

Abstract
Introduction
Arboviral diseases including dengue are increasingly spreading in the tropical/subtropical world including Africa. Updated knowledge on the distribution and abundance of the major vectors Aedes aegypti and Aedes albopictus constitutes crucial surveillance action to prepare African countries such as Cameroon for potential arbovirus outbreaks. Here, we present a nationwide survey in Cameroon to assess the current geographical distribution and prevalence of both vectors including a genetic diversity profiling of Ae. albopictus (invasive species) using mitochondrial DNA.
Methods
Immature stages of Aedes were collected between March and August 2017 in 29 localities across Cameroon following north-south and east-west transects. Larvae and pupae were collected from several containers in each location, reared to adult and morphologically identified. Genetic diversity of Ae. albopictus from 16 locations were analysed using Cytochrome Oxidase I gene (COI).
Results
In total, 30,381 immature stages of Aedes with an average of 646.40±414.21 per location were identified across the country comprising 69.3% of Ae. albopictus and 30.7% of Ae. aegypti. Analysis revealed that Ae. aegypti is still distributed nation widely whereas Ae. albopictus is limited to the southern part, around 6°4’N. However, Ae. albopictus is the most prevalent species in all southern locations where both species are sympatric except in Douala where Ae. aegypti is predominant. This suggests that factors such as climate, vegetation, and building density impact the distribution of both species in Cameroon. Mitochondrial DNA analysis revealed a low genetic diversity in Ae. albopictus populations with a major common haplotype resulting in low haplotype diversity ranging from 0.13 to 0.65 and 0.35 for the total sample. Similarly, low nucleotide diversity was also reported varying from 0.0000 to 0.0017 with an overall index of 0.0008. This low genetic polymorphism is consistent with the recent introduction of Ae. albopictus in Cameroon.
Conclusion
This updated distribution of arbovirus vectors across Cameroon will help in planning vector control programme against possible outbreak of arbovirus related diseases in the country.
Author summary
Aedes albopictus and Ae. aegypti are the most important arbovirus vectors worldwide. Ae. albopictus, native of Asia, was recorded for the first time in early 2000s in Cameroon, central Africa. Previous studies performed a decade ago in Cameroon showed that Ae. albopictus has a geographical distribution limited to the south under 6°N. Whereas the native species Ae. aegypti was present across the country. To update our knowledge in this regards, a nationwide survey was performed in Cameroon to assess the current geographical distribution and prevalence of both vectors including a genetic diversity profiling of Ae. albopictus (invasive species) using mitochondrial DNA. Analysis revealed that Ae. aegypti is still distributed nation widely whereas Ae. albopictus is limited to the southern part, around 6°4’N. However, Ae. albopictus is the most prevalent species in all southern locations where both species are sympatric except in Douala where Ae. aegypti is predominant. This suggests that factors such as climate, vegetation and building density impact the distribution of both species in Cameroon. Mitochondrial DNA analysis revealed a low genetic diversity in Ae. albopictus populations with a major common haplotype detected in almost all locations. This study provides the relevant data that can be helpful to establish the vector surveillance of epidemic arbovirus vectors across the country.
Introduction
Aedes-borne arboviral diseases such as dengue, Zika, and chikungunya are increasing global public health problems due to their rapid geographical spread and rising disease burden [1,2]. The viruses causing these infections are transmitted to vertebrates, including human, mainly by bites of infected mosquito belonging to the Aedes genus. However, transmission by blood transfusion and/or by sexual contacts have also been reported in Zika virus [3]. Two main epidemic vectors of these arboviral diseases, Aedes aegypti Linnaeus 1762 and Aedes albopictus (Skuse) 1894, have different origins. Aedes aegypti, originating from African forests, is currently found in most tropical and subtropical regions worldwide [4,5]. While Ae. albopictus originating from South Asia forest has invaded all the five continents during the past 30–40 years [6,7]. This rapid spread of Ae. albopictus has been mainly facilitated by international commercial exchanges notably international trade of used tires as previously demonstrated [8], and strong ecological and physiological plasticity allowing it to thrive in a wide range of climates and habitats [9]. This invading species was recorded for the first time in central Africa in Cameroon in the early 2000s [10] and became the dominant species over the native species Ae. aegypti in sympatric areas [11]. Interestingly, dengue, Zika, and chikungunya, which were considered to be scarce in central Africa, are emerging in several urban foci simultaneously with the introduction of Ae. albopictus [12–14] suggesting a modification of epidemiology of arboviral diseases. During the last two decades, the circulation of arboviral diseases has been well documented in Cameroon [14–20], but nationwide studies have not been undertaken to get an accurate picture of distribution of these viruses across the country. Meanwhile, the seroprevalence of dengue in general population was assessed in three main cities of Cameroon located in different ecological settings in 2006/2007. This study revealed that 61.4% of people tested had immunoglobulin (Ig) G and 0.3% IgM in Douala, 24.2% IgG and 0.1% IgM in Garoua and 9.8% IgG and no IgM in Yaoundé [21]. More recently, another study in blood donors in population from six cities across Cameroon revealed that the overall seroprevalence of Zika was low around 5%, peaking at 10% and 7.7% in Douala and Bertoua respectively and only 2% in Ngaoundere and Maroua [22]. Indeed, Ae. albopictus has been detected as the main vector in Gabon during concurrent dengue/chikungunya outbreak in 2007 [23,24]. Both Ae. aegypti and Ae. albopictus have been found infected by chikungunya virus during a large outbreak that occurred in the Republic of the Congo in 2011 [25].
The coexistence of both Ae. aegypti and Ae. albopictus has been well documented in several regions throughout the world. In central Africa, both species often share the same larval habitats, although Ae. albopictus preferred man-made containers such as used tires and discarded tanks surrounded by the presence of vegetation whereas Ae. aegypti preferred larval habitats located in a neighborhood with a high building density [26,27]. Competitive displacement between both species has also been well studied in South America and South East Asia revealing that the invasive species often have the competitive advantage over the native species. For example, in Asia, the overall advantage of Ae. aegypti over the resident species Ae. albopictus has been reported [28,29]. In contrast, replacement of Ae. aegypti by invasive species Ae. albopictus was reported in south-eastern USA and Brazil [30–32] and was suspected in Réunion [33] and Mayotte [34]. In central Africa, the dominance of the invading species Ae. albopictus over the native species Ae. aegypti was also reported in the locations where both species were found together [24,35]. The last study conducted in Cameroon on the geographical distribution of Ae. aegypti and Ae. albopictus date more than 10 years. The results of the study revealed that Ae. aegypti was present across the country while the distribution of Ae. albopictus was limited to the south, under 6°N [36,37]. Climatic limitations and the dynamics of invasion have been used to explain the absence of Ae. albopictus beyond 6°N but no temporal study has been performed to assess the dynamic of this distribution and establish whether this species could spread further northwards increasing the risk of arbovirus transmission. Ae. albopictus being more competent to transmit dengue, chikungunya and probably Zika in central Africa [11], it is important to properly define the vector composition in this region to adequately prepare for future outbreaks. This requires updating the data on the geographical distribution and prevalence of Ae. aegypti and Ae. albopictus country-wide. Furthermore, analysis of the genetic diversity of the invasive species is also needed to characterize the population demographic history of this species in Cameroon since its introduction. Previous studies analysing the genetic diversity of Ae. albopictus in Central Africa using the cytochrome oxidase subunit I (COI) gene had revealed a low polymorphism and showed that Cameroonians’ population are related to tropical rather than temperate or subtropical out-groups [9,35]. It remains to establish how this genetic diversity has evolved in the past decade and whether the population of this species has experienced demographic events such as expansion, new colonisation or bottleneck. Here, we present an extensive and nationwide analysis of the current geographical distribution and prevalence of Ae. aegypti and Ae. albopictus in Cameroon as well as the genetic diversity of the invading Ae. albopictus species to improve the entomological surveillance of these vectors.
