

ABSTRACT
The consumption of sugar-free foods is growing because of their low-calorie content and the health concerns about products with high sugar content. Sweeteners that are frequently several hundred thousand times sweeter than sucrose are being consumed as sugar substitutes. Although nonnutritive sweeteners (NNSs) are considered safe and well tolerated, their effects on glucose intolerance, the activation of sweet taste receptors, and alterations to the composition of the intestinal microbiota are controversial. This review critically discusses the evidence supporting the effects of NNSs, both synthetic sweeteners (acesulfame K, aspartame, cyclamate, saccharin, neotame, advantame, and sucralose) and natural sweeteners (NSs; thaumatin, steviol glucosides, monellin, neohesperidin dihydrochalcone, and glycyrrhizin) and nutritive sweeteners (polyols or sugar alcohols) on the composition of microbiota in the human gut. So far, only saccharin and sucralose (NNSs) and stevia (NS) change the composition of the gut microbiota. By definition, a prebiotic is a nondigestible food ingredient, but some polyols can be absorbed, at least partially, in the small intestine by passive diffusion: however, a number of them, such as isomalt, maltitol, lactitol, and xylitol, can reach the large bowel and increase the numbers of bifidobacteria in humans. Further research on the effects of sweeteners on the composition of the human gut microbiome is necessary.
Keywords: nutritive sweeteners, nonnutritive sweeteners, sweetening agents, tabletop sweeteners, microbiota
Introduction
The consumption of sugars, mainly as sucrose and glucose-fructose syrups, has dramatically increased worldwide and growing concerns about their adverse effects on health and metabolic diseases, such as metabolic syndrome, cardiovascular diseases, and type 2 diabetes (T2D), have motivated people to reduce the consumption of free sugars. Sweeteners are sugar substitutes that mimic the sweet taste of sugar but have a negligible impact on energy intake (1, 2).
The sweetness of sweeteners is measured in relation to the reference sugar sucrose. Biologically, the perception of sweetness occurs through the receptors on the taste buds, which are coupled to G proteins [taste receptor types 1 and 2 (T1R1 and T1R2, respectively)] that form part of the C class of proteins (3).
Nonnutritive sweeteners (NNSs) are defined as sweetening agents that have a higher sweetening intensity and lower calorie content per gram compared with caloric or nutritive sweeteners such as sucrose or corn syrup. NNSs can be of synthetic or natural origin, the latter being increasingly consumed (4, 5). Low-calorie sweeteners (LCSs), such as polyols or sugar alcohols and other new sugars, are low-digestible carbohydrates derived from the hydrogenation of their sugar or syrup sources. Sugar alcohols are ∼25–100% as sweet as sugar. Sugar alcohols are slightly lower in calories than sugar and do not promote tooth decay or cause a sudden increase in blood glucose (6).
Both NNSs and LCSs are consumed not only by people with diabetes but also by the general population, because they are used as ingredients in many reduced-calorie foods such as soft drinks, dairy products, powdered drink mixes, baked goods, desserts, candy, chocolates, puddings, canned foods, jams and jellies, and confectionery chewing gums. In addition, they can be used as tabletop sweeteners at home, in cafeterias, and in restaurants (6).
The US FDA approval process for sweeteners includes determining the probable intake amounts, the cumulative effects of the sweetener from all of its uses, and toxicology studies in animals. In addition, in the European Union (EU), the European Food Safety Authority (EFSA), and Codex Alimentarius have evaluated and confirmed that NNSs and LCSs are safe for human consumption and do not cause cancer or other health-related problems as long as they are consumed within the Acceptable Daily Intake (ADI). To date, the FDA has approved 6 high-intensity artificial sweeteners for foods and drinks: acesulfame potassium (acesulfame K), aspartame, neotame, saccharin, sucralose, and advantame. In addition, 3 NNSs of natural origin—steviol glycosides, thaumatin, and luo han guo fruit extracts—have been approved by the FDA (6). The EU EFSA has approved 11 NNSs for human consumption: acesulfame K (E-950), advantame (E-969), aspartame (E-951), aspartame-acesulfame salt (E-962), cyclamic acid and its sodium and calcium salts (E-952), neohesperidin dihydrochalcone (E-959), neotame (E-961), saccharin (E-954), steviol glycosides (E-960, including 10 different glycosides), sucralose (E-955), and thaumatin (E-957) (7).
Food-use–approved polyols are low-calorie carbohydrates with a sweet taste used, volume-for-volume, as a substitute for sucrose and other free sugars. They include erythritol, hydrogenated starch hydrolysates (sometimes listed as maltitol syrup, hydrogenated glucose syrup, polyglycitol, polyglucitol, or simply HSH), isomalt, lactitol, maltitol, mannitol, sorbitol, and xylitol. In the United States, the FDA classifies some polyols as Generally Recognized As Safe, whereas others are approved food additives. The approved LCSs in the EU include the following: sorbitol and sorbitol syrup (E420), mannitol (E-421), isomalt (E-953), polyglycitol syrup (E-964), maltitol and maltitol syrup (E-965), lactitol (E-966), xylitol (E-967), and erythritol (E-968) (7).
Although the FDA, EFSA, Codex Alimentarius, and many national authorities have recognized that both NNSs and LCSs are generally safe and well tolerated, there is controversy about the effects of the sweeteners on human health (2). The consumption of NNSs, mainly in diet sodas, has been related to an increased risk of obesity, metabolic syndrome, and T2D (8–12), although some studies did not find any association (13, 14). The consumption of typically used nonnutritive artificial sweetener formulations drives the development of glucose intolerance through the induction of compositional and functional alterations to the intestinal microbiota (15). In contrast, the consumption of NNSs reduces blood glucose, which is attributed to the lower carbohydrate load rather than the activation of sweet taste receptors (16). In some people, the excessive consumption of polyols may cause gastrointestinal symptoms such as gas or laxative effects, similar to the gastrointestinal reaction to beans and certain high-fiber foods. Such symptoms depend on an individual's sensitivity and the other foods eaten at the same time (17).
Intestinal microbial communities play a significant role in human health and disease; indeed, the intestinal microbiome is involved in metabolism, immunity, growth, and the fermentation of undigested carbohydrates (18). More importantly, the gut microbiota cooperates with the immune system, providing signals to promote the maturation of immune cells and the induction of susceptibility to many pathophysiologic conditions (19). The composition and function of the microbiome are modulated and can be rapidly altered by diet (20). The importance of studying the microbiome as a potential link between NNS/nonnutritive artificial sweetener and LCS consumption and its effects on human health is currently being addressed because of the well-known interactions between human health, diet, and intestinal microbiota. However, there are many gaps in the evidence related to the health effects of NNSs and LCSs in both healthy and nonhealthy populations. Therefore, we critically reviewed the literature describing the impact of NNSs and LCSs on the gut microbiota.
Current Status of Knowledge
Effects of intensive sweeteners on the gut microbiota
Intensive sweeteners have negligible caloric content and high-power sweetening and are used in low quantities in foods. All of them have been classified in synthetic and natural sweeteners (5). Their structures and ADI, as well as their main biological effects, are summarized in Table 1.
TABLE 1.
Structure, ADI, and biological effects of natural and synthetic sweeteners1
| Sweetener | ADI, mg · kg−1 · d−1 | Structure | Biological effects |
|---|---|---|---|
| Acesulfame K (E-950) | 15 | C4H4KNO4S | Acesulfame K undergoes metabolization by the human body, which the majority of studies describe as innocuous. No effects on body weight or glucose tolerance. |
| Aspartame (E-951) | 40 | C14H18N2O5 | Aspartame, a combination of amino acids, namely l-phenylalanine and l-aspartic acid, and connected through methyl ester bonds, is rapidly absorbed. This compound is safe and without toxicity in gene mutations. |
| Neotame (E-961) | 2 | C20H30N2O5 | Neotame is a sweetener with a very similar structure to aspartame. It is safe for patients with phenylketonuria, but also safe for diabetics. With regard to its metabolization, half of the ingested neotame is not absorbed and excreted through the feces, whereas the other half is excreted in the urine as de-esterified neotame. |
| Advantame (E-969) | 5 | C24H30N2O7 | Advantame is obtained through chemical synthesis from aspartame and isovanillin and is a source of phenylalanine. This compound is nontoxic or carcinogenic and there are no risks of its consumption as a food additive. |
| Cyclamate (E-952) | 11 | C6H12NNaO3S | Cyclamate is prepared by the sulfonation of cyclohexylamine (toxic compound). The EU has approved its use in food, although the FDA removed its GRAS status in 1969 and completely banned it in 1970. No effects on body weight or glucose tolerance. |
| Saccharin (E-954) | 5 | C7H5NO3S | Saccharin is excreted through urine and is not metabolized in the body, although it can cross the placenta and can be transferred through breast milk. Its consumption is not recommended for pregnant or breastfeeding women. |
| Sucralose (E-955) | 5 | C12H22O11 | Sucralose is obtained by substitution of the 3-hydroxyl groups in sucrose. Approximately 11–27% of ingested sucralose is absorbed from the gut and is excreted in the kidneys. Sucralose is safe. |
| Steviol glucosides (E-960) | 4 | Variable | Steviol glycosides are molecules extracted from the leaves of Stevia rebaudiana Bertoni. Colonic bacteria converts them into steviol glucoronides to finally be excreted through urine. The consumption of these molecules is safe. |
| Glycyrrhizin | NA | C42H62O16 | Glycyrrhizin is a triterpenoid saponin that is obtained from the roots and rhizome of Glycyrrhiza glabra. In the EU, its consumption is considered safe with a limit of 100 mg/d, given the glucocorticoid effects in the glycyrrhetinic acid present in the extract. |
| Neohesperidine dihydrochalcone (E-959) | 4 | C28H36O15 | Neohesperidin dihydrochalcone is a seminatural sweetener that comes from the skin of the immature fruits of Citrus aurantium L. Approved in the EU since 1994 but not in the United States. |
| Thaumatin (E-957) | 50 | — | Thaumatin is a mixture of compounds extracted from the Thaumatococcus danielli Bennett plant. As a sweetener, it is approved both in the EU and the United States, where it is considered GRAS. |
ADI, Acceptable Daily Intake; EU, European Union; GRAS, Generally Recognized As Safe; NA, not available.
Synthetic sweeteners
Acesulfame K
Acesulfame is an acidic cyclic sulfonamide and acesulfame K (E-950) is the potassium salt of acesulfame. Acesulfame K is metabolized by the human body and has an ADI of 15 mg/kg body weight (5, 21).
