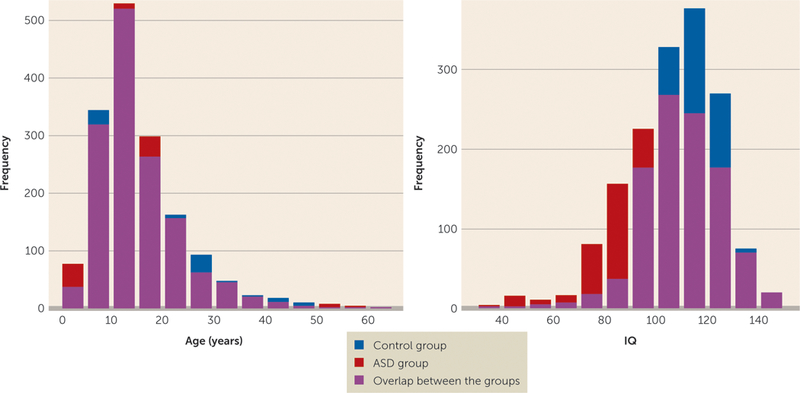
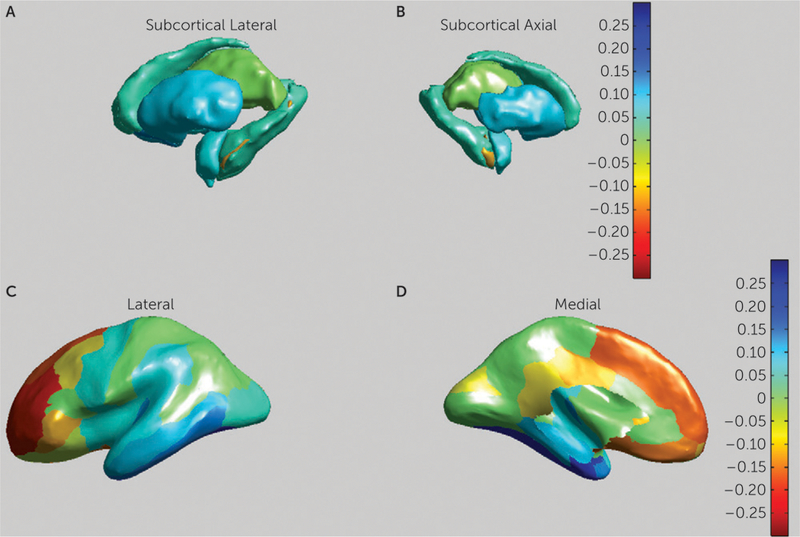
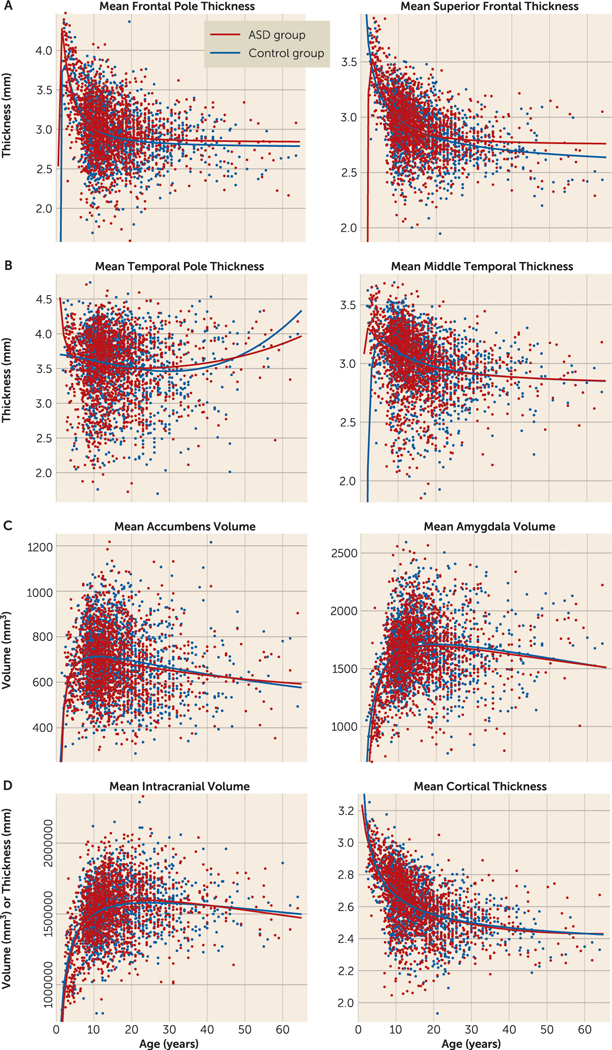
Daan van Rooij
Daan van Rooij, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Evdokia Anagnostou
Evdokia Anagnostou, M.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Celso Arango
Celso Arango, M.D., Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Guillaume Auzias
Guillaume Auzias, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Marlene Behrmann
Marlene Behrmann, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Geraldo F Busatto
Geraldo F Busatto, M.D., Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Sara Calderoni
Sara Calderoni, M.D., Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Eileen Daly
Eileen Daly, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Christine Deruelle
Christine Deruelle, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Adriana Di Martino
Adriana Di Martino, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Ilan Dinstein
Ilan Dinstein, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Fabio Luis Souza Duran
Fabio Luis Souza Duran, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Sarah Durston
Sarah Durston, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Christine Ecker
Christine Ecker, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Damien Fair
Damien Fair, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Jennifer Fedor
Jennifer Fedor, B.S.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Jackie Fitzgerald
Jackie Fitzgerald, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Christine M Freitag
Christine M Freitag, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Louise Gallagher
Louise Gallagher, M.D., Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Ilaria Gori
Ilaria Gori, M.Sc.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Shlomi Haar
Shlomi Haar, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Liesbeth Hoekstra
Liesbeth Hoekstra, M.Sc.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Neda Jahanshad
Neda Jahanshad, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Maria Jalbrzikowski
Maria Jalbrzikowski, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Joost Janssen
Joost Janssen, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Jason Lerch
Jason Lerch, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Beatriz Luna
Beatriz Luna, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Mauricio Moller Martinho
Mauricio Moller Martinho, M.D., Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Jane McGrath
Jane McGrath, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Filippo Muratori
Filippo Muratori, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Clodagh M Murphy
Clodagh M Murphy, M.R.C.Psych., Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Declan GM Murphy
Declan GM Murphy, M.D., F.R.C.Psych.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Kirsten O’Hearn
Kirsten O’Hearn, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Bob Oranje
Bob Oranje, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Mara Parellada
Mara Parellada, M.D., Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Alessandra Retico
Alessandra Retico, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Pedro Rosa
Pedro Rosa, M.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Katya Rubia
