

Abstract
Glutamate responses in cultured rat astrocytes from cerebella of neonatal rats were investigated using the perforated-patch configuration to record membrane currents without rundown of intracellular messenger cascades, and microfluorometric measurements to measure the intracellular Ca2+ concentration ([Ca2+]i) and intracellular pH (pHi) with fura-2 AM and 2′,7′-bis-(2-carboxyethyl)-5,6-carboxyfluorescein acetoxy methylester respectively. In the perforated-patch mode, glutamate evoked single or multiple outward current transients in 82% of the cells, which disappeared when the recording technique was converted into a conventional whole-cell mode. The outward current transients were accompanied by [Ca2+]i transients, whereas pHi fell monophasically, without any sign of oscillation. Pharmacological analysis of the glutamate-induced responses indicated that ionotropic receptor activation evoked an inward current but no outward current transients, and metabotropic receptor activation (of the mGluR1/5 type) elicited outward current transients but no inward current. The outward current transients were reduced in frequency, or even abolished, after depletion of the intracellular Ca2+-stores by the Ca2+-ATPase inhibitor cyclopiaconic acid (10 μm). They reversed near −85 mV and were reduced by tetraethylammonium (10 mm), suggesting that they were caused by K+ channel activation. It is concluded that glutamate evoked these K+ outward current transients by oscillatory Ca2+ release mediated by mGluR activation. The corresponding membrane potential waves across the astroglial syncytium could provide spatial and temporal dynamics to the glial K+ uptake capacity and other voltage-dependent processes.
Keywords: glutamate, perforated patch-clamp, gramicidin, current oscillation, [Ca2+]i oscillations, Ca2+-activated K+ channels, rat
Changes in the intracellular Ca2+ concentration ([Ca2+]i) play a crucial role in many processes involved in the modulation of signal transduction, development, and plasticity in the CNS. At present, multiple pathways have been demonstrated that can lead to [Ca2+]i changes. The involvement of glutamate receptors appears to play a key role. The activation of glutamate receptors can change [Ca2+]iby two distinct mechanisms. First, receptors of the ionotropic subclass, namely NMDA and a subpopulation of Ca2+-permeable AMPA/kainate receptors, directly mediate a Ca2+ influx. Second, receptors of the metabotropic glutamate receptor subclass (mGluR), e.g., the mGluR1 and mGluR5, induce the hydrolysis of phosphatidylinositol that leads to the inositol triphosphate (IP3)-mediated release of Ca2+ from intracellular stores. In addition, the membrane depolarization mediated by ionotropic glutamate receptors could activate voltage-dependent Ca2+-channels that allow Ca2+ influx.
Glial cells express ionotropic glutamate receptors in culture (Bowman and Kimelberg, 1984; Kettenmann et al., 1984) and in situ(Berger et al., 1992; Müller et al., 1992; Seifert and Steinhäuser, 1995). In cultured cerebellar fusiform cells (Burnashev et al., 1992), in cerebellar Bergmann glial cells in situ (Müller et al., 1992), in hippocampal glial cells in the CA1 region of the stratum radiatum (Jabs et al., 1994), and in hilar glial precursor cells (Backus and Berger, 1995), evidence was found that the activation of AMPA/kainate receptors can also result in a Ca2+ influx.
Monophasic and multiple intracellular Ca2+transients were observed in cultured astrocytes after the application of glutamate receptor agonists (Glaum et al., 1990; Jensen and Chiu, 1990, 1991; de Barry et al., 1991; Holzwarth et al., 1994; Brune and Deitmer, 1995). The pharmacological profile of these responses revealed that both Ca2+ influx and intracellular Ca2+ release were responsible for the Ca2+ transients. Recent results suggested that glial cells might communicate with each other, or with neurons, using waves of Ca2+ elevation that spread via gap junctions through the astrocytic syncytium. Ca2+ waves were propagated between glutamate-stimulated cells (Cornell-Bell et al., 1990; Charles et al., 1991), indicating the occurrence of a long-range oscillatory signaling mechanism in glial cells. In hippocampal astrocytes, waves of [Ca2+]i could be induced by the activity of neighboring neurons (Dani et al., 1992). Focal electrical stimulation of astrocytes in mixed cultures of rat forebrain cells induced intercellular Ca2+ waves and large increases in [Ca2+]i in neighboring neurons, suggesting the existence of glial–neuronal signaling pathways (Nedergaard, 1994).
Astrocytes express a large repertoire of voltage- and ligand-gated ion channels, e.g., Ca2+-activated K+channels (Quandt and MacVicar, 1986) or GABAA-receptors (Stelzer et al., 1988), which are sensitive to changes in [Ca2+]i. Because the cytosol is dialyzed rapidly during conventional whole-cell clamp recordings, we used the perforated patch-clamp technique to characterize the membrane responses evoked by glutamate receptor agonists. In combination with the fluorescent imaging technique to measure [Ca2+]i and pHi, we could show in single cells that the application of agonists of glutamate receptors evoked oscillations of membrane currents associated with multiple [Ca2+]i transients.
A preliminary report of some of the results has been published previously in abstract form (Chen et al., 1997).
MATERIALS AND METHODS
Cell culture. For preparing primary cultured astrocytes, the cerebellar hemispheres of newborn rats (P0–P1) were rapidly removed. Cells were isolated and cultured according to the method described by Fischer (1984). Immunohistochemical staining confirmed that the cultures consisted of enriched (>95%) glial fibrillary acidic protein-positive astrocytes (Brune et al., 1994). After the cells had reached confluence, oligodendrocytes and macrophages were removed by a shaking procedure, and the remaining cells were dissociated, plated on glass coverslips coated with poly-d-lysine, and incubated in 7% CO2 at 37°C. The experiments were performed between 2 and 15 d after plating of the astrocytes at room temperature (∼22–24°C).
Experimental setup. For perforated patch-clamp recordings, the culture dish was mounted on the stage of an inverted microscope (Zeiss). For the combined electrophysiological and microfluorometric experiments, the culture dish was mounted on an inverted fluorescence microscope (Diaphot, Nikon, Tokyo, Japan) equipped with a Nikon CF-Fluor 20× objective. Recording patch pipettes were pulled from borosilicate glass, and the tips were fire-polished (resistance 4–6 MΩ, when filled with the pipette solution). Gramicidin–perforated-patch recordings were performed following the method of Kyrozis and Reichling (1995). Briefly, the electrode tip was filled for 1–40 sec with the gramicidin-free pipette solution (see below) to avoid problems with seal formation, and then back-filled with the gramicidin-containing pipette solution. After the formation of a gigaseal (on-cell recording) (Hamill et al., 1981), short steps in holding potential were applied continuously at 2 min intervals to monitor the gradual decrease in series resistance. Drug application was not started until the series resistance decreased below 50 MΩ, which usually lasted 20–30 min. The reference potential for all measurements was the zero-current potential of the pipette in the bath before establishment of the gigaseal.
