

Abstract
Agave, monocotyledonous succulent plants, is endemic to arid regions of North America, exhibiting exceptional tolerance to their xeric environments. They employ various strategies to overcome environmental constraints, such as crassulacean acid metabolism, wax depositions, and protective leaf morphology. Genomic resources of Agave species have received little attention irrespective of their cultural, economic and ecological importance, which so far prevented the understanding of the molecular bases underlying their adaptations to the arid environment. In this study, we aimed to elucidate molecular mechanism(s) using transcriptome sequencing of A. sisalana. A de novo approach was applied to assemble paired-end reads. The expression study unveiled 3,095 differentially expressed unigenes between well-irrigated and drought-stressed leaf samples. Gene ontology and KEGG analysis specified a significant number of abiotic stress responsive genes and pathways involved in processes like hormonal responses, antioxidant activity, response to stress stimuli, wax biosynthesis, and ROS metabolism. We also identified transcripts belonging to several families harboring important drought-responsive genes. Our study provides the first insight into the genomic structure of A. sisalana underlying adaptations to drought stress, thus providing diverse genetic resources for drought tolerance breeding research.
Introduction
Drought is one of the major abiotic stresses, which significantly diminishes the agricultural production and threatens food security worldwide1. Sessile nature of plants limit them to their natural habitat, therefore many species have evolved appropriate mechanisms to cope with the drought stress such as drought escape, avoidance, and tolerance that may act synergistically2. The employed mechanism largely depends on multiple factors e.g plant species, developmental phase, duration and severity of the drought progression3. All these adaptive mechanisms are complex, polygenic in nature, requiring, physio-biochemical and molecular changes in order to survive4. These changes involve a number of drought particular transcripts that can be associated with two broad groups; “functional proteins” versus “regulatory proteins”5. The induction and accumulation of the functional proteins include dehydrins, photosynthesis-related genes, aquaporins, lipid transfer proteins, biosynthesis and transport of various osmoprotectants, protein repair enzymes, proteases, protease inhibitors, and other enzymes, directly guard the cells against the abiotic factors5,6. The regulatory proteins largely involved in the immediate response to drought stress by directing the expression of downstream genes. These proteins include transcription factors (TFs), protein kinases and phosphatases encoding genes, genes involved in the biosynthesis of abscisic acid (ABA) that control the stomatal behavior and other physiological phenomena5,7.
The Agave is predominantly monocarpic, succulent, xerophytic plants belonging to Asparagaceae family. This genus comprehends more than 166 species, native to the arid and semi-arid origin of Mexico8. Presently they are grown in almost every agricultural area of the world because of their extreme ecological adaptation9. In Pakistan, Agave is represented by six cultivated species10. Many of agave species are of great commercial importance for their use in food, fiber, shelter, insecticides, and ornamentals11. Agave tequilana usually known as “blue agave” is useful to prepare alcoholic beverages such as “pulque” and “tequila” which earns $1.7 billion per annum within the United States12. “Sisal” is the sixth most important fiber, harvested from the Agave sisalana Perr. ex. Engelm, representing 2% of the world’s production of plant fibers13. A. sisalana is a hardy plant that displays exceptional drought and temperature tolerance. It grows well all year round in hot and extremely dry climate14.
The leaves and stem of the agave is the rich source of carbohydrates and lignocelluloses and introduced as a lingo-cellulosic bioenergy feedstock. Its average yield falls in the range of 8.5 to 22 Mg ha−1 yr−1 of dry weight under mild climate conditions15,16. Persistent aridity, with no relief of irrigation, harshly damage the yield to 2.0–5.0 Mg ha−1 yr−1 dry mass. However, an adequate level of management and resource input may lead to 38 and 42 Mg ha−1 yr−1 yield for some species17. Its use for bioenergy production could result in higher yield than other energy crops, such Zea mays (15–19 Mg ha−1), miscanthus species (29–38 Mg ha−1), and Panicum virgatum (10–12 Mg ha−1)18.
Remarkable tolerance to abiotic stresses makes the Agave species an ideal plant to explore essential genomic information for abiotic stress traits. The crassulacean acid metabolism (CAM) mechanism makes them possible to utilize the water 4−2x more efficiently12. They have the inbuilt ability to survive more than one season without rainfall and can tolerate extremely hot and low temperatures (−16.1 °C to 61.4 °C)19. Agave has the large, complex genome, estimated between 2940 to 4704 Mbp of DNA in size with a high level of duplication due to polyploidy levels (2x, 3x, 4x, 5x, 6x, and 8x)20. Notwithstanding, its economic and ecological potential towards the abiotic stress, a limited investigation has been carried out yet. There is just a single transcriptome base de novo assembly reported for species A. tequilana and A. deserti12. Therefore, further comprehensive genome-scale studies are lacking to explore out the molecular basis for adaptation of agave to harsh conditions. Whole transcriptome analysis using the Next-generation sequencing (NGS) enables us to understand the expression patterns in response to the environmental stress. In parallel, advancements in computational tools overcome the complication that may arise due to the lack of suitable well-annotated reference genome for non-model plant species21. These tools assemble the raw reads into short DNA de novo sequences, “contigs”, which enables various downstream analyses like gene discovery, mutation detection, and expression analysis. Transcriptomes of non-model organisms via de novo assembly has been reported for numerous plants12,22–25.
In this study, we aim to fill the gap in the existing knowledge on the transcriptional response by the agaves to the drought stress. A de novo assembly of Illumina platform generated reads was carried out to provide a thorough scenario on the A. sisalana transcriptome under drought stress. The study of differential gene expression and their possible pathways analysis should improve the current knowledge to understand the molecular basis behind the adaptation and survival of agaves in a xeric environment. The present work not only enriches the available knowledge about the genome of agave species but also provides an important transcriptomic database for further molecular investigation.
Results
RNA-Seq data overview
To explore the drought tolerance mechanism(s) at the molecular level, we sequenced and analyzed the leaf specific transcriptome of A. sisalana by mRNA sequencing. Six paired-end cDNA libraries were generated from three well irrigated (control: C1, C2, C3) and from three droughts stressed (drought: T1, T2, T3) independent biological samples. The Illumina sequencing platform Hiseq2500 was used for paired-end sequencing at Macrogen Korea with the insert size 101 bp. A total of 276,845,790 reads and 27,961,424,790 nucleotides were sequenced (Supplementary Information 1a) (Table 1). The data of individual biological library were deposited to NCBI SRA database with SRA accession IDs: SRR5137659, SRR5137661, SRR5137662, SRR5137658, SRR5137663, and SRR5137660. Supplementary Information 1b represents the complete workflow and experimental design.
Table 1.
Numerical Summary of the Illumina generated raw reads and denovo assembly statistics.
