

Abstract
Bacteria can utilize diverse sugars as carbon and energy source, but the regulatory mechanisms directing the choice of the preferred substrate are often poorly understood. Here, we analyzed the role of the YugA protein (now designated GlaR—Galactose–lactose operon Regulatory protein) of the RpiR family as a transcriptional activator of galactose (gal genes) and lactose (lac genes) utilization genes in Lactococcus lactis IL1403. In this bacterium, gal genes forming the Leloir operon are combined with lac genes in a single so‐called gal–lac operon. The first gene of this operon is the lacS gene encoding galactose permease. The glaR gene encoding GlaR lies directly upstream of the gal–lac gene cluster and is transcribed in the same direction. This genetic layout and the presence of glaR homologues in the closest neighborhood to the Leloir or gal–lac operons are highly conserved only among Lactococcus species. Deletion of glaR disabled galactose utilization and abrogated or decreased expression of the gal–lac genes. The GlaR‐dependent regulation of the gal–lac operon depends on its specific binding to a DNA region upstream of the lacS gene activating lacS expression and increasing the expression of the operon genes localized downstream. Notably, expression of lacS‐downstream genes, namely galMKTE, thgA and lacZ, is partially independent of the GlaR‐driven activation likely due to the presence of additional promoters. The glaR transcription itself is not subject to catabolite control protein A (CcpA) carbon catabolite repression (CRR) and is induced by galactose. Up to date, no similar mechanism has been reported in other lactic acid bacteria species. These results reveal a novel regulatory protein and shed new light on the regulation of carbohydrate catabolism in L. lactis IL1403, and by similarity, probably also in other lactococci.
Keywords: galactose assimilation, Lactococcus lactis, Leloir pathway, sugar metabolism, transcription regulation
1. INTRODUCTION
Lactose, a disaccharide comprised of galactose linked through a β‐glycosidic bond to the C4 of glucose, is the dominant sugar found in milk. Lactic acid bacteria (LAB) are capable of growth in milk owing to an efficient use of lactose as a carbon source. Because of the high efficiency and economic relevance of lactose fermentation, numerous studies have focused on LAB. Lactose utilization genes have been characterized in many LAB species, and it has been shown that they can take up lactose by two principally different ways including the lactose‐specific phosphotransferase system (lac‐PTS) and secondary transporters such as lactose‐galactose antiporters and lactose‐H+ symporters (reviewed by Aleksandrzak‐Piekarczyk, 2013). The secondary transport systems transfer unphosphorylated lactose via specific permeases of the LacS subfamily (TC No. 2.A.2.2.3) belonging to the 2.A.2 glycoside‐pentoside‐hexuronide (GPH) family (Saier, 2000). After its import, lactose is hydrolyzed by β‐galactosidase to glucose and galactose. Then, glucose is further metabolized via glycolysis, while the galactose moiety can be either released into the medium or converted into glucose‐1‐phosphate (Glc‐1‐P), which enters glycolysis following conversion to Glc‐6‐P. The conversion into Glc‐1‐P is performed by the action of four enzymes that constitute the Leloir pathway (De Vos, 1996; Poolman, 1993; Vaughan, van den Bogaard, Catzeddu, Kuipers, & de Vos, 2001). This pathway, discovered by L. F. Leloir and coworkers in 1950s (reviewed in Frey1996), consists of the crucial enzyme galactokinase (GalK) plus hexose‐1‐P uridylyltransferase (GalT) and UDP‐glucose 4‐epimerase (GalE) that perform the conversion of galactose into glucose‐1‐P. Found more recently, an additional enzyme, the GalM mutarotase (aldose‐1‐epimerase), is involved in the interconversion of the galactose α‐ and β‐anomers (Bouffard, Rudd, & Adhya, 1994).
In LAB, the Leloir pathway genes may be present on their own (gal genes) or combined with genes for lactose metabolism (lac genes) (Grossiord, Vaughan, Luesink, & de Vos, 1998; Vaillancourt, Moineau, Frenette, Lessard, & Vadeboncoeur, 2002). In the latter case, in addition to the galKTEM genes (depending on a LAB species, in a variable genomic organization and order), extra genes such as lacZ (β‐galactosidase) and lacA (thgA; thiogalactoside acetyltransferase) genes are present (gal–lac operon) (Aleksandrzak‐Piekarczyk, Kok, Renault, & Bardowski, 2005; Poolman, Royer, Mainzer, & Schmidt, 1990; Vaillancourt et al., 2002). Directly upstream of these genes encoding enzymes catalyzing lactose hydrolysis and/or galactose conversion, or within this operon, a gene encoding specific permease for lactose or galactose uptake may also be present (Grossiord et al., 1998; Vaillancourt et al., 2002).
The uptake and metabolism of sugars is mastered by numerous regulatory proteins which form a regulatory network detecting environments and setting the catabolic abilities of the cell, thus helping to maintain energy efficiency. Based on their specificity, two groups of regulators are distinguished, general and secondary ones (Guédon, Jamet, & Renault, 2002; Mayo et al., 2010). In most low‐GC gram‐positive bacteria, the main general regulator is catabolite control protein A (CcpA) (Hueck & Hillen, 1995), which acts by binding to 14‐nucleotide DNA target sites known as cre (catabolite responsive elements), conducting carbon catabolite activation (CCA) or repression (CCR) (Weickert & Chambliss, 1990). The cre sites are found in promoter regions of the CCR‐ and CCA‐sensitive genes and the binding by CcpA to them is strongly stimulated by Ser46‐phosphorylated HPr protein (Deutscher, 2008). In Lactococcus lactis strains, CcpA has been shown to repress transcription of different genes associated with the uptake of β‐glucosides, fructose, galactose, and lactose and to activate the glycolytic operon las (Aleksandrzak‐Piekarczyk, Polak, Jezierska, Renault, & Bardowski, 2011; Aleksandrzak‐Piekarczyk et al., 2005; Barrière et al., 2005; Luesink, van Herpen, Grossiord, Kuipers, & de Vos, 1998; Monedero, Kuipers, Jamet, & Deutscher, 2001). Sugar catabolism can also be mastered by specific secondary regulators, common in LAB and acting locally, falling to diverse protein families such as LacI, LysR, AraC, GntR, DeoR, RpiR, or BglG. In lactococci, regulators belonging to some of these families have been shown to positively or negatively control genes directing utilization of sugars such as α‐galactosides, β‐glucosides, fructose, lactose, maltose, sucrose, and xylose (Aleksandrzak‐Piekarczyk, Stasiak‐Różańska, Cieśla, & Bardowski, 2015; Andersson & Rådström, 2002; Bardowski, Ehrlich, & Chopin, 1994; Barrière et al., 2005; Boucher, Vadeboncoeur, & Moineau, 2003; Erlandson et al., 2000; Rauch & de Vos, 1992; Van Rooijen & de Vos, 1990).
The mechanisms of transcriptional regulation of the Leloir pathway genes have been elucidated in some LAB species. Gal–lac operons are frequently regulated by specific transcription regulators, which belong to the LacI type. In Streptococcus (S.) thermophilus and S. mutans, GalR acts as a transcription activator and repressor of the lac and gal operons, respectively (Ajdić & Ferretti, 1998; Vaughan et al., 2001). In both species, the GalR‐encoding galR gene is oriented divergently from the structural genes of the Leloir operon. In Lactobacillus casei, a potential transcription regulatory gene, galR, has been identified in the gal operon and is transcribed in the same direction (Bettenbrock & Alpert, 1998). In Lactobacillus helveticus, the inducible genes lacLM (encoding β‐galactosidase) of the unusually organized gal and lac gene cluster are regulated at the transcriptional level by LacR repressor (Fortina, Ricci, Mora, Guglielmetti, & Manachini, 2003). No specific regulatory genes have been identified for the Leloir operon in L. lactis to date, albeit it has been demonstrated that expression of gal genes is under CcpA‐dependent catabolic repression (Luesink et al., 1998; Zomer, Buist, Larsen, Kok, & Kuipers, 2007).
