

Abstract
The termination of GABA synaptic action by high-affinity, Na+-dependent, neuronal, and glial plasma membrane transporters plays an important role in regulating neuronal activity in physiological and pathological conditions. We have investigated the cellular localization and distribution in the cerebral cortex of adult rats of one GABA transporter (GAT), GAT-3, by immunocytochemistry with affinity-purified polyclonal antibodies directed to its predicted C terminus that react monospecifically with a protein of ∼70 kDa.
Light microscopic studies revealed specific GAT-3 immunoreactivity (ir) in small punctate structures, and it was never observed in fibers or cell bodies. No changes in immunostaining were observed in sections incubated with GAT-3 antibodies preadsorbed with the related rat GAT-1 or mouse GAT-2/BGT-1 C-terminal peptides, whereas in sections incubated with GAT-3 antibodies preadsorbed with rat GAT-3 C-terminal peptide, ir was not present. The highest number of GAT-3-positive puncta was in layer IV and in a narrow band corresponding to layer Vb, followed by layers II and III. Many GAT-3-positive puncta were in close association with pyramidal and nonpyramidal neuron cell bodies. Ultrastructural studies showed that GAT-3 ir was localized exclusively to astrocytic processes, which were found in the neuropil and adjacent to axon terminals having either symmetric or asymmetric specializations. In sections processed by both preembedding labeling for GAT-3 and postembedding immunogold labeling for GABA, only some of the GAT-3-positive astrocytic processes were found close to GABAergic profiles.
These findings on the localization of GAT-3 in the cerebral cortex indicate that this transporter mediates GABA uptake into glial cells, and suggest that glial GABA uptake may function to limit the spread of GABA from the synapse, as well as to regulate overall GABA levels in the neuropil.
Keywords: synaptic transmission, GABA, GABA transporters, neocortex, symmetric and asymmetric synapses, astrocytes
The magnitude and duration of GABA synaptic action are regulated by plasma membrane proteins, termed GABA transporters (GATs), which mediate a high-affinity, Na+/Cl−-dependent, uptake of GABA into presynaptic axon terminals and glial processes (Iversen and Neal, 1968;Iversen and Snyder, 1968; Neal and Iversen, 1969; Iversen, 1971;Iversen and Kelly, 1975). GATs may also release GABA into the extracellular space in a Ca2+-independent, nonvesicular manner (Schwartz, 1982; Pin and Bockaert, 1989; Attwell et al., 1993;Levi and Raitieri, 1993), and they are targets for pharmacological intervention in neurological diseases characterized by GABAergic imbalance, such as epilepsy (Schousboe et al., 1983; Krogsgaard-Larsen et al., 1987; During et al., 1995). To date, four cDNAs encoding highly homologous GATs have been isolated from the rodent and human nervous systems (Guastella et al., 1990; Borden et al., 1992, 1994b; Clark et al., 1992; Liu et al., 1993). GATs have different pharmacological properties (Guastella et al., 1990; Borden et al., 1992, 1994b; Clark et al., 1992; Yamauchi et al., 1992; Liu et al., 1993) and tissue distributions (Ikegaki et al., 1994; Brecha et al., 1995; Durkin et al., 1995; Minelli et al., 1995; Ribak et al., 1996).
In a previous study, we reported that numerous neurons and some astrocytes express GAT-1 mRNA, and that GAT-1 immunoreactivity (ir) is localized to axon terminals forming symmetric synapses as well as to astrocytic processes (Minelli et al., 1995). This indicates that GAT-1, which exhibits a pharmacological profile (Guastella et al., 1990) typical of a “neuronal” transporter (Beart et al., 1972; Iversen and Kelly, 1975; Bowery et al., 1976; Larsson et al., 1983; Mabjeesh et al., 1992), also mediates GABA uptake into glial cells. These findings suggest that the organization of GABA uptake systems in the cerebral cortex is more complex than previously believed on the basis of pharmacological studies, and emphasize the importance of defining further the cellular localization of GATs.
Immunoblot, immunocytochemical, and in situ hybridization studies published to date indicate that GAT-3, a predicted 627-amino-acid protein found only in the nervous system (Borden et al., 1992; Ikegaki et al., 1994), is either absent or very weakly expressed in the cerebral cortex (Clark et al., 1992; Ikegaki et al., 1994;Brecha et al., 1995; Durkin et al., 1995). Because GAT-2 is not present in the cortex and it is expressed only by arachnoid and ependymal cells (Ikegaki et al., 1994; Durkin et al., 1995), these findings imply that glial GABA transport in the cerebral cortex is mediated mainly by GAT-1. However, it seems unlikely that GAT-1 is the sole transporter to mediate glial GABA uptake in the neocortex, because there is significant glial GABA uptake and there is a limited expression of GAT-1 in astrocytes (Minelli et al., 1995). To better understand GABA uptake systems in the cerebral cortex, we have used a new and specific affinity-purified antibody to evaluate the cellular localization and distribution of GAT-3 in the cerebral cortex of adult rats.
MATERIALS AND METHODS
Adult albino rats (Harlan Sprague Dawley, San Diego, CA, and Charles River, Milan, Italy), weighing 180–250 gm, were used in the present studies. Care and handling of animals were approved by the Animal Research Committees of the VAMC-West Los Angeles and of the University of Ancona.
Tissue preparation
For light microscopy, rats were deeply anesthetized with 30% chloral hydrate and perfused transcardially with 0.1 m PBS, pH 7.4, followed by 4% paraformaldehyde (PFA) in 0.1 mphosphate buffer (PB; pH 7.4). For electron microscopy, rats were perfused with 4% PFA plus 1% glutaraldehyde in PB. Brains were post-fixed for 1–2 hr at 4°C in the same fixative used for the perfusion, cut with a vibratome in either coronal or parasagittal plane into 25- to 30-μm-thick sections, which were collected serially in PBS and stored at 4°C until processing. Data were collected from a region of the parietal cortex characterized by the presence of a conspicuous layer IV, with intermingled dysgranular regions, densely packed layers II and III, and a relatively cell-free layer Va. This region corresponds to the first somatic sensory cortex (SI), as defined by Zilles (1985) and Chapin and Lin (1990).
Immunocytochemistry
Antibody production. Affinity-purified rabbit polyclonal antibodies (369-D and 374-E) directed to the predicted C terminus (Borden et al., 1992; Clark et al., 1992) of rat GAT-3 (rGAT-3607-627) were used for these studies. Rabbits were initially immunized with 100 nmol of the GAT-3607-627 conjugated to keyhole limpet hemocyanin (KLH) in complete Freund’s adjuvant, and subsequently immunized at 4–6 week intervals with 50 nmol of the GAT-3607-627 conjugated to KLH in incomplete Freund’s adjuvant. Plasma was harvested at regular intervals after each immunization, and sera were tested for specific immunostaining. Selected sera were affinity-purified using an Epoxy–Sepharose column prepared with the C-terminal sequence of GAT-3 following the manufacturer’s instructions (Pharmica Biotech, Piscataway, NJ). Antibodies were eluted with 3 m KSCN, collected and concentrated with a Centriprep-30 (Amicon, Beverly, MA), and stored in 1% BSA and 0.1 m NaN3 in 0.1 m PB at −70°C.