| Contents | Control Library (C) | Drought Library (T) | Total |
|---|---|---|---|
| RNA-Sequencing Statistics | |||
| Number of clean reads | 152553060 | 124292730 | 276845790 |
| Total base pairs (bp) | 15407859060 | 12553565730 | 27961424790 |
| Q20 percentage (%) | 97.8% | 95.9% | 96.85% |
| N Percentage | 0 | 0 | 0 |
| GC percentage | 48.29% | 47.7% | 48.1% |
| Assembly Statistics | |||
| Contigs | Unigene | ||
| Total number of sequences | 93141 | 67328 | |
| average length | 731 (bp) | 582 (bp) | |
| N50 | 1164 (bp) | 834 (bp) | |
| Min length | 201 (bp) | 201 (bp) | |
| Max length | 9304 (bp) | 9304 (bp) | |
Transcriptome De novo assembly and evaluation statistics
De novo assembly is an efficient and comprehensive way for the discovery of novel transcripts, their expression behavior and new markers in the absence of the whole genome sequencing data. Length distribution pattern of transcripts produced by assemblers (see materials and methods) was generated against the well-annotated Ananas comosus CDS (https://phytozome.jgi.doe.gov) and transcriptome based A. deserti reference sequences (http://datadryad.org/). Trinity generated assembly correlates closest to the reference’s distribution followed by the Trans-ABySS (64 K-mer) and Short Oligonucleotide Analysis Package (SOAP) (Supplementary Dataset 1 S1, S2). The results may vary from one dataset to others, and so the user should optimize their own preferences/settings according to the data type. We have successfully assembled the 276.8 million reads with Trinity into 93,141 contigs (transcripts hereafter) and 67,328 longest isoforms per gene (unigene hereafter) with 68,048,194 and 39,203,184 bp nucleotides in counts respectively (Supplementary Dataset 1-S3) Table 1. The transcripts and unigenes were in-between 200–9304 bp by length span, with an average length of 731 bp and 582 bp respectively. On average, there were about 43,396 transcripts in the range of 200–400 bp, 26,728 in 401–1000 bp, 16,536 in 1001–2000 bp, 4736 in 2001–3800 bp and 398 transcripts hold >4000 bp, while this counts for unigene were 40849, 16788, 6977, 2461 and 253 respectively (Fig. 1A). GC percentage content (45.3%) of A. sisalana assembly was quite similar to the A. deserti (45.1%) than A. tequliana (42.3%) and O. sativa (55%). Further, a Perl supported script orfPerdictor predicts 92,559 (99.3%) and 63,589 (94.3%) sequences having potential readable ORF from the transcripts and unigenes data, respectively. Additionally, BLAST analysis (Blastp with e value 1e−20) against the viridiplantae (Taxon_ID 33090) database returned more than 25000 sequences with significant hits for both queries (contings and unigenes) (Supplementary Dataset 1-S4). The transcriptome completeness and the quality of de novo assembled reference are critical for the accuracy of the downstream analysis like gene identification, differential gene expression analysis, and genetic molecular developments. RMBT and BUSCO V2 16 are the most widely used packages for the assessment of the de novo assembly. Several recent studies have used the BUSCO tool, as the results have been demonstrated to be more solid than the previously used packages like CEGMA (Core Eukaryotic Genes Mapping Approach) and N50 statistics26. Almost 95% of the reads were mapped back to transcriptome by bowtie2 (RMBT) with 83% completeness while duplication percentage was ~21% as by BUSCO analysis (Fig. 1B,C). This intermediate to high duplication level may be due to the higher polyploidy level of the Agave genome. All these indicators supported that we have generated a high-quality transcriptome assembly that could be used for further possible downstream analysis.
Figure 1.

(A) Sequence length distribution of the transcripts and unigenes of the trinity generated de novo assembly driven out of the raw reads from the control and drought stress samples. (B) Graphical representation of the statistics of cleaned raw reads mapping back to the de novo assembled transcripts (RMBT). (C) Benchmarking Universal Single-Copy Orthologs (BUSCO) scores for assembly quality assessment. (D) Homology analysis of the non-redundant unigenes against the publically available databases.
Functional characterization of the assembled transcriptome
The assembled A. sisalana transcriptome features and functional annotations were based on top hits mapping information from nr database (1.0 e−5), then viridiplantae (51.3%), UniProt (38.6%), Arabidopsis thaliana (46.2%), O. sativa (48.2%), Pfam (37.5%), Gene Ontology (GO) (24.01%), PlantTF database(1e−10) (18.8%) and Cluster of Orthologous Groups of proteins (COG) (13.8%) Fig. 1D (Supplementary Dataset 1-S5). In total 37,546 unigenes assigned functions out of the 67,328 (E-value ≤ 1e−5), which may be due to fewer homologous sequences of A. sisalana in the public database. Maximum homology with sequences from the species like Elaeis guineensis (31%), Phoenix dactylifera (27%), and Musa acuminata subsp. Malaccensis (9%) and others were obtained by BLAST search. This similarity index reflects the close genetic relationship with these species (Supplementary Dataset 1-S6a). Though the leaf sampling was performed in the greenhouse from clean tissues, interestingly, we also got hits outside plants domian like Metazoa, bacteria, fungi, and Amoebozoa, etc. (Supplementary Dataset 1-S6b). Further, GeneMARK (http://exon.gatech.edu/GeneMark/) a standalone gene prediction package retrieved 24,797 functional unigenes having minimum 98 amino acid residues. The unpredicted may have less amino acid residues than the predicted or could be the assembler misassembles or novel sequences. The 9307 (14%) unigenes were divided into 25 categories for functional prediction and classification matching the Cluster of Orthologous Group (COG database; e value 1e−5) (Fig. 2). As per GOSlim distribution, most transcripts were related to the biological process (BP) 58.2%, then molecular functions (MF) 43.2% and cellular components (CC) 35.7%. (Supplementary Dataset 2 S1). We obtained 129 biochemical pathways with the involvement of 6338 unigenes based on KEGG database prediction (http://genome.jp/kegg/) under drought stress (Supplementary Dataset 2 S2, S3). These unigenes were further categorized into five diverse functional groups, namely metabolism (93.4%), the organismal system (4.5%), environmental information processing (0.72%), genetic information processing and cellular processes (0.78%) (Fig. 3). The diverse metabolism category had 5876 unigenes, most of which were involved in nucleotide metabolism (21.05%), carbohydrate metabolism (16.9%), metabolism of cofactor and vitamins (14.4%), amino acid metabolism (10.94%), global and overview maps (8.1%) and other eight subcategories (28.54%). Purine and pyrimidine metabolism were the core group in nucleotide metabolism and treated as the housekeeping function within the plant kingdom. Evidence suggests that they involved in the stress protection to abiotic stress tolerance via activation of the ABA metabolism pathway27. In the biosynthesis of secondary metabolites, the most frequent subsets of sequences were Phenylpropanoid biosynthesis (37.06%), Tropane, piperidine and pyridine alkaloid biosynthesis (14.6%) and Novobiocin biosynthesis (14.3%) (Supplementary Dataset 2-S4). Transcription factors are the main upstream regulatory elements that control the gene expression of sessile nature plants through specific binding to the cis-regulatory elements present in the promoter regions. We predicted, 12,676 transcription factors from the unigenes database and their annotation was retrieved from the PlantTFDB. The major families were associated with the bHLH (9.93%) group, followed by the NAC family (7.02%), MYB related group (6.8%), ERF family (5.6%), C2H2 group (5.09%), WRKY (4.33%), FAR1 (4.07%), C3H (3.9%), MYB group (3.6%) (Fig. 4A). All these are considered to be involved in the regulation of metabolic and secondary metabolic biosynthesis in the green plants22,25,28. Heat Shock protein annotation was retrieved based on the Heat Shock Protein information resource database (http://pdslab.biochem.iisc.ernet.in/hspir/) (Fig. 4B).
Figure 2.

Clusters of orthologous group-based classification of all unigenes. The unigenes were aligned to the COG database (1e−5) to understand their possible protein function. In total 9307 unigenes were annotated and grouped into the 26 categories. The capital letter indicates the COG categories listed on the right while numeric represents the total number of unigenes in each category.
Figure 3.

Pathways classification into metabolism, organismal system, environmental information processing, cellular process and genetic processing major groups based on the KEGG analysis.
Figure 4.

Total genes occupied a proportion of the (A) transcription factors and (B) heat shock proteins families in the A. sisalana de novo assembled transcriptome.