We propose that YugA activates expression of lacS and the lac–gal genes localized downstream by binding to the lacS upstream DNA region containing a putative promoter. Because of this newly identified regulatory function of YugA, we propose to re‐name it GlaR (galactose–lactose operon Regulatory protein). To the best of our knowledge, this is the first report exploring a specific GlaR‐dependent regulatory mechanism of the Leloir pathway genes in L. lactis IL1403 at the molecular level. We examined the effects of glaR deletion and found that the lack of GlaR precludes the strain's growth in galactose‐containing media and abolishes lacS gene expression. These results shed new light on the regulation of carbohydrate catabolism in this biotechnologically important bacterium and reveal a new regulatory protein. Notably, the described mechanism of control of galactose and lactose catabolism by enzymes of the Leloir utilization pathway is unique among LAB.
2. MATERIALS AND METHODS
2.1. Bacterial strains, media, and plasmids
Bacterial strains and plasmids used in this study are shown in Table 1. Escherichia coli cells were cultivated in Luria–Bertani (LB) medium (Wood, 1983) at 37°C, and L. lactis was grown in M17 medium (Terzaghi & Sandine, 1975) or in CDM (Sissler et al., 1999). M17 and CMD were supplemented with 1% glucose (G‐M17 or G‐CDM), or 1% cellobiose (C‐M17 or C‐CDM), or 1% galactose (Gal‐M17 or Gal‐CDM), or 1% galactose with 1% cellobiose (GalC‐M17 or GalC‐CDM). When necessary, ampicillin (Amp; 100 μg ml−1 for E. coli) or erythromycin (Em; 100 μg ml−1 for E. coli and 5 μg ml−1 for L. lactis) was added to the medium. Solidified media contained 1.5% agar and, when required for E. coli, 1 mM IPTG (isopropyl β‐D‐thiogalactopyranoside) and 50 μg ml−1 for X‐gal (5‐bromo‐4‐chloro‐3‐indolyl‐β‐D‐galactopyranoside).
Table 1.
Bacterial strains, plasmids, and primers
| Strain, plasmid, or primer pair | Relevant genotypic or phenotypic propertiesa | Source and/or referenceb |
|---|---|---|
| Strains | ||
| L. lactis | ||
| IL1403 | Gal+, plasmid‐free wild‐type, host strain | INRA (Chopin et al.1984) |
| LL302 | L. lactis MG1363 derivative, RepA+ | (Leenhouts et al.1998) |
| IL1403ΔglaR | Gal‐, ΔglaR, Ems , plasmid‐free, IL1403 derivative | This study |
| IL1403ccpA ‐ | Lac+ , CcpA‐ (ISS1), Ems, plasmid‐free, IL1403 derivative | (Aleksandrzak‐Piekarczyk et al., 2005) |
| IL1403ΔglaR‐pGhost9glaR | Gal+, Emr, IL1403ΔglaR derivative carrying pGhost9glaR | This study |
| E. coli | ||
| TG1 | Δ(hsdMS‐mcrB)5 Δ(lac‐proAB) supE thi‐1 F’(traD36 proAB + lacI q ZΔM15) | (Gibson1984) |
| EC1000 | Kmr, RepA+ MC1000 | (Leenhouts et al.1996) |
| BL21 | B F‐ ompT dcm lon hsdSB(rB‐ mB‐) gal [malB+]K‐12(λS) | (Miroux and Walker1996) |
| Plasmids | ||
| pGEM‐T | Ampr, M13ori, linear T‐overhang vector | Promega |
| pGhost9 | Emr, repA (Ts) | INRA (Maguin et al.1996) |
| pIL253 | Emr, high‐copy number lactococcal vector | (Simon and Chopin1988) |
| ptXB1 | Ampr, M13ori, rop, lacI, Mxe GyrA intein | New England Biolabs |
| Recombinant plasmids | ||
| pGhost9ΔglaR | Emr, pGhost9 carrying glaR upstream, and downstream regions | This study |
| pGhost9glaR | Emr , pGhost9 carrying glaR under the control of its promoter | This study |
| Primersc | ||
| For deletion and complementation of the glaR gene | ||
| glaRUPf/glaRUPr | CCATCGATTCAAGTTCCCAAACGCTCC/GGAGAATTCGCCAAGTATAGGATTCAGC | |
| glaRDOWNf/glaRDOWNr | GGAGAATTCCAAGAGTAGTCTTGAGGTG/AAGATGACATAATCCCACCAACAAC | |
| glaRfor/glaRrev | GCTAAGACCGCAGCTTC/GACCAGAAGGCAATGTC | |
| ptXB1for/glaRBamHrev | GTGAGCGGATAACAATTCC/GGATCCTTATTGTTTTAAAGTATAAATGG | |
| For qPCR amplifications | ||
| LlGlaRaF/LlGlaRaR | TGCAACTTTTCCGTAAGCCC/TTGGGATTTTGTCCTTTGGC | |
| LlLacSaF/LlLacSaR | CTGGAACACCACATGAGGATGC/AAGATGACATAATCCCACCAACAAC | |
| LlGalMaF/LlGalMaR | TGACCATCCTTTCTTGTTAGACCAG/CCATGGTGCACTTGCTTTTTC | |
| LlGalKaF/LlGalKaR | AACAAGCCGGTGTCTTGGG/TCCAACTTTGTTGAACCAGAACC | |
| LlGalTaF/LlGalTaR | AAAAAGACCCCAAAGCCATTG/ATTGGAAGCCCCAGTCTTCG | |
| LlThgAaF/LlThgAaR | CCAAATGTTACGATTGACACGG/AGACTCCCTGCGCCAATCAC | |
| LlLacZaF/LlLacZaR | GAAAGCACTTCTTGTTCGTGGAG/TCACACAATTCATACCAGCGTG | |
| LlGalEaF/LlGalEaR | GCCTGATGGAACTTGTATTCGTG/CCTGTTACTTTTCGTGCGGTTTC | |
| LlYufCaF/LlYufCaR | TTGCAGGAGAAACTTTGACGG/TCTGCCCACGGAATAGCAC | |
| LlPurMaF/LlPurMaR | ATTGCGTAGCCATGTGCGTC/CTGTTTCTCCACCAATCAGCG | |
| LlTufaF/LlTufaR | CGTGACCTCTTGAGCGAATACG/GAGTGGTTTGTCAGTGTCGCG | |
| For amplification of nucleotides for EMSA | ||
| glaRfor/glaRrev | GCCAGAGTCCTAATGAAAG/CATGGCTTACTATGCCC | |
| lacSfor/lacSrev | CTAATTGATGCTTACTCC/CTTTCATGGGAATCCTCC | |
| galMfor/galMrev | GCCTATCCTGGTGCAAC/CCATGATATTTCCTAACT | |
| galTfor/galTrev | GTTGTCGGTTATCCAGC/CAAGTGGCTCAATCGTTCC | |
| thgAfor/thgArev | CAGGAAGCAGTTGGAGAAG/CAGCCAGAGCAACAAATGG | |
| galEfor/galErev | GGACATTGGCATCTACTTG/CTGCCACATCGTAACCACG | |
| yufAfor/yufArev | CTTGAAGTGCTTGAAACC/CCATTACATTTTCATGACG | |
aAmp, ampillicin; Em, erythromycin; Km, kanamycin; r, resistance; s, sensitivityl; CcpA, catabolite control protein AbINRA, Institut National de la Recherche Agronomique (Jouy‐en‐Josas, France).cAll primers were designed on the basis of the L. lactis IL1403 genome nucleotide sequence, NCBI with accession no. AE005176 (http://www.ncbi.nlm.nih.gov/genome). To certain primers, restriction sites were added for digestion with EcoRI or ClaI.