Antibody characterization by immunoblotting and immunoblocking. Sprague Dawley rats (150–250 gm) were perfused with cold 4 mm Tris-HCl, pH 7.4, containing 0.32 m sucrose, 1 mm EDTA, 0.5 mmphenylmethylsulphonyl fluoride (PMSF), and 0.5 mmN-ethylmaleimide (NEM). After the meninges were removed, brains were homogenized by glass–Teflon homogenizer in 10 vol of ice-cold buffer (0.32 m sucrose; 4 mm Tris-HCl, pH 7.4; 1 mm EDTA; and 0.25 mm dithiothreitol) (Ikegaki et al., 1994). The homogenate was centrifuged at 1000 × g for 15 min at 4°C. The pellet was discarded, and aliquots of the supernatant (total brain extract) were either used immediately or stored at −80°C. A crude membrane preparation of the cerebral cortex was also made by the same procedure. After removing the low-speed pellet, the remaining supernatant was recentrifuged at 105,000 × g for 1 hr at 4°C and the resulting crude membrane pellet (Thomas and McNamee, 1990) was resuspended in homogenization buffer containing protease inhibitors (1 mmEDTA, 0.5 mm PMSF, 0.5 mm NEM) and either used immediately or stored at −80°C. Protein concentrations were measured according to Bradford (1976), with the Bio-Rad protein assay kit (Bio-Rad Laboratories, Hercules, CA). Aliquots of total brain extract and of crude membrane fraction were mixed with equal volumes of 2× electrophoresis sample buffer with or without 4 m urea (final concentration). The samples were subjected to 10% SDS-PAGE with a 3% stacking gel under reducing conditions, and the separated proteins were electrophoretically transferred to a nitrocellulose filter (0.45 μm) according to the method of Towbin et al. (1979). After the transfer, the gels were stained with Coomassie blue and visually examined for transfer efficiency. The blots were sequentially incubated with the GAT-3 antibody (369D, dilution 1:500) and goat anti-rabbit IgG conjugated to horseradish peroxidase and then reacted with BM chemoluminescence Western blotting kit (Boeh- ringer Mannheim, Mannheim, Germany) following the instructions provided by the manufacturer. Labeled bands were visualized on X-Omat AR films (Eastman Kodak, Rochester, NY). As a control, blots were submitted to the same immunolabeling procedure except that the primary antibody was either omitted or preadsorbed with 10−5m rat GAT-3607-627 peptide.
GAT-3 antibodies were characterized also by immunoblocking with 10−5m or 10−6m rat GAT-3607-627 peptide, with 10−5mrat GAT-1588-599, or mouse GAT-2/BGT-1596-614, C-terminal peptides (Guastella et al., 1990, 1992; Borden et al., 1992;Yamauchi et al., 1992; Liu et al., 1993). In all blocking experiments, the antibody dilution was 1:1000.
To directly compare GAT-3 and GAT-1 immunostaining, sections adjacent to those processed with GAT-3 antisera were incubated with affinity-purified polyclonal antibodies (341-F and 346-J) (Minelli et al., 1995) directed to the predicted C terminus of rat GAT-1 (rGAT-1588-599) (Guastella et al., 1990) peptide.
Procedure. Free-floating sections were preincubated for 1 hr in normal goat serum (NGS; 10% in PBS) with 0.5% Triton X-100 and then incubated overnight at 4°C in GAT-3 or GAT-1 primary antibodies (dilution 1:1000 for both antibodies). The next day, sections were rinsed in PBS and incubated for 15 min in 10% NGS and then in biotinylated anti-rabbit IgG at a dilution of 1:100 in PBS (1 hr at room temperature or overnight at 4°C). Sections were rinsed in PBS, incubated in avidin–biotin–peroxidase complex (ABC) (Hsu et al., 1981) for 30 min, washed several times in PBS, and then incubated first in 50–75 mg DAB/100 ml Tris (0.05 m) and then in fresh DAB with 0.02% H2O2. Sections were washed in PBS, mounted on subbed slides, air-dried, dehydrated, and coverslipped.
For electron microscopy, vibratome sections were pretreated to increase the penetration of immunoreagents by a mild ethanol treatment (10, 25, and 10%; 5 min each). GAT-3 antibodies were used at 1:1000 dilution; Triton X-100 was not used. After completion of the ICC procedure, sections were washed in PB, post-fixed for ∼30 min in 2.5% glutaraldehyde in PB, washed in PB, and post-fixed for 1 hr in 1% OsO4. After dehydration, sections were cleared in propylene oxide, flat-embedded in Epon–Spurr between acetate foils (Aclar; Ted Pella, Redding, CA), and polymerized at 60°C for 36 hr. When polymerization was complete, the embedded sections were examined under a dissecting microscope. Small strips of cortex were excised with razor blades and either glued to cured resin blocks or reembedded in Epon–Spurr. Semithin (1 μm) sections were cut with a Reichert ultramicrotome and collected on glass slides without counterstaining for light microscopical inspection. Ultrathin sections were cut either from the surface or from the edge (i.e., perpendicular to the plane of section), counterstained with uranyl acetate and lead citrate or with lead citrate only, and examined with a Jeol T8 electron microscope.
Some ultrathin sections from different animals were processed first for the visualization of GAT-3 ir and subsequently for the demonstration of GABA ir by a standard postembedding immunogold method (De Biasi et al., 1994) using a rabbit polyclonal GABA antiserum (Sigma, St. Louis, MO). Briefly, thin sections collected on nickel grids were immersed in 1% sodium borohydride (15 min) and, after several rinses in Tris-buffered saline (TBS), incubated at room temperature with 10% NGS (30 min) and with the primary antiserum (1:10,000, overnight). After extensive rinses, grids were incubated in a solution of goat anti-rabbit IgG coupled to 15 nm gold particles (Biocell, Cardiff, UK) diluted 1:30 (1 hr) and then counterstained with uranyl acetate and lead citrate or with lead citrate only. Specificity of the immunogold labeling was evaluated by blocking experiments in which the anti-GABA serum was adsorbed with GABA, in both free and conjugated forms. Method specificity was controlled by the application of rabbit nonimmune serum as well as by processing series of sections omitting various stages of the regular staining sequence. Preadsorption of the antiserum, diluted for tissue reaction, with GABA (free or conjugated) abolished labeling, as did substitution of the primary antiserum with preimmune serum. The pattern of GABA immunogold labeling obtained was the same as that described previously (Beaulieu et al., 1994). Terminals with asymmetric specialization were always devoid of labeling. Nonspecific or background labeling was low in our preparations and, therefore, it was not necessary to perform a statistical analysis of the distribution of gold particles to determine whether a profile was labeled.
The immunogold protocol used to visualize GABA ir allowed the preservation of the DAB reaction product indicating GAT-3 ir. Therefore, in all of the thin sections examined, the following profiles could be identified: (1) profiles containing only the granular electron-dense reaction product of DAB indicating the presence of GAT-3 ir; (2) profiles containing only gold particles indicating the presence of GABA; and (3) profiles unlabeled by either antibodies.
