

This cross-sectional study maps anhedonia in children and probes the specificity of changes against other clinical phenotypes (low mood, anxiety, and attention-deficit/hyperactivity disorder).
Key Points
Question
How do brain functions in children with anhedonia map onto intrinsic and task-related brain imaging measures?
Findings
In this large-scale cross-sectional functional magnetic resonance imaging study that included 2455 children, anhedonia (but not low mood, anxiety, or attention-deficit/hyperactivity disorder) was associated with hypoconnectivity at rest within the cingulo-opercular network and hypoactivation during reward anticipation in the dorsal striatum and cingulo-opercular network.
Meaning
Anhedonia in children was mapped onto perturbed intrinsic reward arousal integration and diminished extrinsic reward anticipation activity.
Abstract
Importance
Anhedonia can present in children and predict detrimental clinical outcomes.
Objective
To map anhedonia in children onto changes in intrinsic large-scale connectivity and task-evoked activation and to probe the specificity of these changes in anhedonia against other clinical phenotypes (low mood, anxiety, and attention-deficit/hyperactivity disorder [ADHD]).
Design, Setting, and Participants
Functional magnetic resonance imaging (fMRI) data were from the first annual release of the Adolescent Brain Cognitive Development study, collected between September 2016 and September 2017 and analyzed between April and September 2018. Cross-sectional data of children aged 9 to 10 years from unreferred, community samples during rest (n = 2455) and during reward anticipation (n = 2566) and working memory (n = 2465) were analyzed.
Main Outcomes and Measures
Alterations in fMRI data during rest, reward anticipation, and working memory were examined, using both frequentist and Bayesian approaches. Functional MRI connectivity within large-scale networks, between networks, and between networks and subcortical regions were examined during rest. Functional MRI activation were examined during reward anticipation and working memory using the monetary incentive delayed and N-back tasks, respectively.
Results
Among 2455 children with adequate-quality resting-state fMRI data (mean [SD] age, 10.04 [0.62] years; 1187 girls [48.35%]), children with anhedonia (215 [8.76%]), compared with those without anhedonia (2222 [90.51%]), showed hypoconnectivity among various large-scale networks and subcortical regions, including within the arousal-related cingulo-opercular network (mean [SD] with anhedonia, 0.27 [0.08] vs without anhedonia, 0.29 [0.08]; t2,435 = 3.14; P = .002; q[false discovery rate] = 0.07; ln[Bayes factor10] = 2.32). Such hypoconnectivity did not manifest among children with low mood (241 of 2455 [9.82%]), anxiety (93 of 2455 [3.79%]), or ADHD (397 of 2455 [16.17%]), suggesting specificity. Similarly, among 2566 children (mean [SD] age, 10.03 [0.62] years; 1257 girls [48.99%]) with high-quality task-evoked fMRI data, children with anhedonia (213 of 2566 [8.3%]) demonstrated hypoactivation during reward anticipation in various areas, including the dorsal striatum and areas of the cingulo-opercular network. This hypoactivity was not found among children with low mood (240 of 2566 [9.35%]), anxiety (83 of 2566 [3.23%]), or ADHD (430 of 2566 [16.76%]). Moreover, we also found context- and phenotype-specific double dissociations; while children with anhedonia showed altered activation during reward anticipation (but not working memory), those with ADHD showed altered activation during working memory (but not reward anticipation).
Conclusions and Relevance
Using the Adolescent Brain Cognitive Development study data set, phenotype-specific alterations were found in intrinsic large-scale connectivity and task-evoked activation in children with anhedonia. The hypoconnectivity at rest and hypoactivation during reward anticipation complementarily map anhedonia onto aberrations in neural-cognitive processes: lack of intrinsic arousal connectivity during rest and diminishment of extrinsic reward-arousal activity during reward anticipation. These findings help delineate the pathophysiological underpinnings of anhedonia in children.
Introduction
Anhedonia, defined as a loss of interest in previously rewarding activities,1 can present in early life and predict detrimental outcomes, including illness severity, treatment refractoriness, and suicidality.2,3,4,5 Efforts to delineate its neural correlates have been greater in adults than youth.3 Here we used the Adolescent Brain Cognitive Development (ABCD) data set6 to (1) assess the association of anhedonia with resting-state functional magnetic resonance imaging (rs-fMRI) connectivity and task-evoked fMRI activation and (2) compare results in other phenotypes.
We first used both rs-fMRI connectivity and task-evoked fMRI activation to understand the brain functional architecture underlying anhedonia in childhood, a key developmental period.2,3,4,5 Resting-state fMRI connectivity and task-evoked fMRI activation may detect distinct sources of individual differences7; thus, investigating both may provide us a richer account than studying either alone. For rs-fMRI connectivity, we used a large-scale network approach.8,9,10 This allowed us to test the long-postulated notion11,12,13,14,15 that anhedonia results from aberrant connectivity among reward-based striatal areas and large-scale networks subserving cognitive processes, such as sustained arousal (cingulo opercular16,17,18), salience detection (salience19), attention orientation (ventral/dorsal attention20), and mind wandering (default mode21). For task-evoked fMRI activation, we used 2 tasks to evoke context-specific processes: reward anticipation and working memory. Specifically, we use the monetary incentive delayed (MID) task22 to probe the hypothesis that anhedonia in preadolescents is associated with reward-anticipation hypoactivation.11,12,23,24 We used the N-back task8 to examine specificity in reward anticipation findings.
We next probed how findings in anhedonia compared with those with other problems, addressing the need to disentangle shared from unique correlates.24,25,26 Evidence from genetics finds shared etiological correlates among many psychiatric problems.27 Attempts to extend these findings through imaging must confront problems associated with small sample sizes28 and other methodological considerations (eg, referral biases29). Here we attempted to address these problems by using a large, community-based data set to compare findings in anhedonia with those in dysphoria, anxiety disorders, and attention-deficit/hyperactivity disorder (ADHD).
Our first comparator was low mood, a cardinal depression symptom. Low mood or dysphoria is a prototypical negative affect, as opposed to motivation-related processes like anhedonia. Moreover, low mood and anhedonia manifest different developmental trajectories and clinical outcomes.1,5,11,30,31 Task-evoked imaging data in older adolescents24 dissociate anhedonia and low mood during reward anticipation. Here we extend these data to children by probing large-scale rs-fMRI networks alongside task-evoked fMRI activation. Accordingly, we expected to differentiate the modulating associations of anhedonia vs low mood in (1) rs-fMRI connectivity between reward-based striatal areas and large-scale networks and (2) task-evoked fMRI activation during reward anticipation.
Two other comparators are anxiety disorders and ADHD. Anxiety is frequently comorbid with depression.32 Moreover, rodent research found an association between anhedonia and anxiety through interactions between reward- and threat-related circuitry.33 While the clinical nexus between ADHD and anhedonia is understudied, considerable work finds shared etiology between depression and ADHD.27,34,35 Particularly, adults with ADHD display brain hypoactivation during reward anticipation, similar to adults with anhedonia,36 but this pattern is mixed in children.37,38,39,40 It is therefore important to examine whether aberrations in both rs-fMRI connectivity and task-evoked activation in children are specific to anhedonia or reflect an expression of shared etiological factors with other child psychiatric problems.
Methods
Participants
We analyzed the ABCD study41 curated annual release 1.0, containing preprocessed, precomputed data from 4524 children.42 This large-scale study was approved by ethics committees from all 21 institutions where data were collected.43
Note that on December 2, 2019, the ABCD study made a public announcement regarding incorrect postprocessing of its previously released resting-state and task-evoked fMRI data (see Issues Identified With Data Release 2.0.1 on the ABCD Data Sharing site at https://abcdstudy.org/scientists/data-sharing/). Briefly, the ABCD study incorrectly specified field maps of the data obtained on Philips scanners. To formally address this concern for our own study, we removed data obtained on Philips scanners.