Acesulfame K decreases glucose fermentation by the cecal microbiota in Cara rats, suggesting that sweeteners might affect glucose transport systems (22). The effects of acesulfame K were not associated with gut microbial functional capability (23).
A study in mice that received distilled water and 15 mg acesulfame K/kg showed that the total bacteria, Firmicutes, Bacteroidetes, and several other genera were similar between the 2 groups, establishing that the consumption of acesulfame K had few effects on gut microbiota and their metabolism in mice (24). In contrast, Bian et al. (25) found the opposite; consuming acesulfame K for 4 wk perturbed the gut microbiota of CD-1 mice. Bacteroides were highly increased in acesulfame K–treated male mice and significant changes in Anaerostipes and Sutterella populations occurred as well. Conversely, in female mice, acesulfame K treatment decreased the relative abundance of Lactobacillus and Clostridium. Those changes in the populations of gut microbiota after the consumption of acesulfame K indicate sex-specific effects (25).
The principal reason for these contradictory results is likely related to the acesulfame K dose administered in each study; in the first study, a dose of 15 body weight mg · kg−1 · d−1 was used (24), and in the second study, a dose of 37.5 body weight mg · kg−1 · d−1 was used (25). With regard to human consumption, the Uebanso et al. (24) study used the maximum ADI level, whereas the Bian et al. (25) study exceeded by more than twice the ADI recommendation. Indeed, this work might be physiologically irrelevant (25).
Aspartame
Aspartame (E-951) is a dipeptide consisting of aspartic acid and phenylalanine, with the carboxyl terminal group of the latter being methylated (N-l-α-aspartyl-l-phenylalanine 1-methyl ester). It is ∼200 times sweeter than sucrose. The metabolism and fate of aspartame are dominated by presystemic hydrolysis to the constituent parts, with little or no parent compound entering the general circulation. According to EU regulation no. 1169/2011, all food that uses aspartame has to have a visible label containing the words “contains aspartame (source of phenylalanine).” The ADI for aspartame is 40 mg/kg body weight (5).
A 400-mg dose of aspartame did not affect the peak insulin concentrations in subjects with or without diabetes but did cause a decrease in plasma glucose concentrations (26). Tordoff and Alleva (27) compared the consumption of aspartame and high-fructose corn syrup and concluded that aspartame reduces sugar intake. Although we have a huge quantity of information with regard to aspartame safety in humans, few of those studies focused on the effects of aspartame intake on the composition of gut microbiota.
In rats, the impact of chronic low-dose aspartame consumption on anthropometric, metabolic, and microbial variables was tested in a diet-induced obesity model. The rats were randomly divided into 4 groups that received the following for 8 wk: a standard feed pellet–diet group (12% of kilocalories from fat) with ad libitum water or 5–7 mg aspartame · kg body weight−1. d−1 in drinking water and a high-fat-diet group (60% of kilocalories from fat) with ad libitum water or 5–7 mg · kg body weight−1 · d−1 in drinking water. Aspartame consumption increased the fasting glucose concentrations in both the standard feed pellet diet and high-fat groups independent of body composition. A metabolomics analysis showed that aspartame was rapidly metabolized and related to SCFA production, especially propionate production. Changes in the microbial composition were observed in animals that received aspartame; the total bacteria and abundance of Enterobacteriaceae and Clostridium leptum increased (28). In addition, mice treated with aspartame for 11 wk developed glucose intolerance, although analyses of the microbiota did not show significant differences between the groups (15).
To our knowledge, there are no data on the potential influences of aspartame on the human gut microbiome. It is hard to understand how aspartame influences the gut microbiota because this NNS is rapidly hydrolyzed in the small intestine. In fact, even with the ingestion of very high doses of aspartame (>200 mg/kg), no aspartame is found in the blood because of its rapid breakdown (29). Upon ingestion, aspartame breaks down into residual components, including aspartic acid, phenylalanine, and methanol and their components, which are readily absorbed so that they do not reach the large bowel (30).
Neotame and advantame
Neotame (E-961) is an artificial sweetener that is between 7000 and 13,000 times sweeter than sucrose with a structure close to that of aspartame [i.e., N-N-(3,3-dimethylbutyl-l-α-aspartyl-l-phenylalanine 1-methyl ester)]. The FDA and EFSA have approved neotame for general use. The suggested ADI is 0.3 mg ⋅ kg body weight−1 ⋅ d−1. Neotame is moderately heat stable, extremely potent, rapidly metabolized, completely eliminated, and does not appear to accumulate in the body. Mice and other test animals fed neotame did not show adverse physical symptoms, water consumption, or clinical pathology evaluations and there were no reports of morbidity, mortality, organ toxicity, or macroscopic or microscopic postmortem findings (31–33).
Advantame (E-969), approved in 2013 by the EU, is an N-substituted derivative of aspartame made from aspartame and vanillin and is ∼20,000 times sweeter than sucrose (34). In 2013, the EFSA panel established an ADI of 5 mg ⋅ kg body weight−1 ⋅ d−1 and recognized this sweetener as nontoxic, noncarcinogenic, and safe for consumption as a food additive (7).
Neither sweetener has been evaluated in animals or in humans because only trace amounts of advantame or neotame are needed to sweeten foods. The amount of methanol derived from the intestinal hydrolysis of neotame is much lower than that found in common foods; therefore, it is improbable that either neotame or advantame would have any influence on the gut microbiota.
Cyclamate
Cyclamate is used in >50 countries; the EU approved cyclamic acid and its sodium and calcium salts for food use (E952), whereas the FDA removed its Generally Recognized As Safe status in 1969 and it was completely banned in 1970 (5). This was because of the detection of bladder tumors in rats fed a cyclamate-saccharin mixture supplemented with cyclohexylamine, a metabolite of cyclamate that is more toxic than cyclamate alone (35, 36). However, these studies were severely criticized because of their designs and doses (37) and cyclamate is being reevaluated. In the EU, the ADI for cyclamate is 7 mg/kg body weight (5, 7).
The first finding of microbiota changes caused by cyclamate was reported in the study by Drasar et al. (38). The authors observed that the conversion of cyclamate to cyclohexylamine in rats does not occur after either parenteral administration of cyclamate or with incubations of cyclamate with the liver, spleen, kidney, or blood preparations. The principal hypothesis was that cyclohexylamine formation occurred solely in the gut as the result of microorganism metabolism (38).
In 1985, Mallett et al. (39) tested the metabolic cyclamate adaptation on rat gut microbiota maintained in vitro in an anaerobic culture system. They found a maximum formation of cycloheximide at 8 wk and increased levels of sulfamatase activity in the fecal content. The authors did not find any taxonomic changes in the fecal microbiota cultured in an in vitro system after the administration of cyclamate.
The presence of cyclamate decreased the fermentation of glucose by the microbiota in Cara rats (22). Cyclamate increases the bacterial sulfatase activity in the intestine (40). To our knowledge, there are no available data on the effects of cyclamate on gut microbiota in humans.
Saccharin
A range of food and beverages are sweetened by saccharin (E-954), which is considered safe despite controversial debate about its potential carcinogenicity. However, studies indicate that the consumption of saccharin might perturb the gut microbiota. Its ADI is the lowest of all the intensive sweeteners (5 mg/kg body weight) (5).
The effect of 7.5% saccharin on aerobic and anaerobic microbial populations from rat cecums over 10 d was tested. The rats consumed ∼90 mg saccharin, which was detected in the cecal contents at the end of the intervention. The presence of saccharin did not alter the total numbers of anaerobic microbes but resulted in the elimination of a specific anaerobic group of microbes from the cecal contents (41).
Saccharin administration inhibited the growth of 6 bacterial strains (3 Lactobacillus species and 3 Escherichia coli strains) isolated from the small intestinal contents in rats that received a 2.5% dose of saccharin; the rats consumed 107.0 mg saccharin in the diet (rat weights were between 200 and 220 g) (42). Saccharin inhibited the fermentation of glucose by the microbiota from Cara rats (22).
Pyrosequencing studies in animals showed that the addition of saccharin plus neohesperidin dihydrochalcone increases the abundance of the Lactobacillus cecal population and increases intraluminal lactic acid concentrations (43). 16S ribosomal RNA gene analyses identified 25 major families encompassing 7 bacterial classes with Bacteroidia, Clostridium, and Bacilli dominating the microbiota. In animals that received saccharin/neohesperidin dihydrochalcone, there were significant shifts in microbial composition, establishing a major influence driving bacterial community dynamics (44).
The deleterious metabolic effects of saccharin in animals were abrogated by antibiotic treatment and were fully transferrable to germ-free mice upon microbiota transplantation. In addition, the altered metabolic pathways were linked with glucose tolerance and dysbiosis in healthy human subjects. In mice fed saccharin, Akkermansia muciniphila, a commensal bacterium that exhibits probiotic properties, was underrepresented (15). Since the study by Suez et al. (15) the scientific focus has moved toward evaluating the impact of saccharin on gut microbiota diversity.
Sweeteners are often used to encourage the consumption of agents such as ethanol and nicotine in laboratory studies that use rodents. Labrecque et al. (45) evaluated the effect of ethanol in either water or saccharin on the fecal microbiome in pregnant and nonpregnant mice. Saccharin reduced Clostridium numbers, even though the total amounts of ethanol consumed were the same for the 2 groups (45).
Inflammation is frequently associated with disruptions to the gut microbiota. Mice treated with 0.3 mg saccharin/mL (a dose equivalent to the FDA-approved ADI for humans) for 6 mo had increased expression of TNF-α and the inducible isoform of NO synthase (iNOS) in their livers. In addition, altered gut bacterial genera were associated with saccharin-induced liver inflammation. These changes in the intestinal microbiota were observed in Ruminococcus, Adlercreutzia, Dorea, Corynebacterium, Roseburia, and Turicibacter (46).
Early studies suggest that artificial sweeteners maintain plasma glucose and peak insulin concentrations without affecting the gut microbiota. However, more recent animal and human studies showed specific changes in the intestinal microbiota related to alterations in the metabolic pathways linked to glucose tolerance and dysbiosis in human subjects, especially with the ingestion of saccharin (Figure 1).
FIGURE 1.
