

Abstract
Pathogenic Salmonella strains that cause gastroenteritis are able to colonize and replicate within the intestines of multiple host species. In general, these strains have retained an ability to form the rdar morphotype, a resistant biofilm physiology hypothesized to be important for Salmonella transmission. In contrast, Salmonella strains that are host-adapted or even host-restricted like Salmonella enterica serovar Typhi, tend to cause systemic infections and have lost the ability to form the rdar morphotype. Here, we investigated the rdar morphotype and CsgD-regulated biofilm formation in two non-typhoidal Salmonella (NTS) strains that caused invasive disease in Malawian children, S. Typhimurium D23580 and S. Enteritidis D7795, and compared them to a panel of NTS strains associated with gastroenteritis, as well as S. Typhi strains. Sequence comparisons combined with luciferase reporter technology identified key SNPs in the promoter region of csgD that either shut off biofilm formation completely (D7795) or reduced transcription of this key biofilm regulator (D23580). Phylogenetic analysis showed that these SNPs are conserved throughout the African clades of invasive isolates, dating as far back as 80 years ago. S. Typhi isolates were negative for the rdar morphotype due to truncation of eight amino acids from the C-terminus of CsgD. We present new evidence in support of parallel evolution between lineages of nontyphoidal Salmonella associated with invasive disease in Africa and the archetypal host-restricted invasive serovar; S. Typhi. We hypothesize that the African invasive isolates are becoming human-adapted and ‘niche specialized’ with less reliance on environmental survival, as compared to gastroenteritis-causing isolates.
Author summary
African clades of nontyphoidal Salmonella cause invasive disease on a daily basis and thousands of deaths each year. Although it is generally accepted that the transmission route for these organisms is fecal-oral, we know very little about their behaviour in the environment between hosts. In this paper, we have identified both a genotype and a phenotype that suggest environmental niche specialization that is distinct from lineages of Salmonella Typhimurium and Salmonella Enteritidis associated with industrialized food supply chains in resource-rich settings. We also compared with strains of Salmonella Typhi, which cause systemic typhoid fever infections exclusively in humans. In each invasive lineage, regulatory or structural gene mutations leading to loss or impairment of biofilm were identified, all associated with curli and cellulose production, the two main structures that comprise the biofilm matrix. This suggests that similar evolutionary pressures are acting on invasive Salmonella isolates. Public health strategies aimed at reducing the burden of invasive Salmonella disease must prevent transmission to vulnerable adults and children via water sanitation and hygiene practices–a process that starts with identification of environmental reservoirs. The results of our study will raise the profile of this neglected aspect of invasive salmonellosis and will challenge researchers and clinicians to search in new places for potential environmental reservoirs of these pathogens.
Introduction
The 2,600 serovars of the genus Salmonella have considerable genetic diversity, which permits them to occupy a wide variety of environmental and animal niches and to cause clinical presentation in humans ranging from asymptomatic carriage through enterocolitis and invasive disease. Most cases of human disease are caused by a few serovars of Salmonella enterica, which are loosely categorized as being invasive/typhoidal (serovars Typhi and Paratyphi A) or nontyphoidal. The nontyphoidal salmonellae (NTS) typically cause self-limiting enterocolitis and include common serovars such as Salmonella Typhimurium and Salmonella Enteritidis [1,2].
This simple clinical distinction breaks down in settings where there is high prevalence of immunosuppressive illness, such as sub-Saharan Africa (sSA). Here, NTS have emerged as a leading cause of bacterial bloodstream infection [3], or invasive nontyphoidal Salmonella (iNTS) disease. In common with typhoid fever, iNTS disease frequently presents without diarrheal symptoms, with non-focal febrile illness being the dominant clinical presentation [4]. This disease is responsible for an estimated 681,000 deaths per year, with nonspecific symptomology, multidrug resistance, and poor clinical outcomes despite correct diagnosis contributing to this significant mortality rate [5]. There is great urgency to better understand iNTS disease and reduce its impact in Africa and other areas of the world [6].
Most NTS infect a wide range of host species and are considered host-generalist pathogens [7]. In contrast, typhoidal serovars are exemplars of host-restricted pathogens, with marked genomic degradation in comparison to NTS serovars [8]. There is evidence that the same process is underway in strains of S. Typhimurium and S. Enteritidis associated with invasive disease in sub-Saharan Africa [9,10]. Many of these gene mutations affect metabolic processes involved in anaerobic respiration on unique carbon sources, a mechanism that is pivotal for the replication and outgrowth of Salmonella in the inflamed intestinal tract [11,12]. Whether this represents a random process related to the different geographical location of these strains, convergent evolution with the typhoidal serovars towards an invasive rather than an enteric “lifestyle”, or adaptation to distinct environmental niches remains an outstanding question.
Irrespective of invasive versus enteric lifestyle, all salmonellae are transmitted via the fecal-oral route, but key questions remain over how invasive strains interact with the environment between host colonization events [13,14]. Biofilm formation is proposed to aid in the survival and persistence of Salmonella cells during this environmental phase of the transmission cycle [15]. The most well-characterized format of Salmonella biofilm physiology has been termed the rdar (red, dry, and rough) morphotype, where a self-produced extracellular matrix interconnects cells and facilitates their adherence to abiotic and biotic surfaces [16–18]. Multiple cues, including ambient temperature, osmolarity, and nutrient availability, act via a complex regulatory network to activate CsgD, a member of the RpoS regulon and the primary transcriptional activator of the rdar morphotype [15,16]. CsgD in turn induces the expression of proteinaceous (curli fimbriae [19]), BapA [20]) and polysaccharide (cellulose [17,21], O-antigen capsule [18,22]) polymers that act as major contributors to the recalcitrant matrix structure associated with this phenotype. The genes for curli fimbriae and cellulose are highly conserved in Salmonella [23–25]; however, almost all Typhi and Paratyphi isolates are phenotypically negative for the rdar morphotype [23,26]. Therefore, loss of the rdar morphotype may represent an additional signature of host adaptation.
Landmark studies revealed genomic degradation within representative iNTS strains S. Typhimurium D23580 [27] and S. Enteritidis D7795 [10]–these observations were consistent with genetic signatures of host adaptation found in typhoidal and paratyphoidal Salmonella species [28,29]. As such, significant attention has focused on evaluating the virulence of these strains in laboratory models of Salmonella infection [9,30–32]. Comparatively less is known about the state of biofilm physiology of iNTS strains [33], and in particular the CsgD-regulated rdar morphotype, which we and other labs have postulated may be important for the persistence of nontyphoidal Salmonella species in non-host environments [15,23,25,26,34].
We wanted to determine if S. Typhimurium D23580 and S. Enteritidis D7795 can form biofilms, and if not, to determine if the corresponding mutations are conserved in other African strains associated with invasive disease. We selected typical Typhimurium and Enteritidis strains that cause enterocolitis to act as controls in our phenotypic screens and to rule out any serovar-specific genetic variations that do not affect biofilm formation. We included the archetypal Typhi strain (CT18) and modern versions of the H58 haplotype [35] to determine if patterns of gene loss or mutation are shared between invasive isolates.
Results
Evaluation of the biofilm phenotype for invasive nontyphoidal and typhoidal Salmonella strains
The rdar morphotype was first described by Römling and colleagues during their research into the expression of curli fimbriae in S. Typhimurium 14028 [16]. For the wildtype strain, the morphotype was reported as a temperature-restricted phenotype, with transcription of curli biosynthesis (csg) genes and characteristic patterns on the colony surface constrained to cells grown at 28˚C in nutrient-rich medium of low osmolarity. These conditions have since been used as a standard for screening Salmonella isolates for the rdar morphotype [26]. After three to five days of growth at 28˚C, nontyphoidal and typhoidal Salmonella strains were easily distinguishable based on the different appearances of their macrocolonies (Fig 1). We added Congo red dye to the agar to visualize the presence of curli fimbriae and cellulose and identify strains that possess the red, dry, and rough (rdar) morphotype [19]. Control strains of NTS were rdar-positive and red in colour, while S. Typhi strains were smooth and white (saw), or rdar-negative. S. Typhimurium D23580 formed pale red colonies with an incomplete wrinkling pattern on the colony’s surface, whereas S. Enteritidis D7795 was rdar-negative. Temperature-based restriction of curli expression can be overcome in some instances; examples of this include when strains carry certain mutations in the csgD promoter [16,36], or when nontyphoidal Salmonella are grown in iron-limiting conditions [16] or in the presence of human bile [37]. However, all NTS strains displayed a temperature-based restriction of curli expression [16], as they did not form rdar colonies at 37°C and curli (csgBAC) transcription was at basal levels (S1 Fig).
Fig 1. The biofilm phenotypes of representative nontyphoidal and typhoidal Salmonella strains.

Salmonella strains that are known to cause gastroenteritis or enterocolitis (green), invasive disease (blue) or typhoid fever (red) were screened: for the ability to form the red, dry, and rough (rdar) morphotype, which presents as the formation of concentric rings and a wrinkled appearance on the surface of macrocolonies (panels 1 and 2, top); for the presence of multicellular, biofilm aggregates and planktonic cells in liquid cultures grown under biofilm-inducing conditions (middle panel; conical tubes); and for cellulose production, visualized as the white and fluorescent appearance of macrocolonies in the presence of calcofluor white dye (bottom panel).
Rdar morphotype-positive Salmonella strains grown in an in vitro flask model of biofilm development will produce two unique subpopulations—multicellular, biofilm aggregates and planktonic cells [38,39]—due to the bistable production of CsgD [40]. We reasoned that multicellular aggregation in liquid cultures would provide a clearer diagnostic for the presence of a functioning biofilm phenotype in both strong and moderate biofilm-producing strains. Cultures of S. Typhimurium D23580 contained visible multicellular aggregates and planktonic cell subpopulations (Fig 1). However, the aggregates made up a lower proportion of the population biomass compared to aggregates from other NTS and had observable differences in their physical structure (S2 Fig), suggesting a moderate but impaired biofilm phenotype. In contrast, cultures of S. Enteritidis D7795 and all typhoidal Salmonella strains consisted solely of planktonic cells. Cellulose production can be specifically assessed by supplementing T agar with calcofluor white dye and illuminating colonies with ultraviolet light [25]. Similar fluorescence intensity was observed for S. Typhimurium D23580 compared to other NTS, indicating the presence of cellulose production despite the partial rdar morphotype (Fig 1). In contrast, cellulose production was comparably minimal or absent from macrocolonies of S. Enteritidis D7795 and all S. Typhi isolates.
Status of CsgD and curli production and RpoS synthesis and activity
The divergent csg operons are central to curli biosynthesis and include genes for curli fimbrial protein subunits (csgBA), transcriptional regulation (csgD), and curli assembly machinery (csgC and csgEFG) [41–43]. CsgD is involved in activating the transcription of both operons [16,19], making detection of curli fimbrial subunits a strong indicator of CsgD activity. We probed the lysates of cell subpopulations emerging from flask cultures for synthesis of CsgD and CsgA, the major subunit of curli fimbriae (Fig 2). We detected both proteins in lysates from multicellular aggregates, including S. Typhimurium D23580. In contrast, neither protein was detected for samples derived from S. Enteritidis D7795 or any of the S. Typhi strains.
Fig 2. Detection of CsgD and CsgA protein synthesis in representative Salmonella strains by Western blot.