Drought responsive transcripts identification and GO tagging
To investigate the differential gene expression among control and drought group, the bioconductor package edgeR was used on the read counts data that was generated by RSEM package. In total 3095 differentially expressed unigenes (DEG) significantly differed between normal and drought conditions with ≥1-fold expression (log2- fold change)| and FDR less than 0.001 confidence interval. Among these, 1195 genes were up-regulated, while 1864 were down-regulated (Supplementary Information 2 S1–S4). Out of these, 2472 (79.3%) unigenes showed homology in the nr database (2682), viridiplantae (2474), Swiss-Prot (2,047), InterPro scan (2,047), Pfam (2,047), GO (1,438), COG (819) and the KEGG (1435) (Supplementary Information 2 S1). KEGG analysis predicted the involvement of DEGs in 114 pathways. Purine metabolism pathway (ko00230;1104 unigenes) was at the top with the highest DEGs involvement, followed by Thiamine metabolism (Ko00730; unigenes 608), Biosynthesis of antibiotics (Ko0079; 481unigenes), starch and sucrose metabolism (Ko00500; 291 unigenes) and Aminobenzoate degradation (Ko00627;168 unigenes). Various significant drought specific pathways and enzymes belonging to metabolism and other groups were also discovered (Supplementary Information 2-S2). On the basis of gene ontology database, 42%, 36.5% and 29% of DEGs were assigned GO terms in the categories of Biological process, Molecular Functions, and Cellular Components respectively. Search against the COG database divided these DEG into 25 functional groups. Carbohydrates transport (16.06%), posttranslational modification (13.5%), chaperones general function prediction only (9.9%), lipid transport and metabolism (8.8%), signal transduction mechanisms (7.7%) were the most frequent categories. Enriched GO terms specific to drought stress were also identified with Singular Enrichment Analysis (SEA) at 0.05 significance interval (Fig. 5). In total 107 significantly enriched GO terms were identified, including response to abiotic stimulus (GO:0009628), photosynthesis (GO:0015979), response to stimulus (GO:0050896), binding (GO:0005488), cell communication (GO:0007154), transcription (GO:0006350), metabolic process (GO:0008152), cellular process (GO:0009987), catalytic activity (GO: 0003824) and others (Supplementary Dataset 3). The role of transcription factors in the plant response to the abiotic stress is critical and have been studied in a variety of species29,30. Here the GO terms for transcription (GO:0006350), transcription regulator activity (GO:0030528) and transcription factor activity (GO:0003700) were significantly enriched indicating enhanced activity under drought stress. Total 1178 DEGs were predicted as the potential TFs under drought stress in A. sisalana transcriptome, and were further classified into 52 subfamilies (Supplementary Dataset 4-S1). Majority of these genes belonged to ERF family (102), bHLH (100), NAC group (86), MYB_releated (84), C2H2 group (58), WRKY family (46), HSFs (33) and others (Fig. 6A). Heat shock proteins (Hsps) are classified into five major categories based on molecular mass. The differential expression of genes within these categories was calculated based on the fold change. Collectively145 differentially expressed HSP genes were identified, and 100 among them were up-regulated (Fig. 6B). Small heat shock family (HSP20) was the major DE group found in this study followed by the HSP70, HSP100 and HSP90 group. We also identified twenty-nine significantly DE unigenes related to the cytochrome (CYP) gene family, while 75 were related to photosynthesis and light reaction function as revealed by fold change analysis. All of them were down-regulated under drought stress including CAB1 (chlorophyll A/B binding protein 1 and 6), LHB1B1 light-harvesting chlorophyll-protein complex II subunit B1, PSAD -2, PSAF, PSAG, PSAH2, PSAK, PSAL, PSAN (involved in photosystem I), PSB group with subunits (components of photosystem II) and others related to ATPase synthesis (Supplementary Dataset 4-S2).
Figure 5.

GO terms enrichment analysis of all the differentially expressed genes was performed by the AgriGO online tool. Percentage of genes that were associated with specific GO terms are shown on left side of the graph.
Figure 6.
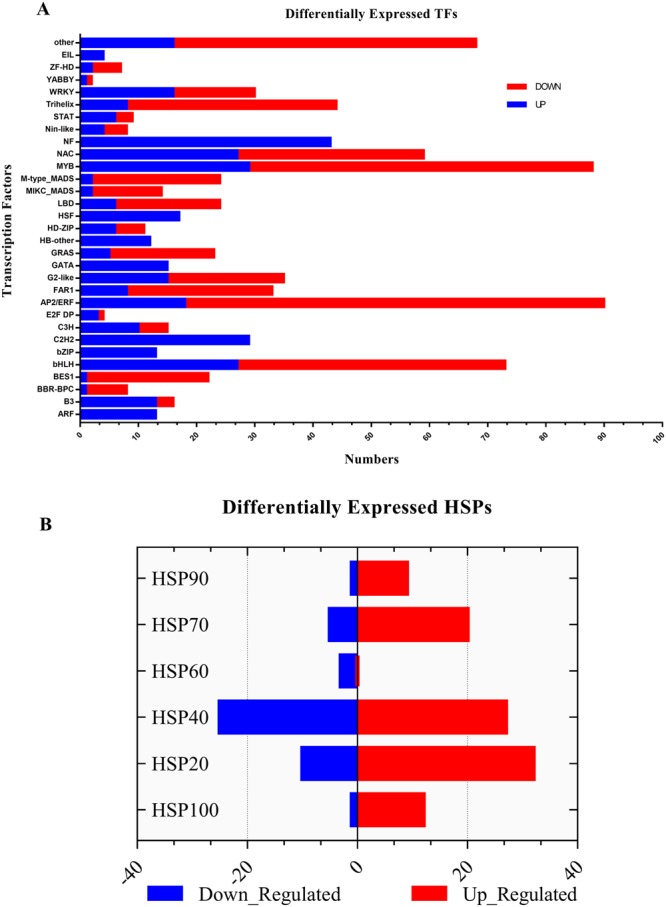
(A) Total number of up and down-regulated transcription factors and their response to drought stress. Within the red bar and blue colors indicating the up-regulated and down-regulated genes respectively. (B) Heat Shock Protein families response to the drought stress.
SSR and SNP detection
The high-throughput transcriptome sequencing provides excellent resources toward the discovery of cost-effective and polymorphic genetic markers (SSRs, SNPs, Indel). We identified total 13,375 SSR markers by using MISA tool within 12,279 unigenes in A. sisalana transcriptome (Supplementary Dataset 5-S1). The average density of microsatellites was found to be one SSR per2.9 kb. Based on the motif repetition, these microsatellites were further categorized into mononucleotide (5318), followed by di-(4347), tri-(3544), tetra-(97), Penta-(37) and hexanucleotides motifs (32), while about 1096 were present in the compound formation (Table 2). (A/T)n motif was the dominant for mononucleotide, (GA/CT/AG)n for di- while (TGC/GAG)n for trinucleotide microsatellites. Specific primers were designed for these SSRs by using Primer3 software and 8164 SSR was verified for a single amplification by in silico PCR with 100-280bp product size (Supplementary Dataset 5-S2). SNPs endure the ability to produces high-density genetic maps, association mapping and molecular markers with the promise of lower cost and error rate. In this study, putative variants were called by aligning the raw reads with the non-redundant de novo assembled reference database. In total 36,525 high confidence variants position were identified includes 35,059 and 1466 for SNPs and indels respectively in 17363 unigenes (Supplementary Dataset 6-S1). An average frequency between all the SNPs in these unigenes was 330 bp. The paleopolyploid nature of A. sisalana may increase the possibility of high counts of SNPs due to the identical paralogous loci in the genome. Large proportion of the unigenes (9576) had the single base shift than di- (3470), tri- (1901) and tetra- (1051), that accounted 27.3%, 9.8%, 5.4% and 2.9% respectively (Supplementary Dataset 6-S2). Transitions and transversion frequencies including six variations are listed in Table 3. The transition between A and G happen most frequently than any other variation. Validation of these SNPs will be required but their annotation indicates potential polymorphism in drought-regulated transcripts.
Table 2.
Statistics of SSRs identified in Agave sisalana.
| SSR mining | |
|---|---|
| Total number of sequences examined | 67,328 |
| Total size of examined sequences (bp) | 39203184 |
| Total number of identified SSRs | 13375 |
| Number of SSR containing sequences | 10729 |
| Number of sequences containing more than one SSR | 2108 |
| Number of SSRs present in compound formation | 1096 |
| Distribution of SSRs in different repeat types | |
| Mono-nucleotide | 5318 (39.7%) |
| Di-nucleotide | 4347 (32.5%) |
| Tri-nucleotide | 3544 (26.4%) |
| Tetra-nucleotide | 97 (0.72%) |
| Penta-nucleotide | 37 (0.28%) |
| Hexa-nucleotide | 32 (0.24%) |
Table 3.
Statistics of identified SNPs.
| Number of SNP | |
|---|---|
| Transition | |
| A<−>G | 10143 (28.9%) |
| C<−>T | 9962 (28.4%) |
| Total | 20105 (57.3%) |
| Transversion | |
| T<−>G | 3817 (10.88%) |
| C<−>G | 3236 (9.23%) |
| A<−>T | 4239 (12.09%) |
| A<−>C | 3662 (10.4%) |
| Total | 14954 (42.6%) |
| Total | 35059 (100%) |
Validation of the RNA-Seq data
To confirm the reliability of expression data, 20 DEGs were studied by using the quantitative real-time PCR (qRT-PCR). The results showed almost same level of fold changes between RNA-Seq expression and qRT-PCR analyses (Fig. 7).
Figure 7.