Materials and methods
Ethics statement
This study was approved by the Cameroonian national ethics committee for human health research N°2017/05/911/CE/CNERSH/SP. Oral consent to inspect the potential breeding sites was obtained in the field in household or garage owners.
Study sites
Mosquitoes were collected in 28 locations across Cameroon, a central African country located between 1°40–13°05N and 8°30–16°10E (Table 1 and Fig 1), following north-south and east-west transects. Cameroon is characterized by a broad range of biotopes varying from the sudano-sahelian climate in the far north to the equatorial guinean forest climate in the south with strong local climate heterogeneities due to huge variations in altitude (0 to 4,000 m above sea level on Mount Cameroon) [36]. The annual precipitation rate vary from 400 mm in the arid areas to 10,000 mm at the foot of mount-Cameroon (>4,000m above sea level). Annual temperatures and relative humidity vary between 18°C to 28°C and 85% to 45%, respectively [38] (Table 1). Mosquito sampling focused mainly on urban settlements that spread along the trade routes throughout the country. This is because invading Aedes species has been directly linked to human activities [39] and important outbreaks usually occur in urban settings.
Table 1. Environmental characteristics of different prospected sites in Cameroon.
| Localities | Lat (N) | Long (E) | Climate | Vegetation | Population |
|---|---|---|---|---|---|
| Maroua | 10°59' | 14°32' | Sahelian Sudano climate | Dense herbaceous savanna | 33,0410 |
| Garoua | 9°32 | 13°39' | Sudanian tropical basin | Tree savanna with shrub deciduous | 26,5583 |
| Mbe | 7°86' | 13°59' | Tropical humid altitude | Tree savanna with shrub deciduous | 17,478 |
| Ngaoundere | 7°33’ | 13°56' | Tropical humid altitude | Wooded savannah deciduous | 26,2747 |
| Banyo | 6°74' | 11°80' | Tropical humid altitude | Wooded savannah deciduous | 93,880 |
| Meiganga | 6°51' | 14°29' | Tropical humid altitude | Post-forest transition and savannah | 88,745 |
| Tibati | 6°47' | 12°61' | Tropical altitude | Post-forest transition and savannah | 72,081 |
| Mayo-Darle | 6°46' | 11°54' | Tropical humid altitude | Wooded savannah deciduous | 23,054 |
| Bankim | 6°08' | 11°48' | Tropical altitude | Savannah forest mosaic | 70,132 |
| Bamenda | 5°96' | 10'15' | Equatorial mountain monsoon climate | Savannah forest mosaic | 322,889 |
| Garoua-Boulai | 5°89' | 14°54' | Guinean subequatorial climate | Savannah forest mosaic | 41,388 |
| Foumban | 5°72' | 10°90' | Equatorial mountain monsoon climate | Savannah forest mosaic | 130,292 |
| Bafoussam | 5°48' | 10°42' | Equatorial mountain monsoon climate | Savannah forest mosaic | 98,339 |
| Bafang | 5°16' | 10°18' | Equatorial mountain monsoon climate | Wooded savannah deciduous | 34,941 |
| Melong | 5°12' | 9°95' | Equatorial monsoon with higher pluviometry | Wet dense forest | 54,279 |
| Bafia | 4°75' | 11°22' | Subequatorial climate under shelter | Wet dense forest | 72,717 |
| Mbalmayo | 3°30' | 11°30' | Guinean subequatorial climate | Wet dense forest | 62,808 |
| Kumba | 4°63' | 9°44' | North Coast Equatorial | Wet dense forest | 166,331 |
| Bertoua | 4°57' | 13°67' | Guinean subequatorial climate | Savannah forest mosaic | 111,986 |
| Buea | 4°15' | 9°26' | North Coast Equatorial | Mountain forest | 131,325 |
| Douala | 4°05' | 9°76' | Equatorial monsoon with higher pluviometry | mangrove swamp and wooded | 1,926,513 |
| Limbe | 4°02' | 9°19' | Equatorial monsoon with higher pluviometry | submontane forest | 118,210 |
| Abong-Mbang | 3°97' | 13°17' | Guinean subequatorial climate | Wet dense forest | 29,005 |
| Edea | 3°79' | 10°13' | Equatorial monsoon with higher pluviometry | Wet dense forest | 8,8481 |
| Akonolinga | 3°78' | 12°24' | Guinean subequatorial climate | Wet dense forest | 47,561 |
| Kribi | 2°94' | 9°91' | Guinean subequatorial climate | Wet dense forest | 93,246 |
| Sangmelima | 2°93' | 11°59' | Guinean subequatorial climate | Wet dense forest | 91,740 |
| Ebolowa | 2°92' | 11°15' | Guinean subequatorial climate | Wet dense forest | 118,267 |
Fig 1. Geographic distribution of Ae. aegypti and Ae. albopictus in Cameroon, March-August 2017.

Mosquito collection and identification
Immature stages of mosquitoes were collected from mid-March to August 2017 corresponding to the rainy season. In most locations located under 6°N where both Ae. aegypti and Ae. albopictus were previously detected [36,37], collections in each town were performed in both the downtown and suburb to assess habitat segregation of both species according to building density and vegetation as demonstrated previously [27,35]. For collection located above 6°N, investigations were performed across the city and pooled together. In each selected location, all the containers with water (potential breeding sites) were inspected and container with at least one larva and/or pupa suspected to belong to Aedes genus (positive breeding sites) was recorded. The number of potential and positive containers were reported. The immature stages were collected, transported to the insectary and pooled together according to the environment (downtown vs. suburban) and location. They were maintained in the insectary until adult emergence. The emerged adults were identified under a binocular magnifying glass by the morphological criteria previously described [40–42], numbered, pooled in a breeding cage according to species and location, and further reared in the controlled conditions (27°C +/- 2°C; relative humidity 80% +/-10%) until generation 1 (G1) for further analysis. G0 adults were stored at -20°C for further molecular and genetic analyses. The prevalence of Ae. aegypti and Ae. albopictus was compared per environment and per location and statistically analysed using the Chi-square test. The Cameroon data Shape files were downloaded from the Global Administrative Areas (GADM) version 2.8 web site and the global positioning system coordinates were projected according to the WGS 84-EPSG 4326 system and the proportion of each Aedes species was generated with the QGIS version 3.4.1-Madeira software.
Mitochondrial DNA sequencing and analysis
The total DNA was extracted from 20 G0 individuals of Ae. albopictus collected in 17 sites (including samples from Yaoundé) using the Livak method as previously described [43].
DNA extracts from each locality were used as templates to amplify a 700-bp fragment of MtCOI gene. The sequences of the primers used are albCOIF 5’-TTTCAACAAATCATAAAGATATTGG-3’ and albCOIR 5’- TAAACTTCTGGA TGACCAAAAAATCA-3’ [44]. Polymerase chain reaction (PCR) amplification was performed using a Gene Touch thermal cycler (Bulldog Bio, Portsmouth, USA) as described previously [44]. Amplicons from the PCR were analysed by agarose gel electrophoresis stained with Midori green and visualized under UV light. Fifteen PCR products from each locality were purified using Exo-SAP protocol according to manufacturer recommendations and sequenced directly.