RESULTS
The specificity of the antibody was examined by immunoblot. The purified antibody against GAT-3 revealed a band of ∼70 kDa both in total brain extract and in the crude membrane fraction of the cerebral cortex (Fig. 1A,B). Omission of the primary antibody or preadsorption with GAT-3607-627 peptide prevented labeling of the blots (Fig.1C).
Fig. 1.

Specificity of the GAT-3 antibody assessed by immunoblotting. 100 μg of protein was loaded for each lane.A, Total brain extract solubilized in sample buffer without urea; a fraction of incompletely solubilized antigen remains at the gel top. B, Crude membrane fraction of the cerebral cortex solubilized in sample buffer containing 4 m urea.C, Crude membrane fraction processed with GAT-3 antibody preadsorbed with 10−5mGAT-3607-627 peptide. Open arrow indicates the gel top.
GAT-3 ir was differentially distributed in the brain: high levels were in the olfactory bulb, thalamus, hypothalamus, and brainstem, whereas low levels were in the caudate-putamen, cerebral cortex, hippocampus, and cerebellum (Fig. 2). In the cerebral cortex, GAT-3 ir was localized exclusively to small punctate structures that were difficult to resolve at the light microscopic level and never appeared as labeled fibers or cell bodies (Figs. 3, 4, 5, 6). Specific GAT-3 ir was prevented when GAT-3 antibodies had been preadsorbed with 10−5 or 10−6m rat GAT-3607-627 (Fig. 4D), whereas no changes were observed in sections incubated with GAT-3 antibodies preadsorbed with 10−5m rat GAT-1588-599 and mouse GAT-2/BGT-1596-614C-terminal peptides (Fig. 4B,C). The same pattern of GAT-3 ir was observed in sections from different animals, and it did not show any significant regional variations along the rostro-caudal or medio-lateral extent of the neocortex.
Fig. 2.

Parasagittal section of the rat brain illustrating the regional distribution of GAT-3 ir. Cb, Cerebellum; CP, caudate–putamen; HF, hippocampal formation; Hy, hypothalamus; T, thalamus. Scale bar, 2 mm.
Fig. 3.
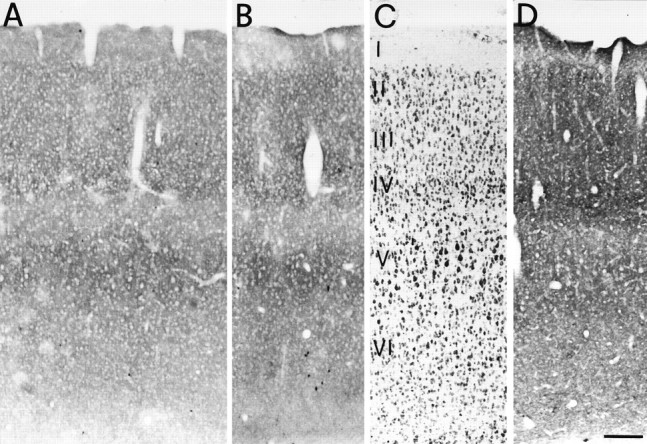
Distribution of GAT-3 ir in SI of adult rats perfused with 4% paraformaldehyde only (A) or with 4% paraformaldehyde and 1% glutaraldehyde (B).C, A Nissl-stained section adjacent to Aand B. Roman numerals indicate cortical layers. D, Distribution of GAT-1 in SI of an adjacent section. Scale bar, 150 μm.
Fig. 4.
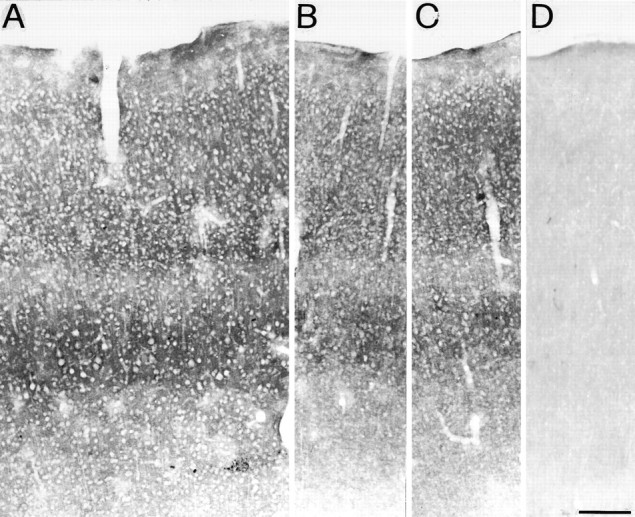
A, GAT-3 ir in SI is localized exclusively to small punctate structures that are densely packed around cell bodies, and is highest in layer IV and Vb. B–D, Adjacent sections processed with GAT-3 antibody preincubated with 10−5m rat GAT-1588-599(B), mouse GAT-2/BGT-1593-602(C), and 10−5m rat GAT-3607-627 (D). GAT-3 ir does not change in B and C, but it is completely abolished in D. Scale bar, 100 μm.
Fig. 5.
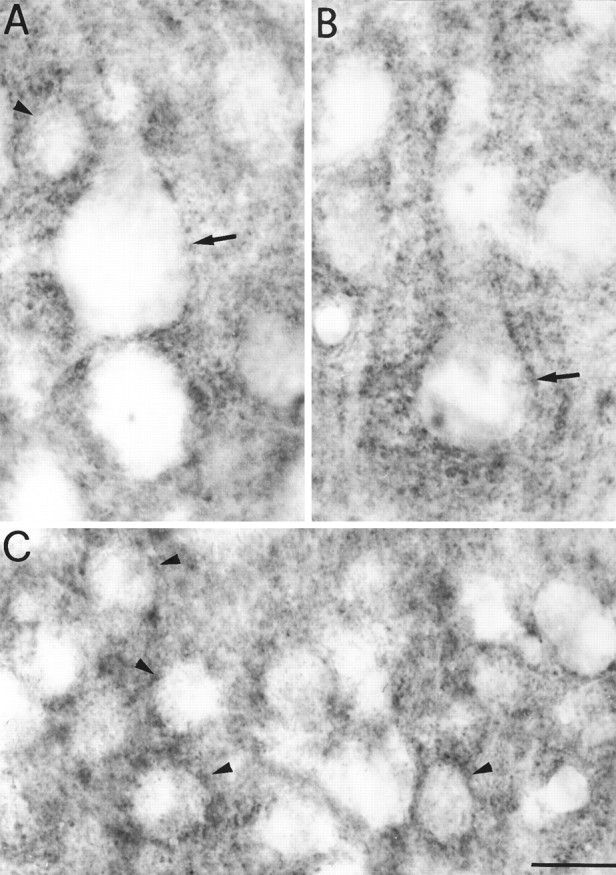
GAT-3 ir within puncta outline large cell bodies that are likely to be pyramidal cells (arrows inA and B) and small cells that are likely to be nonpyramidal neurons (arrowheads inA and C). Scale bar, 15 μm.
Fig. 6.