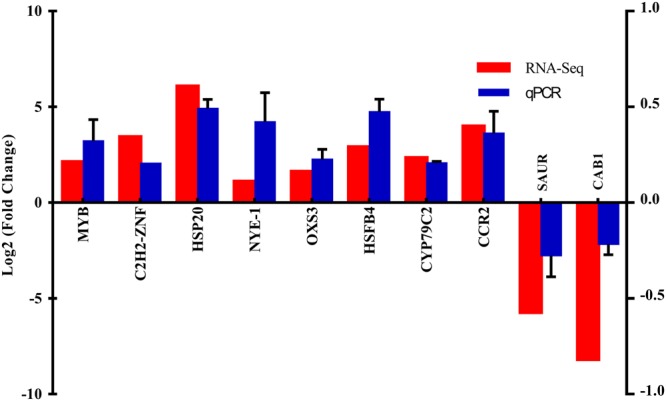
RNA-Seq differentially expressed genes data validation by quantitative real-time PCR (qRT-PCR). (Supplementary File 6 for additional information).
Discussion
Insight into the de novo transcriptome assembly and sequence annotation
Drought tolerance is a multi-pronged mechanism orchestrated by a complex set of gene actions in plants. Its understanding requires a comprehensive approach to explore gene expression and physiological and biochemical pathways. To investigate the dynamic variation of A. sisalana transcriptome to drought conditions, we employed RNA-seq approach using the Illumina platform. We applied ninety days of drought stress which is considered of sufficient length to activate the plant transcriptome under stress as reported in various studies12,31. We observed 18.5% fewer numbers of reads from drought-stressed RNA libraries compared to control data, giving rise to the hypothesis that the A. sisalana genome compromised normal growth processes during the drought with significant up-regulation of signaling and regulatory proteins. In total, 90.3% clean reads were assembled into 67,328 non-redundant unigenes that permitted gene annotation and association of transcripts with biological functions. In addition to functional assignment, the high similarity of unigenes to other plant protein sequences also confirms their integrity32. With BLAST search, 37,546 unigenes out of 67,328 could be functionally assigned with sequences in the nr database, confirming the reliability of the assembly. Insufficient information in public databases and high sequence variations in agave species could arguably be the reasons underlying the high number of un-assigned sequence in A. sisalana genome. GO-based providing information on biological roles33 of transcripts under drought stress was deciding to identify the drought tolerance mechanisms, including the discovery of novel drought stress-related genes in A. sisalana33. Based on differential expression analysis, a number of genes and pathways interactions have been categorized into functional and regulatory groups.
Induction of Functional proteins in drought response
Heat Shock Proteins (HSPs)
The plant’s capability to resist environmental strains is central to development. Protein dysfunctioning is a routine event under the abiotic stress and it is extremely crucial to keep protein functional under the stress. The trigger of the heat shock proteins is the most prominent response to depreciate the cellular injuries and reestablishment of cellular homeostasis. Categorization of these proteins is based on their approximate molecular weight (Hsp100, Hsp90, Hsp70, Hsp60, and HSP20, the small Hsp (sHsp) families)34. In this study, fifty-three heat shock proteins unigenes from six major families includes the sHSP20 group members (HSP17.4, HSP17.6II, HSP18.2, HSP21, ATHSP22.0, and HSP23.6-MITO), HSP70, HSP90.1 and HSP10I were up-regulated under drought stress. HSP-20 was the significantly enriched group with the 8.2 enrichment score including the homolog of Arabidopsis (ATHSP22.0, AT5G51440, AT2G29500, AT1G52560) (Supplementary Dataset 4 S5). sHSPs are the ubiquitous proteins and can be triggered by multiple stresses includes water stress, high temperature, heavy metals, and toxic substances. More than 300-fold expression of small heat shock proteins was observed in S. oleracea under heat stress25. In Arabidopsis, overexpression of GmHsp90s family from Glycine max act as a damage control agent under the abiotic stress35.
Antioxidants response and Osmotic adjustments to dehydration
Over-production of ROS is extremely harmful to plants as it causes lipid peroxidation, DNA damage, and programmed cell death36. The antioxidant enzymes constitute the “first line of defense” against these damages. Here in the study, induced expression of enzymatic and non-enzymatic scavenging molecules indicates the active protection shield against the oxidative stress in A. sisalan leaves. The enriched GO categories like “response to abiotic stimulus” (GO:0009628) and “response to stress” (GO:0009628) give us a strong clue about the active antioxidant enzyme mechanism. Sixteen unigenes were associated in enzymatic scavenging include catalase (CAT1, CAT2), ascorbate peroxidase (APX2, APX4, TL29), peroxidase (PER64, PAP10) and glutathione (Supplementary Dataset 4 S6). Two unigenes (DN17768_c0_g1_i2 and DN19391_c0_g2_i2) encodes the ascorbate peroxidase 2 (APX2) and ascorbate peroxidase 4 (TL29) groups, while others four were homolog to AT1G71695 (Peroxidase superfamily protein), AT2G41480 (PRX25), AT5G66390 (PRX72), and AT4G33420 (PRX47). Ascorbate peroxidase has a significant role in the ascorbate-glutathione detoxification system. GST (ec:2.5.1.18) is critical in glutathione metabolism and is considered as an important indicator for improving the tolerance capability of rice and Arabidopsis37. Genes that encode enzymes (ec:1.8.5.1), glutathione dehydrogenase (ascorbate) and (ec:2.5.1.18) glutathione S-transferase taking part in glutathione metabolism were also detected. Heavy metals accumulation in plants is highly reactive and lethal to living cells. The detoxification transporters and their proteins are well known to detoxify the heavy metals into vacuole of cells and maintain them in a balanced amount38. Total sixteen genes were identified, 6 were associated with the pleiotropic drug resistance-type ATP-binding protein- PDR (PDR4, PDR5, PDR12), two tonoplasts based heavy metal ATPase 2 (HMA2), one was related to farnesylated protein 6 (FP6), others included heavy metal-associated isoprenylated plant protein (HIPP22, HIPP27). In A. thaliana, members of HIPP family involved in cadmium transport play a role in cadmium detoxification36,39.
Osmolytes are the nontoxic small compounds that are synthesized and accumulated in plants under abiotic stress. These include non-toxic macromolecules; organic compounds, sugars, sugar alcohols, starch, lipid peroxidase and free proline40. We also observed significant activities in the metabolism of sugar and starch (non-sugar) related enzymes. The involved pathway was also enriched, “ko00500” with involvement of seven upregulated transcripts. The associated enzymes within this pathway were (ec:3.2.1.21) beta-glucosidase/gentiobiase, (ec:3.2.1.2) saccharogen amylase/beta-amylase, (ec:3.1.3.12) trehalose-6-phosphatase/trehalose-6-phosphate phosphohydrolase and (ec:3.2.1.48) sucrose sucrase/alpha-glucosidase. Trehalose 6-phosphatase (TPP/TPS) is a key player in osmoregulation which strengthens the tolerance in plants to the drought stress25,41. We noted up-regulated six genes “TPS2, TPS3, and TPS6” encoding the trehalose enzymes. Enzyme phosphorylase (ec: 2.4.1.1) was also altered that take part in the decomposition of non-sugar molecules under drought stress. Other enzymes like “(ec:3.1.1.11) - pectinesterase/pectin-demethylase” and “(ec:3.2.1.15)-pectinase/pectin depolymerase were induced under drought stress in this study. They are involved to enhance the cell-to-cell adhesion, cell elongation, the porosity of the wall, disease resistance and ultimately plant growth and development. The role of secondary metabolites like flavonoids, phenylpropanoids are also critical under osmotic stress. Phenylpropanoid biosynthesis pathway (ko00940) was enriched with upregulated enzymes, (ec:2.1.1.146)-O-methyltransferase and (ec:1.11.1.7) peroxidase/lactoperoxidase. Induction of these enzymes indicate the critical role in phenylpropanoid biosynthesis in the osmotic stress. No significant change in expression related to proline biosynthesis was observed.
