

Abstract
Transmitter release at auditory inner hair cell (IHC) ribbon synapses involves exocytosis of glutamatergic vesicles during voltage activation of L-type Cav1.3 calcium channels. At these synapses, the fast and indefatigable release of synaptic vesicles by IHCs is controlled by otoferlin, a six-C2-domain (C2-ABCDEF) protein that functions as a high-affinity Ca2+ sensor. The molecular events by which each otoferlin C2 domain contributes to the regulation of the synaptic vesicle cycle in IHCs are still incompletely understood. Here, we investigate their role using a cochlear viral cDNA transfer approach in vivo, where IHCs of mouse lacking otoferlin (Otof−/− mice of both sexes) were virally transduced with cDNAs of various mini-otoferlins. Using patch-clamp recordings and membrane capacitance measurements, we show that the viral transfer of mini-otoferlin containing C2-ACEF, C2-EF, or C2-DEF partially restores the fast exocytotic component in Otof−/− mouse IHCs. The restoration was much less efficient with C2-ACDF, underlining the importance of the C2-EF domain. None of the mini-otoferlins tested restored the sustained component of vesicle release, explaining the absence of hearing recovery. The restoration of the fast exocytotic component in the transduced Otof−/− IHCs was also associated with a recovery of Ca2+ currents with normal amplitude and fast time inactivation, confirming that the C-terminal C2 domains of otoferlin are essential for normal gating of Cav1.3 channels. Finally, the reintroduction of the mini-otoferlins C2-EF, C2-DEF, or C2-ACEF allowed us to uncover and characterize for the first time a dynamin-dependent ultrafast endocytosis in IHCs.
SIGNIFICANCE STATEMENT Otoferlin, a large six-C2-domain protein, is essential for synaptic vesicle exocytosis at auditory hair cell ribbon synapses. Here, we show that the viral expression of truncated forms of otoferlin (C2-EF, C2-DEF, and C2-ACEF) can partially rescue the fast and transient release component of exocytosis in mouse hair cells lacking otoferlin, yet cannot sustain exocytosis after long repeated stimulation. Remarkably, these hair cells also display a dynamin-dependent ultrafast endocytosis. Overall, our study uncovers the pleiotropic role of otoferlin in the hair cell synaptic vesicle cycle, notably in triggering both ultrafast exocytosis and endocytosis and recruiting synaptic vesicles to the active zone.
Keywords: calcium sensor, deafness, exocytosis, hair cells, otoferlin, ribbon synapses
Introduction
Sound encoding into electrical nerve impulses by inner hair cell (IHC) ribbon synapses is a step crucial to normal hearing. Each of 10 to 20 active zones per IHC is able to drive the spiking rate of its corresponding afferent auditory nerve fiber up to the kilohertz range and maintains its firing rate at several hundred hertz during sustained sound stimulation (Kiang, 1965; Palmer and Russell, 1986; Taberner and Liberman, 2005). To faithfully achieve this encoding process, synaptic vesicle release at the IHC active zones must occur with high temporal precision as well as reach and sustain an extremely high rate of exocytosis, which places an extraordinary demand on the molecular machinery responsible for vesicle recycling (for review, see Moser et al., 2006). Despite its great importance in maintaining membrane homeostasis and sustained vesicle release at the IHC active zone, the precise mechanism by which exocytosed vesicles are rapidly recycled through endocytosis remains largely unknown.
In CNS synapses, synaptotagmin I and II—proteins with one N-terminal transmembrane region and two C-terminal C2 domains (C2-A and C2-B)—are known to serve as essential calcium sensors for vesicle exocytosis and transmitter release (Chapman, 2008; Südhof, 2012). Although synaptotagmins are transiently expressed in immature spiking IHCs (before the onset of hearing in mice), these neuronal Ca2+ sensors are not involved in mature IHCs (Beurg et al., 2010). It is otoferlin, a large protein having six C2 domains (C2-ABCDEF) and a single transmembrane domain at its C-terminal end (Fig. 1), that serves as a Ca2+ sensor for vesicle exocytosis in cochlear (Roux et al., 2006; Michalski et al., 2017) and vestibular hair cells (Dulon et al., 2009). However, the precise molecular events by which each otoferlin C2 domain contributes to the synaptic vesicle cycle is still incompletely understood.
Figure 1.

Comparative structure of the native otoferlin and the four different mini-otoferlins inserted in the AAV vector to evaluate their role in hearing and IHC exocytosis. A, Otoferlin is a large protein (∼2000 amino acids) that contains a single TM domain at its C-terminal part, a FerA domain, and six C2 domains (C2-A to C2-F) that potentially bind calcium (Ca2+) and interact with phospholipids. Dysferlin, a dystrophy-associated Fer-1-like protein, contains seven C2 domains, a FerA domain, and a DysF domain. The dotted lines in the otoferlin and dysferlin sequences indicate a putative calpain cleavage site (Lek et al., 2012; Redpath et al., 2014). Synaptotagmins, the major Ca2+ sensors in CNS synapses, which are absent in IHCs synapses (Beurg et al., 2010), contain only two C2 domains. Note that in humans, a short isoform of otoferlin with three C2-domains (C2-DEF) and a FerA domain with unknown function has been identified (Yasunaga et al., 1999). The putative function of these truncated mini-otoferlins, C2-EF and C2-DEF, in IHC exocytosis remains unknown. Also, two abridged otoferlin-like containing C2-ACEF or C2-ACDF constructs (∼30% smaller than the native otoferlin) were tested. The AAV8 sequence contains a promoter CB6 that allows the expression of the short otoferlins (C2-EF, C2-DEF, C2-ACEF or C2-ACDF domains) and the eGFP reporter (only for C2-EF) in IHCs. (ITR as Inverted Terminal Repeat sequences). B, AAVs are injected in vivo through the round window of the cochlea at P1–P3. C, For C2-EF, the successful AAV infection of IHCs was verified with the fluorescent reporter eGFP. D, Rates of transfection for IHCs and OHCs, respectively. Error bars indicate SE. E, The expression of the mini-otoferlin C2-DEF in Otof−/− IHCs was verified by immunoconfocal imaging with a C-terminal otoferlin antibody. Note that the expression of C2-DEF was similar to that of the native otoferlin in WT IHCs.
Otoferlin has also been suggested to have a role in vesicle clathrin-dependent endocytosis, based on its interaction with the AP-2 adaptor protein complex (Duncker et al., 2013; Jung et al., 2015; Selvakumar et al., 2017) and on a reduced membrane trafficking at the ribbon active zone in Otof−/− mice (Revelo et al., 2014). Different modes of endocytosis seem to coexist in IHCs because different kinetics for membrane retrieval have been observed: one fast (with a time constant of several hundreds of milliseconds; Neef et al., 2014; Vincent et al., 2014; Michalski et al., 2017) and one slow (with a 10–20 s time constant; Moser and Beutner, 2000; Beutner et al., 2001; Cho et al., 2011). Neither of these modes of endocytosis is likely sufficiently fast to support the extremely high rate of exocytosis occurring in IHCs. A much faster compensatory endocytosis, which remains to be identified, must take place in IHCs to maintain the active zone's original surface area and prevent depletion of vesicle pools.
Recent studies from invertebrate neuromuscular junctions (Watanabe et al., 2013a) and mouse hippocampal synapses (Watanabe et al., 2013b, 2014), using high-pressure freezing electron microscopy paired with optogenetic stimulation, showed that synaptic vesicles can be recycled extremely rapidly via clathrin-independent ultrafast endocytosis, with a time constant in the range of tens of milliseconds. Here, we examined whether such an ultrafast endocytosis, which has been shown to be dynamin and actin dependent (Watanabe et al., 2014; Delvendahl et al., 2016), occurs in mouse IHCs. Furthermore, we explored the role of various C2 domain combinations in IHC exocytosis/endocytosis by using an in vivo cochlear viral cDNA transfer of various mini-otoferlin constructs in mice lacking otoferlin (Fig. 1). We found that the viral expression of mini-otoferlins that contain the C-terminal C2-E and C2-F domains, induced a restoration of the fast component of synaptic vesicle exocytosis in Otof−/− IHCs but not the sustained component during repeated stimulations. Also, the reintroduction of these mini-otoferlins allowed us to uncover a novel ultrafast endocytosis mechanism in IHCs.
Materials and Methods
Viral vector constructs.
The murine cDNA corresponding to the mini-Otof C2-EF (1896 bp; amino acids 1364–1996), C2-DEF (2763 bp; amino acids 960-1996), C2-ACDF (3465 bp), and C2-ACEF (3432 bp) domains of otoferlin were cloned into an AAV8.CB6.PI.rBG plasmid (p1045; Penn Vector Core; Fig. 1A). The C2-ACEF construct contains three disruptions: 125r394 (in amino acids) containing the C2-B domain, 946r1490 containing the C2-D domain, and 1695r1731 between the C2-E and C2-F domains. The C2-ACDF construct contains two disruptions: 125r394 containing the C2-B domain and 1110r1679 containing the C2-E domain. These mini-Otof C2-ACEF or C2-ACDF sequences were designed with a similar structure-based method developed for dysferlin (Llanga et al., 2017). Each C2 domains was analyzed by predicted β strands, potential Ca2+-binding residues, C2-domain topology and length, and continuity of the hydrophobic packing in the core of the domain. Then, the otoferlin sequence was edited in silico by defining excision sites. The absolute boundaries for each of the six C2 domains of the mouse otoferlin protein were selected based on several factors. The approximate location of each C2 domain could be delineated based on the output from the SMART database. This server (http://smart.embl-heidelberg.de/) can identify the presence of each domain; however, the structural boundaries are not necessarily clear. With the approximate boundaries defined, each domain was aligned and sorted against an extensive library of known type 1 and type 2 C2 domain ferlins. The resulting phylogenetic tree sorted each otoferlin C2 domain according to its most likely topology. Once the topology was defined, each of the eight β-strands was located by aligning the known “in-out-in-out” periodicity characteristic of C2-domain β-strands. This time-consuming method reduces the risk of misdefining a large loop insertion as a α- or β-secondary structure that may actually be unique to an individual C2 domain. Once the accurate boundaries of each C2 were established, two mini-Otofs were constructed by excluding individual domains and linker regions. Only C2-A, the central FerA domain, C2-F, and the transmembrane span were left intact. Two shorter genes that could be accommodated within an adeno-associated virus (AAV) were constructed using these rules: Otof C2-ACEF and Otof C2-ACDF. The coding sequence of each mini-Otof was synthesized (GeneCust Europe), subcloned into an AAV2 genome, and packaged in an AAV8 capsid as described by Emptoz et al. (2017).
The expressions of these different mini-Otof genes were all driven by the CB6 promoter (1 kb). Because of the 4.8 kb packaging limit of the AAV8 vector, only the sequence of the mini-Otof C2-EF was followed by an internal ribosome entry site (IRES; 0.6 kb) and the eGFP reporter gene (0.7 kb), allowing a direct monitoring of transfected IHCs (Fig. 1B). The transduction of mini-Otof C2-EF and C2-DEF was assessed by confocal immunomicroscopy, whereas the expression of mini-Otof C2-ACEF and C2-ACDF was determined by RT-PCR (see below, RT-PCR to detect C2-ACEF and C2-ACDF transcripts).
Round window membrane injection.
The AAV8 vector containing the mini-Otof sequences was injected in the cochlea at a titer of 3.21 × 1013 genome copies (gc)/ml, and the AAV8.CB6.eGFP with no inserted mini-otoferlin gene was used as control at a titer of 3.34 × 1013 gc/ml. Protocols were approved by the Animal Care and Use Committee of the Institut Pasteur and the University of Bordeaux. Intracochlear viral transduction was performed as described previously (Akil et al., 2015; Emptoz et al., 2017; Dulon et al., 2018). Mice were anesthetized using hypothermia exposure. A left postauricular incision was made to expose the otic bulla and to visualize the cochlea. A glass micropipette containing 1 μl of AAV vector preparation was inserted through the round window membrane and injected into the cochlea. The pipette was removed, the hole in the membrane was covered with connective tissue, and the incision was sealed with biological glue (3M Vetbond). Standard postoperative care was applied.
Auditory brainstem responses.