GAT-3 (A) and GAT-1 (B) ir in semithin sections. With both antisera, labeling is in punctate structures scattered throughout the neuropil among unlabeled cell bodies. GAT-3-immunoreactive puncta are small and evenly distributed (A), whereas some of the GAT-1-immunoreactive puncta are large (arrows) and surround neuronal cell bodies and major dendrites (B). The apparent GAT-3 labeling of perivascular structures has not been confirmed at the electron microscopic level. Phase-contrast microscopy; noncounterstained sections. Layer V. Scale bar, 25 μm.
GAT-3 ir was present throughout the depth of SI, but the apparent density of positive puncta varied in the different layers. The highest level of GAT-3 ir was observed in layer IV and in a narrow band corresponding to the lower portion of layer V (Vb; Figs.3A,B, 4A). In these layers, the neuropil was filled by numerous, heavily stained, GAT-3-positive puncta, which were densely packed around unstained cell body profiles (Figs. 3A, 4A). In layers II and III, GAT-3 ir exhibited the same features but was less intense. Ir was faint in layers I, Va, and VI, where positive puncta were sparser and less intensely stained than in the other layers (Figs. 3A,4A). Comparison between GAT-1 and GAT-3 ir showed that GAT-3 ir, although less intense, displayed a laminar pattern similar to that of GAT-1 ir (Fig. 3C).
GAT-3 ir was not evenly distributed in SI: in all cortical layers, but particularly in layers VI and IV, restricted areas of tissue were observed, in which the expression of GAT-3 was very low (Figs.3A, 4A). Patchy regions exhibited clear-cut borders (Figs. 3A, 4A) and were observed in all sections from all animals.
GAT-3-positive puncta were small in size (<1 μm in diameter) and were observed both in the neuropil and in close relationship with cell bodies. In the latter case, GAT-3-positive puncta formed a continuous sheet around the somata of both pyramidal and nonpyramidal neurons (Fig. 6A–C). In layers II, III, and V, GAT-3-positive puncta were also in close association with the proximal portion of basal (Fig. 5A) and apical (Fig.5B) dendrites of pyramidal cells. GAT-3-positive puncta were different from GAT-1-positive puncta since (1) they were more numerous in the neuropil, and (2) they were smaller than the majority of GAT-1-immunoreactive puncta. However, the distribution of GAT-3-positive puncta around somata and dendrites of cortical neurons (Fig. 5) is similar to that of GAT-1 (Fig. 6) (see also Fig. 8 in Minelli et al., 1995).
Fig. 8.
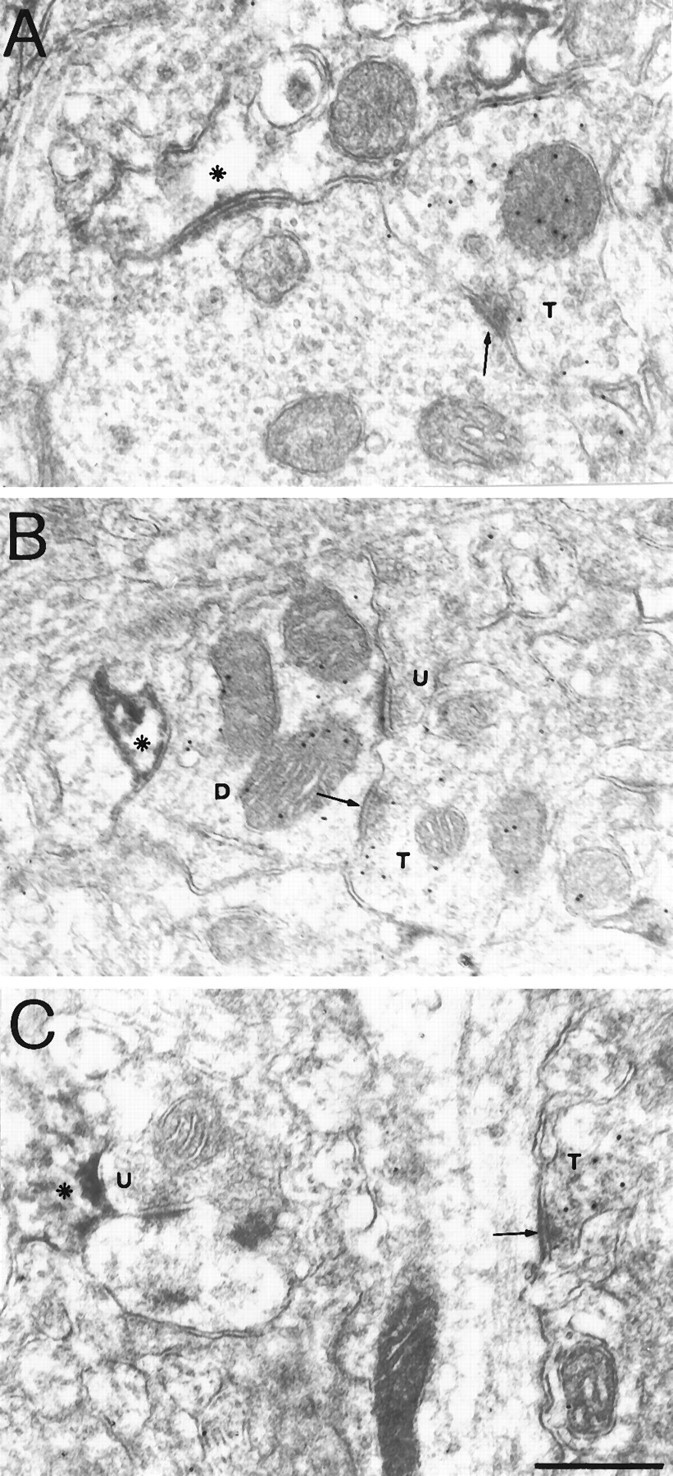
Simultaneous subcellular localization of GAT-3 and GABA ir. A, A GAT-3-labeled astrocytic process (asterisk) is adjacent to a GABA-containing axon terminal (T) forming a symmetric synapse (arrow) on an unlabeled dendrite. B, A GAT-3-labeled astrocytic process (asterisk) is adjacent to a GABA-immunogold-labeled dendrite (D) that receives a symmetric synapse (arrow) from a GABA-immunogold-labeled terminal (T) and an asymmetric synapse from an unlabeled axon terminal (U). C, A GAT-3-labeled astrocytic process (asterisk) is adjacent to an unlabeled axon terminal (U) with asymmetric specialization. A GABA-immunogold-labeled terminal (T) makes a symmetric synapse (arrow) on a dendrite. Uranyl acetate and lead citrate counterstaining. Scale bar, 0.5 μm.
The ultrastructural pattern of GAT-3 labeling was the same in all layers of the cortex. GAT-3 ir was found exclusively in distal astrocytic processes, characterized by an irregular contour, whereas astrocytic cell bodies and neuronal profiles were always unlabeled (Fig. 7). Electron-dense reaction product was present along the cytoplasmic side of the plasma membrane and in cytoplasmatic granular patches. Labeled astrocytic processes were evenly distributed throughout the neuropil, as they were found adjacent to axon terminals making either symmetric (Fig. 7A,B) or asymmetric (Fig. 7C,D) synaptic contact with cell bodies (Fig. 7A), dendrites (Fig. 7B), dendritic spines (Fig. 7C,D), or close to neuronal profiles that did not form synaptic contacts in the plane of section (not shown). Analysis of patchy regions devoid of GAT-3 ir showed that cortical tissue was normally preserved, but lacked GAT-3 ir.