Currents were recorded by an EPC-7 (List, Darmstadt, Germany) patch-clamp amplifier. Before digitization (sampling rate 0.5–1 kHz), currents were filtered at 3 kHz with a three-pole low-pass Bessel filter. Data were stored and evaluated with the aid of the PCLAMP hardware and software package (Axon Instruments, Foster City, CA) for a personal computer. To determine the agonist-induced current amplitudes, the maximal deflection from the baseline was used. The difference of two samples was tested using the two-tailed t test.
Microfluorometric recordings. The fluorescence microscope was equipped with a dual excitation fluorometric imaging system (PTI, Wedel, Germany). The illumination was generated by a 75 W xenon bulb. Monochromator settings, chopper frequency, and complete data acquisition were controlled by software (PTI) for microcomputer systems. Astrocytes were loaded by incubating the cell cultures in 2′,7′-bis-(2-carboxyethyl)-5,6-carboxyfluorescein-acetoxy-methylester (BCECF-AM; 25 μm) for 20 min or fura-2 AM (5 μm) for 60 min at room temperature. Dye-loaded astrocytes were excited by monochromatic light at wavelengths 350 and 380 nm (fura-2 AM) to measure [Ca2+]i or at 440 and 495 nm (BCECF-AM) to measure pHi. The fluorescence emission of the cell under perforated-patch control, of another single cell, and the emission of a selected area with a group of 5–10 cells on the coverslip was recorded simultaneously with a video camera (SIT C-2400, Hamamatsu, Garching, Germany), using a 495 nm longpass filter for fura-2 AM and a 520 nm longpass filter for BCECF-AM. The signals were sampled at 3 Hz and computed into relative ratio units. Drug-induced changes of [Ca2+]i were measured by determining the changes of the fluorescence ratio of 350 nm: 380 nm. The BCECF-AM ratio of 440 nm: 495 nm was converted into pH units according to calibration described by Brune et al. (1994).
Solutions and drug application. The pipette solution for perforated patch-clamp recordings contained (in mm): KCl 140, NaCl 5, CaCl2 0.5, MgCl2 1, EGTA 5, HEPES 10; pH adjusted to 7.2 with KOH. Gramicidin (Sigma, St. Louis, MO), 5 mg/ml, was dissolved in dimethylsulfoxide (DMSO), vortexed for 1 min, sonicated for 20 sec, and then added to the pipette solution to give a final concentration of 25–50 μg/ml. In some cells the conventional whole-cell configuration was established after recording in the gramicidin–perforated-patch configuration. This resulted in a dialysis of the cell interior with the pipette solution listed above.
During the experiments the cell cultures were superfused continuously with a HEPES-buffered saline containing (in mm): NaCl 145, KCl 5, CaCl2 2, MgCl2 1, glucose 10, HEPES 10; pH adjusted to 7.4 with NaOH. In a few experiments we used a Ca2+-free salt solution in which CaCl2was replaced by EGTA (1 mm). Stock solutions of glutamate, quisqualate (100 mm in aqua bidest; both from Sigma), and kainate (100 mm in 100 mm NaOH; Sigma) were prepared. Stock solutions of cyclopiazonic acid (CPA, 50 mm; Sigma) and 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX, 50 mm; Tocris Cookson, Bristol, UK) were dissolved in DMSO and stored at −20°C. Kynurenic acid (Sigma) was dissolved in the saline in the final concentration. Stock solutions oftrans-(±)-1-aminocyclopentane-1S,3R-dicarboxylic acid (t-ACPD; 20 mm in 50 mm NaOH), (s) 3,5 dihydroxyphenyl-glycine (DHPG; 100 mm in aqua bidest), andl-2-amino-4-phosphonobutyric acid (l-AP4; 20 mm in 50 mm NaOH) were prepared. t-ACPD, DHPG, and l-AP4 were purchased from Tocris Cookson. All other drugs were obtained from Sigma. Drugs were added to the saline shortly before use in defined concentrations. Fura-2 AM and BCECF-AM were obtained from Molecular Probes (Eugene, OR).
RESULTS
Glutamate evokes membrane current oscillations
The whole-cell membrane current of cultured cerebellar astrocytes was recorded in the gramicidin–perforated-patch configuration at a holding potential (Vh) of −70 mV. In these cells 1 mm glutamate, applied by bath superfusion for 10 sec, evoked different current response patterns (Fig.1). All of them showed a distinct inward current (100% of the cells; n = 62), but they differed in the frequency, shape, and time course of oscillating outward current transients (Fig. 1A–C, E) that were superimposed in most cells (82%; n = 51).
Fig. 1.

Glutamate-induced current response patterns in cultured cerebellar astrocytes. A–C, The membrane current was recorded in the gramicidin–perforated-patch configuration (PPC), and glutamate (Glu, 1 mm) was applied by bath application for 10 sec as indicated. Glutamate evoked an inward current that was superimposed by multiple (A), a single (B), or no outward current transients (C).D, In the conventional whole-cell clamp configuration (WCC), glutamate exclusively induced an inward current without showing any outward current transients. E, Gramicidin–perforated-patch recording that demonstrates the persistency of outward current transients in time.
In 29 cells, multiple outward current transients (2–8) (Fig.1A,E) were observed after glutamate application. Although the first outward current transient usually had the largest amplitude (41 ± 25 pA; mean ± SD), the subsequent transients showed a continuous but slow decrease in amplitude with time. When the glutamate application was prolonged until up to 3 min, the outward current transients persisted for several minutes (data not shown). In some cells (16 of 62), only a single outward current transient with a mean amplitude of 22 ± 14 pA was superimposed onto the rising phase of the inward current component (Fig.1B). No outward current transients were present in 11 of 62 cells, i.e., in 18% of the astrocytes (Fig. 1C). A few astrocytes (n = 6) showed a mixed response where single and multiple outward current transients could change from one to the next glutamate application and vice versa. The presence of multiple current oscillations in astrocytes showed a remarkable persistency, because they could be observed for up to 1 hr without any major changes in shape, number, and amplitude of the current transients (Fig.1E).
Membrane current oscillations induced by glutamate, however, could be observed only in the gramicidin–perforated-patch configuration. Astrocytes exposed to glutamate (1 mm; 10 sec) in the conventional whole-cell clamp configuration responded with an inward current of 28 ± 7.5 pA (n = 35) (Fig.1D) that was significantly (p< 0.01) larger than that recorded in the gramicidin–perforated-patch configuration but never showed any outward current oscillations. In eight cells that showed outward current transients after glutamate application in the gramicidin–perforated-patch configuration, the membrane patch was ruptured carefully to establish the conventional whole-cell clamp configuration. After this procedure the glutamate-induced outward current transients disappeared completely (Figs. 2, 3, top traces), indicating the requirement of a functionally intact cytosol for the occurrence of current oscillations in astrocytes as revealed by the gramicidin–perforated-patch configuration.
Fig. 2.

Simultaneous recording of membrane current and [Ca2+]i using the digital imaging technique in fura-2 AM-loaded cerebellar astrocytes showing multiple outward current transients. Left column, Gramicidin–perforated-patch configuration: glutamate (Glu, 1 mm), applied for 10 sec as indicated, induced an inward membrane current that was superimposed by three outward current transients (top). Each outward current transient was preceded by a transient increase in [Ca2+]i (middle). Thebottom trace represents the averaged change in [Ca2+]i induced by glutamate recorded from a group of cells in the vicinity of the cell displayed above.Right column, Responses of the same cells to glutamate after establishment of the conventional whole-cell clamp configuration.
