

Abstract
Objective
To test whether targeting hippocampal-cortical brain networks with high-frequency transcranial magnetic stimulation in older adults influences behavioral and neural measures characteristic of age-related memory impairment.
Methods
Fifteen adults aged 64 to 80 years (mean = 72 years) completed a single-blind, sham-controlled experiment. Stimulation targets in parietal cortex were determined based on fMRI connectivity with the hippocampus. Recollection and recognition memory were assessed after 5 consecutive daily sessions of full-intensity stimulation vs low-intensity sham stimulation using a within-subjects crossover design. Neural correlates of recollection and recognition memory formation were obtained via fMRI, measured within the targeted hippocampal-cortical network vs a control frontal-parietal network. These outcomes were measured approximately 24 hours after the final stimulation session.
Results
Recollection was specifically impaired in older adults compared to a young-adult control sample at baseline. Relative to sham, stimulation improved recollection to a greater extent than recognition. Stimulation increased recollection fMRI signals throughout the hippocampal-cortical network, including at the targeted location of the hippocampus. Effects of stimulation on fMRI recollection signals were greater than those for recognition and were greater in the targeted network compared to the control network.
Conclusions
Age-related recollection impairments were causally related to hippocampal-cortical network function in older adults. Stimulation selectively modified neural and behavioral hallmarks of age-related memory impairment, indicating effective engagement of memory intervention targets in older adults.
Memory decline is the predominant cognitive complaint in older adults, is indicative of mild cognitive impairment, and presages conversion to Alzheimer disease.1 Memory decline during normal aging is most severe for recollection of associative information, as typically measured using paired-associate tests, with relative preservation of other types of memory, such as recognition of individual items.2 Age-related recollection decline is thought to arise from disruption of a distributed cortical network interacting with the hippocampus.3,4
Multisession high-frequency transcranial magnetic stimulation (TMS) targeting the hippocampal-cortical network improves recollection and influences network function in young adults.5–7 However, it is unknown whether these methods are suitable for older adults, given that structural and/or functional changes with age could diminish the ability to generate network-level responses to stimulation.
We performed a pilot study to determine whether it is possible to rescue age-related recollection impairments and alter related brain activity in older adults. We hypothesized that stimulation would improve recollection accuracy and increase fMRI activity signals of recollection memory formation in the targeted network. We hypothesized that these effects would be selective, with greater effects for recollection than for recognition, with more coherent6 recollection fMRI activity in the targeted hippocampal-cortical network,7 without effects in a control frontal-parietal network.
Methods
Fifteen cognitively normal older adults (mean age 72.46 years, age range 64–80 years, 11 women; data available from Dryad, table e-1, doi.org/10.5061/dryad.p2qq2d0) completed a sham-controlled, single-blind, counterbalanced experiment. TMS was applied to participant-specific lateral parietal locations based on their fMRI connectivity with hippocampus (figure 1A).5–7 fMRI was acquired while participants learned objects paired with scenes and with locations (data available from Dryad, figure e-1, doi.org/10.5061/dryad.p2qq2d0). Recognition of objects and recollection of object associations were assessed before, approximately 24 hours after, and 1 week after full-intensity stimulation and low-intensity sham (figure 1B). See data available from Dryad (supplementary materials, doi.org/10.5061/dryad.p2qq2d0).
Figure 1. Stimulation increased recollection accuracy.

(A) Participant-specific stimulation locations (one sphere shown per participant) were selected based on high seed-based resting-state fMRI connectivity with hippocampal target locations (one sphere shown per participant). (B) Before and approximately 24 hours after 5 consecutive daily sessions of full-intensity or sham stimulation, participants completed fMRI memory assessments. Stimulation and sham were administered in all participants in counterbalanced order using a within-subjects crossover design. Approximately 1 week following the post-stim and post-sham sessions, participants returned for follow-up assessments. Stimulation-induced electrical field for each stimulation condition is displayed for a representative participant with warmer colors representing peak intensity (range: 1–119 V/m). (C) Effects of stimulation on recollection and on recognition calculated for stimulation vs sham at the 24-hour assessment. (D) Recollection changes due to stimulation and sham. (E) Each bar represents a single-participant change in recollection and recognition for stimulation and sham, demonstrating consistent improvement due to stimulation particularly for recollection. Error bars indicate SEM. *p ≤ 0.05, **p ≤ 0.01.