2.2. Construction of glaR deletion mutant and complementing plasmid
Lactococcus lactis IL1403 glaR deletion strain (L. lactis IL1403ΔglaR) was generated by double crossover between pGhost9 carrying DNA fragments flanking the glaR gene and the corresponding chromosomal region. The glaR upstream and downstream DNA fragments were amplified with, respectively, the glaRUPf/glaRUPr and glaRDOWNf/glaRDOWNr primer pairs (Table 1). The obtained DNA fragments were cloned in the proper orientation in the integrative vector pGhost9, producing pGhost9ΔglaR. This deletion plasmid was transported into L. lactis IL1403 and homologous recombination was enforced by 10−3 dilution of an overnight culture and incubation at nonpermissive temperature (38°C). Cells harboring pGhost9ΔglaR in the chromosome were cultivated at 38°C on G‐M17Em. Removal from the chromosome and elimination of pGhost9 from L. lactis were performed by growing the integrants in G‐M17 without antibiotic for at least 100 generations at the permissive temperature (28°C). The genomic organization of the resulting glaR deletion strain (L. lactis IL1403ΔglaR) was confirmed by determining its sensitivity to Em and by sequencing of the mutated region.
To complement the glaR deletion, the glaR gene containing its putative promoter region was amplified using glaRUPf and glaRDOWNr primers (Table 1) and ExTaq polymerase giving the glaR(A) insert. The insert was introduced by TA cloning into pGhost9, as described by Radziwill‐Bienkowska et al. (2016). Shortly, pGhost9 was blunt‐linearized with EcoRV and, to add 3′ thymidine overhangs, treated with terminal deoxynucleotidyl transferase (TdT; Thermo Fisher Scientific, USA) and 2′,3′‐dideoxythymidine‐5′‐triphosphate (ddTTP; Affymetrix, USA). The obtained pGhost9(T) was ligated with the glaR(A) insert and cloned in E. coli EC1000. The pGhost9glaR plasmid was isolated, verified by sequencing of the glaR insert, and transformed into L. lactis IL1403ΔglaR to give L. lactis IL1403ΔglaR‐pGhost9glaR.
2.3. Quantification of gene expression by reverse transcription‐quantitative PCR (RT‐qPCR)
RNA was isolated following manufacturer's instructions with the use of GeneMATRIX Universal RNA Purification Kit (EURx, Poland) from 10 ml of L. lactis IL1403 and L. lactis IL1403ΔglaR cultures grown in G‐M17, C‐M17, Gal‐M17 or GalC‐M17 and collected from midexponential phase (OD600 = 0.6). RNA was isolated from at least three independent cultures.
First‐strand cDNA was obtained from DNAse I (Sigma‐Aldrich, USA)‐treated RNA with random primers by the use of the RevertAid(TM) First‐Strand cDNA Synthesis Kit (Thermo Fisher Scientific) according to manufacturer's instructions. qPCR assays on the cDNA were carried out in a 7500 Real‐Time PCR System (Applied Biosystems, USA) and following the previously described methodology (Aleksandrzak‐Piekarczyk et al., 2015). Specific primers for genes (Table 1) were created with Primer Express software (Applied Biosystems). The results were normalized to the L. lactis IL1403 reference genes tuf and purM coding for elongation factor TU and phosphoribosylaminoimidazole synthetase, respectively.
2.4. Growth testing for carbon source utilization
Growth tests were performed using a Microbiology Reader Analyser, Bioscreen C (Oy Growth Curves Ab Ltd, Finland) in 200 μl of CDM with the required sugars (glucose, galactose or cellobiose). OD600 of the bacterial cultures was recorded every 60 mins of growth up to 40 hr at 30°C. The assays were carried out in triplicate.
2.5. Overproduction and purification of GlaR
The self‐cleavable IMPACTTM affinity tag system (New England Biolabs, USA) was used to purify the GlaR protein. E. coli BL21 competent cells were transformed with the pTXB1 plasmid carrying the glaR gene. The obtained transformants were verified by colony PCR, with specific primers ptXB1for and glaRBamHrev. LB medium (600 ml) containing 100 μg/ml ampicillin was inoculated with a freshly grown colony and incubated at 37°C with shaking until an OD660 of 0.5 was reached. After induction of the glaR gene expression using 0.3 mM IPTG, the culture was incubated overnight at 18°C. Then, the cells were pelleted by centrifugation (3000xg, 10 min, 4°C) and stored at −20°C until use. All subsequent purification steps were carried out at 4°C. The frozen cells were resuspended in 10 ml of column buffer A (25 mM Tris‐HCl, pH 8.0; 500 mM NaCl; 10% glycerol) and were disrupted by High Pressure Homogenizer Emulsiflex (Avestin Inc., Canada). After centrifugation (15,000×g, 30 min, 4°C), 5 ml of the clear supernatant was loaded (at 0.5–1 ml/min) onto a polypropylene column (Qiagen, Germany) with 2 ml of chitin beads (New England Biolabs) previously equilibrated with 20 ml of chitin column buffer A. Next, the resin was washed with 20 ml of the same buffer and then with 3 ml of the cleavage buffer B (25 mM Tris‐HCl, pH 8.0; 100 mM KCl; 50 mM DTT; 1 mM MgCl2; 10% glycerol). The GlaR protein was released from the chitin beads after 16 hrs of incubation at 23°C. The eluted GlaR protein, with no extra residues, was concentrated using an Amicon filter device (Millipore, USA) and analyzed on 15% SDS‐polyacrylamide gel with Coomassie staining.
The protein band was cut out from the gel, reduced with 100 mM DTT (30 min, 56°C), alkylated at darkroom with 0.5 M iodoacetamide (45 min, RT), and digested overnight with trypsin (37°C 10 ng μl−1; Promega, USA). Peptide mixture was concentrated, desalted on a RP‐C18 precolumn (Waters, USA), and separated on a nano‐Ultra Performance Liquid Chromatography (UPLC) RP‐C18 column (BEH130 C18; Waters), using a 160‐min gradient from 5% to 30% of acetonitrile. Measurements were taken with the Orbitrap Velos spectrophotometer (Thermo Fisher Scientific), working in the regime of data‐dependent MS to MS/MS switch with HCD type peptide fragmentation. Identification of proteins was performed using the Mascot search engine with the probability‐based algorithm. Data were searched with automatic decoy database and filtered to obtain a false discovery rate below 1%.
Protein concentration was determined using Bradford assay on a NanoDrop spectrophotometer (Thermo Scientific).
2.6. Electrophoretic mobility‐shift assay (EMSA)
EMSA was performed using 1 nM of double‐stranded DNA fragments (~300 bp) generated by PCR with specific primer pairs (Table 1) multiplying the upstream DNA regions of selected genes (glaR, lacS, galM, galT, thgA, and galE). The PCR product of the yufA upstream DNA region served as a negative control. The DNA fragments were incubated with increasing quantities of the GlaR protein (0; 1; 2; 2.5; 3; 3.5; 4; 4.5; 5, and 6 μM) in 10× binding buffer (10 mM Tris‐HCl, pH 8.5; 10 mM MgCl2; 100 mM KCl; 0.1 mg/ml BSA), supplemented with 250 mM galactose in a total volume of 20 μl. After 20 mins of incubation at 37°C, the samples were separated on 5% polyacrylamide gel in 0.5× Tris‐borate‐EDTA buffer (TBE). The amount of GlaR protein giving best results was evaluated as 4 μM. The final EMSA for all upstream DNA sequences of selected genes was performed in previously described conditions, using the optimized amount of the protein (Figure 4).
Figure 4.