Cuticle, wax biosynthesis, cell wall metabolism under drought stress
Wax accumulation on outer surface of plant cuticle provides the hydrophobic protection against water loss under osmotic stress42. Biosynthesis of wax begins in epidermal cells of plastids with a C16-C18 long chain of fatty acid with cofactor acyl carrier protein. β-ketoacyl-CoA synthase (KCS), β-ketoacyl-CoA reductase (KCR), β-hydroxy acyl-CoA dehydratase (HCD), and enoyl-CoA reductase (ECR) catalyzed the long chain to produce very long chain fatty acid (VLCFAs). In this investigation, all core-mentioned enzymes that take part in wax biosynthesis and regulation were found in the differentially expressed database except the HCD (Supplementary Dataset 4-S12). The expression of KCS6, GPAT1 (Glycerol-3-phosphate acyltransferase 1) and LTP3 (Lipid transfer proteins 3) were induced under drought stress along with CER1 (Eceriferum /trans-2-enoyl-CoA reductase 1) and EXL 2 and 3 (EXORDIUM like 2). (ec:2.3.1.75) long-chain- alcohol O-fatty-acyltransferase; and (ec:2.3.1.20) palmitoyl-CoA-sn-1,2-diacylglycerol acyltransferase. These enzymes take part in cutin, suberine, and wax biosynthesis pathway (Ko00073) with (ec:1.14.13.8 – monooxygenase) were also upregulated under drought stress. Surprisingly a long list of wax biosynthesis genes was also down-regulated under drought stress (Supplementary Dataset 4-S12). Muthusamy et al. and Ni et al. also reported a high proportion of downregulation of wax biosynthesis transcripts under drought stress43,44.
The ABC transporter G subfamily has been reported to be involved in the export of mature fatty acids in A. thaliana45,46 (Supplementary Dataset 4-S8). The involvement of ERF/AP2 has been extensively reported in the cuticle biosynthesis, especially regulation, accumulation, and transport in response to the abiotic stresses23. MYB TFs are also characterized for their role in the cuticle metabolism44. These factors in combination with other regulatory genes in A. sisalana may act as the coordinator for leaf cuticle synthesis. Identification of these wax related genes would assist further to understand the biosynthesis and functions of the cuticular wax under drought stress.
Signaling and Regulatory Proteins Response to Drought Stress
Ca+ Signaling and activation of kinases (PK & RLK)
Activation of various signaling transduction pathways is key phenomena that happen mostly across the cell membranes to initiate a series of self-protective mechanisms under unfavorable conditions within the cells. Proteins and receptor kinases are the sensors on the membrane of the cell that perceive extracellular signals and transmit them to target genes for the activation of specific stress response. Abundance of the kinases is expected as their domain is actively involved in a number of cellular processes. In this study, seventy-eight significantly differentially expressed transcripts were associated with the protein and receptor kinase group under drought stress conditions. Majority of them belong to PK and RLK superfamily, like Leucine-rich repeat protein kinase and Leucine-rich receptor-like protein kinase family protein RLKs (BAM 1 & 2, BRII, CLVI, ER, and FSL2). The other includes adenosine kinase (ADK 1 & 2, CBL and CBL) interacting protein kinases (CIPK1 & 3, CRCK2), SNF1-related (SNRK2 0.1), Serine/Threonine kinase catalytic domain protein (NEK5) and other (Supplementary Dataset 4-S4). Leucine-rich receptor-like kinases (RLKs) are one of the major group that managed the meristem proliferation, reproduction, organ initiation, specification and hormonal signal cascade. There are several reports that revealed their response towards the drought tolerance e.g. in Arabidopsis abrupt increased of RLKs was observed towards the osmotic stress47.
The abrupt increase in the calcium ions happens in plants under abiotic stress conditions, is a sign of activation of the stress-responsive cascades. In this study, nine significantly enriched unigenes that belong to the calcium transport signaling group includes, calcium ion binding protein (SUB, SUB1), calcium exchanger (CAX3, CAX5, and CAX7) and tonoplast calcium sensor (CBL3) have been identified (Supplementary Dataset 4-S3). The induced response of these proteins under drought stress stabilized the structural rigidity of cell wall. The CAX group of genes has been discovered in a number of plant species and act ubiquitously as they regulate the tonoplast localized Ca 2+/H + antiport activities. Furthermore, the interaction among different protein phosphatases like HAI2, HAI3 and kinases such as serine/threonine-protein kinase (NEK5), CBL-interacting protein kinase initiated the protein phosphorylation cascade which take part in cell signal recognition and transduction in the responses to abiotic stress48. SNF1-related protein kinase 2.1 (SNRK2.1) also act as a positive regulator of the hormonal (ABA) signaling. In A. thaliana complex association between the Calcineurin like proteins (CBL4/CIPK) are associated with the sodium ions release from the cells and absorption of K+ by the root surface, that regulates the stomatal behavior under osmotic stress49.
Phytohormones pathways gene to drought stress
To combat various environmental stresses, novel and dynamic approaches should be devised, and phytohormone engineering could be a method of choice to improve the productivity including drought resistance50. Plant hormones improve the resistance to osmotic stress by regulating the physiological process. The abscisic acid (ABA) is a key plant growth regulator that directly involved in the responses to abiotic stresses50. Here, we noted twenty-three up-regulated unigenes related to ABA-induced protein phosphatase 2 and 3 (HAI2 and HAI3) (ec:3.1.3.16), one protein phosphatase 2CA group (PP2CA) (ec:3.1.3.16), homolog of ABI2 (HAB, HAB2), while four with protein phosphatase 2C families (ABI1) that were homologous to AT3G62260, AT3G63320, AT1G18030, AT3G12620 IDs. A higher number of up regulations of the ABA encodes unigenes including ABA receptor family (PYL4) is an indication of accumulation of ABA due to the decreased cellular water contents under drought stress (Supplementary Dataset 4-S9). Protein phosphatases are the chief regulators and are considered to mediate the ABA triggered signaling pathways. Induced PP2C and PP3C (Protein phosphatases) level in association with the ABA pathway indicated its hyper response to the drought stress in A. sisalana, which is a conserved mechanism in the metabolism of ABA. The differential expression of these genes may regulate the guard’s cell of stomata for gaseous exchange and activation of ABA-dependent regulatory elements, such as MYB factors.
Auxin biosynthesis and transport are essential in regulating the response to environmental stresses, including drought, salinity, and pathogen attack. Changes in Indole-3-acetic acid (IAA) biosynthesis in response to external stimulus regulate the stomatal closure via cross-talk with other plant hormones like ABA and others. The IAA mutant plant of Arabidopsis exhibited significant induced water loss than the normal plants51. In this study, gene enrichment analysis showed the number of genes contributing to the growth under drought stress related to auxin hormones including auxin-induced protein (IAA13, IAA16, IAA33), auxin response factor (ARF-1,9,11,19) that were homologous to AT1G19220, AT4G23980, AT1G59750, AT2G46530, GH3 and 4, auxin efflux carrier family protein (PIN1and EIR1), like-LAX2 related gene, auxin- responsive factor AUX/IAA-like protein (NPH4) and auxin binding ABP like proteins (Supplementary Dataset 4-S10). Several positively regulated induced auxins genes is an indication of there important role in A. sisalana against drought stress.
We also observed thirty-seven DE unigenes related to the cytochrome p450s gene family (Supplementary Dataset 4-S11). Cytochrome is one of the largest and central superfamilies in plants, so far encoding about 1% of the protein coding sequences that act in hormonal control mechanisms including biosynthesis and catabolism of primary and secondary metabolites52. Several members of this group like CYP71 are known to catalyze the production of aliphatic and aromatic nitriles suggesting their possible role in the defense to the biotic stress53,54. Members of CYP86, CYP94, CYP96, and CYP704 are also known as candidates for cuticle biosynthesis55,56. The detection of these cytochromes in our dataset may indicate their potential role for cuticle biosynthetic . The promoter region of various cytochrome genes has the affinity for the drought-induced TF includes MYB/MYC, TGA, and W-box for the WRKY. The appearance of high number unigenes associated with these TFs and the CYPs-450 might be a strategy to combat stress. Biosynthesis of jasmonic acid (JA) and Brassinosteroids (BR) hormones are also stressed sensitive. In our data, we noted two differentially expressed genes involved in the alpha-Linolenic acid metabolism that regulate the JA biosynthesis57. There are several reports that confirmed their involvements to improve the stress tolerance ability of drought-tolerant cultivars58,59. The transcription factor-like MYC2 is a key regulator of JA response and their upregulation in this study indicates its regulatory role in this process and act as a mediator in cross-talk along with WRKY and MYB TFs.
