

Abstract
Parkinson's disease (PD) is a common neurodegenerative disorder characterized by selective loss of dopamine neurons in the substantia nigra pars compacta of the midbrain. Reports from postmortem studies in the human PD brain, and experimental PD models reveal that endoplasmic reticulum (ER) stress is implicated in the pathogenesis of PD. In times of stress, the unfolded or misfolded proteins overload the folding capacity of the ER to induce a condition generally known as ER stress. During ER stress, cells activate the unfolded protein response (UPR) to handle increasing amounts of abnormal proteins, and recent evidence has demonstrated the activation of the ER chaperone GRP78/BiP (78 kDa glucose-regulated protein/binding immunoglobulin protein), which is important for proper folding of newly synthesized and partly folded proteins to maintain protein homeostasis. Although the activation of this protein is essential for the initiation of the UPR in PD, there are inconsistent reports on its expression in various PD models. Consequently, this review article aims to summarize current knowledge on neuroprotective agents targeting the expression of GRP78/BiP in the regulation of ER stress in experimental PD models.
1. Introduction
Parkinson's disease (PD) is a neurological disorder characterized by degeneration of dopaminergic neurons in the substantia nigra pars compacta (SNpc) of the midbrain, resulting in loss of dopamine in the striatum. In patients with PD, there are four primary motor symptoms which include tremor at rest, postural instability, rigidity, and bradykinesia [1]. PD was previously considered to be a condition that affects only the motor system, but with more research, it is now known to be a multifaceted disorder with diverse clinical features that include sleep, cognitive, and neuropsychiatric disorders [2, 3].
Although the etiology of the disease is not entirely understood, reports indicate that several factors such as oxidative and endoplasmic reticulum (ER) stress promote neuronal degeneration. The ER is regarded as the largest organelle in the cell with multiple functions such as protein, steroid, and phospholipid synthesis, storage of calcium, and metabolism of carbohydrates [4–8]. In the ER, chaperones such as 78 kDa glucose-regulated protein (GRP78), also known as binding immunoglobulin protein (BiP) or heat shock 70 kDa protein 5 (HSPA5) and other stress sensor proteins, are needed to maintain quality control of proteins. These proteins are activated to ensure proper handling and to prevent aggregation of misfolded/unfolded proteins [9]. Thus, when there is a disturbance in function, oxidative damage, or disruption of glucose or calcium homeostasis, the unfolded/misfolded proteins exceeds the folding capacity of the ER resulting to a condition commonly known as ER stress [10, 11]. The induction of ER stress and the consequent aggregation of misfolded or unfolded proteins have been implicated in PD pathogenesis [12, 13].
Existing treatment options for PD are inadequate as drugs are focused mainly on relieving symptoms. For example, levodopa is exceptionally effective for regulating PD symptoms, especially those linked to bradykinesia [14], and its combination with carbidopa improves the beneficial effects of levodopa. In cases where PD patients are sensitive to minor side effects such as nausea and vomiting, lodosyn may be taken with the routine carbidopa/levodopa therapy [15]. Other treatment options include dopamine agonists such as pramipexole [16], ropinirole [17, 18], and apomorphine [19, 20] while nondopaminergic drugs treatments include anticholinergics and amantadine [15] as well as entacapone [21] and tolcapone [22] catechol-o-methyl-transferase inhibitors.
Since there is no treatment for PD, there is an ever-increasing need to identify neuroprotective strategies with the ability to slow down or halt the advancement of PD. This search for new drug treatment options has paved the way for the discovery of such natural products as medicinal herbs, plant extracts, and their bioactive compounds. Some of these compounds are under clinical investigations owing to their remarkable potential as neuroprotective treatment options in PD [23, 24]. In this regard, while drug researchers are currently focused on discovering new remedies, plant-derived bioactive compounds targeting ER stress and its pathways could help in the identification and validation of novel treatment options in PD. Hence, this review presents an outline of the scientific literature on the research of plant-derived bioactive compounds and other neuroprotective agents targeting GRP78/BiP in experimental models of PD.
2. Endoplasmic Reticulum Stress Pathway and Disease
The ER stress pathway or unfolded protein response (UPR) is known to handle growing quantities of aberrant proteins in the ER [25]. This response program is tasked with the reduction of misfolded/abnormal proteins through various mechanisms (Figure 1). Firstly, GRP78/BiP disassociates from the ER stress sensors, namely, protein kinase RNA-like endoplasmic reticulum kinase (PERK), activating transcription factor 6 (ATF6), and inositol-requiring enzyme 1 (IRE1) to initiate the ER stress response. Following dissociation of GRP78/BiP, autophosphorylation and activation of PERK facilitate the phosphorylation of eukaryotic translation initiation factor 2a (eIF2a) to inhibit further protein synthesis and translation [26–28]. ATF6 is cleaved in the Golgi after translocation from the ER and then migrates into the nucleus to upregulate ER chaperones such as GRP78/BiP and 94 kDa glucose-regulated protein (GRP94) which enhances the folding capacity of the ER [29]. Also, IRE1 is involved in endoribonuclease activity and activates X-box binding protein 1 (XBP-1) to promote ER-associated degradation [30–32].
Figure 1.

Simplified diagram highlighting the regulation of ER stress signaling pathways.
The extent and degree of ER stress and UPR activation may determine if the ER stress response is either anti- or prosurvival (Figure 2). Certain aspects of the ER stress response such as increased expression of chaperones would appear to be advantageous by lessening the burden of misfolded proteins [33, 34], while other ER stress responses may be advantageous for a limited amount of time, thus leading to degeneration if sustained. Sustained activation of the UPR under stress would lead to apoptosis via the activation of ER-specific caspases, c-Jun amino-terminal kinase (JNK) and apoptosis signal-regulating kinase 1 (ASK1), induction of CCAAT-enhancer-binding protein homologous protein (CHOP), and the activation of p53 upregulated modulator of apoptosis (PUMA), BAX, and NOXA [35].
Figure 2.
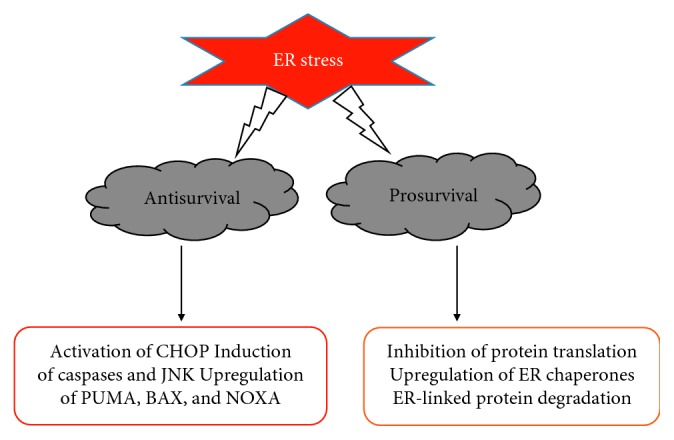
Important events during cellular response to ER stress.
3. ER Stress Response in Parkinson's Disease
GRP78/BiP is a key chaperone essential for proper functioning of the ER and in various cellular processes [36–38]. Most notable is its dual role of regulating protein folding and the initiation of UPR signaling in the ER [39]. In PD, there are inconsistent reports on the expression of GRP78/BiP in various experimental models. For instance, treatment of MN9D cells with a neurotoxin 1-methyl-4-phenylpyridinium (MPP+; Figure 3) resulted in a reduction of GRP78/BiP expression, while treatment of SH-SY5Y cells with a different neurotoxin 6-hydroxydopamine (6-OHDA; Figure 3) increased its expression [40, 41]. In a PD model using MPP+-treated rabbits, Ghribi and colleagues revealed the translocation of GRP78/BiP to the nucleus and cytosol from the ER as well as a significant decrease in TH-positive cells in the SNpc [42]. In a different study, Shimoke and coworkers demonstrated an increase in the expression of GRP78/BiP after exposure to tunicamycin; however, they observed no increase in the expression of GRP78/BiP in PC12 cells after treatment with a neurotoxin, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP; Figure 3), for 24 hours [43]. Duan and Mattson utilized the MPTP-treated mouse model of PD to demonstrate that the upregulation of GRP78/BiP by 2-deoxy-d-glucose significantly prevented loss of dopamine neurons [44].
Figure 3.
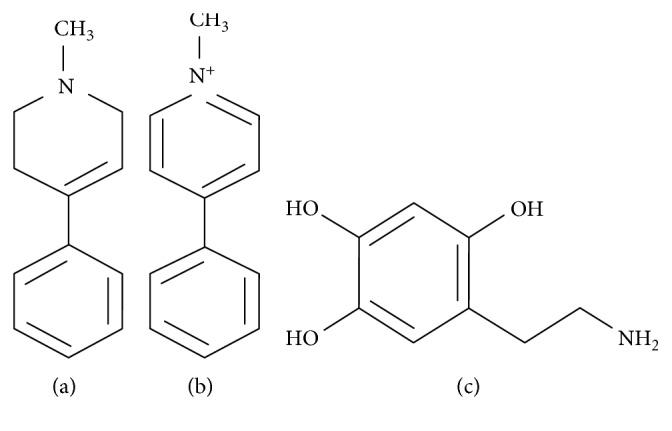
Diagram showing the chemical structure of PD toxins: (a) MPTP; (b) MPP+; (c) 6-OHDA.