Fig. 7.
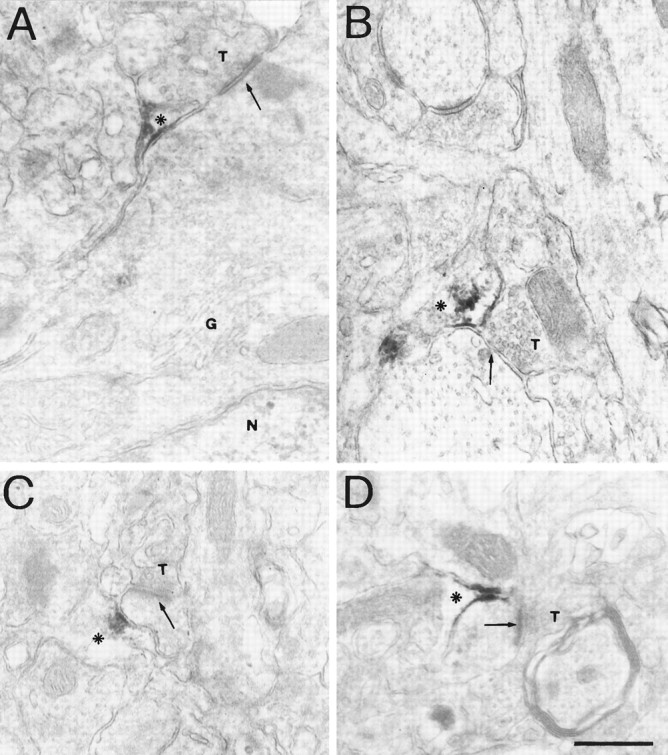
Subcellular localization of GAT-3 ir.A, A GAT-3-labeled astrocytic process (asterisk) is adjacent to an axon terminal (T) making a symmetric synapse (arrow) on a neuronal cell body. N, Nucleus; G, Golgi complex. B, A GAT-3-labeled astrocytic process (asterisk) is adjacent to an axon terminal (T) making a symmetric synapse (arrow) on a dendrite. C,D, GAT-3-labeled astrocytic processes (asterisks) are adjacent to axon terminals (T) making asymmetric synapses (arrows) on dendritic spines. Lead citrate counterstaining. Scale bar, 0.5 μm.
In sections reacted for postembedding immunogold visualization of GABA, some of the GAT-3-stained astrocytic processes were found adjacent either to terminals with symmetric specialization (Fig.8A) or to dendrites (Fig.8B) that were selectively enriched in gold particles coding for GABA. However, most of the GAT-3-stained astrocytic processes were not associated with GABA-positive processes: of these, some were close to terminals forming asymmetric synapses (Fig.8C). None of these astrocytic processes contained GABA ir (Fig. 8A–C).
DISCUSSION
The present study shows that in the cerebral cortex of adult rats GAT-3 is localized exclusively to astrocytic processes, and that the distribution of GAT-3 ir processes is compatible with a role of GAT-3 in the regulation of GABA levels in the extracellular space.
Antibody specificity and comparison with previous studies
To visualize the distribution of GAT-3, affinity-purified polyclonal antibodies were raised against a synthetic peptide corresponding to the predicted C terminus of rat GAT-3. The characterization of these antibodies by immunoblot analysis of rat brain homogenates and crude membrane fractions of cerebral cortex showed that they monospecifically react with a protein of ∼70 kDa, consistent with the molecular mass predicted by the analysis of cloned cDNA (Borden et al., 1992; Clark et al., 1992). The specificity of these antibodies was demonstrated further by immunoblocking studies, which revealed that GAT-3 ir was prevented when GAT-3 antibodies were preadsorbed with their specific cognate peptide, but not with other related GABA transporter C-terminal peptides.
Overall, the distribution of GAT-3 ir described here is in agreement with previous immunocytochemical (Ikegaki et al., 1994) and in situ hybridization (Clark et al., 1992; Durkin et al., 1995) studies, which also showed that the highest levels of GAT-3 expression are in olfactory bulb, hypothalamus, thalamus, and brainstem. A major difference between earlier descriptions and the present study concerns only the level of GAT-3 expression in the cerebral cortex. These studies indicate that GAT-3 is either absent or very weakly expressed in the cerebral cortex (Clark et al., 1992; Ikegaki et al., 1994;Brecha et al., 1995; Durkin et al., 1995), whereas our results show that neocortex exhibits low, but not negligible, levels of GAT-3. Given the specificity of the antibodies used in the present study, it is likely that the level of GAT-3 mRNA or protein was underestimated in earlier studies, perhaps because of the very high levels of GAT-3 expression elsewhere in the brain.
In the cerebral cortex, GAT-3 is expressed only by astrocytes
GAT-3 ir is localized in numerous, small punctate structures throughout the cerebral cortex. Ultrastructural studies revealed that neuronal cell bodies, dendrites, and axon terminals were always unlabeled, and they showed that GAT-3 ir was exclusively localized to many small processes. Their irregular contours conformed to outlines of surrounding neuropilar elements, and previous studies have shown that these features are typical of astrocytic processes (Peters et al., 1991). Thus, although we cannot rule out the possibility that some cortical neurons synthesize GAT-3 at such low levels that they cannot be visualized by current immunocytochemical procedure, the present study indicates that in the cerebral cortex GAT-3 is expressed exclusively in astrocytes and it is localized to their processes.
The existence of GABA uptake systems associated with glial cells was demonstrated in the cerebral cortex in early autoradiographic studies, which showed a significant [3H]β-alanine uptake in cortical slices and synaptosome preparations (Schon and Kelly, 1975), and in pharmacological studies showing that GABA transport in cortical astrocytic membranes is highly sensitive to β-alanine (Mabjeesh et al., 1992).
The present anatomical results on the cellular localization of GAT-3 are in agreement with previous data. For instance, heterologous cells transfected with GAT-3 and “glial” GABA transport have similar pharmacological properties (Borden et al., 1992; Clark et al., 1992;Clark and Amara, 1994). That is, GABA uptake by GAT-3 is strongly inhibited by β-alanine and β-proline (Borden et al., 1992; Clark et al., 1992; Clark and Amara, 1994), and it is not inhibited by the “neuronal” GABA transport inhibitors ACHC and DABA (Iversen and Kelly, 1975; Neal and Bowery, 1977; Gavrilovic et al., 1984; Kanner and Bendahan, 1990; Mabjeesh et al., 1992). Furthermore, GAT-3 is expressed by rat retinal Muller cells (Brecha et al., 1995), by hippocampal astrocytic processes (Ribak et al., 1996), and by cultured astrocytes (Borden et al., 1994a). The reported expression of GAT-3 in neurons located in the brainstem and in a few amacrine cells of the retina (Clark et al., 1992; Brecha et al., 1995) does not contradict the present findings: rather, it suggests that there is a regional heterogeneity in GAT-3 cellular expression. Whether other neurotransmitter transporters display a similar regional heterogeneity in their cellular expression pattern remains an interesting challenge for future investigations.