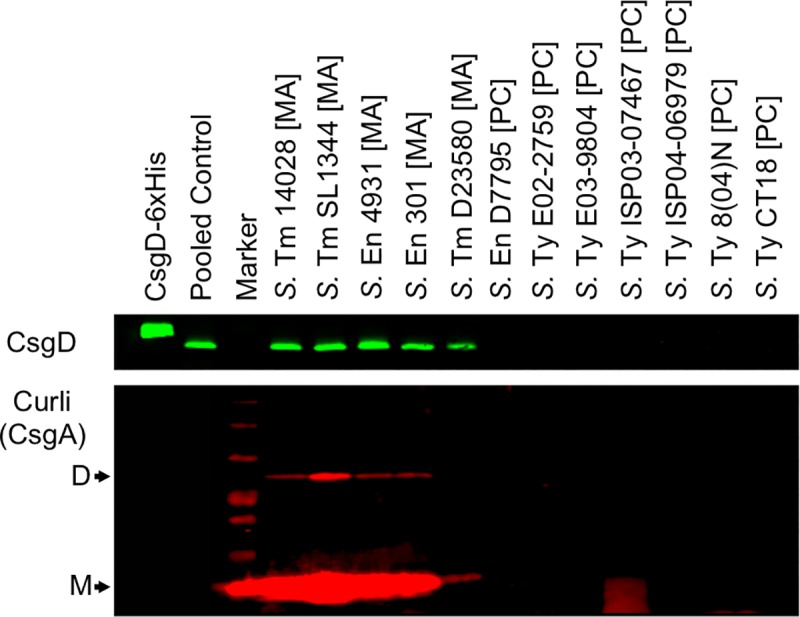
Whole-cell lysates were derived from multicellular aggregates [MA] or planktonic cells [PC] harvested from flask cultures of nontyphoidal and typhoidal Salmonella. Lysates used for CsgD detection (top panel) were normalized based on total protein concentration. Purified CsgD-6xHis recombinant protein was used as a technical control for CsgD detection. Pooled control samples were derived from combining lysates obtained from the multicellular aggregates of S. Typhimurium 14028 and SL1344 and S. Enteritidis 4931 and 301 strains. Black arrows indicate the detection of CsgA subunit dimers (D) and monomers (M) (bottom panel). The data shown is representative of two biological replicates.
We also probed the lysates for RpoS, the sigma factor that controls csgD expression [19] (S3A Fig). RpoS protein was detected in all nontyphoidal Salmonella strains, including both biofilm and planktonic cell subpopulations derived from biofilm-producing strains. S. Enteritidis D7795 appeared to have lower levels of RpoS compared to the other NTS strains. For the S. Typhi strains, there was significant variation in protein levels at the 24-hour time point used for sampling, with low levels of RpoS observed for S. Ty E03-9804 and S. Ty 8(04)N. Bacterial luciferase reporter technology allows for the systematic comparison of transcriptional activity across a wide variety of strains [24,44]. To evaluate RpoS activity, we tracked the expression of a synthetic, RpoS-dependent promoter-luciferase reporter construct in each strain during growth in microbroth cultures [15] (S3B–S3D Fig). All strains in our panel exceeded the standard RpoS activity threshold identified in our previous work with other Salmonella strains (i.e. 10,000 CPS) [24], except for S. Enteritidis D7795.
Screening for serovar- and strain-specific cis or trans variation in biofilm gene regulation
We hypothesized that variation in csgD expression levels could account for differences in the biofilm phenotypes we observed. The sequence of the 755-bp intergenic region between the csgBAC and csgDEFG operons was compared between each strain in our panel (Fig 3A). For S. Enteritidis D7795 and S. Typhimurium D23580, unique single nucleotide polymorphisms (SNPs) were identified within the intergenic region (Fig 3B). For S. Enteritidis D7795, a C-to-T SNP was observed in the regulatory region 47 bp upstream of the csgD transcriptional start site. For S. Typhimurium D23580, independent C-to-A and G-to-A transversion mutations were identified 80 bp and 189 bp upstream of the csgD transcriptional start site.
Fig 3. Evaluation of the csgDEFG-csgBAC intergenic region for changes in promoter sequence and activity.

(A) Diagram representing the divergent csg operons. Transcriptional start sites are indicated as black elbow arrows. The square bracket indicates the region analyzed in (B). (B) Multiple sequence alignment of the intergenic and 5ʹ untranslated regions for the csgDEFG and csgBAC operons from Salmonella strains in this study. A neighbour-joining dendrogram was established based on bootstrapping parameters set to 1,000 replicates and a support threshold of 70%. Transcription factor binding sites that have been experimentally verified in Salmonella are indicated above the consensus sequence. Single nucleotide polymorphisms (SNPs) within each strain’s DNA sequence are indicated as coloured rectangles within grey tracks (red, adenine; blue, cytosine; yellow, guanine; green, thymine). Red stars and yellow vertical blocks highlight SNP positions unique to S. Typhimurium D23580 and S. Enteritidis D7795. (C) Promoter-reporter fusion constructs were generated using csgD and csgB promoter sequences derived from the Salmonella strains. The activity of each construct was evaluated in S. Typhimurium 14028 cells during 48 hours of growth. Graphed values represent the maximum reporter activity recorded in this period and is reported as counts per second (CPS). Statistical significance: *, P < 0.05; **, P < 0.01. (D) Constructs derived from S. Typhimurium 14028 csgD and csgB promoter sequences were introduced into each Salmonella strain. Letters above the bars indicate mean values that were statistically similar to (black font) or different from (red font) other mean values. #, values below the activity threshold as established in [24]. Each bar represents the mean value from three to five biological replicates. Error bars represent standard deviations.
To assess the effects of sequence or ‘cis’ differences in the csg intergenic region, we generated transcriptional reporters for the csgD and csgB promoters of all twelve strains. The reporters were transformed into S. Typhimurium 14028 to ensure that promoter activity was measured in a consistent cellular environment. Maximum expression levels were similar for all csgD or all csgB promoter-reporter constructs generated from NTS control strains and S. Typhi strains, suggesting that serovar-specific polymorphisms did not have a significant impact on promoter functionality (Fig 3C). We noted slightly lower activity from the csgB promoter from S. Typhimurium SL1344, but considered the promoter functional based on its overall expression profile (S4 Fig). In contrast, the activity of csgD promoters from S. Typhimurium D23580 and S. Enteritidis D7795 were significantly lower than native S. Typhimurium 14028 promoters (Fig 3C). S. Typhimurium D23580 had peak expression that was approximately three-fold lower than S. Typhimurium 14028, whereas S. Enteritidis D7795 had near background levels of expression (~1,000 counts per second) and appeared to be non-functional. In contrast, there was minimal difference between the activities of csgB promoters from the African NTS strains and the panel of control strains.
To determine if strains harboured mutations in trans-acting regulatory elements, each strain was transformed with a set of biofilm-associated transcriptional reporters derived from S. Typhimurium 14028. The csgD promoter was expressed in all Salmonella strains, though its activity was six-fold lower in S. Enteritidis D7795, S. Ty E02-2759, and S. Ty E03-9804 (Fig 3D). This result suggested that mutations in the regulatory network upstream of csgD could be partially responsible for loss of the rdar morphotype in these three strains. However, sequence analysis of the promoters and genes of six common regulators of csgD transcription did not reveal any unique sequence changes in these strains (i.e., positive regulators OmpR, MlrA, IHF, RstA; negative regulators CpxR, H-NS [45,46]). Expression of the csgB promoter showed a clearer distinction, as all control NTS strains and S. Typhimurium D23580 had significantly greater csgB promoter expression than rdar-negative S. Enteritidis D7795 and the S. Typhi strains (Fig 3D). A similar pattern was observed for the promoters of adrA, a gene which regulates cellulose production, and cpxRA, which is part of a two-component system that responds to envelope stress (S5 Fig). These trans patterns of gene expression correlated with a lack of CsgD in S. Enteritidis D7795 and S. Typhi strains.
A single, non-coding nucleotide shuts off the rdar morphotype for Salmonella serovar Enteritidis D7795
S. Enteritidis D7795 possessed a unique C-to-T transition in the csgD promoter region and also had a non-functional csgD promoter. We performed genome engineering to replace this SNP with corresponding sequence from rdar-positive S. Typhimurium 14028 (see Materials and Methods). We were able to track the presence of the SNP by digestion of the csgD promoter region with the restriction endonuclease, PsiI. Strains containing the SNP (i.e., TTATAA) had a unique 148 bp fragment compared to strains with the SNP corrected (i.e., TTACAA) (Fig 4A). Strains of S. Enteritidis D7795 with the SNP corrected (-47T>C) had their biofilm phenotypes restored, as judged by the formation of rdar colonies on TCR agar and cellulose production on agar supplemented with calcofluor white (Fig 4B), in addition to the formation of biofilm aggregates when grown in liquid culture (S6 Fig). In contrast, S. Enteritidis D7795 strains that had retained the SNP after genome engineering were negative for the rdar morphotype and multicellular aggregate formation. As another test of the importance of this promoter SNP, we introduced the change into S. Typhimurium 14028 (-47C>T), which resulted in the loss of rdar colony morphology and cellulose production (Fig 4B).
Fig 4. Biofilm phenotypes and csgD promoter activities following chromosomal replacement of strain-specific csgD promoter sequences in S. Enteritidis D7795 and S. Typhimurium 14028.

(A) Genome engineering was used to replace part of the native csgD promoter sequence in S. Enteritidis D7795 with sequence from S. Typhimurium 14028. The same process was also used to replace part of the native sequence in S. Typhimurium 14028 cells with sequence from S. Enteritidis D7795. The 780-bp csgD promoter region was PCR amplified from the strains and clones listed, followed by digestion with PsiI, which has a recognition site overlapping the S. Enteritidis D7795 SNP-containing region. (B) Macrocolony phenotypes of S. Enteritidis D7795 and S. Typhimurium 14028 clones that either contain or do not contain the identified ‘T’ promoter SNP at position -47. (C) The csgD promoter from each S. Enteritidis D7795 clone was used to generate promoter luciferase reporters that were transformed into S. Typhimurium 14028 cells. Each line represents one biological replicate culture (n = 22) with measured promoter activity (CPS, counts per second) plotted versus time. (D) Maximum reporter activity recorded from csgD promoter luciferase constructs derived from S. Typhimurium 14028 clones and transformed into wildtype S. Typhimurium 14028 cells (n = 11 per reporter construct). The activity of wildtype (WT) S. Typhimurium 14028 and S. Enteritidis D7795 csgD promoter-reporter constructs were included in the assay (n = 4 per each construct). Violin plots show the frequency distribution of the data, with the dotted line representing the median value. ****, P < 0.0001.
To determine if the phenotypic changes observed were directly linked to changes in csgD promoter activity, we generated promoter-reporter constructs from each engineered strain. Presence of the ‘T’ SNP at position -47 correlated with functional inactivation of the csgD promoter, shown as a 48-hour time course for S. Enteritidis D7795 strains (Fig 4C) and a statistically significant drop in maximum promoter expression for the S. Typhimurium 14028 strains (Fig 4D). Together, these experiments provided evidence that a single polymorphism in a non-coding region of the Salmonella genome was capable of shutting off CsgD-regulated biofilm phenotypes.
Unique SNPs reduce csgD promoter activity in invasive NTS strain Salmonella serovar Typhimurium D23580
S. Typhimurium D23580 possessed two unique SNPs in the csgD promoter region and the promoter had reduced activity when compared to S. Typhimurium 14028. We used genome engineering to replace the SNP at the -80 bp position (D23580 -80A>C), which boosted csgD promoter activity approximately two-fold as compared to the D23580 parent strain (Fig 5A). Replacement of the SNP at -189 bp position (D23580 -80A>C, -189A>G) resulted in an additional boost in promoter activity to reach similar levels as the csgD promoter from wildtype S. Typhimurium 14028 (Fig 5A). Despite the increase in promoter activity, replacement of both csgD promoter SNPs (S. Typhimurium D23580-PcsgD -80A>C, -189A>G) was unable to restore the rdar morphotype, and although the strain appeared to bind more Congo red than the parent S. Typhimurium D23580 strain, the difference was hard to quantitate (Fig 5B).
Fig 5. Biofilm phenotypes and csgD promoter activities following chromosomal replacement of strain-specific csgD promoter sequences or bcsG mutations in S. Typhimurium D23580 or S. Typhimurium 14028.
