor-like protein kinase reduces expression of numerous genes involved in plant pathogen defense. Plant Physiol Biochem. 2016;100:130–140. doi: 10.1016/j.plaphy.2016.01.005. [DOI] [PubMed] [Google Scholar]
- 73.Yamaguchi Y, Pearce G, Ryan CA. The cell surface leucine-rich repeat receptor for AtPep1, an endogenous peptide elicitor in Arabidopsis, is functional in transgenic tobacco cells. Proc Natl Acad Sci U S A. 2006;103(26):10104–10109. doi: 10.1073/pnas.0603729103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Yamaguchi Y, Huffaker A, Bryan AC, Tax FE, Ryan CA. PEPR2 is a second receptor for the Pep1 and Pep2 peptides and contributes to defense responses in Arabidopsis. Plant Cell. 2010;22(2):508–522. doi: 10.1105/tpc.109.068874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Hou S, Wang X, Chen D, Yang X, Wang M, Turra D, Di Pietro A, Zhang W. The secreted peptide PIP1 amplifies immunity through receptor-like kinase 7. PLoS Pathog. 2014;10(9):e1004331. doi: 10.1371/journal.ppat.1004331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Gomez-Gomez L, Boller T. Flagellin perception: a paradigm for innate immunity. Trends Plant Sci. 2002;7(6):251–256. doi: 10.1016/S1360-1385(02)02261-6. [DOI] [PubMed] [Google Scholar]
- 77.Yoshimura S, Yamanouchi U, Katayose Y, Toki S, Wang ZX, Kono I, Kurata N, Yano M, Iwata N, Sasaki T. Expression of Xa1, a bacterial blight-resistance gene in rice, is induced by bacterial inoculation. Proc Natl Acad Sci U S A. 1998;95(4):1663–1668. doi: 10.1073/pnas.95.4.1663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Dang FF, Wang YN, Yu L, Eulgem T, Lai Y, Liu ZQ, Wang X, Qiu AL, Zhang TX, Lin J, et al. CaWRKY40, a WRKY protein of pepper, plays an important role in the regulation of tolerance to heat stress and resistance to Ralstonia solanacearum infection. Plant Cell Environ. 2013;36(4):757–774. doi: 10.1111/pce.12011. [DOI] [PubMed] [Google Scholar]
- 79.Hiratsu K, Matsui K, Koyama T, Ohme-Takagi M. Dominant repression of target genes by chimeric repressors that include the EAR motif, a repression domain, in Arabidopsis. Plant J. 2003;34(5):733–739. doi: 10.1046/j.1365-313X.2003.01759.x. [DOI] [PubMed] [Google Scholar]
- 80.Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1. The phylogenetic relationship of CaLRR-RLK1 protein with other plant LRR-RLKs. An unrooted neighbor-joining tree was built by MEGA 4.0. (TIF 30 kb)
Figure S2. Transient overexpression of HA-CaLRR-RLK1 in pepper leaves detected by immunoblot. (TIF 36 kb)
Figure S3. The expression of CaLRR-RLK1 in representative T3 transgenic tobacco plants was checked by RT-PCR. Wild-type (WT) tobacco plants as the blank control, and NtEF1α served as an endogenous control. (TIF 21 kb)
Table S1. Pepper primers used for qPCR in this study. Table S2. Tobacco primers used for qPCR in this study. Table S3. Primers used in these experiments. (DOCX 21 kb)
Data Availability Statement
All data generated or analysed during this study are included in this published article (and its Additional file 4).