Recruited through the school systems, participants were aged 9 to 10 years from 21 sites across the United States. The ABCD investigators obtained written and oral informed consent from parents and children, respectively. Demographics of children are largely consistent with a national survey.44 We selected participants who had no missing value and passed ABCD’s extensive quality control in every fMRI run.45 For rs-fMRI, we excluded data for any participant who had a missing value in at least 1 of the rs-fMRI indices (ie, listwise deletion), and for task-evoked fMRI, we excluded data for participants from statistical analyses separately for each brain region based on a missing value in the particular region (ie, pairwise deletion). This difference is due to the use of “ComBat” to harmonize scanner-related variance in rs-fMRI data (see below) that requires listwise deletion. Using a binary cutoff from the Kiddie Schedule for Affective Disorders and Schizophrenia for DSM-5,46,47 we identified children with anhedonia, low mood, anxiety, and ADHD as those either in the past or currently. We used children’s self-report for the internalizing problems (anhedonia, low mood, and anxiety) and parent report for the externalizing problem (ADHD) following previous recommendations.48 See eTable 1 in Supplement 1 for detailed demographics.
Overall Analyses
We ran analyses on parcellated regions: 333 cortical surface49 and 19 subcortical volumetric.50 We computed (1) rs-fMRI connectivity representing associations among the large-scale cortical-surface networks and subcortical regions and (2) task-evoked fMRI activation for reward anticipation and working memory in cortical areas belonging to each large-scale network and subcortical regions (see below). We used these overall measures of rs-fMRI connectivity and task-evoked activation to examine specific associations with anhedonia compared with low mood, anxiety, and ADHD. For both rs-fMRI and task-evoked fMRI, we used Analysis of Functional NeuroImages (AFNI)51 for preprocessing, R statistical software, version 3.4.3 (R Project for Statistical Computing), with BayesFactor version 0.9.2 (https://richarddmorey.github.io/BayesFactor/) for statistical analyses, and Python version 3.6 with Nilearn52 for visualization.
Resting-State fMRI Connectivity
Using rs-fMRI allowed us to investigate alterations in intrinsic, large-scaled functional connectivity. Details about the acquisition and preprocessing were described elsewhere.41,45,53 (Please note that the ABCD consortium raised concerns about potential problems with the rs-fMRI data that was included in curated annual release 1.0.54 During the revision of this article, we obtained the updated data from the consortium and revised our article accordingly.) Briefly, children viewed a crosshair for 20 minutes while the rs-fMRI data were collected. To quantify connectivity strength during rs-fMRI, we applied a seed-based, correlational approach on parcellated regions.49,55,56 Using a functional atlas,49 we grouped the cortical-surface regions into 12 predefined large-scaled networks49: auditory, cingulo-opercular, cingulo-parietal, default-mode, dorsal-attention, fronto-parietal, retrosplenial-temporal, salience, sensorimotor-hand, sensorimotor-mouth, ventral-attention, and visual networks. After discarding regions that were not fit with these large-scale network definitions,49 we computed rs-fMRI connectivity-strength indices using Fisher r to z transformation of the mean correlations between pairs of regions within each large-scale network (12), between large-scale networks (66), and between large-scale networks and subcortical regions (228). This resulted in 306 connectivity indices. To harmonize scanner-related variance among 27 scanners used, we applied the Empirical-Bayes “ComBat” method57,58,59,60 using the command: ComBatHarmonization::combat(dat = dat, batch = batch), in which dat is a matrix of all rs-fMRI indices by participants and batch is a vector of scanner identifications.60
Task-Evoked fMRI Activation
Using task-evoked fMRI allowed us to investigate alterations in extrinsic, context-specific neural processes: reward anticipation in the MID tasks22 and working memory in the N-back tasks.8 Details about the acquisition and preprocessing of the task fMRI data have been previously published.42,45 Task-evoked activation was modeled using AFNI’s 3dDeconvolve.51 Recent research shows that using within-individual contrasts can mitigate scanner-related variance in the ABCD task-evoked fMRI data.57 Accordingly, we did not apply the ComBat method on the task-evoked fMRI data and instead computed contrasts between conditions of interest.
The MID task started with a cue, indicating possible earnings: large reward ($5), small reward ($0.20), neutral ($0), small punishment ($0.20), and large punishment ($5). After a variable period following the cue, the children were shown a brief target. To either earn reward or avoid punishment, children needed to respond before the target disappeared. Following the response, children saw the outcome feedback of the trial. To focus on reward anticipation, we investigated the contrast that maximizes this process—the large reward vs neutral cue.
For the N-back task, depending on the condition, children needed to respond whether the stimulus was the same as (1) the one shown 2 trials earlier (2 back) or (2) the target stimulus shown at the beginning (0 back). Stimuli included houses and emotional and nonemotional faces. To focus on working memory, we investigated the contrast between the 2 back vs 0 back conditions regardless of stimulus type.
Associations With Anhedonia and Other Clinical Phenotypes
To investigate the modulating effects of anhedonia, we conducted 2-tailed independent-samples t tests on each rs-fMRI index and task-evoked fMRI region between children with and without anhedonia regardless of whether the children had any other clinical phenotypes. To avoid outliers, we excluded data points that deviated over 1.5 interquartile ranges from the nearer quartile for each index or region. We used Benjamini-Hochberg false discovery rate (FDR)61 to adjust for multiple comparisons across indices or regions. Besides frequentist t tests, we applied Bayesian hypothesis testing using Jeffreys-Zellner-Siow–prior Bayesian t tests62 with a Cauchy prior (r scale = 0.707) on each index or region. These Bayesian t tests provided Bayes Factor10 (BF10), which expresses the likelihood of the observed data under the alternative (of there being a difference), relative to the null (of there being no difference), hypothesis. Thus, BF10 allows us to quantify evidence for the difference due to having anhedonia relative to evidence for the lack of the difference, which is informative especially because the sample size is large.3 Based on Jeffreys,63,64 natural-log transformed (ln) BF10 greater than 1.1 and less than −1.2 (ie, nontransformed BF10 > 3 and < 0.3) are interpreted as substantial evidence for alternative and null hypotheses, respectively.
To demarcate anhedonia vs other phenotypes, we first repeated the same analyses done on anhedonia on low mood, anxiety, and ADHD. This is to investigate whether low mood, anxiety, and ADHD modulated similar rs-fMRI indices and task-evoked regions to anhedonia. If the modulation were different, it would rule out the possibility that the associations with anhedonia were due to comorbidity with these clinical phenotypes. Second, if anhedonia significantly modulated any rs-fMRI indices or task-evoked regions, we would conduct follow-up, pairwise analyses to directly compare between children with anhedonia vs with another phenotype on these indices or regions. That is, in these follow-up analyses, we only included the indices or regions that differed (q[FDR] < .05 or ln[BF10] > 1.1) between children with vs without anhedonia and further investigated if these indices or regions were differed between children with anhedonia vs with another phenotype. We also excluded children who had both anhedonia and the compared phenotype from these follow-up analyses.
Results
For rs-fMRI, of 4524 children, 3091 children had at least 4 rs-fMRI runs that passed the quality control. Among those who passed the quality control, 2726 children were not scanned with a Philips scanner. Removing children with a missing value in any rs-fMRI indices, which was required for ‘ComBat’, left 2455 children (mean [SD] age, 10.04 [0.62] years; 1187 girls [48.35%]). Some children had anhedonia (215 [8.76%]), low mood (241 [9.82%]), anxiety (93 [3.79%]), and ADHD (397 [16.17%]). For the MID task, there were task-evoked fMRI data from 3255 children available. The data from 2874 children passed the quality control in all runs of the MID task. Among those whose data passed the quality control, 2566 children (mean [SD] age, 10.03 [0.62] years; 1257 girls [48.99%]) were not scanned with a Philips scanner. Some children had anhedonia (213 [8.3%]), low mood (240 [9.35%]), anxiety (83 [3.23%]), and ADHD (430 [16.76%]). For the N-back task, there were task-evoked fMRI data from 3099 children available. The data from 2745 children passed the quality control in all runs. Among those whose data passed the quality control, 2465 children (mean [SD] age, 10.04 [0.62] years; 1120 girls [46.57%]) were not scanned with a Philips scanner. Some children had anhedonia (207 [8.4%]), low mood (219 [8.88%]), anxiety (83 [3.37%]), and ADHD (409 [16.59%]).