Fig. 3.

Simultaneous recording of membrane current and [Ca2+]i using the digital imaging technique in cerebellar astrocytes showing a single outward current transient. Left column, Gramicidin–perforated-patch configuration: glutamate (Glu, 1 mm), applied for 10 sec as indicated, induced an inward membrane current that was superimposed by a single outward current transient (top). A transient increase in [Ca2+]i was followed by a slowly decaying [Ca2+]i decrease (middle). The bottom trace represents the averaged change in [Ca2+]i induced by glutamate recorded from a group of cells in the vicinity of the cell displayed above. Right column, Responses of the same cells to glutamate after establishment of the conventional whole-cell clamp configuration.
Simultaneous measurement of [Ca2+]i, pHi, and membrane current oscillations
Recently it was reported that the application of glutamate on astrocytes could evoke transient and complex oscillatory changes in [Ca2+]i (Cornell-Bell et al., 1990;Glaum et al., 1990; Jensen and Chiu, 1990, 1991; de Barry et al., 1991;Dani et al., 1992; Holzwarth et al., 1994; Brune and Deitmer, 1995) and considerable increases in the intracellular H+concentration (Brune and Deitmer, 1995). Because these experiments were performed exclusively in intact cells, we supposed that the glutamate-induced current oscillation we observed in the gramicidin–perforated-patch configuration could be mediated by changes in [Ca2+]i or pHi, or both. To examine this issue, the changes in [Ca2+]i or pHi were recorded simultaneously with the membrane current in the perforated-patch and conventional whole-cell mode.
Glutamate (1 mm) evoked a large inward current that was superimposed by a number of distinct outward current transients (Fig.2). In the same cell we observed an equal number of transient [Ca2+]i changes that were temporally associated with the outward current transients. In addition, a small increase in basal [Ca2+]i was observed that followed roughly the time course of the deactivating inward current component. In cells that responded only with a single outward current transient, only a single large Ca2+transient was also apparent (Fig. 3). When the whole-cell clamp configuration was subsequently established in the same cell (n = 4), both outward current and multiple Ca2+ transients disappeared, leaving a strongly reduced single initial Ca2+ transient and a small tonic increase in basal [Ca2+]i(Figs. 2, 3, top right and middle right traces). Also in two cells in which glutamate failed to induce current oscillations in the perforated-patch configuration, no Ca2+ transients were observed.
Changes in [Ca2+]i in a group of neighboring astrocytes, which were not in contact with the patch pipette, are shown in the bottom traces of Figures 2 and 3. In these cells glutamate also induced changes in [Ca2+]i, but oscillations were not observed, indicating that the fura-2 AM fluorescence signal reflected the integrated response of many cells that were not producing oscillations in phase. Rather, these presumed [Ca2+]i oscillations showed up as an initial transient and a sustained rise for >1 min (Fig. 2) or a monophasic [Ca2+]i rise (Fig. 3).
A small decrease in pHi could be measured in single astrocytes in which 1 mm glutamate evoked outward current transients, but pH oscillations were never observed (Fig.4). Similar results were obtained in single astrocytes or in groups of “unpatched” cells (n = 8), regardless of whether they showed multiple or single outward current transients (Fig. 4). These results suggested that there was probably no coupling between changes in pHiand the different current and [Ca2+]iresponse patterns in these astrocytes.
Fig. 4.

Simultaneous recording of membrane current in the gramicidin–perforated-patch configuration and pHi using the digital imaging technique in BCECF-AM-loaded cerebellar astrocytes.Top, Current responses induced by glutamate (Glu, 1 mm, applied for 10 sec as indicated) of an astrocyte showing multiple outward current transients (A) and another showing a single outward current transient (B). Second row from top, Corresponding glutamate-induced pHi changes to the current traces on top. Third row from top, Simultaneous measurement of the glutamate-induced pHichange in another single cell in the vicinity that was not under perforated-patch control. Bottom, Simultaneous measurement of the glutamate-induced pHi change averaged over a group of cells in the vicinity. A decrease in pHi is displayed as an upward deflection.
Effects of glutamate receptor ligands
The absence of glutamate-induced outward current transients and [Ca2+]i oscillations in the whole-cell clamp configuration suggested the involvement of an intracellular messenger system, which was dialyzed in the conventional whole-cell mode but persisted in the perforated patch-clamp mode. Because we had observed a correlation in time between [Ca2+]i oscillations and outward current transients, we attempted to identify the glutamate receptor subtype and the mechanism underlying the membrane current oscillations by recording the membrane current in the gramicidin–perforated-patch configuration. The effects of different glutamate receptor ligands on [Ca2+]i and pHi in rat astrocytes had been studied previously (de Barry et al., 1991;Holzwarth et al., 1994; Brune and Deitmer, 1995).
Glutamate is a mixed agonist at both ionotropic receptors and mGluRs. Therefore, we tested separately an involvement of ionotropic and metabotropic receptors in the induction of current oscillations. Kainate (400 μm, 10 sec), an agonist of the non-NMDA glutamate receptor subtype, evoked an inward current, but was unable to elicit outward current oscillations as did glutamate in the same cell (n = 5) (Fig.5A). Kynurenic acid (1 mm; n = 4) (Fig. 5B), a broad-spectrum glutamate antagonist of ionotropic receptors (Perkins and Stone, 1982), and CNQX (50 μm; n = 4) (Fig. 5C), a selective blocker of the AMPA/kainate receptor subtype (Honoré et al., 1988), reduced the glutamate-evoked inward current but failed to block the glutamate-induced outward current transients (Fig. 5B,C).
Fig. 5.