Data availability
Data are archived in a repository publicly available with registration at nunda.northwestern.edu.
Standard protocol approvals, registrations, and patient consents
All participants gave written informed consent. The institutional review board at Northwestern University approved all study procedures.
Results
Relative to a sample of younger adults performing the same task,6 older adults in the current experiment were impaired prior to receiving stimulation for recollection (t30 = 2.17, p = 0.038, Cohen d = 0.77) but not for recognition (t30 = 0.28, p = 0.78). Stimulation improved older adults' recollection more than recognition at the 24-hour assessment relative to sham (t14 = 2.77, p = 0.02, Cohen d = 0.71; figure 1C). There was robust recollection improvement (t14 = 3.25, p < 0.01, Cohen d = 0.84) and weak recognition improvement (t14 = 2.25, p = 0.04, Cohen d = 0.58). Relative to baseline, the recollection improvement due to stimulation was 31.1% on average (t14 = 3.10, p < 0.01, Cohen d = 0.80; figure 1D) with nonsignificant change of −3.1% due to sham (t14 = 0.38, p = 0.71). In contrast, there was nonsignificant recognition improvement of 2.8% (t14 = 1.55, p = 0.14) and with nonsignificant change of −2.9% due to sham (t14 = 1.57, p = 0.14). Recollection improvements were highly consistent across participants due to stimulation but not for sham (figure 1E). Following stimulation, recollection no longer differed significantly for older adults relative to younger adults in the control sample (t30 = 0.78, p = 0.44).
The same patterns of effects were identified when analyzed using raw values rather than percent-change values. Stimulation did not affect performance of a battery of other cognitive abilities (data available from Dryad, tables e-2 and e-3, doi.org/10.5061/dryad.p2qq2d0), with no indication of impairment for any measure. Recollection remained significantly elevated at approximately 1-week follow-up testing relative to baseline but without significant differentiation from sham, indicating that significant gains measured at 24 hours did not persist relative to sham (data available from Dryad, supplementary materials, doi.org/10.5061/dryad.p2qq2d0).
fMRI activity correlates of memory formation were measured for the targeted hippocampal-cortical network and a control frontal-parietal network, defined a priori based on previous reports of age-related impairments4 (figure 2A). Stimulation increased recollection more than recognition activity in the targeted vs the control network (t14 = 2.10, p = 0.05, Cohen d = 0.54; figure 2B). Relative to sham, activity increases due to stimulation were greater for recollection than for recognition in the targeted network (t14 = 2.14, p = 0.05, Cohen d = 0.55; figure 2B) but not in the control network (t14 = 0.29, p = 0.78), reflecting significant and consistent increase in recollection activity for the targeted network (t14 = 2.38, p = 0.03, Cohen d = 0.61) but not the control network (t14 = 0.55, p = 0.59), relative to sham (figure 2C). Recognition activity did not change significantly in either network (targeted: t14 = 1.71, p = 0.11; control: t14 = 0.07, p = 0.95). Furthermore, stimulation did not significantly affect recollection activity locally for the stimulation location (data available from Dryad, supplementary materials, doi.org/10.5061/dryad.p2qq2d0). Thus, stimulation selectively increased fMRI activity of recollection memory formation throughout the targeted hippocampal-cortical network, relative to recognition and to the control network.
Figure 2. Stimulation coherently increased hippocampal-cortical network activity during memory formation.

(A) Mean fMRI activity was extracted from a priori selected network regions of the targeted hippocampal-cortical network and the control frontal-parietal network during object-location memory formation. (B) Mean fMRI activity changes (post-stim vs post-sham) for recollection compared to recognition in the targeted network vs the control network, and for recollection alone in the targeted and control networks. (C) Each bar is a single participant's mean recollection activity change in the targeted and control networks. (D) Mean recollection fMRI activity changes (post-stim vs post-sham) for each region of interest (ROI) in each network (see data available from dryad, table e-4, doi.org/10.5061/dryad.p2qq2d0, for region labels and for raw values for the post-stim and post-sham conditions). (E) Mean coherence change due to stimulation within the targeted network, the control network, and between the targeted and control networks. Error bars indicate SEM. *p ≤ 0.05.