To record auditory brainstem responses (ABRs; which represent the sound-evoked synchronous firing of the auditory cochlear nerve fibers) as well as distortion product otoacoustic emissions [DPOAEs; which reflect the amplification provided by outer hair cells (OHCs)], mice were anesthetized with intraperitoneal injection of a mixture of xylazine (6 mg/ml, Rompun catalog #KP091XW, Centravet) and ketamine (80 mg/ml, Virbac catalog #4C14, Centravet) diluted in physiological saline. The mouse body temperature was kept constant at 37°C. For stimulus generation and data acquisition, we used a TDT RZ6/BioSigRZ system (Tucker-Davis Technologies). ABR signals were averaged after the presentation of a series of 512 stimulations. ABR thresholds were defined as the lowest stimuli for recognizable waves I and II. The amplitude of ABR wave I was estimated by measuring the voltage difference between the positive and negative peaks of wave I. Pure-tone stimuli were used at frequencies of 4, 8, 16, 24, and 32 kHz. Sound intensities of 10 to 90 dB SPL, in 10 dB step, were tested (Fig. 2).
Figure 2.

Viral transfer of mini-otoferlins to IHCs failed to restore ABRs in Otof−/− mice. The auditory function was tested 2 and 3 weeks (P15–P21) after the viral injection. A, Example of click-ABR waves at different sound intensities (10–90 dB SPL) in WT (black trace) and WT mice injected with AAV8-eGFP (sham-injected, green trace). Note the absence of ABR waves in both noninjected Otof−/−mice (red trace) and AAV8–C2-EF-IRES-eGFP-injected mice (light blue trace). B, C, Mean ABR wave I amplitude as a function of sound intensities (B) and tone ABR thresholds at sound frequencies from 4 to 32 kHz (C) also indicated the absence of ABR waves in injected Otof−/− mice with Otof-C2-EF (light blue) or Otof-C2-ACEF (dark blue). Note that similar results were obtained with the other mini-otoferlins, C2-DEF or C2-ACDF (not shown for clarity). Error bars indicate SE.
Tissue preparation of the organ of Corti.
Experiments were performed in accordance with the guidelines of the Animal Care Committee of the European Communities Council Directive (86/609/EEC) and were approved by the ethics committee of the University of Bordeaux (Animal Facility Agreement No. C33-063-075). All mice (C57BL6 of either sex) were anesthetized by intraperitoneal injection of a xylazin (6 mg/ml) and ketamine (80 mg/ml) mixture (Rompun, catalog #KP091XW; Vibrac, catalog #4C14) diluted in physiological saline. Electrophysiological recordings from IHCs were obtained in whole-mount organ of corti (OC) from mice at postnatal day 15 (P15)–P21, that is, after the onset of hearing. The OC was freshly dissected under binocular microscopy in an extracellular solution maintained at 4°C containing the following (in mm): 135 NaCl, 5.8 KCl, 1.3 CaCl2, 0.9 MgCl2, 0.7 NaH2PO4, 5.6 glucose, 2 Na pyruvate, and 10 HEPES, pH 7.4, 305 mOsm. The OC was then placed in a recording chamber and visualized under a Nikon 60× water immersion (WI) objective (CFI Fluor 60× Near InfraRed, Working Distance 2.0mm and Numerical Aperture 1) attached to an upright Nikon FN1 microscope coupled to a C2 confocal fluorescent imaging system. The extracellular solution was complemented with 0.25 μm of apamin (Latoxan, catalog #L8407) and 1 μm of XE-991 (Tocris Bioscience, catalog #2000) to block SK channels and KCNQ4 channels, respectively. The external Ca2+ concentration was increased from 1.3 to 5 mm to enhance the amplitude of Ca2+ currents to levels nearby body temperature. All experiments were performed at room temperature (22°C–24°C).
Patch-clamp recording and capacitance measurement.
All IHC recordings were performed in the 20–40% normalized distance from the apex, an area coding for frequencies ranging from 8 to 16 kHz, using an EPC10 amplifier controlled by Patchmaster pulse software (HEKA Elektronik). For the mini-Otof C2-EF experiments, only eGFP-expressing IHCs were recorded. For the other mini-Otof experiments, in the absence of eGFP fluorescent signaling, IHCs were randomly recorded in the tissue preparation. Patch pipettes were pulled with a micropipette puller (P-97 Flaming/Brown; Sutter Instrument) and fire polished with an MF-830 microforge (Narishige) to obtain a resistance range of 2 to 4 MΩ. Patch pipettes were filled with an intracellular cesium-based solution containing the following (in mm): 145 CsCl, 1 MgCl2, 5 HEPES, 1 EGTA, 20 TetraEthylAmmonium chloride, 2 ATP, and 0.3 GTP, pH 7.2, 300 mOsm. Changes in cell membrane capacitance (ΔCm) were used to monitor fusion of synaptic vesicles during exocytosis. The Cm was measured accordingly to the Lindau and Neher (1988) technique by using the lock-in amplifier Patchmaster software (HEKA) and applying a 1 kHz command sine wave (20 mV amplitude) at holding potential (−80 mV) before and after the pulse experiment. Because recording conditions can greatly influence capacitance measurements, only IHC patch-clamp recordings with low series resistance below 10 mΩ and a maximum leak current of 25 pA (at Vh = −80 mV) were considered in the present study.
Voltage stimulation.
Three protocols were used. First, readily releasable pool (RRP) exocytosis was recorded during a protocol in which IHCs were depolarized from −80 to −10 mV with increasing durations from 5 to 40 ms in 5 ms increments. Second, RRP exocytosis was also recorded during a protocol of five consecutive stimulations of 20 ms each, separated by 100 ms interpulses. Third, a protocol probing the replenishment of the synaptic zone in vesicles where IHCs were depolarized during a train of 100 ms pulses from −80 to −10 mV was used.
Intracellular Ca2+ uncaging.
To trigger a fast increase in intracellular Ca2+ concentration from the caged Ca2+chelator DM-Nitrophen (Interchim, catalog #317210), we used 100 ms brief flashes from a 365 nm UV light source delivered by a Mic-LED-365 (350 mW; Prizmatix). The UV LED was directly connected to the epi-illumination port at the rear of the upright Nikon FN1 microscope and illumination focalized through the 60× objective (CFI Fluor 60× W NIR, WD = 2.0 mm, NA = 1). Hair cells were loaded with the following (in mm[scap]): 145 CsCl, 5 HEPES, 20 TEA, 10 DM-Nitrophen, and 10 CaCl2. After rupture of the patch, we waited systematically for 2 min at a holding potential of −70 mV to load and equilibrate the cells with the intrapipette solution. Upon UV photolysis, the changes in [Ca2+]i, continuously measured with a C2 confocal system and NIS-Elements imaging software (Nikon) coupled to the FN1 microscope, reached nearly a mean maximum value of 20 ± 5 μm within 15–20 ms (Michalski et al., 2017), a value that was not different between wild-type (WT) and Otof−/− IHCs, as shown previously by Vincent et al. (2015).
Dyngo-4a application.
For each set of experiments, during voltage stimulation or Ca2+ uncaging, control recordings (ICa and ΔCm) were obtained in the same organ of Corti by first bathing in normal perilymph and then adding 30 μm of Dyngo-4a (dynamin inhibitor from Abcam, catalog #ab120689) for 20 min before recording its effect on ICa and ΔCm.
Immunostaining and confocal microscopy.
Rapidly dissected P15–P21 mouse cochleae were fixed with cold 4% paraformaldehyde at 4°C overnight. Once washed with cold PBS, they were decalcified for several hours in 10% EDTA buffered with PBS. After PBS washing, whole bones were removed and Reissner and tectorial membranes were carefully removed to open scala media and allow efficient antibodies access to the OC. Tissues were first incubated with PBS containing 30% normal horse serum and 0.5% Triton X-100 for 1 h at room temperature. Then, they were incubated with primary antibodies diluted with PBS (1:200) containing 5% horse serum and 0.1% Triton X-100.
Synaptic ribbons were labeled with a goat polyclonal anti-CtBP2 (Santa Cruz Biotechnology, catalog #SC-5966; RRID:AB_2086774); the otoferlin C-ter part was labeled with a rabbit polyclonal antibody (Roux et al., 2006). Once washed with PBS, tissues were incubated with a suitable mix of secondary antibodies at 1:500 for 2 h at room temperature: donkey anti-rabbit Alexa 488 (Abcam, catalog #ab150061; RRID:AB_2571722) and donkey anti-goat Fluoprobes 547H (Interchim, catalog #FP-SB2110; RRID:AB_2744533). To identify and localize the hair cells in our surface preparation of the organ of Corti, F-actin (highly concentrated in the hair cell stereocilia) was stained using tagged Phalloidin Fluoprobe 405 (1:200; Interchim, catalog #FP-CA9870). To better visualize the ribbons and otoferlin in our images, the fluorescent channel of F-actin labeling was not shown.
After PBS washing, OC surface preparations were mounted on Superfrost slides in ProLong Antifade mounting medium (Invitrogen, catalog # P7481) and kept in the dark at −20°C until observation. All comparative immunoreactions were performed simultaneously on the same days in rigorously identical conditions.
Confocal imaging was performed with a Leica SP8 confocal laser-scanning microscope (Bordeaux Imaging Center) with a 63× oil-immersion objective (NA = 1.4) and white light laser (470 to 670 nm). Phalloidin was imaged using a diode laser at 405 nm also mounted on the microscope. Stack images were acquired with the following parameters: laser power, 60%; scan rate, 700 Hz; scans averaged per XY section, four; step size, 800 nm; pixel size, 103 nm, giving an XY image size of 53 × 53 μm (512 × 512 pixels). Images were analyzed with ImageJ software (W. S. Rasband, NIH).
RT-PCR to detect C2-ACEF and C2-ACDF transcripts.
Two organ of Corti apical turns from either P50 injected with otoferlin C2-ACEF or C2-ACDF construct were freshly dissected and stored in separate low-DNA-binding tubes containing l μl of RNase inhibitor (Amresco, catalog #E633) maintained in dry ice. Total RNA was extracted from tissues using the RNAzol protocol (Molecular Research Center, catalog #RN 190). mRNA were purified using the Dynabeads mRNA Purification Kit (ThermoFisher Scientific, catalog # 61006) and reverse transcribed to cDNA by using the oligodT primers linked to the magnetic beads and AffinityScript Multiple Temperature Reverse Transcriptase (Stratagene, catalog #600107). Reverse transcription was performed at 37°C overnight. Specific PCR amplification was performed by using 0.45 μg of cDNA and Herculase II Fusion DNA Polymerase (Stratagene, catalog #600677). Specific sense and antisense primers (Eurofins Genomics) are summarized in Table 1.
Table 1.
Sense and antisense primers used
| Name of primer | Primer sequence (5′–3′) | Product size (bp) for C2-ACEF | Product size (bp) for C2-ACDF |
|---|---|---|---|
| 231s | gatagggaccttctgcatgg | 454 | 454 |
| 1494as | gaagacgacctgctcattcc | ||
| 1478s | atgagcaggtcgtcttcac | 1142 | 1142 |
| 2720 as | ggcataggcgatacgtttgt | ||
| 1478s | atgagcaggtcgtcttcac | 1146 | |
| 4563as | gatgtcggtcttgcctaact | ||
| 1478s | atgagcaggtcgtcttcac | 1341 | |
| 2919as | ggcttgcttcttggtgtag |
Data analysis.
Data were analyzed using ImageJ, Patchmaster (HEKA Elektronik), and Origin 9.1 (OriginLab). Statistical analyses were performed using independent two-sample Student t tests and, for multiple comparisons, either one-way ANOVA (two-way ANOVA in the case of Fig. 3B,C,G) followed by Tukey's post hoc test for normally distributed data (Shapiro test) or the nonparametric Kruskal–Wallis test followed by Dunn's test for not normally distributed data. Means are expressed as ±SEM. In the figures, asterisks indicate statistical significance with p < 0.05. A statistical F test was used to compare the exocytotic curve efficiency in Figure 3E.
Figure 3.