Katya Rubia, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Devon Shook
Devon Shook, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Margot Taylor
Margot Taylor, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Paul M Thompson
Paul M Thompson, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Michela Tosetti
Michela Tosetti, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Gregory L Wallace
Gregory L Wallace, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Fengfeng Zhou
Fengfeng Zhou, Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1,
Jan K Buitelaar
Jan K Buitelaar, M.D., Ph.D.
1Department of Cognitive Neuroscience, Donders Institute for Brain, Cognition, and Behavior, Donders Center for Cognitive Neuroimaging, Radboud University Medical Center, Nijmegen, the Netherlands; the KarakterChildand AdolescentPsychiatryUniversity Center, Nijmegen; the Bloorview Research Institute, University of Toronto, Toronto; the Child and Adolescent Psychiatry Department, Gregorio Maranon General University Hospital, School of Medicine, Universidad Complutense, liSGM, CIBERSAM, Madrid; Institut de Neurosciences de la Timone, Aix Marseille Université, CNRS, Marseille, France; the Department of Psychology, Carnegie Mellon University, Pittsburgh; the IRCCS Stella Maris Foundation, Pisa, Italy; the Department of Forensic and Neurodevelopmental Sciences and the Sackler Institute for Translational Neurodevelopment, Institute of Psychiatry, Psychology, and Neuroscience, King’s College London; the Institute for Pediatric Neuroscience, Child Study Center, NYU Langone Health, New York; the Department of Psychology and Department of Brain and Cognitive Sciences, Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Beer Sheva, Israel; Brain Center Rudolf Magnus, Department of Psychiatry, University Medical Center Utrecht, the Netherlands; the Department of Child and Adolescent Psychiatry, Psychosomatics, and Psychotherapy, University Hospital, Goethe University Frankfurt am Main, Frankfurt, Germany; the Department of Behavioral Neuroscience, Oregon Health and Science University, Portland; the Department of Psychiatry, University of Pittsburgh, Pittsburgh; the Department of Psychiatry, School of Medicine, and the Trinity College Institute of Neuroscience, Trinity College, Dublin; the National Institute for Nuclear Physics, Pisa Division, Pisa, Italy; the Mouse Imaging Centre and the Department of Diagnostic Imaging, Hospital for Sick Children, University of Toronto, Toronto; the Department of Clinical and Experimental Medicine, University of Pisa, Pisa, Italy; the Behavioural Genetics Clinic, Adult Autism Service, Behavioural and Developmental Psychiatry Clinical Academic Group, South London and Maudsley Foundation NHS Trust, London; the Laboratory of Neuroimaging, University of Sao Paulo Medical School, Sao Paulo, Brazil; the Imaging Genetics Center, Markand Mary Stevens Institution for Neuroimaging and Informatics, University of Southern California, Marina del Rey; the Department of Speech, Language, and Hearing Sciences, George Washington University, Washington, D.C.; the College of Computer Science and Technology and the Key Laboratory of Symbolic Computation and Knowledge Engineering of the Ministry of Education, Jilin University, Changchun, Jilin, China.
1