Changes in activity coherence6 evaluated the consistency of stimulation effects among regions comprising each network, relative to sham (figure 2, D and E). Stimulation increased coherence within the targeted network (t7 = 2.85, p = 0.02). This increase was greater than coherence changes between the targeted and control networks (t24 = 2.68, p = 0.01). Thus, stimulation coherently increased fMRI signals of recollection memory formation among regions comprising the targeted network, and these activity changes were decoupled from the control network.
Following previous evidence for effects of stimulation on the targeted hippocampal segment,7 stimulation increased fMRI activity at the a priori hippocampal target (figure 1A) for recollection (t14 = 2.90, p = 0.01, Cohen d = 0.75) but not for recognition (t14 = 0.43, p = 0.68). To assess spatial selectivity, stimulation effects were assessed along sequential segments of the anterior and posterior hippocampus relative to the target in each hemisphere (figure 3A). For the left hippocampus, recollection activity was greater following stimulation than sham (F1,14 = 11.06, pcorr = 0.02, ηp2 = 0.44) and this relative increase varied by segment (F8, 112 = 2.74, pcorr = 0.03, ηp2 = 0.16). Post hoc tests indicated significantly increased activity at the target and posterior hippocampus segments for stimulation relative to sham (figure 3B). In contrast, there were no effects for recollection in the right hippocampus or for recognition in the left or right hippocampus (data available from Dryad, supplementary materials, doi.org/10.5061/dryad.p2qq2d0).
Figure 3. Stimulation increased hippocampal activity during memory formation.
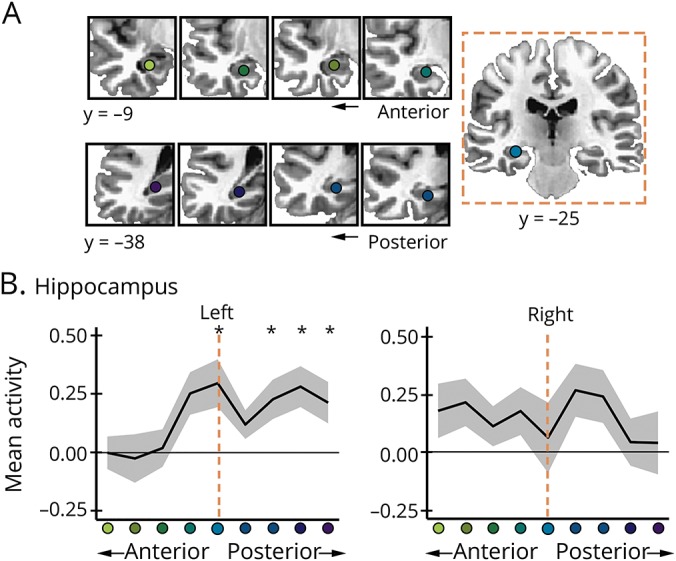
(A) Segments along the anterior-posterior long axes of the left and right hippocampus relative to the average a priori target in the left hippocampus (y = −25). (B) Mean difference in recollection fMRI activity for each segment post-stim minus post-sham. Error bars indicate SEM. *p ≤ 0.05.
Discussion
Network-targeted TMS selectively improved recollection and increased fMRI correlates of recollection memory formation in the hippocampal-cortical network. These effects on recollection were greater than the effects on recognition and there were no effects on fMRI activity in the control frontal-parietal network. Stimulation effects on recollection fMRI signals were coherent throughout the hippocampal-cortical network and occurred at the targeted location of the hippocampus. This is notable given that recollection is strongly associated with hippocampal2 and hippocampal-cortical network function.8 These findings demonstrate a causal link between recollection and the hippocampal-cortical network in older adults, thereby supporting the hypothesis1,3,4 that dysfunction of this network with age causes memory impairment. Although recognition may have been less sensitive to stimulation than recollection because of differences in difficulty or performance, these findings nonetheless indicate that the hallmark recollection deficits of aging were improved by stimulation.
In animal models, improved memory function has been associated with greater hippocampal excitability, especially in aging.9 It is possible that increased activity due to stimulation indicated heightened network-specific excitability (i.e., greater response to the same category of visual stimuli). Although stimulation rescued older adults' recollection impairment such that they no longer differed significantly from younger adults, further testing is needed to test an increased excitability hypothesis and to evaluate efficacy in individuals with more severe damage of the hippocampal-cortical network.