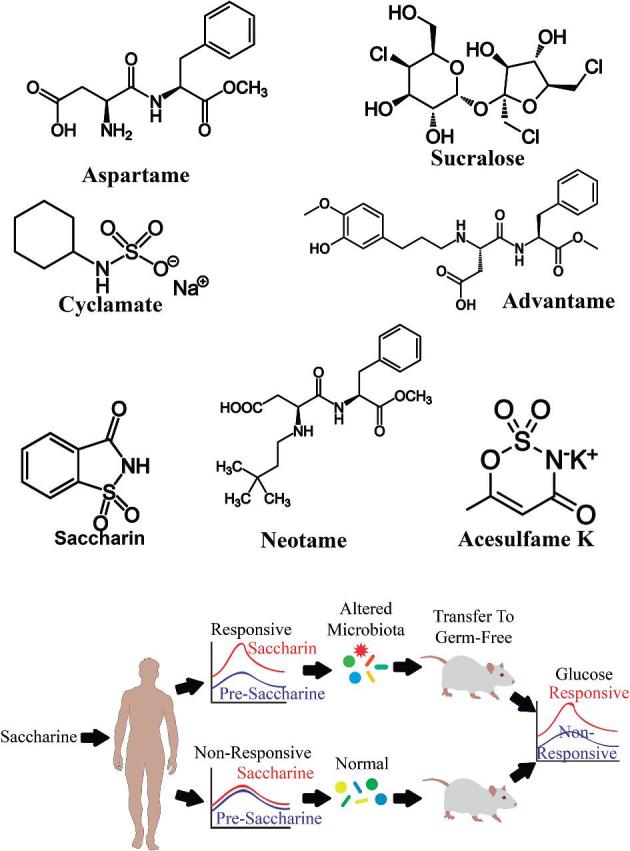
Effects of artificial sweeteners and saccharin on gut microbiota. Animal studies have reported specific shifts in the intestinal microbiota related to alterations in the metabolic pathways linked to glucose tolerance after ingestion of saccharin.
Sucralose
Sucralose (E-955) is a synthetic sweetener derived by the substitution of the 3 hydroxyl groups in sucrose and is ∼320–1000 times sweeter than sucrose (47). Its ADI is 5 mg/kg body weight. The first study that evaluated sucralose on the intestinal microbiota was performed in 2008 with the use of fecal samples from Sprague-Dawley rats that received the sweetener for 12 wk. The consumption of sucralose decreased the total number of anaerobic and aerobic bacteria, bifidobacteria, lactobacilli, Bacteroides, and Clostridium (48). The administration of 15 mg sucralose/kg affected the relative abundance of the Clostridium cluster XIVa in mice (49).
More recently, the administration of sucralose in mice produced modifications in the intestinal microbiota at 14 different taxonomic levels, including Turicibacteraceae, Lachnospiraceae, Ruminococcaceae, Verrucomicrobiaceae, Staphylococcaceae, Streptococcaceae, Dehalobacteriaceae, Dehalobacterium, Lachnospiraceae, Clostridiaceae, Christensenellaceae, Peptostreptococcaceae, Erysipelotrichaceae, and the order Bacillales, and changes in the synthesis and regulation of amino acids. These variations were related to inflammation in the host (50). The main reported effects of synthetic sweeteners on the gut microbiota are listed in Table 2.
TABLE 2.
Effects of synthetic sweeteners on gut microbiota1
| Sweetener and study (reference) | Model | Dose tested | Method of microbial analysis | Main outcomes | Magnitude of change |
|---|---|---|---|---|---|
| Acesulfame K (E-950) | |||||
| Pfeffer et al. (22) | Rats | 3% acesulfame | Inhibitory activity in cecal content | Acesulfame K might act on glucose transport systems. | Marginally inhibited |
| Frankenfeld et al. (23) | Human trial | 1.7–33.2 mg · kg−1 · d−1 | 16S rRNA | Consumption was not associated with the functional capability of the gut microbiota. | Reduction in bacterial diversity from 24 to 7 phyla |
| Uebanso et al. (24) | Mice | 15 mg · kg body weight−1 · d−1 | PCR denaturing gradient gel electrophoresis | Scarce effects on the gut microbiota and its metabolism. | Marginal changes |
| Bian et al. (25) | Mice | 37.5 mg · kg body weight−1 · d−1 | 16S rRNA and GC | The population of Bacteroides was highly increased in acesulfame K–treated male mice, with significant changes in the Anaerostipes and Sutterella populations. Conversely, in female mice, acesulfame K decreased the Lactobacillus and Clostridium populations. | The bacterial genera increased or decreased more than twice |
| Aspartame (E-951) | |||||
| Horwitz et al. (26) | Human trial | 400 mg | Ingestion and analysis of AUC | Plasma glucose declined and the peak insulin concentrations in subjects treated with aspartame; no effects on gut microbiota. | No changes |
| Tordoff and Alleva (27) | Human trial | 590 mg | Ingestion and dietary record | Aspartame reduced sugar intake; no effects on gut microbiota. | No changes |
| Palmnäs et al. (28) | Rats | 60 mg/L drinking water | qRT-PCR analysis | Increased numbers of Enterobacteriaceae and Clostridium leptum. | More than 10% increase |
| Suez et al. (15) | Mice | 4% aspartame | 16S rRNA | No change in the intestinal microbiota. | No changes |
| Cyclamate (E-952) | |||||
| Drasar et al. (38) | Rats | 100 mg calcium cyclamate | 14C-analysis | No effects on the gut microbiota. | No changes |
| Mallett et al. (39) | In vitro | 25–75% cyclamate concentration in medium | Two-stage continuous culture system | No effects on the gut microbiota. | No changes |
| Pfeffer et al. (22) | In vitro | 5% cyclamate | Inhibitory activity in cecal content | Cyclamate decreased glucose fermentation. | Marginally inhibited |
| Saccharin (E-954) | |||||
| Anderson et al. (41) | Rats | 7.5% sodium saccharin | Enzymatic activity and microbiology analyses | Saccharin did not alter the total numbers of anaerobic microbes but deleted a specific anaerobic microbe in the cecal contents. | Marginally inhibited |
| Naim et al. (42) | Rats | 2.5% sodium saccharin | Enzymatic activity and microbiology analyses | Saccharin inhibited the growth of 3 Lactobacillus strains and 3 Escherichia coli strains. | Almost 40% of growth inhibition |
| Pfeffer et al. (22) | In vitro | 0.5% saccharin | Inhibitory activity in cecal content | Saccharin inhibited glucose fermentation by the gut microbiota in Cara rats. | Marginally inhibited |
| Daly et al. (43) | Piglets | 0.015% (wt:wt) saccharin and neohesperidin dihydrochalcone | 16S rRNA | Neohesperidin dihydrochalcone/saccharin increased the cecal populations of Lactobacillus and the intraluminal lactic acid concentration. | Increased by 3 times the lactobacilli population |
| Daly et al. (44) | Piglets | 0.015% (wt:wt) saccharin and neohesperidin dihydrochalcone | 16S rRNA | Saccharin caused significant shifts in microbial composition. | Increased lactobacilli twice and decreased Ruminococceae and Veillonellaceae by almost 50% |
| Suez et al. (15) | Mice/human trial | 120 mg saccharin | 16S rRNA | Alterations in metabolic pathways linked to glucose tolerance and dysbiosis in healthy human subjects. | The magnitude of the difference was >30% |
| Labrecque et al. (45) | Mice | 0.066% (wt:vol) saccharin in water | qRT-PCR analysis | Eubacteria were increased in the pregnant group that received ethanol plus saccharin and the presence of saccharin reduced Clostridium counts. | Reduction in Clostridium was almost 50% |
| Bian et al. (46) | Mice | 0.3 mg/mL in drinking water | 16S rRNA | Altered gut bacterial genera were associated with the saccharin-induced liver inflammation. | iNOS and TNF-α increased by 3 and 2 times, respectively. Intestinal microbiota changes were observed in Ruminococcus, Adlercreutzia, Dorea, Corynebacterium, Roseburia, and Turicibacter, increasing by more than twice |
| Neotame (E-961) | — | — | — | No effects on gut microbiota. | No changes |
| Advantame (E-969) | — | — | — | No effects on gut microbiota. | No changes |
| Sucralose (E-955) | |||||
| Abou-Donia et al. (48) | Rats | 100, 300, 500, or 1000 mg/kg | Bacteriological analyses | The consumption of sucralose decreased the total anaerobes and aerobic bacteria, bifidobacteria, lactobacilli, Bacteroides, and Clostridium. | The decrease was >2-fold |
| Uebanso et al. (49) | Mice | 15 mg · kg body weight−1 · d−1 | PCR denaturing gradient gel electrophoresis | Sucralose administration produced modifications in Clostridium cluster XIVa. | The inhibition was >50% |
iNOS, inducible NO synthase; 16S rRNA, 16S ribosomal RNA.
Effects of natural sweeteners on the gut microbiota
Natural sweeteners are sweeter than sucrose, contribute few calories, have no carcinogenic effects, and do not affect insulin production (5).
Steviol glucosides
Stevia rebaudiana is a shrub belonging to the family Ateracea (native to South America), whose leaves contain diterpene glycosides such as stevioside, steviolbioside, rubusoside, dulcoside A, and rebaudiosides A, B, C, D, E, F, and M. Its extracts are used as natural noncaloric sweeteners because it is 250 times sweeter than sucrose (51), although only highly purified steviol glycosides are approved for use in food in the EU (7).
Stevioside extracts from S. rebaudiana are not carcinogenic in the adult population (52). Steviol glycosides are sweet, low in calories, and noncarcinogenic, but consuming more than the ADI limit of 4 mg ⋅ kg body weight−1 ⋅ d−1 is unsafe (EU regulation 1129/2011) (53, 54).
Several in vitro studies have investigated how the components of stevia extract are metabolized. The data show that the microbiota (no differences found between humans and rats) are able to degrade the main components, stevioside and rebaudioside A, to steviol (55, 56). Therefore, neither stevioside nor rebaudioside A is absorbed in the upper gastrointestinal tract (30).
Bacteroides are the most efficient group of bacteria at hydrolyzing stevioside and rebaudioside A to steviol (56). Other bacterial groups, such as lactobacilli, bifidobacteria, Clostridium, coliforms, and enterococci species, were tested. None of the tested bacteria were able to hydrolyze and use steviol glycosides as a usable substrate (56). These tested bacterial groups are the major types of bacteria found in the gastrointestinal tracts of animals and humans (57).