In PD patients, GRP78/BiP was reported to be more expressed in the cingulate gyrus and parietal cortex when compared to healthy controls [45]. The upregulation of GRP78/BiP in the cingulate gyrus was linked to an increase in α-synuclein expression, thus providing an association between GRP78/BiP and α-synuclein toxicity. This observation is confirmed by a report demonstrating that the knockdown of GRP78/BiP aggravates the toxicity of α-synuclein in rats [46] and in another study showing that miRNA-induced reduction of GRP78/BiP enhanced cell death induced by a neurotoxin-rotenone [47]. In contrast to studies mentioned above, reports demonstrate that the upregulation of GRP78/BiP suppresses α-synuclein aggregation and toxicity in PD models [48, 49]. For example, Gorbatyuk and colleagues in a rat model of PD induced by an elevated level of human α-synuclein demonstrated that although the accumulation of α-synuclein induced the expression of apoptosis-regulating ATF4, the upregulation of GRP78/BiP inhibited α-synuclein toxicity by regulating ER stress signaling pathways [49].
Leucine-rich repeat kinase 2 (LRRK2) is the most significant gene mutated in PD [50]. LRRK2 pathogenesis has been associated with ER stress as it partly localizes in the ER in dopaminergic neurons of individuals with PD [51]. Reports show that the neuroprotective activity of LRRK2 against 6-OHDA or α-synuclein induced neurodegeneration in the nematode; C. elegans is attributed to the activity of GRP78/BiP via signaling through the p38 mitogen-activated protein kinase (MAPK) pathway [26, 52]. In confirmation of these reports, Samann and colleagues reported that LRRK2 mutant C. elegans were highly vulnerable to ER stress and developed spontaneous neurodegeneration [53, 54]. Furthermore, ageing is the greatest risk factor for PD [55, 56], and various age-related changes in cellular structure and function are observed in PD patients. To corroborate these observations, studies reveal that ageing results in a significant reduction in the activity and expression of GRP78/BiP in the brain of old versus young rodents [57–59]. From the aforementioned, GRP78/BiP is undoubtedly an essential component of the UPR, and proper regulation of GRP78/BiP could prove valuable in identifying new treatment options in PD.
4. Regulation of GRP78/BiP by Therapeutic Agents in PD Models
Over the years, the use of neurotoxin-based experimental models of PD has contributed extensively to the understanding of PD and human health. For instance, such neurotoxins as MPTP, MPP+, 6-OHDA, paraquat, and rotenone have been utilized in the search, identification, and development of novel therapeutic agents in PD [60]. Also, the MPTP mouse and 6-OHDA rat models of PD have contributed immensely to the translation of animal experimentation into clinical practice and are still very much important for investigating different mechanisms of neuronal degeneration in PD. Considerable evidence shows that some experimental therapeutic agents have substantial antioxidant and anti-inflammatory activities, thus demonstrating an inhibitory effect in the oxidative and inflammatory mechanisms linked to neuronal loss in PD [61, 62].
The plant-derived bioactive compounds and other therapeutic agents highlighted in this review demonstrate significant neuroprotective effects and also regulate the activity of GRP78/BiP in experimental models of PD. One such compound is luteolin (3′, 4′, 5′, 7′-tetrahydroxyflavone), a naturally occurring flavonoid present in several herbs, fruits, and vegetables [63, 64]. It is a very potent antioxidant and is usually the most effective when compared to other flavonoids [65]. Plants containing luteolin have been utilized for the inhibition and treatment of such diseases as cancer and hypertension [66, 67]. Also, reports show that luteolin crosses the blood-brain barrier and has multiple biological, pharmacological, anticancer, anti-inflammatory, antibacterial, antiamnesic, and neuroprotective activities [68–71].
While luteolin is structurally composed of hydroxyl groups at carbons 5, 7, 3′, and 4′ positions (Figure 4), the presence of 2-3 double bonds are linked to its multiple biological activities [72]. In a study, Hu and colleagues investigated the neuroprotective activity of luteolin in PC12 cells treated with 6-OHDA using RT-Q-PCR and western blot techniques [73]. They reported that luteolin attenuated the 6-OHDA-induced upregulation of GRP78/BiP and downregulated UPR, leading to the reduction of phospho-eIF2a, ATF4, and CHOP [73]. Based on these findings, the authors attributed the neuroprotective activity of luteolin to the regulation of GRP78/BiP and other UPR related proteins.
Figure 4.
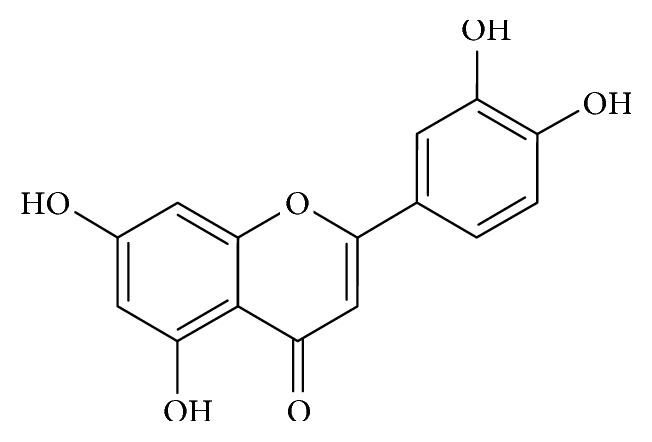
Diagram showing the chemical structure of luteolin.
Salidroside (p-hydroxyphenethyl-β-D-glucoside; C14H20O7; Figure 5), a phenol glycoside extracted as an active constituent from Rhodiola rosea L., is widely used in traditional folk medicine in Asia and Europe [74, 75]. In China, it is commonly used as an antifungal herb and as a supplement to improve kidney function, stimulate blood circulation, and clear chest congestion [76].
Figure 5.
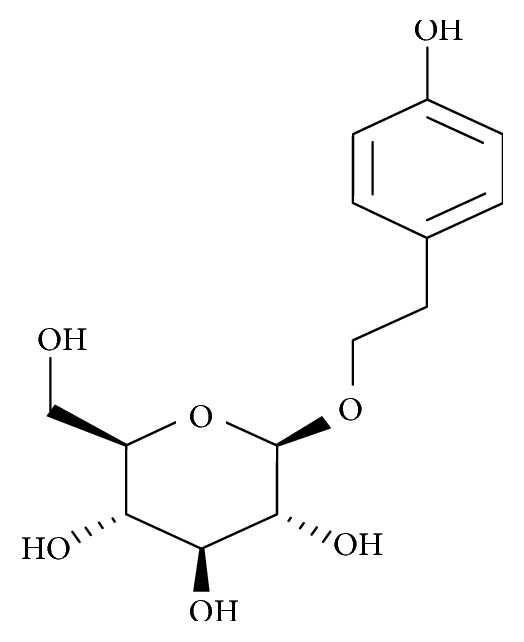
Diagram showing the chemical structure of salidroside.
Salidroside exhibits a wide range of pharmacological activities including antioxidative, antiageing, anticancer, anti-inflammatory antitumour, antidepressive, antifatigue, adaptogenic, cardioprotective, and hepatoprotective effects [77–80]. In addition, reports show that salidroside is effective against cognitive decline during ageing and can protect neurons from apoptosis as well as mitochondrial dysfunction in experimental models of neurodegeneration [81–83]. To investigate salidroside's ability to regulate GRP78/BiP in an experimental model of PD, Tao and coworkers treated SN4741 cells with 6-OHDA after pretreatment with salidroside. Findings revealed that salidroside reduced the expression levels of GRP78/BiP and other ER stress markers (p-PERK and p-IRE1) when compared with cells treated with 6-OHDA only [84]. From the study, they demonstrated that the protective effect of salidroside against the toxicity of 6-OHDA was partly due to the regulation of GRP78/BiP and other ER stress markers.
Lithospermic acid (C27H22O12) is a key component of Salvia miltiorrhiza, a Chinese medicinal herb widely used to increase blood flow and treat diabetic as well as cardiovascular problems in humans [85]. Lithospermic acid shares a similar structure with salvianolic acid B (Figure 6) and is reported to have multiple pharmacological activities which include antihypertensive, antidiabetic, antiapoptotic, and antioxidant effects [86–88].
Figure 6.
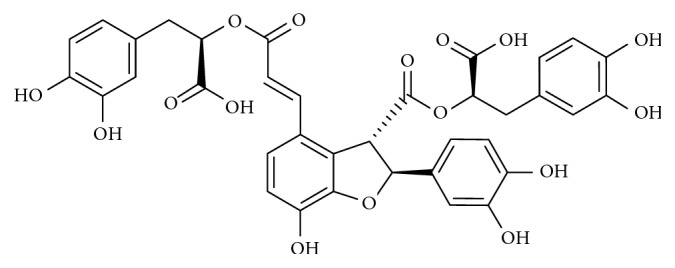
Diagram showing the chemical structure of salvianolic acid B.