Sequences were visualized and corrected manually when necessary using BioEdit software version 7.0.5.3 and aligned using Clustal W [45]. Sequences were numbered based on the reference sequence downloaded from GenBank (Accession number KU738429.1) that originated from China. The number of haplotypes (h), the number of polymorphism sites (S), haplotype diversity (Hd), nucleotide diversity (π), were computed with DnaSP 5.10 [46]. The statistical tests of Tajima [47], Fu and Li [48] were estimated with DnaSP in order to establish non-neutral evolution and deviation from mutation-drift equilibrium. Different haplotypes detected were compared to previous COI region sequences published in GenBank from populations that originated from China, USA, Singapore, Thailand, Italy, Japan and Congo [49,50]. These COI sequences were used to construct the maximum likelihood phylogenetic tree using MEGA 7.0 [51]. Genealogical relationship between haplotype detected across Cameroon was assessed using TCS [52] and tcsBU [53] software.
Results
Larval habitats prospected per location
A total of 4,054 potential breeding sites was inspected in 28 locations across Cameroon, out of which 1,103 (27.20%) were found containing immature stages of Aedes (positive breeding sites). Detected breeding sites were grouped in six categories: used tires, discarded tanks; miscellaneous; water storage tanks; natural and recycled tires commonly used by the locals to protect wells (Table 2). Used tires were the most potential breeding sites discovered at 87.53% (3,545/4,050) and mostly infested (85.31%, 941/1,103) by Aedes larvae. The prevalence of other breeding sites was very low with 0.45% (5/1,103) in natural breeding sites, 3.0% (33/1,103) in tires covering water wells, 1.45% (16/1,103) in miscellaneous and 5.07% (56/1,103) in discarded tanks (Table 2).
Table 2. Containers prospected per site in Cameroon, mid-March to August 2017.
| Location | Used tires n (%) |
Tires on well n (%) | Water storage n (%) |
Discarded tanks n (%) |
Natural n (%) |
Miscellaneous n (%) |
Total n (%) |
|---|---|---|---|---|---|---|---|
| Maroua | 72 (43.1) | 0 (NC) | 2 (50.0) | 1 (100) | 0 (NC) | 2 (50) | 77 (44.2) |
| Garoua | 136 (27.2) | 0 (NC) | 2 (50) | 2 (100) | 0 (NC) | 3 (33.3) | 143 (29.4) |
| Mbe | 22 (22.7) | 0 (NC) | 5 (20) | 7 (100) | 2 (50) | 0 (NC) | 36 (38.9) |
| Meiganga | 54 (27.8) | 0 (NC) | 38 (5.3) | 0 (NC) | 0 (NC) | 9 (44.4) | 99 (21.2) |
| Banyo | 68 (14.7) | 0 (NC) | 0 (NC) | 4 (25) | 0 (NC) | 7 (100) | 79 (22.8) |
| Mayo-Darle | 119 (13.4) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 1 (100) | 120 (14.2) |
| Bankim | 150 (24.7) | 8 (87.5) | 0 (NC) | 4 (50) | 0 (NC) | 0 (NC) | 162 (28.4) |
| Ngaoundere downtown | 73 (42.8) | 0 (NC) | 0 (NC) | 19 (10.5) | 0 (NC) | 0 (NC) | 92 (35.9) |
| Ngaoundere suburban | 43 (27.9) | 0 (NC) | 0 (NC) | 3 (66.7) | 0 (NC) | 7 (100) | 53 (39.6) |
| Tibati downtown | 153 (28.8) | 0 (NC) | 0 (NC) | 4 (50) | 0 (NC) | 7 (42.9) | 164 (29.9) |
| Tibati suburban | 71 (35.2) | 12 (58.3) | 0 (NC) | 23 39.1) | 0 (NC) | 2 (0) | 108 (37.9) |
| Foumban downtown | 113 (20.4) | 0 (NC) | 0 (NC) | 9 (66.7) | 0 (NC) | 6 (66.7) | 128 (25.8) |
| Foumban suburban | 85 (23.5) | 0 (NC) | 0 (NC) | 1 (0) | 0 (NC) | 0 (NC) | 86 (23.3) |
| Bafoussam downtown | 358 (2.2) | 0 (NC) | 2 (0) | 2 (0) | 0 (NC) | 3 (0) | 365 (2.2) |
| Bafoussam suburban | 237 (10.6) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 3 (0) | 240 (10.4) |
| Bafang downtown | 33 (54.5) | 4 (100) | 0 (NC) | 3 (0) | 0 (NC) | 1 (0) | 41 (53.6) |
| Bafang suburban | 6 (66.7) | 0 (NC) | 8 (12.5) | 9 (11.1) | 0 (NC) | 20 (25) | 43 (25.6) |
| Bamenda downtown | 241 (9.1) | 0 (NC) | 0 (NC) | 1 (0) | 0 (NC) | 2 (0) | 244 (9.0) |
| Bamenda suburban | 98 (15.3) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 98 (15.3) |
| Bafia downtown | 82 (26.8) | 0 (NC) | 0 (NC) | 2 (0) | 0 (NC) | 1 (0) | 85 (25.9) |
| Bafia suburban | 68 (8.8) | 0 (NC) | 4 (0) | 30 (13.3) | 1 (100) | 27 (0) | 130 (8.4) |
| Mbalmayo downtown | 36 (22.2) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 1 (100) | 36 (25.0) |
| Mbalmayo suburban | 65 (58.5) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 65 (58.5) |
| Akonolinga suburban | 31 (22.6) | 0 (NC) | 0 (NC) | 3 (100) | 0 (NC) | 12 (66.7) | 46 (39.1) |
| Ebolowa downtown | 57 (52.6) | 0 (NC) | 1 (100) | 2 (50) | 0 (NC) | 2 (50) | 62 (53.2) |
| Ebolowa suburban | 37 (59.5) | 0 (NC) | 0 (NC) | 4 (25) | 2 (0) | 12 (41.7) | 55 (50.9) |
| Sangmelima downtown | 64 (46.9) | 0 (NC) | 1 (100) | 3 (100) | 0 (NC) | 4 (75) | 72 (51.4) |
| Sangmelima suburban | 15 (33.3) | 0 (NC) | 5 (60) | 6 (50) | 0 (NC) | 12 (50) | 36 (52.8) |
| Garoua-Boulai downtown | 117 (25.6) | 2 (50) | 1 (100) | 1 (0) | 0 (NC) | 2 (0) | 123 (26.01) |
| Garoua-Boulai suburban | 2 (100) | 17 (47.05) | 0 (NC) | 12 (8.3) | 0 (NC) | 0 (NC) | 31 (35.5) |
| Bertoua downtown | 62 (61.3) | 2 (50) | 0 (NC) | 2 (50) | 0 (NC) | 0 (NC) | 66 (60.6) |
| Bertoua suburban | 56 (42.9) | 6 (33.3) | 0 (NC) | 1 (0) | 0 (NC) | 0 (NC) | 63 (41.)3 |
| Abong-Mbang | 31 (58.1) | 1 (100) | 0 (NC) | 3 (0) | 3 (100) | 1 (100) | 39 (59.0) |
| Kumba downtown | 35 (25.7) | 0 (NC) | 0 (NC) | 7 (57.1) | 0 (NC) | 3 (33.3) | 45 (33.3) |
| Kumba suburban | 20 (55.0) | 1 (100) | 3 (33.3) | 4 (25.0) | 1 (0) | 7 (42.9) | 36 (47.2) |
| Limbe downtown | 40 (65.0) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 40 (65.0) |
| Limbe suburban | 95 (47.4) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 2 (100) | 97 (48.5) |
| Buea downtown | 88 (23.9) | 0 (NC) | 0 (NC) | 2 (0) | 0 (NC) | 3 (0) | 93 (22.6) |
| Buea suburban | 100 (26.0) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 100 (26.0) |
| Melong downtown | 70 (34.3) | 2 (50) | 2 (0) | 5 (20) | 0 (NC) | 11 (63.6) | 90 (36.7) |
| Melong suburban | 52 (32.7) | 0 (NC) | 0 (NC) | 2 (0) | 0 (NC) | 21 (14.3) | 75 (26.7) |
| Douala downtown | 36 (91.7) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 36 (91.7) |
| Douala suburban | 100 (26) | 0 (NC) | 0 (NC) | 1 (100) | 0 (NC) | 0 (NC) | 101 (26.73) |
| Edea downtown | 24 (87.5) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 24 (87.5) |
| Edea suburban | 30 (36.66) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 0 (NC) | 30 (30.66) |
| Total | 3,545 (26.5) | 40 (82.5) | 63 (19.04) | 176 (31.8) | 9 (55.6) | 188 (39.4) | 4,054 (27.2) |
N, number of containers found with water; (%), the percentage of positive containers; NC, not computed.