In addition, compared with glucose, 24 h incubation of mixed fecal bacteria from volunteers with stevioside and rebaudioside A caused a slight alteration of the human microbiota (56). Stevioside weakly inhibits anaerobic bacteria, whereas rebaudioside A weakly inhibits aerobic bacteria, in particular over coliforms.
The roots of S. rebaudiana contain inulin and fructans, functional food ingredients that have a positive effect on human health (30). The fermentation capacity of fructans as a substrate for microbiota is strain specific. Fructans derived from S. rebaudiana, especially those with a polymerization degree of <6 (carbohydrates with different-size chain), improved the growth of select microbial strains (bifidobacteria and lactobacilli) that are important for bowel function (58).
Glycyrrhizin
Glycyrrhizin comes from the roots and rhizome of Glycyrrhiza glabra. It is 30–200 times sweeter than sucrose and is considered safe if <100 mg/d is ingested. Glycyrrhizin has anticancer, anti-inflammatory, antioxidant, antiviral, and hepatoprotective properties. However, it has potential hypertensive effects and an intense aftertaste (59).
In the gut, glycyrrhizin is de-glycosylated to glycyrrhetic acid (the major product) by Eubacterium spp. and Bacteroides J-37 and to 18β-glycyrrhetic acid 3-O-monoglucuronide (the minor product) by Bacteroides J-37 and Streptococcus LJ-22. The conversion of 18β-glycyrrhetic acid 3-O-monoglucuronide to glycyrrhetic acid can also be mediated by Eubacterium spp. (59–61). These glycyrrhizin metabolites (especially 18β-glycyrrhetinic acid) are potent cytotoxicity agents against tumor cells and they exert potent inhibitory effects on rotavirus infection and antiplatelet aggregation activity (62).
Some data suggest that the relation between glycyrrhizin and the intestinal microbiota exerts positive effects on the host (60, 61, 63). Better-designed studies are needed to determine if this is truly the case and what the implications of its metabolism and its mechanism of action and effects are on the composition of the intestinal microbiota.
Neohesperidin dihydrochalcone, thaumatin, and monellin
Neohesperidin dihydrochalcone is a natural sweetener obtained from the skin of immature citrus fruits and is only ∼1500 times sweeter than sucrose. Neohesperidin dihydrochalcone is metabolized by intestinal microbiota to innocuous products (5, 53). Thaumatin is a sweet protein isolated from the fruit of Thaumatococcus daniellii Benth, a plant native to tropical West Africa. Thaumatin is 100,000 times sweeter than sucrose (5). Monellin is a sweet protein, naturally extracted from the fruit of the serendipity berry shrub (64). To our knowledge, there are no ongoing or past studies ascertaining the potential effects of those natural sweeteners on the intestinal microbiota.
In summary, natural sweeteners have only a few studies associating their consumption with changes in the intestinal microbiota. Stevia extracts have the most information with regard to their effects on the gut microbiota composition, although the current effects of stevia on Bacteroides need further study (Figure 2).
FIGURE 2.

Natural sweeteners and their effects on gut microbiota. Stevia extracts have been described as capable of changing the gut microbiota composition, although the current effects of stevia on Bacteroides need further study.
Effects of nutritive LCSs on the gut microbiota
Polyols
Polyols are a specific group of compounds used as food additives. They are stable at high temperatures and through pH changes and do not intervene in Maillard reactions. A number of polyols are naturally present in some fruits, vegetables, and mushrooms. Their industrial production started in the last century with the hope of solving health problems related to excessively consumed NNSs. Polyols are noncariogenic, do not induce salivation, and do not interfere with insulin concentrations or increase the blood glucose response; therefore, they are used in “light” foods. The FDA, Codex Alimentarius, and EFSA have approved 8 different polyols—erythritol, hydrogenated starch hydrolysates, isomalt, lactitol, maltitol, mannitol, sorbitol, and xylitol—for use as bulk sweeteners in human foods (5, 7, 65, 66).
The excessive consumption of polyols causes gastrointestinal symptoms and laxative effects in healthy patients. Polyols also induce dose-dependent symptoms of flatulence, abdominal discomfort, and laxative effects when consumed by both healthy volunteers and patients with irritable bowel syndrome (IBS). In addition, moderate doses of polyols increase the number of bifidobacteria in the microbiomes of healthy individuals and may therefore be beneficial as a prebiotic, but the data are limited to patients with a number of intestinal diseases, including IBS (66). It is important to know the impact of polyol consumption on gut microbiota both in healthy and diseased humans.
As with all food additives, the safety of polyols is being re-evaluated by the EFSA. In June 2017, the EFSA called for technical and toxicologic data on sweeteners authorized as food additives in the EU. This re-evaluation will be completed by the end 2020 (67).
Erythritol
Erythritol (E-968), a four-carbon sugar alcohol, occurs widely in nature and in foods such as wine, beer, mushrooms, pears, grapes, and soy sauce (68). Industrially, it is produced from glucose by an osmophilic yeast and subsequently, by separation and purification, yields a crystalline product with a purity of 99% (69). The estimated daily intake of erythritol is 1.24 g (53). Oral microorganisms do not metabolize erythritol and the in vitro incubation of erythritol with Streptococci species does not produce lactic acid or other organic acids (70, 71).
Erythritol is rapidly absorbed in the small intestine by passive diffusion, it is scattered widely through tissues with minimum metabolism, and finally, it is quantitatively excreted in the urine (68). Hence, erythritol does not affect plasma glucose or insulin concentrations or gut microbiota (72, 73). Despite the adjustment and consideration of all the fermentation variables (e.g., gas production, hydrogen accumulation, pH, SCFA production, and substrate degradation), erythritol is completely nonfermentable by freshly collected human fecal microbiota within a period of 24 h (71). Although there is no evidence on the effects of erythritol on gut microbiota in humans in clinical trials, erythritol is considered a safe additive after many specific tests on its toxicity, carcinogenicity, and reproductive hazards were found to be negative (5).
Isomalt
Hydrogenated isomaltose, isomaltitol, or isomalt (E-953) is a polyol used worldwide as a sugar replacement with technological properties comparable to those of sucrose. Isomalt is used in bubble gums, gelatins, chocolate, coatings, baked goods, and yogurts. Isomalt is obtained through the enzymatic transformation of sucrose, is stable at high temperatures, and has a very low hygroscopic value. Moreover, it is low in energy, noncariogenic, and is as sweet as other polyols. Undigested or unabsorbed portions of isomalt reach the colon and are fermented by the gut microbiota; the fermented fraction of ingested isomalt is ∼90% (5, 17, 74, 75). Isomalt is considered a prebiotic carbohydrate that might contribute to a healthy luminal colonic mucosal environment. During 4-wk periods in a double-blind, placebo-controlled, crossover-design clinical trial, 19 healthy volunteers consumed a basal diet enriched with either 30 g isomalt or 30 g sucrose/d and found that isomalt has beneficial effects on the gut microbiota (75). Later, it was reported that isomalt fermented in the gut increased bifidobacteria and decreased bacterial β-glucosidase, whereas β-glucuronidase, sulfatase, nitroreductase, and urease remained unchanged. Fecal SCFAs, lactate, bile acids, neutral sterols, nitrogen, ammonia, phenol, and p-cresol were not affected by isomalt consumption. In addition, in vitro, several bifidobacteria strains were capable of metabolizing the isomalt and generated high butyrate concentrations (76). Moreover, at the end of each test phase, rectal biopsy samples were taken and gene expression was measured by microarray and qRT-PCR. Dietary intervention with low digestible isomalt compared with digestible sucrose did not affect gene expression in the rectal mucosa lining (77). Hence, isomalt is a polyol with bifidogenic properties that might contribute to a healthy colonic environment.
Lactitol
Lactitol (E-966) is a non–naturally occurring sugar alcohol, which is obtained by the hydrogenation of lactose. Compared with the other polyols, its sweetening power is limited and consequently it is usually used in combination with intense sweeteners (5). This disaccharide is normally not absorbed in the small intestine (78) because of the lack of a suitable β-galactosidase and, as a result, it reaches the lower gut where it is fermented (79). Therefore, it makes a caloric contribution of <2% (31). However, higher lactitol intakes can produce laxative effects; studies showed that lactitol promotes the growth of bifidobacteria and lactobacilli. On the other hand, fermentation of lactitol by saccharolytic bacteria has also been shown to decrease counts of proteolytic bacteria, such as Bacteroides, coliforms, Enterobacteria, and Enterococci (80). In vitro, lactitol (2 mg/L) reduces the Enterobacteriaceae population in feline fecal cultures and exerts a prebiotic effect on feline intestinal microbiota (81). Ballongue et al. (82) showed that lactitol treatment over 9 wk decreased bacterial populations of Bacteroides, Clostridium, coliforms, and Eubacterium. Lactitol also causes a decrease in fecal pH (82). In rats, lactitol increases the production of butyrate and IgA secretion without signs of mucosal inflammation (83). In the elderly, the consumption of lactitol as a synbiotic, combined with Lactobacillus acidophilus NCFM, improves some intestinal microbiota markers and mucosal functions (84). In addition, the synbiotic products of L. acidophilus NCFM and lactitol decrease the abundance of the Blautia coccoides–Eubacterium rectale bacterial group and Clostridium cluster XIVab counts (85). Another study by Finney et al. (86) showed that low doses of lactitol consumed as a sweetener (10 g) beneficially affect the fecal microbiota, increasing bifidobacteria and concentrations of the volatile propionic and butyric acids, without eliciting gross symptoms of intolerance; it is a prebiotic (86). These data suggest that lactitol is useful as a prebiotic for enhancing the gut microbiota, is noncariogenic, and is of mild sweetness. For many, compared with lactulose or other sweeteners, the low sweetness of lactitol is an advantage of lactitol supplementation compared with others such as lactulose (80).
Maltitol
Maltitol (E965) is obtained by the hydrolysis, reduction, and hydrogenation of starch, resulting in a sweetener with ∼90% sweetening capacity. Maltitol most resembles the flavor of sugar, is noncariogenic, and safe for diabetics. It has a similar solubility and hygroscopicity to sucrose and is the preferred sugar for use in the production of no-sugar-added–labeled chocolate (5, 87). Maltitol has a very slow digestion rate because it is fermented in the colon. Therefore, it is expected that it could be fermented by the gut microbiota. In a human study, 40 volunteers consumed a test chocolate containing 22.8 g maltitol, maltitol plus polydextrose, or maltitol plus resistant starch for 14 consecutive days. The doses of the test chocolates were doubled every 2 wk over a 6-wk period. The authors evaluated the impact of confectionary sweeteners on the composition of gut microbiota and, at an optimal dose of 34.2 g for maltitol plus polydextrose, the numbers of fecal bifidobacteria, lactobacilli, and SCFAs significantly increased after the ingestion of maltitol compared with the ingestion of sucrose (88). However, to date, there are not enough data to determine the specific effects of maltitol on gut microbiota.