In a study by Lin and colleagues, MPP+-treated CATH.a cells were utilized as a model of PD to investigate the role of lithospermic acid on ER stress [89]. Findings from western blots revealed that MPP+ triggered ER stress in CATH.a cells by increasing the expression of GRP78/BiP, while lithospermic acid treatment inhibited the upregulation of GRP78/BiP, thus acting as a neuroprotective agent [89].
Basic fibroblast growth factor (bFGF), a member of the FGF family, is an essential protein with multiple physiological roles in the peripheral and central nervous system (CNS) [90, 91]. It is involved in a series of neurotrophic activities contributing to CNS repair and cell survival [92]. Reports indicate that bFGF shares receptors and influences a range of biological activities such as inhibition of apoptosis, cellular proliferation, and morphogenesis [93–95]. Previous studies show that bFGF exhibits neuroprotective activities in PD models; for instance, bFGF protected against rotenone-triggered dopaminergic cell loss in SH-SY5Y cells and enhanced survival of dopaminergic cells in human fetal tissue strands transplanted into immunosuppressed rats injected with 6-OHDA [96, 97]. In a study by Cai and coworkers, bFGF was found to suppress 6-OHDA-triggered upregulation of ER stress response proteins in Sprague–Dawley rats. Immunohistochemical and western blot findings revealed that bFGF treatment significantly inhibited 6-OHDA-induced increase in the expression of GRP78/BiP and CHOP, thus providing evidence on the regulation of GRP78/BiP as a neuroprotective mechanism in PD [98].
Ulinastatin (C13H16O3) is a glycoprotein and Kunitz-type serine protease inhibitor obtained by separation and purification from the urine of healthy men [99, 100]. Reports indicate that ulinastatin plays an important anti-inflammatory role through the inhibition of inflammatory cytokines and proteases [101]. For this reason, it is commonly used in Japan, Korea, and China for the management and treatment of severe pancreatitis, rheumatoid arthritis, and other inflammatory disorders [102–104]. Other pharmacological evidence reveals that ulinastatin has a protective role in multiple organ dysfunction syndrome, acute respiratory distress syndrome, and acute lung injury [105, 106]. To investigate the role of ulinastatin on ER stress in an in vivo model of PD, Li and colleagues observed that paraquat triggered a higher expression of GRP78/BiP and apoptosis in treated rats when compared to rats in the control group. However, they observed that ulinastatin-treated rats showed lower expression of GRP78/BiP when compared to rats treated with paraquat only. These findings demonstrated that the regulation of GRP78/BiP by ulinastatin was partly responsible for its overall protective effect observed in the PD model [107].
Salubrinal (C21H17Cl3N4OS; Figure 7) is a synthetic compound that was discovered in a screening of compounds with protective activity on ER stress-mediated cell death [108]. It is used experimentally to investigate stress response studies linked to eIF2α [109], and as a mechanism of action in ER stress, it inhibits the dephosphorylation of eIF2α, leading to a reduction in protein synthesis and inhibition of protein translation [110, 111]. Reports indicate that salubrinal is protective against cell death induced by tunicamycin, arsenic, cyclosporine, cadmium, hypoxia, and paraquat [112–115].
Figure 7.
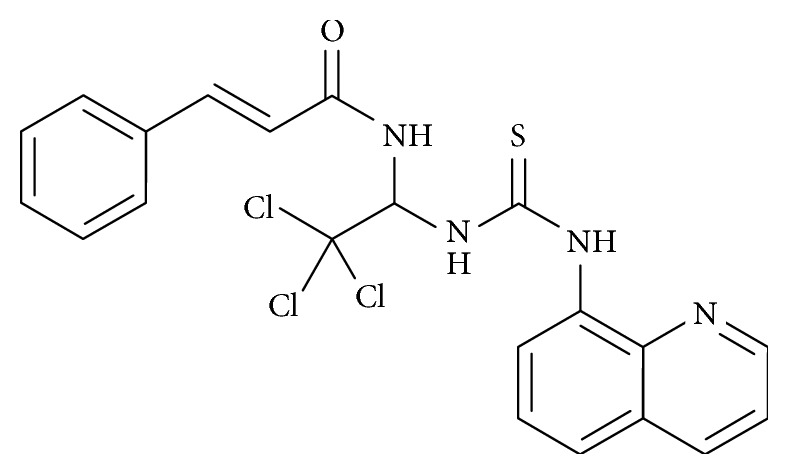
Diagram showing the chemical structure of salubrinal.
As a neuroprotective agent in PD, salubrinal prevented MN9D cells from MPP+ and 6-OHDA-induced toxicity [116]. It is believed that the protective activity of salubrinal can be attributed to the functional groups in its molecular structure except for the quinolone moiety [117]. To demonstrate the neuroprotective mechanism of salubrinal on paraquat-induced ER stress in SH-SY5Y cells, Yang and colleagues showed that salubrinal inhibited the activation of GRP78/BiP and other ER stress sensors IRE1, ASK1, JNK, and CHOP [118]. In a different study by Goswami and colleagues, treatment of neuro-2A cells with rotenone increased the expression of GRP78/BiP and CHOP [119]; however, pretreatment of the cells with salubrinal reduced the expression of GRP78/BiP and CHOP. The authors highlighted the inhibition of eIF2α dephosphorylation and the regulation of GRP78/BiP as a neuroprotective mechanism of salubrinal in rotenone-induced toxicity in PD [119].
Echinacoside (C35H46O20; Figure 8) is a primary component of phenylethanoid glycoside isolated from Cistanche salsa, a Chinese herbal medicine known for its antifatigue and antisenile properties [120, 121]. Reports show that echinacoside has potent antioxidant effects, scavenges for free radicals, and inhibits macrophage-induced generation of nitric oxide [122–124]. Other biological effects include anti-inflammatory, antiageing, antitumour, and hepatoprotective activities [125, 126].
Figure 8.
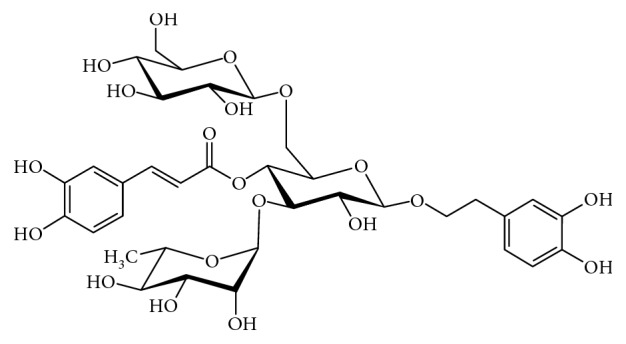
Diagram showing the chemical structure of echinacoside.
The role of echinacoside in neurodegenerative disorders has also been reported; for instance, studies show that echinacoside treatment significantly protected PC12 and SH-SY5Y cells against H2O2 and TNF-α triggered cell death, respectively [127, 128]. In another study, echinacoside prevented dopaminergic neuronal loss in rats injected with 6-OHDA and mitigated the reduction of dopamine and its intermediates [129, 130]. Geng and colleagues demonstrated that echinacoside improved behavioural deficits, prevented loss of dopamine neurons, and reduced the activation of caspase 3/8 in in vivo and in vitro models of PD [131]. Also, Wang and coworkers reported that echinacoside prevented 6-OHDA-triggered loss of dopamine neurons via the attenuation of ROS generation and mitochondrial dysfunction [121]. To elucidate the role of echinacoside in the regulation of GRP78/BiP in an experimental model of PD, Zhang and colleagues revealed that echinacoside attenuated the upregulation of GRP78/BiP in 6-OHDA-treated PC12 cells and Sprague–Dawley rats injected with 6-OHDA, thus highlighting the neurotherapeutic potential of echinacoside in experimental models of PD [132].
Rifampicin is derived from rifamycins, a class of antibiotics obtained from Nocardia mediterranei through a process of fermentation [133]. It is commonly used against Mycobacterium tuberculosis and other mycobacterial infections [133, 134]. Its hydroxyl radical scavenging properties are ascribed to the naphthohydroquinone ring in its chemical structure (Figure 9), while its lipophilic ansa chain is believed to help in its transport into the brain parenchyma across the blood-brain barrier [135, 136]. Pharmacological reports show that rifampicin has immunosuppressive and antioxidant properties [137–139] and inhibits β-amyloid accumulation and neurotoxicity [140]. It also prevents lipopolysaccharide-triggered upregulation of proinflammatory mediators, decreases NF-κB and MAPK signaling [134, 141], attenuates apoptosis in focal ischemic stroke, and inhibits loss of dopaminergic neurons in PD models [133, 142, 143].
Figure 9.
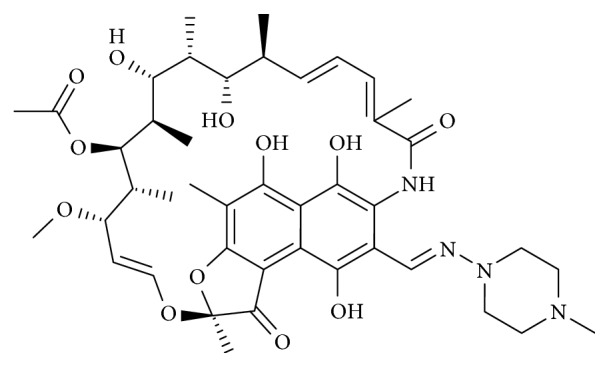
Diagram showing the chemical structure of rifampicin.