Resting-State fMRI Connectivity: Modulation by Anhedonia
We found evidence for the difference (q[FDR] < .05 or ln[BF10] > 1.1) between children with and without anhedonia in several rs-fMRI connectivity indices (Figure 1 and eTable 2 in Supplement 1). First, compared with children without anhedonia, the cingulo-opercular network of children with anhedonia exhibited weaker within-network connectivity (ln[BF10] = 2.32) and weaker positive correlations with the brain stem (ln[BF10] = 1.2). Their brain stem also had weaker positive correlations with the sensorimotor-hand network (ln[BF10] = 2.85), which had weaker positive correlations with the right hippocampus (ln[BF10] = 1.81). Conversely, for children with, compared with without, anhedonia, the brain stem had stronger positive correlations with the cingulo-parietal network (ln[BF10] = 3.37), which had weaker positive correlations with the right pallidum (ln[BF10] = 1.97). Similarly, in children with anhedonia, the salience network showed weaker anticorrelations with the left ventral diencephalon50 (ln[BF10] = 2.81). The dorsal attention network of children with vs without anhedonia also displayed weaker anticorrelations with the default mode network (ln[BF10] = 3.29) and left hippocampus (ln[BF10] = 5.06). Lastly, the retrosplenial-temporal network of children with anhedonia showed weaker within-network connectivity (ln[BF10] = 3.99) and weaker positive correlations with the right cerebellum (ln[BF10] = 2.15).
Figure 1. Group Differences in Resting-State Functional Magnetic Resonance Imaging (rs-fMRI) Connectivity z[r].

The arrows depict the differences (q[false discovery rate] < .05 or ln[Bayes Factor10] > 1.1) between children with vs without anhedonia. Green arrows depict averaged positive correlations between 2 nodes across participants, while red arrows depict averaged negative (ie, anti) correlations between 2 nodes. The comparison between children and another clinical phenotype only included the rs-fMRI indices that differed (q[false discovery rate] < .05 or ln[Bayes Factor10] > 1.1) between children with vs without anhedonia. ADHD indicates attention-deficit/hyperactivity disorder.
Task-Evoked fMRI Activation: Modulation by Anhedonia
We found evidence for the difference (q[FDR] < .05 or ln[BF10] > 1.1) between children with and without anhedonia in task-based activation during reward anticipation in the MID task (Figure 2, Figure 3, and eTable 6 in Supplement 1). Children with anhedonia showed hypoactivation at various cortical-surface regions, many of which were part of the large-scale networks that were significant in the rs-fMRI analyses. For instance, children with anhedonia had hypoactivation in areas in the cingulo-opercular (eg, the midcingulate cortex and right superior-marginal gyrus), salience (eg, the anterior-cingulate cortex), dorsal-attention (eg, the superior parietal cortex), default-mode (eg, the medial-prefrontal cortex), sensorimotor-hand and -mouth (eg, the right supplementary-motor cortex and bilateral precentral and postcentral gyri) networks. As for subcortical-volumetric areas, we found hypoactivation in the putamen, which is part of the dorsal striatum. We found fewer regions, mainly at the default-mode network, passing the threshold for the difference (q[FDR] < .05 or ln[BF10] > 1.1) between children with and without anhedonia in task-evoked activation on the N-back task (Figure 4 and eTable 10 in Supplement 1).
Figure 2. Group Differences in Task-Evoked Functional Magnetic Resonance Imaging Activation During Reward Anticipation.
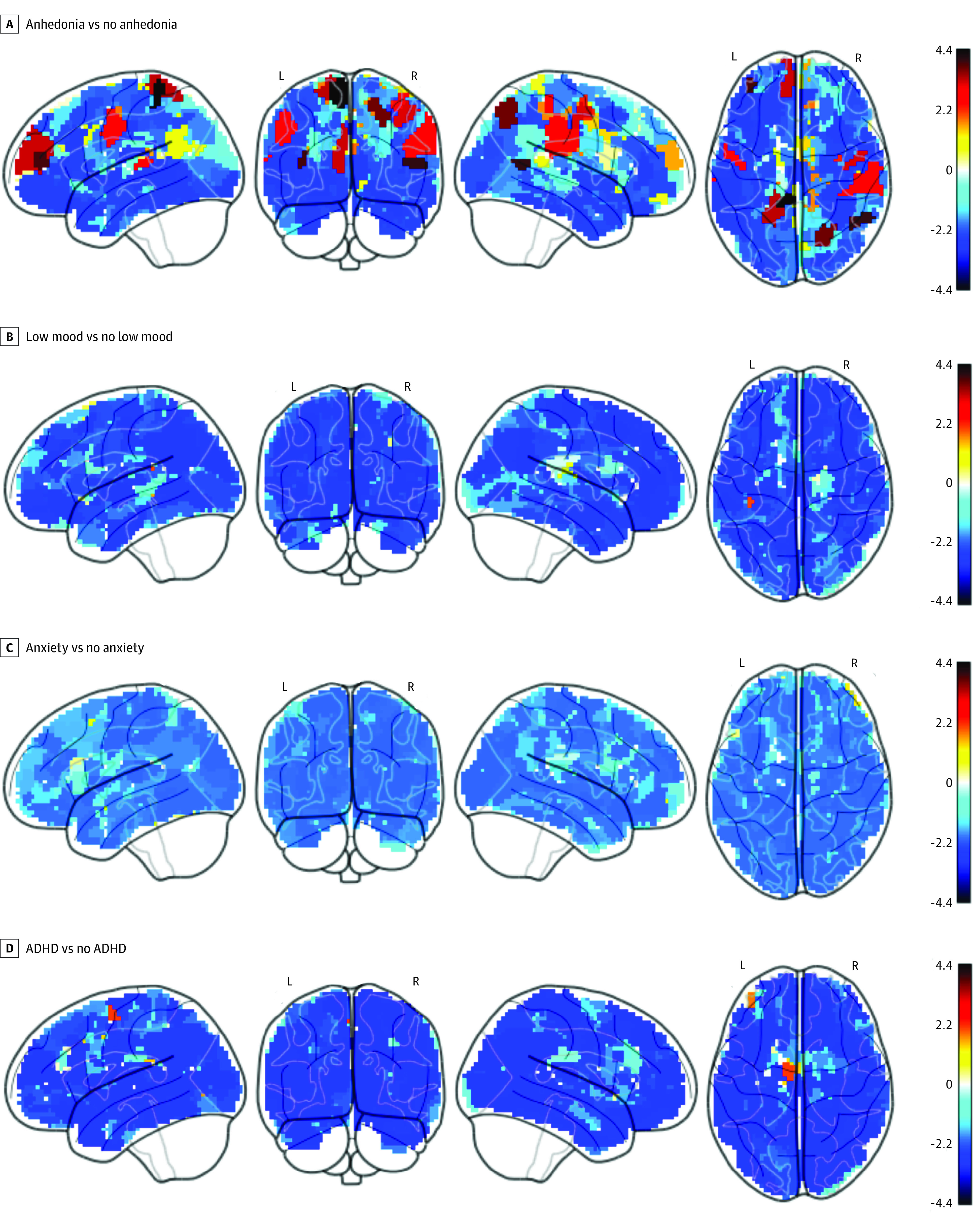
The activation was computed from the [large reward > neutral cue] contrast in the monetary incentive delayed task at 333 cortical-surface regions. Ln(BF10) depicts the natural log of Bayes Factor10. The value of ln(BF10) greater than 1.1 and less than −1.2 are interpreted as substantial evidence for the alternative (of there being a difference) and null hypotheses (of there being no difference), respectively. ADHD indicates attention-deficit/hyperactivity disorder.