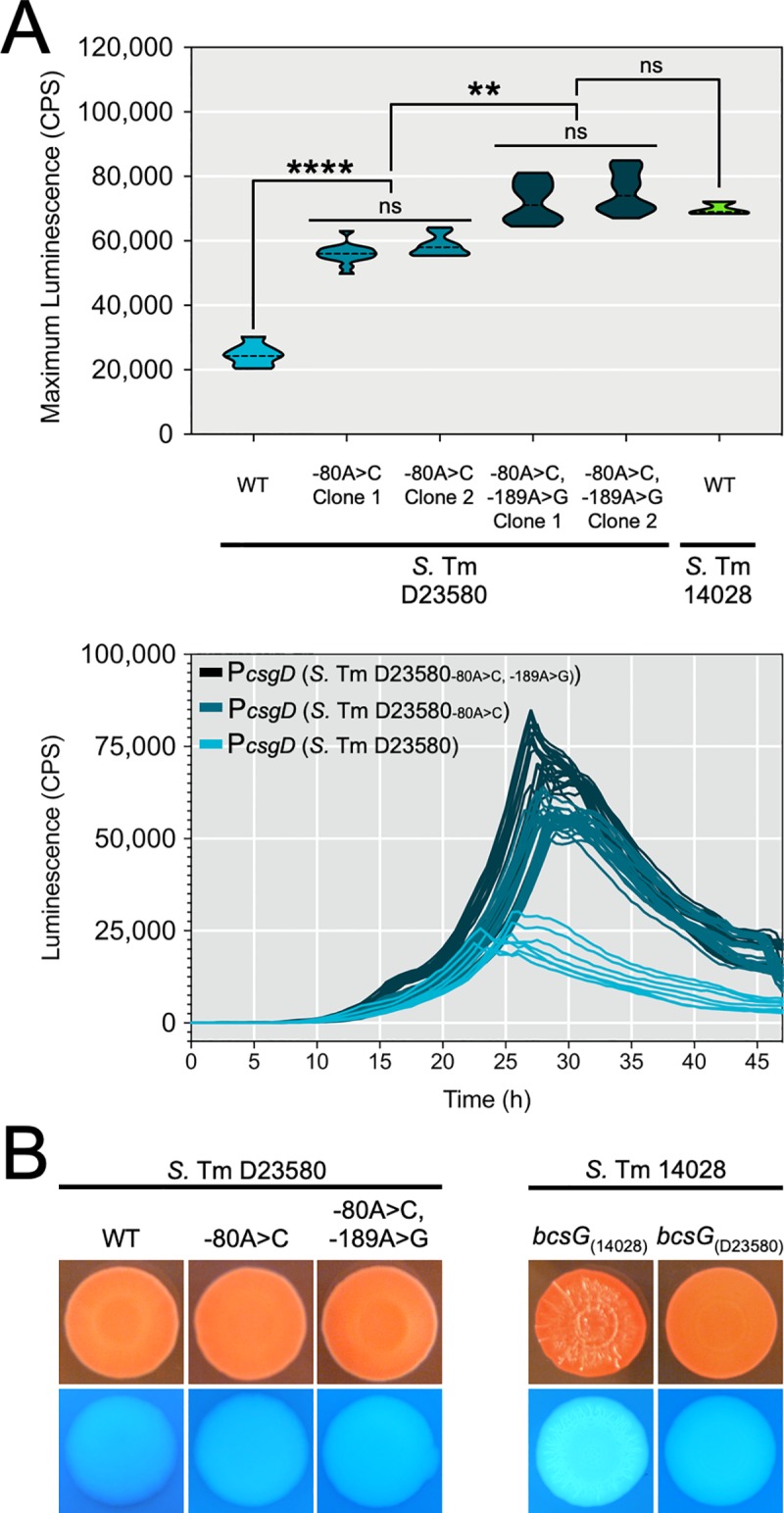
(A) Top panel: Maximum activity (CPS; counts per second) was recorded for csgD promoter luciferase reporters derived from S. Typhimurium D23580 clones generated by genome engineering that either contained or did not contain the ‘A’ SNP at position -80 and ‘A’ SNP at position -189 relative to the csgD transcriptional start site. Reporters were transformed into S. Typhimurium 14028 and activity monitored during 48 hours of growth (n = 28 per reporter construct). Wildtype (WT) reporters from S. Typhimurium D23580 (n = 24) and S. Typhimurium 14028 (n = 12) were included as controls. Violin plots show the frequency distribution of the data, with the dotted line representing the median value; **, P < 0.01; ****, P < 0.0001. Bottom panel: 48 h time-course expression profiles for csgD promoter luciferase reporters analyzed in the top panel, except for S. Typhimurium 14028; each line represents one biological replicate culture. (B) Macrocolony phenotypes of S. Typhimurium D23580 and S. Typhimurium 14028 clones containing native or replacement csgD promoter sequences (left panel) or bcsG alleles (right panel).
S. Typhimurium D23580 was first characterized as being rdar-negative or -intermediate due to the presence of a premature stop codon in bcsG, and the rdar morphotype was restored by over-expressing bcsG [47]. To evaluate the impact of the bcsG mutation, we introduced this SNP into the rdar-positive S. Typhimurium 14028 strain, which caused a loss of pattern formation on the colony surface and a drop in calcofluor binding intensity (Fig 6B). We concluded from this data that S. Typhimurium D23580 possessed multiple mutations that influence curli and cellulose production, leading to a reduced biofilm phenotype.
Fig 6. Conservation of csgD promoter and bcsG single nucleotide polymorphisms in invasive S. Enteritidis and S. Typhimurium lineages.

Maximum likelihood phylogenic trees were constructed from bacterial genome sequences: (A) S. Enteritidis isolates from Feasey et al. [10], and (B) S. Typhimurium isolates from Okoro et al [48], keeping the same general tree shape for comparison purposes. (A) S. Enteritidis isolates were divided into the Central/East African clade (167 isolates) and global epidemic clade (250 isolates), with the distinct region of isolation shown along with presence or absence of the ‘T’ SNP. (B) S. Typhimurium isolates were divided into gastroenteritis-associated and invasive lineages (I and II), with the presence or absence of csgD promoter and bcsG polymorphisms shown.
Conservation of biofilm-altering SNPs in invasive lineages of S. Typhimurium and S. Enteritidis
We performed in silico screening of additional serovar Enteritidis and Typhimurium strains isolated from Africa and other areas of the world [10,48] to determine how widespread the biofilm-altering SNPs were. For S. Enteritidis, all 167 strains in the central/east African clade (i.e., HierBAPs-predicted clade from [10]), which includes D7795, possessed the inactivating csgD promoter SNP (Fig 6A). In contrast, 100% of the 250 isolates from the HierBAPs-predicted global clade associated with human enterocolitis and/or poultry farming did not have this SNP (Fig 6A; S3 Table). For S. Typhimurium, the csgD promoter mutation at position -189 was conserved in all 50 lineage I and 71 lineage II strains analyzed (Fig 6B; S4 Table), that were isolated from people in sSA within the past 30 years [48]. Lineage II, which includes strain D23580, is thought to have arisen from lineage I due to increased selection pressure from heavy antibiotic use [48]. Therefore, we predicted that lineage II isolates would carry some unique mutations. Consistent with this, all lineage II isolates possessed the csgD promoter SNP at position -80 as well as the premature stop codon in bcsG. Importantly, none of the 63 S. Typhimurium strains of other sequence types (i.e., ST-19, -34, -98, -128 or -568) possessed the biofilm-altering mutations (Fig 6B; S4 Table).
Loss of the rdar morphotype and multicellular aggregation in Salmonella serovar Typhi strains is due to truncated CsgD
Salmonella Typhi represents the best characterized group of strains that cause a more invasive disease than enterocolitis. All six S. Typhi strains that we analyzed had functional csgD promoters, and at least four strains appeared to have upstream regulatory components intact (Fig 3). In the S. Typhi CT18 genome sequence, Parkhill et al. identified a premature stop codon at the 3’ end of csgD, which would result in truncation of eight amino acids from the C-terminal end of CsgD [49]. To analyze the functionality of this truncated csgD allele (i.e., csgD^CT18) compared to full-length csgD (i.e., csgD^14028), we performed complementation experiments in a S. Typhimurium 14028 ΔcsgD strain background. The presence of csgD^14028 in a multi-copy plasmid resulted in a strain with characteristic rdar morphotype colonies, whereas the p3xFLAG control strain was smooth and white (Fig 7A). The presence of csgD^CT18 resulted in a strain that formed red colonies with rdar-intermediate morphology, indicating that the truncated CsgD protein was partially functional. When grown in liquid culture, the csgD^CT18 and p3xFLAG cultures were devoid of multicellular aggregates, whereas the csgD^14028 complemented culture produced both cell types (Fig 7A). We detected robust CsgD-3xFLAG synthesis in the csgD^14028 culture, whereas only a faint CsgD band could be detected in the csgD^CT18 culture (Fig 7B). When expression of csgD^CT18 was induced by addition of IPTG, it caused a boost in curli (csgBAC) gene promoter expression (Fig 7B) and aggregates were formed in liquid culture (Fig 7C). Expression of the csgB promoter in the induced csgD^CT18 cultures reached levels similar to, but still below the levels in the csgD^14028 uninduced cultures (Fig 6C). We were unable to measure csgB promoter activity in induced csgD^14028 cultures because growth ceased upon addition of IPTG. Together, these experiments demonstrated that truncated CsgD from S. Typhi CT18 had reduced functionality compared to full-length CsgD, but was able to restore both rdar and other CsgD-regulated biofilm phenotypes if expressed at higher levels. As a final test of functionality, we used genome engineering to introduce the premature stop codon in csgD into S. Typhimurium 14028, resulting in a strain that was rdar-negative with minimal Congo red binding and reduced cellulose production (Fig 7D). This experiment showed that the SNP at the 3’ end of csgD was sufficient to disrupt CsgD-regulated biofilm phenotypes in S. Ty CT18.
Fig 7. Functional analysis of the S. Typhi CT18 csgD allele.

(A) The csgD alleles from S. Typhimurium 14028 and S. Typhi CT18 were cloned into p3xFLAG and transformed into S. Typhimurium 14028 ΔcsgD cells. Colony morphology and flask cultures of uninduced cells were evaluated for biofilm phenotypes. (B) Top panel: Whole cell lysates were generated from cells acquired from flask cultures and probed for synthesis of CsgD via Western blot. Bottom panel: A csgB promoter-reporter construct was used to evaluate CsgD activity in S. Typhimurium 14028 ΔcsgD cells harbouring p3xFLAG-csgD^CT18 with or without IPTG induction. (C) Top panel: Presence or absence of multicellular aggregates in flask cultures of S. Typhimurium 14028 ΔcsgD cells containing p3xFLAG constructs (+/- IPTG) or p3xFLAG alone. Bottom panel: Maximum promoter activity of a csgB promoter-reporter construct measured in microbroth cultures corresponding to the tubes shown in the top panel. (D) Biofilm phenotypes of S. Typhimurium 14028 strains that contained the native csgD^14028 allele or the truncated csgD allele from S. Typhi CT18 following genome engineering.
We wanted to determine if there were other mutations that could potentially affect curli or cellulose production within our panel of twelve Salmonella strains. For curli, we performed sequence alignment of the entire 4,450 bp region containing the divergent csgBAC and csgDEFG operons (S7 Fig). Seventeen unique SNPs were identified in each of the six S. Typhi strains, however the only clear nonsynonymous mutation was the premature stop codon in csgD that was previously described [49]. Several serovar Typhimurium- and Enteritidis-specific SNPs were identified in the intergenic region and in the csg coding regions, however since the majority of these SNPs were found in biofilm-positive strains, we concluded that they likely do not pose a significant effect on csg function. For cellulose, we performed sequence alignment of the 14,273 bp region containing the divergent bcsRQABZC and bcsEFG operons (S8 Fig). Overall, this DNA region was less conserved than the csg region. All six S. Typhi strains shared numerous SNPs, including 21 non-synonymous changes in the bcs coding regions, and four premature stop codons in bcsC. S. Enteritidis D7795 had one unique SNP in bcsC, which resulted in a non-synonymous change that was shared with S. Typhi strains, and S. Typhimurium D23580 had one unique SNP leading to a premature stop codon in bcsG, as previously described [47].