To investigate the primary mechanism by which rifampicin promotes neuronal survival, Jing and colleagues revealed a dose-dependent activation of GRP78/BiP in rifampicin-treated PC12 cells [144]. Upon silencing of the GRP78/BiP gene, they investigated if rifampicin-induced GRP78/BiP activation protected against toxicity in rotenone-treated PC12 cells. Western blots and morphological evaluation revealed that cells without the GRP78/BiP gene were more prone to rotenone-triggered damage when compared to cells with the GRP78/BiP gene irrespective of rifampicin treatment [144]. These findings show that silencing of the GRP78/BiP gene mitigated rifampicin-induced protection and thus confirmed that the regulation and activation of GRP78/BiP was responsible for the neuroprotective activity of rifampicin in the PD model.
5. Conclusion
Protein misfolding and aggregation is implicated in the pathogenesis of PD, and the regulation of GRP78/BiP is critical for proper functioning of the UPR. As highlighted in this review, several studies have attempted to unravel the mechanism behind ER stress by targeting GRP78/BiP and the UPR as a way of halting dopaminergic neuronal loss in PD. Although it is established that GRP78/BiP is an essential chaperone in the UPR, studies discussed in this review indicate that the expression of GRP78/BiP is altered in various models of PD depending on the cell type and toxin used in inducing neuronal damage. Consequently, various neuroprotective agents induce the upregulation or downregulation of GRP78/BiP in response to the ER stress-inducing agent in these PD models to promote the survival of dopaminergic neurons. Also, evidence from this review indicate that a translational potential exists for the regulation of GRP78/BiP activity; however, further investigations are needed to properly understand the involvement of GRP78/BiP in the protection of neurons against degeneration in PD. This knowledge would be valuable in designing novel remedies targeted at combating PD and other neurodegenerative disorders linked to the aggregation of misfolded proteins.
Conflicts of Interest
The authors have no conflicts of interest to declare.
References
- 1.Jankovic J. Parkinson’s disease: clinical features and diagnosis. Journal of Neurology, Neurosurgery & Psychiatry. 2008;79(4):368–376. doi: 10.1136/jnnp.2007.131045. [DOI] [PubMed] [Google Scholar]
- 2.Chaudhuri K. R., Sauerbier A. Unravelling the nonmotor mysteries of Parkinson disease. Nature Reviews Neurology. 2016;12(1):10–11. doi: 10.1038/nrneurol.2015.236. [DOI] [PubMed] [Google Scholar]
- 3.Krüger R., Klucken J., Weiss D., et al. Classification of advanced stages of Parkinson’s disease: translation into stratified treatments. Journal of Neural Transmission. 2017;124(8):1015–1027. doi: 10.1007/s00702-017-1707-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Reid D. W., Nicchitta C. V. Diversity and selectivity in mRNA translation on the endoplasmic reticulum. Nature Reviews Molecular Cell Biology. 2015;16(4):221–231. doi: 10.1038/nrm3958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Braakman I., Hebert D. N. Protein folding in the endoplasmic reticulum. Cold Spring Harbor Perspectives in Biology. 2013;5(5) doi: 10.1101/cshperspect.a013201.a013201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Fagone P., Jackowski S. Membrane phospholipid synthesis and endoplasmic reticulum function. Journal of Lipid Research. 2009;50:311–316. doi: 10.1194/jlr.r800049-jlr200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hebert D. N., Garman S. C., Molinari M. The glycan code of the endoplasmic reticulum: asparagine-linked carbohydrates as protein maturation and quality-control tags. Trends in Cell Biology. 2005;15(7):364–370. doi: 10.1016/j.tcb.2005.05.007. [DOI] [PubMed] [Google Scholar]
- 8.Clapham D. E. Calcium signaling. Cell. 2007;131(6):1047–1058. doi: 10.1016/j.cell.2007.11.028. [DOI] [PubMed] [Google Scholar]
- 9.Faitova J., Krekac D., Hrstka R., Vojtesek B. Endoplasmic reticulum stress and apoptosis. Cellular & Molecular Biology Letters. 2006;11(4):p. 488. doi: 10.2478/s11658-006-0040-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ron D., Walter P. Signal integration in the endoplasmic reticulum unfolded protein response. Nature Reviews Molecular Cell Biology. 2007;8(7):519–529. doi: 10.1038/nrm2199. [DOI] [PubMed] [Google Scholar]
- 11.Freeman O. J., Mallucci G. R. The UPR and synaptic dysfunction in neurodegeneration. Brain Research. 2016;1648:530–537. doi: 10.1016/j.brainres.2016.03.029. [DOI] [PubMed] [Google Scholar]
- 12.Omura T., Kaneko M., Okuma Y., Matsubara K., Nomura Y. “Endoplasmic reticulum stress and Parkinson’s disease: the role of HRD1 in averting apoptosis in neurodegenerative disease. Oxidative Medicine and Cellular Longevity. 2013;2013:7. doi: 10.1155/2013/239854.239854 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Tsujii S., Ishisaka M., Hara H. Modulation of endoplasmic reticulum stress in Parkinson’s disease. European Journal of Pharmacology. 2015;765:154–156. doi: 10.1016/j.ejphar.2015.08.033. [DOI] [PubMed] [Google Scholar]
- 14.Jankovic J. Levodopa strengths and weaknesses. Neurology. 2002;58(1):19–32. doi: 10.1212/wnl.58.suppl_1.s19. [DOI] [PubMed] [Google Scholar]
- 15.Jankovic J., Aguilar L. G. Current approaches to the treatment of Parkinson’s disease. Neuropsychiatric Disease and Treatment. 2008;4(4):p. 743. doi: 10.2147/ndt.s2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Weiner W. J., Factor S. A., Jankovic J., et al. The long-term safety and efficacy of pramipexole in advanced Parkinson’s disease. Parkinsonism & Related Disorders. 2001;7(2):115–120. doi: 10.1016/s1353-8020(00)00031-6. [DOI] [PubMed] [Google Scholar]
- 17.Korczyn A. D., Brunt E. R., Larsen J. P., Nagy Z., Poewe W. H., Ruggieri S. A 3-year randomized trial of ropinirole and bromocriptine in early Parkinson’s disease. Neurology. 1999;53(2):p. 364. doi: 10.1212/wnl.53.2.364. [DOI] [PubMed] [Google Scholar]
- 18.Rascol O., Brooks D. J., Korczyn A. D., De Deyn P. P., Clarke C. E., Lang A. E. A five-year study of the incidence of dyskinesia in patients with early Parkinson’s disease who were treated with ropinirole or levodopa. New England Journal of Medicine. 2000;342(20):1484–1491. doi: 10.1056/nejm200005183422004. [DOI] [PubMed] [Google Scholar]
- 19.Ondo W., Hunter C., Almaguer M., Gancher S., Jankovic J. Efficacy and tolerability of a novel sublingual apomorphine preparation in patients with fluctuating Parkinsonʼs disease. Clinical Neuropharmacology. 1999;22(1):1–4. doi: 10.1097/00002826-199901000-00001. [DOI] [PubMed] [Google Scholar]
- 20.Poewe W., Wenning G. K. Apomorphine: an underutilized therapy for Parkinson’s disease. Movement Disorders. 2000;15(5):789–794. doi: 10.1002/1531-8257(200009)15:5<789::aid-mds1005>3.0.co;2-h. [DOI] [PubMed] [Google Scholar]
- 21.Schrag A. Entacapone in the treatment of Parkinson’s disease. The Lancet Neurology. 2005;4(6):366–370. doi: 10.1016/s1474-4422(05)70098-3. [DOI] [PubMed] [Google Scholar]
- 22.Lees A. J., Ratziu V., Tolosa E., Oertel W. H. Safety and tolerability of adjunctive tolcapone treatment in patients with early Parkinson’s disease. Journal of Neurology, Neurosurgery & Psychiatry. 2007;78(9):944–948. doi: 10.1136/jnnp.2006.097154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Saklani A., Kutty S. K. Plant-derived compounds in clinical trials. Drug Discovery Today. 2008;13(3-4):161–171. doi: 10.1016/j.drudis.2007.10.010. [DOI] [PubMed] [Google Scholar]
- 24.More S. V., Kumar H., Kang S. M., Song S.-Y., Lee K., Choi D.-K. Advances in neuroprotective ingredients of medicinal herbs by using cellular and animal models of Parkinson’s disease. Evidence-Based Complementary and Alternative Medicine. 2013;2013:15. doi: 10.1155/2013/957875.957875 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Levy O. A., Malagelada C., Greene L. A. Cell death pathways in Parkinson’s disease: proximal triggers, distal effectors, and final steps. Apoptosis. 2009;14(4):478–500. doi: 10.1007/s10495-008-0309-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Harding H. P., Zhang Y., Ron D. Protein translation and folding are coupled by an endoplasmic-reticulum-resident kinase. Nature. 1999;397(6716):271–274. doi: 10.1038/16729. [DOI] [PubMed] [Google Scholar]