Figure 3. Group Differences Between Children With and Without Anhedonia in Task-Evoked Functional Magnetic Resonance Imaging Activation During Reward Anticipation.
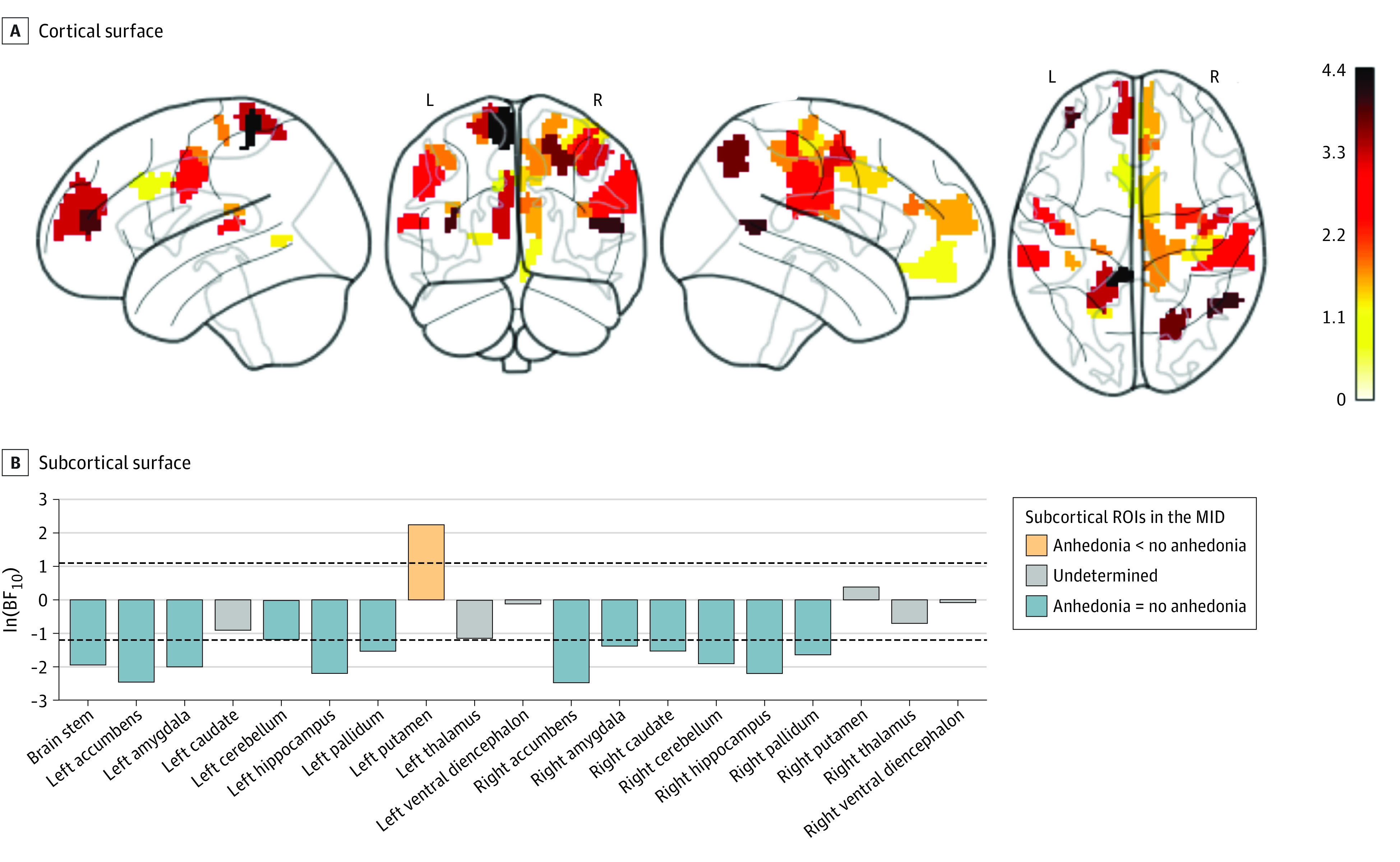
The natural log of Bayes Factor10 (BF10) on cortical regions in the monetary incentive delayed (MID) task for anhedonia > no anhedonia. The activation was computed from the large reward > neutral cue contrast in the MID task on 333 cortical surface (A) and 19 subcortical-volumetric (B) regions. For the cortical surface (A), we only show the regions with either q[false discovery rate] < 0.05 or ln(BF10) > 1.1 that suggests evidence for weaker activation in children with anhedonia compared with children without anhedonia. For the subcortical surface (B), the area between the 2 dotted lines indicates the ln(BF10) threshold between −1.2 and 1.1, at which the evidence is not strong enough to support either the alternative or null hypothesis. ROI indicates region of interest.
Figure 4. Group Differences in Task-Evoked Functional Magnetic Resonance Imaging Activation During Working Memory.
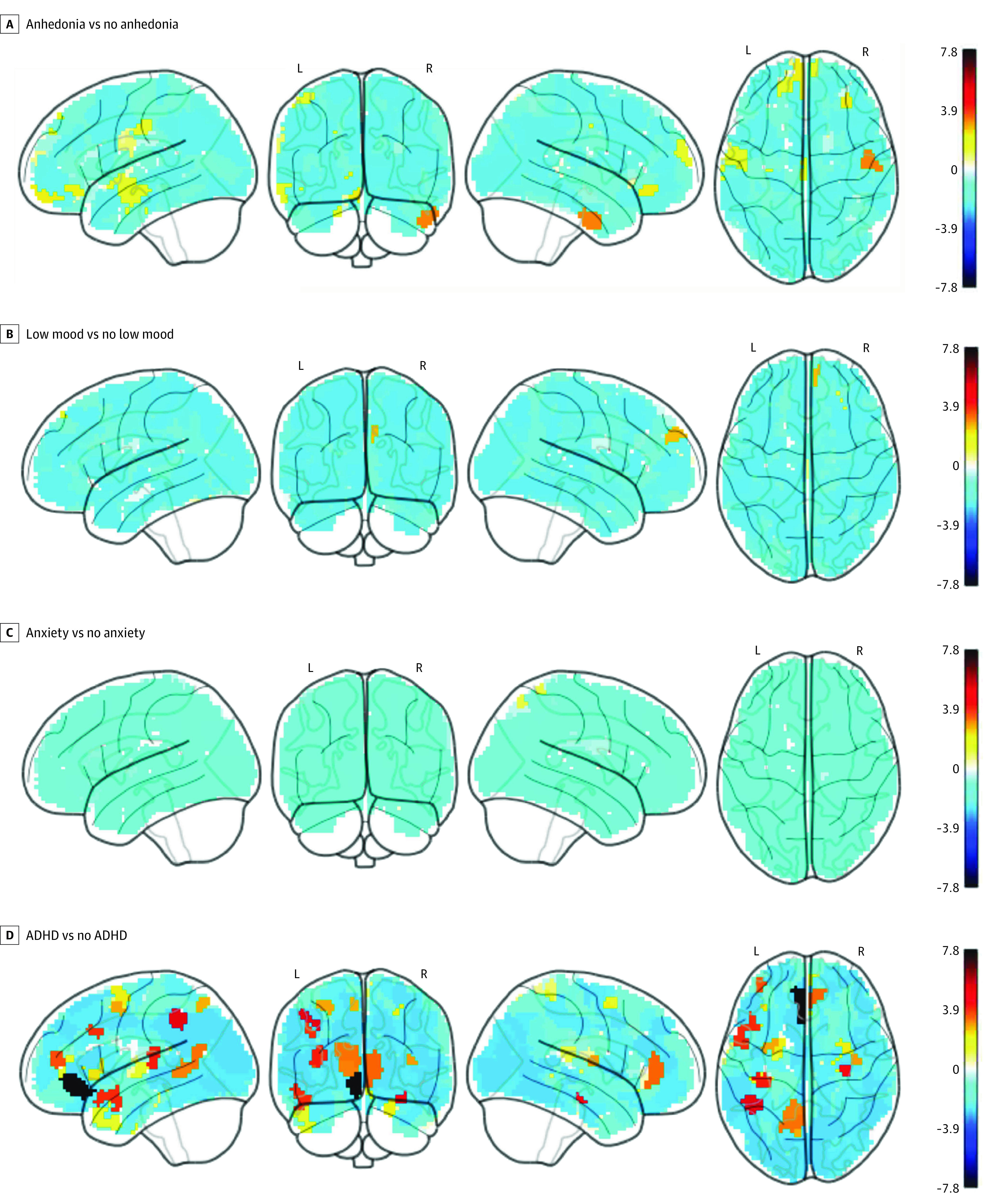
The activation was computed from the [2 back>0 back] contrast in the N-back task on 333 cortical surface regions.49 Ln(BF10) depicts the natural log of BF10. The value of ln(BF10) >1.1 and <−1.2 are interpreted as substantial evidence for the alternative (of there being a difference), and null hypotheses (of there being no difference), respectively. ADHD indicates attention-deficity/hyperactivity disorder; BF10, Bayes Factor10.