Sorbitol
Sorbitol (E-420), also known as d-glucitol, is an isomeric polyol whose production is based on the catalytic hydrogenation of glucose with subsequent purification. Sorbitol is found naturally in apples, pears, peaches, apricots, and some vegetables (65); and although there is no evidence of sorbitol toxicity, a possible association of sorbitol with genotoxicity and shifts in metabolism in rats fed sorbitol has been reported. In healthy people, 71% show malabsorption after the consumption of 10 g sorbitol and 20% had gastrointestinal symptoms (89). In addition, patients with IBS have adverse gastrointestinal reactions to polyols, especially sorbitol and mannitol (independent of the absorption patterns of each molecule). Although sorbitol can be of concern for patients with IBS, it seems to be safe for healthy individuals, although there are reports of laxative effects when consumed in high doses (5, 90). Most healthy individuals tolerate ∼10 g sorbitol/d with only mild gastrointestinal discomfort, such as flatulence or bloating. However, doses of 20 g sorbitol/d can evoke more distressing symptoms of abdominal pain and diarrhea (66, 91). Sorbitol is usually less tolerated than lactitol because sorbitol exerts a greater osmotic load in the gastrointestinal tract, leading to an increased concentration of water in the colon and, consequently, greater laxative effects (92, 93). However, to date, there are not enough data to definitively determine the effects of sorbitol on gut microbiota.
Mannitol
Mannitol (E-421) is an isomer of sorbitol and is obtained from the hydrogenation of glucose and its consequent purification. Although less sweet than sorbitol, mannitol is also used in food because it has a high metabolization ratio (the ratio of a drug metabolite concentration to its parent drug concentration, expressed as a range) of ∼75%; the other 25% is absorbed before being excreted in the urine. Because it is virtually inert (i.e., it does not react with active components of drugs) and confers a cool sweet taste, apart from being used in the food industry it is also widely used in dental hygiene products, drug fillers, and as a diuretic in intravenous fluids (5, 17, 31, 94, 95). To our knowledge, no data on the effects of mannitol on the gut microbiota are available.
Xylitol
Xylitol (E-967), a five-carbon polyol, obtained by the hydrogenation of d-xylose is naturally found in fruits, berries, vegetables, oats, and mushrooms and a small percentage is also produced by the human body. Xylitol is widely used in various pharmaceutical products in addition to sugar-free candies and chewing gums. Xylitol was first synthesized in 1891 and is ∼95% as sweet as sucrose (5, 66).
The effects of intakes of 40 and 200 mg xylitol · kg body weight−1 · d−1 on the composition of gut microbiota and lipid metabolism in mice have been reported. Xylitol reduced the abundance of fecal Bacteroidetes and Barnesiella and increased the abundance of Firmicutes and Prevotella in mice fed a high-fat diet with medium-dose dietary xylitol (49). Because xylitol is capable of modifying the gut microbiota in mice, a study by Tamura et al. (96) compared the fecal microbiome of mice after being fed either a 0.05% isoflavone and 5% xylitol diet or a 0.05% isoflavone-only diet as a control. The Bacteroides concentration was higher in the control diet than in the xylitol-rich diet (96). In addition, xylitol ingestion shifted the rodent fecal microbiome population from Gram-negative to Gram-positive bacteria. In human volunteers, a similar shift occurs after a single 30-g oral dose of xylitol (97). The effect of soluble low-digestible carbohydrates, including xylitol, on butyrate production and the prebiotic potential of these substances has been assessed using in vitro human fecal cultures. l-Sorbose and xylitol cause prebiotic stimulation of the growth and metabolic activity of Anaerostipes spp. in the human colon (98). The correlation between bacterial translocation with morphologic changes in the intestinal mucosa and shifts of the intestinal microbiota in hamsters with Clostridium difficile infection show that the combination of lactobacilli (probiotic) and xylitol (prebiotic) had a protective effect against C. difficile infection (99).
To conclude, although some polyols, such as erythritol, sorbitol, and mannitol, do not affect the composition of gut microbiota, moderate doses of polyols could induce shifts in the gut microbiome in healthy people. The laxative effects of polyols need to be considered when they are consumed by patients with inflammatory bowel disease. The effects of polyols are described in Table 3. In addition, Figure 3 summarizes the effects of polyols on gut microbiota.
TABLE 3.
Effects of polyols on gut microbiota1
| Sweetener and study (reference) | Sources | Fermented fraction | Model | Dose tested | Method of microbial analysis | Main outcomes | Adverse effects | Magnitude of change |
|---|---|---|---|---|---|---|---|---|
| Erythritol (E-968) | Wine, beer, mushrooms, pears, grapes, and soy sauce | 10% reaches the colon | — | — | — | — | — | — |
| Arrigoni et al. (71) | — | — | In vitro | NA | pH, total gas, H2, and SCFA production in feces | Erythritol is completely nonfermentable | None observed | No changes |
| Isomalt (E-953) | Bubble gums, gelatins, chocolate, coatings, baked goods, and yogurts | 90% reaches the colon | — | — | — | — | — | — |
| Gostner et al. (75) | — | — | Human trial | 30 g isomalt | 16S/23S rRNA | Increased populations of bifidobacteria, decreased bacterial β-glucosidase and fecal SCFAs | None observed | 0.2–0.3 log cells/g feces |
| Lactitol (E-966) | A nonnaturally occurring sugar alcohol obtained by the hydrogenation of lactose | Not absorbed in the small intestine because of a lack of β-galactosidase | — | — | — | — | — | — |
| Patil et al. (78) | — | — | RCT | 10, 30, 60, and 100 mmol lactitol/d; 70–130 g/d | Galactose content (galactose dehydrogenase) | Reaches the lower gut where it is fermented | None observed | 40 g lactitol/d is well tolerated |
| Ballongue et al. (82) | — | — | RCT | 20 g lactitol/d | Microbiology determined in agar medium | Lactitol decreased populations of Bacteroides, Clostridium, coliforms, and Eubacterium; decreased fecal pH | None observed | Bacteroides, Clostridium, coliforms, and Eubacterium were decreased by 1.5, 1.2, 1, and 1.9 log units |
| Pinna et al. (81) | — | — | In vitro | 2 g/L for 24 h | Fluorescence in situ hybridization | Reduced the population of Enterobacteriaceae in feline fecal culture at 2 g/L, exerting prebiotic effect on feline intestinal microbiota | None observed | Clostridium perfringens: +1.6 log cells/g; Enterobacteriaceae –0.3 log cells/g |
| Peuranen et al. (83) | — | — | Rats | 2% (wt:wt) | Flow cytometry and 16S rRNA sequencing | Increased the production of butyrate and IgA secretion without signs of mucosal inflammation | None observed | Butyric acid: 22.2% IgA: 996% (lactitol + polydextrose) |
| Ouwehand et al. (84) | — | — | RCT | 5–5.5 g | Flow cytometry and 16S rRNA sequencing | Lactitol as a synbiotic combined with Lactobacillus acidophilus NCFM may improve some markers of the intestinal microbiota | None observed | Synbiotic: 7.8 × 109 CFUs/g ; placebo: 3.8 × 109 CFUs/g |
| Björklund et al. (85) | — | — | RCT | 2 × 1010L. acidophilus and 10 g lactitol | qPCR (percent guanine-plus-cytosine) | L. acidophilus NCFM and lactitol decrease the Blautia coccoides and Eubacterium rectale bacterial group levels | None observed | B. coccoides: 1.83 × 1010 to 1.34 × 1010; E. rectale 1.19 × 1010 to 7.34 × 109 |
| Finney et al. (86) | — | — | RCT | 10 g sucrose:lactitol (ratios: 10:0, 5:5, 0:10) | Microbiology determined in agar medium | 10 g lactitol can beneficially affect the fecal microbiota, increasing bifidobacteria and concentrations of propionic and butyric acids | None observed | 10 g lactitol increased from 9.37 to 10.06 bifidobacteria log CFUs |
| Maltitol (E-965) | Obtained by the hydrolysis, reduction, and hydrogenation of starch | — | — | — | — | — | — | — |
| Beards et al. (88) | — | — | RCT | 22.8 g | 16S rRNA sequencing | Numbers of fecal bifidobacteria significantly increased after maltitol treatment | None observed | 0.8 log cells/g feces |
| Sorbitol (E-420) | Obtained by catalytic hydrogenation of glucose with subsequent purification and is found naturally in apples, pears, peaches, apricots, and some vegetables | — | — | — | — | — | — | — |
| Yao et al. (90) | — | — | — | — | — | Adverse gastrointestinal reactions to sorbitol in IBD patients | None observed | — |
| Mannitol (E-421) | Mannitol is obtained from hydrogenation of glucose and purification | Similar absorption rate to sorbitol | — | — | — | No effects on gut microbiota | None observed | — |
| Xylitol (E-967) | Fruits, berries, vegetables, oats, and mushrooms; a small percentage is also produced by the human body | — | — | — | — | — | — | — |
| Uebanso et al. (49) | — | — | Mice | 40 and 200 mg xylitol · kg body weight−1 · d−1 | 16S rRNA sequencing | Reduced the abundance of fecal Bacteroidetes and the genus Barnesiella and increased Firmicutes and the genus Prevotella | None observed | 194 mg xylitol · kg−1 · d−1 reduced |
| Tamura et al. (96) | — | — | Mice | 5% xylitol diet for 28 d | Detected by T-RFLP analysis, based on PCR amplification | The concentration of Bacteroides was higher in the control diet than in the xylitol-rich diet | None observed | 50% |
| Naaber et al. (99) | — | — | Mice | Synbiotic: Lactobaccillus rhamnosus and xylitol (1 mL of 20% solution) | Diffusion method and blood agar | Treatment of L. rhamnosus and xylitol had some effects against Clostridium difficile in a mouse model | None observed | Translocation effects |
IBD, inflammatory bowel disease; NA, not available; RCT, randomized clinical trial; rRNA, ribosomal RNA; T-RFLP, terminal restriction fragment length polymorphism.
FIGURE 3.