- 27.Rutkowski D. T., Kaufman R. J. A trip to the ER: coping with stress. Trends in Cell Biology. 2004;14(1):20–28. doi: 10.1016/j.tcb.2003.11.001. [DOI] [PubMed] [Google Scholar]
- 28.Shi Y., Vattem K. M., Sood R., et al. Identification and characterization of pancreatic eukaryotic initiation factor 2 α-subunit kinase, PEK, involved in translational control. Molecular and Cellular Biology. 1998;18(12):7499–7509. doi: 10.1128/mcb.18.12.7499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wang M., Wey S., Zhang Y., Ye R., Lee A. S. Role of the unfolded protein response regulator GRP78/BiP in development, cancer, and neurological disorders. Antioxidants & Redox Signaling. 2009;11(9):2307–2316. doi: 10.1089/ars.2009.2485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Calfon M., Zeng H., Urano F., et al. Erratum: IRE1 couples endoplasmic reticulum load to secretory capacity by processing the XBP-1 mRNA. Nature. 2002;415(6867):92–96. doi: 10.1038/415092a. [DOI] [PubMed] [Google Scholar]
- 31.Lee A.-H., Iwakoshi N. N., Glimcher L. H. XBP-1 regulates a subset of endoplasmic reticulum resident chaperone genes in the unfolded protein response. Molecular and Cellular Biology. 2003;23(21):7448–7459. doi: 10.1128/mcb.23.21.7448-7459.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Yoshida H., Matsui T., Yamamoto A., Okada T., Mori K. XBP1 mRNA is induced by ATF6 and spliced by IRE1 in response to ER stress to produce a highly active transcription factor. Cell. 2001;107(7):881–891. doi: 10.1016/s0092-8674(01)00611-0. [DOI] [PubMed] [Google Scholar]
- 33.Ahn T.-B., Jeon B. S. Protective role of heat shock and heat shock protein 70 in lactacystin-induced cell death both in the rat substantia nigra and PC12 cells. Brain Research. 2006;1087(1):159–167. doi: 10.1016/j.brainres.2006.02.097. [DOI] [PubMed] [Google Scholar]
- 34.Dong Z., Wolfer D. P., Lipp H.-P., Büeler H. Hsp70 gene transfer by adeno-associated virus inhibits MPTP-induced nigrostriatal degeneration in the mouse model of Parkinson disease. Molecular Therapy. 2005;11(1):80–88. doi: 10.1016/j.ymthe.2004.09.007. [DOI] [PubMed] [Google Scholar]
- 35.Li J., Lee B., Lee A. S. Endoplasmic reticulum stress-induced apoptosis. Journal of Biological Chemistry. 2006;281(11):7260–7270. doi: 10.1074/jbc.m509868200. [DOI] [PubMed] [Google Scholar]
- 36.Ni M., Lee A. S. ER chaperones in mammalian development and human diseases. FEBS Letters. 2007;581(19):3641–3651. doi: 10.1016/j.febslet.2007.04.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hendershot L. M. The ER function BiP is a master regulator of ER function. The Mount Sinai Journal of Medicine. 2004;71(5):289–297. [PubMed] [Google Scholar]
- 38.Lee A. S. The ER chaperone and signaling regulator GRP78/BiP as a monitor of endoplasmic reticulum stress. Methods. 2005;35(4):373–381. doi: 10.1016/j.ymeth.2004.10.010. [DOI] [PubMed] [Google Scholar]
- 39.Gorbatyuk M. S., Gorbatyuk O. S. The molecular chaperone GRP78/BiP as a therapeutic target for neurodegenerative disorders: a mini review. Journal of Genetic Syndrome & Gene Therapy. 2013;4(2) doi: 10.4172/2157-7412.1000128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Holtz W. A., O’Malley K. L. Parkinsonian mimetics induce aspects of unfolded protein response in death of dopaminergic neurons. Journal of Biological Chemistry. 2003;278(21):19367–19377. doi: 10.1074/jbc.M211821200. [DOI] [PubMed] [Google Scholar]
- 41.Chen G., Bower K. A., Ma C., Fang S., Thiele C. J., Luo J. Glycogen synthase kinase 3β (GSK3β) mediates 6-hydroxydopamine-induced neuronal death. The FASEB Journal. 2004;18(10):1162–1164. doi: 10.1096/fj.04-1551fje. [DOI] [PubMed] [Google Scholar]
- 42.Ghribi O., Herman M. M., Pramoonjago P., Savory J. MPP+ induces the endoplasmic reticulum stress response in rabbit brain involving activation of the ATF-6 and NF-κB signaling pathways. Journal of Neuropathology & Experimental Neurology. 2003;62(11):1144–1153. doi: 10.1093/jnen/62.11.1144. [DOI] [PubMed] [Google Scholar]
- 43.Shimoke K., Kudo M., Ikeuchi T. MPTP-induced reactive oxygen species promote cell death through a gradual activation of caspase-3 without expression of GRP78/Bip as a preventive measure against ER stress in PC12 cells. Life Sciences. 2003;73(5):581–593. doi: 10.1016/s0024-3205(03)00351-5. [DOI] [PubMed] [Google Scholar]
- 44.Duan W., Mattson M. P. Dietary restriction and 2-deoxyglucose administration improve behavioral outcome and reduce degeneration of dopaminergic neurons in models of Parkinson’s disease. Journal of Neuroscience Research. 1999;57(2):195–206. doi: 10.1002/(sici)1097-4547(19990715)57:2<195::aid-jnr5>3.0.co;2-p. [DOI] [PubMed] [Google Scholar]
- 45.Baek J.-H., Whitfield D., Howlett D., et al. Unfolded protein response is activated in lewy body dementias. Neuropathology and Applied Neurobiology. 2016;42(4):352–365. doi: 10.1111/nan.12260. [DOI] [PubMed] [Google Scholar]
- 46.Salganik M., Sergeyev V. G., Shinde V., et al. The loss of glucose-regulated protein 78 (GRP78) during normal aging or from siRNA knockdown augments human alpha-synuclein (α-syn) toxicity to rat nigral neurons. Neurobiology of Aging. 2015;36(6):2213–2223. doi: 10.1016/j.neurobiolaging.2015.02.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Jiang M., Yun Q., Shi F., et al. Downregulation of miR-384-5p attenuates rotenone-induced neurotoxicity in dopaminergic SH-SY5Y cells through inhibiting endoplasmic reticulum stress. American Journal of Physiology-Cell Physiology. 2016;310(9):755–763. doi: 10.1152/ajpcell.00226.2015. [DOI] [PubMed] [Google Scholar]
- 48.Jiang P., Gan M., Lin W.-L., Yen S.-H. C. Nutrient deprivation induces α-synuclein aggregation through endoplasmic reticulum stress response and SREBP2 pathway. Frontiers in Aging Neuroscience. 2014;6:p. 268. doi: 10.3389/fnagi.2014.00268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Gorbatyuk M. S., Shabashvili A., Chen W., et al. Glucose regulated protein 78 diminishes α-synuclein neurotoxicity in a rat model of Parkinson disease. Molecular Therapy. 2012;20(7):1327–1337. doi: 10.1038/mt.2012.28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Cabral-Miranda F., Hetz C. ER stress in neurodegenerative disease: from disease mechanisms to therapeutic interventions. Endoplasmic Reticulum Stress in Diseases. 2017;4(1):11–26. doi: 10.1515/ersc-2017-0002. [DOI] [Google Scholar]
- 51.Vitte J., Traver S., Maués De Paula A., et al. Leucine-rich repeat kinase 2 is associated with the endoplasmic reticulum in dopaminergic neurons and accumulates in the core of Lewy bodies in Parkinson disease. Journal of Neuropathology & Experimental Neurology. 2010;69(9):959–972. doi: 10.1097/nen.0b013e3181efc01c. [DOI] [PubMed] [Google Scholar]
- 52.Yuan Y., Cao P., Smith M. A., et al. Dysregulated LRRK2 signaling in response to endoplasmic reticulum stress leads to dopaminergic neuron degeneration in C. elegans. PloS One. 2011;6(8) doi: 10.1371/journal.pone.0022354.e22354 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Sämann J., Hegermann J., von Gromoff E., Eimer S., Baumeister R., Schmidt E. Caenorhabditits elegansLRK-1 and PINK-1 act antagonistically in stress response and neurite outgrowth. Journal of Biological Chemistry. 2009;284(24):16482–16491. doi: 10.1074/jbc.m808255200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Varma D., Sen D. Role of the unfolded protein response in the pathogenesis of Parkinson’s disease. Acta Neurobiologiae Experimentalis. 2015;75(1):1–26. [PubMed] [Google Scholar]