Mini-Otof constructs partially restore RRP exocytosis but not its vesicle resupply in P15–P21 Otof−/− mice. A, Example traces of vesicular exocytosis (ΔCm) and Ca2+ currents in IHCs triggered by 15 and 25 ms depolarizing steps from −80 to −10 mV in WT, Otof−/−, and C2-ACEF-, C2-EF-, C2-DEF-, and C2-ACDF-injected Otof−/− mice. B, Comparative kinetics of RRP exocytosis (depolarizing steps from −80 to −10 mV with increasing duration from 5 to 40 ms) for Otof-C2-ACEF-treated (dark blue) and C2-EF-treated (light-blue) Otof−/− IHCs. C, RRP kinetics in C2-DEF (purple) and C2-ACDF (green). For B and C, note a significant restoration of the exocytotic response compared with sham Otof−/− IHCs. *p < 0.05 (two-way ANOVA). Kinetics were best fitted with a single exponential with R2 > 0.94 except for Otof-C2-ACDF and Otof−/−. D, Comparative rate of RRP exocytosis obtained by fitting with a linear function the first four time–data points of the kinetics shown in Figure 4, B and C (linear slope in femtofarad per millisecond). The expression of the mini-Otof constructs C2-ACEF, C2-EF, and C2-DEF allowed a significant increase in the rate of exocytosis compared with sham-injected Otof−/−. *p < 0.05 (one-way ANOVA; see individual p values in Table 2). E, Ca2+ efficiency of RRP exocytosis (ΔCm/QCa) where the loading charge in Ca2+ (QCa) was obtained by integrating the Ca2+ current evoked during a voltage step from −80 to −10 mV for different brief pulses lasting 5, 10, 15, and 20 ms (full line). The slope of Otof-C2-ACEF IHCs was not significantly different of the WT slope (2.77 ± 0.21 and 2.09 ± 0.28 fF/pC, respectively; F test with F = 2.18 and p = 0.23), which is larger than in Otof−/− mice (0.39 ± 0.20 fF/pC; p = 5.8 10−3). For longer pulses (25, 30, 35, and 40 ms; dashed lines), exocytosis Ca2+ efficiency (ΔCm/QCa) in Otof-C2-ACEF IHCs is decreased, with a slope similar to that for noninjected Otof−/− mice (0.56 ± 0.14 and 0.50 ± 0.30 fF/pC, respectively; F test with F = 0.015 and p = 0.98). F, To evaluate the replenishment of the IHC active zone in vesicles, exocytosis was recorded during a train of 100 ms pulses from −80 to −10 mV, each stimulation separated by 100 ms. Examples of exocytotic traces of vesicular recruitment in control WT, Otof−/−, and Otof-C2-EF-treated IHCs. G, Comparative mean recruitment in Otof-C2-EF-, Otof-C2-DEF-, and Otof-C2-ACEF-treated IHCs. Note that in Otof−/− IHCs (red), the vesicles recruitment is severely impaired. When mini-Otof constructs are expressed in Otof−/− IHCs, the vesicle supply is only slightly, but significantly, restored compared to Otof−/− (two-way ANOVA, p = 4.3 10−8; Tukey's post hoc test, p < 10−4 for each as indicated by the asterisks), except for Otof-C2-ACDF (data not shown; p = 0.16).
Results
Viral transfer of mini-Otof cDNAs in hair cells lacking otoferlin
A single intracochlear viral injection of various AAV8 vectors containing different mini-otoferlin cDNA constructs (mini-Otofs) was performed in WT control mice and in mice lacking otoferlin (Otof−/− mice) on P1–P3 (Fig. 1A,B). Four mini-Otofs were designed and tested with regard to the following criteria: first, a synaptotagmin-like structure, that is, a mini-Otof containing only two C2 domains (C2-EF) attached to the C-terminal transmembrane domain; second, a structure similar to the short human otoferlin isoform, that is, containing the three last C-terminal C2 domains (C2-DEF) but lacking the FerA domain (Yasunaga et al., 2000); and third, two predicted abridged otoferlin-like containing C2-ACEF or C2-ACDF domain, designed using 3D structural modeling similar to that for nano-dysferlin (Llanga et al., 2017).
The rate of hair cell transduction efficacy of the mini-Otof C2-EF-IRES-eGFP was first evaluated on postnatal days P15–P21 by counting eGFP-expressing hair cells (Fig. 1C,D). It is to be noted that to avoid a possible inhibitory effect of eGFP on the function of mini-C2-EF otoferlin, as observed previously when eGFP is attached to Syt 1 (Han et al., 2005), we added an IRES between the two coding sequences in the viral vector (Fig. 1A). This IRES sequence allowed then a separate expression of the mini-Otof C2-EF and eGFP under the CB6 promoter. We found that, all along the cochlear partition, the eGFP transduction was essentially associated with the sensory hair cells of the organ of Corti, with a much higher rate for IHCs compared to OHCs: 80.4 ± 2.3% and 29.5 ± 3.5%, respectively (unpaired t test, p < 10−4; n = 22 cochleae; Fig. 1D). It is to be noted that the eGFP transduction in IHCs reached a higher percentage (above 90%) at the apical portion of the organ of Corti. In addition, the expression of the mini-Otofs C2-EF and C2-DEF in Otof−/− hair cells was directly assessed under confocal immunofluorescence microscopy at P15–P21 by using a specific antibody against the C terminal of otoferlin (Roux et al., 2006). At the apical turn of the injected cochlea, where most of electrophysiological recordings were performed, an average of 95 ± 5% of IHCs were found to express the mini-Otofs C2-EF (n = 3 mice) and C2-DEF (n = 14 mice). The subcellular distribution of these mini-Otof proteins was similar to that of the native otoferlin in wild-type mice, with a preferential higher expression in the basolateral area of IHCs where the ribbon synapses are located (Fig. 1E). Because the C2-ACEF and C2-ACDF constructs contain various disruptions in their encoding amino acid sequences (see Materials and Methods), rendering their immunodetection impossible with the various otoferlin antibodies currently in our hands, their expression in the organ of Corti was positively assessed at the mRNA level by RT-PCR (see Materials and Methods; data not shown).
Absence of auditory function recovery by cochlear injection of mini-Otof cDNAs
Mice lacking otoferlin (Otof−/− mice) are profoundly deaf due to a defect in synaptic vesicle release at the IHC ribbon synapse (Roux et al., 2006; Fig. 2). Measurements of ABRs were performed to evaluate a possible recovery in auditory function in AAV8-mini-Otof-injected Otof−/− mice. As a sham, we first verified that cochlear injection of AAV8-eGFP not containing the mini-Otof cDNAs did not affect hearing sensitivity of WT mice (Fig. 2A, green trace). The input–output function (click ABR-wave-I amplitude as a function of sound intensity; Fig. 2B) and the thresholds at different frequencies were similar between control WT sham-injected mice and WT noninjected mice (Fig. 2C). Furthermore, we found that hearing thresholds of WT mice were not affected for up to 3 months after cochlear injection, indicating the innocuity of the surgical and viral procedures, as reported previously (Emptoz et al., 2017; Dulon et al., 2018). The DPOAEs of WT mice, which probe the activity of OHCs, also remained unaffected by viral injections (data not shown). Cochlear injection of the various AAV8-mini-Otofs tested, C2-EF (n = 22 mice), C2-DEF (n = 42 mice), C2-ACEF (n = 13 mice), C2-ACDF (n = 8 mice), and C2-ACEF together with C2-ACDF (n = 5 mice), did not rescue the hearing phenotype in Otof−/− mice. These mice were tested for up to 3 months after injection. These results suggested that the cochlear injection of the various mini-Otofs tested did not restore normal synaptic exocytotic properties of Otof−/− IHCs.
Viral transfer of mini-Otofs in Otof−/− IHCs partially restores exocytosis of the readily releasable pool of vesicles but not its vesicle resupply
Although the measurement of auditory evoked potential (ABRs) is a good method to determine the overall integrity of the auditory system, this method is not precise enough to detect subtle changes in auditory function, notably those associated with certain synaptopathies produced by noise exposure and aging, which are thus classified as “hidden hearing loss” (Liberman, 2017). In this context, and in a reverse way, it is possible that subtle synaptic recovery after mini-Otof injections in Otof−/− mice cannot be detected in ABRs recordings. To assess whether IHC exocytosis was somewhat improved in mini-Otof-injected Otof−/− mice, we directly recorded IHCs using whole-cell patch-clamp experiments. The study of the presynaptic exocytotic function in IHCs was performed at P15–P21 in organ of Corti explants dissected either from WT (Otof +/+) or Otof−/− mice injected with various AAV8-mini-Otofs (Fig. 1). All IHC patch-clamp recordings were performed in the 20–40% normalized distance from the apex, an area encoding sound frequencies ranging from 8 to 16 kHz.
First, we controlled the innocuity of the viral construct on IHC exocytosis. Recordings from P15–P21 WT IHCs of the cochlea injected with AAV8-eGFP (mice with no inserted mini-Otof are sham-injected mice) displayed similar exocytotic responses as P15–P21 WT IHCs from noninjected cochleae. Indeed, for a 15 ms voltage step, from −80 to −10 mV, AAV8-eGFP-transduced IHCs showed a mean capacitance jump of 6.0 ± 1.4 fF (n = 6), a value comparable to that for WT IHCs from noninjected mouse cochleae (6.7 ± 1.0 fF; n = 6; unpaired t test, p = 0.71; data not shown). We also found no significant difference in the exocytotic responses between IHCs from WT sham-injected and WT noninjected mice for longer and repetitive depolarizing steps that involved vesicle recruitment (data not shown). As expected, we observed no rescue effect on exocytosis in Otof−/− IHCs. For a 15 ms voltage step, from −80 to −10 mV, Otof−/− IHCs transduced with AAV8-eGFP showed a mean capacitance jump of 0.83 ± 0.5 fF (n = 6), a value comparable to that for Otof−/− IHCs from noninjected mouse cochleae (1.5 ± 0.4 fF; n = 13; unpaired t test, p = 0.38). Because IHCs from sham-injected mice displayed exocytotic properties similar to those of IHCs from noninjected mice, these data were combined and used as controls. Controls were named WT or Otof−/− in the rest of the study.
Conversely, recordings from Otof−/− IHCs of the cochleae injected with AAV-Otof-C2-EF-IRES-eGFP displayed a partial rescue of exocytosis when stimulated with brief depolarization impulses lasting <20 ms (Fig. 3A,B,D). These brief voltage impulses are believed to mobilize the RRP of vesicles at the active zones of the IHCs (Moser and Beutner, 2000; Michalski et al., 2017). For a 25 ms impulse from −80 to −10 mV, exocytosis in Otof-C2-EF-transduced IHCs, directly visualized with eGFP fluorescence, averaged 3.8 ± 0.8 fF (n = 14; from six mice issued from three different litters), a value significantly larger than that for littermate noninjected and sham (control) Otof−/− mice (1.5 ± 0.4 fF; one-way ANOVA, p = 2.25 10−10; post hoc Tukey's test, p = 0.045) but significantly smaller than that for control WT IHCs (10.3 ± 1.4 fF; one-way ANOVA, post hoc Tukey's test, p = 4.6 10−6; Fig. 3A–C). The kinetics of exocytosis Otof-C2-EF-transduced IHCs were significantly higher than in Otof−/− IHCs (two-way ANOVA, p = 6.8 10−13; post hoc Tukey's test, p = 1.8 10−8; Fig. 3B).
A similar partial rescue of exocytosis was obtained in Otof−/− IHCs from Otof-C2-DEF virus-injected cochlea (Fig. 3A–C; data from four mice of three different litters). Note that in this latter set of experiments with Otof-C2-DEF, in the absence of eGFP (because of the size limit of the cDNA insertion in the AAV vector; see Materials and Methods), randomly selected IHCs were recorded in the organ of Corti. Only one of seven IHCs did not show a significant increase in exocytosis compared with noninjected Otof−/− mice. This high rate of exocytosis rescue in AAV-Otof-C2-DEF-injected cochleae was consistent with the high percentage (>90%) of IHCs expressing the mini-otoferlin protein (Fig. 1E). The kinetics of exocytosis for Otof-C2-DEF-transduced IHCs were significantly higher than in Otof−/− IHCs (two-way ANOVA, p = 6.8 10−13; post hoc Tukey's test, p = 1.7 10−3; Fig. 3C).