Phenotype Specificity in rs-fMRI Connectivity
We first examined brain function in each other phenotype. We then contrasted findings in these phenotypes with those in anhedonia.
We found evidence for the difference (q[FDR] < .05 or ln[BF10] > 1.1) associated with ADHD in some rs-fMRI connectivity indices (eTable 5 in Supplement 1) that were different from those associated with anhedonia. For instance, compared with children without ADHD, children with ADHD had weaker within-network connectivity in the fronto-parietal (ln[BF10] = 3.37) and default-mode (ln[BF10] = 3.63) networks. Additionally, children with ADHD had stronger positive correlations between the cingular-parietal network and amygdala (ln[BF10] = 4.45) and between the cingulo-opercular and retrosplenial-temporal networks (ln[BF10] = 7.20). They also had stronger anticorrelations between the auditory network and brain stem (ln[BF10] = 5.60). Fewer rs-fMRI connectivity indices demonstrated evidence for the difference (q[FDR] < .05 or ln[BF10] > 1.1) between children with and without low mood and between children with and without anxiety (eTables 3 and 4 in Supplement 1).
Figure 1 and eTable 17 in Supplement 1 show follow-up analyses that directly compared rs-fMRI connectivity between children with anhedonia and those with another phenotype. Briefly, children with anhedonia differed from those with low mood and with ADHD in many connectivity indices. These involved the cingulo-opercular, salience, cingulo-parietal, and sensorimotor-hand networks. Compared with children with anxiety, those with anhedonia exhibited weaker anticorrelations between the dorsal-attention network and left hippocampus.
Phenotype Specificity in Task-Evoked fMRI Activation
As with rs-fMRI above, we first show the modulation by each of the other phenotypes on their own. We then contrast them to anhedonia.
Findings in ADHD markedly differed from those in anhedonia. Compared with anhedonia findings, we found fewer regions that demonstrated differences (q[FDR] > .05 or ln[BF10] > 1.1) between children with and without ADHD in task-evoked activation during reward anticipation (Figure 2 and eTable 9 in Supplement 1). However, we found more regions that demonstrated differences (q[FDR] < .05 or ln[BF10] > 1.1) between children with and without ADHD in task-evoked activation during working memory (Figure 4 and eTable 13 in Supplement 1). Here, children with ADHD showed alterations in many areas, eg, the left inferior-frontal, middle-frontal and right super-marginal gyri, anterior-cingulate cortex, bilateral inferior-parietal lobes, and bilateral hippocampi among others. As for low mood and anxiety, we found only a few regions that demonstrated phenotype-associated differences (q[FDR] > .05 or ln[BF10] > 1.1) in task-evoked activation both during reward anticipation and working memory (Figure 2, Figure 4; and eTables 7-8 and 11-12 in Supplement 1).
Of note, anhedonia modulated task-evoked activation during reward anticipation, more so than during working memory. As such, we conducted pairwise, follow-up analyses during reward anticipation, directly comparing task-evoked activation between children with anhedonia vs those with another phenotype. These pairwise analyses yielded differences (q[FDR] > .05 or ln[BF10] > 1.1) between children with anhedonia and those with anxiety in several regions, including areas in the cingulo-opercular network, but yielded differences in only a few regions in other comparison pairs (eTables 14-16 in Supplement 1).
Discussion
This study had 2 aims, namely to (1) compare rs-fMRI connectivity and task-evoked fMRI activation in children with or without anhedonia and (2) probe the specificity of alterations in anhedonia against other clinical phenotypes. For the first aim, we identified functional brain alterations in children with anhedonia in both rs-fMRI connectivity and task-evoked activation. For rs-fMRI, we found changes in various connectivity indices, including within the cingulo-opercular network, which is associated with sustained arousal18,65 and is often coactivated with the striatum during tasks requiring high alertness.18,65 Thus, their altered connectivity with the cingulo-opercular network suggests an altered arousal in children with anhedonia.
Children with anhedonia also demonstrated context specificity, expressed as task-specific alterations. Here, children with anhedonia showed hypoactivation during reward anticipation in the dorsal striatum (putamen) and cingulo-opercular network. These areas of hypoactivation overlap with those of a meta-analysis of anhedonia during reward anticipation23 and those of recent work on arousal during reward anticipation.66 Importantly, this hypoactivation pattern in anhedonia only emerged during reward anticipation but not working memory, thus indicating context specificity. Altogether, hypoconnectivity at rest and hypoactivation during reward processing complementarily map anhedonia onto aberrations in neural cognitive processes: lack of intrinsic arousal connectivity and diminishment of reward-arousal activity while anticipating rewards.
The second aim concerns the phenotypic specificity—whether neural correlates of anhedonia could be demarcated against other phenotypes. Several patterns emerged from investigating the modulation by low mood, anxiety, and ADHD. Regarding associations unique to anhedonia, anhedonia was associated with various changes in connectivity indices linked to the cingulo-opercular network, many of which were significantly different from low mood and ADHD. Similarly, from task-evoked activation, anhedonia was the only phenotype among the 4 to show aberrations during reward anticipation, while ADHD was the only one to show aberrations during working memory, in keeping with prior theory and findings.37 This double dissociation is consistent with research linking anhedonia to diminished reward anticipation11,12,23,24 and ADHD to deficits in working memory.67 However, other findings provided less compelling evidence of specificity. Namely, in some instances, findings occurring in anhedonia were not found with anxiety, although contrasts of children with anhedonia and anxiety revealed differences. Regardless, overall patterns did provide evidence of specificity. In particular, the data linked anhedonia specifically to aberrant connectivity within the arousal-related cingulo-opercular network and hypoactivity during reward anticipation.