Uniqueness of the identified SNPs influencing rdar morphotype biofilm formation in Salmonella
We wanted to determine if the SNPs identified in S. Enteritidis D7795, S. Typhimurium D23580 and S. Typhi were unique to these lineages or could be detected in isolates from other S. enterica subspecies enterica serovars. We analyzed the genomes of 248 isolates from 55 serovars, including representatives of the most common serovars associated with human disease [50] and host-adapted serovars such as Dublin, Choleraesuis and Gallinarum (S5 Table). The -47 C>T mutation of S. Enteritidis D7795 was found in one strain of serovar Weltevreden, one strain of serovar Anatum had a 110-bp deletion in the csgD promoter region comprising both the -47 C>T and -80 G>T mutations, and the -189 C>T mutation of S. Typhimurium D23580 was found in one strain of serovar Hillingdon and two strains of serovar Typhimurium (S5 Table). The SNP leading to a premature stop codon in csgD was unique to Typhi strains, and the G>A mutation causing a premature stop codon in bcsG was not detected in any strains (S5 Table). We also screened 82 West African serovar Enteritidis strains associated with invasive human disease [10] but none of the mutations were detected (S6 Table). With just a few exceptions, the SNPs identified in our biofilm screening appeared to be unique to the invasive lineages of Salmonella where they were originally detected.
Discussion
In this study, we identified critical changes that disrupt or reduce the rdar morphotype and other CsgD-regulated biofilm phenotypes in three invasive Salmonella lineages. For S. Enteritidis D7795, a single promoter mutation was responsible for inactivating the csgD promoter and shutting off the rdar morphotype. The polymorphism was in one of the two most conserved bases in the OmpR recognition site (ACNTTTNGNTA’C’ANNTAT; [51]). This is predicted to knock out OmpR binding to a region which has been shown to be a major activating factor for csgD transcription [52]. Restoring biofilm formation was as simple as replacing this SNP, which re-activated the csgD promoter, overcoming low RpoS activity in S. Enteritidis D7795. We have observed mutations in this OmpR binding site before, in two strains of Salmonella serovar Arizonae that had lost biofilm formation, strains that we speculated were adapted for living within the snake intestine [24]. Conservation of the csgD promoter SNP in all 167 strains of the Central/East African clade of S. Enteritidis from sub-Saharan Africa, and lack of the SNP in 250 ‘global’ S. Enteritidis isolates, is strong evidence that loss of this CsgD-regulated biofilm phenotype has being selectively maintained in the invasive population since the most recent common ancestor, circa 1945 [10].
S. Typhimurium D23580 had multiple mutations that influenced the rdar morphotype. Two SNPs were identified in the csgD promoter region, each causing a reduction in transcriptional activity. We hypothesized that reduced csgD promoter activity in S. Typhimurium D23580 could explain the reduced levels of curli production measured in the biofilm flask model. The third mutation was a premature stop codon in bcsG that was first identified by Singletary et al. in 2016 [47]. Recent work has shown that deletion of bcsG shuts off cellulose production [53]; however, in our hands, S. Typhimurium D23580 still produced measurable amounts of cellulose. BcsG has at least two functions, to add phosphatidylethanolamine (PE) to monomers of the growing cellulose chain [54], and to stabilize integration of BcsA into the inner membrane, a role that has been ascribed to the N-terminal half of the protein [53]. In S. Typhimurium D23580, the premature stop codon occurs at amino acid 247 in BcsG. If the N-terminal region of BcsG is synthesized, it would allow BcsA, the cellulose synthase enzyme, to reach native levels within the cytoplasmic membrane [53]. This could explain why S. Typhimurium D23580 can still produce moderate levels of cellulose. The effects of the csgD promoter mutations and bcsG truncation could have a great impact in the natural lifecycle of Salmonella, since they would reduce both the amount of biofilm produced and alter the physical structure of any multicellular aggregates. This could result in isolates that do not survive as well in the environment, as recently demonstrated for invasive S. Typhimurium isolates in Mali [33]. Conservation of all three polymorphisms in 100% of African S. Typhimurium lineage II isolates, collected from human patients in sSA within the past 30 years [48], is evidence of sustained selection against the rdar morphotype in this lineage.
For Salmonella serovar Typhi, we showed that a premature stop codon, resulting in loss of eight amino acids from the C-terminus of CsgD, was sufficient to shut off the rdar morphotype. The truncated CsgD protein had reduced activity, but was able to activate rdar-like morphologies and multicellular aggregation when expressed at higher levels. Therefore, any increases in csgD transcription, which can be caused by known promoter mutations [16,24] or potentially host-related environmental signals such as iron limitation [16] or the presence of human bile [37], may be enough to restore CsgD-regulated biofilm phenotypes in S. Typhi. To explain why CsgD was not detectable in S. Typhi, we hypothesize that the reduced activity of truncated CsgD was not able to activate the genetic feed-forward loop that normally amplifies CsgD production [39,55]. Key dimerization domains exist in the N-terminal half of CsgD [56], but it is not yet clear why the C-terminal truncation would reduce its activity. Despite the lack of a rdar morphotype, it should be noted that S. Typhi can form biofilms on human gallstones [57,58], a process which has been simulated with S. Typhimurium in a mouse model of chronic carriage; however, the nature of the extracellular matrix in such biofilms is still under investigation [37,59,60]. Our analysis highlighted the presence 4 premature stop codons in the bcsC of the S. Typhi strains included in our panel [23]. BcsC is an essential enzyme in cellulose biosynthesis [61], with a C-terminal ‘pore’ domain that guides the growing cellulose chain out of the cell [62]. This suggests that S. Typhi strains could be negative for cellulose production irrespective of reduced CsgD function.
The impacts on the curli and cellulose systems in all three lineages of invasive S. enterica isolates provides strong evidence that parallel evolution has occurred [63]. With the association of phenotypic changes to mutations in promoter regions [64–66] or in transcriptional regulatory proteins that can act as bistable switches [40,67,68], we present further evidence that changes in gene expression can drive specialization or ecological divergence without significant changes in gene content [69,70]. We recently reported the increased expression of over 780 genes during the development of multicellular aggregates in flask cultures of S. Typhimurium 14028 [39]. The shift in the transcriptome of Salmonella cells contrasts sharply with both the small regulon of genes directly controlled by CsgD as well as the complex but limited number of regulatory factors that influence expression and synthesis of CsgD itself [34,45]. Variation in cis regulatory regions is hypothesized to have a reduced fitness cost compared to changes in coding sequence, since genetic plasticity is retained [71]. Within the three lineages of invasive Salmonella isolates, both regulatory and structural mutations have played a prominent role in loss or impairment of the rdar morphotype. It is possible that the accumulation of genetic changes has been aided by replication and circulation of African strains within immunocompromised hosts [72].
We do not fully understand what the selective pressures are that have led to loss or impairment of biofilm formation in invasive Salmonella isolates. The changes we identified were generally specific to the invasive lineages that were investigated and were not conserved across a wider variety of serovars and isolates. We know that enterocolitis-causing isolates replicate to high numbers in the intestine before passing out of the host and into the environment [7]. Replication is aided by Salmonella-induced inflammation, which destabilizes the normal microbiota and provides Salmonella with a selective advantage due to specific metabolic adaptations [30,73]. In contrast, Salmonella strains that cause systemic disease tend to have a stealth and persistence strategy and remain associated with the host for a longer duration of time [8]. The transition from the intestinal to systemic niche is thought to represent an evolutionary bottleneck for Salmonella [28], with significant losses in the functional gene repertoire consistently observed for invasive variants [29]. Bistable genetic networks in bacteria, such as the one described for CsgD, are often associated with the formation of two distinguishable phenotypes within a clonal population [74], which is thought to allow genotypes to persist in fluctuating environments [75]. It would make sense for enterocolitis-causing isolates to retain the CsgD network, because the presence of persistent (CsgD-ON) biofilm cells and virulent (CsgD-OFF) planktonic cells would likely improve the odds for future transmission events [15,23,26,39]. Systemic isolates might not require the CsgD network because they are increasingly reliant on human carriers for transmission, as noted for S. Typhi [28,57,58]. In general, the biofilm-altering SNPs identified in the invasive isolates were not found in lineages of S. Enteritidis or S. Typhimurium that typically cause enterocolitis in association with zoonotic transmission.
An alternative explanation for selection against the rdar morphotype is that the biofilm surface structures themselves are targets of the host immune system. Several independent studies have shown that S. Typhimurium strains that have lost curli and cellulose production are able to invade tissue culture cells more efficiently [76,77] and spread systemically in vivo [78,79]. Curli have been established as potent stimulators of the innate immune system that are recognized by Toll-like receptor 1 and 2 complexes [80,81], as well as intracellular NOD-like receptors [82]. Curli also have the ability to stimulate T-helper cell 17 (Th17) differentiation and increase expression of pro-inflammatory interleukin (IL) cytokines IL-17A, IL-22 and IL-1ß [82,83]. Some questions still remain, however. Although there is evidence that cellulose can be produced in vivo [78,84], it has yet to be conclusively established if this constitutes a biofilm phenotype. It is also difficult to extrapolate how much of an immune response could be generated against Salmonella biofilms in immunocompromised, often HIV-positive patients in sSA.
Both csgD expression and the rdar morphotype are highly conserved across Salmonella enterica and E. coli [19,23,24,85], with notable exceptions including Salmonella serovars associated with host restriction and systemic disease (S. Typhi, S. Paratyphi A, and S. Gallinarum) and enteroinvasive E. coli and Shigella [23,26]. Loss of the rdar morphotype in Salmonella has been correlated with invasion of the intestinal epithelial lining [23]. There are numerous examples of Salmonella biofilm formation providing a survival or persistence advantage under conditions of stress, such as desiccation, nutrient deprivation and disinfection [24,25,86–88]. The impairment or inactivation of the rdar morphotype in the African invasive lineages suggests that their lifestyle could be distinct from lineages of S. Typhimurium and S. Enteritidis associated with industrialized food supply chains in resource rich settings. Although it is generally accepted that the transmission route for these invasive organisms is fecal-oral, we know very little about their behaviour in the environment between hosts. Based on the data presented, we hypothesize that the African invasive NTS isolates are becoming human-adapted, as has been speculated by other researchers [89,90]. Increased knowledge about the ecological niches that harbor these specialized strains as well as their transmission patterns will be critical for developing public health measures to reduce the morbidity and mortality associated with invasive salmonellosis.
Materials and methods
Bacterial strains, media, and growth conditions
The bacterial strains used in this study are listed in S1 Table. For standard growth, strains were inoculated from frozen stocks onto LB agar (lysogeny broth, 1% NaCl, 1.5% agar) and grown overnight at 37˚C. One isolated colony was used to inoculate 5 mL LB broth and the culture was incubated for 18 hours at 37˚C with agitation at 200 RPM. For colony morphology assays, overnight cultures of each strain were normalized to an optical density of 1.0 at 600 nm and 2 μL were spotted onto 1% tryptone medium containing 1.5% Difco agar (T agar) [36]. To visualize the rdar morphotype, T agar was supplemented with 40–60 μg mL-1 Congo red. To visualize cellulose production, T agar was supplemented with calcofluor white (fluorescent brightener 28; Sigma-Aldrich Canada) at a final concentration of 200 μg mL-1 [25]. To analyze liquid culture growth under biofilm-inducing conditions, 1 x 109 CFU were inoculated into 100 mL of 1% tryptone, pH 7.4, and incubated at 28˚C for 24 or 48 hours with agitation at 200 rpm.
Generation of bacterial luciferase reporters
The pCS26 and pU220 reporter plasmids containing csgDEFG, csgBAC, and adrA promoter sequences from S. Typhimurium 14028 fused to the luxCDABE operon from Photorhabdus luminescens have been described previously [15,24]. The RpoS-dependent reporter plasmid sig38H4 contains the luxCDABE operon preceded by a synthetic promoter designed based on the alignment of multiple RpoS-controlled promoters [15]. To generate the pCS26-cpxR promoter-luxCDABE construct, the cpx intergenic region was PCR amplified from S. Typhimurium 14028 genomic DNA using primers cpxR1 and cpxR2 (S2 Table) and Phusion high-fidelity DNA polymerase (New England BioLabs), with reaction conditions outlined by the manufacturer. The desired PCR product was purified (Geneaid PCR cleanup kit), sequentially digested with XhoI and BamHI (New England BioLabs), and ligated using T4 DNA ligase (New England BioLabs) into the pCS26 vector cut with XhoI and BamHI. To generate csgDEFG and csgBAC luciferase reporters from each Salmonella strain, the csg intergenic region was PCR amplified from genomic DNA using primers agfD1 and agfD2 (S2 Table). The PCR products were then ligated into either pCS26 (XhoI-BamHI) or pU220 (BamHI-XhoI) to generate the csgB and csgD promoter-reporter constructs, respectively.