Remarkably, recordings from Otof−/− IHCs of AAV-Otof-C2-ACEF-injected cochlea displayed a more efficient rescue of exocytosis when stimulated with brief depolarizing impulses lasting <20 ms (Fig. 3A,B). Only 1 of 10 IHCs recorded from AAV-Otof-C2-ACEF-injected mice did not produce a ΔCm response different from that of Otof−/− IHCs of noninjected cochleae, again a result in good agreement with the high percentage of transduced IHCs (Fig. 1C,D). The kinetics of exocytosis for Otof-C2-ACEF were significantly higher than in Otof−/− IHCs (two-way ANOVA, p = 6.8 10−13, p < 10−4; Fig. 3B). The kinetics of exocytosis with Otof-C2-ACDF were also found to be significantly higher than in Otof−/− IHCs (two-way ANOVA, p = 6.8 10−13, p = 2.2 10−3; Fig. 3C).
The most common method to quantify the RRP is to fit the data with a single exponential function, with time constant (τ) giving 63.2% of the RRP value. For WT control IHCs, we found a RRP value of 19.3 ± 2.6 fF with τ = 29.5 ± 6.5 ms. In our experimental conditions, whereas the data for Otof-C2-EF (RRP = 5.0 ± 0.8 fF with τ = 15.5 ± 7.3 ms), Otof-C2-ACEF (RRP = 5.9 ± 0.3 fF with τ = 8.5 ± 1.9 ms), and Otof-C2-DEF (RRP = 4.5 ± 0.5 fF with τ = 19.0 ± 5.2 ms) could be fitted with a single exponential, we could not accurately fit the data for Otof-C2-ACDF and Otof−/− with such a function and therefore determine RRP values for these conditions. Therefore, to be able to statistically compare all mini-Otof conditions and controls within a single one-way ANOVA, we instead compared the initial rate of RRP exocytosis. This RRP rate was obtained for each condition by fitting the first four time points of each of the kinetics with a linear function. All mini-Otofs, except Otof-C2-ACDF, showed a significant increase in the rate of the RRP exocytosis compared with Otof−/− IHCs (Fig. 3D, Table 2).
Table 2.
Summary of exocytosis properties (with p values indicating individual comparison with Otof−/−)
| Function | WT | Otof−/− + C2-EF | Otof−/− + C2-DEF | Otof−/− + C2-ACEF | Otof−/− + C2-ACDF | Otof−/− | |
|---|---|---|---|---|---|---|---|
| Fig. 3D, one-way ANOVA, p = 1.15 10−10 | RRP rate (fF/ms) | 0.51 ± 0.07 (n = 12) | 0.35 ± 0.06 (n = 14) | 0.27 ± 0.05 (n = 6) | 0.49 ± 0.07 (n = 10) | 0.17 ± 0.02 (n = 9) | 0.028 ± 0.012 (n = 19) |
| *p = 2.2 10−5 | *p = 0.047 | *p = 2.7 10−8 | p = 0.308 | ||||
| Fig. 3E, F test | RRP Ca2+ efficiency (fF/pC; full line) | 2.77 ± 0.21 (n = 10) | 1.49 ± 0.23 (n = 10) | 0.47 ± 0.12 (n = 6) | 2.09 ± 0.28 (n = 9) | 0.50 ± 0.24 (n = 9) | 0.39 ± 0.20 (n = 10) |
| p = 0.058 | p = 0.97 | *p = 0.018 | p = 0.92 | ||||
| Fig. 4C, one-way ANOVA, p = 1.35 10−3 | ICa total (pA) | 205 ± 13 (n = 13) | 204 ± 7 (n = 20) | 207 ± 9 (n = 7) | 203 ± 7 (n = 10) | 202 ± 7 (n = 10) | 159 ± 14 (n = 16) |
| *p = 0.006 | *p = 0.019 | *p = 0.025 | *p = 0.022 | ||||
| Fig. 4D, Kruskal–Wallis, p = 1.5 10−6 | Fast ICa inactivation (pA) | 20.2 ± 4.0 (n = 12) | 11.6 ± 2.7 (n = 19) | 18.7 ± 3.1 (n = 7) | 8.1 ± 3.2 (n = 10) | 0.6 ± 0.4 (n = 10) | 5.0 ± 1.8 (n = 17) |
| p = 0.249 | *p = 0.030 | p = 0.999 | p = 0.999 |
* indicates significantly different with p < 0.05.
The Ca2+ efficiency of RRP exocytosis (ΔCm/QCa; QCa is defined as the integral of the Ca2+ current) in Otof-C2-ACEF IHCs was also found to be similar to that in WT controls for brief depolarizations lasting up to 15 ms, with mean slopes of 2.77 ± 0.21 fF/pC and 2.09 ± 0.28 fF/pC, respectively (Fig. 3E; F test with F = 2.18, p = 0.23). These results indicate that the abridged short otoferlin, Otof-C2-ACEF, was able to trigger fast vesicular membrane fusion as efficiently as the native six-C2 otoferlin.
However, Otof−/− IHCs treated with the mini-Otof, although showing partial recovery of the RRP release, could not efficiently sustain exocytosis for voltage impulses during a train of stimulations compared with WT IHCs (Fig. 3F,G), suggesting a defect in the vesicle pool replenishment process. The Ca2+ efficiency of exocytosis (ΔCm/QCa) measured for voltage steps beyond 20 ms up to 40 ms was significantly reduced from 1.69 ± 0.29 fF/pC in WT to 0.56 ± 0.14 fF/pC in Otof-C2-ACEF IHCs (Fig. 3E; F test with F = 16.0, p = 0.0039). The replenishment rate, calculated during a train of depolarizations, was reduced by a factor of 3 to 5 compared with WT IHCs, although it was significantly larger, except for Otof-C2-ACDF, than for Otof−/− IHCs from noninjected mice (Fig. 3G). This defect in the vesicle pool resupply to the RRP, found also in the pachanga mouse model with a point mutation in C2-F (Pangrsic et al., 2010), explains the absence of ABR waves in these mini-Otof-injected mice (Fig. 2).
Mini-Otof constructs partially restore the fast Ca2+ current inactivation in Otof−/− IHCs
Cochlear IHCs have been shown to express long and short C-terminal Cav1.3 isoforms (Scharinger et al., 2015, Vincent et al., 2017). Short C-terminal Cav1.3 channel (Cav1.3S) isoforms lacking the C-terminal modulator (Bock et al., 2011) are rapidly inactivated by calmodulin (CaM) and are thought to carry the fast component of Ca2+ current inactivation in IHCs (representing 15–30% of the total calcium current; Vincent et al., 2017; Fig. 4A). The distal and proximal C-terminal regulatory domains (DCRD and PCRD, respectively) of long Cav1.3 (Cav1.3L) isoforms compete with CaM, and these channels do not present fast inactivation. These Cav1.3L isoforms likely support the major part of the Ca2+ current in IHCs (Fig. 4A; Vincent et al., 2017). Unlike WT IHCs, IHCs lacking otoferlin display Ca2+ currents with weak or no inactivation, suggesting that otoferlin is required for the functional expression of Cav1.3S isoforms in IHCs (Vincent et al., 2017; Fig. 4B,4D). WT IHCs displayed a fast inactivating component with an amplitude of 20 ± 4 pA, with τ = 20.3 ± 3.3 ms, when fitting the first 50 ms of the current trace with a single exponential function, an amplitude significantly larger than that for Otof−/− IHCs (5 ± 2 pA; Kruskal–Wallis, p = 1.5 10−6; Dunn's post hoc test with p = 3.7 10−3; with similar τ = 20.4 ± 3.5 ms). Also, the peak amplitude of the Ca2+ currents was smaller in P15–P21 Otof−/− IHCs compared with P15–P21 WT IHCs (Fig. 4C; one-way ANOVA, p = 1.3 10−3; Tukey's post hoc test, p = 5.3 10−3). Remarkably, the expression of all the mini-Otofs tested, even Otof-C2-ACDF, which poorly rescues RRP exocytosis, restored the amplitude of Ca2+ currents in P15–P21 Otof−/− IHCs to levels comparable to those of WT IHCs (Fig. 4C, Table 2). However, only Otof-C2-DEF restored the fast inactivating component of the Ca2+ current in Otof−/− IHCs (mean amplitude of 19 ± 3 pA with τ = 20.6 ± 3.2 ms) compared to Otof−/− IHCs (5 ± 2 pA with τ = 20.4 ± 3.5 ms), when IHCs were stepped from −80 to −10 mV (Table 2, Fig. 4D). These results are in good agreement with previous in vitro protein–protein interaction assays showing that otoferlin can interact through its C-terminal C2 domains, notably C2-D, with the Cav1.3 channels (Ramakrishnan et al., 2009; Hams et al., 2017). Furthermore, they confirm that otoferlin is essential for the functional expression of the fast inactivating Cav1.3S in IHCs (Vincent et al., 2017), likely through a functional interaction between C2-DEF and Cav1.3S. Note that none of the combinations of the four C2 domain constructs C2-ACEF or C2-ACDF resulted in a significant recovery of the fast inactivating component of Ca2+ current (Fig. 4D). We recall that to insert the four C2 domains of the mini-Otof into the AAV construct, we truncated some of the inter-C2 sequence between the domains C2-A–C2-C and C2-E–C2-F for ACEF (Fig. 1A; Materials and Methods). A similar truncation was done for ACDF. It is therefore possible that these inter-C2 sequences are important for Ca2+ current inactivation while leaving intact the membrane expression level of the Ca2+ channels.
Figure 4.

AAV gene transfer of mini-Otof restores the fast inactivating component of the Ca2+ current in P15–P21 Otof−/− IHCs. A, IHCs are thought to express long and short C-terminal Cav1.3 isoforms (Scharinger et al., 2015, Vincent et al., 2017). Short C-terminal Cav1.3 channel isoforms are rapidly inactivated by calmodulin and are thought to carry the fast component of Ca2+ current inactivation (Vincent et al., 2017). C-terminal regulatory domains (PCRD and DCRD) of long Cav1.3 isoforms compete with CaM, and these channels, representing the major isoforms, do not present fast inactivation (IQ indicates the isoleucine-glutamine motif domain interacting with Ca2+/CaM). B, Unlike WT IHCs (black trace), IHCs lacking otoferlin (red trace) do not display Ca2+ current with fast inactivation (Vincent et al., 2017). C, Peak ICa (ICa total) is significantly diminished in Otof−/− mice compared with WT mice, but the expression of all mini-otoferlins allows the recovery of the ICa current (one-way ANOVA; see Table 2). D, Only when the mini-otoferlin C2-DEF is virally expressed in IHCs is the amplitude of the inactivating component of the Ca2+ current (fitted with a single exponential over the first 50 ms of the current trace) significantly recovered compared with Otof−/− (Kruskal–Wallis; see Table 2). IHCs were depolarized for 100 ms from −80 to −10 mV. *p < 0.05. Error bars indicate SE.
Mini-Otof constructs partially restore the number of synaptic ribbons in Otof−/− IHCs
As described previously by Vincent et al. (2017), we found that IHCs from Otof−/− mice displayed a significantly lower number of synaptic ribbons at P15–P17 compared to WT IHCs (Fig. 5A,C): 10.7 ± 0.3 ribbons per cell (n = 70 Otof−/− IHCs from three mice) and 16.2 ± 0.3 (n = 40 WT IHCs from three mice; one-way ANOVA, p < 10−4; Tukey's post hoc test, p < 10−4), respectively. This comparative quantification of the ribbon numbers was done in the same cochlear area where the patch-clamp recordings were performed (20–40% normalized distance from the cochlear apex corresponding to encoding frequencies between 8 and 16 kHz). Note that the mean number of ribbons per WT IHC is consistent with previous findings (Meyer et al., 2009; Vincent et al., 2014). We found that the viral expression of Otof-C2-EF in P15–P17 Otof−/− IHCs leads to a significant increase in the number of ribbons per IHC compared with noninjected and sham Otof−/− IHCs: 11.9 ± 0.4 ribbons per IHC (n = 43 IHCs from three mice; one-way ANOVA, p = 0.022; Fig. 5C). The number of synaptic ribbons per IHC in control untreated Otof−/− mice continued to decrease to 7.1 ± 0.3 ribbons (n = 56 IHCs) at P30–P32 compared to P15–P17 (unpaired t test, p < 10−4; Fig. 5C,D). Again, cochlear injection of C2-ACEF in Otof−/− mice led to a significant preservation of about two ribbons per IHC at P30–P32 compared to untreated control Otof−/− mice (8.8 ± 0.2; n = 31 IHCs; Kruskal–Wallis, p = 6.7 10−9; Dunn's post-test, p = 1.8 10−3; Fig. 5D). The incomplete preservation of the ribbons with C2-EF and C2-ACEF was in line with the partial restoration of the RRP responses (Fig. 3A,B).