Distribution and prevalence of Ae. aegypti and Ae. albopictus across the country
30,381 immature stages of Ae. albopictus and Ae. aegypti species were collected in 29 localities between mid-March and August 2017 in Cameroon (Table 3). Several other species were found in association with Ae. aegypti and Ae. albopictus. These include Aedes simpsoni Theobald 1905, Aedes vittatus Bigot 1861, Anopheles gambiae s.l. Giles 1902, Culex tigripes De Grandpré and De Charmoy 1900, Culex pipiens quinquefasciatus Say, 1823, Culex perfuscus Edwards 1914, Culex duttoni Theobald 1901, Culex antennatus Beker 1903, Culex sp Linnaeus 1758, Eretmapodites brevipalpis Ingram and De Meillon 1927, and Toxorhynchites brevipalpis Ribeiro 1991.
Table 3. Relative abundance of Ae. albopictus and Ae. aegypti across the prospected location in Cameroon.
| Location |
Ae. albopictus n (%) |
Ae. aegypti n (%) |
|---|---|---|
| Kumba downtown | 717 (96.24) | 28 (3.76) |
| Kumba suburban | 157 (80.51) | 38 (19.49) |
| Buea downtown | 743 (88.45) | 97 (11.55) |
| Buea suburban | 697 (88.23) | 93 (11.77) |
| Limbe downtown | 313 (41.62) | 439 (58.38) |
| Limbe suburban | 863 (63.74) | 491 (36.26) |
| Melong downtown | 347 (60.66) | 225 (39.34) |
| Melong suburban | 156 (60.47) | 102 (39.53) |
| Douala downtown | 319 (31.71) | 687 (68.29) |
| Douala suburban | 214 (35.73) | 385 (64.27) |
| Edea downtown | 164 (47.81) | 179 (52.19) |
| Edea suburban | 168 (97.11) | 5 (2.89) |
| Bamenda downtown | 245 (83.62) | 48 (16.38) |
| Bamenda suburban | 247 (62.85) | 146 (37.15) |
| Bafoussam downtown | 272 (99.27) | 2 (0.73) |
| Bafoussam suburban | 294 (53.55) | 255 (46.45) |
| Bafang downtown | 1032 (91.17) | 100 (8.83) |
| Bafang suburban | 182 (81.98) | 40 (18.02) |
| Foumban downtown | 442 (67.69) | 211 (32.31) |
| Foumban suburban | 335 (64.67) | 183 (35.33) |
| Bafia downtown | 844 (93.99) | 54 (6.01) |
| Bafia suburban | 306 (48.73) | 322 (51.27) |
| Mbalmayo downtown | 504 (83.17) | 102 (16.83) |
| Mbalmayo suburban | 1055 (93.86) | 69 (6.14) |
| Akonolinga* | 1449 (94.89) | 78 (5.11) |
| Ebolowa downtown | 305 (97.44) | 8 (2.56) |
| Ebolowa suburban | 566 (98.43) | 9 (1.57) |
| Sangmelima downtown | 679 (96.86) | 22 (3.14) |
| Sangmelima suburban | 437 (92.78) | 34 (7.22) |
| Kribi downtown | 495 (97.44) | 13 (2.6) |
| Kribi suburban | 1597 (77.94) | 452 (22.06) |
| Abong-Mbang* | 656 (77.18) | 194 (22.82) |
| Bertoua downtown | 769 (99.1) | 7 (0.9) |
| Bertoua suburban | 867 (97.97) | 18 (2.03) |
| Garoua-Boulaï downtown | 59 (28.92) | 145 (71.08) |
| Garoua-Boulai suburban | 430 (92.47) | 35 (7.53) |
| Tibati downtown | 733 (58.5) | 520 (41.5) |
| Tibati suburban | 359 (60.44) | 235 (39.56) |
| Bankim* | 1031 (69.57) | 451 (30.43) |
| Ngaoundere downtown | 0 | 556 (100) |
| Ngaoundere suburban | 0 | 147 (100) |
| Mbe* | 0 | 249 (100) |
| Garoua* | 0 | 812 (100) |
| Maroua* | 0 | 146 (100) |
| Meiganga* | 0 | 484 (100) |
| Banyo* | 0 | 292 (100) |
| Mayo-Darle* | 0 | 125 (100) |
| Total | 21,048 (69.28) | 9,333 (30.72) |
n, number of adult mosquitoes identified; %, percentage
*, mosquitoes collected across the city and pool together.
Aedes aegypti was found across the country in all the sites investigated whereas Ae. albopictus distribution was limited to the southern part of the country under 6°4’N (Fig 1 and Table 3). Overall, Ae albopictus was more prevalent (69.28%) than Ae. aegypti (30.72%)(Table 3). In all the locations in which both species were found together Ae. albopictus was found to be more abundant except in Douala where Ae. aegypti was predominant in downtown and suburban. When analyses were done according to the environment (suburban vs downtown) in each location, Ae. albopictus was found to be the dominant species in the suburban and downtown in all the sympatric areas except in Garoua-Boulai, Douala, Limbe, and Edea where Ae. aegypti was predominant in downtown (Table 3). Analysis also revealed that in some locations such as Bertoua, Kribi, Sangmelima, Ebolowa and Bafoussam, Ae. albopictus is highly prevalent in all areas and nearly excluding the native species which sometimes represents less than 3%.
Genetic diversity of Ae. albopictus with Mitochondrial DNA
In total, 226 individuals of Ae. albopictus from 17 localities throughout Cameroon were analysed with the mtCOI gene (Table 4). Sequences analysed based on 636 nucleotides revealed a low polymorphism, with four substitution sites defining five haplotypes resulting in low haplotype diversity (hd) ranging from 0.13 to 0.65 with overall hd of 0.32. Similarly, low nucleotide diversity (π) index was recorded varying from 0.0000 to 0.0017 with 0.00075 for total sample. The predominant haplotype H1 (79.7% in total sample) was recorded in all the locations with frequency ranging from 35.7% to 100% (Fig 2, S1 Table). This haplotype was also the most prevalent in almost all locations excepted in Bafia and Kribi where it is rather H2 (57.1%) and H3 (54.5%) haplotypes, respectively (S1 Table). The predominant H1 haplotype matches perfectly with the reference sequence downloaded from GenBank originating from China (KU738429.1) and corresponds to the predominant haplotype detected recently in a neighbouring country, the Republic of the Congo [50]. All the three haplotypes isolated previously in the Republic of the Congo were also detected with the same primer sets in Cameroon. Analysis of haplotype network revealed that each haplotype is isolated from the others by one mutational step. Overall, all the Tajima statistics estimated were negatives (D = -0.43, D* = -0.43, Fs = -0.93, and F* = -040), but not statistically significant (Table 4). Phylogenetic analysis of the 636bp fragment showed that Cameroonian and Congolese Ae. albopictus populations have the same origin as they cluster together on the Maximum likelihood tree (Fig 3). Nucleotide sequences of five haplotypes detected across Cameroon have been deposited in the GenBank database (accession numbers: MH921568, MH921569, MH921570, MH921571 and MH921572).