Effects of polyols on gut microbiota composition. Erythritol, sorbitol, and mannitol do not affect the composition of the gut microbiota; however, lactitol, isomalt, xylitol, and maltitol provoke shifts in the gut microbiome, increasing bifidobacteria numbers in healthy people. The laxative effects of polyols need to be considered when they are consumed by patients with inflammatory bowel disease. C. difficile, Clostridium difficile; N/A, not available.
Effects of sweeteners on the gut microbiota in human trials
Sweeteners are used in many food processes, and the impact of the consumption of these kinds of compounds affects health status and microbiota composition. Today, the potential modifications of the intestinal microbiome, mediated by specific sweeteners, in healthy adults or children are a matter of concern. However, there are few clinical studies.
The study by Suez et al. (15) in 2014 showed modifications in the intestinal microbiota after the administration of some sweeteners [especially noncaloric artificial sweeteners (NASs)] from data collected on 172 randomly selected individuals. They found positive correlations between NAS consumption and the Enterobacteriaceae family, the Deltaproteobacteria class, and the Actinobacteria phylum. In addition, they followed 7 healthy volunteers who did not normally consume NASs or NAS-containing foods for 1 wk. In that week, the volunteers consumed the maximal ADI of saccharin (5 mg/kg). Compared with their individual glycemic response on days 1–4, the volunteers in the NAS group showed decreased glycemic responses at days 5 and 7 (15). The magnitude of the difference was >30%. These findings suggest that NAS consumption, and especially saccharin at a maximum dose, might have a deleterious effect on glucose tolerance through changes to the intestinal microbiota. There is a current controversy in the scientific field with regard to the Suez et al. study because of the control groups, the use of antibiotics, and the fecal transplantations that were used (100). Nevertheless, the simple message from this study is that dietary sugar alternatives meant to stave off the risk of obesity and diabetes might increase the risk of those diseases.
In another study, 31 adults completed a 4-d food record and provided a fecal sample on the fifth day. Their intestinal microbiota were analyzed by pyrosequencing. The abundance profiles were not associated with sweetener consumption, especially with aspartame. However, the overall bacterial diversity varied across both consumers and nonconsumers of sweeteners (23).
Conclusions
Growing concerns about the increased prevalence of obesity and its metabolic comorbidities have led to a reduced consumption of simple sugars and an increase in the intake of NNSs and LCSs. Thus, sweeteners, which appear as sugar alternatives, have been critically evaluated by the FDA, EFSA, and Codex Alimentarius and are considered safe and well tolerated. However, some long-term prospective studies raise the concern that the consumption of artificial sweeteners might actually contribute to the development of metabolic derangements that lead to obesity, T2D, and cardiovascular disease (101). In addition, there are gaps in the evidence related to the effects of NNSs on appetite, short-term intake, and risk of cancer and diabetes (2). The effects of sweeteners on gut microbiota have not been completely elucidated. Within NNSs, only saccharin and sucralose shift the populations of gut microbiota. The ingestion of saccharin by animals and humans showed alterations in metabolic pathways linked to glucose tolerance and dysbiosis in humans. However, more human studies are needed to clarify these preliminary observations. Within nutritive sweeteners, only stevia extracts may affect gut microbiota composition. Finally, polyols, as they reach the colon, can induce dose-dependent flatulence, especially in patients with inflammatory bowel disease. Several polyols, including isomalt and maltitol, increase bifidobacteria numbers in healthy subjects, and these polyols may have prebiotic actions. On the other hand, different human clinical trials showed that lactitol decreases the populations of Bacteroides, Clostridium, coliforms, and Eubacterium. In addition, lactitol increases the production of butyrate and IgA secretion without signs of mucosal inflammation and presents symbiotic effects. Xylitol reduces the abundance of fecal Bacteroidetes and the genus Barnesiella, increases Firmicutes and the genus Prevotella, and affects C. difficile in mice.
Further studies are needed to elucidate whether the changes observed in the intestinal microbiota in animals are present in humans and to study the effects of sweeteners for which evidence is not available so far. In this regard, there is an actual need to perform well-designed, long-term, double-blind, placebo-controlled, randomized clinical trials with appropriated doses and adequate subject sizes to evaluate the potential impact of both NNSs and LCSs on intestinal microbiota and how they could affect major outcomes and risk biomarkers related to chronic diseases.
Acknowledgments
FJR-O, JP-D, and AG are part of the Plan Propio de Investigación 2016, Unit of Excellence on Exercise and Health (UCEES), University of Granada. The authors’ responsibilities were as follows—FJR-O, JP-D, MJS-L, and AG: contributed to the design, analysis, and presentation of the results and drafted the manuscript; and all authors: discussed, revised, and read and approved the final manuscript.
Notes
Published in a supplement to Advances in Nutrition. Presented at the International Union of Nutritional Sciences (IUNS) 21st International Congress of Nutrition (ICN) held in Buenos Aires, Argentina, October 15–20, 2017. The International Union of Nutritional Sciences (IUNS) thanks Mead Johnson Nutrition and Herbalife Nutrition for generously providing grants to support the publication and distribution of the present supplement from the 21st International Union of Nutritional Sciences. The contents of this supplement are solely the responsibility of the authors and do not necessarily represent official views of the IUNS. The supplement coordinators were Angel Gil and Alfredo Martinez. The supplement coordinators had no conflicts of interest to disclose.The authors reported no funding received for this study.
Author disclosures: FJR-O, JP-D, MJS-L, and AG, no conflicts of interest.Publication costs for this supplement were defrayed in part by the payment of page charges. This publication must therefore be hereby marked “advertisement” in accordance with 18 USC section 1734 solely to indicate this fact. The opinions expressed in this publication are those of the authors and are not attributable to the sponsors or the publisher, Editor, or Editorial Board of Advances in Nutrition.
Abbreviations used: ADI, Acceptable Daily Intake; EFSA, European Food Safety Authority; EU, European Union; IBS, irritable bowel syndrome; LCS, low-calorie sweetener; NAS, noncaloric artificial sweetener; NNS, nonnutritive sweetener; T2D, type 2 diabetes.
References
- 1. Stanhope KL. Sugar consumption, metabolic disease and obesity: the state of the controversy. Crit Rev Clin Lab Sci 2016;53:52–67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Lohner S, Toews I, Meerpohl JJ. Health outcomes of non-nutritive sweeteners: analysis of the research landscape. Nutr J 2017;16:55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Shallenberger RS, Acree TE. Chemical structure of compounds and their sweet and bitter tas te. In: Handbook of sensory physiology. Berlin:Springer-Verlag; 1971;51:221–77. [Google Scholar]
- 4. Chattopadhyay S, Raychaudhuri U, Chakraborty R. Artificial sweeteners—a review. J Food Sci Technol 2014;51(4):611–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Carocho M, Morales P, Ferreira ICFR. Sweeteners as food additives in the XXI century: a review of what is known, and what is to come. Food Chem Toxicol 2017;107(Part A):302–17. [DOI] [PubMed] [Google Scholar]
- 6. FDA. High-intensity sweeteners. 2015. Available from: www.fda.gov/food/ingredientspackaginglabeling/foodadditivesingredients/ucm397716.htm, Accessed 2014. [Google Scholar]
- 7. Food Standards Agency. Cu rrent EU approved additives and their E numbers. 2016. [cited 2018 Jan 13]. Available from: https://www.food.gov.uk/science/additives/enumberlist#toc-4. [Google Scholar]
- 8. Fowler SP, Williams K, Resendez RG, Hunt KJ, Hazuda HP, Stern MP. Fueling the obesity epidemic? Artificially sweetened beverage use and long-term weight gain. Obesity (Silver Spring) 2008;16:1894–900. [DOI] [PubMed] [Google Scholar]
- 9. Lutsey PL, Steffen LM, Stevens J. Dietary intake and the development of the metabolic syndrome: the Atherosclerosis Risk in Communities Study. Circulation 2008;117:754–61. [DOI] [PubMed] [Google Scholar]
- 10. Nettleton JA, Lutsey PL, Wang Y, Lima JA, Michos ED, Jacobs DR Jr. Diet soda intake and risk of incident metabolic syndrome and type 2 diabetes in the Multi-Ethnic Study of Atherosclerosis (MESA). Diabetes Care 2009;32:688–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Fagherazzi G, Vilier A, Saes Sartorelli D, Lajous M, Balkau B, Clavel-Chapelon F. Consumption of artificially and sugar-sweetened beverages and incident type 2 diabetes in the Etude Epidemiologique aupres des femmes de la Mutuelle Generale de l'Education Nationale—European Prospective Investigation into Cancer and Nutrition cohort. Am J Clin Nutr 2013;97:517–23. [DOI] [PubMed] [Google Scholar]
- 12. Pepino MY. Metabolic effects of non-nutritive sweeteners. Physiol Behav 2015;152:450–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Schulze MB, Manson JE, Ludwig DS, Colditz GA, Stampfer MJ, Willett WC, Hu FB. Sugarsweetened beverages, weight gain, and incidence of type 2 diabetes in young and middle-aged women. JAMA 2004;292:927–34. [DOI] [PubMed] [Google Scholar]
- 14. Swithers SE. Artificial sweeteners produce the counterintuitive effect of inducing metabolic derangements. Trends Endocrinol Metab 2013;24:431–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Suez J, Korem T, Zeevi D, Zilberman-Schapira G, Thaiss CA, Maza O, Israeli D, Zmora N, Gilad S, Weinberger A et al.. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 2014;514:181–6. [DOI] [PubMed] [Google Scholar]
- 16. Tucker RM, Tan SY. Do non-nutritive sweeteners influence acute glucose homeostasis in humans? A systematic review. Physiol Behav 2017;182:17–26. [DOI] [PubMed] [Google Scholar]
- 17. Livesey G. Health potential of polyols as sugar replacers, with emphasis on low glycaemic properties. Nutr Res Rev 2003;16:163e191. [DOI] [PubMed] [Google Scholar]