The demonstration that cortical astrocytes express GAT-3 together with previous findings on astrocytic expression of GAT-1 (Minelli et al., 1995) raises some interesting questions. First, are GAT-1 and GAT-3 coexpressed? Although GAT-3 is expressed more robustly than GAT-1 in astrocytes, astrocytes expressing GAT-1 and GAT-3 display the same laminar segregation and, at least in part, an overlapping distribution; it is possible, therefore, that a subpopulation of cortical astrocytes may coexpress both GAT-1 and GAT-3. Second, what is the relative contribution of each of these transporters to overall GABA uptake by glial cells in the cerebral cortex? There are no experimental studies addressing this question. However, because GAT-1 and GAT-3 transport exhibit different ionic dependences and inhibitor sensitivities in heterologous cell systems (Guastella et al., 1990; Borden et al., 1992;Clark et al., 1992; Keynan et al., 1992), it is conceivable that the relative contribution of GAT-1 and GAT-3 to glial GABA uptake is determined by different regulatory mechanisms, e.g., different responses to the composition of the extracellular milieuand/or different regulation of GAT’s activity and/or expression. For instance, protein kinase C (PKC) activators, which increase GABA uptake in cells transfected with GAT-1 (Corey et al., 1994), reduce glial GABA transport in primary astrocyte cultures (Gomeza et al., 1991). A differential regulation of astrocytic GAT-1 and GAT-3 would allow a great adaptability in the fine regulation of GABA extracellular levels under various physiological conditions.
The distribution of GAT-3 in the cerebral cortex is compatible with a dual physiological role
The major features of the distribution of GAT-3 ir in the cerebral cortex, as revealed for the first time by this study, are as follows: (1) GAT-3 ir astrocytic processes display a laminar segregation similar to that of GAT-1 ir puncta (Minelli et al.;, 1995) and of GABAergic axon terminals (Ribak, 1978; Houser et al., 1984); and (2) GAT-3-positive astrocytic processes are not exclusively associated with GABAergic profiles, as shown by double-labeling with GABA.
Axon terminals releasing GABA are densest in layer IV, followed by layers II and III, and in layer Vb GABAergic terminals outline unstained pyramidal cell bodies (Ribak, 1978; Freund et al., 1983;Houser et al., 1984). These cortical layers or sublayers are also the richest in GAT-1-positive puncta (Minelli et al., 1995). Here we have shown that the distribution of GAT-3-positive puncta is similar to that of axon terminals releasing GABA and of GAT-1-positive puncta. This correlation implies that GAT-3 is highly expressed in the very layers in which GABA is prominently released and, therefore, highly concentrated. The presence of numerous “patches” devoid of GAT-3 ir in neocortex indicates some differences in GAT-3 expression and activity. The functional meaning of this organizational feature of GAT-3 ir is at present unknown; it is worth noting, however, that an analogous distribution has been reported for a glial glutamate (Glu) transporter, GLT-1 (Rothstein et al., 1994).
GAT-3-positive puncta are in close relationship with neuronal cell bodies and proximal dendrites, the major targets of GABAergic axon terminals (Freund et al., 1983; Houser et al., 1984). To evaluate directly the relationship between GAT-3-positive astrocytic processes and GABA-releasing axon terminals, we combined preembedding ICC for GAT-3 with postembedding immunogold labeling for the visualization of GABA. The results of these experiments show that some GAT-3-stained astrocytic processes are in direct contact with GABA-containing axon terminals that are likely to release GABA. This observation suggests that GAT-3 uptake mediates GABA reaccumulation in glial cells surrounding GABAergic synapses, thus contributing to both termination and regulation of GABAergic synaptic transmission (Isaacson et al., 1993).
GAT-3-positive puncta, however, are also (1) scattered in the neuropil and (2) close to axon terminals making asymmetric synapses. Their functional role can be envisaged better bearing in mind that (1) GABA may diffuse in the neuropil and therein act in a paracrine fashion and (2) that this paracrine effect is potentiated by SKF-89976A, a competitive blocker of GABA transport (Dingledine and Korn, 1985;Thompson and Gahwiler, 1992; Isaacson et al., 1993). Because the blocker used by Isaacson et al. (1993) is more potent at GAT-1 than at GAT-3 (Borden et al., 1994b), these findings suggest that the diffuse action of GABA is modulated by GAT-1, but this does not rule out that GAT-3 may also play a role. The availability of GAT-3-specific blockers [e.g., (S)-SNAP-5114; Borden et al., 1994b] will allow for further evaluation of GAT-3’s modulation of GABA’s diffuse action. Such a function of GAT-3 is strongly suggested by the presence of numerous GAT-3-positive astrocytic processes in the neuropil. The results presented here show that some GAT-3-positive astrocytic processes are adjacent to axon terminals forming asymmetric synapses, which contain and presumably release Glu (Conti and Minelli, 1996). Interestingly, Isaacson et al. (1993) showed that inhibition of GABA uptake enhances the presynaptic action of GABA on presumed GABAB receptors located on excitatory nerve terminals. GAT-3-positive astrocytic processes adjacent to axon terminals forming asymmetric synapses could contribute, therefore, to the regulation of GABA action on excitatory synaptic transmission.
Finally, it has been shown that GATs may release GABA in a nonvesicular, Ca2+-independent manner (Schwartz, 1982; Attwell et al., 1983; Levi and Raiteri, 1993). These data, together with the present demonstration of a widespread distribution of GAT-3-positive astrocytic processes in the neuropil, suggest that GAT-3 may mediate GABA release in the cerebral cortex.
Footnotes
Received December 1, 1995; revised June 27, 1996; accepted July 8, 1996.
This work was supported by NATO (CRG 910273), Consiglio Nazionale delle Ricerche (AI 90-01371, 91-00731, CT04), Ministero dell’Università e della Ricerca Scientifica e Tecnologica, National Institutes of Health (EY04067), VA Medical Research Funds and Morphology Imaging CORE DK 41301. We thank Dr. Catia Sternini for helpful comments and discussions, Ty K. Chen and Katherine Wen for assistance in antibody production, purification, and characterization, and Francesca Natalini for help with histology.
Correspondence should be addressed to Fiorenzo Conti, Istituto di Fisiologia Umana, Università di Ancona, Via Ranieri, Monte d’Ago, I-60131 Ancona, Italy.