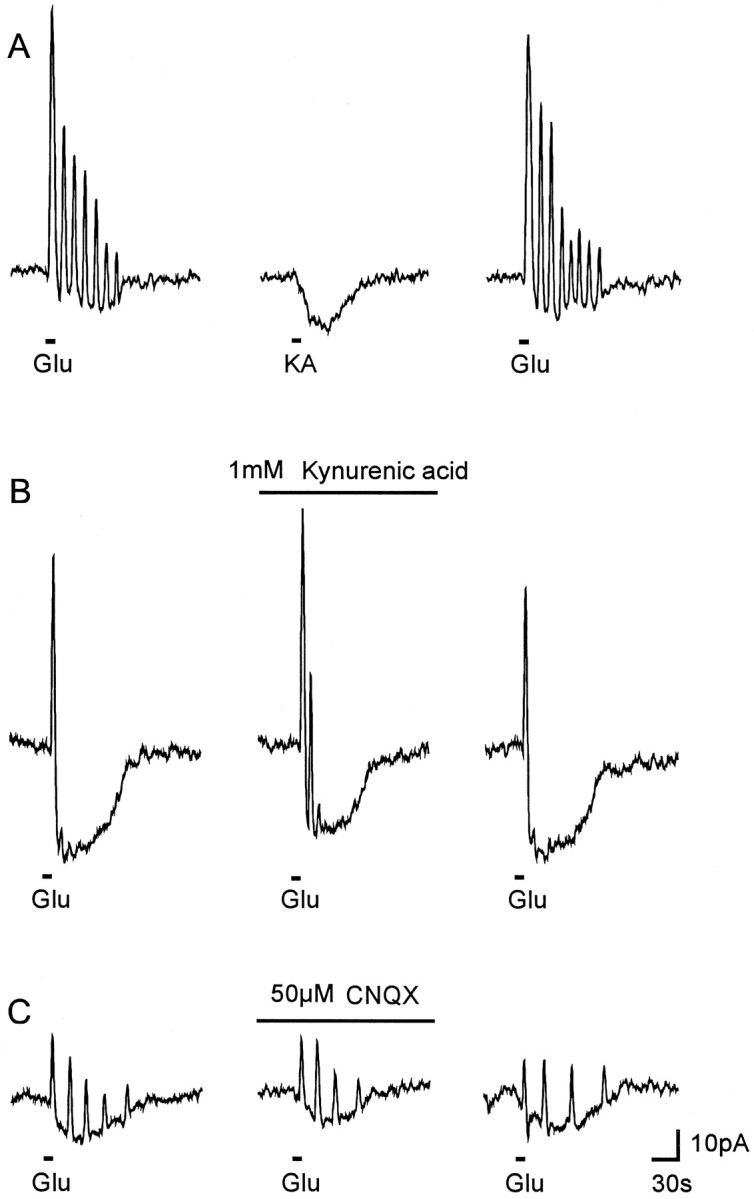
Effect of ionotropic glutamate receptor ligands.A, Kainate (KA, 400 mm, applied for 10 sec as indicated) did not elicit any current oscillation. B, Kynurenic acid (1 mm, preapplication time: 5 min), and C, CNQX (50 μm, preapplication time: 5 min) reduced the glutamate-induced inward current but not the outward current transients.
Quisqualate, which is known to be an agonist of AMPA/kainate as well as of mGluRs, evoked outward current transients but no detectable inward current when applied in a concentration of 200 μm for 10 sec (n = 5) (Fig.6A). The metabotropic agonist t-ACPD (30 μm), an agonist reported to activate receptors of the mGluR2/3 subtype but that also exerted weak agonistic activity at mGluR1/5 receptors (Nakanishi, 1992; Tanabe et al., 1993), failed to evoke an inward current but elicited one or multiple outward current transients (n = 5) (Fig. 6B). DHPG (100 μm), an agonist reported to be selective for mGluRs including the mGluR1/5 subtypes, also mimicked the glutamate-evoked outward current transients (n = 5) (Fig. 6C). The agonist of receptors of the mGluR4/6–8 subtypes, l-AP4 (Nakanishi, 1992; Tanabe et al., 1993), however, had no effect on the membrane current in cultured astrocytes when applied at 200 μm for 10 sec (n = 5) (Fig. 6D). These results indicate that the outward current transients evoked by glutamate were attributable to the activation of mGluRs. The pharmacological profile of the glutamate-evoked outward current transients suggested the activation of mGluRs of the mGluR1/5 or mGluR2/3 subtypes.
Fig. 6.

The effect of metabotropic glutamate receptor agonists. A, Quisqualate (QQ, 200 μm, applied for 10 sec as indicated), B, t-ACPD (30 μm, 10 sec), and C, DHPG (100 μm, 10 sec) evoked outward current transients but no detectable inward currents. D, l-AP4 (200 μm, 10 sec) had no effect on the membrane current of cultured cerebellar astrocytes.
Contribution of intracellular Ca2+ mobilization
Glutamate receptors of the mGluR1/5 subtype are coupled via a G-protein to the hydrolysis of phosphatidylinositol, which results in the production of IP3 and the subsequent release of Ca2+ from IP3-sensitive intracellular stores (Berridge and Irvine, 1989). To test whether an intracellular Ca2+ release was involved in the activation of the glutamate-evoked outward current transients, intracellular Ca2+ stores were depleted using CPA (10 μm), which was preapplied for 5 min and then in combination with glutamate (1 mm; n = 5). CPA alone elicited a transient [Ca2+]irise, indicating reversible loss of Ca2+ from intracellular stores (n = 6).
In the presence of CPA the outward current transients evoked by glutamate were strongly reduced, usually to a single transient (Fig.7A), or when it was preapplied for 10 min, they were totally abolished (Fig. 7B). The inward current component, however, remained unaffected by CPA (Fig. 7). These results indicate that the current oscillations were likely mediated by the release of Ca2+ from intracellular, CPA-sensitive stores.
Fig. 7.

Effect of CPA on glutamate-induced currents in cultured cerebellar astrocytes. CPA (10 μm, preapplied for 5 min) inhibited the outward current transients induced by glutamate (1 mm, applied for 10 sec as indicated) in cells responding with multiple oscillations (A) or with a single oscillation (B), but had no effect on the inward current component.
The contribution of an influx of extracellular Ca2+to the glutamate-induced current oscillations was examined by comparing the current responses evoked in standard saline with those in a Ca2+-free saline that contained 1 mmEGTA. The initial outward current transient observed in standard salt solution was still prominent when glutamate was applied in Ca2+-free salt solution (n = 8; not shown). In cells expressing multiple current oscillations, however, no consistent effect of the Ca2+-free salt solution was observed. Usually the oscillatory outward current transients were reduced or disappeared completely (n = 6). In these cells, readdition of extracellular Ca2+ did not result in a recovery of outward current oscillations. The results suggest that the sustained outward current transients required the presence of extracellular Ca2+ to continue, but the large initial transient apparently did not depend on Ca2+ influx and was presumably caused by Ca2+ release from intracellular stores.
Identification of the outward current transients
To identify the type of ion channel underlying the glutamate-induced outward current transients we determined their reversal potential. Therefore, 1 mm glutamate was applied when Vhr was altered to more hyperpolarized values during the glutamate-induced inward current. As shown in Figure8A,B, the outward current transients decreased or reversed their polarity at a holding potential of −90 or −100 mV. The current amplitudes were fitted by linear regression to yield a rough estimate of the reversal potential of these outward current transients (Fig. 8C). The resulting reversal potential of −84.9 mV (n = 4) was close to the calculated K+ equilibrium potential (EK+ = −83 mV), indicating the involvement of K+ channels during the glutamate-evoked current transients.
Fig. 8.