- 18. Bokulich NA, Blaser MJ. A bitter aftertaste: unintended effects of artificial sweeteners on the gut microbiome. Cell Metab 2014;20:701–3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Clemente JC, Ursell LK, Parfrey LW, Knight R. The impact of the gut microbiota on human health: an integrative view. Cell 2012;148:1258–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Suez J, Korem T, Zilberman-Schapira G, Segal E, Elinav E. Non-caloric artificial sweeteners and the microbiome: findings and challenges. Gut Microbes 2015;6:149–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Renwick AG. The metabolism of intense sweeteners. Xenobiotica 1986;16:1057–71. [DOI] [PubMed] [Google Scholar]
- 22. Pfeffer M, Ziesenitz SC, Siebert G. Acesulfame K, cyclamate and saccharin inhibit the anaerobic fermentation of glucose by intestinal bacteria. Z Ernahrungswiss 1985;24:231–5. [DOI] [PubMed] [Google Scholar]
- 23. Frankenfeld CL, Sikaroodi M, Lamb E, Shoemaker S, Gillevet PM. High-intensity sweetener consumption and gut microbiome content and predicted gene function in a cross-sectional study of adults in the United States. Ann Epidemiol 2015;25:736–42, e4. [DOI] [PubMed] [Google Scholar]
- 24. Uebanso T, Ohnishi A, Kitayama R, Yoshimoto A, Nakahashi M, Shimohata T, Mawatari K, Takahashi A. Effects of low-dose non-caloric sweetener consumption on gut microbiota in mice. Nutrients 2017;9:E560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Bian X, Chi L, Gao B, Tu P, Ru H, Lu K. The artificial sweetener acesulfame potassium affects the gut microbiome and body weight gain in CD-1 mice. PLoS One 2017;12:e0178426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Horwitz DL, McLane M, Kobe P. Response to single dose of aspartame or saccharin by NIDDM patients. Diabetes Care 1988;11:230–4. [DOI] [PubMed] [Google Scholar]
- 27. Tordoff MG, Alleva AM. Effect of drinking soda sweetened with aspartame or high-fructose corn syrup on food intake and body weight. Am J Clin Nutr 1990;51:963–9. [DOI] [PubMed] [Google Scholar]
- 28. Palmnäs MS, Cowan TE, Bomhof MR, Su J, Reimer RA, Vogel HJ, Hittel DS, Shearer J. Low-dose aspartame consumption differentially affects gut microbiota-host metabolic interactions in the diet-induced obese rat. PLoS One 2014;9:e109841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Magnuson BA, Burdock GA, Doull J, Kroes RM, Marsh GM, Pariza MW, Spencer PS, Waddell WJ, Walker R, Williams GM. Aspartame: a safety evaluation based on current use levels, regulations, and toxicological and epidemiological studies". Crit Rev Toxicol 2007;37:629–727. [DOI] [PubMed] [Google Scholar]
- 30. Magnuson BA, Carakostas MC, Moore NH, Poulos SP, Renwick AG. Biological fate of low-calorie sweeteners. Nutr Rev 2016;74(11):670–89. [DOI] [PubMed] [Google Scholar]
- 31. Mitchell H. Sweeteners and sugar alternatives in food technology. Oxford (United Kingdom): Blackwell Publishing; 2006. [Google Scholar]
- 32. Nofre C, Tinti J. Neotame: discovery, properties, utility. Food Chem 2000;69:245, e257. [Google Scholar]
- 33. Zhu L, Wang G, Dong B, Peng CC, Tian YY, Gong LM. Effects on sweetener neotame on diet preference, performance and hematological and biochemical parameters of weaned piglets. Anim Feed Sci Technol 2016;214:86e94. [Google Scholar]
- 34. Otabe A, Fujieda T, Masuyama T. In vitro and in vivo assessment of the mutagenic activity of N-[N-[3-(3-hydroxy-4-methoxyphenyl) propyl]-α-aspartyl]-L-phenylalanine 1-methyl ester, monohydrate (advantame). Food Chem Toxicol 2011;49(Suppl 1):S30–4. [DOI] [PubMed] [Google Scholar]
- 35. Price JM, Biava CG, Oser BL, Vogin EE, Steinfeld J, Ley HL. Bladder tumors in rats fed cyclohexylamine or high doses of a mixture of cyclamate and saccharin. Science 1970;167:1131–2. [DOI] [PubMed] [Google Scholar]
- 36. Oser BL, Carson S, Cox GE, Vogin EE, Sternberg SS. Chronic toxicity study of cyclamate: saccharin (10:1) in rats. Toxicology 1975;4:315–30. [PubMed] [Google Scholar]
- 37. Ahmed FE, Thomas DB. Assessment of the carcinogenicity of the nonnutritive sweetener cyclamate. Crit Rev Toxicol 1992;22:81–118. [DOI] [PubMed] [Google Scholar]
- 38. Drasar BS, Renwick AG, Williams RT. The role of the gut flora in the metabolism of cyclamate. Biochem J 1972;129:881–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Mallett AK, Rowland IR, Bearne CA, Purchase R, Gangolli SD. Metabolic adaptation of rat faecal microflora to cyclamate in vitro. Food Chem Toxicol 1985;23:1029–34. [DOI] [PubMed] [Google Scholar]
- 40. Goldin BR. Intestinal microflora: metabolism of drugs and carcinogens. Ann Med 1990;22:43–8. [DOI] [PubMed] [Google Scholar]
- 41. Anderson RL, Kirkland JJ. The effect of sodium saccharin in the diet on caecal microflora. Food Cosmet Toxicol 1980;18:353–5. [DOI] [PubMed] [Google Scholar]
- 42. Naim M, Zechman JM, Brand JG, Kare MR, Sandovsky V. Effects of sodium saccharin on the activity of trypsin, chymotrypsin, and amylase and upon bacteria in small intestinal contents of rats. Proc Soc Exp Biol Med 1985;178:392–401. [DOI] [PubMed] [Google Scholar]
- 43. Daly K, Darby AC, Hall N, Nau A, Bravo D, Shirazi-Beechey SP. Dietary supplementation with lactose or artificial sweetener enhances swine gut Lactobacillus population abundance. Br J Nutr 2014;111:S30–5. [DOI] [PubMed] [Google Scholar]
- 44. Daly K, Darby AC, Hall N, Wilkinson MC, Pongchaikul P, Bravo D, Shirazi-Beechey SP. Bacterial sensing underlies artificial sweetener-induced growth of gut Lactobacillus. Environ Microbiol 2016;18:2159–71. [DOI] [PubMed] [Google Scholar]
- 45. Labrecque MT, Malone D, Caldwell KE, Allan AM. Impact of ethanol and saccharin on fecal microbiome in pregnant and non-pregnant mice. J Pregnancy Child Health 2015;2:1000193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Bian X, Tu P, Chi L, Gao B, Ru H, Lu K. Saccharin induced liver inflammation in mice by altering the gut microbiota and its metabolic functions. Food Chem Toxicol 2017;107:530–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Roberts A, Renwick AG, Sims J, Snodin DJ. Sucralose metabolism and pharmakinetics in man. Food Chem Toxicol 2008;38:S31eS41. [DOI] [PubMed] [Google Scholar]
- 48. Abou-Donia MB, El-Masry EM, Abdel-Rahman AA, McLendon RE, Schiffman SS. Splenda alters gut microflora and increases intestinal p-glycoprotein and cytochrome p-450 in male rats. J Toxicol Environ Health A 2008;71:1415–29. [DOI] [PubMed] [Google Scholar]
- 49. Uebanso T, Kano S, Yoshimoto A, Naito C, Shimohata T, Mawatari K, Takahashi A. Effects of consuming xylitol on gut microbiota and lipid metabolism in mice. Nutrients 2017;9:E756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Bian X, Chi L, Gao B, Tu P, Ru H, Lu K. Gut microbiome response to sucralose and its potential role in inducing liver inflammation in mice. Front Physiol 2017;8:487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Soejima A, Tanabe AS, Takayama I, Kawahara T, Watanabe K, Nakazawa M, Mishima M, Yahara T. Phylogeny and biogeography of the genus Stevia (Astareaceae: Eupatorieae): an example of diversification in the Asteraceae in the new world. J Plant Res 2017;130:953–72. [DOI] [PubMed] [Google Scholar]
- 52. Ferrazzano GF, Cantile T, Alcidi B, Coda M, Ingenito A, Zarrelli A, Di Fabio G, Pollio A. Is Stevia rebaudiana Bertoni a non cariogenic sweetener? A review. Molecules 2015;21:E38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. European Food Safety Authority. Scientific opinion on the safety of steviol glycosides for the proposed uses as a food additive. EFSA Panel of Food Additive and Nutrients Sources added to Food (ANS). EFSA J 2010;8(4):1537. [Google Scholar]
- 54. European Food Safety Authority. Scientific opinion on the safety of the proposed amendment of the specifications for steviol glycosides (E960) as a food additive. EFSA Panel of Food Additive and Nutrient Sources added to Food (ANS). EFSA J 2015;13(12):4316. [Google Scholar]
- 55. Koyama E, Kitazawa K, Ohori Y, Izawa O, Kakegawa K, Fujino A, Ui M. In vitro metabolism of the glycosidic sweeteners, stevia mixture and enzymatically modified stevia in human intestinal microflora. Food Chem Toxicol 2003;41:359–74. [DOI] [PubMed] [Google Scholar]
- 56. Gardana C, Simonetti P, Canzi E, Zanchi R, Pietta P. Metabolims of stevioside and rebaudioside A from Stevia rebaudiana extracts by human microflora. J Agric Food Chem 2003;51:6618–22. [DOI] [PubMed] [Google Scholar]
- 57. Renwick AG, Tarka SM. Microbial hydrolysis of steviol glycosides. Food Chem Toxicol 2008;46:S70–4. [DOI] [PubMed] [Google Scholar]
- 58. Sanches Lopes SM, Francisco MG, Higashi B, de Almeida RTR, Krausová G, Pilau EJ, Gonçalves JE, Gonçalves RAC, Oliveira AJB. Chemical characterization and prebiotic activity of fructo-oligosaccharides from Stevia rebaudiana (Bertoni) roots and in vitro adventitious root cultures. Carbohydr Polym 2016;152:718–25. [DOI] [PubMed] [Google Scholar]
- 59. Roohbakhsh A, Iranshahy M, Iranshahi M. Glycyrrhetinic acid and its derivatives: anti-cancer and cancer chemopreventive properties, mechanisms of action and structure-cytotoxic activity relationship. Curr Med Chem 2016;23:498–517. [DOI] [PubMed] [Google Scholar]
- 60. Kim YS, Kim JJ, Cho KH, Jung WS, Moon SK, Park EK, Kim DH. Biotransformation of ginsenoside Rb1, crocin, amygdalin, geniposide, puerarin, ginsenoside Re, hesperidin, poncirin, glycyrrhizin, and baicalin by human fecal microflora and its relation to cytotoxicity against tumor cells. J Microbiol Biotechnol 2008;18:1109–14. [PubMed] [Google Scholar]
- 61. Yu K, Chen F, Li C. Absorption, disposition, and pharmacokinetics of saponins from Chinese medicinal herbs: what do we know and what do we need to know more? Curr Drug Metab 2012;13:577–98. [DOI] [PubMed] [Google Scholar]
- 62. Kim DH, Hong SW, Kim BT, Bae EA, Park HY, Han MJ. Biotransformation of glycyrrhizin by human intestinal bacteria and its relation to biological activities. Arch Pharm Res 2000;23:172–7. [DOI] [PubMed] [Google Scholar]
- 63. Yim JS, Kim YS, Moon SK, Bae HS, Kim JJ, Park EK, Dim DH. Metabolic activities of ginsenoside Rb1, baicalin, glycyrrhizin and geniposide to their bioactive compounds by human intestinal microflora. Biol Pharm Bull 2004;10:1580–3. [DOI] [PubMed] [Google Scholar]
- 64. Leone S, Picone D. Molecular dynamics driven design of pH-stabilized mutants of MNEI, a sweet protein. PloS One 2016;11:e0158372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Grembecka M. Sugar alcohols—their role in the modern world of sweeteners: a review. Eur Food Res Technol 2015;241:1–14. [Google Scholar]
- 66. Lenhart A, Chey WD. A systematic review of the effects of polyols on gastrointestinal health and irritable bowel syndrome. Adv Nutr 2017;8(4):587–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. European Food Safety Authority. Call for technical and toxicological data on sweeteners authorised as food additives in the EU. 2017. Available from: https://www.efsa.europa.eu/en/data/call/170621.