This study was intended to test for selective neural and behavioral target engagement, not to evaluate clinical efficacy. The participant sample was relatively small, all testing was performed at the same site, and there was no active control stimulation. However, we formulated strong a priori hypotheses with high specificity to age-related decline and identified specific and reliable effects of stimulation across participants, consistent with previous findings in young adults.5–7 Furthermore, effects were measured after approximately 24 hours to reduce potential confounds associated with the subjective experience of stimulation. In comparison to other approaches in older adults in which effects are measured during or immediately after stimulation,10 this regimen produced long-lasting improvements in targeted memory function, which is of practical significance given the varying and unpredictable memory demands of daily living.
Network-targeted noninvasive stimulation can selectively engage behavioral and neural characteristics of age-related memory impairment. These findings motivate future studies to optimize the effectiveness of noninvasive stimulation for treatment of age-related memory impairment and to improve mechanistic understanding of the hippocampal-cortical networks that support episodic memory across the lifespan.
Acknowledgment
The authors thank Donna Bridge, Brennan Durr, Elise Gagnon, Melissa Gunlogson, Molly Hermiller, Sungshin Kim, Valerie McDonald, Melissa McSweeney, Jonathan O'Neil, Robert Palumbo, John Walker, Mallory Ward, and Kristen Warren for their support of this project. Neuroimaging was performed at the Northwestern University Center for Translational Imaging, supported by Northwestern University Department of Radiology. This research was also supported in part through the computational resources and staff contributions provided for Quest, the high-performance computing facility at Northwestern University, and by resources provided by Northwestern University Clinical and Translational Sciences Institute, funded by the National Center for Advancing Translational Sciences (UL1TR001422).
Glossary
- TMS
transcranial magnetic stimulation
Appendix. Authors
Study funding
This research was funded by the National Institute on Aging (T32AG20506, F31AG057109, R01AG049002) and a pilot project grant from the Northwestern University Cognitive Neurology and Alzheimer's Disease Center (P30AG13854). None of the sponsoring organizations participated in the design, conduct, analysis, manuscript preparation, or decision to publish. The content of this manuscript is solely the responsibility of the authors and does not necessarily represent the official view of the NIH.
Disclosure
The authors report no disclosures relevant to the manuscript. Go to Neurology.org/N for full disclosures.
References
- 1.Brayne C. The elephant in the room—healthy brains in later life, epidemiology and public health. Nat Rev Neurosci 2007;8:233–239. [DOI] [PubMed] [Google Scholar]
- 2.Yonelinas AP. The nature of recollection and familiarity: a review of 30 years of research. J Mem Lang 2002;46:441–517. [Google Scholar]
- 3.Chhatwal JP, Sperling RA. Functional MRI of mnemonic networks across the spectrum of normal aging, mild cognitive impairment, and Alzheimer's disease. J Alzheimers Dis 2012;31(suppl 3):S155–S167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Andrews-Hanna JR, Snyder AZ, Vincent JL, et al. Disruption of large-scale brain systems in advanced aging. Neuron 2007;56:924–935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nilakantan AS, Bridge DJ, Gagnon EP, VanHaerents SA, Voss JL. Stimulation of the posterior cortical-hippocampal network enhances precision of memory recollection. Curr Biol 2017;27:465–470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kim S, Nilakantan AS, Hermiller MS, Palumbo RT, VanHaerents S, Voss JL. Selective and coherent activity increases due to stimulation indicate functional distinctions between episodic memory networks. Sci Adv 2018;4:eaar2768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wang JX, Rogers LM, Gross EZ, et al. Targeted enhancement of cortical-hippocampal brain networks and associative memory. Science 2014;345:1054–1057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.King DR, de Chastelaine M, Elward RL, Wang TH, Rugg MD. Recollection-related increases in functional connectivity predict individual differences in memory accuracy. J Neurosci 2015;35:1763–1772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Yu XW, Oh MM, Disterhoft JF. CREB, cellular excitability, and cognition: implications for aging. Behav Brain Res 2017;322:206–211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hsu WY, Ku Y, Zanto TP, Gazzaley A. Effects of noninvasive brain stimulation on cognitive function in healthy aging and Alzheimer's disease: a systematic review and meta-analysis. Neurobiol Aging 2015;36:2348–2359. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data are archived in a repository publicly available with registration at nunda.northwestern.edu.