Limitations
Our study has some limitations. First, participants were drawn from the community and therefore were not necessarily representative of clinical cases. However, this approach avoids referral biases68 and may provide advantages associated with generalizability. Second, our definition of anhedonia relies on a measure generated from criterion standard clinical interviews (Kiddie Schedule for Affective Disorders and Schizophrenia for DSM-5). While this is an advantage, it also has the limitation of only generating a categorical definition of anhedonia. It is therefore reassuring that our findings are consistent with previous studies in anhedonia.23 Third, despite the large sample size, there were too few cases of major depressive disorder and schizophrenia at this age group to examine the modulation by anhedonia within those disorders. Both illnesses have been associated with striatal reward-network aberrations37,42,69 and these may be due to anhedonia. Future waves of ABCD should capture the age-related increase in prevalence of both disorders.
Conclusions
Anhedonia in youth is associated with hypoconnectivity at rest and hypoactivation during reward anticipation. This suggests perturbed intrinsic arousal connectivity and diminished extrinsic reward-arousal activity in the reward-anticipation context. Detecting such anhedonia-specific alterations helps differentiate the pathophysiological underpinnings of anhedonia from other phenotypes.
eTable 1. Demographics of participants who passed the quality control for resting-state fMRI (2455 children)
eTable 2. Group differences between children with vs. without anhedonia in the resting-state fMRI connectivity z[r]
eTable 6. Group differences between children with vs. without anhedonia during reward anticipation [Large Reward > Neutral Cue] in the MID task-fMRI activation
eTable 10. Group differences between children with vs. without anhedonia during working-memory [Two-Back > Zero-Back] in the N-Back task-fMRI activation
eTable 5. Group differences between children with vs. without ADHD in the resting-state fMRI connectivity z[r]
eTable 3. Group differences between children with vs. without low mood in the resting-state fMRI connectivity z[r]
eTable 4. Group differences between children with vs. without Anxiety in the resting-state fMRI connectivity z[r]
eTable 9. Group differences between children with vs. without ADHD during reward anticipation [Large Reward > Neutral Cue] in the MID task-fMRI activation
eTable 13. Group differences between children with vs. without ADHD during working-memory [Two-Back > Zero-Back] in the N-Back task-fMRI activation
eTable 7. Group differences between children with vs. without low mood during reward anticipation [Large Reward > Neutral Cue] in the MID task-fMRI activation
eTable 8. Group differences between children with vs. without anxiety during reward anticipation [Large Reward > Neutral Cue] in the MID task-fMRI activation
eTable 11. Group differences between children with vs. without low mood during working-memory [Two-Back > Zero-Back] in the N-Back task-fMRI activation
eTable 12. Group differences between children with vs. without anxiety during working-memory [Two-Back > Zero-Back] in the N-Back task-fMRI activation
eTable 14. Group differences between anhedonia and low-mood during reward-anticipation [Large Reward > Neutral Cue] in the MID task-fMRI activation
eTable 15. Group differences between anhedonia and anxiety during reward-anticipation [Large Reward > Neutral Cue] in the MID task-fMRI activation
eTable 16. Group differences between anhedonia and ADHD during reward-anticipation [Large Reward > Neutral Cue] in the MID task-fMRI activation
eTable 17. Follow-up Analysis
eReferences.
Retracted article with errors highlighted
Replacement article with corrections highlighted
References
- 1.American Psychiatric Association . Diagnostic and Statistical Manual of Mental Disorders. 5th ed. Washington, DC: American Psychiatric Association; 2013. [Google Scholar]
- 2.Gutkovich Z, Morrissey RF, Espaillat RK, Dicker R. Anhedonia and pessimism in hospitalized depressed adolescents. Depress Res Treat. 2011;2011:795173. doi: 10.1155/2011/795173 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Luby JL, Mrakotsky C, Heffelfinger A, Brown K, Spitznagel E. Characteristics of depressed preschoolers with and without anhedonia: evidence for a melancholic depressive subtype in young children. Am J Psychiatry. 2004;161(11):1998-2004. doi: 10.1176/appi.ajp.161.11.1998 [DOI] [PubMed] [Google Scholar]
- 4.Robbins DR, Alessi NE. Depressive symptoms and suicidal behavior in adolescents. Am J Psychiatry. 1985;142(5):588-592. doi: 10.1176/ajp.142.5.588 [DOI] [PubMed] [Google Scholar]
- 5.McMakin DL, Olino TM, Porta G, et al. Anhedonia predicts poorer recovery among youth with selective serotonin reuptake inhibitor treatment-resistant depression. J Am Acad Child Adolesc Psychiatry. 2012;51(4):404-411. doi: 10.1016/j.jaac.2012.01.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Aims and Scope. Dev Cogn Neurosci. 2018;21:IFC. doi: 10.1016/S1878-9293(18)30129-4 [DOI] [Google Scholar]
- 7.Gratton C, Laumann TO, Nielsen AN, et al. Functional brain networks are dominated by stable group and individual factors, not cognitive or daily variation. Neuron. 2018;98(2):439-452. doi: 10.1016/j.neuron.2018.03.035 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Barch DM, Burgess GC, Harms MP, et al. ; WU-Minn HCP Consortium . Function in the human connectome: task-fMRI and individual differences in behavior. Neuroimage. 2013;80:169-189. doi: 10.1016/j.neuroimage.2013.05.033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Williams LM. Precision psychiatry: a neural circuit taxonomy for depression and anxiety. Lancet Psychiatry. 2016;3(5):472-480. doi: 10.1016/S2215-0366(15)00579-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Menon V. Large-scale brain networks and psychopathology: a unifying triple network model. Trends Cogn Sci. 2011;15(10):483-506. doi: 10.1016/j.tics.2011.08.003 [DOI] [PubMed] [Google Scholar]
- 11.Treadway MT, Zald DH. Reconsidering anhedonia in depression: lessons from translational neuroscience. Neurosci Biobehav Rev. 2011;35(3):537-555. doi: 10.1016/j.neubiorev.2010.06.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Pizzagalli DA. Depression, stress, and anhedonia: toward a synthesis and integrated model. Annu Rev Clin Psychol. 2014;10(1):393-423. doi: 10.1146/annurev-clinpsy-050212-185606 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mulders PC, van Eijndhoven PF, Schene AH, Beckmann CF, Tendolkar I. Resting-state functional connectivity in major depressive disorder: a review. Neurosci Biobehav Rev. 2015;56:330-344. doi: 10.1016/j.neubiorev.2015.07.014 [DOI] [PubMed] [Google Scholar]
- 14.Tu P-C, Hsieh J-C, Li C-T, Bai Y-M, Su T-P. Cortico-striatal disconnection within the cingulo-opercular network in schizophrenia revealed by intrinsic functional connectivity analysis: a resting fMRI study. Neuroimage. 2012;59(1):238-247. doi: 10.1016/j.neuroimage.2011.07.086 [DOI] [PubMed] [Google Scholar]
- 15.Orliac F, Naveau M, Joliot M, et al. Links among resting-state default-mode network, salience network, and symptomatology in schizophrenia. Schizophr Res. 2013;148(1-3):74-80. doi: 10.1016/j.schres.2013.05.007 [DOI] [PubMed] [Google Scholar]
- 16.Sadaghiani S, Hesselmann G, Kleinschmidt A. Distributed and antagonistic contributions of ongoing activity fluctuations to auditory stimulus detection. J Neurosci. 2009;29(42):13410-13417. doi: 10.1523/JNEUROSCI.2592-09.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Coste CP, Kleinschmidt A. Cingulo-opercular network activity maintains alertness. Neuroimage. 2016;128:264-272. doi: 10.1016/j.neuroimage.2016.01.026 [DOI] [PubMed] [Google Scholar]
- 18.Sadaghiani S, D’Esposito M. Functional characterization of the cingulo-opercular network in the maintenance of tonic alertness. Cereb Cortex. 2015;25(9):2763-2773. doi: 10.1093/cercor/bhu072 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Seeley WW, Menon V, Schatzberg AF, et al. Dissociable intrinsic connectivity networks for salience processing and executive control. J Neurosci. 2007;27(9):2349-2356. doi: 10.1523/JNEUROSCI.5587-06.2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Corbetta M, Patel G, Shulman GL. The reorienting system of the human brain: from environment to theory of mind. Neuron. 2008;58(3):306-324. doi: 10.1016/j.neuron.2008.04.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Raichle ME, MacLeod AM, Snyder AZ, Powers WJ, Gusnard DA, Shulman GL. A default mode of brain function. Proc Natl Acad Sci U S A. 2001;98(2):676-682. doi: 10.1073/pnas.98.2.676 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Knutson B, Westdorp A, Kaiser E, Hommer D. FMRI visualization of brain activity during a monetary incentive delay task. Neuroimage. 2000;12(1):20-27. doi: 10.1006/nimg.2000.0593 [DOI] [PubMed] [Google Scholar]