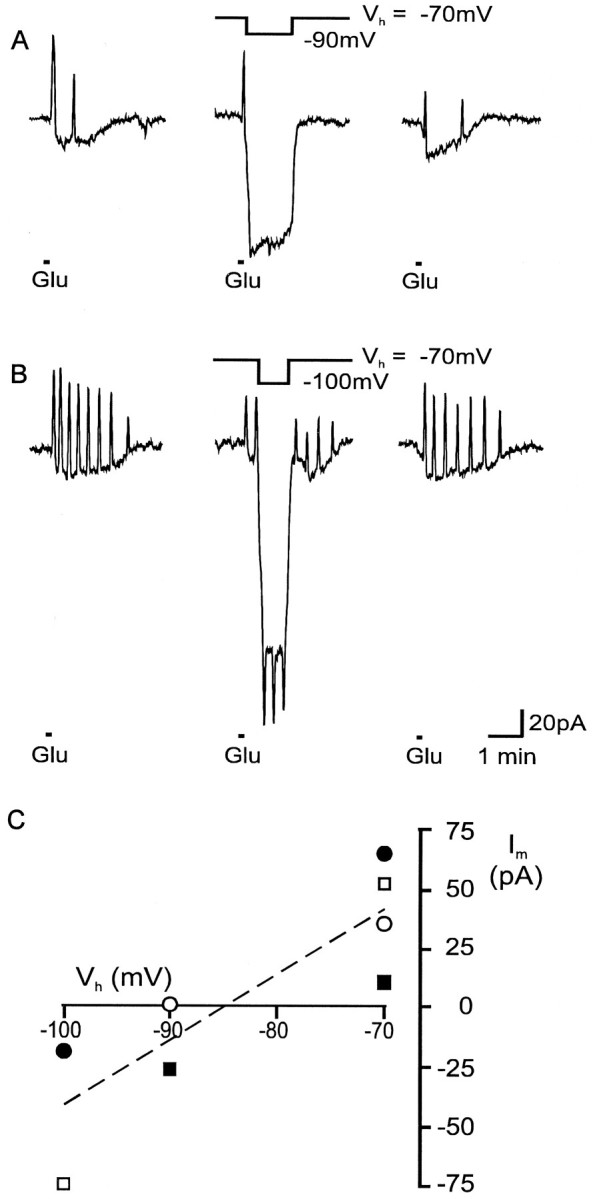
Determination of the reversal potential of glutamate-evoked outward current transients. When the holding potential was changed during a glutamate-induced current response from −70 to −90 mV (A) or from −70 to −100 mV (B) as indicated, the outward current transients reversed their polarity. C, The current amplitude of glutamate-induced current oscillations was plotted as a function of the membrane potential. To estimate the reversal potential of the current transients, all data points were fitted by linear regression (reversal potential: −84.9 mV; n = 4; the differentsymbols represent the different cells).
This was confirmed by the sensitivity to tetraethylammonium (TEA) of the outward current transients. TEA (10 mm;n = 5), a K+ channel blocker (Rudy, 1988), reduced the outward current transients evoked by glutamate in cells showing multiple oscillations (Fig.9A) and also in those with a single outward current transient (Fig. 9B).
Fig. 9.

The effect of TEA on glutamate-induced outward current transients. TEA (10 mm, preapplied for 5 min) reduced the glutamate-induced outward current transients in cerebellar astrocytes expressing multiple current oscillations (A) as well as a single current oscillation (B), indicating that they were mediated by K+ channels.
DISCUSSION
Perforated-patch recordings reveal glutamate-induced current oscillations
To minimize the washout of regulatory molecules that could modulate ion channel activity (Korn et al., 1991), we have used perforated-patch recordings with gramicidin, which forms pores selective for monovalent cations but impermeable for anions and divalent cations (Myers and Haydon, 1972). The use of this technique revealed that glutamate receptor agonists evoked an inward current that was superimposed by a single or multiple outward current transients. Many preceding studies on glutamate receptors in glial cells in culture (Sontheimer et al., 1988; Usowicz et al., 1989; Wyllie and Cull-Candy, 1994) and in brain slices (Berger et al., 1992; Müller et al., 1992; Jabs et al., 1994; Backus and Berger, 1995; Seifert and Steinhäuser, 1995) missed the current oscillations, presumably because of the use of the conventional whole-cell clamp configuration. In another study in which the perforated-patch technique was used in hilar glial precursor cells, Ca2+-dependent K+ channels were observed after Ca2+ influx through Ca2+-permeable AMPA/kainate receptors (Backus et al., 1995).
Identification of the mGluR subtype
The application of the ionotropic agonist kainate evoked an inward current but no superimposed outward current transients, whereas the mixed agonist quisqualate and agonists selective for mGluR subtypes, DHPG and t-ACPD, induced outward current transients but no inward current component. These findings, together with the lack of effect of blockers of ionotropic glutamate receptors, kynurenic acid and CNQX, indicate that the outward current transients were caused by the activation of mGluRs. The inward current component was evoked only by glutamate or kainate, in either conventional or perforated patch-clamp mode, but not by t-ACPD, DHPG, or l-AP4, indicating that it was mediated by ionotropic glutamate receptors, most likely of the AMPA/kainate subtype, which are expressed in astrocytes in culture (Sontheimer et al., 1988; Backus et al., 1989; Usowicz et al., 1989;Burnashev et al., 1992; Wyllie and Cull-Candy, 1994; Telgkamp et al., 1996) and in brain slices (Berger et al., 1992; Müller et al., 1992; Jabs et al., 1994; Backus and Berger, 1995; Seifert and Steinhäuser, 1995).
The pharmacological profile of mGluR subtypes (Nakanishi, 1992; Tanabe et al., 1993; Pin and Duvoisin, 1995) suggests that the induction of outward current transients by glutamate is mediated by the mGluR1/5 subtypes, because their selective agonist DHPG was effective. t-ACPD, an agonist of mGluR2/3 and mGluR1/5 subtypes (Pin and Duvoisin, 1995), was also effective. The selective mGluR4/6–8 agonist l-AP4 did not induce any response. Because the depletion of intracellular Ca2+-stores by CPA led to a reduction of outward current transients, it is likely that the mGluR subtype involved in the mechanism underlying the outward current oscillations is coupled to intracellular Ca2+ release. At present, mGluR1/5 receptors are the only mGluR subtypes that are linked to the IP3-mediated intracellular Ca2+mobilization (Abe et al., 1992; Aramori and Nakanishi, 1992). Therefore, mGluR1/5 receptors are likely involved in the induction of these outward current oscillations.
Glutamate-induced [Ca2+]i changes and oscillations
Glial cells respond to various neurotransmitters with changes of [Ca2+]i. Monophasic changes in [Ca2+]i are caused by an elevation of the extracellular K+ concentration or by the activation of ionotropic receptor agonists, e.g., kainate (Jensen and Chiu, 1990; Salm and McCarthy, 1990; Brune and Deitmer, 1995) or GABA (Kirchhoff and Kettenmann, 1992). These responses are thought to be caused by an influx of Ca2+ through Ca2+-permeable ionotropic receptors or voltage-dependent channels (Finkbeiner, 1995). Polyphasic responses evoked by glutamate receptor agonists are characterized by sustained [Ca2+]i oscillations preceded by an initial transient [Ca2+]i rise that is caused primarily by a release of Ca2+ from IP3-sensitive intracellular stores (cf. Finkbeiner, 1995;Verkhratsky and Kettenmann, 1996).