- 55.Driver J. A., Logroscino G., Gaziano J. M., Kurth T. Incidence and remaining lifetime risk of Parkinson disease in advanced age. Neurology. 2009;72(5):432–438. doi: 10.1212/01.wnl.0000341769.50075.bb. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Collier T. J., Kanaan N. M., Kordower J. H. Ageing as a primary risk factor for Parkinson’s disease: evidence from studies of non-human primates. Nature Reviews Neuroscience. 2011;12(6):359–366. doi: 10.1038/nrn3039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Erickson R. R., Dunning L. M., Holtzman J. L. The effect of aging on the chaperone concentrations in the hepatic, endoplasmic reticulum of male rats: the possible role of protein misfolding due to the loss of chaperones in the decline in physiological function seen with age. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences. 2006;61(5):435–443. doi: 10.1093/gerona/61.5.435. [DOI] [PubMed] [Google Scholar]
- 58.Gavilan M. P., Vela J., Castaño A., et al. Cellular environment facilitates protein accumulation in aged rat hippocampus. Neurobiology of Aging. 2006;27(7):973–982. doi: 10.1016/j.neurobiolaging.2005.05.010. [DOI] [PubMed] [Google Scholar]
- 59.Naidoo N. The endoplasmic reticulum stress response and aging. Reviews in the Neurosciences. 2009;20(1):23–38. doi: 10.1515/revneuro.2009.20.1.23. [DOI] [PubMed] [Google Scholar]
- 60.Bové J., Perier C. Neurotoxin-based models of Parkinson’s disease. Neuroscience. 2012;211:51–76. doi: 10.1016/j.neuroscience.2011.10.057. [DOI] [PubMed] [Google Scholar]
- 61.Joseph J., Cole G., Head E., Ingram D. Nutrition, brain aging, and neurodegeneration. Journal of Neuroscience. 2009;29(41):12795–12801. doi: 10.1523/jneurosci.3520-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Hamazaki T., Hashimoto M. Neuroprotective and ameliorative actions of polyunsaturated fatty acids against neuronal diseases-evidence from basic to clinical studies: preface. Journal of Pharmacological Sciences. 2011;116(2):p. 149. doi: 10.1254/jphs.11r02fm. [DOI] [PubMed] [Google Scholar]
- 63.Sun T., Xu Z., Wu C.-T., Janes M., Prinyawiwatkul W., No H. K. Antioxidant activities of different colored sweet bell peppers (Capsicum annuum L.) Journal of Food Science. 2007;72(2):98–102. doi: 10.1111/j.1750-3841.2006.00245.x. [DOI] [PubMed] [Google Scholar]
- 64.Nabavi S. F., Braidy N., Gortzi O., et al. Luteolin as an anti-inflammatory and neuroprotective agent: a brief review. Brain Research bulletin. 2015;119:1–11. doi: 10.1016/j.brainresbull.2015.09.002. [DOI] [PubMed] [Google Scholar]
- 65.Seelinger G., Merfort I., Wölfle U., Schempp C. Anti-carcinogenic effects of the flavonoid luteolin. Molecules. 2008;13(10):2628–2651. doi: 10.3390/molecules13102628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Middleton E., Kandaswami C., Theoharides T. C. The effects of plant flavonoids on mammalian cells: implications for inflammation, heart disease, and cancer. Pharmacological Reviews. 2000;52(4):673–751. [PubMed] [Google Scholar]
- 67.Lopez-Lazaro M. Distribution and biological activities of the flavonoid luteolin. Mini-Reviews in Medicinal Chemistry. 2009;9(1):31–59. doi: 10.2174/138955709787001712. [DOI] [PubMed] [Google Scholar]
- 68.Dirscherl K., Karlstetter M., Ebert S., et al. Luteolin triggers global changes in the microglial transcriptome leading to a unique anti-inflammatory and neuroprotective phenotype. Journal of Neuroinflammation. 2010;7(1):p. 3. doi: 10.1186/1742-2094-7-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Lin Y., Shi R., Wang X., Shen H.-M. Luteolin, a flavonoid with potential for cancer prevention and therapy. Current Cancer Drug Targets. 2008;8(7):634–646. doi: 10.2174/156800908786241050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Liu R., Gao M., Qiang G.-F., et al. The anti-amnesic effects of luteolin against amyloid β25-35 peptide-induced toxicity in mice involve the protection of neurovascular unit. Neuroscience. 2009;162(4):1232–1243. doi: 10.1016/j.neuroscience.2009.05.009. [DOI] [PubMed] [Google Scholar]
- 71.Guo D.-J., Li F., Yu P. H.-F., Chan S.-W. Neuroprotective effects of luteolin against apoptosis induced by 6-hydroxydopamine on rat pheochromocytoma PC12 cells. Pharmaceutical Biology. 2013;51(2):190–196. doi: 10.3109/13880209.2012.716852. [DOI] [PubMed] [Google Scholar]
- 72.Pandurangan A. K., Esa N. M. Luteolin, a bioflavonoid inhibits colorectal cancer through modulation of multiple signaling pathways: a review. Asian Pacific Journal of Cancer Prevention. 2014;15(14):5501–5508. doi: 10.7314/apjcp.2014.15.14.5501. [DOI] [PubMed] [Google Scholar]
- 73.Hu L.-W., Yen J.-H., Shen Y.-T., Wu K.-Y., Wu M.-J. Luteolin modulates 6-hydroxydopamine-induced transcriptional changes of stress response pathways in PC12 cells. PLoS One. 2014;9(5) doi: 10.1371/journal.pone.0097880.e97880 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Grech-Baran M., Sykłowska-Baranek K., Pietrosiuk A. Biotechnological approaches to enhance salidroside, rosin and its derivatives production in selected Rhodiola spp. in vitro cultures. Phytochemistry Reviews. 2015;14(4):657–674. doi: 10.1007/s11101-014-9368-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Zhang B., Li Q., Chu X., Sun S., Chen S. “Salidroside reduces tau hyperphosphorylation via up-regulating GSK-3β phosphorylation in a tau transgenic drosophila model of Alzheimer’s disease. Translational Neurodegeneration. 2016;5(1):p. 21. doi: 10.1186/s40035-016-0068-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Luo X., Bao N., Chen L., Sun J. Pharmacological activities and progress in structure modification of salidroside. Medicinal Chemistry. 2017;7(3):818–823. doi: 10.4172/2161-0444.1000434. [DOI] [Google Scholar]
- 77.Guan S., Feng H., Song B., et al. Salidroside attenuates LPS-induced pro-inflammatory cytokine responses and improves survival in murine endotoxemia. International Immunopharmacology. 2011;11(12):2194–2199. doi: 10.1016/j.intimp.2011.09.018. [DOI] [PubMed] [Google Scholar]
- 78.Wang J., Li J.-Z., Lu A.-X., Zhang K.-F., Li B.-J. Anticancer effect of salidroside on A549 lung cancer cells through inhibition of oxidative stress and phospho-p38 expression. Oncology Letters. 2014;7(4):1159–1164. doi: 10.3892/ol.2014.1863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Zhang B., Wang Y., Li H., et al. “Neuroprotective effects of salidroside through PI3K/Akt pathway activation in Alzheimer’s disease models. Drug Design, Development and Therapy. 2016;10:p. 1335. doi: 10.2147/DDDT.S99958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Skopińska-Rózewska E., Malinowski M., Wasiutyński A., et al. The influence of rhodiola quadrifida 50% hydro-alcoholic extract and salidroside on tumor-induced angiogenesis in mice. Polish Journal of Veterinary Sciences. 2008;11(2):97–104. [PubMed] [Google Scholar]
- 81.Jin H., Pei L., Shu X., et al. Therapeutic intervention of learning and memory decays by salidroside stimulation of neurogenesis in aging. Molecular Neurobiology. 2016;53(2):851–866. doi: 10.1007/s12035-014-9045-6. [DOI] [PubMed] [Google Scholar]
- 82.Li X., Ye X., Li X., et al. Salidroside protects against MPP+-induced apoptosis in PC12 cells by inhibiting the NO pathway. Brain Research. 2011;1382:9–18. doi: 10.1016/j.brainres.2011.01.015. [DOI] [PubMed] [Google Scholar]
- 83.Wang S., He H., Chen L., Zhang W., Zhang X., Chen J. Protective effects of salidroside in the MPTP/MPP+ induced model of Parkinson’s disease through ROS-NO-related mitochondrion pathway. Molecular Neurobiology. 2015;51(2):718–728. doi: 10.1007/s12035-014-8755-0. [DOI] [PubMed] [Google Scholar]
- 84.Tao K., Wang B., Feng D., et al. Salidroside protects against 6-hydroxydopamine-induced cytotoxicity by attenuating ER stress. Neuroscience Bulletin. 2016;32(1):61–69. doi: 10.1007/s12264-015-0001-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.She G. M., Xu C., Liu B., Shi R. B. Polyphenolic acids from mint (the aerial of Mentha haplocalyx Briq.) with DPPH radical scavenging activity. Journal of Food Science. 2010;75(4):359–362. doi: 10.1111/j.1750-3841.2010.01603.x. [DOI] [PubMed] [Google Scholar]