Table 4. Genetic diversity indices in Aedes albopictus from Cameroon.
| Location | N | Hp | S | HpD | π (k) | D | D* | Fs | F* |
|---|---|---|---|---|---|---|---|---|---|
| Abong-Mbang | 9 | H1, H2 | 2 | 0.22 | 0.0007 (0.444) | -1.36 | -1.51 | 0.67 | -1.50 |
| Bafang | 13 | H1, H2 | 2 | 0.15 | 0.0005 (0.308) | -1.47 | -1.78 | 0.36 | -1.75 |
| Bafoussam | 14 | H1 | 0 | NC | 0.0000 (0.000) | NC | 0.00 | 0.00 | NC |
| Bamenda | 13 | H1 | 0 | NC | 0.0000 (0.000) | NC | 0.00 | 0.00 | NC |
| Bankim | 14 | H1 | 0 | NC | 0.0000 (0.000) | NC | 0.00 | 0.00 | NC |
| Bertoua | 15 | H1 | 0 | NC | 0.0000 (0.000) | NC | 0.00 | 0.00 | NC |
| Bafia | 14 | H1, H2, H3 | 2 | 0.58 | 0.0016 (1.022) | 1.70 | 0.94 | 0.91 | 1.17 |
| Buea | 14 | H1, H3 | 1 | 0.36 | 0.0006 (0.363) | 0.32 | 0.72 | 0.64 | 0.64 |
| Douala | 10 | H1, H3 | 1 | 0.36 | 0.0006 (0.355) | 0.015 | 0.80 | 0.42 | 0.63 |
| Ebolowa | 14 | H1, H3, H4 | 2 | 0.47 | 0.0009 (0.582) | -0.20 | -0.45 | -0.21 | -0.40 |
| Garoua-Boulai | 15 | H1, H3 | 1 | 0.13 | 0.0002 (0.133) | -1.16 | -1.43 | -0.65 | -1.41 |
| Kribi | 11 | H1, H3 | 1 | 0.55 | 0.0009 (0.545) | 1.44 | 0.78 | 1.14 | 0.96 |
| Kumba | 13 | H1, H2, H3 | 2 | 0.62 | 0.0017 (1.077) | 1.88 | 0.95 | 0.93 | 1.23 |
| Limbe | 14 | H1 | 2 | 0.26 | 0.0008 (0.527) | -0.44 | 0.94 | 1.25 | 0.60 |
| Melong | 15 | H1, H3, H5 | 2 | 0.36 | 0.0006 (0.381) | -1.00 | -0.48 | -0.92 | -0.64 |
| Tibati | 14 | H1 | 0 | NC | 0.0000 (0.000) | NC | 0.00 | 0.00 | NC |
| Yaounde | 14 | H1, H2, H3 | 2 | 0.65 | 0.0013 (0.802) | 0.75 | 0.93 | 0.40 | 0.92 |
| Total | 226 | 5 | 4 | 0.35 | 0.0008 (0.503) (0.503) | -0.43 | -0.43 | -0.93 | -0.51 |
N: Number of sequences analysed; S: Number of segregating sites; π: Nucleotide diversity per site; k: Average number of nucleotide differences; Hp: Number of haplotypes; HpD: Haplotype diversity; Fs: Fu’s Statistic; D: Tajima’s statistic; D* and F*: Fu and Li’s statistics NC: Not Computed.
Fig 2. Genetic diversity of the COI gene across Cameroonian populations of Ae. albopictus.
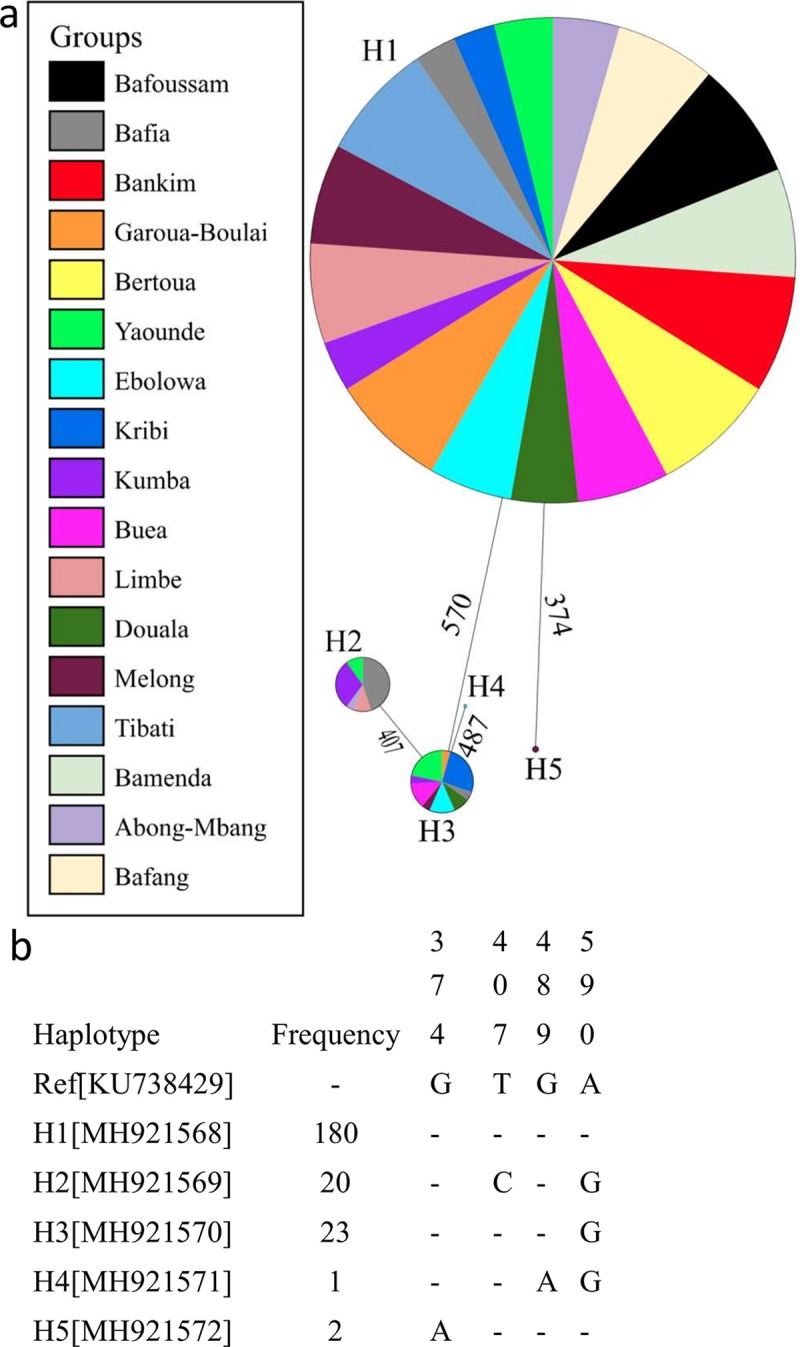
a, Haplotype network showing the genealogic relationship between five haplotypes detected across Cameroon. b, COI haplotypes found across Cameroon. Only polymorphic positions are shown and are numbered with reference (Ref) to the published Ae. albopictus sequences for COI (JF309317; China). Dots represent identity with respect to the reference.
Fig 3. Molecular phylogenetic analysis of Ae. albopictus by maximum likelihood method.

The evolutionary history was inferred by using the Maximum Likelihood method based on the Tamura 3-parameter model. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. There was a total of 636 positions in the final dataset.
Discussion
This study presents an extensive profiling of the prevalence and geographical distribution of Ae. albopictus and Ae. aegypti in Cameroon updating data generated more than 10 years ago. This current report reveals that Ae. albopictus distribution continues to be restricted to the southern part of the country, around 6°N latitude while Ae. aegypti is found throughout the country. The predominance of the invading species (Ae. albopictus) over the native species (Ae. aegypti) is also reported in almost all locations where both species are sympatric.