Gramicidin–perforated-patch recordings in combination with fura-2 AM digital imaging disclosed several types of [Ca2+]i changes in cultured cerebellar astrocytes. In a subpopulation of cells, polyphasic changes in [Ca2+]i were prominent and were characterized by two to eight transient rises in [Ca2+]i and a simultaneous increase in the [Ca2+]i baseline. In a small group of cells (<10) glutamate application resulted in an initial transient rise of [Ca2+]i that was followed by a sustained plateau reflecting the averaged oscillatory response of a group of cells that closely resembled the response pattern reported recently for a large group of cells (>20) (Brune and Deitmer, 1995). The nonoscillatory increase in baseline Ca2+ might be caused by a Ca2+ influx through Ca2+-permeable AMPA receptors that are expressed in cultured rat cerebellar astrocytes (Telgkamp et al., 1996).
In another subpopulation of cerebellar astrocytes, glutamate induced an initial transient rise that was followed by a plateau without showing distinct oscillations (Fig. 3). In these cells the initial increase in [Ca2+]i might have been insufficient to induce subsequent oscillations; however, the response type of glutamate-induced [Ca2+]i changes strongly depends on the mGluR subtype. In stably transfected cell lines expressing recombinant receptors of the mGluR1α subtype, monophasic changes in [Ca2+]i were observed, whereas in cells expressing the mGluR5a subtype, oscillatory responses with an increasing number of [Ca2+]itransients were found (Kawabata et al., 1996). Therefore, different populations of cerebellar astrocytes might exist: one population expressing primarily the mGluR1α subtype and another the mGluR5a subtype.
Glutamate-induced changes in pHi
Intracellular acidifications induced by glutamate were small and non-oscillatory and therefore could not be correlated to either the current or the [Ca2+]i oscillations. The pHi measurements might have been impaired by the H+ permeability of the gramicidin pores (Myers and Haydon, 1972), which could have caused a dampening of the pHi changes. The mechanisms of this acidification are discussed elsewhere (Brune and Deitmer, 1995; Rose and Ransom, 1996;Deitmer and Schneider, 1997).
Is there a link between [Ca2+]iand outward current oscillations?
The [Ca2+]i oscillations observed in the astrocytes likely induced the outward current oscillations, because (1) the number of Ca2+ oscillations always exactly matched the number of outward current transients (Fig. 2) and (2) the Ca2+ oscillations always started shortly before the corresponding outward current transient, suggesting that an increase of [Ca2+]i directly caused the current transients. Because the outward current transients were blocked by TEA and reversed close to the estimated K+ equilibrium potential, we conclude that they were mediated by Ca2+-dependent K+channels.
The withdrawal of extracellular Ca2+ did not affect the initial outward current transient in most astrocytes, suggesting that it was mediated by an intracellular Ca2+release. In addition, maintained oscillatory outward current transients required the presence of extracellular Ca2+. Recent findings have indeed shown that the initial transient rise in [Ca2+]i was mediated by an intracellular Ca2+ release and that the sustained [Ca2+]i oscillations required the presence of extracellular Ca2+ (Cornell-Bell et al., 1990; Glaum et al., 1990; Jensen and Chiu, 1990; de Barry et al., 1991;Holzwarth et al., 1994; Kim et al., 1994; Brune and Deitmer, 1995). In contrast, nonglutamate-evoked oscillations in [Ca2+]i also persisted in the absence of extracellular Ca2+ (mechanical stimulation:Charles et al., 1991; P2Y-purinergic stimulation:Kastritsis et al., 1992); however, it cannot be discounted that the reduction of oscillatory outward current transients was attributable to a depletion of intracellular Ca2+ stores caused by the withdrawal of extracellular Ca2+.
Kainate did not induce current oscillations, although it did evoke distinct changes in [Ca2+]i in astrocytes (Enkvist et al., 1989; Jensen and Chiu, 1990, 1991;Holzwarth et al., 1994; Brune and Deitmer, 1995). A monophasic increase in [Ca2+]i, however, would give rise to a monophasic Ca2+-activated K+ current. Therefore, the kainate-induced current represents the sum of a large inward current through kainate-activated AMPA receptor/channels and a K+ outward current. Perforated-patch recordings in hilar glial precursor cells in situ in acute hippocampal slice preparations have shown that kainate could also induce biphasic current responses that were composed of an AMPA receptor/channel-mediated inward current and a subsequent delayed Ca2+-activated K+ current (Backus et al., 1995).
This is the first report of glutamate-induced current oscillations in glial cells. Glutamate-induced depolarizations of the membrane potential have been reported previously in cultured rat brain astrocytes (Bowman and Kimelberg, 1984; Kettenmann et al., 1984) and were attributed to the activation of non-NMDA receptors (Backus et al., 1989). In these studies the changes in membrane potential were measured with sharp intracellular microelectrodes, which should not affect [Ca2+]i transients. Although our experimental conditions are comparable, none of these studies has shown any oscillatory changes in membrane potential after glutamate application. In cultured cortical rat astrocytes, however, quisqualate induced biphasic changes in membrane potential (compare Fig. 1C of Backus et al., 1989), characterized by an initial transient hyperpolarization and a delayed depolarization mediated by non-NMDA receptors. Moreover, oscillations of the membrane potential were observed in glial cells of rat hippocampal slices (Walz and MacVicar, 1986) and in astrocytes of kainic acid-lesioned hippocampal slices after the application of a phorbol ester (MacVicar et al., 1987).
Oscillatory changes in [Ca2+]i can spread across the glial syncytium like waves (cf. Finkbeiner, 1995) and thus by activation of Ca2+-activated K+ channels could give rise to wave-like changes of the glial membrane potential. In astrocytes such potential waves could arise at sites of increased neuronal activity, where an elevation of the extracellular K+ concentration and the exposure to glutamate depolarize the membrane potential and evoke an increase in [Ca2+]i. K+ uptake into the astrocytic syncytium could travel in phase with a hyperpolarizing potential wave, thus providing spatial dynamics to the glial K+ buffer capacity.
Footnotes
This study was supported by a grant of the Deutsche Forschungsgemeinschaft to J.W.D. and K.H.B. (De 231/11-1). We thank C. Lohr for critically reading this manuscript. We are grateful to Sandra Bergstein for her excellent technical assistance.
Reprint requests should be addressed to Dr. J. W. Dietmer, Abteilung für Allgemeine Zoologie, FB Biologie, Universität Kaiserslautern, Postfach 3049, D-67653 Kaiserslautern, Germany.
Dr. Chen’s present address: II. Physiologisches Institut de Universität Heidelberg, Im Neuenheimer Feld 326, D-69120 Heidelberg, Germany.