- 68. Bernt WO, Borzelleca JF, Flamm G, Munro IC. Erythritol: a review of biological and toxicological studies. Regul Toxicol Pharmacol 1996;24:S191–7. [DOI] [PubMed] [Google Scholar]
- 69. Goossens J, Röper H. Erythritol: a new sweetener. Vilvoorde (Belgium): Cerestar R&D, Euro Centre Food; 1994. [Google Scholar]
- 70. Kawanabe J, Hirasawa M, Takeuchi T, Oda T, Ikeda T. Noncariogenicity of erythritol as a substrate. Caries Res 1992;26:358–62. [DOI] [PubMed] [Google Scholar]
- 71. Arrigoni E, Brouns F, Amadò R. Human gut microbiota does not ferment erythritol. Br J Nutr 2005;94:643–6. [DOI] [PubMed] [Google Scholar]
- 72. Bornet FR, Blayo A, Dauchy F, Slama G. Gastrointestinal response and plasma and urine determinations in human subjects given erythritol. Regul Toxicol Pharmacol 1996;24:S296–302. [DOI] [PubMed] [Google Scholar]
- 73. Ishikawa M, Miyashita M, Kawashima Y, Nakamura T, Saitou N, Modderman J. Effects of oral administration of erythritol on patients with diabetes. Regul Toxicol Pharmacol 1996;24:S303–8. [DOI] [PubMed] [Google Scholar]
- 74. Van Ommen B, de Bie B, Bär A. Disposition of 14C-erythritol in germfree and conventional rats. Regul Toxicol Pharmacol 1996;24:S198–205. [DOI] [PubMed] [Google Scholar]
- 75. Gostner A, Schäffer V, Theis S, Menzel T, Lührs H, Melcher R, Schauber J, Kudlich T, Dusel G, Dorbath D et al.. Effects of isomalt consumption on gastrointestinal and metabolic parameters in healthy volunteers. Br J Nutr 2005;94(4):575–81. [DOI] [PubMed] [Google Scholar]
- 76. Gostner A, Blaut M, Schäffer V, Kozianowski G, Theis S, Klingeberg M, Dombrowski Y, Martin D, Ehrhardt S, Taras D et al.. Effect of isomalt consumption on faecal microflora and colonic metabolism in healthy volunteers. Br J Nutr 2006;95(1):40–50. [DOI] [PubMed] [Google Scholar]
- 77. Schauber J, Weiler F, Gostner A, Melcher R, Kudlich T, Lührs H, Scheppach W. Human rectal mucosal gene expression after consumption of digestible and non-digestible carbohydrates. Mol Nutr Food Res 2006;50:1006–12. [DOI] [PubMed] [Google Scholar]
- 78. Patil DH, Grimble GK, Silk DB. Lactitol, a new hydrogenated lactose derivative: intestinal absorption and laxative threshold in normal human subjects. Br J Nutr 1987;57(2):195–9. [DOI] [PubMed] [Google Scholar]
- 79. Piva A, Panciroli A, Meola E, Formigoni A. Lactitol enhances short-chain fatty acid and gas production by swine cecal microflora to a greater extent when fermenting low rather than high fiber diets. Nutrient Metab 1996;126(1):280–9. [DOI] [PubMed] [Google Scholar]
- 80. Ackerman DL, Craft KM, Townsend SD. Infant food applications of complex carbohydrates: structure, synthesis, and function. Carbohydr Res 2017;437:16–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Pinna C, Stefanelli C, Biagi G. In vitro effect of dietary protein level and nondigestible oligosaccharides on feline fecal microbiota. J Anim Sci 2014;92:5593–602 [DOI] [PubMed] [Google Scholar]
- 82. Ballongue J, Schumann C, Quignon P. Effects of lactulose and lactitol on colonic microflora and enzymatic activity. Scand J Gastroenterol Suppl 1997;222:41–4. [DOI] [PubMed] [Google Scholar]
- 83. Peuranen S, Tiihonen K, Apajalahti J, Kettunen A, Saarinen M, Rautonen N. Combination of polydextrose and lactitol affects microbial ecosystem and immune responses in rat gastrointestinal tract. Br J Nutr 2004;91:905–14. [DOI] [PubMed] [Google Scholar]
- 84. Ouwehand AC, Tiihonen K, Saarinen M, Putaala H, Rautonen N. Influence of a combination of Lactobacillus acidophilus NCFM and lactitol on healthy elderly: intestinal and immune parameters. Br J Nutr 2009;101:367–75. [DOI] [PubMed] [Google Scholar]
- 85. Björklund M, Ouwehand AC, Forssten SD, Nikkilä J, Tiihonen K, Rautonen N, Lahtinen SJ. Gut microbiota of healthy elderly NSAID users is selectively modified with the administration of Lactobacillus acidophilus NCFM and lactitol. Age (Dordr) 2012;34(4):987–99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Finney M, Smullen J, Foster HA, Brokx S, Storey DM. Effects of low doses of lactitol on faecal microflora, pH, short chain fatty acids and gastrointestinal symptomology. Eur J Nutr 2007;46:307–14. [DOI] [PubMed] [Google Scholar]
- 87. Joshi K, Kumari A, Arora S, Singh AK. Development of an analytical protocol for the estimation of maltitol from yoghurt, burfi and flavoured milk. Food Sci Tech nol 2016;70:41, e45. [Google Scholar]
- 88. Beards E, Tuohy K, Gibson G. A human volunteer study to assess the impact of confectionery sweeteners on the gut microbiota composition. Br J Nutr 2010;104:701–8. [DOI] [PubMed] [Google Scholar]
- 89. Hyams JS. Sorbitol intolerance: an unappreciatedcause of functional gastrointestinal complaints. Gastroenterology 1983;84:30–3. [PubMed] [Google Scholar]
- 90. Yao CK, Tan HL, van Langenberg DR, Barrett JS, Rose R, Liels K, Gibson PR, Muir JG. Dietary sorbitol and mannitol: food content and distinct absorption patterns between healthy individuals and patients with irritable bowel syndrome. J Hum Nutr Diet 2014;27:263, e275. [DOI] [PubMed] [Google Scholar]
- 91. Corazza GR, Strocchi A, Rossi R, Sirola D, Gasbarrini G. Sorbitol malabsorption in normal volunteers and in patients with coeliac disease. Gut 1988;29:44–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Zumbé A, Lee A, Storey D. Polyols in confectionery: the route to sugarfree, reduced sugar and reduced calorie confectionery. Br J Nutr 2001;85(Suppl 1):S31–45. [DOI] [PubMed] [Google Scholar]
- 93. Symons P, Jones MP, Kellow JE. Symptom provocation in irritable bowel syndrome: effects of differing doses of fructose-sorbitol. Scand J Gastroenterol 1992;27:940–4. [DOI] [PubMed] [Google Scholar]
- 94. Biesiekierski JR, Rosella O, Rose R, Liels K, Barrett JS, Shepherd SJ, Gibson PR, Muir JG. Quantification of fructans, galacto-oligosacharides and other short-chain carbohydrates in processed grains and cereals. J Hum Nutr Diet 2011;24:154, e176. [DOI] [PubMed] [Google Scholar]
- 95. Lee J. Sorbitol, Rubus fruit, and misconception. Food Chem 2015;166:616, e622. [DOI] [PubMed] [Google Scholar]
- 96. Tamura M, Hoshi C, Hori S. Xylitol affects the intestinal microbiota and metabolism of daidzein in adult male mice. Int J Mol Sci 2013;14:23993–4007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Salminen S, Salminen E, Koivistoinen P, Bridges J, Marks V. Gut microflora interactions with xylitol in the mouse, rat and man. Food Chem Toxicol 1985;23:985–90. [DOI] [PubMed] [Google Scholar]
- 98. Sato T, Kusuhara S, Yokoi W, Ito M, Miyazaki K. Prebiotic potential of L-sorbose and xylitol in promoting the growth and metabolic activity of specific butyrate-producing bacteria in human fecal culture. FEMS Microbiol Ecol 2017;93(1):227. [DOI] [PubMed] [Google Scholar]
- 99. Naaber P, Mikelsaar RH, Salminen S, Mikelsaar M. Bacterial translocation, intestinal microflora and morphological changes of intestinal mucosa in experimental models of Clostridium difficile infection. J Med Microbiol 1998;47(7):591–8. [DOI] [PubMed] [Google Scholar]
- 100. Greenhill C. Gut microbiota: not so sweet—artificial sweeteners can cause glucose intolerance by affecting the gut microbiota. Nat Rev Endocrinol 2014;10:637. [DOI] [PubMed] [Google Scholar]
- 101. Swithers SE, Shearer J. Obesity: sweetener associated with increased adiposity in young adults. Nat Rev Endocrinol 2017;13:443–4. [DOI] [PubMed] [Google Scholar]