Transcriptional regulatory network induced a response to drought stress
TFs are the key regulatory switches that directly regulate the signal transduction pathways60. In eukaryotes, especially in plants, TFs are highly conserved and represented by various multigene families to perform specific functions. The number of genes encoding these families may vary due to origin, expansion, and tissue-specific functions. In the current study, 372 transcription factors belonging to the ERF (E2F3) family, bHLH, NAC, HSF, MYB and Zinc finger-like protein and others were found to be differentially upregulated under drought stress (Supplementary Dataset 6 S1). In addition, we also found two transcription factors of the GRAS family, PAT1, and SCL7 (homologs of AT5G41920). GRAS plays a critical function in plant growth and environmental adaptations, especially in the modulation of plant tolerance to stress61. In A.thaliana, up-regulation of the SCL7 and SCL 23 TFs has enhanced tolerance to the salt and drought stress58. Heat Shock TFs (HSF) are central facilitator for expression of the genes responsive to various abiotic stress conditions. Here, eight induced DEG got annotation to HSFs group, including HSF3, HSFA3, HSF-A4A, HSFB2A, HSFB4, and HSFC1. NAC proteins are plant-specific TFs that are considered important for plant development, abiotic stress responses as well as for ABA signaling. The Arabidopsis and rice genome hold 106 and 149 NAC proteins respectively. Here we found 27 induced AsNAC related TFs to drought stress. Overexpression of NAC proteins enhanced the longevity and abiotic stress tolerance efficiency in Arabidopsis, Oryza sativa, Zea mays (ZmNAC55) and cicer (CarNAC4)62.
Conclusion
To the best of our knowledge, here we reported the first transcriptome study of Agave sisalana with the objective to identify the functional genes associated with drought tolerance. 67328 unigenes were de novo assembled, and 37546 were functionally annotated. Further differential gene expression provides the clear understanding of responsive mechanism to drought stress. In addition, the identified genetic marker will provide the source for marker development in this species. This study may not only provide the insights to genomics of adaptation of drought tolerance in agave but also excellent genetic resources for drought tolerance crop development.
Materials and Methods
Plant material, stress conditions, and tissue sampling
The offshoots of similar age/height from 1-year-old mature adult A. sisalana plants, which were asexually propagated from a single “the mother plant,” were used for the current study (Supplementary Information 3-S1). These offshoots were further grown in pots (one per pot) having the soil mixture of peat moss, vermiculite and sandy soil in the ratio (1:1:1) in the greenhouse. After 90 days of propagation, we divided these plants into two groups randomly with three replicates each. A control group (C); watered regularly while the other treated group (T); no water was applied until the leaf sampling. The newly emerged middle leaves of the A. sisalana rosette (Supplementary Information 3-S2) were harvested from each plant of the group, immersed into liquid nitrogen, ground well to a very fine powder and stored at −80 °C till further molecular investigation.
RNA isolation, cDNA library preparation, and Illumina sequencing
Total RNA was isolated from the stored ground leaves by using the Trizol method with column based purification as described by34. Genomic DNA contamination was removed by RNase free amplification grade DNase I kit (AMPD1-sigma). TruSeq RNA Sample Prep Kit v2 (Illumina, Inc. San Diego, CA, USA) protocol was used for library constructions. Six paired-end cDNA libraries were constructed and sequenced on the Illumina HiSeqTM2500 platform with the 101 bp insert size at Macrogen Inc. (Korea). The FastQC (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/) application and NGS QC Toolkit (http://www.nipgr.res.in/ngsqctoolkit.html) was used to ensure the standard quality statistic for the FASTQ files.
Transcriptome de novo assembly and evaluation
The high quality, adapter free reads were used to construct the de novo assembly with assemblers including Trinity v 2.3.1 (https://github.com/trinityrnaseq/trinityrnaseq/wiki) under the default settings (25 K-mer), Trans-ABySS (http://www.bcgsc.ca/platform/bioinfo/software/trans-abyss) with multi K-mer adjustment to include odd numbers, i.e., 23 k-mer, 25, 27 and so on up to 63 k-mer, and Short Oligonucleotide Analysis Package (SOAP) (http://soap.genomics.org.cn/SOAPdenovo-Trans.html). Quality evaluation of assemblies was considered with major bioinformatics indicators like contigs mean length, GC percentage, N50, and N25 value. We also compared the mean distribution plot of the contigs produced by aforementioned assemblers with the Ananas comosus V3 partial genome assembly obtained from (https://phytozome.jgi.doe.gov/pz/portal.html#!info?alias=Org_Acomosus_er) and A. deserti transcriptome data (http://datadryad.org/resource/doi:10.5061/dryad.h5t68). Based on these indicators, the Trinity developed assembly was selected for further downstream investigations. Further unigene decontamination was done using MEGAN version 6.01 (http://ab.inf.uni-tuebingen.de/software/megan/) on taxonomy ID basis. The longest isoform (unigenes) was generated by combining the contigs having consensus sequences by Perl script obtained from google group “Trinity RNAseq-user” by Brian Haas (https://groups.google.com/forum/#!forum/trinityrnaseq-users). Assembly quality was assessed using three approaches. (i) a Perl script ORFpredictor (ii) Reads mapping back to transcriptome (RMBT) (iii) BUSCO v1.161 (Benchmarking Universal Single-Copy Orthologs) analysis.
Transcript annotation and functional classification
Cleaned contigs were annotated using the NCBI standalone local BLASTX Programme with the cutoff e-value 10−5 against the NCBI nr database (ftp://ftp.ncbi.nlm.nih.gov/blast/db/FASTA /), SwissProt/uniport (http://web.expasy.org/docs/swiss-prot_guideline.html), Viridiplantae database taxonomy ID (tax ID: 33090) (http://www.uniprot.org/taxonomy/33090), COG database https://www.ncbi.nlm.nih.gov/COG/ and plant TFdatabase (http://planttfdb.cbi.pku.edu.cn/) by homology search. The Kyoto Encyclopedia of Genes and Genomes (http://www.genome.jp/kegg/) was accessed for biochemical pathway identification based on the assigned enzyme codes. The best BLAST hits were used to choose the downstream analysis direction. GO analysis was performed in the standalone Blast2GO v3.2 (https://www.blast2go.com) with value 1e−3, annotation cutoff filter 55, code set to 0.8 to assign the GO terms to each transcript regarding molecular functions (MFs), biological processes (BPs), and cellular components (CCs). GO enrichment analysis was carried out by AgriGO software with FDR value not less than <0.05.
Transcript count and differentially expressed gene identification
First, the abundance of each transcript was calculated by bowtie2 and RSEM (RNA-Seq by Expectation-Maximization-http://deweylab.github.io/RSEM/package) for each library. Differentially expressed genes (DEGs) among the drought and control treated libraries were calculated by using the Empirical Analysis of Digital Gene Expression (edgeR) (http://bioconductor.org/packages/release/bioc/html/edgeR.html) statistical package. The trimmed mean of M-values (TMM) method was used to calculate the normalization factors. The threshold p-value < 0.05 and false discovery rate (FDR) < 0.001 was adjusted to identify the differentially expressed genes by fold change ( ≥ 1).
Simple Sequence Repeat (SSR) and Single-Nucleotide Polymorphism (SSR) Calling
Perl supported script MISA (MIcroSAtellite identification tool- http://pgrc.ipk-gatersleben.de/misa/) was used to mining the SSR repeats with di-, tri-, tetra-, penta- and hexanucleotide motifs present in the A. sisalana assembly. The latest version of PRIMER3 with modifies Perl scripts (p3_in_v2.pl p3_out_v2.pl https://gist.github.com/ascatanach/7a562621b9c86c7b5e81973136e6419f) was used for primer designing. Clean reads from the six libraries were aligned back to the unigenes by short read aligner (bowtie2) with default parameter63. Further SNPs and indel calling was carried out using the mpileup function of SAMtools (http://samtools.sourceforge.net/) and VarScan (http://varscan.sourceforge.net/) mpileup v0.1.7a64,65.
Quantitative RT-qPCR validation
We randomly selected 10 annotated DEGs to verify the RNA-Seq expression data. Gene-specific primers were designed from the selected unigene sequences using Primer 6.0 software (http://www.premierbiosoft.com/primerdesign/) (Supplementary Information 4). Relative fold expression (RT-qPCR) was carried out on the IQ5 system (BioRad) by using the SYBR® Green PCR Master Mix (cat#4309155). Thermal settings included the following conditions: 95 °C for 3 min, followed by 40 cycles at 95 °C for 30 s, then 60 °C for 30 s and at 72 °C for 30 s. All this study was carried out on three independent biological and technical replicates. Relative Expression Software Tool (REST) (http://www.gene-quantification.com/rest-2009.html) was used for relative fold expression calculation.