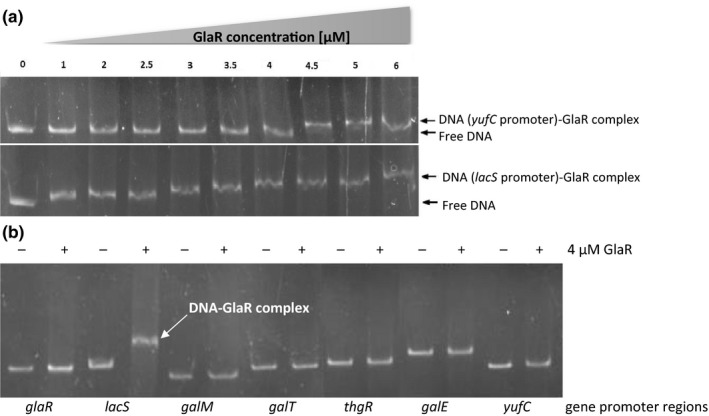
GlaR binding to potential promoter regions of gal–lac operon genes and glaR. The test was performed by electrophoretic mobility‐shift assays (EMSA) of (A) the GlaR protein gradient and lacS or yufC (negative control) ca. 200 nt putative promoter regions (B) and GlaR at concentration of 4 μM and ca. 200 nt putative promoter regions of selected genes of the Leloir operon plus yufC. “Free DNA” indicates DNA without bound GlaR; “DNA‐complex” indicates DNA with bound GlaR
3. RESULTS
3.1. Structural characterization of DNA region following the glaR gene
Figure 1 illustrates the chromosomal region of the glaR gene (formerly denoted yugA) in L. lactis IL1403. The product of glaR, the GlaR protein, is highly similar with other transcription regulators of the RpiR family as it is a two‐domain protein and comprises a 59‐residue N‐terminal DNA‐binding helix‐turn‐helix (HTH) domain and a 99‐residue sugar isomerase (SIS) motif at its C‐terminus (http://pfam.sanger.ac.uk/). The YugA amino acid sequence is 100% identical with its orthologues encoded in L. lactis subsp. lactis genomes, 98% with L. lactis subsp. cremoris and only 70% with Lactococcus garvieae. The glaR gene is preceded by a putative promoter, of which the ‐10 region is in full agreement with the promoter consensus sequence, defined as TATAAT (Browning & Busby, 2004), and a potential CcpA‐binding cre site with two mismatches with the cre consensus, in L. lactis defined as WGWAARCGYTWWMA (Zomer et al., 2007) (Figure 1). The glaR gene is followed by a potential rho‐independent terminator (Figure 1) with a free energy value (ΔG) of −15 kcal/mol. The presence of these transcriptional signals indicates that glaR may form a single‐gene operon regulated by the CcpA protein.
Figure 1.
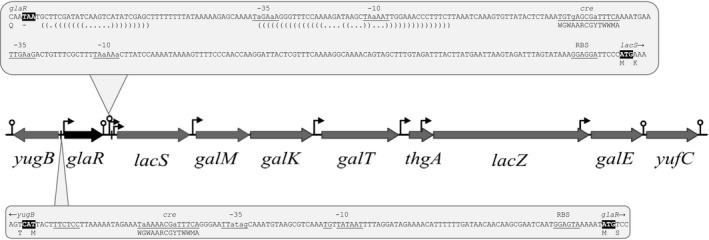
Organization of the gal–lac operon with surrounding genes in L. lactis IL1403. Lollipops and brackets indicate potential transcription terminators. Bent arrows and underlined nucleotides indicate potential promoter regions. White‐on‐black font marks START and STOP codons. RBS indicates ribosome‐binding site. Vertical lines and underlined cre indicate catabolite responsive element with cre consensus sequence (WGWAARCGYTWWMA). The picture presents the correct sequence of lacS, which at the NCBI database is annotated as a pseudogene due to the deletion of an adenine 446. We confirmed correctness of lacS by its sequencing
Located downstream of glaR and transcribed in the same direction are the genes of Leloir pathway cluster, which in L. lactis IL1403 in addition to the four galactose genes contains a sugar permease gene (lacS) and predicted genes for lactose assimilation such as lacZ (β‐galactosidase) and thgA (thiogalactoside acetyltransferase). An in silico analysis identified several putative promoters preceding the lacS, galMKT, thgA, lacZ, and galE genes (Figure 1) suggesting multiple transcription start sites within the operon. Two of the identified potential −10 regions, those upstream of lacS and galE, are in full agreement with the promoter consensus sequence, whereas none of the promoters found contains a sequence identical to the −35 consensus TTGACA (Browning & Busby, 2004). Downstream of the galE gene, a potential rho‐independent terminator with a ΔG value of −11.4 kcal/mol, was identified.
3.2. The genetic organization of glaR followed by the Leloir operon is highly conserved among lactococci
Among the 39 fully sequenced Lactococcus spp. genomes deposited in the GenBank database (as of January, 2018), glaR homologues were identified in 36 strains of L. lactis and L. garvieae, but were absent from Lactococcus piscium and Lactococcus raffinolactis. In all the cases, glaR lies directly upstream of the Leloir operon and is transcribed in the same direction. Further comparative analyses revealed that this genetic layout is specific for lactococci only, as in other species of the order Lactobacillales, in some of which more distant glaR homologues are present (over 30% amino acid sequence identity), this gene is never adjacent to the Leloir cluster.
3.3. GlaR is crucial for L. lactis IL1403 growth on galactose
To assess the possible role of GlaR, a L. lactis IL1403ΔglaR mutant strain was constructed lacking the glaR gene and its growth was tested in CDM with different sugars and compared with its parental wild‐type IL1403 strain. No significant differences were found between the growth of these two strains in G‐CDM or C‐CDM, but in a galactose‐supplemented medium, the mutant lacking GlaR was unable to grow completely (Figure 2).
Figure 2.
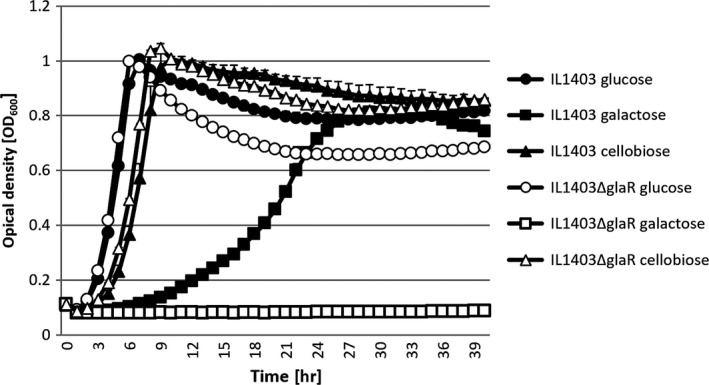
Kinetics of L. lactis IL1403 wild‐type and ΔglaR strains in CDM supplemented with different sugars
Transformation of pGhost9glaR into IL1403ΔglaR that led to the creation of the L. lactis IL1403ΔglaRpGhost9glaR strain, fully reversed the effect of the glaR deletion, restoring the mutant's growth in medium supplemented with galactose.
3.4. GlaR is a transcriptional activator of the gal–lac genes
To define the influence of GlaR on the expression of the gal–lac operon genes in response to various sugars, using RT‐qPCR, we compared mRNA levels of individual genes in L. lactis IL1403 wild‐type and IL1403ΔglaR grown in C‐M17, G‐M17, Gal‐M17 or GalC‐M17. Cellobiose in GalC‐M17 allowed L. lactis IL1403ΔglaR to grow in the presence of galactose, as this mutant is incapable to use galactose as a carbon source. In the presence of galactose, the expression of most of the Leloir operon genes was significantly lower in L. lactis IL1403ΔglaR, whereas in cellobiose‐ or glucose‐supplemented media, they were expressed at a similar level in both the strains (Figure 3a). The most pronounced difference between the strains concerned lacS, whose mRNA was virtually undetectable in L. lactis IL1403ΔglaR grown in the presence of any sugar tested, but was abundant in L. lactis IL1403 grown in galactose‐containing media (Gal‐M17 and GalC‐M17) (Figure 3a). Notably, lacS was not expressed in wild‐type L. lactis IL1403 grown without galactose. Some GlaR‐dependent activation in the presence of galactose was also observed for other gal–lac operon genes. The GlaR activation coefficient (calculated as the ratio of gene expression level in IL1403 to that in IL1403ΔglaR) for those genes varied between 2.6 (galE) and 8 (galM) (Figure 3b). For the negative control yufC, the GlaR activation coefficient was close to 1, indicating—as expected—a lack of GlaR‐dependent activation (Figure 3b).