- 23.Zhang B, Lin P, Shi H, et al. Mapping anhedonia-specific dysfunction in a transdiagnostic approach: an ALE meta-analysis. Brain Imaging Behav. 2016;10(3):920-939. doi: 10.1007/s11682-015-9457-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Stringaris A, Vidal-Ribas Belil P, Artiges E, et al. ; IMAGEN Consortium . The brain’s response to reward anticipation and depression in adolescence: dimensionality, specificity, and longitudinal predictions in a community-based sample. Am J Psychiatry. 2015;172(12):1215-1223. doi: 10.1176/appi.ajp.2015.14101298 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Segarra N, Metastasio A, Ziauddeen H, et al. Abnormal frontostriatal activity during unexpected reward receipt in depression and schizophrenia: relationship to anhedonia. Neuropsychopharmacology. 2016;41(8):2001-2010. doi: 10.1038/npp.2015.370 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hägele C, Schlagenhauf F, Rapp M, et al. Dimensional psychiatry: reward dysfunction and depressive mood across psychiatric disorders. Psychopharmacology (Berl). 2015;232(2):331-341. doi: 10.1007/s00213-014-3662-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lee SH, Ripke S, Neale BM, et al. ; Cross-Disorder Group of the Psychiatric Genomics Consortium; International Inflammatory Bowel Disease Genetics Consortium (IIBDGC) . Genetic relationship between five psychiatric disorders estimated from genome-wide SNPs. Nat Genet. 2013;45(9):984-994. doi: 10.1038/ng.2711 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kraemer HC. Statistical issues in assessing comorbidity. Stat Med. 1995;14(8):721-733. doi: 10.1002/sim.4780140803 [DOI] [PubMed] [Google Scholar]
- 29.Feinstein AR, Horwitz RI. A critique of the statistical evidence associating estrogens with endometrial cancer. Cancer Res. 1978;38(11 Pt 2):4001-4005. [PubMed] [Google Scholar]
- 30.Pine DS, Cohen E, Cohen P, Brook J. Adolescent depressive symptoms as predictors of adult depression: moodiness or mood disorder? Am J Psychiatry. 1999;156(1):133-135. doi: 10.1176/ajp.156.1.133 [DOI] [PubMed] [Google Scholar]
- 31.Insel T, Cuthbert B, Garvey M, et al. Research domain criteria (RDoC): toward a new classification framework for research on mental disorders. Am J Psychiatry. 2010;167(7):748-751. doi: 10.1176/appi.ajp.2010.09091379 [DOI] [PubMed] [Google Scholar]
- 32.Winer ES, Bryant J, Bartoszek G, Rojas E, Nadorff MR, Kilgore J. Mapping the relationship between anxiety, anhedonia, and depression. J Affect Disord. 2017;221:289-296. doi: 10.1016/j.jad.2017.06.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Bolton JL, Molet J, Regev L, et al. Anhedonia following early-life adversity involves aberrant interaction of reward and anxiety circuits and is reversed by partial silencing of amygdala corticotropin-releasing hormone gene. Biol Psychiatry. 2018;83(2):137-147. doi: 10.1016/j.biopsych.2017.08.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Cross-Disorder Group of the Psychiatric Genomics Consortium . Identification of risk loci with shared effects on five major psychiatric disorders: a genome-wide analysis. Lancet. 2013;381(9875):1371-1379. doi: 10.1016/S0140-6736(12)62129-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Power RA, Tansey KE, Buttenschøn HN, et al. ; CONVERGE Consortium, CARDIoGRAM Consortium, GERAD1 Consortium . Genome-wide association for major depression through age at onset stratification: major depressive disorder working group of the psychiatric genomics consortium. Biol Psychiatry. 2017;81(4):325-335. doi: 10.1016/j.biopsych.2016.05.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Plichta MM, Scheres A. Ventral-striatal responsiveness during reward anticipation in ADHD and its relation to trait impulsivity in the healthy population: a meta-analytic review of the fMRI literature. Neurosci Biobehav Rev. 2014;38:125-134. doi: 10.1016/j.neubiorev.2013.07.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Demurie E, Roeyers H, Wiersema JR, Sonuga-Barke E. No evidence for inhibitory deficits or altered reward processing in ADHD: data from a new integrated monetary incentive delay go/no-go task. J Atten Disord. 2016;20(4):353-367. doi: 10.1177/1087054712473179 [DOI] [PubMed] [Google Scholar]
- 38.Ma I, van Holstein M, Mies GW, et al. Ventral striatal hyperconnectivity during rewarded interference control in adolescents with ADHD. Cortex. 2016;82:225-236. doi: 10.1016/j.cortex.2016.05.021 [DOI] [PubMed] [Google Scholar]
- 39.Lemiere J, Danckaerts M, Van Hecke W, et al. Brain activation to cues predicting inescapable delay in adolescent attention deficit/hyperactivity disorder: an fMRI pilot study. Brain Res. 2012;1450:57-66. doi: 10.1016/j.brainres.2012.02.027 [DOI] [PubMed] [Google Scholar]
- 40.Paloyelis Y, Mehta MA, Faraone SV, Asherson P, Kuntsi J. Striatal sensitivity during reward processing in attention-deficit/hyperactivity disorder. J Am Acad Child Adolesc Psychiatry. 2012;51(7):722-732. doi: 10.1016/j.jaac.2012.05.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Casey BJ, Cannonier T, Conley MI, et al. ; ABCD Imaging Acquisition Workgroup . The Adolescent Brain Cognitive Development (ABCD) study: imaging acquisition across 21 sites. Dev Cogn Neurosci. 2018;32:43-54. doi: 10.1016/j.dcn.2018.03.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Keren H, O'Callaghan G, Vidal-Ribas P, et al. Reward processing in depression: a conceptual and meta-analytic review across fMRI and EEG studies. Am J Psychiatry. 2018:appiajp201817101124. doi: 10.1176/appi.ajp.2018.17101124 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Auchter AM, Hernandez Mejia M, Heyser CJ, et al. A description of the ABCD organizational structure and communication framework. Dev Cogn Neurosci. 2018;32:8-15. doi: 10.1016/j.dcn.2018.04.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Garavan H, Bartsch H, Conway K, et al. Recruiting the ABCD sample: design considerations and procedures. Dev Cogn Neurosci. 2018;32:16-22. doi: 10.1016/j.dcn.2018.04.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hagler DJ, Hatton SN, Makowski C, et al. Image processing and analysis methods for the Adolescent Brain Cognitive Development Study. Bio Rxiv. Image processing and analysis methods for the Adolescent Brain Cognitive Development Study. Published November 2018. Accessed February 8, 2019. [DOI] [PMC free article] [PubMed]
- 46.Kobak K, Kratochvil C, Stanger C, Kaufman J. Computerized Screening of comorbidity in adolescents With Substance or Psychiatric Disorders. La Jola, CA: Anxiety Disorders and Depression; 2013. [Google Scholar]
- 47.Barch DM, Albaugh MD, Avenevoli S, et al. Demographic, physical and mental health assessments in the adolescent brain and cognitive development study: Rationale and description. Dev Cogn Neurosci. 2018;32:55-66. doi: 10.1016/j.dcn.2017.10.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.van der Ende J, Verhulst FC, Tiemeier H. Agreement of informants on emotional and behavioral problems from childhood to adulthood. Psychol Assess. 2012;24(2):293-300. doi: 10.1037/a0025500 [DOI] [PubMed] [Google Scholar]
- 49.Gordon EM, Laumann TO, Adeyemo B, Huckins JF, Kelley WM, Petersen SE. Generation and evaluation of a cortical area parcellation from resting-state correlations. Cereb Cortex. 2016;26(1):288-303. doi: 10.1093/cercor/bhu239 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Fischl B, Salat DH, Busa E, et al. Whole brain segmentation: automated labeling of neuroanatomical structures in the human brain. Neuron. 2002;33(3):341-355. doi: 10.1016/S0896-6273(02)00569-X [DOI] [PubMed] [Google Scholar]
- 51.Cox RW. AFNI: software for analysis and visualization of functional magnetic resonance neuroimages. Comput Biomed Res. 1996;29(3):162-173. doi: 10.1006/cbmr.1996.0014 [DOI] [PubMed] [Google Scholar]
- 52.Petersen AC, Crockett L, Richards M, Boxer A. A self-report measure of pubertal status: reliability, validity, and initial norms. J Youth Adolesc. 1988;17(2):117-133. doi: 10.1007/BF01537962 [DOI] [PubMed] [Google Scholar]
- 53.NIMH Data Archive. National database for autism research. https://ndar.nih.gov/study.html?id=500. Accessed January 25, 2019.