REFERENCES
- 1.Abe T, Sugihara H, Nawa H, Shigemote R, Mizuno N, Nakanishi S. Molecular characterization of a novel metabotropic glutamate receptor mGluR5 coupled to inositol phosphate/Ca2+ signal transduction. J Biol Chem. 1992;267:13361–13368. [PubMed] [Google Scholar]
- 2.Aramori I, Nakanishi S. Signal transduction and pharmacological characteristics of a metabotropic glutamate receptor, mGluR1, in transfected CHO cells. Neuron. 1992;8:757–765. doi: 10.1016/0896-6273(92)90096-v. [DOI] [PubMed] [Google Scholar]
- 3.Backus KH, Berger T. Developmental variation of the Ca2+ permeability of AMPA receptors in hilar glial precursor cells. Pflügers Arch. 1995;431:244–252. doi: 10.1007/BF00410197. [DOI] [PubMed] [Google Scholar]
- 4.Backus KH, Kettenmann H, Schachner M. Pharmacological characterization of the glutamate receptor in cultured astrocytes. J Neurosci Res. 1989;22:274–282. doi: 10.1002/jnr.490220307. [DOI] [PubMed] [Google Scholar]
- 5.Backus KH, Elsen FP, Berger T. Secondary effects of kainate in glial cells. Verh Dtsch Zool Ges. 1995;88.1:3. [Google Scholar]
- 6.Berger T, Walz W, Schnitzer J, Kettenmann H. GABA- and glutamate-activated currents in glial cells of the mouse corpus callosum slice. J Neurosci Res. 1992;31:21–27. doi: 10.1002/jnr.490310104. [DOI] [PubMed] [Google Scholar]
- 7.Berridge MJ, Irvine RF. Inositol phosphates and cell signaling. Nature. 1989;341:197–205. doi: 10.1038/341197a0. [DOI] [PubMed] [Google Scholar]
- 8.Bowman CL, Kimelberg HK. Excitatory amino acids directly depolarize rat brain astrocytes in primary culture. Nature. 1984;311:656–659. doi: 10.1038/311656a0. [DOI] [PubMed] [Google Scholar]
- 9.Brune T, Deitmer JW. Intracellular acidification and Ca2+ transients in cultured rat cerebellar astrocytes evoked by glutamate agonists and noradrenaline. Glia. 1995;14:153–161. doi: 10.1002/glia.440140210. [DOI] [PubMed] [Google Scholar]
- 10.Brune T, Fetzer S, Backus KH, Deitmer JW. Evidence for electrogenic sodium-bicarbonate cotransport in cultured rat cerebellar astrocytes. Pflügers Arch. 1994;429:64–71. doi: 10.1007/BF02584031. [DOI] [PubMed] [Google Scholar]
- 11.Burnashev N, Khodorova A, Jonas P, Helm PJ, Wisden W, Monyer H, Seeburg PH, Sakmann B. Calcium-permeable AMPA-kainate receptors in fusiform cerebellar glial cells. Science. 1992;256:1566–1570. doi: 10.1126/science.1317970. [DOI] [PubMed] [Google Scholar]
- 12.Charles AC, Merrill JE, Dirksen ER, Sanderson MJ. Intercellular signaling in glial cells: calcium waves and oscillations in response to mechanical stimulation and glutamate. Neuron. 1991;6:983–992. doi: 10.1016/0896-6273(91)90238-u. [DOI] [PubMed] [Google Scholar]
- 13.Chen J, Backus KH, Deitmer JW. Glutamate evokes intracellular calcium transients and K+ current oscillations in cultured rat astrocytes. Proceedings of the 25nd Göttingen Neurobiology Conference, Vol II (Elsner N, Wässle H, eds), p 880. Thieme Verlag; Stuttgart: 1997. [Google Scholar]
- 14.Cornell-Bell AH, Finkbeiner SM, Cooper MS, Smith SJ. Glutamate induces calcium waves in cultured astrocytes: long-range glial signaling. Science. 1990;247:470–473. doi: 10.1126/science.1967852. [DOI] [PubMed] [Google Scholar]
- 15.Dani JW, Chernjavsky A, Smith SJ. Neuronal activity triggers calcium waves in hippocampal astrocyte networks. Neuron. 1992;8:429–440. doi: 10.1016/0896-6273(92)90271-e. [DOI] [PubMed] [Google Scholar]
- 16.de Barry J, Ogura A, Kudo Y. Ca2+ mobilisation in cultured rat cerebellar cells: astrocytes are activated by t-ACPD. Eur J Neurosci. 1991;3:1146–1154. doi: 10.1111/j.1460-9568.1991.tb00049.x. [DOI] [PubMed] [Google Scholar]
- 17.Deitmer JW, Schneider HP. Intracellular acidification in the leech giant glial cell induced by glutamate and aspartate. Glia. 1997;19:111–122. [PubMed] [Google Scholar]
- 18.Enkvist MOK, Holopainen I, Akerman KEO. Glutamate receptor-linked changes in membrane potential and intracellular Ca2+ in primary rat astrocytes. Glia. 1989;2:397–402. doi: 10.1002/glia.440020602. [DOI] [PubMed] [Google Scholar]
- 19.Finkbeiner SM. Modulation and control of intracellular calcium. In: Kettenmann H, Ransom BR, editors. Neuroglia. Oxford UP; New York: 1995. pp. 273–288. [Google Scholar]
- 20.Fischer G. Growth requirements of immature astrocytes in serum-free hormonally defined media. J Neurosci Res. 1984;12:543–552. doi: 10.1002/jnr.490120403. [DOI] [PubMed] [Google Scholar]
- 21.Glaum SR, Holzwarth JA, Miller RJ. Glutamate receptors activate Ca2+ mobilization and Ca2+ influx into astrocytes. Proc Natl Acad Sci USA. 1990;87:3454–3458. doi: 10.1073/pnas.87.9.3454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hamill OP, Marty A, Neher E, Sakmann B, Sigworth FJ. Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches. Pflügers Arch. 1981;391:85–100. doi: 10.1007/BF00656997. [DOI] [PubMed] [Google Scholar]
- 23.Holzwarth JA, Gibbons SJ, Brorson JR, Philipson LH, Miller RJ. Glutamate receptor agonists stimulate diverse calcium responses in different types of cultured rat cortical glial cells. J Neurosci. 1994;14:1879–1891. doi: 10.1523/JNEUROSCI.14-04-01879.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Honoré T, Davies SN, Drejer J, Fletcher EJ, Jacobsen P, Lodge D, Nielsen FE. Quinoxalinediones: potent competitive non-NMDA glutamate receptor antagonists. Science. 1988;241:701–703. doi: 10.1126/science.2899909. [DOI] [PubMed] [Google Scholar]
- 25.Jabs R, Kirchhoff F, Kettenmann H, Steinhäuser C. Kainate activates Ca2+-permeable glutamate receptors and blocks voltage-gated K+ currents in glial cells of mouse hippocampal slices. Pflügers Arch. 1994;426:310–319. doi: 10.1007/BF00374787. [DOI] [PubMed] [Google Scholar]