REFERENCES
- 1.Attwell D, Barbour B, Szatkowski M. Nonvesicular release of neurotransmitter. Neuron. 1993;11:401–407. doi: 10.1016/0896-6273(93)90145-h. [DOI] [PubMed] [Google Scholar]
- 2.Beaulieu C, Campistron G, Crevier C. Quantitative aspects of the GABA circuitry in the primary visual cortex of the adult rat. J Comp Neurol. 1994;339:559–572. doi: 10.1002/cne.903390407. [DOI] [PubMed] [Google Scholar]
- 3.Beart PM, Johnston GAR, Uhr ML. Competitive inhibition of GABA uptake in rat brain slices by some GABA analogues of restricted conformation. J Neurochem. 1972;19:1855–1861. doi: 10.1111/j.1471-4159.1972.tb01474.x. [DOI] [PubMed] [Google Scholar]
- 4.Borden LA, Smith KE, Hartig PR, Brancheck TA, Weinshank RL. Molecular heterogeneity of the γ-aminobutyric acid (GABA) transport system. J Biol Chem. 1992;267:21098–21104. [PubMed] [Google Scholar]
- 5.Borden LA, Smith KE, Vaysse PJ-J, Weinshank RL, Branchek TA. GABA transporters in neuronal and glial cell cultures: correlation of pharmacological activity with mRNA localization. Soc Neurosci Abstr. 1994a;20:919. [Google Scholar]
- 6.Borden LA, Dhar TGM, Smith KE, Branchek TA, Glucowski C, Weinshank RL. Cloning of the human homologue of the GABA transporter GAT-3 and identification of a novel inhibitor with selectivity for this site. Receptors Channels. 1994b;2:207–213. [PubMed] [Google Scholar]
- 7.Bowery NG, Jones GP, Neal MJ. Selective inhibition of neuronal GABA uptake by cis -1,3-aminocyclohexane carboxylic acid (ACHC). Nature. 1976;264:281–284. doi: 10.1038/264281a0. [DOI] [PubMed] [Google Scholar]
- 8.Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 9.Brecha NC, Johnson J, Chen T, Conti F, Minelli A, DeBiasi S, Ribak C. GABA transporter expression in the rat nervous system. Presynaptic mechanisms of neurotransmission. Soc Neurosci Abstr. 1995;124:1. [Google Scholar]
- 10.Chapin JK, Lin C-S. The somatic sensory cortex of rat. In: Kolb B, Tees RG, editors. The cerebral cortex of the rat. MIT; Cambridge: 1990. pp. 341–380. [Google Scholar]
- 11.Clark JA, Amara SG. Stable expression of a neuronal γ-aminobutyric acid transporter, GAT-3, in mammalian cells demonstrates unique pharmacological properties and ion dependence. Mol Pharmacol. 1994;46:550–557. [PubMed] [Google Scholar]
- 12.Clark JA, Deutch AY, Gallipoli PZ, Amara SG. Functional expression and CNS distribution of a β-alanine-sensitive neuronal GABA transporter. Neuron. 1992;9:337–348. doi: 10.1016/0896-6273(92)90172-a. [DOI] [PubMed] [Google Scholar]
- 13.Conti F, Minelli A. The anatomy of glutamatergic transmission in the cerebral cortex. In: Conti F, Hicks TP, editors. Excitatory amino acids & the cerebral cortex. MIT; Cambridge: 1996. pp. 81–98. [Google Scholar]
- 14.Corey JL, Davidson N, Lester HA, Brecha NC, Quick MW. Protein kinase C modulates the activity of a cloned γ-aminobutyric acid transporter expressed in Xenopus oocytes via regulated subcellular redistribution of the transporter. J Biol Chem. 1994;269:14759–14767. [PubMed] [Google Scholar]
- 15.De Biasi S, Vitellaro-Zuccarello L, Bernardi P, Valtschanoff JG, Weinberg RJ. Ultrastructural and immunocytochemical characterization of primary afferent terminals in the rat cuneate nucleus. J Comp Neurol. 1994;347:275–287. doi: 10.1002/cne.903470209. [DOI] [PubMed] [Google Scholar]
- 16.Dingledine R, Korn SJ. γ-Aminobutyric acid uptake and the termination of inhibitory synaptic potentials in the rat hippocampal slices. J Physiol (Lond) 1985;366:387–409. doi: 10.1113/jphysiol.1985.sp015804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.During MJ, Ryder KM, Spencer DD. Hippocampal GABA transporter function in temporal lobe epilepsy. Nature. 1995;376:174–177. doi: 10.1038/376174a0. [DOI] [PubMed] [Google Scholar]
- 18.Durkin MM, Smith KE, Borden LA, Weinshank RL, Branchek TA, Gustafson EL. Localization of messenger RNAs encoding three GABA transporters in rat brain: an in situ hybridization study. Mol Brain Res. 1995;33:7–21. doi: 10.1016/0169-328x(95)00101-w. [DOI] [PubMed] [Google Scholar]
- 19.Freund TF, Martin KAC, Smith AD, Somogyi P. Glutamate decarboxylase-immunoreactive terminals of Golgi-impregnated axoaxonic cells and presumed basket cells in synaptic contacts with pyramidal neurons of the cat’s visual cortex. J Comp Neurol. 1983;221:263–278. doi: 10.1002/cne.902210303. [DOI] [PubMed] [Google Scholar]
- 20.Gavrilovic J, Raff M, Cohen G. GABA uptake by purified rat Swann cells in culture. Brain Res. 1984;303:183–185. doi: 10.1016/0006-8993(84)90226-9. [DOI] [PubMed] [Google Scholar]
- 21.Gomeza J, Casado M, Gimenez C, Aragon C. Inhibition of high-affinity γ-aminobutyric acid uptake in primary astrocyte cultures by phorbol esters and phospholipase C. Biochem J. 1991;275:435–439. doi: 10.1042/bj2750435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Guastella J, Nelson N, Nelson H, Czyzyk L, Keynan S, Miedel MC, Davidson N, Lester HA, Kanner BI. Cloning and expression of a rat brain GABA transporter. Science. 1990;249:1303–1306. doi: 10.1126/science.1975955. [DOI] [PubMed] [Google Scholar]
- 23.Guastella J, Brecha NC, Weigman C, Lester HA, Davidson N. Cloning, expression, and localization of a rat brain high-affinity glycine transporter. Proc Natl Acad Sci USA. 1992;89:7189–7193. doi: 10.1073/pnas.89.15.7189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Houser CR, Vaughn GE, Hendry SHC, Jones EG, Peters A (1984) GABA neurons in the cerebral cortex. In: Cerebral cortex, Vol 2, Functional properties of cortical cells (Jones EG, Peters A, eds), pp 63–90. New York: Plenum.