The current distribution of Ae. aegypti and Ae. albopictus is similar to the previous distribution reported in Cameroon in 2003 [36] and 2007 [37] and in the Central African Republic in 2012 [35] where the distribution of Ae. albopictus was shown to be restricted to the South around 6°N. This boundary has been suggested to be due to the unfavourable climatic conditions for the establishment of this species in the northern part of central Africa above 6°5’N rather than the dynamics of invasion process that is still ongoing as suggested previously [36]. Indeed, mean annual temperatures in Cameroon vary between 20 to 28° C and increase from the south towards the north. Although, both Ae. aegypti and Ae. albopictus have desiccant-resistant eggs, previous studies showed that Ae. aegypti eggs are more tolerant to high temperatures than those of Ae. albopictus [54].
Consequently, the climatic conditions in the southern part of Cameroon (i.e., equatorial climate, average annual temperature <26.5°C) are favourable to the development of Ae. albopictus. A higher prevalence of Ae. albopictus was observed in some sympatric areas in both suburban and downtown environment, which is in contrast to the previous results in central Africa. Ae. aegypti has often been the prevalent species in downtown with high building density whereas Ae. albopictus was predominant in suburban area surrounded by vegetation [27,35]. In some southern locations such as Limbe and Edea, Ae. albopictus was found to be more prevalent in suburb whereas it is Ae. aegypti that was found more in downtown. Ae. aegypti was also the prevalent species in both suburb and downtown sites in Douala located in the coastal region in the southern part of Cameroon. This observation is consistent with a previous report in Douala in 2006 suggesting that the prevailing climate in Douala is not favourable for the propagation of the invading species [37]. All these observations suggest that the differences in the proportions of both species found in different locations in southern Cameroon may probably reflect differences in environmental factors such as climate, vegetation and building density as suggested previously [37].
Due to the fact that both species exploit the same ecological niches and resources (larval habitat, blood source), Ae. aegypti and Ae. albopictus have competitive interactions [30,32,55] that may result in the replacement of one species by another in a given environment. Indeed, several studies carried out around the world have shown the changes of the range and abundance of the native species after the introduction of Ae. albopictus [31,33,34]. In Cameroon, where both species exploit the same types of resources, it is possible that such competitive phenomena are in progress. The mechanisms for the competition are not well known, but many authors believe that it could occur at the pre-imaginal phase and that several factors such as temperature, precipitation, response to symbionts, parasites, predators, and chemical interferences delaying growth could be the main driving forces [30,32]. In addition, other studies demonstrated that mating interference in favour to Ae. albopictus, called satyrization, is one of the probable cause of the competitive displacement of resident Ae. aegypti by the invasive Ae. albopictus where they co-exist [56,57]. On the other hand, the coexistence of Ae. aegypti and Ae. albopictus was reported in certain locations in Florida (USA) two decades after competitive displacement [58].
Used tires were the containers mostly found and mostly positive as breeding sites across the country. This is consistent with previous studies in central Africa demonstrating that used tires are the main productive for both Ae. aegypti and Ae. albopictus [11,35–37]. The propensity of Ae. aegypti and Ae. albopictus to colonize used tires may be due to the fact that these species are native to the forest and breed mainly in natural tree holes, which share the characteristics of tires, as the dark colour and the dark interior provide attractive resting or oviposition site for Aedes spp. as previously suggested [35]. Nevertheless, in this study, sampling was targeted mainly at garages and used tire shops to increase the chances of finding the immature stages of Aedes spp. It is important to highlight the remarkable presence of tires as they are used to protect water wells in certain locations such as Garoua Boulai, Bertoua, Bankim and Tibati. This important observation could help raise the awareness in the populations of inadvertently providing larval habitats for arbovirus vectors due to this habit potentially helping to fight against the arboviral diseases.
The presence and higher prevalence of Ae. albopictus in southern part of Cameroon can have a significant impact on the epidemiology of mosquito-borne arboviral diseases since Ae. albopictus has been found competent to transmit about 22 arboviruses [59]. Interestingly, the emergence of dengue and chikungunya viruses in domesticated environments in central Africa coincides with the introduction of Ae. albopictus in this area [11,12,24]. In addition, Ae. albopictus was found infected by zika virus in natural conditions in Gabon in Central Africa [60]. It was also demonstrated that Ae. albopictus from Bangui in the Central African Republic is able to transmit enzootic chikungunya virus strain [61]. This suggests Ae. albopictus can serve as bridge to transfer viruses from sylvan area to urban in central Africa whether this species become dominant in wild settings. Further studies assessing the spread of the invading species Ae. albopictus in sylvan and rural environments are also needed.
The discrepancy observed in the distribution of Ae. albopictus and Ae. aegypti across the country notably the restriction of Ae. albopictus in the southern part suggest that the implementing of vector control programme should take into account the specificity of each area. However, data collected across the country show that a good system of waste management in the domesticated environment including the destroying of the used tires could contribute to reduce the density of both Ae. aegypti and Ae. albopictus in Cameroon and indirectly reduce the risk of transmission of diseases transmitted by these mosquitoes.
MtDNA analysis using the COI gene in Ae. albopictus populations from Cameroon revealed a low polymorphism with only five haplotypes detected across the country. Among these haplotypes, three (H1, H2 and H3) of them have been detected previously in the Republic of the Congo with the same primers [50]. This low polymorphism reported in Cameroon and Republic of the Congo is in accordance with the previous studies using another portion of the COI gene in areas newly colonised by Ae. albopictus including Central African countries [9,35,62]. It was previously suggested that this low polymorphism is mainly due to the recent introduction of Ae. albopictus from a founder population [35]. Indeed, Ae. albopictus was reported for the first time in Cameroon in 1999. Phylogenetic analysis showed that the haplotype sequences from Cameroon are very close to other sequences isolated to the populations originating from China and Congo, suggesting Cameroonian’s and Congolese populations could have the same origin. Meanwhile, the fact that more haplotypes are found in Cameroon suggests that this country could have been the entrance point of Ae. albopictus in Central Africa potentially through the Port of Douala which is the major one in Central Africa. Primers used in this current study were not the same as those used in the previous study in Cameroon, Central African Republic, and Sao Tome Island. Thus, it was not possible to compare the haplotypes detected in this study with the previous ones detected in Central Africa. Nevertheless, the current results support previous findings suggesting that it is likely that the invading population which colonized Central Africa originated mainly from other tropical regions of the world [9,35,62]. Further studies, including samples from all central African region using other markers such as double digest RAD sequencing, are required to assess the genetic structure and the level of the gene flow between central African Ae. albopictus populations.
This study shows that for the past 10 years the distribution of Ae. albopictus is still restricted to southern Cameroon below 6°5’N latitude while Ae. aegypti is present across the country. This suggests that the prevailing climate in the northern part of Cameroon is not conducive to the invading species Ae. albopictus in this part of the country. However, the invading species is more prevalent in almost all locations in sympatric with the native species suggesting replacement of native species Ae. aegypti is ongoing in some locations.
Supporting information
(DOCX)
Acknowledgments
We would like to thank the populations from different collection sites for their collaboration during the field works.