Figure 5.

Mini-Otof constructs partially rescued the synaptic ribbon numbers the synaptic ribbon numbers in Otof−/− IHCs. A, Using immunofluorescent confocal imaging, we quantified the number of synaptic ribbons (red dots) at P15–P17 in IHCs from WT, Otof−/− expressing the otoferlin C2-EF domain, and Otof−/− mice. The green signal indicates otoferlin expression in WT and C2-EF but not in noninjected Otof−/− IHCs. B, Immunolabeling examples of synaptic ribbons (at larger magnification; without otoferlin immunolabeling) in P30–P32 IHCs from a WT (control) mouse, an Otof−/− mouse injected with C2-ACEF, and a noninjected Otof−/− mouse. The dashed white line delineates the cell contour at the basal synaptic area of one IHC for each category. C, Comparative ribbon histogram at P15–P17 for C2-EF: IHCs of Otof−/− mice displayed a largely reduced number of ribbons (10.7 ± 0.3; n = 70 IHCs; 3 mice) compared with WT mice (16.2 ± 0.3, n = 40 IHCs, one-way ANOVA, p < 10−4). The Otof-C2-EF expression in Otof−/− IHCs had a significantly larger number of ribbons per cell (11.9 ± 0.4; n = 43 IHCs) compared with control, noninjected Otof−/− mice (10.7 ± 0.3; n = 70 IHCs; one-way ANOVA, p = 0.022). D, Comparative histograms at P30–P32 for C2-ACEF: IHCs of C2-ACEF-injected mice also displayed a significant preservation of synaptic ribbons (8.8 ± 0.2, n = 31 IHCs) compared with control, untreated Otof−/− mice (7.1 ± 0.3; n = 56 IHCs; Kruskal–Wallis, p = 1.8 10−3). E, The resting size of Otof−/− IHCs, measured in whole-cell recordings at −70 mV, was diminished compared to the WT IHC resting size (8.8 ± 0.09 pF, n = 53; 10.79 ± 0.14 pF, n = 43 respectively; one-way ANOVA, p < 10−4). None of the mini-Otofs significantly restored the resting size of IHCs. The asterisks indicate significantly different with p < 0.05 (for each compared to Otof−/− mice, one-way ANOVA). Error bars indicate SE.
Also, as described previously (Vincent et al., 2014, 2015, 2017), we found that the resting whole-cell membrane capacitance of IHCs from Otof−/− mice, measured at VH = −70 mV, was significantly reduced compared with WT IHCs: 8.8 ± 0.09 pF (n = 53) and 10.79 ± 0.14 pF (n = 43; one-way ANOVA, p = 2.9 10−7; Tukey's post hoc test, p = 8.7 10−7), respectively (Fig. 5E). This reduction in surface membrane area of IHCs lacking otoferlin can be explained by a reduced resting vesicular exocytosis and a sustained otoferlin-independent slow endocytosis (Vincent et al., 2014). The viral expression of all mini-Otof in Otof−/− IHCs, did not produced a significant change in the resting IHC membrane capacitance compared with Otof−/− IHCs (one-way ANOVA; Tukey's post hoc test, p > 0.05; Fig. 5E).
Mini-Otof constructs reveal ultrafast endocytosis in Otof−/− IHCs
Dyngo-4a, by blocking endocytosis, increases exocytosis evoked by voltage activation of Ca2+ channels
Dyngo-4a is a highly potent inhibitor of dynamin, a GTP binding protein essential for ultrafast endocytosis (Watanabe et al., 2013a, b). Dynamin is thought to polymerize around the neck of the synaptic vesicles that are fused with the plasma membrane, leading to a fast membrane fission and pinching off of the vesicle from the plasma membrane active zone. Both dynamin 1 and 3 are expressed in IHCs (Neef et al., 2014; Shen et al., 2015). We first tested the effect of external application of 30 μm Dyngo-4a on the mobilization of the RRP evoked by five consecutive brief 20 ms depolarizing pulses, with an interpulse interval of 100 ms, in P15–P21 WT IHCs (Fig. 6A). Remarkably, the dynamin inhibitor largely increased the RRP responses (Fig. 6A,C). The increase in ΔCm responses was not the result of increased Ca2+ currents since similar amplitudes were found in the presence or absence of the endocytotic inhibitor (Fig. 6D). If we consider that the ΔCm responses result from the net difference between exocytosis and endocytosis, the increasing effect of Dyngo-4a on the ΔCm response suggested that a 20 ms time scale endocytosis in IHCs was blocked, a time scale in the range of the ultrafast endocytosis described in central synapses (Watanabe et al., 2013b). Notably, application of Dyngo-4a had no effect on the ΔCm responses recorded in Otof−/− IHCs, suggesting that the small residual otoferlin-independent exocytosis was not large enough to trigger measurable ultrafast endocytosis (Fig. 6B,C). Alternatively, this result suggested that otoferlin is required for ultrafast endocytosis.
Figure 6.

The effect of a dynamin inhibitor on RRP exocytosis. A, B, We tested the effect of 30 μm Dyngo-4a (dynamin inhibitor) on the ΔCm response in P15–P21 IHCs from WT (A) and in Otof−/− mice (B) by applying five consecutive brief 20 ms depolarizing pulses. C, The presence of Dyngo-4a increased the net ΔCm responses in WT IHCs by blocking endocytosis. *p < 0.05 (unpaired t test). Dyngo-4a did not affect the ΔCm responses in Otof−/− IHCs. D, The increase in ΔCm responses in WT IHCs was not the result of augmented Ca2+ currents since similar amplitudes were found in the presence or absence of the endocytotic inhibitor. Error bars indicate SE.
Dyngo-4a increases the ΔCm responses triggered upon intracellular Ca2+ uncaging
The effect of Dyngo-4a was also tested on ΔCm responses evoked during intracellular Ca2+ uncaging experiments (Fig. 7). In these conditions, in WT IHCs, we observed a significant increase in the rate of the ΔCm responses, calculated as the first derivative at each point of the ΔCm response curve, in the presence of Dyngo-4a responses (maximal dCm/dt of 42.8 ± 2.2 fF/ms and 88.4 ± 3.9 fF/ms, respectively, without and with Dyngo-4a; unpaired t test, p = 0.019). Also, the maximum amplitude was significantly larger in the presence of Dyngo-4a (from 1293 ± 26 fF to 1813 ± 57 fF; unpaired t test, p = 0.027; Fig. 7A), again suggesting that ultrafast endocytosis occurs concomitantly with exocytosis. Subtracting the ΔCm responses produced by Ca2+ uncaging in the presence of Dyngo-4a from those obtained in the absence of Dyngo-4a indicated an ultrafast endocytosis with a time constant of τ = 10.9 ± 0.5 ms, a maximal amplitude of 737.1 ± 24.9 fF, and a maximal rate (dCm/dt) of 70.7 fF/ms (Fig. 7B, Table 3). Of note, Dyngo-4a had no significant effect on the rate of ΔCm responses evoked by Ca2+ uncaging in noninjected Otof−/− IHCs (maximal rates of 2.0 ± 0.1 fF/ms and 2.2 ± 0.1 fF/ms, with and without Dyngo-4a, respectively; unpaired t test, p = 0.74), again indicating that the rate of exocytosis in the absence of otoferlin is too slow to trigger ultrafast endocytosis and/or that otoferlin is required for endocytosis (Fig. 7C,D).
Figure 7.

Dyngo-4a effects on exocytosis evoked by intracellular Ca2+ uncaging A, We tested the effect of Dyngo-4a on the exocytotic responses evoked by intracellular Ca2+ uncaging in WT P15–P21 IHCs. In the presence of Dyngo-4a (dark blue), we observed an increased in the kinetics and amplitude of the ΔCm responses. B, The subtraction of the ΔCm responses reveals the presence of an ultrafast endocytosis (best fitted with a single exponential with τ = 10.9 ± 0.5 ms). C, D, The presence of Dyngo-4a has no effect on exocytosis in Otof−/− IHCs. E, When Dyngo-4a was added to Otof-C2-EF IHCs, the initial rate of the exocytotic response was transiently increased. F, The subtraction of the ΔCm responses revealed a transient ultrafast endocytosis (τ = 7.6 ± 1.0 ms) followed by exocytosis.
Table 3.
Summary of ultrafast endocytosis
| Subtraction | Amplitude (fF) | Time constant (ms) | Mean maximal rate (dCm/dt, in fF/ms) |
|---|---|---|---|
| Fig. 7B | |||
| WT (n = 6) − [WT (n = 6) + Dyngo-4a] | 737.1 | 10.9 ± 0.5 | 70.7 |
| Fig. 7D | |||
| Otof−/− (n = 6) − [Otof−/− (n = 4) + Dyngo-4a] | — | — | — |
| Fig. 7F | |||
| C2-EF (n = 6) − [C2-EF (n = 6) + Dyngo-4a] | 44.7 | 7.6 ± 1.0 | 7.9 |
| Fig. 8D | |||
| [Otof−/− (n = 6) + C2-EF] − Otof −/− (n = 7) | 38.2 | 10.7 ± 1.3 | 5.0 |
| Fig. 8B | |||
| [Otof−/− (n = 11) + C2-DEF] − Otof −/− (n = 7) | 35.8 | 16.0 ± 4.7 | 7.0 |
| Fig. 8F | |||
| [Otof−/− (n = 6) + C2-ACEF] − Otof −/− (n = 6) (exocytosis) | 57.3 | 5.2 ± 2.5 | 6.9 |
| [Otof−/− (n = 6) + C2-ACEF] − Otof −/− (n = 6) (endocytosis) | 65.7 | 87.2 ± 12.9 | 0.8 |
| Data not shown | |||
| [Otof−/− (n = 8) + C2-ACDF] − Otof −/− (n = 7) | 32.7 | 21.5 ± 10.7 | 1.6 |
A dash indicates a nonmeasurable endocytosis.
We then tested the effects of Dyngo-4a on Ca2+ uncaging-evoked exocytosis in Otof−/− IHCs expressing Otof-C2-EF. Remarkably, the presence of Dyngo-4a significantly increased the initial mean rate of exocytosis during the first 20 ms after the UV flash (from 0.9 ± 0.1 fF/ms without Dyngo-4a and 2.8 ± 0.1 fF/ms with Dyngo-4a; unpaired t test, p = 0.02; Fig. 7E). Subtracting the traces in the presence of Dyngo-4a from those in its absence again indicated a transient fast initial decrease in ΔCm, that is, an ultrafast endocytosis with a maximal amplitude of 44.7 ± 4.3 fF, a time constant of 7.6 ± 1.0 ms, and a maximal rate of 7.9 fF/ms (Fig. 7F, Table 3). Surprisingly, the endocytotic response was transient and followed by an increase in membrane capacitance (Fig. 7F). This was likely due to the fact that in the presence of Dyngo-4a, the exocytotic response depressed more rapidly with time than that for the untreated control Otof−/− IHCs (Fig. 7E). This suggests that vesicle endocytosis could participate in local fast recycling of synaptic vesicles, and this process contributes to sustain release.
It is unlikely that the IHC capacitance responses measured during Ca2+ uncaging result or is contaminated by thermal excitation of the mechanoelectrical transduction apparatus and depolarization (Azimzadeh et al., 2018). In this later study, the power density of the UV laser used to generate thermal effect in hair cells is extremely high, more than three orders of magnitude larger than our UV power density generated with our UV LED. Our UV LED flash produced no significant thermal effect to stimulate the hair cells. Strong arguments for this assumption come from the absence of sustained repetitive Cm responses once intracellular DM-Nitrophen is photolyzed, when applying repetitive consecutive UV flashes, or the absence of Cm response when flashing the IHCs with the UV LED in the absence of intracellular DM-Nitrophen (our unpublished data). Furthermore, recordings in whole-cell voltage-clamp conditions should minimize UV-induced membrane depolarization in our study.
Why was endocytosis, unmasked by Dyngo-4a, first transiently observed during Ca2+ uncaging in Otof−/− IHCs Otof-C2-EF? One possible explanation is that endocytosis, a Ca2+-dependent process (Yamashita et al., 2010), is more Ca2+ sensitive compared with exocytosis but is inhibited at high Ca2+ concentrations at which exocytosis is still activated, as proposed previously at synaptic terminals of bipolar neurons by von Gersdorff and Matthews (1994a,b). It is worth noting that our UV-LED-Ca2+ uncaging system was measured to rise the concentration of intracellular Ca2+ in IHCs to 20 μm within 15–20 ms (Michalski et al., 2017).