- 26.Jensen AM, Chiu SY. Fluorescence measurement of changes in intracellular calcium induced by excitatory amino acids in cultured cortical astrocytes. J Neurosci. 1990;10:1165–1175. doi: 10.1523/JNEUROSCI.10-04-01165.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Jensen AM, Chiu SY. Differential intracellular calcium responses to glutamate in type 1 and type 2 cultured brain astrocytes. J Neurosci. 1991;11:1674–1684. doi: 10.1523/JNEUROSCI.11-06-01674.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kastritsis CHC, Salm AK, McCarthy K. Stimulation of P2Y purinergic receptor on type 1 astroglia results in inositol phosphate formation and calcium mobilization. J Neurochem. 1992;58:1277–1284. doi: 10.1111/j.1471-4159.1992.tb11339.x. [DOI] [PubMed] [Google Scholar]
- 29.Kawabata S, Tsutsumi R, Kohara A, Yamaguchi T, Nakanishi S, Okada M. Control of calcium oscillations by phosphorylation of metabotropic glutamate receptors. Nature. 1996;383:89–92. doi: 10.1038/383089a0. [DOI] [PubMed] [Google Scholar]
- 30.Kettenmann H, Backus KH, Schachner M. Aspartate, glutamate and γ-aminobutyric acid depolarize cultured astrocytes. Neurosci Lett. 1984;52:25–29. doi: 10.1016/0304-3940(84)90345-8. [DOI] [PubMed] [Google Scholar]
- 31.Kim WT, Rioult MG, Cornell-Bell AH. Glutamate-induced calcium signaling in astrocytes. Glia. 1994;11:173–184. doi: 10.1002/glia.440110211. [DOI] [PubMed] [Google Scholar]
- 32.Kirchhoff F, Kettenmann H. GABA triggers a [Ca2+]i increase in murine precursor cells of the oligodendrocyte lineage. Eur J Neurosci. 1992;4:1049–1058. doi: 10.1111/j.1460-9568.1992.tb00131.x. [DOI] [PubMed] [Google Scholar]
- 33.Korn SJ, Marty A, Connor JA, Horn R. Perforated patch recording. In: Conn PM, editor. Electrophysiology and microinjection. Academic; San Diego: 1991. pp. 364–373. [Google Scholar]
- 34.Kyrozis A, Reichling DB. Perforated-patch recording with gramicidin avoids artifactual changes in intracellular chloride concentration. J Neurosci Methods. 1995;57:27–35. doi: 10.1016/0165-0270(94)00116-x. [DOI] [PubMed] [Google Scholar]
- 35.MacVicar BA, Crichton SA, Burnard DM, Tse FWY. Membrane conductance oscillations in astrocytes induced by phorbol ester. Nature. 1987;329:242–243. doi: 10.1038/329242a0. [DOI] [PubMed] [Google Scholar]
- 36.Müller T, Möller T, Berger T, Schnitzer J, Kettenmann H. Calcium entry through kainate receptors and resulting potassium-channel blockade in Bergmann glial cells. Science. 1992;256:1563–1566. doi: 10.1126/science.1317969. [DOI] [PubMed] [Google Scholar]
- 37.Myers VB, Haydon DA. Ion transfer across lipid membranes in the presence of gramicidin A. II. The ion selectivity. Biochim Biophys Acta. 1972;274:313–322. doi: 10.1016/0005-2736(72)90179-4. [DOI] [PubMed] [Google Scholar]
- 38.Nakanishi S. Molecular diversity of glutamate receptors and implications for brain functions. Science. 1992;258:597–603. doi: 10.1126/science.1329206. [DOI] [PubMed] [Google Scholar]
- 39.Nedergaard M. Direct signaling from astrocytes to neurons in cultures of mammalian brain cells. Science. 1994;263:1768–1771. doi: 10.1126/science.8134839. [DOI] [PubMed] [Google Scholar]
- 40.Perkins MN, Stone TW. An iontophoretic investigation of the actions of convulsant kynurenines and their interaction with the endogenous excitant quinolinic acid. Brain Res. 1982;247:184–187. doi: 10.1016/0006-8993(82)91048-4. [DOI] [PubMed] [Google Scholar]
- 41.Pin JP, Duvoisin R. The metabotropic glutamate receptors: structure and functions. Neuropharmacology. 1995;34:1–26. doi: 10.1016/0028-3908(94)00129-g. [DOI] [PubMed] [Google Scholar]
- 42.Quandt FN, MacVicar BA. Calcium activated potassium channels in cultured astrocytes. Neuroscience. 1986;19:29–41. doi: 10.1016/0306-4522(86)90003-5. [DOI] [PubMed] [Google Scholar]
- 43.Rose CR, Ransom BR. Mechanisms of H+ and Na+ changes induced by glutamate, kainate, and d-aspartate in rat hippocampal astrocytes. J Neurosci. 1996;16:5393–5404. doi: 10.1523/JNEUROSCI.16-17-05393.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Rudy B. Diversity and ubiquity of K channels. Neuroscience. 1988;25:729–749. doi: 10.1016/0306-4522(88)90033-4. [DOI] [PubMed] [Google Scholar]
- 45.Salm AK, McCarthy KD. Norepinephrine-evoked calcium transients in cultured cerebral type 1 astroglia. Glia. 1990;3:529–538. doi: 10.1002/glia.440030612. [DOI] [PubMed] [Google Scholar]
- 46.Seifert G, Steinhäuser C. Glial cells in the mouse hippocampus express AMPA receptors with an intermediate Ca2+ permeability. Eur J Neurosci. 1995;7:1872–1881. doi: 10.1111/j.1460-9568.1995.tb00708.x. [DOI] [PubMed] [Google Scholar]
- 47.Sontheimer H, Kettenmann H, Backus KH, Schachner M. Glutamate opens Na+/K+ channels in cultured astrocytes. Glia. 1988;1:328–336. doi: 10.1002/glia.440010505. [DOI] [PubMed] [Google Scholar]
- 48.Stelzer A, Kay AR, Wong RKS. GABAA-receptor function in hippocampal cells is maintained by phosphorylation factors. Science. 1988;241:339–341. doi: 10.1126/science.2455347. [DOI] [PubMed] [Google Scholar]
- 49.Tanabe Y, Nomura A, Masu M, Shigemoto R, Mizuno N, Nakanishi S. Signal transduction, pharmacological properties, and expression patterns of two rat metabotropic glutamate receptors, mGluR3 and mGluR4. J Neurosci. 1993;13:1373–1378. doi: 10.1523/JNEUROSCI.13-04-01372.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Telgkamp P, Backus KH, Deitmer JW. Blockade of AMPA receptors by nickel in rat astrocytes. Glia. 1996;16:140–146. doi: 10.1002/(SICI)1098-1136(199602)16:2<140::AID-GLIA6>3.0.CO;2-7. [DOI] [PubMed] [Google Scholar]
- 51.Usowicz MM, Gallo V, Cull-Candy SG. Multiple conductance channels in type-2 cerebellar astrocytes activated by excitatory amino acids. Nature. 1989;339:380–383. doi: 10.1038/339380a0. [DOI] [PubMed] [Google Scholar]
- 52.Verkhratsky A, Kettenmann H. Calcium signaling in glial cells. Trends Neurosci. 1996;19:346–352. doi: 10.1016/0166-2236(96)10048-5. [DOI] [PubMed] [Google Scholar]
- 53.Walz W, MacVicar BA. Electrophysiological properties of glial cells: comparison of brain slices with primary cultures. Brain Res. 1986;443:321–324. doi: 10.1016/0006-8993(88)91626-5. [DOI] [PubMed] [Google Scholar]
- 54.Wyllie DJA, Cull-Candy SG. A comparison of non-NMDA receptor channels in type-2 astrocytes and granule cells from rat cerebellum. J Physiol (Lond) 1994;475:95–114. doi: 10.1113/jphysiol.1994.sp020052. [DOI] [PMC free article] [PubMed] [Google Scholar]