Data Availability
Nucleotide sequences of five haplotypes detected across Cameroon have been deposited in the GenBank database (accession numbers: MH921568, MH921569, MH921570, MH921571 and MH921572)
Funding Statement
This study was supported by a Wellcome Trust Training Fellowship in Public Health and Tropical Medicine (204862) awarded to BK. The funders had no role in study design, data collection or analysis, decision to publish or preparation of the manuscript
References
- 1.Leta S, Beyene TJ, De Clercq EM, Amenu K, Kraemer MUG, et al. (2018) Global risk mapping for major diseases transmitted by Aedes aegypti and Aedes albopictus. Int J Infect Dis 67: 25–35. 10.1016/j.ijid.2017.11.026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Paixao ES, Teixeira MG, Rodrigues LC (2018) Zika, chikungunya and dengue: the causes and threats of new and re-emerging arboviral diseases. BMJ Glob Health 3: e000530 10.1136/bmjgh-2017-000530 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Sharma A, Lal SK (2017) Zika virus: transmission, detection, control, and prevention. Front Microbiol 8: 110 10.3389/fmicb.2017.00110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Mattingly PF (1957) Genetical aspects of the Aedes aegypti problem. Annals of Tropical Medicine & Parasitology 51: 392–408. [PubMed] [Google Scholar]
- 5.Powell JR, Tabachnick WJ (2013) History of domestication and spread of Aedes aegypti—a review. Mem Inst Oswaldo Cruz 108 Suppl 1: 11–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Paupy C, Delatte H, Bagny L, Corbel V, Fontenille D (2009) Aedes albopictus, an arbovirus vector: from the darkness to the light. Microbes Infect 11: 1177–1185. 10.1016/j.micinf.2009.05.005 [DOI] [PubMed] [Google Scholar]
- 7.Benedict MQ, Levine RS, Hawley WA, Lounibos LP (2007) Spread of the tiger: global risk of invasion by the mosquito Aedes albopictus. Vector Borne Zoonotic Dis 7: 76–85. 10.1089/vbz.2006.0562 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Reiter P (1998) Aedes albopictus and the world trade in used tires: 1988–1995 the shape of things to come? Journal of the America Mosquito Control Association 14: 83–94. [PubMed] [Google Scholar]
- 9.Kamgang B, Brengues C, Fontenille D, Njiokou F, Simard F, et al. (2011) Genetic structure of the tiger mosquito, Aedes albopictus, in Cameroon (Central Africa). PLoS One 6: e20257 10.1371/journal.pone.0020257 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fontenille D, Toto JC (2001) Aedes (Stegomyia) albopictus (Skuse), a potential new Dengue vector in southern Cameroon. Emerg Infect Dis 7: 1066–1067. 10.3201/eid0706.010631 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ngoagouni C, Kamgang B, Nakoune E, Paupy C, Kazanji M (2015) Invasion of Aedes albopictus (Diptera: Culicidae) into central Africa: what consequences for emerging diseases? Parasit Vectors 8: 191 10.1186/s13071-015-0808-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mombouli JV, Bitsindou P, Elion DO, Grolla A, Feldmann H, et al. (2013) Chikungunya virus infection, Brazzaville, Republic of Congo, 2011. Emerg Infect Dis 19: 1542–1543. 10.3201/eid1909.130451 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Leroy EM, Nkoghe D, Ollomo B, Nze-Nkogue C, Becquart P, et al. (2009) Concurrent chikungunya and dengue virus infections during simultaneous outbreaks, Gabon, 2007. Emerg Infect Dis 15: 591–593. 10.3201/eid1504.080664 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Peyrefitte CN, Rousset D, Pastorino BA, Pouillot R, Bessaud M, et al. (2007) Chikungunya virus, Cameroon, 2006. Emerg Infect Dis 13: 768–771. 10.3201/eid1305.061500 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Maurice D, Alain SM, Christophe V, Rene N, Irene KT, et al. (2015) Molecular characterization of chikungunya virus from three regions of Cameroon. Virol Sin 30: 470–473. 10.1007/s12250-015-3663-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Demanou M, Antonio-Nkondjio C, Ngapana E, Rousset D, Paupy C, et al. (2010) Chikungunya outbreak in a rural area of Western Cameroon in 2006: A retrospective serological and entomological survey. BMC Res Notes 3: 128 10.1186/1756-0500-3-128 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ndip LM, Bouyer DH, Travassos Da Rosa AP, Titanji VP, Tesh RB, et al. (2004) Acute spotted fever rickettsiosis among febrile patients, Cameroon. Emerg Infect Dis 10: 432–437. 10.3201/eid1003.020713 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kuniholm MH, Wolfe ND, Huang CY, Mpoudi-Ngole E, Tamoufe U, et al. (2006) Seroprevalence and distribution of Flaviviridae, Togaviridae, and Bunyaviridae arboviral infections in rural Cameroonian adults. Am J Trop Med Hyg 74: 1078–1083. [PubMed] [Google Scholar]
- 19.Yousseu FBS, Nemg FBS, Ngouanet SA, Mekanda FMO, Demanou M (2018) Detection and serotyping of dengue viruses in febrile patients consulting at the New-Bell District Hospital in Douala, Cameroon. PLoS One 13: e0204143 10.1371/journal.pone.0204143 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Monamele GC, Demanou M (2018) First documented evidence of dengue and malaria co-infection in children attending two health centers in Yaounde, Cameroon. Pan Afr Med J 29: 227 10.11604/pamj.2018.29.227.15316 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Demanou M, Pouillot R, Grandadam M, Boisier P, Kamgang B, et al. (2014) Evidence of dengue virus transmission and factors associated with the presence of anti-dengue virus antibodies in humans in three major towns in Cameroon. PLoS Negl Trop Dis 8: e2950 10.1371/journal.pntd.0002950 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Gake B, Vernet MA, Leparc-Goffart I, Drexler JF, Gould EA, et al. (2017) Low seroprevalence of Zika virus in Cameroonian blood donors. Braz J Infect Dis 21: 481–483. 10.1016/j.bjid.2017.03.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Pages F, Peyrefitte CN, Mve MT, Jarjaval F, Brisse S, et al. (2009) Aedes albopictus mosquito: the main vector of the 2007 Chikungunya outbreak in Gabon. PLoS One 4: e4691 10.1371/journal.pone.0004691 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Paupy C, Ollomo B, Kamgang B, Moutailler S, Rousset D, et al. (2010) Comparative role of Aedes albopictus and Aedes aegypti in the emergence of Dengue and Chikungunya in central Africa. Vector Borne Zoonotic Dis 10: 259–266. 10.1089/vbz.2009.0005 [DOI] [PubMed] [Google Scholar]
- 25.Moyen N, Thiberville SD, Pastorino B, Nougairede A, Thirion L, et al. (2014) First reported chikungunya fever outbreak in the republic of Congo, 2011. PLoS One 9: e115938 10.1371/journal.pone.0115938 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Weetman D, Kamgang B, Badolo A, Moyes CL, Shearer FM, et al. (2018) Aedes Mosquitoes and Aedes-Borne Arboviruses in Africa: Current and Future Threats. Int J Environ Res Public Health 15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kamgang B, Yougang AP, Tchoupo M, Riveron JM, Wondji C (2017) Temporal distribution and insecticide resistance profile of two major arbovirus vectors Aedes aegypti and Aedes albopictus in Yaounde, the capital city of Cameroon. Parasit Vectors 10: 469 10.1186/s13071-017-2408-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Gilotra SK, Rozeboom LE, Bhattacharya NC (1967) Observations on possible competitive displacement between populations of Aedes aegypti Linnaeus and Aedes albopictus Skuse in Calcutta. Bull World Health Organ 37: 437–446. [PMC free article] [PubMed] [Google Scholar]
- 29.Rudnick A, Chan YC (1965) Dengue Type 2 Virus in Naturally Infected Aedes albopictus Mosquitoes in Singapore. Science 149: 638–639. 10.1126/science.149.3684.638 [DOI] [PubMed] [Google Scholar]