Electronic supplementary material
Acknowledgements
The authors are thankful to Dr. Khalid Mehmood (AU) and Mukhtar Ahmed (CEMB) for their support in data analysis. The authors are also thankful to the Higher Education Commission (HEC) Pakistan for the provision of funds for this study.
Author Contributions
Conceived and designed the Experiment: M.B.S., B.R., Z.A. Analyzed the data: M.B.S., M.L., I.N. Contributed to the writing of the research article: M.B.S., P.L.G., S.H., T.A. and T.H. approved the final draft. All authors reviewed and approved the final manuscript.
Competing Interests
The authors declare no competing interests.
Footnotes
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Supplementary information accompanies this paper at 10.1038/s41598-018-35891-6.
References
- 1.Wang W, Vinocur B, Altman A. Plant responses to drought, salinity and extreme temperatures: towards genetic engineering for stress tolerance. Planta. 2003;218:1–14. doi: 10.1007/s00425-003-1105-5. [DOI] [PubMed] [Google Scholar]
- 2.Blum, A. In Drought tolerance in higher plants: genetical, physiological and molecular biological analysis 57–70 (Springer, 1996).
- 3.Pinheiro C, Chaves M. Photosynthesis and drought: can we make metabolic connections from available data? Journal of experimental botany. 2010;62:869–882. doi: 10.1093/jxb/erq340. [DOI] [PubMed] [Google Scholar]
- 4.Pieczynski M, et al. Genomewide identification of genes involved in the potato response to drought indicates functional evolutionary conservation with Arabidopsis plants. Plant biotechnology journal. 2018;16:603–614. doi: 10.1111/pbi.12800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Shinozaki K, Yamaguchi-Shinozaki K. Gene networks involved in drought stress response and tolerance. Journal of experimental botany. 2007;58:221–227. doi: 10.1093/jxb/erl164. [DOI] [PubMed] [Google Scholar]
- 6.Ramanjulu S, Bartels D. Drought‐and desiccation‐induced modulation of gene expression in plants. Plant, cell & environment. 2002;25:141–151. doi: 10.1046/j.0016-8025.2001.00764.x. [DOI] [PubMed] [Google Scholar]
- 7.Seki M, Umezawa T, Urano K, Shinozaki K. Regulatory metabolic networks in drought stress responses. Current opinion in plant biology. 2007;10:296–302. doi: 10.1016/j.pbi.2007.04.014. [DOI] [PubMed] [Google Scholar]
- 8.Nobel PS. Water relations and photosynthesis of a desert CAM plant, Agave deserti. Plant Physiology. 1976;58:576–582. doi: 10.1104/pp.58.4.576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Mcdaniel R. Field evaluation of Agave parryi and A. americana in Arizona. Univ. of Arizona, Tucson, USA Desert Plant Dept., Ornamental Horti. Abst. 1985;7R(2):57–60. [Google Scholar]
- 10.Kanwal H, Hameed M, Nawaz T, Ahmad MA, Younis A. Structural adaptations for adaptability in some exotic and naturalized species of Agavaceae. Pak. J. Bot. 2012;44:129–134. [Google Scholar]
- 11.Delgado-Lemus A, Casas A, Téllez O. Journal of ethnobiology and ethnomedicine. 2014. Distribution, abundance and traditional management of Agave potatorum in the Tehuacán Valley, Mexico: bases for sustainable use of non-timber forest products; p. 63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Gross SM, et al. De novo transcriptome assembly of drought tolerant CAM plants, Agave deserti and Agave tequilana. BMC genomics. 2013;14:563. doi: 10.1186/1471-2164-14-563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.FAO. Future Fibers (http://www.fao.org/economic/futurefibres/home/en/) (2018).
- 14.Nikam T, Bansude G, Kumar KA. Somatic embryogenesis in sisal (Agave sisalana Perr. ex. Engelm) Plant cell reports. 2003;22:188–194. doi: 10.1007/s00299-003-0675-9. [DOI] [PubMed] [Google Scholar]
- 15.Davis SC, Kuzmick ER, Niechayev N, Hunsaker DJ. Productivity and water use efficiency of Agave americana in the first field trial as bioenergy feedstock on arid lands. Gcb Bioenergy. 2017;9:314–325. doi: 10.1111/gcbb.12324. [DOI] [Google Scholar]
- 16.Davis SC, LeBauer DS, Long SP. Light to liquid fuel: theoretical and realized energy conversion efficiency of plants using Crassulacean Acid Metabolism (CAM) in arid conditions. Journal of experimental botany. 2014;65:3471–3478. doi: 10.1093/jxb/eru163. [DOI] [PubMed] [Google Scholar]
- 17.Nobel P, García‐Moya E, Quero E. High annual productivity of certain agaves and cacti under cultivation. Plant, Cell & Environment. 1992;15:329–335. doi: 10.1111/j.1365-3040.1992.tb00981.x. [DOI] [Google Scholar]
- 18.Heaton EA, Dohleman FG, Long SP. Meeting US biofuel goals with less land: the potential of Miscanthus. Global change biology. 2008;14:2000–2014. doi: 10.1111/j.1365-2486.2008.01662.x. [DOI] [Google Scholar]
- 19.NOBEL PS, SMITH SD. High and low temperature tolerances and their relationships to distribution of agaves. Plant, Cell & Environment. 1983;6:711–719. [Google Scholar]
- 20.Zhou W-Z, Zhang Y-M, Lu J-Y, Li J-F. Construction and evaluation of normalized cDNA libraries enriched with full-length sequences for rapid discovery of new genes from sisal (Agave sisalana Perr.) different developmental stages. International journal of molecular sciences. 2012;13:13150–13168. doi: 10.3390/ijms131013150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Schliesky S, Gowik U, Weber AP, Bräutigam A. RNA-seq assembly–are we there yet? Frontiers in plant science. 2012;3:220. doi: 10.3389/fpls.2012.00220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Li H, Yao W, Fu Y, Li S, Guo Q. De novo assembly and discovery of genes that are involved in drought tolerance in Tibetan Sophora moorcroftiana. PloS one. 2015;10:e111054. doi: 10.1371/journal.pone.0111054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ma X, et al. De novo transcriptome sequencing and comprehensive analysis of the drought-responsive genes in the desert plant Cynanchum komarovii. BMC genomics. 2015;16:753. doi: 10.1186/s12864-015-1873-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Talukder, S. et al. De novo assembly and characterization of tall fescue transcriptome under water stress. The Plant Genome8 (2015). [DOI] [PubMed]
- 25.Yan J, et al. De novo transcriptome sequencing and gene expression profiling of spinach (Spinacia oleracea L.) leaves under heat stress. Scientific reports. 2016;6:19473. doi: 10.1038/srep19473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Moreton, J., Izquierdo, A. & Emes, R. D. Assembly, assessment, and availability of de novo generated eukaryotic transcriptomes. Frontiers in genetics6 (2015). [DOI] [PMC free article] [PubMed]
- 27.Watanabe S, et al. The purine metabolite allantoin enhances abiotic stress tolerance through synergistic activation of abscisic acid metabolism. Plant, cell & environment. 2014;37:1022–1036. doi: 10.1111/pce.12218. [DOI] [PubMed] [Google Scholar]
- 28.Zhou Z, et al. RNA-seq Reveals Complicated Transcriptomic Responses to Drought Stress in a Nonmodel Tropic Plant, Bombax ceiba L. Evolutionary bioinformatics online. 2015;11:27. doi: 10.4137/EBO.S20620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Fujita Y, Fujita M, Shinozaki K, Yamaguchi-Shinozaki K. ABA-mediated transcriptional regulation in response to osmotic stress in plants. Journal of plant research. 2011;124:509–525. doi: 10.1007/s10265-011-0412-3. [DOI] [PubMed] [Google Scholar]
- 30.Lata C, Prasad M. Role of DREBs in regulation of abiotic stress responses in plants. Journal of experimental botany. 2011;62:4731–4748. doi: 10.1093/jxb/err210. [DOI] [PubMed] [Google Scholar]
- 31.Jain M, Ghanashyam C, Bhattacharjee A. Comprehensive expression analysis suggests overlapping and specific roles of rice glutathione S-transferase genes during development and stress responses. BMC genomics. 2010;11:73. doi: 10.1186/1471-2164-11-73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Wu, B. et al. Transcriptome analysis of hexaploid hulless oat in response to salinity stress. PLOS ONE12, 10.1371/journal.pone.0171451 (2017). [DOI] [PMC free article] [PubMed]
- 33.Jensen, L. J. & Bork, P. Ontologies in quantitative biology: a basis for comparison, integration, and discovery. PLoS biology8, e1000374 (2010). [DOI] [PMC free article] [PubMed]
- 34.Sarwar, M. B. et al. Integration and expression of heat shock protein gene in segregating population of transgenic cotton for drought tolerance. Pakistan Journal of Agricultural Sciences51 (2014).