Luciferase reporter assays
LB overnight cultures of Salmonella strains were diluted 1 in 600 in a final volume of 150 μL of 1% tryptone broth supplemented with 50 μg mL-1 Kn in 96-well clear-bottom black plates (Costar #9520; Corning Life Sciences). To minimize evaporation of the medium during the assay, cultures were overlaid with 50 μL of mineral oil. Cultures were assayed for absorbance (590 nm, 0.1 s) and luminescence (1s; in counts per second [CPS]) every 30 min during growth at 28˚C with agitation in a Victor X3 multilabel plate reader (Perkin-Elmer).
SDS-PAGE and western blotting
For planktonic cell samples, approximately 5 x 1010 cells were sedimented by centrifugation (11,000 x g; 10 min; 4˚C). For biofilm aggregate samples, approximately 30 mg samples were resuspended in 1 mL of water and homogenized with a glass tissue grinder (Product #7727–2, Corning Life Sciences) for 25 dounces, prior to centrifugation (10,000 x g; 1 min) to sediment the cell material. Sedimented samples were resuspended in 1 mL of SDS-PAGE sample buffer without 2-mercaptoethanol and bromophenol blue and boiled for 5 min. Using the DC protein assay (Bio-Rad Laboratories), cell lysates were normalized to a final protein concentration of 3 mg/mL. Bromophenol blue (0.0002% final concentration) and 2-mercaptoethanol (0.2% final concentration) were added to each lysate before loading 15 μg of total protein per lane. SDS-PAGE was performed with a 5% stacking gel and a 12 or 15% resolving gel. Proteins were transferred to nitrocellulose for 40 min at 25 V using a Trans-Blot SD semi-dry transfer cell (Bio-Rad Laboratories) in tris-glycine buffer supplemented with methanol. To detect curli fimbriae, cell debris was sedimented following boiling in SDS-PAGE sample buffer, washed twice with 500 μL of distilled water, suspended in 250 μL of 90% formic acid, frozen and lyophilized [36]. The dried samples were resuspended in SDS-PAGE sample buffer and loaded directly without boiling into each SDS-PAGE gel lane. CsgD was detected using a CsgD-specific monoclonal antibody at a 1-in-6 dilution of tissue culture supernatant (ImmunoPrecise Antibodies Ltd., Victoria, BC). To detect RpoS protein, a commercially available mouse polyclonal immune serum recognizing epitope 33 to 256 of E. coli RpoS was used at a 1-in-2000 dilution (BioLegend; 1RS1). CsgA, the major subunit of curli fimbriae, was detected by using a rabbit polyclonal serum raised against whole purified curli [36]. GroEL was used as a protein-loading control and was detected with a 1-in-60,000 dilution of rabbit polyclonal immune serum (Sigma-Aldrich; G6532). Secondary antibodies IRDye 800CW Goat anti-Mouse immunoglobulin G (IgG) or 680RD Goat anti-rabbit IgG (Mandel Scientific) were used at a 1-in-10,000 dilution and detected using the the Odyssey CLx imaging system and Image Studio 4.0 software package (Li-Cor Biosciences).
Reference genome sequences
Whole genome sequences were obtained from the National Centre for Biotechnology Information (NCBI) via the following accession numbers: NC_016810 (S. Typhimurium SL1344); NC_016854 (S. Typhimurium D23580); NC_003198 (S. Typhi CT18). Mapped assemblies for S. Typhi H58 haplotype strains E02-2759, E03-9804, ISP03-07467, ISP04-06979 were available from http://www.sanger.ac.uk/Projects/S_typhi [28]. The S. Typhi 8(04)N genome was available from the European Nucleotide Archive under the name SGB112 and associated with the assembly number GCA_001362315.1. The sequence for S. Enteritidis D7795 was available from the Public Health England Pathogens BioProject on NCBI (accession number PRJNA248792).
Chromosomal DNA isolation, genome sequencing and sequence alignments
Overnight liquid cultures of Salmonella serovar Enteritidis strains 301 and ATCC 4931 were sub-cultured 1 in 100 in 200 mL LB broth and grown at 37˚C for 2.5 hours. Approximately 7 x 108 cells were centrifuged (6000 x g, 10 minutes, 4˚C), resuspended in 25 mM Tris, 1 mM EDTA solution (pH 8.0) to a total volume of 3.5 mL, and then treated with 10 mg lysozyme (Sigma-Aldrich; L6876) and 200 units of mutanolysin (Sigma-Aldrich; M9901) for 1 hour at 37˚C. For cell lysis, each sample was treated with 50 μL of 25% SDS, 1 mg proteinase K (Applied Biosystems; AM23548), and 125 μL of a 5M sodium chloride solution and incubated at 65˚C for 30 minutes. Nucleic acid was isolated from cell lysates using a series of phenol:chloroform:isoamyl and phenol:chloroform extractions, precipitated by the addition of ammonium acetate (at a final concentration of 2M), washed with ethanol, and resuspended in Tris-EDTA solution (10 mM Tris, 1 mM EDTA, pH 8.0). To remove RNA, RNase A was added to each sample at a final concentration of 0.2 mg/mL and incubated for 1 hour at 37˚C. Samples were purified once more by phenol:chloroform:isoamyl extraction, precipitated with ammonium acetate, washed with ethanol, and resuspended in a final volume of 200 μL of Tris-EDTA solution.
Purified chromosomal DNA samples were fragmented by cup horn sonication with a high-intensity ultrasonic processor (Vibra-Cell, Danbury, CT) for 10 cycles of a 30-second pulses and 2 minute rest. DNA libraries were prepared from 1 μg of fragmented DNA using the NEBNext Ultra DNA Library Prep Kit for Illumina (New England BioLabs; E7645S) and NEBNext Multiplex Oligos for Illumina (Index Primer Set 2) (New England BioLabs; E7500S) according to the manufacturer’s protocols. Adaptor-ligated DNA was size-selected between 400 and 500 bp total library size (length of insert sequence + adaptor sequence) following kit instructions. DNA samples were assessed for quality, purity, and integrity using a NanoDrop ND-1000 spectrophotometer (Fisher Scientific) and an Agilent 2100 Bioanalyzer with a High Sensitivity DNA chip (Agilent Technologies; 5067–4626). To ensure efficient adaptor ligation, samples were analyzed by quantitative PCR using the KAPA Library Quantification Kit for the Illumina platform (KAPA Biosystems; KK4824). Samples were sequenced using the MiSeq Reagent Kit version 3, 600 cycles (2 x 300 bp read length) (Illumina; MS-102-3003).
Genome sequencing assembly and DNA sequence alignments were performed using the Geneious Pro v8.1.5 software package (Biomatters). Paired-end sequence reads from Entertidis strains 4931 and 301 were assembled into contigs based on mapping to the S. Enteritidis reference genome P125109 (NC_011294). Sequence alignments were performed using ClustalW and an IUB cost matrix (gap open cost of 15, gap extend cost of 6.66). A phylogenetic tree for strains included in this study was constructed based on the csg operon region by using the Geneious Tree Builder program [91], with the Tamura-Nei model of genetic substitution and the neighbour-joining algorithm with bootstrapping (1000 replicates and support threshold of 70%).
Genome engineering in S. Enteritidis D7795, S. Typhimurium D23580 and S. Typhimurium 14028
The I-SceI suicide plasmid system developed by Victor de Lorenzo and colleagues [92] was used for genome engineering. The following fragments were PCR amplified: 1) csg intergenic region from S. Typhimurium 14028 or S. Enteritidis D7795 using primers agfD3-FWD and agfD4-REV (S2 Table); 2) csg intergenic region from S. Typhimurium 14028 using primers agfD5-FWD and agfD6-REV (S2 Table); 3) bcsG-containing region from S. Typhimurium 14028 or S. Typhimurium D23580 using primers bcsG-checkF and bcsG-checkR (S2 Table); and 4) csgD-containing region from S. Typhi CT18 using primers csgDORFstartPstI and csgDreplaceREV (S2 Table). Phusion high-fidelity DNA polymerase was used for amplification, following reaction conditions outlined by the manufacturer (New England BioLabs). PCR products were purified, digested with BamHI and PstI (New England Biolabs) and ligated into BamHI/PstI-digested pSEVA212 [94]. Clones corresponding to each product were selected in E. coli S17-1 (λpir) and mating was performed to move the plasmid constructs into S. Enteritidis D7795, S. Typhimurium D23580 or S. Typhimurium 14028. Merodiploid strains with pSEVA212 plasmid constructs inserted into the genome were selected by growth on M9 minimal agar supplemented with 1mM MgSO4, 0.2% glucose and 100 μg mL-1 kanamycin (M9-Glc-Kn100) and confirmed by re-streaking onto M9-Glc-Kn100 agar. Purified pSEVA628S (200–300 ng) [93] was transformed into merodiploid strains by electroporation with selection on Luria agar supplemented with 1 mM m-toluate and 20 μg mL-1 gentamicin. Resulting colonies were re-streaked onto Luria agar supplemented with 20 μg mL-1 gentamicin, streaked on Luria agar supplemented with 50 μg mL-1 kanamycin to confirm loss of the pSEVA212 plasmid, and streaked on TCR plates (1% tryptone, 1.5% agar, 40 μg mL-1 Congo red) to check the biofilm phenotype. Colonies were selected from TCR plates and grown at 37°C for two overnight growth steps without gentamicin to generate cells that lack pSEVA628S. Final colonies were streaked onto (1) Luria agar, (2) Luria agar + 50 μg mL-1 Kn, (3) Luria agar + 20 μg mL-1 gentamicin, and (4) TCR plates to select the desired phenotypes. To confirm the genotypes, csgD promoter-, bcsG- or csgD-containing regions were PCR-amplified from resulting strains and Sanger sequencing was performed (Eurofins MWG Operon; Louisville, Kentucky, USA).
Generating p3xFLAG vector constructs
The pFLAG-CTC expression vector (Sigma Aldrich #E8408) was maintained in E. coli DH10B. Purified pFLAG-CTC was digested with SalI and XhoI restriction enzymes. A synthetic polylinker generated from phosphorylated oligonucleotides 3xFLAG-linkerA and 3xFLAG-linkerB (S2 Table) was ligated into the digested vector to generate p3xFLAG. The csgD open reading frame was PCR amplified from S. Typhimurium 14028 or S. Typhi CT18 genomic DNA using primers csgD-ORF-start and csgD-ORF-end (S2 Table), followed by digestion with XhoI and BamHI. The csgD fragments were ligated into XhoI- and BglII-digested plasmid to generate p3xFLAG/csgD^14028 and p3xFLAG/csgD^CT18. Insertion of the 3xFLAG polylinker and csgD ORF was confirmed by DNA sequencing. The csgD^CT18 allele does not have the 3xFLAG linker attached because of the premature stop codon in the 3’ end of csgD. The resulting p3xFLAG vectors were transformed into S. Typhimurium ΔcsgD prior to analysis of biofilm formation.
In silico detection of identified polymorphisms in Salmonella serovar Enteritidis and serovar Typhimurium strains
S. Enteritidis D77 is one of 167 isolates recently sequenced and described as being part of a distinct clade of S. Enteritidis featuring genomic degradation and geographical restriction to central and eastern Africa [10]. Similarly, S. Typhimurium D23580 is part of a unique lineage of S. Typhimurium, consisting of isolates from sSA [48]. Genome assemblies of the S. Enteritidis and S. Typhimurium isolates were investigated for the presence or absence of csgD promoter or bcsG SNPs through in silico PCR. Primers of the csgD sequence with a SNP in the promoter region were searched for in the genome assemblies using in_silico_pcr.py script (https://github.com/simonrharris/in_silico_pcr), allowing zero changes for a match. Names and accession numbers for each strain included in the SNP screening are listed in S3 and S4 Tables. A similar approach was followed to screen the genome sequences of 248 diverse S. enterica isolates [94], as well as 82 strains of serovar Enteritidis that are part of the West Africa lineage of strains originally described by Feasey et al. [10].