- 86.Kamata K., Noguchi M., Nagai M. Hypotensive effects of lithospermic acid B isolated from the extract of Salviae miltiorrhizae Radix in the rat. General Pharmacology: The Vascular System. 1994;25(1):69–73. doi: 10.1016/0306-3623(94)90011-6. [DOI] [PubMed] [Google Scholar]
- 87.Soung D. Y., Rhee S. H., Kim J. S., et al. Peroxynitrite scavenging activity of lithospermate B from Salvia miltiorrhiza. Journal of Pharmacy and Pharmacology. 2003;55(10):1427–1432. doi: 10.1211/0022357021891. [DOI] [PubMed] [Google Scholar]
- 88.Lee B.-W., Chun S. W., Kim S. H., et al. Lithospermic acid B protects beta-cells from cytokine-induced apoptosis by alleviating apoptotic pathways and activating anti-apoptotic pathways of Nrf2-HO-1 and Sirt1. Toxicology and Applied Pharmacology. 2011;252(1):47–54. doi: 10.1016/j.taap.2011.01.018. [DOI] [PubMed] [Google Scholar]
- 89.Lin Y.-L., Tsay H.-J., Lai T.-H., Tzeng T.-T., Shiao Y.-J. Lithospermic acid attenuates 1-methyl-4-phenylpyridine-induced neurotoxicity by blocking neuronal apoptotic and neuroinflammatory pathways. Journal of Biomedical Science. 2015;22(1):p. 37. doi: 10.1186/s12929-015-0146-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Baird A. Fibroblast growth factors: activities and significance of non-neurotrophin neurotrophic growth factors. Current Opinion in Neurobiology. 1994;4(1):78–86. doi: 10.1016/0959-4388(94)90035-3. [DOI] [PubMed] [Google Scholar]
- 91.Bikfalvi A., Klein S., Pintucci G., Rifkin D. B. Biological roles of fibroblast growth factor-2. Endocrine Reviews. 1997;18(1):26–45. doi: 10.1210/er.18.1.26. [DOI] [PubMed] [Google Scholar]
- 92.Roceri M., Molteni R., Fumagalli F., et al. Stimulatory role of dopamine on fibroblast growth factor-2 expression in rat striatum. Journal of Neurochemistry. 2001;76(4):990–997. doi: 10.1046/j.1471-4159.2001.00088.x. [DOI] [PubMed] [Google Scholar]
- 93.Itoh N., Ornitz D. M. Fibroblast growth factors: from molecular evolution to roles in development, metabolism and disease. Journal of Biochemistry. 2011;149(2):121–130. doi: 10.1093/jb/mvq121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Beenken A., Mohammadi M. The FGF family: biology, pathophysiology and therapy. Nature Reviews Drug Discovery. 2009;8(3):235–253. doi: 10.1038/nrd2792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Xiao J., Lv Y., Lin S., et al. Cardiac protection by basic fibroblast growth factor from ischemia/reperfusion-induced injury in diabetic rats. Biological & Pharmaceutical Bulletin. 2010;33(3):444–449. doi: 10.1248/bpb.33.444. [DOI] [PubMed] [Google Scholar]
- 96.Hsuan S.-L., Klintworth H. M., Xia Z. Basic fibroblast growth factor protects against rotenone-induced dopaminergic cell death through activation of extracellular signal-regulated kinases 1/2 and phosphatidylinositol-3 kinase pathways. Journal of Neuroscience. 2006;26(17):4481–4491. doi: 10.1523/jneurosci.4922-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Chadi G., Silva C., Maximino J. R., Fuxe K., da Silva G. O. Adrenalectomy counteracts the local modulation of astroglial fibroblast growth factor system without interfering with the pattern of 6-OHDA-induced dopamine degeneration in regions of the ventral midbrain. Brain Research. 2008;1190:23–38. doi: 10.1016/j.brainres.2007.11.024. [DOI] [PubMed] [Google Scholar]
- 98.Cai P., Ye J., Zhu J., et al. Inhibition of endoplasmic reticulum stress is involved in the neuroprotective effect of bFGF in the 6-OHDA-induced Parkinson’s disease model. Aging and Disease. 2016;7(4):p. 336. doi: 10.14336/ad.2016.0117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Zhang Y., Zeng Z., Cao Y., Du X., Wan Z. Effect of urinary protease inhibitor (ulinastatin) on cardiopulmonary bypass: a meta-analysis for China and Japan. Plos One. 2014;9(12) doi: 10.1371/journal.pone.0086397.e113973 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Liu W., Pang G., Wang S., Sun A. Protective effect of ulinastatin on severe pulmonary infection under immunosuppression and its molecular mechanism. Experimental and Therapeutic Medicine. 2017;14(4):3583–3588. doi: 10.3892/etm.2017.4993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Shigetomi H., Onogi A., Kajiwara H., et al. Anti-inflammatory actions of serine protease inhibitors containing the Kunitz domain. Inflammation Research. 2010;59(9):679–687. doi: 10.1007/s00011-010-0205-5. [DOI] [PubMed] [Google Scholar]
- 102.Chen H., He M.-y., Li Y.-m. Treatment of patients with severe sepsis using ulinastatin and thymosin alpha1: a prospective, randomized, controlled pilot study. Chinese Medical Journal. 2009;122(8):883–888. [PubMed] [Google Scholar]
- 103.Kanai T., Ishiwata T., Kobayashi T., et al. Ulinastatin, a urinary trypsin inhibitor, for the initial treatment of patients with Kawasaki disease: a retrospective study. Circulation, Circulationaha. 2011;111 doi: 10.1161/CIRCULATIONAHA.111.028423.028423 [DOI] [PubMed] [Google Scholar]
- 104.Lagoo J. Y., D’Souza M. C., Kartha A., Kutappa A. M. Role of ulinastatin, a trypsin inhibitor, in severe acute pancreatitis in critical care setting: a retrospective analysis. Journal of Critical Care. 2018;45:27–32. doi: 10.1016/j.jcrc.2018.01.021. [DOI] [PubMed] [Google Scholar]
- 105.Sun R., Li Y., Chen W., Zhang F., Li T. Total ginsenosides synergize with ulinastatin against septic acute lung injury and acute respiratory distress syndrome. International Journal of Clinical and Experimental Pathology. 2015;8(6):p. 7385. [PMC free article] [PubMed] [Google Scholar]
- 106.Atal S. S., Atal S. Ulinastatin–a newer potential therapeutic option for multiple organ dysfunction syndrome. Journal of Basic and Clinical Physiology and Pharmacology. 2016;27(2):91–99. doi: 10.1515/jbcpp-2015-0003. [DOI] [PubMed] [Google Scholar]
- 107.Li H.-f., Zhao S.-x., Xing B.-p., Sun M.-l. Ulinastatin suppresses endoplasmic reticulum stress and apoptosis in the hippocampus of rats with acute paraquat poisoning. Neural Regeneration Research. 2015;10(3):p. 467. doi: 10.4103/1673-5374.153698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Boyce M., Py B. F., Ryazanov A. G., et al. A pharmacoproteomic approach implicates eukaryotic elongation factor 2 kinase in ER stress-induced cell death. Cell Death & Differentiation. 2008;15(3):589–599. doi: 10.1038/sj.cdd.4402296. [DOI] [PubMed] [Google Scholar]
- 109.Nakka V. P., Gusain A., Raghubir R. Endoplasmic reticulum stress plays critical role in brain damage after cerebral ischemia/reperfusion in rats. Neurotoxicity Research. 2010;17(2):189–202. doi: 10.1007/s12640-009-9110-5. [DOI] [PubMed] [Google Scholar]
- 110.Boyce M., Bryant K. F., Jousse C., et al. A selective inhibitor of eIF2 dephosphorylation protects cells from ER stress. Science. 2005;307(5711):935–939. doi: 10.1126/science.1101902. [DOI] [PubMed] [Google Scholar]
- 111.Wiseman R. L., Balch W. E. A new pharmacology-drugging stressed folding pathways. Trends in Molecular Medicine. 2005;11(8):347–350. doi: 10.1016/j.molmed.2005.06.011. [DOI] [PubMed] [Google Scholar]
- 112.Paschen W. Endoplasmic reticulum: a primary target in various acute disorders and degenerative diseases of the brain. Cell Calcium. 2003;34(4-5):365–383. doi: 10.1016/s0143-4160(03)00139-8. [DOI] [PubMed] [Google Scholar]
- 113.Gasparetto M., Gentry T., Sebti S., et al. Identification of compounds that enhance the anti-lymphoma activity of rituximab using flow cytometric high-content screening. Journal of Immunological Methods. 2004;292(1-2):59–71. doi: 10.1016/j.jim.2004.06.003. [DOI] [PubMed] [Google Scholar]
- 114.Chou Y. H., Chao P. L., Tsai M. J., et al. Arsenite-induced cytotoxicity in dorsal root ganglion explants. Free Radical Biology and Medicine. 2008;44(8):1553–1561. doi: 10.1016/j.freeradbiomed.2007.12.014. [DOI] [PubMed] [Google Scholar]