The amplitude of endocytosis, revealed by Ca2+ uncaging and the effect of Dyngo-4a, was much smaller in Otof−/− IHCs expressing the mini-Otof constructs C2-EF compared with WT IHCs. However, the endocytotic time constant was similarly ultrafast, respectively 7.6 and 10.9 ms, suggesting that this Dyngo-4a-sensitive process was similar. The difference in rate and amplitude could be explained by the fact that the mini-otoferlin C2-EF incompletely restores the rate of vesicle fusion and/or, more importantly, the vesicle replenishment, as indicated in Figure 3.
Mini-Otof constructs reveal ultrafast endocytosis during intracellular Ca2+ uncaging
When compared with a new set of noninjected Otof−/− IHCs, uncaging intracellular Ca2+ in P15–P21 IHCs from mouse injected with AAV8-C2-DEF displayed an initial decrease in the rate of the ΔCm responses from 1.83 ± 0.01 fF/ms to 1.10 ± 0.01 fF/ms (unpaired t test, p = 0.036, Fig. 8A). The subtraction of the traces gave a mean ultrafast endocytosis amplitude of 35.8 ± 4.6 fF, a time constant of 16.0 ± 4.7 ms and maximal rate dCm/dt of 7.0 fF/ms (Fig. 8B, Table 3). Similar ultrafast endocytosis was obtained with Otof-C2-EF (Fig. 8C–D, Table 3) and Otof-C2-ACDF (curve not shown; Table 3). The initial rates of endocytosis produced by Otof-C2-DEF, Otof-C2-EF, and Otof-C2-ACDF may have been underestimated because the change in membrane capacitance upon Ca2+ uncaging likely results at each point of the curve from the balance of exocytosis and endocytosis and we cannot rule out that these two events occur concomitantly. Remarkably, the initial endocytosis was rapidly followed by exocytosis in IHCs from Otof-C2-DEF and Otof-C2-EF injected mice but not in Otof-C2-ACDF (data not shown), in agreement with their respective efficiency in restoring exocytosis when voltage-stepping the cells (Fig. 3A–C). During the voltage-activation of Ca2+ currents in Figure 3A–C, it is likely that we missed the initial transient ultrafast endocytosis because of the recording blind window inherent to the single sine-wave Cm measurement technique during the Ca2+ current activation.
Figure 8.

AAV gene transfer of mini-otoferlins revealed otoferlin-dependent ultrafast endocytosis in IHCs. A, Uncaging intracellular Ca2+ in P15–P21 IHCs from a mouse injected with AAV8-Otof-C2-DEF (purple trace) displayed an initial drop in their ΔCm responses compared with noninjected Otof−/− IHCs (red trace). B, Subtracting the ΔCm responses of C2-DEF-injected IHCs from those of the noninjected Otof−/− IHCs indicated a transient ultrafast endocytosis that could be fitted with a single exponential with τ = 16.0 ± 4.7 ms. This transient endocytosis was followed by exocytosis. C, D, C2-EF injection (C, light blue trace) also showed a transient initial ultrafast endocytosis (τ = 10.7 ± 1.3 ms). E, IHCs from a mouse injected with AAV8-Otof-C2-ACEF (dark blue trace) showed an initial transient increase of the exocytotic response that was followed by a rapid endocytosis compared with noninjected Otof−/− IHCs (red trace). Note that the insets in A, C, E expand the Y axis of the Cm responses to see more details at the onset. F, The subtraction of the ΔCm responses (black trace) indicated a transient exocytosis (τ = 5.2 ± 2.5 ms) followed by a fast endocytotic response (τ = 87.2 ± 12.9 ms). Note that for each condition, new Otof−/− IHCs were simultaneously recorded in the same set of experiments. For each condition, traces are means from n IHCs, indicated in brackets in A, C, E.
Uncaging intracellular Ca2+ in IHCs from mouse injected with AAV8-C2-ACEF showed an initial increase of the exocytotic response from 2.31 ± 0.03 fF/ms to 3.90 ± 0.14 fF/ms (Fig. 8E; unpaired t test, p = 0.022). Subtracting the ΔCm responses of Otof-C2-ACEF-injected IHCs from noninjected Otof−/− IHCs (new set of Otof−/− IHCs recorded simultaneously to Otof-C2-ACEF-injected IHCs) indicated first a transient exocytosis with mean amplitude of 57.3 ± 6.4 fF, a time constant of 5.2 ± 2.5 ms and maximal rate dCm/dt of 6.9 fF/ms (Fig. 8F, Table 3). This initial exocytotic response was transient and followed by a fast Cm decrease likely reflecting ultrafast endocytosis with a mean amplitude of 65.7 ± 4.1 fF, a time constant of 87.2 ± 12.9 ms and maximal rate dCm/dt of 0.8 fF/ms. Why Ca2+ uncaging in Otof−/− IHCs expressing Otof-C2-ACEF first produced a transient fast exocytosis? This is possibly due to the higher efficiency of Otof-C2-ACEF to restore RRP exocytosis compared to Otof-C2-EF or Otof-C2-DEF (Fig. 3). Then, endocytosis in Otof-C2-ACEF IHCs was only revealed secondary to exocytosis when the vesicular RRP pool was exhausted because of the absence of an efficient vesicle replenishment. Alternatively Otof-C2-ACEF, while being highly Ca2+ efficient for exocytosis could be much less effective for endocytosis as indicated by its lower rate and time constant compared to Otof-C2-EF.
Discussion
Restoration of RRP exocytosis with mini-otoferlins
We showed that expression of the mini-otoferlins Otof-C2-EF, Otof-C2-DEF, and Otof-C2-ACEF can partially restore RRP exocytosis in Otof−/− IHCs. Otof-C2-ACEF was the most efficient, with restoration of exocytosis to levels similar to that for WT mice for stimulations <15 ms (Fig. 3B). C2-EF and C2-DEF constructs also restored exocytosis but only partially, up to 40% of the WT level. Interestingly, Otof-C2-ACDF was the less efficient mini-Otof and therefore revealed the importance of C2-E domain, or its association with C2-F domain at the C-terminal region of otoferlin. The C2-E domain has been shown to potentially bind to the t-SNARE protein syntaxin 1A in vitro (Ramakrishnan et al., 2009), a protein that is expressed in IHCs (Safieddine and Wenthold, 1999) and is generally implicated in the docking of synaptic vesicles with the presynaptic plasma membrane. The importance of the C2-F domain of otoferlin has been emphasized in the pachanga mouse model bearing a substitution point mutation in this domain (Pangrsic et al., 2010). In pachanga mice, IHCs have normal initial vesicle fusion of docked/primed vesicles but a strong defect in vesicle recruitment at the active zone. The C2-F domain can interact in vitro with both t-SNARE proteins (syntaxin-1A and SNAP-25) and phosphatidylinositol 4,5-bisphosphate (PIP2) at the plasma membrane (Roux et al., 2006; Ramakrishnan et al., 2009; Padmanarayana et al., 2014; Fig. 9), similarly to the C2-B domain of synaptotagmin (Wang et al., 2016). We propose that the tandem C2-EF domain of otoferlin is at least necessary for vesicle fusion at the active zone of IHCs, presumably by interacting with the vesicle-attached t-SNARE proteins and the plasma membrane. Notably, truncated forms of otoferlin retaining the C-terminal C2-F domain were also shown to rescue the otoferlin knock-out phenotype in zebrafish (Chatterjee et al., 2015).
Figure 9.
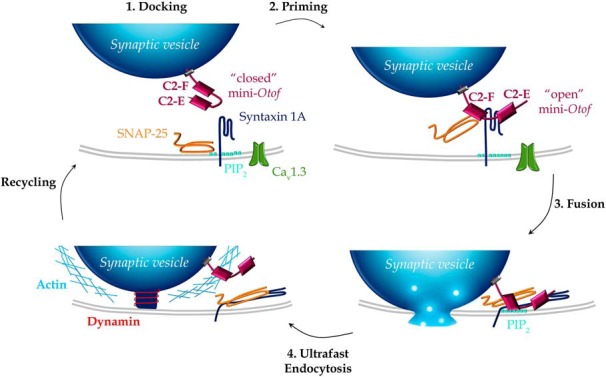
Schematic model for the engagement of the mini-otoferlin C2-EF in the synaptic vesicle cycle (exocytosis and endocytosis) at the IHC active zone. 1, Docking. The mini-Otof C2-EF, attached by its C-terminal transmembrane domain to the glutamatergic docked vesicles, is not yet engaged in direct interactions with the t-SNAREs (syntaxin 1A and SNAP-25) and the phospholipids PIP2 at the plasma membrane. At low intracellular Ca2+ concentration, when nearby Ca2+ channels are not voltage activated, the mini-otoferlin is folded, and its C2-E and C2-F domains interact with each other. 2, Priming. During voltage activation of the Ca2+ channels, [Ca2+]i increases and vesicles undergo a priming reaction where mini-Otof conformation shift from “closed” to “open” and interact with t-SNARE, forming a complex. Syntaxin 1A has been proposed to interact with C2-E and C2-F domains and SNAP-25 only with C2-F (Ramakrishnan et al., 2014). This step of the vesicle cycle is poorly restored by Otof-C2-EF, Otof-C2-DEF, Otof-C2-ACEF, and Otof-C2-ACDF (recruitment, Fig. 3G), explaining the absence of hearing restoration (Fig. 2). The molecular interactions of the mini-Otof complex with the Cav1.3 channels are not represented. 3, Fusion. When Ca2+ ions bind to the C2 domains of mini-Otof, the synaptic vesicle membrane and the plasma membrane are forced into close proximity by the SNARE complex, triggering partial insertion of the C2-F domain into PIP2 and resulting in vesicle fusion. This step of vesicle fusion is partially restored by Otof-C2-EF, Otof-C2-DEF, and Otof-C2-ACEF. 4, Ultrafast vesicle endocytosis. This is mediated by dynamin, which pinches off the vesicular membrane with the help of filamentous F-actin for initializing the membrane curvature. This ultrafast endocytosis was efficiently observed with Otof-C2-EF, Otof-C2-DEF, Otof-C2-ACEF, and Otof-C2-ACDF that could interact with the dynamin process.
Interestingly, the C2-EF domain of another Ferlin-like C2-domain protein, dysferlin, is considered essential for Ca2+-triggered vesicle–plasma membrane fusion in skeletal muscle cells and cardiomyocytes (Lek et al., 2012). A “mini-dysferlin,” constituted by C2-EF domains and the C-terminal transmembrane domain, can be produced by intracellular calpain-1 digestion. This release of C2-EF was found to recruit vesicles to the sites of membrane injury faster than the long native form (Lek et al., 2013). A calpain-1 cleavage was also found in otoferlin, forming a possible mini-otoferlin with C2-EF and transmembrane (TM) domains (Redpath et al., 2014). Previous studies hypothesized that C-terminal dual-C2 domains (synaptotagmin-like modules) released through enzymatic cleavage of ferlin proteins have a specialized vesicle-fusion modality (Redpath et al., 2014). However, our observation that the tandem C2-EF domain of otoferlin only poorly restored RRP exocytosis suggests that this mini-otoferlin does not have a crucial function in IHCs.
Remarkably, addition of C2-A and C2-C domains to the Otof-C2-EF construct led to a more efficient recovery of RRP exocytosis in Otof−/− IHCs. These C2 domains can also potentially interact with syntaxin 1A, SNAP-25, PIP2, and the L-type voltage-gated Ca2+ Cav1.3 channels (Ramakrishnan et al., 2009; Padmanarayana et al., 2014). Addition of C2-A and C2-C to C2-EF could therefore allow increased synaptic protein interactions such as a multiple binding to SNARE proteins simultaneously (Hams et al., 2017). Furthermore, C2-A and C2-C domains associated to C2-EF, forming a longer construct, could also facilitate protein folding and self C2–C2 interactions (Fig. 9). Indeed, the Otof-C2-ACEF could more easily form a folding tertiary structure “closed” at low Ca2+ concentrations that could “open” as Ca2+ increases to promote vesicle fusion, this model being proposed for the native otoferlin by Ramakrishnan et al. (2014). Note that open and closed states of C2-A and C2-B domains have also been proposed for synaptotagmins (Fuson et al., 2007; Evans et al., 2016). We propose that the C2-ACEF domains of otoferlin are necessary and sufficient to promote efficient Ca2+-evoked vesicle fusion in IHCs.