Statistical analysis
Statistical analysis was performed using GraphPad Prism versions 7.0c and 8.0.2. Data collected from promoter-reporter luciferase assays was reported as the maximum luciferase expression measured during the course of the experiment, and was expressed as the mean ± the standard deviation. This data was logarithmically transformed and evaluated for normal distribution using the Shapiro-Wilk normality test. If the data was normally distributed, comparisons of the mean maximum luciferase expression levels obtained from multiple experiments were performed using an ordinary one-way ANOVA with post-hoc analysis via Holm-Sidak’s multiple comparisons test with statistical significance set at p < 0.05. If the data was not normally distributed, comparisons were performed using the Kruskal-Wallis test with post-hoc analysis via Dunn’s multiple comparison test with statistical significance set at a p value of 0.05.
Data availablility
All numerical data and statistical analysis has been deposited in figshare (https://figshare.com/) and is publicly available at doi: https://doi.org/10.6084/m9.figshare.8220866.v1. The Illumina paired end sequence reads comprising the genome sequences of S. enterica suspecies enterica serovar Enteritidis strains have been deposited in the Sequence Read Archives (strain 301—SAMN11956692; strain ATCC 4931—SAMN11956691).
Supporting information
(A) Morphological comparison of colonies grown for five days at 28˚C or 37˚C on T agar supplemented with 40 μg mL-1 Congo red. (B) Maximum expression from a curli-specific reporter (csgBAC::luxCDABE) in each strain grown for 48 h at 28˚C or 37˚C. Luciferase expression was measured every 30 min during continuous culture.
(TIF)
Multicellular aggregates and planktonic cells formed from liquid cultures of S. Typhimurium 14028 and S. Typhimurium D23580. Images match those presented in Fig 1, but have been enhanced to emphasize aggregates within the samples. Aggregates formed by S. Typhimurium D23580 cells appear structurally distinct from aggregates formed by S. Typhimurium 14028 cells.
(TIF)
(A) Whole cell lysates were generated from multicellular aggregates and planktonic cells isolated from flask cultures of Salmonella strains after 24 hours of growth and probed for synthesis of RpoS. Lysates were normalized by total protein concentration. GroEL was used as a loading control to ensure that equal amounts of protein were loaded into each sample lane. (B) RpoS activity was evaluated by measuring luminescence from a synthetic, RpoS-dependent promoter-reporter construct expressed in each Salmonella strain during 48 hours of growth. Graphed values represent the maximum reporter activity recorded and is reported as counts per second (CPS). (C) Absorbance measurements represent strain growth in microaerophilic conditions in 96-well microtiter plates. Each curve represents one biological replicate; n = 4 per strain. (D) Time course of RpoS-dependent promoter activity in each indicated strain.
(TIF)
Each curve represents one biological replicate; n = 3.
(TIF)
Promoter-reporter constructs derived from S. Typhimurium 14028 adrA and cpxRA promoter sequences were introduced into each of the strains. Letters above the bars indicate mean values that were statistically similar to (black font) or different from (red font) other mean values. #, values below the activity threshold as established in [24]. Each bar represents the mean value from 3–5 independent biological replicates and error bars represent the standard deviations.
(TIF)
Clones that contain (-47T>T) or do not contain (-47T>C) the identified ‘T’ promoter SNP at position -47 were grown in flasks of 1% tryptone for 24 hours before being evaluated for the ability to form aggregates in liquid cultures.
(TIF)
Simplified multiple sequence alignment of the csgDEFG and csgBAC operons highlighting serovar- and strain-specific single nucleotide polymorphisms representing both nonsynonymous and synonymous mutations: S. Typhimurium and D23580 (yellow), S. Enteritidis and D7795 (light blue), and S. Typhi (pink). Other highlighted changes (black) include the SNP in D7795 that inactivates csgD transcription, and the SNP in S. Typhi strains that introduces a premature stop codon in csgD, yielding a CsgD protein that is truncated by 8 amino acids. The long black bar represents the DNA region with nucleotide position numbers listed above and csg genes shown below as yellow-boxed arrows. Special sequence features involved in operon regulation are highlighted above and below the black bar: -35 and -10 promoter regions (black elbow arrows), H-NS binding region (grey box), CpxR-binding sites (black bars), and OmpR binding regions (hatched boxes).
(TIF)
Simplified multiple sequence alignment of the bcsRQABZC and bcsEFG operons highlighting non-synonymous serovar- and strain-specific single nucleotide polymorphisms. Non-synonymous SNPs are shown in yellow; black bars indicate SNPs that yield premature stop codons (i.e. bcsG in D23580; four SNPs in bcsC in S. Typhi strains). The long black bar represents the DNA region with nucleotide position numbers listed above and bcs genes shown below as yellow-boxed arrows. bcsR, bcsF and a small hypothetical protein are shown without their names listed.
(TIF)
(PDF)
(PDF)
(XLSX)
(XLSX)
(XLSX)
(XLSX)
Acknowledgments
The authors wish to thank Gordon Dougan, Robert Kingsley and members of their laboratories for sending Salmonella strains; Victor de Lorenzo for providing I-SceI plasmids; Nicolas Wenner and Jay Hinton for providing the I-SceI genome engineering protocols; Bill Kay and Mike Surette for critical reading of the manuscript. Published as VIDO manuscript #844.
Data Availability
All raw numerical data and statistical analysis has been deposited in figshare (https://figshare.com/) and is publicly available at: doi: https://doi.org/10.6084/m9.figshare.8220866.v1.
Funding Statement
This work was supported by the Natural Sciences and Engineering Research Council of Canada, http://www.nserc-crsng.gc.ca/ (Grant #2017-05737 to APW; Alexander Graham Bell Canada Graduate Scholarship to KDM; Undergraduate Research Award to EGH), the Jarislowsky Chair in Biotechnology (APW), Saskatchewan Health Research Foundation (3866 to KDM); the University of Saskatchewan (Integrated Training Program in Infectious Diseases to MBP; Biomedical Research Award to DJH). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Kirk MD, Pires SM, Black RE, Caipo M, Crump JA, Devleesschauwer B, et al. World Health Organization Estimates of the Global and Regional Disease Burden of 22 Foodborne Bacterial, Protozoal, and Viral Diseases, 2010: A Data Synthesis. PLOS Med. 2015. December 3;12(12):e1001921 10.1371/journal.pmed.1001921 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Majowicz SE, Musto J, Scallan E, Angulo FJ, Kirk M, O'Brien SJ, et al. The global burden of nontyphoidal Salmonella gastroenteritis. Clin Infect Dis. 2010. March 15;50(6):882–9. 10.1086/650733 [DOI] [PubMed] [Google Scholar]
- 3.Reddy EA, Shaw AV, Crump JA. Community-acquired bloodstream infections in Africa: a systematic review and meta-analysis. Lancet Infect Dis. 2010;10(6):417–32. 10.1016/S1473-3099(10)70072-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Feasey NA, Dougan G, Kingsley RA, Heydermann RS, Gordon MA. Invasive non-typhoidal Salmonella disease: an emerging and neglected tropical disease in Africa. The Lancet. 2012. June 30;379(9835):2489–99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ao TT, Feasey NA, Gordon MA, Keddy KH, Angulo FJ, Crump JA. Global Burden of Invasive Nontyphoidal Salmonella Disease, 2010. Emerg Infect Dis. 2015;21(6):941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Haselbeck AH, Panzner U, Im J, Baker S, Meyer CG, Marks F. Current perspectives on invasive nontyphoidal Salmonella disease. Curr Opin Infect Dis. 2017;30(5). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bäumler A, Fang FC. Host specificity of bacterial pathogens. Cold Spring Harb Perspect Med. 2013. December 1;3(12):a010041–1. 10.1101/cshperspect.a010041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dougan G, Baker S. Salmonella enterica Serovar Typhi and the Pathogenesis of Typhoid Fever. Annu Rev Microbiol. 2014. September 8;68(1):317–36. [DOI] [PubMed] [Google Scholar]
- 9.Okoro CK, Barquist L, Connor TR, Harris SR, Clare S, Stevens MP, et al. Signatures of Adaptation in Human Invasive Salmonella Typhimurium ST313 Populations from Sub-Saharan Africa. PLoS Negl Trop Dis. 2015. March 24;9(3):e0003611 10.1371/journal.pntd.0003611 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Feasey NA, Hadfield J, Keddy KH, Dallman TJ, Jacobs J, Deng X, et al. Distinct Salmonella Enteritidis lineages associated with enterocolitis in high-income settings and invasive disease in low-income settings. Nat Genet. 2016. August 22;48(10):1211–7. 10.1038/ng.3644 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Winter SE, Thiennimitr P, Winter MG, Butler BP, Huseby DL, Crawford RW, et al. Gut inflammation provides a respiratory electron acceptor for Salmonella. Nature. 2010. September 23;467(7314):426–9. 10.1038/nature09415 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Nuccio S-P, Bäumler AJ. Comparative Analysis of Salmonella Genomes Identifies a Metabolic Network for Escalating Growth in the Inflamed Gut. mBio. 2014. May 1;5(2):e00929–14. 10.1128/mBio.00929-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lawley TD, Bouley DM, Hoy YE, Gerke C, Relman DA, Monack DM. Host Transmission of Salmonella enterica Serovar Typhimurium Is Controlled by Virulence Factors and Indigenous Intestinal Microbiota. Infect Immun. 2008. January 1;76(1):403 10.1128/IAI.01189-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Winfield MD, Groisman EA. Role of Nonhost Environments in the Lifestyles of Salmonella and Escherichia coli. Appl Environ Microbiol. 2003. July 1;69(7):3687 10.1128/AEM.69.7.3687-3694.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.White AP, Gibson DL, Kim W, Kay WW, Surette MG. Thin Aggregative Fimbriae and Cellulose Enhance Long-Term Survival and Persistence of Salmonella. J Bacteriol. 2006. April 17;188(9):3219–27. 10.1128/JB.188.9.3219-3227.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Römling U, Sierralta WD, Eriksson K, Normark S. Multicellular and aggregative behaviour of Salmonella typhimurium strains is controlled by mutations in the agfD promoter. Mol Microbiol. 1998. April;28(2):249–64. [DOI] [PubMed] [Google Scholar]
- 17.Zogaj X, Nimtz M, Rohde M, Bokranz W, Römling U. The multicellular morphotypes of Salmonella typhimurium and Escherichia coli produce cellulose as the second component of the extracellular matrix. Mol Microbiol. 2001. March;39(6):1452–63. [DOI] [PubMed] [Google Scholar]
- 18.Gibson DL, White AP, Snyder SD, Martin S, Heiss C, Azadi P, et al. Salmonella Produces an O-Antigen Capsule Regulated by AgfD and Important for Environmental Persistence. J Bacteriol. 2006. November 1;188(22):7722–30. 10.1128/JB.00809-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Römling U, Bian Z, Hammar M, Sierralta WD, Normark S. Curli fibers are highly conserved between Salmonella typhimurium and Escherichia coli with respect to operon structure and regulation. J Bacteriol. 1998. February;180(3):722–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Latasa C, Roux A, Toledo-Arana A, Ghigo J-M, Gamazo C, Penadés JR, et al. BapA, a large secreted protein required for biofilm formation and host colonization of Salmonella enterica serovar Enteritidis. Mol Microbiol. 2005. October 14;58(5):1322–39. 10.1111/j.1365-2958.2005.04907.x [DOI] [PubMed] [Google Scholar]
- 21.Zakikhany K, Harrington CR, Nimtz M, Hinton JCD, Römling U. Unphosphorylated CsgD controls biofilm formation in Salmonella enterica serovar Typhimurium. Mol Microbiol. 2010. July 6;77(3):771–86. 10.1111/j.1365-2958.2010.07247.x [DOI] [PubMed] [Google Scholar]