Figure 3.
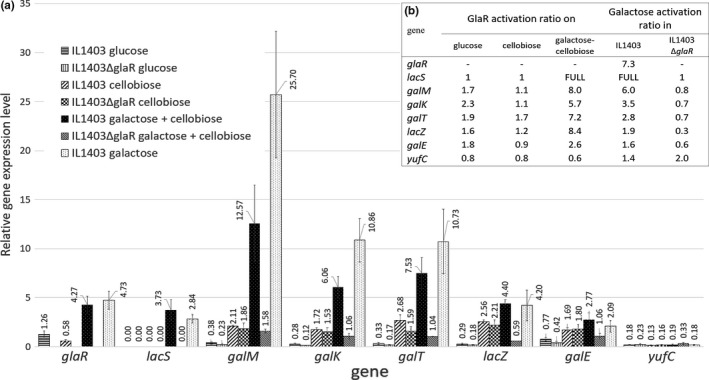
The relative gene expression levels in L. lactis IL1403 wild‐type and IL1403ΔglaR. (A) mRNA levels determined by RT‐qPCR in relation to tuf and purM. (B) GlaR and galactose activation ratios calculated, respectively, as a quotient of relative gene expression in IL1403 and IL1403ΔglaR, and as a quotient of relative gene expression in strains grown in GalC‐M17 and C‐M17. “FULL” indicates a high induction from a gene expression from the non‐detectable level
The lowest transcript levels of the genes studied were detected in L. lactis IL1403 wild‐type and IL1403ΔglaR growing under repressive conditions (G‐M17), most likely due to the downregulation of gal–lac genes by CcpA, as described previously (Luesink et al., 1998). Expression of most of the Leloir genes increased in both strains in the medium supplemented with cellobiose (Figure 3a) indicating a release from catabolic repression. Notably, lacS mRNA was not detected in either of these media in either of the strains. In comparison with cellobiose, higher transcript levels of the gal–lac operon genes were detected when the wild‐type strain was grown in media supplemented with galactose (Gal‐M17 or GalC‐M17) (Figure 3a). The activation by galactose calculated as the ratio of expression in GalC‐M17 and in C‐M17 was the highest for the lacS gene, and for the other gal–lac genes, it varied from 1.6 (galE) to 6 (galM) (Figure 3b). In the glaR mutant downstream of lacS, these ratios were ca. 1 indicating that in the absence of GlaR, the galactose‐dependent activation of the gal–lac genes does not occur. Also for the negative control gene yufC, its expression levels with and without galactose were similar in both the wild‐type strain and in the glaR mutant further confirming that it is not subject to galactose induction (Figure 3B).
3.5. GlaR activates expression of the Leloir operon by binding to the lacS promoter region
To identify the genomic region to which the GlaR protein binds specifically, an in vitro EMSA test was performed with selected upstream regions containing potential promoters of the Leloir operon genes (lacS, galM, galT, thgA, and galE) and of glaR and purified GlaR protein. An unrelated dsDNA containing the yufA upstream region was used as a control to test for nonspecific binding. No nonspecific interactions were detected at GlaR concentrations up to 4 μM; therefore, this concentration was used to investigate specific binding (Figure 4a). At this concentration, GlaR bound to the putative lacS promoter but it did not form specific complexes with any other putative promoters tested (Figure 4b). Notably, GlaR bound to the lacS dsDNA also at lower concentrations (1–3.5 μM) (Figure 4a), indicating that the interaction is fairly strong.
3.6. GlaR expression is inducible by galactose but insensitive to CcpA‐mediated catabolite repression
CcpA is a master transcriptional regulator controlling carbohydrate utilization and metabolism genes in gram‐positive bacteria including L. lactis (Hueck & Hillen, 1995; Zomer et al., 2007). As the promoter region of glaR contains a potential cre sequence (Figure 1) that could be recognized by CcpA, we sought to determine the role of CcpA in the transcriptional regulation of this gene. glaR expression in the presence of different sugars (glucose, galactose, or cellobiose) was compared between the wild‐type strain and a ccpA mutant (IL1403ccpA ‐). The lack of the CcpA regulatory protein had no effect on glaR expression in any of the media tested, indicating that glaR is not under CcpA‐dependent catabolite repression. On the other hand, the transcription of glaR in IL1403 was elevated ca. sevenfold in galactose‐containing media compared to its expression in glucose‐ or cellobiose‐supplemented media (Figure 3b), and indicating a possible autoregulation of the glaR gene.
4. DISCUSSION
Because of the substantial biotechnological relevance of galactose, especially in the dairy industry, where unmetabolized galactose is associated with poor product quality (Baskaran & Sivakumar, 2003; Hutkins, Halambeck, & Morris, 1986; Michel & Martley, 2001), galactose metabolism and its regulation have been thoroughly studied in several LAB species. The crucial role of the Leloir pathway in the utilization of nonphosphorylated galactose is well documented. In several species of the genera Lactobacillus and Streptococcus, the Leloir or gal–lac operons are known to be regulated transcriptionally mainly by repressor proteins belonging to the LacI family, but the regulatory mechanism of the Leloir pathway genes in L. lactis, important dairy industry bacterium, remained unexplored to date. In this study, we demonstrate that in L. lactis, the regulation of the gal–lac operon differs from that in other LAB species as it is under a positive control of the RpiR‐family transcriptional regulator GlaR.
The L. lactis IL1403 genome carries eight genes encoding RpiR‐family members: GlaR (previously named YugA), ClaR (previously YebF), GntR, YecA, YfeA, YidA, YljC, and YleF (retrieved from http://www.kegg.jp/kegg/ssdb/). Thus far, only one RpiR‐member, ClaR, has been characterized in L. lactis and shown to function as an activator of cellobiose and lactose metabolism genes bglS and celB (Aleksandrzak‐Piekarczyk et al., 2015). In other species distantly related to L. lactis, members of the RpiR family have been found to function as regulators targeting genes involved in the metabolism of diverse carbon sources. Thus, GlvR is a positive regulator of maltose metabolism in B. subtilis (Yamamoto, Serizawa, Thompson, & Sekiguchi, 2001), HexR, IolR, MurR, and RpiR act as repressors of glucose, inositol, N‐acetylmuramic acid, ribose or central carbon metabolism in several gram‐negative bacteria (Antunes et al., 2016; Jaeger & Mayer, 2008; Kohler, Choong, & Rossbach, 2011; Sørensen & Hove‐Jensen, 1996), and HexR is a dual‐mode pleiotropic regulator of the central carbohydrate metabolism in proteobacteria (Leyn et al., 2011). Thus far, none of the RpiR regulators has been implicated in modulating galactose metabolism.