- 54.NIMH Data Archive. National database for autism research. https://ndar.nih.gov/study.html?id=576. Accessed January 25, 2019.
- 55.Van Dijk KRA, Hedden T, Venkataraman A, Evans KC, Lazar SW, Buckner RL. Intrinsic functional connectivity as a tool for human connectomics: theory, properties, and optimization. J Neurophysiol. 2010;103(1):297-321. doi: 10.1152/jn.00783.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Seibert TM, Brewer JB. Default network correlations analyzed on native surfaces. J Neurosci Methods. 2011;198(2):301-311. doi: 10.1016/j.jneumeth.2011.04.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Nielson DM, Pereira F, Zheng CY, et al. Detecting and harmonizing scanner differences in the ABCD study: annual release 1.0. Bio Rxiv. https://www.biorxiv.org/content/10.1101/309260v1. Published 2018. Accessed January 24, 2019.
- 58.Yu M, Linn KA, Cook PA, et al. Statistical harmonization corrects site effects in functional connectivity measurements from multi-site fMRI data. Hum Brain Mapp. 2018;39(11):4213-4227. doi: 10.1002/hbm.24241 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Johnson WE, Li C, Rabinovic A. Adjusting batch effects in microarray expression data using empirical Bayes methods. Biostatistics. 2007;8(1):118-127. doi: 10.1093/biostatistics/kxj037 [DOI] [PubMed] [Google Scholar]
- 60.Fortin J-P, Parker D, Tunç B, et al. Harmonization of multi-site diffusion tensor imaging data. Neuroimage. 2017;161:149-170. doi: 10.1016/j.neuroimage.2017.08.047 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc B. 1995;57(1):289-300. [Google Scholar]
- 62.Rouder JN, Speckman PL, Sun D, Morey RD, Iverson G. Bayesian t tests for accepting and rejecting the null hypothesis. Psychon Bull Rev. 2009;16(2):225-237. doi: 10.3758/PBR.16.2.225 [DOI] [PubMed] [Google Scholar]
- 63.Wagenmakers EJ, Wetzels R, Borsboom D, van der Maas HLJ. Why psychologists must change the way they analyze their data: the case of psi: comment on Bem (2011). J Pers Soc Psychol. 2011;100(3):426-432. doi: 10.1037/a0022790 [DOI] [PubMed] [Google Scholar]
- 64.Jefferys WH. Bayesian analysis of random event generator data. J Sci Explor. 1990;4(2):153-169. [Google Scholar]
- 65.Sadaghiani S, Scheeringa R, Lehongre K, Morillon B, Giraud A-L, Kleinschmidt A. Intrinsic connectivity networks, alpha oscillations, and tonic alertness: a simultaneous electroencephalography/functional magnetic resonance imaging study. J Neurosci. 2010;30(30):10243-10250. doi: 10.1523/JNEUROSCI.1004-10.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Schneider M, Leuchs L, Czisch M, Sämann PG, Spoormaker VI. Disentangling reward anticipation with simultaneous pupillometry/fMRI. Neuroimage. 2018;178:11-22. doi: 10.1016/j.neuroimage.2018.04.078 [DOI] [PubMed] [Google Scholar]
- 67.Martinussen R, Hayden J, Hogg-Johnson S, Tannock R. A meta-analysis of working memory impairments in children with attention-deficit/hyperactivity disorder. J Am Acad Child Adolesc Psychiatry. 2005;44(4):377-384. doi: 10.1097/01.chi.0000153228.72591.73 [DOI] [PubMed] [Google Scholar]
- 68.Berkson J. Limitations of the application of fourfold table analysis to hospital data. Biometrics. 1946;2(3):47-53. doi: 10.2307/3002000 [DOI] [PubMed] [Google Scholar]
- 69.Pan PM, Sato JR, Salum GA, et al. Ventral striatum functional connectivity as a predictor of adolescent depressive disorder in a longitudinal community-based sample. Am J Psychiatry. 2017;174(11):1112-1119. doi: 10.1176/appi.ajp.2017.17040430 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
eTable 1. Demographics of participants who passed the quality control for resting-state fMRI (2455 children)
eTable 2. Group differences between children with vs. without anhedonia in the resting-state fMRI connectivity z[r]
eTable 6. Group differences between children with vs. without anhedonia during reward anticipation [Large Reward > Neutral Cue] in the MID task-fMRI activation
eTable 10. Group differences between children with vs. without anhedonia during working-memory [Two-Back > Zero-Back] in the N-Back task-fMRI activation
eTable 5. Group differences between children with vs. without ADHD in the resting-state fMRI connectivity z[r]
eTable 3. Group differences between children with vs. without low mood in the resting-state fMRI connectivity z[r]
eTable 4. Group differences between children with vs. without Anxiety in the resting-state fMRI connectivity z[r]
eTable 9. Group differences between children with vs. without ADHD during reward anticipation [Large Reward > Neutral Cue] in the MID task-fMRI activation
eTable 13. Group differences between children with vs. without ADHD during working-memory [Two-Back > Zero-Back] in the N-Back task-fMRI activation
eTable 7. Group differences between children with vs. without low mood during reward anticipation [Large Reward > Neutral Cue] in the MID task-fMRI activation
eTable 8. Group differences between children with vs. without anxiety during reward anticipation [Large Reward > Neutral Cue] in the MID task-fMRI activation
eTable 11. Group differences between children with vs. without low mood during working-memory [Two-Back > Zero-Back] in the N-Back task-fMRI activation
eTable 12. Group differences between children with vs. without anxiety during working-memory [Two-Back > Zero-Back] in the N-Back task-fMRI activation
eTable 14. Group differences between anhedonia and low-mood during reward-anticipation [Large Reward > Neutral Cue] in the MID task-fMRI activation
eTable 15. Group differences between anhedonia and anxiety during reward-anticipation [Large Reward > Neutral Cue] in the MID task-fMRI activation
eTable 16. Group differences between anhedonia and ADHD during reward-anticipation [Large Reward > Neutral Cue] in the MID task-fMRI activation
eTable 17. Follow-up Analysis
eReferences.
Retracted article with errors highlighted
Replacement article with corrections highlighted