- 35.Xu J, et al. Overexpression of GmHsp90s, a heat shock protein 90 (Hsp90) gene family cloning from soybean, decrease damage of abiotic stresses in Arabidopsis thaliana. PLoS One. 2013;8:e69810. doi: 10.1371/journal.pone.0069810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Mittler R, et al. ROS signaling: the new wave? Trends in plant science. 2011;16:300–309. doi: 10.1016/j.tplants.2011.03.007. [DOI] [PubMed] [Google Scholar]
- 37.Chen, J.-H. et al. Drought and salt stress tolerance of an Arabidopsis glutathione S-transferase U17 knockout mutant are attributed to the combined effect of glutathione and abscisic acid. Plant Physiol. 158, 340–351 (2012). [DOI] [PMC free article] [PubMed]
- 38.Ueno, D. et al. Gene limiting cadmium accumulation in rice. Proceedings of the National Academy of Sciences107(38), 16500–16505 (2010). [DOI] [PMC free article] [PubMed]
- 39.Mittler R, Vanderauwera S, Gollery M, Van Breusegem F. Reactive oxygen gene network of plants. Trends in plant science. 2004;9:490–498. doi: 10.1016/j.tplants.2004.08.009. [DOI] [PubMed] [Google Scholar]
- 40.Slama I, Abdelly C, Bouchereau A, Flowers T, Savoure A. Diversity, distribution and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. Annals of Botany. 2015;115:433–447. doi: 10.1093/aob/mcu239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Yan Q, et al. GmCYP82A3, a soybean cytochrome P450 family gene involved in the jasmonic acid and ethylene signaling pathway, enhances plant resistance to biotic and abiotic stresses. PloS one. 2016;11:e0162253. doi: 10.1371/journal.pone.0162253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Yeats, T. H. & Rose, J. K. The formation and function of plant cuticles. Plant physiology, pp. 113.222737 (2013). [DOI] [PMC free article] [PubMed]
- 43.Muthusamy, M., Uma, S., Backiyarani, S., Saraswathi, M. S. & Chandrasekar, A. Transcriptomic Changes of Drought-Tolerant and Sensitive Banana Cultivars Exposed to DroughtStress. Frontiers in plant science 7 (2016). [DOI] [PMC free article] [PubMed]
- 44.Ni Y, Guo N, Zhao Q, Guo Y. Identification of candidate genes involved in wax deposition in Poa pratensis by RNA-seq. BMC genomics. 2016;17:314. doi: 10.1186/s12864-016-2641-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Bessire M, et al. A member of the PLEIOTROPIC DRUG RESISTANCE family of ATP binding cassette transporters is required for the formation of a functional cuticle in Arabidopsis. The Plant Cell. 2011;23:1958–1970. doi: 10.1105/tpc.111.083121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Panikashvili D, et al. The Arabidopsis DESPERADO/AtWBC11 transporter is required for cutin and wax secretion. Plant physiology. 2007;145:1345–1360. doi: 10.1104/pp.107.105676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Osakabe Y, Yamaguchi-Shinozaki K, Shinozaki K, Tran L-SP. Sensing the environment: key roles of membrane-localized kinases in plant perception and response to abiotic stress. Journal of experimental botany. 2013;64:445–458. doi: 10.1093/jxb/ers354. [DOI] [PubMed] [Google Scholar]
- 48.Luan S. The CBL–CIPK network in plant calcium signaling. Trends in plant science. 2009;14:37–42. doi: 10.1016/j.tplants.2008.10.005. [DOI] [PubMed] [Google Scholar]
- 49.Kudla J, Batisti O, Hashimoto K. Calcium signals: the lead currency of plant information processing. The Plant Cell. 2010;22:541–563. doi: 10.1105/tpc.109.072686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sah, S. K., Reddy, K. R. & Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Frontiers in Plant Science 7, 10.3389/fpls.2016.00571 (2016). [DOI] [PMC free article] [PubMed]
- 51.Shi H, et al. Modulation of auxin content in Arabidopsis confers improved drought stress resistance. Plant Physiology and Biochemistry. 2014;82:209–217. doi: 10.1016/j.plaphy.2014.06.008. [DOI] [PubMed] [Google Scholar]
- 52.Mizutani M. Impacts of diversification of cytochrome P450 on plant metabolism. Biological and Pharmaceutical Bulletin. 2012;35:824–832. doi: 10.1248/bpb.35.824. [DOI] [PubMed] [Google Scholar]
- 53.Frisch T, Møller BL. Possible evolution of alliarinoside biosynthesis from the glucosinolate pathway in Alliaria petiolata. The FEBS journal. 2012;279:1545–1562. doi: 10.1111/j.1742-4658.2011.08469.x. [DOI] [PubMed] [Google Scholar]
- 54.Nelson D, Werck‐Reichhart D. A P450‐centric view of plant evolution. The Plant Journal. 2011;66:194–211. doi: 10.1111/j.1365-313X.2011.04529.x. [DOI] [PubMed] [Google Scholar]
- 55.Wellesen K, et al. Functional analysis of the LACERATA gene of Arabidopsis provides evidence for different roles of fatty acid ω-hydroxylation in development. Proceedings of the National Academy of Sciences. 2001;98:9694–9699. doi: 10.1073/pnas.171285998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Xiao F, et al. Arabidopsis CYP86A2 represses Pseudomonas syringae type III genes and is required for cuticle development. The EMBO journal. 2004;23:2903–2913. doi: 10.1038/sj.emboj.7600290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Pan H, et al. Structural studies of cinnamoyl-CoA reductase and cinnamyl-alcohol dehydrogenase, key enzymes of monolignol biosynthesis. The Plant Cell. 2014;26:3709–3727. doi: 10.1105/tpc.114.127399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Bai ZY, et al. Whole-transcriptome sequence analysis of differentially expressed genes in Phormium tenax under drought stress. Sci Rep. 2017;7:41700. doi: 10.1038/srep41700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Lenka SK, Katiyar A, Chinnusamy V, Bansal KC. Comparative analysis of drought responsive transcriptome in Indica rice genotypes with contrasting drought tolerance. Plant biotechnology journal. 2011;9:315–327. doi: 10.1111/j.1467-7652.2010.00560.x. [DOI] [PubMed] [Google Scholar]
- 60.Gupta K, Agarwal PK, Reddy M, Jha B. SbDREB2A, an A-2 type DREB transcription factor from extreme halophyte Salicornia brachiata confers abiotic stress tolerance in Escherichia coli. Plant cell reports. 2010;29:1131–1137. doi: 10.1007/s00299-010-0896-7. [DOI] [PubMed] [Google Scholar]
- 61.Hirsch S, Oldroyd GE. GRAS-domain transcription factors that regulate plant development. Plant signaling & behavior. 2009;4:698–700. doi: 10.4161/psb.4.8.9176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Marques DN, dos Reis SP, de Souza CR. Plant NAC transcription factors responsive to abiotic stresses. Plant Gene. 2017;11:170–179. doi: 10.1016/j.plgene.2017.06.003. [DOI] [Google Scholar]
- 63.Langmead B, Salzberg SL. Fast gapped-read alignment with Bowtie 2. Nature methods. 2012;9:357. doi: 10.1038/nmeth.1923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Li H, et al. The sequence alignment/map format and SAMtools. Bioinformatics. 2009;25:2078–2079. doi: 10.1093/bioinformatics/btp352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Li H, Durbin R. Fast and accurate short read alignment with Burrows–Wheeler transform. bioinformatics. 2009;25:1754–1760. doi: 10.1093/bioinformatics/btp324. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.