Members of the RpiR family harbor a DNA‐binding HTH domain and a phospho‐sugar‐binding SIS motif, respectively, at their N‐ and C‐terminal regions (Bateman, 1999; Teplyakov, Obmolova, Badet‐Denisot, Badet, & Polikarpov, 1998). The SIS domain is found in numerous proteins that regulate expression of genes dedicated to the synthesis of phospho‐sugars (Aleksandrzak‐Piekarczyk et al., 2015; Bateman, 1999; Daddaoua, Krell, & Ramos, 2009; Jaeger & Mayer, 2008; Sørensen & Hove‐Jensen, 1996; Teplyakov et al., 1998; Yamamoto et al., 2001) but here, we show that a protein from this family can also be engaged in regulating of an operon involved in the metabolism of a nonphosphorylated sugar galactose. This mode of regulation seems to be restricted to the genus Lactococcus as well‐conserved GlaR homologues occur only in these bacteria and their genes are always localized directly upstream of the gal–lac or Leloir operons.
Using two L. lactis strains differing by the presence of glaR and growing them in media with different sugars as the sole carbon source, we showed that the gal–lac operon genes are maximally expressed only when both galactose and GlaR are available. This effect was absolute for the lacS gene, as its transcript was virtually undetectable in the absence of galactose or GalR. For the other genes located downstream of lacS, the GlaR‐dependent induction by galactose was less spectacular and, notably, its extent decreased with increasing distance from lacS. These results suggested that the genes in question form a single operon with the promoter preceding the lacS gene. Indeed, using EMSA, we found that GlaR does bind specifically to a region upstream of lacS, but not to the putative promoters of the other genes downstream of lacS. Notably, the presence of GlaR‐independent promoter‐like regions upstream of these genes explains why they were expressed at a submaximal level even in the absence of galactose/or GlaR. We additionally confirmed that functional expression of lacS requires the action of GlaR by showing that the L. lactis IL1403ΔglaR was unable to grow in galactose medium. LacS permease is the main transporter used for galactose (but not for lactose; Aleksandrzak‐Piekarczyk et al., 2005) uptake in IL1403 cells, and its inactivation leads to the gal ‐ phenotype (our unpublished data).
Remarkably, also the transcription of glaR was induced substantially in galactose‐containing medium in comparison with cellobiose, which could in part explain the effect of galactose on the GlaR‐dependent expression of the gal–lac operon. It also suggested possible autoregulation of glaR expression by GlaR. Autoregulation is frequent in prokaryotic gene regulation strategies and has been reported for numerous transcription regulators (Gerlach, Valentin‐Hansen, & Bremer, 1990; Meng, Kilstrup, & Nygaard, 1990; Morel, Lamarque, Bissardon, Atlan, & Galinier, 2001; Vaughan et al., 2001; Weickert & Adhya, 1993). However, we could not confirm a direct involvement of GlaR in glaR activation as no GlaR binding to the glaR promoter region was found by EMSA (Figure 4b). A plausible explanation includes an indirect control by GlaR (e.g., via an alternative regulator under the control of GlaR) or the action of another galactose‐dependent but GlaR‐independent mechanism.
Both lacS and glaR are preceded by cre boxes suggesting that their expression is under CcpA‐driven carbon catabolite repression (CCR). Indeed, in the presence of glucose, transcriptional arrest of all the genes under the control of the lacS promoter was detected, whereas cellobiose or galactose caused a relief from CCR. This phenomenon has already been studied in another L. lactis strain, MG1363 (Luesink et al., 1998), in which the Leloir operon differs from the one of IL1403 but is also subject to CcpA‐driven catabolic repression. In contrast, we found that that CcpA is not engaged in the regulation of glaR expression in L. lactis IL1403. One reason for this could be the two‐nucleotide deviation of the cre sequence upstream of glaR (TaAAAACGaTTTCA) form the cre consensus WGWAARCGYTWWMA (Zomer et al., 2007). The two adenine mismatches may prevent or impair CcpA interaction with its operator and thus allow of the glaR transcription also in repressive conditions (glucose).
In summary, here, we have documented unusual mechanism of gal–lac operon activation in L. lactis IL1403 and, by similarity, probably also in other Lactococcus spp. No similar mechanism has been reported in other LAB species. This regulation relies on galactose‐inducible and GlaR‐dependent transcriptional activation of the lacS promoter inducing the lacS gene itself and the other lac and Leloir pathway genes located downstream.
ETHICAL STATEMENT
This article does not contain any studies with human or animals performed by any of the authors.
CONFLICT OF INTEREST
The authors declare that they have no conflict of interest.
ACKNOWLEDGMENTS
RT‐qPCR assays amplification were performed at the Genetic Modifications Analysis Lab, IBB PAS. Proteomic analyses were performed at the Laboratory of Mass Spectrometry, IBB PAS. The equipment used for proteomic analysis was funded in part by the Centre for Preclinical Research and Technology (CePT), a project cosponsored by the European Regional Development Fund and Innovative Economy, The National Cohesion Strategy of Poland.
Aleksandrzak‐Piekarczyk T, Szatraj K, Kosiorek K. GlaR (YugA)—a novel RpiR‐family transcription activator of the Leloir pathway of galactose utilization in Lactococcus lactis IL1403. MicrobiologyOpen. 2019;8:e714 10.1002/mbo3.714
DATA ACCESSIBILITY
The authors declare that all data generated or analyzed during this study are included in this article.
REFERENCES
- Ajdić, D. , & Ferretti, J. J. (1998). Transcriptional regulation of the Streptococcus mutans gal operon by the GalR repressor. Journal of Bacteriology, 180, 5727–5732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aleksandrzak‐Piekarczyk, T (2013). Lactose and β‐glucosides metabolism and its regulation in Lactococcus lactis: A review In Kongo JM. (ed) Lactic Acid Bacteria ‐ R & D for Food, Health and Livestock Purposes. IntechOpen, UNITED KINGDOM. 10.5772/50889 [DOI] [Google Scholar]
- Aleksandrzak‐Piekarczyk, T. , Kok, J. , Renault, P. , & Bardowski, J. (2005). Alternative lactose catabolic pathway in Lactococcus lactis IL1403. Applied and Environment Microbiology, 71, 6060–6069. 10.1128/AEM.71.10.6060-6069.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aleksandrzak‐Piekarczyk, T. , Polak, J. , Jezierska, B. , Renault, P. , & Bardowski, J. (2011). Genetic characterization of the CcpA‐dependent, cellobiose‐specific PTS system comprising CelB, PtcB and PtcA that transports lactose in Lactococcus lactis IL1403. International Journal of Food Microbiology, 145, 186–194. 10.1016/j.ijfoodmicro.2010.12.011 [DOI] [PubMed] [Google Scholar]