Surprisingly, the Otof-C2-ACEF construct failed to restore hearing in mice lacking otoferlin, although partially rescuing RRP exocytosis. The lack of hearing restoration could be easily explained at the IHC level by the absence of efficient vesicle recruitment to the RRP vesicle component, as similar defects were also found in pachanga mice (Pangrsic et al., 2010). Our study confirms that the full length of otoferlin is required for efficient Ca2+-dependent recruitment of the synaptic vesicles to the active zone. Full-length otoferlin, by promoting additional C2–C2 interactions, could promote the formation of long tethers holding and priming the synaptic vesicles at the IHC active zones, a crucial structure for vesicle replenishment (Vogl et al., 2016).
Another unexpected result was that the C2-DEF mini-otoferlin did not produce a better exocytosis restoration compared with C2-EF. Notably, this C2-DEF construct, although lacking the FerA domain, closely corresponds to the short cDNA isoform found in humans but not in mice (Yasunaga et al., 2000). Therefore, our study suggests either that the short human-like otoferlin isoform does not function as an efficient Ca2+ sensor for exocytosis in mouse IHCs or that the FerA domain, a four helix bundle that can bind to lipids in a Ca2+-dependent manner (Harsini et al., 2018), missing in our C2-DEF construct, is essential for an efficient fast vesicular fusion.
Otoferlin and Ca2+ channel interaction
At the IHC active zone, the Cav1.3 L-type Ca2+ channels, governing Ca2+-triggered synaptic exocytosis in IHCs, are thought to be expressed as long and short C-terminal isoforms (Scharinger et al., 2015; Vincent et al., 2017). The short isoforms (Cav1.3S), with fast inactivation, are essential to drive the RRP exocytosis in IHCs, whereas the long isoforms (Cav1.3L) with slow inactivation regulate vesicle recruitment (Vincent et al., 2017). Notably, Cav1.3S isoforms, but not Cav1.3L isoforms, associate in functional cooperative clusters of two or more channels, to facilitate Ca2+ currents in hippocampal neurons (Moreno et al., 2016). Similarly, Cav1.3S could aggregate in cooperative clusters to potentiate Ca2+ influx at the active zone of IHCs.
The absence of otoferlin in IHCs is associated with a lack of the fast inactivating component of the Ca2+ currents and a decreased expression of Cav1.3S (Vincent et al., 2017), suggesting a functional interaction between otoferlin and Cav1.3S channels at the IHC synapses. Direct interactions between the II–III cytoplasmic loop of Cav1.3 and otoferlin, via the C2-D or C2-ABDF domains of otoferlin, have been suggested (Ramakrishnan et al., 2009; Hams et al., 2017). Thus, the restoration of the fast inactivating component of the Ca2+ currents in Otof−/− IHCs with Otof-C2-DEF is in agreement with the literature and confirms that the C-terminal C2 domains of otoferlin interact with Cav1.3S channels. Surprisingly, the Otof-C2-ACEF construct did not produce a significant recovery of the fast inactivating component of ICa. Notably this construct, contains a small deletion revealed by 3D structural modeling between the C2-E and C2-F (1695aar1731aa), which could alter the interaction with Cav1.3 channels.
Otoferlin and ultrafast endocytosis
A striking finding of our study was the expression of ultrafast endocytosis in auditory IHCs. Although different slow modes of endocytosis, likely clathrin dependent, have been described in IHCs (Beutner et al., 2001; Cho et al., 2011; Neef et al., 2014; Jung et al., 2015), they remain too slow to support the unusually high rate of exocytosis in auditory IHCs. Previous studies have revealed the existence of ultrafast endocytosis with a time constant of 50 ms in mouse hippocampal synapses (Watanabe et al., 2013b, 2014). The molecular mechanisms underlying ultrafast endocytosis are not fully understood, but its dependence on dynamin to pinch off the endocytic vesicle is well recognized. Consistent with these previous studies, we here report a novel dynamin-dependent ultrafast endocytosis in IHCs that can compensate exocytosis at a 20 ms time scale. This time scale is too fast for the dynamin-dependent clathrin-mediated endocytosis known to occur in IHCs at time scales ranging from 300 ms to 30 s, and we found that ultrafast endocytosis was absent in IHCs lacking otoferlin. We propose that otoferlin functions as a dual Ca2+ sensor for exocytosis and endocytosis, much like synaptotagmin I (Poskanzer et al., 2003; Yao et al., 2011).
Ultrafast endocytosis in neuronal synapses is sensitive to latrunculin-A, a sponge toxin that disrupts the F-actin filaments at synaptic active zones (Watanabe et al., 2013b). The F-actin cytoskeleton is thought to be essential for the polymerization and constriction of dynamin molecules around the membrane neck of the vesicles before fission and scission. Interestingly, latrunculin-A has also been shown previously to disrupt F-actin at the IHC ribbon active zones and increase the rate of exocytosis (Vincent et al., 2015; Guillet et al., 2016). We believe that increased exocytotic rates produced by latrunculin-A could also be, at least in part, due to inhibition of ultrafast endocytosis, resulting in a positive effect on exocytosis similar to that observed with Dyngo-4a.
Conclusions
Our study shows the importance of the C-terminal C2 domains of otoferlin in triggering RRP exocytosis and regulating ultrafast endocytosis in IHCs. However, these mini-otoferlins did not restore vesicle replenishment at the IHC active zones. This process possibly requires cooperative intramolecular interactions between C2 domains of otoferlin, similar to those of the tandem C2 domains of synaptotagmin-1 (Evans et al., 2016).
Footnotes
This work was supported by a grant from the Fondation Pour l'Audition (D.D.), European Commission Grant ERC-2011-ADG_294570, LabEx Lifesenses Grant ANR-10-LABX-65, and French government funds managed by the Agence Nationale de la Recherche (EargenCure).
The authors declare no competing financial interests.
References
- Akil O, Rouse SL, Chan DK, Lustig LR (2015) Surgical method for virally mediated gene delivery to the mouse inner ear through the round window membrane. J Vis Exp. Advance online publication. Retrieved March 16, 2015. doi: 10.3791/52187. 10.3791/52187 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Azimzadeh JB, Fabella BA, Kastan NR, Hudspeth AJ (2018) Thermal excitation of the mechanotransduction apparatus of hair cells. Neuron 97:586–595.e4. 10.1016/j.neuron.2018.01.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beurg M, Michalski N, Safieddine S, Bouleau Y, Schneggenburger R, Chapman ER, Petit C, Dulon D (2010) Control of exocytosis by synaptotagmins and otoferlin in auditory hair cells. J Neurosci 30:13281–13290. 10.1523/JNEUROSCI.2528-10.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beutner D, Voets T, Neher E, Moser T (2001) Calcium dependence of exocytosis and endocytosis at the cochlear inner hair cell afferent synapse. Neuron 29:681–690. 10.1016/S0896-6273(01)00243-4 [DOI] [PubMed] [Google Scholar]
- Bock G, Gebhart M, Scharinger A, Jangsangthong W, Busquet P, Poggiani C, Sartori S, Mangoni ME, Sinnegger-Brauns MJ, Herzig S, Striessnig J, Koschak A (2011) Functional properties of a newly identified C-terminal splice variant of Cav1.3 L-type Ca2+ channels. J Biol Chem 286:42736–42748. 10.1074/jbc.M111.269951 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chapman ER. (2008) How does synaptotagmin trigger neurotransmitter release? Annu Rev Biochem 77:615–641. 10.1146/annurev.biochem.77.062005.101135 [DOI] [PubMed] [Google Scholar]
- Chatterjee P, Padmanarayana M, Abdullah N, Holman CL, LaDu J, Tanguay RL, Johnson CP (2015) Otoferlin deficiency in zebrafish results in defects in balance and hearing: rescue of the balance and hearing phenotype with full-length and truncated forms of mouse otoferlin. Mol Cell Biol 35:1043–1054. 10.1128/MCB.01439-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cho S, Li GL, von Gersdorff H (2011) Recovery from short-term depression and facilitation is ultrafast and Ca2+ dependent at auditory hair cell synapses. J Neurosci 31:5682–5692. 10.1523/JNEUROSCI.5453-10.2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delvendahl I, Vyleta NP, von Gersdorff H, Hallermann S (2016) Fast, temperature-sensitive and clathrin-independent endocytosis at central synapses. Neuron 90:492–498. 10.1016/j.neuron.2016.03.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dulon D, Safieddine S, Jones SM, Petit C (2009) Otoferlin is critical for a highly sensitive and linear calcium-dependent exocytosis at vestibular hair cell ribbon synapses. J Neurosci 29:10474–10487. 10.1523/JNEUROSCI.1009-09.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dulon D, Papal S, Patni P, Cortese M, Vincent PF, Tertrais M, Emptoz A, Tlili A, Bouleau Y, Michel V, Delmaghani S, Aghaie A, Pepermans E, Alegria-Prevot O, Akil O, Lustig L, Avan P, Safieddine S, Petit C, El-Amraoui A (2018) Clarin-1 gene transfer rescues auditory synaptopathy in model of Usher syndrome. J Clin Invest 128:3382–3401. 10.1172/JCI94351 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duncker SV, Franz C, Kuhn S, Schulte U, Campanelli D, Brandt N, Hirt B, Fakler B, Blin N, Ruth P, Engel J, Marcotti W, Zimmermann U, Knipper M (2013) Otoferlin couples to clathrin-mediated endocytosis in mature cochlear inner hair cells. J Neurosci 33:9508–9519. 10.1523/JNEUROSCI.5689-12.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Emptoz A, Michel V, Lelli A, Akil O, Boutet de Monvel J, Lahlou G, Meyer A, Dupont T, Nouaille S, Ey E, Franca de Barros F, Beraneck M, Dulon D, Hardelin JP, Lustig L, Avan P, Petit C, Safieddine S (2017) Local gene therapy durably restores vestibular function in a mouse model of usher syndrome type 1G. Proc Natl Acad Sci U S A 114:9695–9700. 10.1073/pnas.1708894114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans CS, He Z, Bai H, Lou X, Jeggle P, Sutton RB, Edwardson JM, Chapman ER (2016) Functional analysis of the interface between the tandem C2 domains of synaptotagmin-1. Mol Biol Cell 27:979–989. 10.1091/mbc.E15-07-0503 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuson KL, Montes M, Robert JJ, Sutton RB (2007) Structure of human synaptotagmin 1 C2AB in the absence of Ca2+ reveals a novel domain association. Biochemistry 46:13041–13048. 10.1021/bi701651k [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guillet M, Sendin G, Bourien J, Puel JL, Nouvian R (2016) Actin filaments regulate exocytosis at the hair cell ribbon synapse. J Neurosci 36:649–654. 10.1523/JNEUROSCI.3379-15.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hams N, Padmanarayana M, Qiu W, Johnson CP (2017) Otoferlin is a multivalent calcium-sensitive scaffold linking SNAREs and calcium channels. Proc Natl Acad Sci U S A 114:8023–8028. 10.1073/pnas.1703240114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han W, Rhee JS, Maximov A, Lin W, Hammer RE, Rosenmund C, Südhof TC (2005) C-terminal ECFP fusion impairs synaptotagmin 1 function: crowding out synaptotagmin 1. J Biol Chem 280:5089–5100. 10.1074/jbc.M408757200 [DOI] [PubMed] [Google Scholar]