- 22.White AP, Gibson DL, Collinson SK, Banser PA, Kay WW. Extracellular Polysaccharides Associated with Thin Aggregative Fimbriae of Salmonella enterica Serovar Enteritidis. J Bacteriol. 2003. August 29;185(18):5398–407. 10.1128/JB.185.18.5398-5407.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Römling U, Bokranz W, Rabsch W, Zogaj X, Nimtz M, Tschäpe H. Occurrence and regulation of the multicellular morphotype in Salmonella serovars important in human disease. Int J Med Microbiol. 2003. January 1;293(4):273–85. 10.1078/1438-4221-00268 [DOI] [PubMed] [Google Scholar]
- 24.White AP, Surette MG. Comparative Genetics of the rdar Morphotype in Salmonella. J Bacteriol. 2006. December 4;188(24):8395–406. 10.1128/JB.00798-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Solano C, García B, Valle J, Berasain C, Ghigo J-M, Gamazo C, et al. Genetic analysis of Salmonella enteritidis biofilm formation: critical role of cellulose. Mol Microbiol. 2002. February 28;43(3):793–808. [DOI] [PubMed] [Google Scholar]
- 26.MacKenzie KD, Palmer MB, Köster WL, White AP. Examining the Link between Biofilm Formation and the Ability of Pathogenic Salmonella Strains to Colonize Multiple Host Species. Front Vet Sci. 2017. August 25;4:307–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kingsley RA, Msefula CL, Thomson NR, Kariuki S, Holt KE, Gordon MA, et al. Epidemic multiple drug resistant Salmonella Typhimurium causing invasive disease in sub-Saharan Africa have a distinct genotype. Genome Res. 2009. December 1;19(12):2279–87. 10.1101/gr.091017.109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Holt KE, Parkhill J, Mazzoni CJ, Roumagnac P, Weill F-X, Goodhead I, et al. High-throughput sequencing provides insights into genome variation and evolution in Salmonella Typhi. Nat Genet. 2008. August;40(8):987–93. 10.1038/ng.195 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Holt KE, Thomson NR, Wain J, Langridge GC, Hasan R, Bhutta ZA, et al. Pseudogene accumulation in the evolutionary histories of Salmonella enterica serovars Paratyphi A and Typhi. BMC Genomics. 2009;10(1):36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Yang J, Barrila J, Roland KL, Kilbourne J, Ott CM, Forsyth RJ, et al. Characterization of the Invasive, Multidrug Resistant Non-typhoidal Salmonella Strain D23580 in a Murine Model of Infection. PLoS Negl Trop Dis. 2015. June 19;9(6):e0003839 10.1371/journal.pntd.0003839 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Carden S, Okoro C, Dougan G, Monack D. Non-typhoidal Salmonella Typhimurium ST313 isolates that cause bacteremia in humans stimulate less inflammasome activation than ST19 isolates associated with gastroenteritis. FEMS Pathogens and Disease. 2014. December 24;73(4):2247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Parsons BN, Humphrey S, Salisbury AM, Mikoleit J, Hinton JCD, Gordon MA, et al. Invasive Non-Typhoidal Salmonella Typhimurium ST313 Are Not Host-Restricted and Have an Invasive Phenotype in Experimentally Infected Chickens. PLoS Negl Trop Dis. 2013. October 10;7(10):e2487 10.1371/journal.pntd.0002487 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ramachandran G, Aheto K, Shirtliff ME, Tennant SM. Poor biofilm-forming ability and long-term survival of invasive Salmonella Typhimurium ST313. FEMS Pathog Dis. 2016. June 23;74(5):ftw049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Steenackers H, Hermans K, Vanderleyden J, De Keersmaecker SCJ. Salmonella biofilms: An overview on occurrence, structure, regulation and eradication. Food Res Int. 2012. February 29;45(2):502–31. [Google Scholar]
- 35.Wong VK, Baker S, Pickard DJ, Parkhill J, Page AJ, Feasey NA, et al. Phylogeographical analysis of the dominant multidrug-resistant H58 clade of Salmonella Typhi identifies inter- and intracontinental transmission events. Nat Genet. 2015. May 11;47(6):632–9. 10.1038/ng.3281 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Collinson SK, Emödy L, Müller KH, Trust TJ, Kay WW. Purification and characterization of thin, aggregative fimbriae from Salmonella enteritidis. J Bacteriol. 1991. August 1;173(15):4773–81. 10.1128/jb.173.15.4773-4781.1991 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.González JF, Tucker L, Fitch J, Wetzel A, White P, Gunn JS. Human bile-mediated regulation of Salmonella curli fimbriae. J Bacteriol. 2019. April 1:1–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.White AP, Gibson DL, Grassl GA, Kay WW, Finlay BB, Vallance BA, et al. Aggregation via the Red, Dry, and Rough Morphotype Is Not a Virulence Adaptation in Salmonella enterica Serovar Typhimurium. Infect Immun. 2008. February 21;76(3):1048–58. 10.1128/IAI.01383-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.MacKenzie KD, Wang Y, Shivak DJ, Wong CS, Hoffman LJL, Lam S, et al. Bistable Expression of CsgD in Salmonella enterica Serovar Typhimurium Connects Virulence to Persistence. Infect Immun. 2015. May 12;83(6):2312–26. 10.1128/IAI.00137-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Grantcharova N, Peters V, Monteiro C, Zakikhany K, Römling U. Bistable expression of CsgD in biofilm development of Salmonella enterica serovar Typhimurium. J Bacteriol. 2010. January;192(2):456–66. 10.1128/JB.01826-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Hammar MR, Arnqvist A, Bian Z, Olsén A, Normark S. Expression of two csg operons is required for production of fibronectin- and Congo red-binding curli polymers in Escherichia coli K-12. Mol Microbiol. 1995. November;18(4):661–70. [DOI] [PubMed] [Google Scholar]
- 42.Barnhart MM, Chapman MR. Curli Biogenesis and Function. Annu Rev Microbiol. 2006. October;60(1):131–47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Taylor JD, Zhou Y, Salgado PS, Patwardhan A, McGuffie M, Pape T, et al. Atomic Resolution Insights into Curli Fiber Biogenesis. Structure. 2011. September 7;19(9):1307–16. 10.1016/j.str.2011.05.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Shivak DJ, MacKenzie KD, Watson NL, Pasternak JA, Jones BD, Wang Y, et al. A Modular, Tn 7-Based System for Making Bioluminescent or Fluorescent Salmonella and Escherichia coli Strains. Appl Environ Microbiol. 2016. July 29;82(16):4931–43. 10.1128/AEM.01346-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Ogasawara H, Yamada K, Kori A, Yamamoto K, Ishihama A. Regulation of the Escherichia coli csgD promoter: interplay between five transcription factors. Microbiology. 2010. August 2;156(8):2470–83. [DOI] [PubMed] [Google Scholar]
- 46.Ogasawara H, Yamamoto K, Ishihama A. Regulatory role of MlrA in transcription activation of csgD, the master regulator of biofilm formation in Escherichia coli. FEMS Microbiol Lett. 2010. September 27;312(2):160–8. 10.1111/j.1574-6968.2010.02112.x [DOI] [PubMed] [Google Scholar]
- 47.Singletary LA, Karlinsey JE, Libby SJ, Mooney JP, Lokken KL, Tsolis RM, et al. Loss of Multicellular Behavior in Epidemic African Nontyphoidal Salmonella enterica Serovar Typhimurium ST313 Strain D23580. mBio. 2016. May 4;7(2):e02265–15. 10.1128/mBio.02265-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Okoro CK, Kingsley RA, Connor TR, Harris SR, Parry CM, Al-Mashhadani MN, et al. Intracontinental spread of human invasive Salmonella Typhimurium pathovariants in sub-Saharan Africa. Nat Genet. 2012. November;44(11):1215–21. 10.1038/ng.2423 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Parkhill J, Dougan G, James KD, Thomson NR, Pickard D, Wain J, et al. Complete genome sequence of a multiple drug resistant Salmonella enterica serovar Typhi CT18. Nature. 2001. October 25;413(6858):848–52. 10.1038/35101607 [DOI] [PubMed] [Google Scholar]
- 50.Dieckmann R, Malorny B. Rapid Screening of Epidemiologically Important Salmonella enterica subsp. enterica Serovars by Whole-Cell Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry. Appl Environ Microbiol. 2011. June 7;77(12):4136–46. 10.1128/AEM.02418-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Huang K-J, Igo MM. Identification of the Bases in the ompF Regulatory Region, which Interact with the Transcription Factor OmpR. J Mol Biol. 1996;262(5):615–28. 10.1006/jmbi.1996.0540 [DOI] [PubMed] [Google Scholar]
- 52.Gerstel U, Park C, Römling U. Complex regulation of csgD promoter activity by global regulatory proteins. Mol Microbiol. 2004. January 28;49(3):639–54. [DOI] [PubMed] [Google Scholar]
- 53.Sun L, Vella P, Schnell R, Polyakova A, Bourenkov G, Li F, et al. Structural and Functional Characterization of the BcsG Subunit of the Cellulose Synthase in Salmonella typhimurium. J Mol Biol. 2018;430(18, Part B):3170–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Thongsomboon W, Serra DO, Possling A, Hadjineophytou C, Hengge R, Cegelski L. Phosphoethanolamine cellulose: A naturally produced chemically modified cellulose. Science. 2018. January 19;359(6373):334 10.1126/science.aao4096 [DOI] [PubMed] [Google Scholar]
- 55.Gualdi L, Tagliabue L, Landini P. Biofilm formation-gene expression relay system in Escherichia coli: modulation of sigmaS-dependent gene expression by the CsgD regulatory protein via sigmaS protein stabilization. J Bacteriol. 2007. November;189(22):8034–43. 10.1128/JB.00900-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Wen Y, Ouyang Z, Devreese B, He W, Shao Y, Lu W, et al. Crystal structure of master biofilm regulator CsgD regulatory domain reveals an atypical receiver domain. Prot Sci. Aug 22;26(10):2073–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Gunn JS, Marshall JM, Baker S, Dongol S, Charles RC, Ryan ET. Salmonella chronic carriage: epidemiology, diagnosis, and gallbladder persistence. Trends Microbiol. 2014. November 1;22(11):648–55. 10.1016/j.tim.2014.06.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Crawford RW, Rosales-Reyes R, Ramirez-Aguilar MDLL, Chapa-Azuela O, Alpuche-Aranda C, Gunn JS. Gallstones play a significant role in Salmonella spp. gallbladder colonization and carriage. Proc Natl Acad Sci USA. 2010. March 2;107(9):4353–8. 10.1073/pnas.1000862107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Neiger MR, González JF, Gonzalez-Escobedo G, Kuck H, White P, Gunn JS. Pathoadaptive alteration of Salmonella biofilm formation in response to the gallbladder environment. J Bacteriol. 2019. April 8;:1–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Crawford RW, Reeve KE, Gunn JS. Flagellated but Not Hyperfimbriated Salmonella enterica Serovar Typhimurium Attaches to and Forms Biofilms on Cholesterol-Coated Surfaces. J Bacteriol. 2010. May 26;192(12):2981–90. 10.1128/JB.01620-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Wong HC, Fear AL, Calhoon RD, Eichinger GH, Mayer R, Amikam D, et al. Genetic organization of the cellulose synthase operon in Acetobacter xylinum. Proc Natl Acad Sci USA. 1990. October 1;87(20):8130 10.1073/pnas.87.20.8130 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Römling U, Galperin MY. Bacterial cellulose biosynthesis: diversity of operons, subunits, products, and functions. Trends Microbiol. 2015. September 1;23(9):545–57. 10.1016/j.tim.2015.05.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Stern DL. The genetic causes of convergent evolution. Nat Rev Genet. 2013. November;14(11):751–64. 10.1038/nrg3483 [DOI] [PubMed] [Google Scholar]
- 64.Hammarlöf DL, Kröger C, Owen SV, Canals R, Lacharme-Lora L, Wenner N, et al. Role of a single noncoding nucleotide in the evolution of an epidemic African clade of Salmonella. Proc Natl Acad Sci USA. 2018. March 13;115(11):E2614 10.1073/pnas.1714718115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Osborne SE, Walthers D, Tomljenovic AM, Mulder DT, Silphaduang U, Duong N, et al. Pathogenic adaptation of intracellular bacteria by rewiring a cis-regulatory input function. Proc Natl Acad Sci USA. 2009. March 10;106(10):3982 10.1073/pnas.0811669106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Ilyas B, Mulder DT, Little DJ, Elhenawy W, Banda MM, Pérez-Morales D, et al. Regulatory Evolution Drives Evasion of Host Inflammasomes by Salmonella Typhimurium. Cell Reports. 2018;25(4):825–5. 10.1016/j.celrep.2018.09.078 [DOI] [PubMed] [Google Scholar]