- Aleksandrzak‐Piekarczyk, T. , Stasiak‐Różańska, L. , Cieśla, J. , & Bardowski, J. (2015). ClaR—a novel key regulator of cellobiose and lactose metabolism in Lactococcus lactis IL1403. Applied Microbiology and Biotechnology, 99, 337–347. 10.1007/s00253-014-6067-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andersson, U. , & Rådström, P. (2002). Physiological function of the maltose operon regulator, MalR, in Lactococcus lactis . BMC Microbiology, 2, 28 10.1186/1471-2180-2-28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Antunes, A. , Golfieri, G. , Ferlicca, F. , Giuliani, M. M. , Scarlato, V. , & Delany, I. (2016). HexR controls glucose‐responsive genes and central carbon metabolism in Neisseria meningitidis . Journal of Bacteriology, 198, 644–654. 10.1128/JB.00659-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bardowski, J. , Ehrlich, S. D. , & Chopin, A. (1994). BglR protein, which belongs to the BglG family of transcriptional antiterminators, is involved in beta‐glucoside utilization in Lactococcus lactis . Journal of Bacteriology, 176, 5681–5685. 10.1128/jb.176.18.5681-5685.1994 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barrière, C. , Veiga‐da‐Cunha, M. , Pons, N. , Guedon, E. , van Hijum, S. A. F. T. , Kok, J. , … Renault, P. (2005). Fructose utilization in Lactococcus lactis as a model for low‐GC Gram‐positive bacteria: Its regulator, signal, and DNA‐binding site. Journal of Bacteriology, 187, 3752–3761. 10.1128/JB.187.11.3752-3761.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baskaran, D. , & Sivakumar, S. (2003). Galactose concentration in pizza cheese prepared by three different culture techniques. International Journal of Dairy Technology, 56, 229–232. 10.1046/j.1471-0307.2003.00109.x [DOI] [Google Scholar]
- Bateman, A. (1999). The SIS domain: A phosphosugar‐binding domain. Trends in Biochemical Sciences, 24, 94–95. 10.1016/S0968-0004(99)01357-2 [DOI] [PubMed] [Google Scholar]
- Bettenbrock, K. , & Alpert, C.‐A. (1998). The gal genes for the Leloir pathway of Lactobacillus casei 64H. Applied and Environment Microbiology, 64, 2013–2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boucher, I. , Vadeboncoeur, C. , & Moineau, S. (2003). Characterization of genes involved in the metabolism of α‐galactosides by Lactococcus raffinolactis . Applied and Environment Microbiology, 69, 4049–4056. 10.1128/AEM.69.7.4049-4056.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouffard, G. G. , Rudd, K. E. , & Adhya, S. L. (1994). Dependence of lactose metabolism upon mutarotase encoded in the gal operon in Escherichia coli . Journal of Molecular Biology, 244, 269–278. 10.1006/jmbi.1994.1728 [DOI] [PubMed] [Google Scholar]
- Browning, D. F. , & Busby, S. J. W. (2004). The regulation of bacterial transcription initiation. Nature Reviews Microbiology, 2, 57–65. 10.1038/nrmicro787 [DOI] [PubMed] [Google Scholar]
- Chopin, A. , Chopin, M. C. , Moillo‐Batt, A. , & Langella, P. (1984). Two plasmid determined restriction and modification systems in Streptococcus lactis . Plasmid, 11, 260–263. 10.1016/0147-619X(84)90033-7 [DOI] [PubMed] [Google Scholar]
- Daddaoua, A. , Krell, T. , & Ramos, J.‐L. (2009). Regulation of glucose metabolism in Pseudomonas: The phosphorylative branch and Entner‐Doudoroff enzymes are regulated by a repressor containing a sugar isomerase domain. Journal of Biological Chemistry, 284, 21360–21368. 10.1074/jbc.M109.014555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Vos, W. M. (1996). Metabolic engineering of sugar catabolism in lactic acid bacteria. Antonie van Leeuwenhoek, 70, 223–242. 10.1007/BF00395934 [DOI] [PubMed] [Google Scholar]
- Deutscher, J. (2008). The mechanisms of carbon catabolite repression in bacteria. Current Opinion in Microbiology, 11, 87–93. 10.1016/j.mib.2008.02.007 [DOI] [PubMed] [Google Scholar]
- Erlandson, K. A. , Park, J.‐H. , Wissam, El Khal , Kao, H.‐H. , Basaran, P. , Brydges, S. , & Batt, C. A. (2000). Dissolution of xylose metabolism in Lactococcus lactis . Applied and Environment Microbiology, 66, 3974–3980. 10.1128/AEM.66.9.3974-3980.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fortina, M. G. , Ricci, G. , Mora, D. , Guglielmetti, S. , & Manachini, P. L. (2003). Unusual organization for lactose and galactose gene clusters in Lactobacillus helveticus . Applied and Environment Microbiology, 69, 3238–3243. 10.1128/AEM.69.6.3238-3243.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frey, P. A. (1996). The Leloir pathway: A mechanistic imperative for three enzymes to change the stereochemical configuration of a single carbon in galactose. FASEB Journal, 10, 461–470. 10.1096/fasebj.10.4.8647345 [DOI] [PubMed] [Google Scholar]
- Gerlach, P. , Valentin‐Hansen, P. , & Bremer, E. (1990). Transcriptional regulation of the cytR repressor gene of Escherichia coli: Autoregulation and positive control by the cAMP/CAP complex. Molecular Microbiology, 4, 479–488. 10.1111/j.1365-2958.1990.tb00614.x [DOI] [PubMed] [Google Scholar]
- Gibson, T. J . (1984). Studies on the Epstein‐Barr virus genome [Ph.D. thesis]. Cambridge, United Kingdom: Cambridge University
- Grossiord, B. , Vaughan, E. E. , Luesink, E. , & de Vos, W. M. (1998). Genetics of galactose utilisation via the Leloir pathway in lactic acid bacteria. Le Lait, 78, 77–84. 10.1051/lait:1998110 [DOI] [Google Scholar]
- Guédon, E. , Jamet, E. , & Renault, P. (2002). Gene regulation in Lactococcus lactis: The gap between predicted and characterized regulators. Antonie van Leeuwenhoek, 82, 93–112. 10.1023/A:1020680926267 [DOI] [PubMed] [Google Scholar]
- Hueck, C. J. , & Hillen, W. (1995). Catabolite repression in Bacillus subtilis: A global regulatory mechanism for the gram‐positive bacteria? Molecular Microbiology, 15, 395–401. 10.1111/j.1365-2958.1995.tb02252.x [DOI] [PubMed] [Google Scholar]
- Hutkins, R. , Halambeck, S. M. , & Morris, H. A. (1986). Use of galactose‐fermenting Streptococcus thermophilus in the manufacture of Swiss, Mozzarella, and short‐method Cheddar cheese. Journal of Dairy Science, 69, 1–8. 10.3168/jds.S0022-0302(86)80361-7 3700799 [DOI] [Google Scholar]
- Jaeger, T. , & Mayer, C. (2008). The transcriptional factors MurR and catabolite activator protein regulate N‐acetylmuramic acid catabolism in Escherichia coli . Journal of Bacteriology, 190, 6598–6608. 10.1128/JB.00642-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kohler, P. R. A. , Choong, E.‐L. , & Rossbach, S. (2011). The RpiR‐like repressor IolR regulates inositol catabolism in Sinorhizobium meliloti . Journal of Bacteriology, 193, 5155–5163. 10.1128/JB.05371-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leenhouts, K. , Bolhuis, A. , Venema, G. , & Kok, J. (1998). Construction of a food‐grade multiple‐copy integration system for Lactococcus lactis . Applied Microbiology and Biotechnology, 49, 417–423. 10.1007/s002530051192 [DOI] [PubMed] [Google Scholar]
- Leenhouts, K. , Buist, G. , Bolhuis, A. , ten Berge, A. , Kiel, J. , Mierau, I. , … Kok, J. (1996). A general system for generating unlabelled gene replacements in bacterial chromosomes. Molecular and General Genetics, 253, 217–224. 10.1007/s004380050315 [DOI] [PubMed] [Google Scholar]
- Leyn, S. A. , Li, X. , Zheng, Q. , Novichkov, P. S. , Reed, S. , Romine, M. F. , … Rodionov, D. A. (2011). Control of proteobacterial central carbon metabolism by the HexR transcriptional regulator: A case study in Hewanella oneidensis . Journal of Biological Chemistry, 286, 35782–35794. 10.1074/jbc.M111.267963 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luesink, E. J. , van Herpen, R. E. M. A. , Grossiord, B. P. , Kuipers, O. P. , & de Vos, W. M. (1998). Transcriptional activation of the glycolytic las operon and catabolite repression of the gal operon in Lactococcus lactis are mediated by the catabolite control protein CcpA. Molecular Microbiology, 30, 789–798. 10.1046/j.1365-2958.1998.01111.x [DOI] [PubMed] [Google Scholar]