- 30.Lounibos LP (2002) Invasions by insect vectors of human disease. Annual review of entomology 47: 233–266. 10.1146/annurev.ento.47.091201.145206 [DOI] [PubMed] [Google Scholar]
- 31.O'Meara GF, Evans LF Jr., Gettman AD, Cuda JP (1995) Spread of Aedes albopictus and decline of Ae. aegypti (Diptera: Culicidae) in Florida. J Med Entomol 32: 554–562. [DOI] [PubMed] [Google Scholar]
- 32.Juliano SA, Lounibos LP, O'Meara GF (2004) A field test for competitive effects of Aedes albopictus on A. aegypti in South Florida: differences between sites of coexistence and exclusion? Oecologia 139: 583–593. 10.1007/s00442-004-1532-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Bagny L, Delatte H, Quilici S, Fontenille D (2009) Progressive decrease in Aedes aegypti distribution in Reunion Island since the 1900s. J Med Entomol 46: 1541–1545. [DOI] [PubMed] [Google Scholar]
- 34.Bagny L, Delatte H, Elissa N, Quilici S, Fontenille D (2009) Aedes (Diptera: Culicidae) vectors of arboviruses in Mayotte (Indian Ocean): distribution area and larval habitats. J Med Entomol 46: 198–207. [DOI] [PubMed] [Google Scholar]
- 35.Kamgang B, Ngoagouni C, Manirakiza A, Nakoune E, Paupy C, et al. (2013) Temporal patterns of abundance of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) and mitochondrial DNA analysis of Ae. albopictus in the Central African Republic. PLoS Negl Trop Dis 7: e2590 10.1371/journal.pntd.0002590 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Simard F, Nchoutpouen E, Toto JC, Fontenille D (2005) Geographic Distribution and Breeding Site Preference of Aedes albopictus and Aedes aegypti (Diptera: Culicidae) in Cameroon, Central Africa. Journal of Medical Entomology 42: 726–731. [DOI] [PubMed] [Google Scholar]
- 37.Kamgang B, Happi JY, Boisier P, Njiokou F, Herve JP, et al. (2010) Geographic and ecological distribution of the dengue and chikungunya virus vectors Aedes aegypti and Aedes albopictus in three major Cameroonian towns. Med Vet Entomol 24: 132–141. 10.1111/j.1365-2915.2010.00869.x [DOI] [PubMed] [Google Scholar]
- 38.Olivry JC (1986) Fleuves et rivières du Cameroun. 781 p. [Google Scholar]
- 39.Simberloff D, Martin JL, Genovesi P, Maris V, Wardle DA, et al. (2013) Impacts of biological invasions: what's what and the way forward. Trends Ecol Evol 28: 58–66. 10.1016/j.tree.2012.07.013 [DOI] [PubMed] [Google Scholar]
- 40.Jupp PG (1996) Mosquitoes of southern Africa. Culicinae and Toxorhynchitinae: Ekogilde, Hartebeespoort, South Africa. [Google Scholar]
- 41.Edwards FW (1941) Mosquitoes of the Ethiopian region III Culicine adults and pupae. British Museum (Natural History), London, United Kingdom: 499. [Google Scholar]
- 42.Gillies MT, De Meillon B (1968) The Anophelinae of Africa south of the Sahara (Ethiopian Zoogeographical Region). Johannesburg: South African Institute for Medical Research Institute; 54. [Google Scholar]
- 43.Livak KJ (1984) Organization and mapping of a sequence on the Drosophila melanogaster X and Y chromosomes that is transcribed during spermatogenesis. Genetics 107: 611–634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Maynard AJ, Ambrose L, Cooper RD, Chow WK, Davis JB, et al. (2017) Tiger on the prowl: Invasion history and spatio-temporal genetic structure of the Asian tiger mosquito Aedes albopictus (Skuse 1894) in the Indo-Pacific. PLoS Negl Trop Dis 11: e0005546 10.1371/journal.pntd.0005546 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 22: 4673–4680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Librado P, Rozas J (2009) DnaSP v5: a software for comprehensive analysis of DNA polymorphism data. Bioinformatics 25: 1451–1452. 10.1093/bioinformatics/btp187 [DOI] [PubMed] [Google Scholar]
- 47.Tajima F (1989) Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 123: 585–595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Fu YX, Li WH (1993) Statistical tests of neutrality of mutations. Genetics 133: 693–709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Zhong D, Lo E, Hu R, Metzger ME, Cummings R, et al. (2013) Genetic analysis of invasive Aedes albopictus populations in Los Angeles County, California and its potential public health impact. PLoS One 8: e68586 10.1371/journal.pone.0068586 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kamgang B, Wilson-Bahun TA, Irving H, Kusimo MO, Lenga A, et al. (2018) Geographical distribution of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) and genetic diversity of invading population of Ae. albopictus in the Republic of the Congo. Wellcome Open Res 3: 79 10.12688/wellcomeopenres.14659.3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol 30: 2725–2729. 10.1093/molbev/mst197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Clement M, Posada D, Crandall KA (2000) TCS: a computer program to estimate gene genealogies. Mol Ecol 9: 1657–1659. [DOI] [PubMed] [Google Scholar]
- 53.Murias dos Santos A, Cabezas MP, Tavares AI, Xavier R, Branco M (2016) tcsBU: a tool to extend TCS network layout and visualization. Bioinformatics 32: 627–628. 10.1093/bioinformatics/btv636 [DOI] [PubMed] [Google Scholar]
- 54.Juliano SA, O'Meara GF, Morrill JR, Cutwa MM (2002) Desiccation and thermal tolerance of eggs and the coexistence of competing mosquitoes. Oecologia 130: 458–469. 10.1007/s004420100811 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Chan YC, Chan KL, Ho BC (1971) Aedes aegypti (L.) and Aedes albopictus (Skuse) in Singapore City. 1. Distribution and density. Bull World Health Organ 44: 617–627. [PMC free article] [PubMed] [Google Scholar]
- 56.Bargielowski IE, Lounibos LP, Carrasquilla MC (2013) Evolution of resistance to satyrization through reproductive character displacement in populations of invasive dengue vectors. Proc Natl Acad Sci U S A 110: 2888–2892. 10.1073/pnas.1219599110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Bargielowski IE, Lounibos LP (2016) Satyrization and satyrization-resistance in competitive displacements of invasive mosquito species. Insect Sci 23: 162–174. 10.1111/1744-7917.12291 [DOI] [PubMed] [Google Scholar]
- 58.Lounibos LP, Bargielowski I, Carrasquilla MC, Nishimura N (2016) Coexistence of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in Peninsular Florida Two Decades After Competitive Displacements. J Med Entomol 53: 1385–1390. 10.1093/jme/tjw122 [DOI] [PubMed] [Google Scholar]
- 59.Gratz NG (2004) Critical review of the vector status of Aedes albopictus. Med Vet Entomol 18: 215–227. 10.1111/j.0269-283X.2004.00513.x [DOI] [PubMed] [Google Scholar]
- 60.Grard G, Caron M, Mombo IM, Nkoghe D, Mboui Ondo S, et al. (2014) Zika virus in Gabon (Central Africa)—2007: a new threat from Aedes albopictus? PLoS Negl Trop Dis 8: e2681 10.1371/journal.pntd.0002681 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Ngoagouni C, Kamgang B, Kazanji M, Paupy C, Nakoune E (2017) Potential of Aedes aegypti and Aedes albopictus populations in the Central African Republic to transmit enzootic chikungunya virus strains. Parasit Vectors 10: 164 10.1186/s13071-017-2101-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Reis S, Cornel AJ, Melo M, Pereira H, Loiseau C (2017) First record of Aedes albopictus (Skuse 1894) on Sao tome island. Acta Trop 171: 86–89. 10.1016/j.actatropica.2017.03.035 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
Data Availability Statement
Nucleotide sequences of five haplotypes detected across Cameroon have been deposited in the GenBank database (accession numbers: MH921568, MH921569, MH921570, MH921571 and MH921572)