- Harsini FM, Chebrolu S, Fuson KL, White MA, Rice AM, Sutton RB (2018) FerA is a membrane-associating four-helix bundle domain in the ferlin family of membrane-fusion proteins. Sci Rep 8:10949. 10.1038/s41598-018-29184-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jung S, Maritzen T, Wichmann C, Jing Z, Neef A, Revelo NH, Al-Moyed H, Meese S, Wojcik SM, Panou I, Bulut H, Schu P, Ficner R, Reisinger E, Rizzoli SO, Neef J, Strenzke N, Haucke V, Moser T (2015) Disruption of adaptor protein 2μ (AP-2μ) in cochlear hair cells impairs vesicle reloading of synaptic release sites and hearing. EMBO J 34:2686–2702. 10.15252/embj.201591885 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiang N. (1965) Discharge patterns of single fibers in the cat's auditory nerve. Cambridge, MA: MIT. [Google Scholar]
- Lek A, Evesson FJ, Sutton RB, North KN, Cooper ST (2012) Ferlins: regulators of vesicle fusion for auditory neurotransmission, receptor trafficking and membrane repair. Traffic 13:185–194. 10.1111/j.1600-0854.2011.01267.x [DOI] [PubMed] [Google Scholar]
- Lek A, Evesson FJ, Lemckert FA, Redpath GM, Lueders AK, Turnbull L, Whitchurch CB, North KN, Cooper ST (2013) Calpains, cleaved mini-dysferlinC72, and L-type channels underpin calcium-dependent muscle membrane repair. J Neurosci 33:5085–5094. 10.1523/JNEUROSCI.3560-12.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liberman MC. (2017) Noise-induced and age-related hearing loss: new perspectives and potential therapies. F1000Res. 6:927. 10.12688/f1000research.11310.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lindau M, Neher E (1988) Patch-clamp techniques for time-resolved capacitance measurements in single cells. Pflugers Arch 411:137–146. 10.1007/BF00582306 [DOI] [PubMed] [Google Scholar]
- Llanga T, Nagy N, Conatser L, Dial C, Sutton RB, Hirsch ML (2017) Structure-based designed nano-dysferlin significantly improves dysferlinopathy in BLA/J mice. Mol Ther 25:2150–2162. 10.1016/j.ymthe.2017.05.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer AC, Frank T, Khimich D, Hoch G, Riedel D, Chapochnikov NM, Yarin YM, Harke B, Hell SW, Egner A, Moser T (2009) Tuning of synapse number, structure and function in the cochlea. Nat Neurosci 12:444–453. [DOI] [PubMed] [Google Scholar]
- Michalski N, Goutman JD, Auclair SM, Boutet de Monvel J, Tertrais M, Emptoz A, Parrin A, Nouaille S, Guillon M, Sachse M, Ciric D, Bahloul A, Hardelin JP, Sutton RB, Avan P, Krishnakumar SS, Rothman JE, Dulon D, Safieddine S, Petit C (2017) Otoferlin acts as a Ca2+ sensor for vesicle fusion and vesicle pool replenishment at auditory hair cell ribbon synapses. Elife 6:e31013. 10.7554/eLife.31013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moreno CM, Dixon RE, Tajada S, Yuan C, Opitz-Araya X, Binder MD, Santana LF (2016) Ca(2+) entry into neurons is facilitated by cooperative gating of clustered Cav1.3 channels. Elife 5:e15744. 10.7554/eLife.15744 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moser T, Beutner D (2000) Kinetics of exocytosis and endocytosis at the cochlear inner hair cell afferent synapse of the mouse. Proc Natl Acad Sci U S A 97:883–888. 10.1073/pnas.97.2.883 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moser T, Neef A, Khimich D (2006) Mechanisms underlying the temporal precision of sound coding at the inner hair cell ribbon synapse. J Physiol 576:55–62. 10.1113/jphysiol.2006.114835 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neef J, Jung S, Wong AB, Reuter K, Pangrsic T, Chakrabarti R, Kügler S, Lenz C, Nouvian R, Boumil RM, Frankel WN, Wichmann C, Moser T (2014) Modes and regulation of endocytic membrane retrieval in mouse auditory hair cells. J Neurosci 34:705–716. 10.1523/JNEUROSCI.3313-13.2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Padmanarayana M, Hams N, Speight LC, Petersson EJ, Mehl RA, Johnson CP (2014) Characterization of the lipid binding properties of otoferlin reveals specific interactions between PI(4,5)P2 and the C2C and C2F domains. Biochemistry 53:5023–5033. 10.1021/bi5004469 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palmer AR, Russell IJ (1986) Phase-locking in the cochlear nerve of the guinea-pig and its relation to the receptor potential of inner hair-cells. Hear Res 24:1–15. 10.1016/0378-5955(86)90002-X [DOI] [PubMed] [Google Scholar]
- Pangrsic T, Lasarow L, Reuter K, Takago H, Schwander M, Riedel D, Frank T, Tarantino LM, Bailey JS, Strenzke N, Brose N, Müller U, Reisinger E, Moser T (2010) Hearing requires otoferlin-dependent efficient replenishment of synaptic vesicles in hair cells. Nat Neurosci 13:869–876. 10.1038/nn.2578 [DOI] [PubMed] [Google Scholar]
- Poskanzer KE, Marek KW, Sweeney ST, Davis GW (2003) Synaptotagmin I is necessary for compensatory synaptic vesicle endocytosis in vivo. Nature 426:559–563. 10.1038/nature02184 [DOI] [PubMed] [Google Scholar]
- Ramakrishnan NA, Drescher MJ, Drescher DG (2009) Direct interaction of otoferlin with syntaxin 1A, SNAP-25, and the L-type voltage-gated calcium channel Cav1.3. J Biol Chem 284:1364–1372. 10.1074/jbc.M803605200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramakrishnan NA, Drescher MJ, Morley BJ, Kelley PM, Drescher DG (2014) Calcium regulates molecular interactions of otoferlin with soluble NSF attachment protein receptor (SNARE) proteins required for hair cell exocytosis. J Biol Chem 289:8750–8766. 10.1074/jbc.M113.480533 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Redpath GM, Woolger N, Piper AK, Lemckert FA, Lek A, Greer PA, North KN, Cooper ST (2014) Calpain cleavage within dysferlin exon 40a releases a synaptotagmin-like module for membrane repair. Mol Biol Cell 25:3037–3048. 10.1091/mbc.e14-04-0947 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Revelo NH, Kamin D, Truckenbrodt S, Wong AB, Reuter-Jessen K, Reisinger E, Moser T, Rizzoli SO (2014) A new probe for super-resolution imaging of membranes elucidates trafficking pathways. J Cell Biol 205:591–606. 10.1083/jcb.201402066 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roux I, Safieddine S, Nouvian R, Grati M, Simmler MC, Bahloul A, Perfettini I, Le Gall M, Rostaing P, Hamard G, Triller A, Avan P, Moser T, Petit C (2006) Otoferlin, defective in a human deafness form, is essential for exocytosis at the auditory ribbon synapse. Cell 127:277–289. 10.1016/j.cell.2006.08.040 [DOI] [PubMed] [Google Scholar]
- Safieddine S, Wenthold RJ (1999) SNARE complex at the ribbon synapses of cochlear hair cells: analysis of synaptic vesicle- and synaptic membrane-associated proteins. Eur J Neurosci 11:803–812. 10.1046/j.1460-9568.1999.00487.x [DOI] [PubMed] [Google Scholar]
- Scharinger A, Eckrich S, Vandael DH, Schönig K, Koschak A, Hecker D, Kaur G, Lee A, Sah A, Bartsch D, Benedetti B, Lieb A, Schick B, Singewald N, Sinnegger-Brauns MJ, Carbone E, Engel J, Striessnig J (2015) Cell-type-specific tuning of Cav1.3 Ca(2+)-channels by a C-terminal automodulatory domain. Front Cell Neurosci 9:309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selvakumar D, Drescher MJ, Deckard NA, Ramakrishnan NA, Morley BJ, Drescher DG (2017) Dopamine D1A directly interacts with otoferlin synaptic pathway proteins: Ca2+ and phosphorylation underlie an NSF-to-AP2mu1 molecular switch. Biochem J 474:79–104. 10.1042/BCJ20160690 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen J, Scheffer DI, Kwan KY, Corey DP (2015) SHIELD: an integrative gene expression database for inner ear research. Database (Oxford) 2015:bav071 10.1093/database/bav071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Südhof TC. (2012) Calcium control of neurotransmitter release. Cold Spring Harb Perspect Biol 4:a011353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taberner AM, Liberman MC (2005) Response properties of single auditory nerve fibers in the mouse. J Neurophysiol 93:557–569. 10.1152/jn.00574.2004 [DOI] [PubMed] [Google Scholar]
- Vincent PF, Bouleau Y, Safieddine S, Petit C, Dulon D (2014) Exocytotic machineries of vestibular type I and cochlear ribbon synapses display similar intrinsic otoferlin-dependent Ca2+ sensitivity but a different coupling to Ca2+ channels. J Neurosci 34:10853–10869. 10.1523/JNEUROSCI.0947-14.2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vincent PF, Bouleau Y, Petit C, Dulon D (2015) A synaptic F-actin network controls otoferlin-dependent exocytosis in auditory inner hair cells Elife 4:e10988 10.7554/eLife.10988 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vincent PF, Bouleau Y, Charpentier G, Emptoz A, Safieddine S, Petit C, Dulon D (2017) Different CaV1.3 channel isoforms control distinct components of the synaptic vesicle cycle in auditory inner hair cells. J Neurosci 37:2960–2975. 10.1523/JNEUROSCI.2374-16.2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vogl C, Panou I, Yamanbaeva G, Wichmann C, Mangosing SJ, Vilardi F, Indzhykulian AA, Pangršič T, Santarelli R, Rodriguez-Ballesteros M, Weber T, Jung S, Cardenas E, Wu X, Wojcik SM, Kwan KY, Del Castillo I, Schwappach B, Strenzke N, Corey DP, et al. (2016) Tryptophan-rich basic protein (WRB) mediates insertion of the tail-anchored protein otoferlin and is required for hair cell exocytosis and hearing. EMBO J 35:2536–2552. 10.15252/embj.201593565 [DOI] [PMC free article] [PubMed] [Google Scholar]
- von Gersdorff H, Matthews G (1994a) Dynamics of synaptic vesicle fusion and membrane retrieval in synaptic terminals. Nature 367:735–739. 10.1038/367735a0 [DOI] [PubMed] [Google Scholar]
- von Gersdorff H, Matthews G (1994b) Inhibition of endocytosis by elevated internal calcium in a synaptic terminal. Nature 370:652–655. 10.1038/370652a0 [DOI] [PubMed] [Google Scholar]
- Wang S, Li Y, Ma C (2016) Synaptotagmin-1 C2B domain interacts simultaneously with SNAREs and membranes to promote membrane fusion. Elife 5:e14211. 10.7554/eLife.14211 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watanabe S, Liu Q, Davis MW, Hollopeter G, Thomas N, Jorgensen NB, Jorgensen EM (2013a) Ultrafast endocytosis at caenorhabditis elegans neuromuscular junctions. Elife 2:e00723. 10.7554/eLife.00723 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watanabe S, Rost BR, Camacho-Pérez M, Davis MW, Söhl-Kielczynski B, Rosenmund C, Jorgensen EM (2013b) Ultrafast endocytosis at mouse hippocampal synapses. Nature 504:242–247. 10.1038/nature12809 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watanabe S, Trimbuch T, Camacho-Pérez M, Rost BR, Brokowski B, Söhl-Kielczynski B, Felies A, Davis MW, Rosenmund C, Jorgensen EM (2014) Clathrin regenerates synaptic vesicles from endosomes. Nature 515:228–233. 10.1038/nature13846 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamashita T, Eguchi K, Saitoh N, von Gersdorff H, Takahashi T (2010) Developmental shift to a mechanism of synaptic vesicle endocytosis requiring nanaodomain Ca2+. Nat Neurosci 13:838–844. 10.1038/nn.2576 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yao J, Kwon SE, Gaffaney JD, Dunning FM, Chapman ER (2011) Uncoupling the roles of synaptotagmin I during endo- and exocytosis of synaptic vesicles. Nat Neurosci 15:243–249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yasunaga S, Grati M, Cohen-Salmon M, El-Amraoui A, Mustapha M, Salem N, El-Zir E, Loiselet J, Petit C (1999) A mutation in OTOF, encoding otoferlin, a FER-1-like protein, causes DFNB9, a nonsyndromic form of deafness. Nat Genet 21:363–369. 10.1038/7693 [DOI] [PubMed] [Google Scholar]
- Yasunaga S, Grati M, Chardenoux S, Smith TN, Friedman TB, Lalwani AK, Wilcox ER, Petit C (2000) OTOF encodes multiple long and short isoforms: genetic evidence that the long ones underlie recessive deafness DFNB9. Am J Hum Genet 67:591–600. 10.1086/303049 [DOI] [PMC free article] [PubMed] [Google Scholar]