- Maguin, E. , Prévost, H. , Ehrlich, S. D. , & Gruss, A. (1996). Efficient insertional mutagenesis in lactococci and other Gram‐positive bacteria. Journal of Bacteriology, 178(3), 931–935. 10.1128/jb.178.3.931-935.1996 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayo, B. , Aleksandrzak‐Piekarczyk, T. , Aleksandrzak‐Piekarczyk, T. , Fernndez, M. , Kowalczyk, M. , lvarez‐Martn, P. , Bardowski, J. . (2010). Updates in the metabolism of lactic acid bacteria In Mozzi F., Raya R. R., Vignolo G. M. (Eds.), Biotechnology of Lactic Acid Bacteria (pp 3–33). Oxford, UK: Wiley‐Blackwell; 10.1002/9780813820866.ch1 [DOI] [Google Scholar]
- Meng, L. M. , Kilstrup, M. , & Nygaard, P. (1990). Autoregulation of PurR repressor synthesis and involvement of PurR in the regulation of purB, purC, purL, purMN and guaBA expression in Escherichia coli . European Journal of Biochemistry, 187, 373–379. 10.1111/j.1432-1033.1990.tb15314.x [DOI] [PubMed] [Google Scholar]
- Michel, V. , & Martley, F. G. (2001). Streptococcus thermophilus in cheddar cheese‐production and fate of galactose. Journal of Dairy Research, 68, 317–325. 10.1017/S0022029901004812 [DOI] [PubMed] [Google Scholar]
- Miroux, B. , & Walker, J. E. (1996). Over‐production of proteins in Escherichia coli: mutant hosts that allow synthesis of some membrane proteins and globular proteins at high levels. Journal of Molecular Biology, 260, 289–298. 10.1006/jmbi.1996.0399 [DOI] [PubMed] [Google Scholar]
- Monedero, V. , Kuipers, O. P. , Jamet, E. , & Deutscher, J. (2001). Regulatory functions of serine‐46‐phosphorylated HPr in Lactococcus lactis . Journal of Bacteriology, 183, 3391–3398. 10.1128/JB.183.11.3391-3398.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morel, F. , Lamarque, M. , Bissardon, I. , Atlan, D. , & Galinier, A. (2001). Autoregulation of the biosynthesis of the CcpA‐like protein, PepR1, in Lactobacillus delbrueckii subsp. bulgaricus . Journal of Molecular Microbiology and Biotechnology, 3, 63–66. [PubMed] [Google Scholar]
- Poolman, B. (1993). Energy transduction in lactic acid bacteria. FEMS Microbiology Reviews, 12, 125–147. 10.1111/j.1574-6976.1993.tb00015.x [DOI] [PubMed] [Google Scholar]
- Poolman, B. , Royer, T. J. , Mainzer, S. E. , & Schmidt, B. F. (1990). Carbohydrate utilization in Streptococcus thermophilus: Characterization of the genes for aldose 1‐epimerase (mutarotase) and UDPglucose 4‐epimerase. Journal of Bacteriology, 172, 4037–4047. 10.1128/jb.172.7.4037-4047.1990 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radziwill‐Bienkowska, J. M. , Le, D. T. L. , Szczesny, P. , Duviau, M.‐P. , Aleksandrzak‐Piekarczyk, T. , Loubière, P. , … Kowalczyk, M. (2016). Adhesion of the genome‐sequenced Lactococcus lactis subsp. cremoris IBB477 strain is mediated by specific molecular determinants. Applied Microbiology and Biotechnology, 100, 9605–9617. 10.1007/s00253-016-7813-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rauch, P. J. , & de Vos, W. M. (1992). Transcriptional regulation of the Tn5276‐located Lactococcus lactis sucrose operon and characterization of the sacA gene encoding sucrose‐6‐phosphate hydrolase. Gene, 121, 55–61. 10.1016/0378-1119(92)90161-H [DOI] [PubMed] [Google Scholar]
- Saier, M. H. (2000). Families of transmembrane sugar transport proteins. Molecular Microbiology, 35, 699–710. 10.1046/j.1365-2958.2000.01759.x [DOI] [PubMed] [Google Scholar]
- Simon, D. , & Chopin, A. (1988). Construction of a vector plasmid family and its use for molecular cloning in Streptococcus lactis . Biochimie, 70, 559–566. 10.1016/0300-9084(88)90093-4 [DOI] [PubMed] [Google Scholar]
- Sissler, M. , Delorme, C. , Bond, J. , Ehrlich, S. D. , Renault, P. , & Francklyn, C. (1999). An aminoacyl‐tRNA synthetase paralog with a catalytic role in histidine biosynthesis. Proceedings of the National Academy of Sciences of the United States of America, 96, 8985–8990. 10.1073/pnas.96.16.8985 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sørensen, K. I. , & Hove‐Jensen, B. (1996). Ribose catabolism of Escherichia coli: Characterization of the rpiB gene encoding ribose phosphate isomerase B and of the rpiR gene, which is involved in regulation of rpiB expression. Journal of Bacteriology, 178, 1003–1011. 10.1128/jb.178.4.1003-1011.1996 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teplyakov, A. , Obmolova, G. , Badet‐Denisot, M.‐A. , Badet, B. , & Polikarpov, I. (1998). Involvement of the C terminus in intramolecular nitrogen channeling in glucosamine 6‐phosphate synthase: Evidence from a 1.6 a crystal structure of the isomerase domain. Structure, 6, 1047–1055. 10.1016/S0969-2126(98)00105-1 [DOI] [PubMed] [Google Scholar]
- Terzaghi, B. E. , & Sandine, W. E. (1975). Improved medium for lactic streptococci and their bacteriophages. Applied Microbiology, 29, 807–813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vaillancourt, K. , Moineau, S. , Frenette, M. , Lessard, C. , & Vadeboncoeur, C. (2002). Galactose and lactose genes from the galactose‐positive bacterium Streptococcus salivarius and the phylogenetically related galactose‐negative bacterium Streptococcus thermophilus: Organization, sequence, transcription, and activity of the gal gene products. Journal of Bacteriology, 184, 785–793. 10.1128/JB.184.3.785-793.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Rooijen, R. J. , & de Vos, W. M. (1990). Molecular cloning, transcriptional analysis, and nucleotide sequence of lacR, a gene encoding the repressor of the lactose phosphotransferase system of Lactococcus lactis . Journal of Biological Chemistry, 265, 18499–18503. [PubMed] [Google Scholar]
- Vaughan, E. E. , van den Bogaard, P. T. C. , Catzeddu, P. , Kuipers, O. P. , & de Vos, W. M. (2001). Activation of silent gal genes in the lac‐gal regulon of Streptococcus thermophilus . Journal of Bacteriology, 183, 1184–1194. 10.1128/JB.183.4.1184-1194.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weickert, M. J. , & Adhya, S. (1993). Control of transcription of gal repressor and iso repressor genes in Escherichia coli . Journal of Bacteriology, 175, 251–258. 10.1128/jb.175.1.251-258.1993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weickert, M. J. , & Chambliss, G. H. (1990). Site‐directed mutagenesis of a catabolite repression operator sequence in Bacillus subtilis . Proceedings of the National Academy of Sciences of the United States of America, 87, 6238–6242. 10.1073/pnas.87.16.6238 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wood, E. (1983). Molecular cloning. A laboratory manual. Biochemical Education, 11, 82 10.1016/0307-4412(83)90068-7 [DOI] [Google Scholar]
- Yamamoto, H. , Serizawa, M. , Thompson, J. , & Sekiguchi, J. (2001). Regulation of the glv operon in Bacillus subtilis: yfiA (GlvR) is a positive regulator of the operon that Is repressed through CcpA and cre . Journal of Bacteriology, 183, 5110–5121. 10.1128/JB.183.17.5110-5121.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zomer, A. L. , Buist, G. , Larsen, R. , Kok, J. , & Kuipers, O. P. (2007). Time‐resolved determination of the CcpA regulon of Lactococcus lactis subsp. cremoris MG1363. Journal of Bacteriology, 189, 1366–1381. 10.1128/JB.01013-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The authors declare that all data generated or analyzed during this study are included in this article.