- 115.Guo H., Jiang C., Sun X. Therapeutical effects and mechanism of salubrinal combined with ulinastatin on treating paraquat poisoning. Cell Biochemistry and Biophysics. 2014;70(3):1559–1563. doi: 10.1007/s12013-014-0095-1. [DOI] [PubMed] [Google Scholar]
- 116.Huang Y., Xu J., Liang M., et al. RESP18 is involved in the cytotoxicity of dopaminergic neurotoxins in MN9D cells. Neurotoxicity Research. 2013;24(2):164–175. doi: 10.1007/s12640-013-9375-6. [DOI] [PubMed] [Google Scholar]
- 117.Long K., Boyce M., Lin H., Yuan J., Ma D. Structure-activity relationship studies of salubrinal lead to its active biotinylated derivative. Bioorganic & Medicinal Chemistry Letters. 2005;15(17):3849–3852. doi: 10.1016/j.bmcl.2005.05.120. [DOI] [PubMed] [Google Scholar]
- 118.Yang W., Tiffany-Castiglioni E., Koh H. C., Son I.-H. Paraquat activates the IRE1/ASK1/JNK cascade associated with apoptosis in human neuroblastoma SH-SY5Y cells. Toxicology Letters. 2009;191(2-3):203–210. doi: 10.1016/j.toxlet.2009.08.024. [DOI] [PubMed] [Google Scholar]
- 119.Goswami P., Gupta S., Biswas J., et al. Endoplasmic reticulum stress plays a key role in rotenone-induced apoptotic death of neurons. Molecular Neurobiology. 2016;53(1):285–298. doi: 10.1007/s12035-014-9001-5. [DOI] [PubMed] [Google Scholar]
- 120.Lei L., Yang F., Zhang T., Tu P., Wu L., Ito Y. Preparative isolation and purification of acteoside and 2’-acetyl acteoside from Cistanches salsa (C.A. Mey.) G. Beck by high-speed counter-current chromatography. Journal of Chromatography A. 2001;912(1):181–185. doi: 10.1016/s0021-9673(01)00583-0. [DOI] [PubMed] [Google Scholar]
- 121.Wang Y.-H., Xuan Z.-H., Tian S., Du G.-H. Echinacoside protects against 6-hydroxydopamine-induced mitochondrial dysfunction and inflammatory responses in PC12 cells via reducing ROS production. Evidence-Based Complementary and Alternative Medicine. 2015;2015:9. doi: 10.1155/2015/189239.189239 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Sloley B. D., Urichuk L. J., Tywin C., Coutts R. T., Pang P. K. T., Shan J. J. Comparison of chemical components and antioxidant capacity of different Echinacea species. Journal of Pharmacy and Pharmacology. 2001;53(6):849–857. doi: 10.1211/0022357011776009. [DOI] [PubMed] [Google Scholar]
- 123.Dalby-Brown L., Barsett H., Landbo A.-K. R., Meyer A. S., Mølgaard P. Synergistic antioxidative effects of alkamides, caffeic acid derivatives, and polysaccharide fractions from Echinacea purpureaon in vitro oxidation of human low-density lipoproteins. Journal of Agricultural and Food Chemistry. 2005;53(24):9413–9423. doi: 10.1021/jf0502395. [DOI] [PubMed] [Google Scholar]
- 124.Xiong Q., Tezuka Y., Kaneko T., et al. Inhibition of nitric oxide by phenylethanoids in activated macrophages. European Journal of Pharmacology. 2000;400(1):137–144. doi: 10.1016/s0014-2999(00)00354-x. [DOI] [PubMed] [Google Scholar]
- 125.Wu Y., Li L., Wen T., Li Y.-Q. Protective effects of echinacoside on carbon tetrachloride-induced hepatotoxicity in rats. Toxicology. 2007;232(1-2):50–56. doi: 10.1016/j.tox.2006.12.013. [DOI] [PubMed] [Google Scholar]
- 126.He W., Fang T., Tu P. Research progress on pharmacological activities of echinacoside. China Journal of Chinese Materia Medica. 2009;34(4):476–479. doi: 10.4268/cjcmm20160203. [DOI] [PubMed] [Google Scholar]
- 127.Kuang R., Sun Y., Yuan W., Lei L., Zheng X., Food Z. Protective effects of echinacoside, one of the phenylethanoid glycosides, on H2 O2-induced cytotoxicity in PC12 cells. Planta Medica. 2009;75(14):1499–1504. doi: 10.1055/s-0029-1185806. [DOI] [PubMed] [Google Scholar]
- 128.Deng M., Zhao J. Y., Tu P. F., Jiang Y., Li Z. B., Wang Y. H. Echinacoside rescues the SHSY5Y neuronal cells from TNFα-induced apoptosis. European Journal of Pharmacology. 2004;505(1-3):11–18. doi: 10.1016/j.ejphar.2004.09.059. [DOI] [PubMed] [Google Scholar]
- 129.Chen H., Jing F. C., Li C. L., Tu P. F., Zheng Q. S., Wang Z. H. Echinacoside prevents the striatal extracellular levels of monoamine neurotransmitters from diminution in 6-hydroxydopamine lesion rats. Journal of Ethnopharmacology. 2007;114(3):285–289. doi: 10.1016/j.jep.2007.07.035. [DOI] [PubMed] [Google Scholar]
- 130.Chen L.-W., Wang Y.-Q., Wei L.-C., Shi M., Chan Y.-S. Chinese herbs and herbal extracts for neuroprotection of dopaminergic neurons and potential therapeutic treatment of Parkinson’s disease. CNS & Neurological Disorders-Drug Targets (Formerly Current Drug Targets-CNS & Neurological Disorders) 2007;6(4):273–281. doi: 10.2174/187152707781387288. [DOI] [PubMed] [Google Scholar]
- 131.Geng X., Tian X., Tu P., Pu X. Neuroprotective effects of echinacoside in the mouse MPTP model of Parkinson’s disease. European Journal of Pharmacology. 2007;564(1-3):66–74. doi: 10.1016/j.ejphar.2007.01.084. [DOI] [PubMed] [Google Scholar]
- 132.Zhang Y., Long H., Zhou F., et al. Echinacoside’s nigrostriatal dopaminergic protection against 6-OHDA-induced endoplasmic reticulum stress through reducing the accumulation of seipin. Journal of Cellular and Molecular Medicine. 2017;21(12):3761–3775. doi: 10.1111/jcmm.13285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Yulug B., Kilic Ü., Kilic E., Bähr M. Rifampicin attenuates brain damage in focal ischemia. Brain Research. 2004;996(1):76–80. doi: 10.1016/j.brainres.2003.10.012. [DOI] [PubMed] [Google Scholar]
- 134.Bi W., Zhu L., Jing X., et al. Rifampicin improves neuronal apoptosis in LPS-stimulated co-cultured BV2 cells through inhibition of the TLR-4 pathway. Molecular Medicine Reports. 2014;10(4):1793–1799. doi: 10.3892/mmr.2014.2480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Mindermann T., Landolt H., Zimmerli W., Rajacic Z., Gratzl O. Penetration of rifampicin into the brain tissue and cerebral extracellular space of rats. Journal of Antimicrobial Chemotherapy. 1993;31(5):731–737. doi: 10.1093/jac/31.5.731. [DOI] [PubMed] [Google Scholar]
- 136.Kilic L., Kilic E., Lingor P., Yulug B., Bähr M. Rifampicin inhibits neurodegeneration in the optic nerve transection model in vivo and after 1-methyl-4-phenylpyridinium intoxication in vitro. Acta Neuropathologica. 2004;108(1):65–68. doi: 10.1007/s00401-004-0867-6. [DOI] [PubMed] [Google Scholar]
- 137.Nilsson B. S. Rifampicin: an immunosuppressant? The Lancet. 1971;298(7720):p. 374. doi: 10.1016/s0140-6736(71)90087-0. [DOI] [PubMed] [Google Scholar]
- 138.Dajani B. M., Canady M. S., Thompson J. S., Kasik J. E. Rifampicin: an immunosuppressant? The Lancet. 1972;300(7786):p. 1094. doi: 10.1016/s0140-6736(72)92390-2. [DOI] [PubMed] [Google Scholar]
- 139.Yulug B., Hanoglu L., Kilic E., Schabitz W. R. Rifampicin: an antibiotic with brain protective function. Brain Research Bulletin. 2014;107:37–42. doi: 10.1016/j.brainresbull.2014.05.007. [DOI] [PubMed] [Google Scholar]
- 140.Namba Y., Kawatsu K., Izumi S., Ueki A., Ikeda K. Neurofibrillary tangles and senile plaques in brain of elderly leprosy patients. The Lancet. 1992;340(8825):p. 978. doi: 10.1016/0140-6736(92)92870-l. [DOI] [PubMed] [Google Scholar]
- 141.Bi W., Zhu L., Wang C., et al. Rifampicin inhibits microglial inflammation and improves neuron survival against inflammation. Brain Research. 2011;1395:12–20. doi: 10.1016/j.brainres.2011.04.019. [DOI] [PubMed] [Google Scholar]
- 142.Oida Y., Kitaichi K., Nakayama H., et al. Rifampicin attenuates the MPTP-induced neurotoxicity in mouse brain. Brain Research. 2006;1082(1):196–204. doi: 10.1016/j.brainres.2006.01.116. [DOI] [PubMed] [Google Scholar]
- 143.Chen S., Sun Y., Zeng Z., Tao E. Rifampicin inhibits apoptosis in rotenone-induced differentiated Pc12 cells by ameliorating mitochondrial oxidative stress∗⋆. Neural Regeneration Research. 2010;5(4):251–256. [Google Scholar]
- 144.Jing X., Shi Q., Bi W., et al. Rifampicin protects PC12 cells from rotenone-induced cytotoxicity by activating GRP78 via PERK-eIF2α-ATF4 pathway. PloS One. 2014;9(3) doi: 10.1371/journal.pone.0092110.e92110 [DOI] [PMC free article] [PubMed] [Google Scholar]