- 25.Hsu SM, Raine L, Fanger H. Use of avidin-biotin-peroxidase complex (ABC) in immunoperoxidase techniques: a comparison between ABC and unlabelled antibody (PAP) procedures. J Histochem Cytochem. 1981;29:557–580. doi: 10.1177/29.4.6166661. [DOI] [PubMed] [Google Scholar]
- 26.Ikegaki N, Saito N, Hashima M, Tanaka C. Production of specific antibodies against GABA transporter subtypes (GAT-1, GAT-2, GAT-3) and their application to immunocytochemistry. Mol Brain Res. 1994;26:47–54. doi: 10.1016/0169-328x(94)90072-8. [DOI] [PubMed] [Google Scholar]
- 27.Isaacson JS, Solis JM, Nicoll RA. Local and diffuse synaptic actions of GABA in the hippocampus. Neuron. 1993;10:165–175. doi: 10.1016/0896-6273(93)90308-e. [DOI] [PubMed] [Google Scholar]
- 28.Iversen LL. Role of transmitter uptake mechanisms in synaptic neurotransmission. Br J Pharmacol. 1971;41:571–591. doi: 10.1111/j.1476-5381.1971.tb07066.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Iversen LL, Kelly JS. Uptake and metabolism of γ-aminobutyric acid by neurons and glial cells. Biochemical Pharmacol. 1975;24:933–938. doi: 10.1016/0006-2952(75)90422-0. [DOI] [PubMed] [Google Scholar]
- 30.Iversen LL, Neal MJ. The uptake of [3H]GABA by slices of rat cerebral cortex. J Neurochem. 1968;15:1141–1149. doi: 10.1111/j.1471-4159.1968.tb06831.x. [DOI] [PubMed] [Google Scholar]
- 31.Iversen LL, Snyder SH. Synaptosomes: different population storing catecholamines and gamma-aminobutyric acid in homogenate of rat brain. Nature. 1968;220:796–798. doi: 10.1038/220796a0. [DOI] [PubMed] [Google Scholar]
- 32.Kanner BI, Bendahan A. Two pharmacologically distinct sodium- and chloride-coupled high-affinity γ-aminobutyric acid transporters are present in plasma membrane vesicles and reconstituted preparations from rat brain. Proc Natl Acad Sci USA. 1990;87:2550–2554. doi: 10.1073/pnas.87.7.2550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Keynan S, Suh Y-J, Kanner BI, Rudnick G. Expression of a cloned γ-aminobutyric acid transporter in mammalian cells. Biochemistry. 1992;31:1974–1979. doi: 10.1021/bi00122a011. [DOI] [PubMed] [Google Scholar]
- 34.Krogsgaard-Larsen P, Falch E, Larsson OM, Schousboe A. GABA uptake inhibitors: relevance to antiepileptic drug research. Epilepsy Res. 1987;1:77–93. doi: 10.1016/0920-1211(87)90012-x. [DOI] [PubMed] [Google Scholar]
- 35.Larsson OM, Johnston GAR, Schousboe A. Differences in uptake kinetics of cis -3-aminocyclohexane carboxylic acid into neurons and astrocytes in primary cultures. Brain Res. 1983;260:279–285. doi: 10.1016/0006-8993(83)90681-9. [DOI] [PubMed] [Google Scholar]
- 36.Levi G, Raitieri M. Carrier-mediated release of neurotransmitters. Trends Neurosci. 1993;16:415–418. doi: 10.1016/0166-2236(93)90010-j. [DOI] [PubMed] [Google Scholar]
- 37.Liu Q-R, Lopez-Còrcuera B, Mandiyan S, Nelson H, Nelson N. Molecular characterization of four pharmacologically distinct γ-aminobutyric acid transporters in mouse brain. J Biol Chem. 1993;268:2106–2112. [PubMed] [Google Scholar]
- 38.Mabjeesh NJ, Frese M, Rauen T, Jeserich G, Kanner BI. Neuronal and glial γ-aminobutyric acid transporters are distinct proteins. FEBS Lett. 1992;299:99–102. doi: 10.1016/0014-5793(92)80109-t. [DOI] [PubMed] [Google Scholar]
- 39.Minelli A, Brecha NC, Karschin C, DeBiasi S, Conti F. GAT-1, a high-affinity GABA plasma membrane transporter, is localized to neurons and astroglia in the cerebral cortex. J Neurosci. 1995;15:7734–7746. doi: 10.1523/JNEUROSCI.15-11-07734.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Neal MJ, Bowery NG. Cis -3-aminocyclohexane carboxylic acid: a substrate for the neuronal GABA transport system. Brain Res. 1977;138:169–174. doi: 10.1016/0006-8993(77)90793-4. [DOI] [PubMed] [Google Scholar]
- 41.Neal MJ, Iversen LL. Subcellular distribution of endogenous and [3H]γ-aminobutyric acid in rat cerebral cortex. J Neurochem. 1969;16:1245–1252. doi: 10.1111/j.1471-4159.1969.tb05972.x. [DOI] [PubMed] [Google Scholar]
- 42.Peters A, Palay SL, Webster H deF H deF. Oxford UP; New York: 1991. The fine structure of the nervous system: neurons and their supporting cells. [Google Scholar]
- 43.Pin J-P, Bockaert J. Two distinct mechanisms, differentially affected by excitatory amino acids, trigger GABA release from fetal mouse striatal neurons in primary cultures. J Neurosci. 1989;9:648–656. doi: 10.1523/JNEUROSCI.09-02-00648.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Ribak CE. Aspinous and sparsely-spinous stellate neurons in the visual cortex of rat contain glutamic acid decarboxylase. J Neurocytol. 1978;7:461–478. doi: 10.1007/BF01173991. [DOI] [PubMed] [Google Scholar]
- 45.Ribak CE, Tong W, Brecha NC. The GABA plasma membrane transporters, GAT-1 and GAT-3, display different distributions in the rat hippocampus. J Comp Neurol. 1996;367:595–606. doi: 10.1002/(SICI)1096-9861(19960415)367:4<595::AID-CNE9>3.0.CO;2-#. [DOI] [PubMed] [Google Scholar]
- 46.Rothstein JD, Martin L, Levey AI, Dykes-Hoberg M, Jin L, Wu D, Nash M, Kuncl RW. Localization of neuronal and glial glutamate transporters. Neuron. 1994;13:713–725. doi: 10.1016/0896-6273(94)90038-8. [DOI] [PubMed] [Google Scholar]
- 47.Schon F, Kelly JS. Selective uptake of [3H]β-alanine by glia: association with the glial uptake system for GABA. Brain Res. 1975;86:243–257. doi: 10.1016/0006-8993(75)90700-3. [DOI] [PubMed] [Google Scholar]
- 48.Schousboe A, Larsson OM, Wood JD, Krogsgaard-Larsen P. Transport and metabolism of γ-aminobutyric acid in neurons and glia: implications for epilepsy. Epilepsia. 1983;24:531–538. doi: 10.1111/j.1528-1157.1983.tb03417.x. [DOI] [PubMed] [Google Scholar]
- 49.Schwartz EA. Calcium-independent release of GABA from isolated horizontal cells of the toad retina. J Physiol (Lond) 1982;323:211–227. doi: 10.1113/jphysiol.1982.sp014069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Thomas CT, McNamee MG. Purification of membrane proteins. Methods Enzymol. 1990;182:499–520. doi: 10.1016/0076-6879(90)82040-9. [DOI] [PubMed] [Google Scholar]
- 51.Thompson SM, Gahwiler BH. Effects of the GABA uptake inhibitor tiagabine on inhibitory synaptic potentials in rat hippocampal slice cultures. J Neurophysiol. 1992;67:1698–1670. doi: 10.1152/jn.1992.67.6.1698. [DOI] [PubMed] [Google Scholar]
- 52.Towbin H, Staehelin T, Gordon J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci USA. 1979;76:4350–4354. doi: 10.1073/pnas.76.9.4350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Yamauchi A, Uchida S, Kwon HM, Preston AS, Robey RB, Garcia-Perez A, Burg MB, Handler JS. Cloning of a Na+- and Cl−-dependent betaine transporter that is regulated by hypertonicity. J Biol Chem. 1992;267:649–652. [PubMed] [Google Scholar]
- 54.Zilles K. Springer; Berlin: 1985. The cortex of rat. . [Google Scholar]