- 67.Chin CY, Tipton KA, Farokhyfar M, Burd EM, Weiss DS, Rather PN. A high-frequency phenotypic switch links bacterial virulence and environmental survival in Acinetobacter baumannii. Nat Microbiol. 2018;3(5):563–9. 10.1038/s41564-018-0151-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Turner KH, Vallet-Gely I, Dove SL. Epigenetic Control of Virulence Gene Expression in Pseudomonas aeruginosa by a LysR-Type Transcription Regulator. PLoS Genet. 2009. December 18;5(12):e1000779 10.1371/journal.pgen.1000779 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Konstantinidis KT, Ramette A, Tiedje JM. The bacterial species definition in the genomic era. Philos Trans R Soc Lond B Biol Sci. 2006. November 29;361(1475):1929–40. 10.1098/rstb.2006.1920 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Cohan FM. Transmission in the Origins of Bacterial Diversity, From Ecotypes to Phyla. Microbiol Spectr. 2017. October 3;5(5):1–26. [DOI] [PubMed] [Google Scholar]
- 71.Mayo AE, Setty Y, Shavit S, Zaslaver A, Alon U. Plasticity of the cis-Regulatory Input Function of a Gene. PLoS Biol. 2006. March 28;4(4):e45 10.1371/journal.pbio.0040045 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Klemm EJ, Gkrania-Klotsas E, Hadfield J, Forbester JL, Harris SR, Hale C, et al. Emergence of host-adapted Salmonella Enteritidis through rapid evolution in an immunocompromised host. Nat Microbiol. 2016. March;1(3):15023 10.1038/nmicrobiol.2015.23 [DOI] [PubMed] [Google Scholar]
- 73.Thiennimitr P, Winter SE, Bäumler AJ. Salmonella, the host and its microbiota. Curr Opin Microbiol. 2012. February 1;15(1):108–14. 10.1016/j.mib.2011.10.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Veening J-W, Smits WK, Kuipers OP. Bistability, Epigenetics, and Bet-Hedging in Bacteria. Annu Rev Microbiol. 2008. October;62(1):193–210. [DOI] [PubMed] [Google Scholar]
- 75.Ackermann M. A functional perspective on phenotypic heterogeneity in microorganisms. Nat Rev Microbiol. 2015. August;13(8):497–508. 10.1038/nrmicro3491 [DOI] [PubMed] [Google Scholar]
- 76.Ahmad I, Lamprokostopoulou A, Le Guyon S, Streck E, Barthel M, Peters V, et al. Complex c-di-GMP Signaling Networks Mediate Transition between Virulence Properties and Biofilm Formation in Salmonella enterica Serovar Typhimurium. PLoS ONE. 2011. December 2;6(12):e28351–15. 10.1371/journal.pone.0028351 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Lamprokostopoulou A, Monteiro C, Rhen M, Römling U. Cyclic di-GMP signalling controls virulence properties of Salmonella enterica serovar Typhimurium at the mucosal lining. Environ Microbiol. 2010. January;12(1):40–53. 10.1111/j.1462-2920.2009.02032.x [DOI] [PubMed] [Google Scholar]
- 78.Pontes MH, Lee E-J, Choi J, Groisman EA. Salmonella promotes virulence by repressing cellulose production. Proc Natl Acad Sci USA. 2015. April 21;112(16):5183–8. 10.1073/pnas.1500989112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Oppong GO, Rapsinski GJ, Newman TN, Nishimori JH, Biesecker SG, Tükel Ç. Epithelial Cells Augment Barrier Function via Activation of the Toll-Like Receptor 2/Phosphatidylinositol 3-Kinase Pathway upon Recognition of Salmonella enterica Serovar Typhimurium Curli Fibrils in the Gut. Infect Immun. 2013. February 1;81(2):478 10.1128/IAI.00453-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Tükel Ç, Raffatellu M, Humphries AD, Wilson RP, Andrews-Polymenis HL, Gull T, et al. CsgA is a pathogen-associated molecular pattern of Salmonella enterica serotype Typhimurium that is recognized by Toll-like receptor 2. Mol Microbiol. 2005. August 26;58(1):289–304. 10.1111/j.1365-2958.2005.04825.x [DOI] [PubMed] [Google Scholar]
- 81.Tükel Ç, Nishimori JH, Wilson RP, Winter MG, Keestra AM, Van Putten JPM, et al. Toll-like receptors 1 and 2 cooperatively mediate immune responses to curli, a common amyloid from enterobacterial biofilms. Cell Microbiol. 2010. May 18;12(10):1495–505. 10.1111/j.1462-5822.2010.01485.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Rapsinski GJ, Wynosky-Dolfi MA, Oppong GO, Tursi SA, Wilson RP, Brodsky IE, et al. Toll-Like Receptor 2 and NLRP3 Cooperate To Recognize a Functional Bacterial Amyloid, Curli. Infect Immun. 2015. January 14;83(2):693–701. 10.1128/IAI.02370-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Nishimori JH, Newman TN, Oppong GO, Rapsinski GJ, Yen J-H, Biesecker SG, et al. Microbial Amyloids Induce Interleukin 17A (IL-17A) and IL-22 Responses via Toll-Like Receptor 2 Activation in the Intestinal Mucosa. Infect Immun. 2012. December 1;80(12):4398 10.1128/IAI.00911-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Amarasinghe JJ, D 039 Hondt RE, Waters CM, Mantis NJ. Exposure of Salmonella enterica Serovar Typhimurium to a Protective Monoclonal IgA Triggers Exopolysaccharide Production via a Diguanylate Cyclase-Dependent Pathway. Infect Immun. 2013. March 1;81(3):653 10.1128/IAI.00813-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Smith RD, Price EJ, Burby EP, Blanco PL, Chamberlain J, Chapman RM. The Production of Curli Amyloid Fibers Is Deeply Integrated into the Biology of Escherichia coli. Biomolecules. 2017;7(4). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Anriany YA, Weiner RM, Johnson JA, De Rezende CE, Joseph SW. Salmonella enterica Serovar Typhimurium DT104 Displays a Rugose Phenotype. Appl Environ Microbiol. 2001. September 1;67(9):4048 10.1128/AEM.67.9.4048-4056.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Scher K, Römling U, Yaron S. Effect of Heat, Acidification, and Chlorination on Salmonella enterica Serovar Typhimurium Cells in a Biofilm Formed at the Air-Liquid Interface. Appl Environ Microbiol. 2005. March 1;71(3):1163 10.1128/AEM.71.3.1163-1168.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Finn S, Condell O, McClure P, Amézquita A, Fanning S. Mechanisms of survival, responses and sources of Salmonella in low-moisture environments. Front Microbiol. 2013;4:331 10.3389/fmicb.2013.00331 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Kariuki S, Onsare RS. Epidemiology and Genomics of Invasive Nontyphoidal Salmonella Infections in Kenya. Clin Infect Dis. 2015. November 1;61 Suppl 4(suppl 4):S317–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Kariuki S, Revathi G, Kariuki N, Kiiru J, Mwituria J, Muyodi J, et al. Invasive multidrug-resistant non-typhoidal Salmonella infections in Africa: zoonotic or anthroponotic transmission? J Med Microbiol. 2006;55(5):585–91. [DOI] [PubMed] [Google Scholar]
- 91.Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 2012. June 11;28(12):1647–9. 10.1093/bioinformatics/bts199 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Martínez-García E, de Lorenzo V. Engineering multiple genomic deletions in Gram-negative bacteria: analysis of the multi-resistant antibiotic profile of Pseudomonas putida KT2440. Environ Microbiol. 2011. August 24;13(10):2702–16. 10.1111/j.1462-2920.2011.02538.x [DOI] [PubMed] [Google Scholar]
- 93.Silva-Rocha R, Martínez-García E, Calles B, Chavarría M, Arce-Rodríguez A, las Heras de A, et al. The Standard European Vector Architecture (SEVA): a coherent platform for the analysis and deployment of complex prokaryotic phenotypes. Nucleic Acids Res. 2012. November 22;41(D1):D666–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Wang Y, Chen X, Hu Y, Zhu G, White AP, Köster W. Evolution and Sequence Diversity of FhuA in Salmonella and Escherichia. Infect Immun. 2018. November;86(11):181–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(A) Morphological comparison of colonies grown for five days at 28˚C or 37˚C on T agar supplemented with 40 μg mL-1 Congo red. (B) Maximum expression from a curli-specific reporter (csgBAC::luxCDABE) in each strain grown for 48 h at 28˚C or 37˚C. Luciferase expression was measured every 30 min during continuous culture.
(TIF)
Multicellular aggregates and planktonic cells formed from liquid cultures of S. Typhimurium 14028 and S. Typhimurium D23580. Images match those presented in Fig 1, but have been enhanced to emphasize aggregates within the samples. Aggregates formed by S. Typhimurium D23580 cells appear structurally distinct from aggregates formed by S. Typhimurium 14028 cells.
(TIF)
(A) Whole cell lysates were generated from multicellular aggregates and planktonic cells isolated from flask cultures of Salmonella strains after 24 hours of growth and probed for synthesis of RpoS. Lysates were normalized by total protein concentration. GroEL was used as a loading control to ensure that equal amounts of protein were loaded into each sample lane. (B) RpoS activity was evaluated by measuring luminescence from a synthetic, RpoS-dependent promoter-reporter construct expressed in each Salmonella strain during 48 hours of growth. Graphed values represent the maximum reporter activity recorded and is reported as counts per second (CPS). (C) Absorbance measurements represent strain growth in microaerophilic conditions in 96-well microtiter plates. Each curve represents one biological replicate; n = 4 per strain. (D) Time course of RpoS-dependent promoter activity in each indicated strain.
(TIF)
Each curve represents one biological replicate; n = 3.
(TIF)
Promoter-reporter constructs derived from S. Typhimurium 14028 adrA and cpxRA promoter sequences were introduced into each of the strains. Letters above the bars indicate mean values that were statistically similar to (black font) or different from (red font) other mean values. #, values below the activity threshold as established in [24]. Each bar represents the mean value from 3–5 independent biological replicates and error bars represent the standard deviations.
(TIF)
Clones that contain (-47T>T) or do not contain (-47T>C) the identified ‘T’ promoter SNP at position -47 were grown in flasks of 1% tryptone for 24 hours before being evaluated for the ability to form aggregates in liquid cultures.
(TIF)
Simplified multiple sequence alignment of the csgDEFG and csgBAC operons highlighting serovar- and strain-specific single nucleotide polymorphisms representing both nonsynonymous and synonymous mutations: S. Typhimurium and D23580 (yellow), S. Enteritidis and D7795 (light blue), and S. Typhi (pink). Other highlighted changes (black) include the SNP in D7795 that inactivates csgD transcription, and the SNP in S. Typhi strains that introduces a premature stop codon in csgD, yielding a CsgD protein that is truncated by 8 amino acids. The long black bar represents the DNA region with nucleotide position numbers listed above and csg genes shown below as yellow-boxed arrows. Special sequence features involved in operon regulation are highlighted above and below the black bar: -35 and -10 promoter regions (black elbow arrows), H-NS binding region (grey box), CpxR-binding sites (black bars), and OmpR binding regions (hatched boxes).
(TIF)
Simplified multiple sequence alignment of the bcsRQABZC and bcsEFG operons highlighting non-synonymous serovar- and strain-specific single nucleotide polymorphisms. Non-synonymous SNPs are shown in yellow; black bars indicate SNPs that yield premature stop codons (i.e. bcsG in D23580; four SNPs in bcsC in S. Typhi strains). The long black bar represents the DNA region with nucleotide position numbers listed above and bcs genes shown below as yellow-boxed arrows. bcsR, bcsF and a small hypothetical protein are shown without their names listed.
(TIF)
(PDF)
(PDF)
(XLSX)
(XLSX)
(XLSX)
(XLSX)
Data Availability Statement
All raw numerical data and statistical analysis has been deposited in figshare (https://figshare.com/) and is publicly available at: doi: https://doi.org/10.6084/m9.figshare.8220866.v1.