

Abstract
In epithelial-derived cancers, altered regulation of cell-cell adhesion facilitates the disruption of tissue cohesion that is central to the progression to malignant disease. Although numerous intercellular adhesion molecules participate in epithelial adhesion, the immunoglobulin superfamily (IgSF) member activated leukocyte cell adhesion molecule (ALCAM), has emerged from multiple independent studies as a central contributor to tumor progression. ALCAM is an archetypal member of the IgSF with conventional organization of five Ig-like domains involved in homo-and heterotypic adhesions. Like many IgSF members, ALCAM is broadly expressed and involved in cellular adhesion across many cellular processes. While the redundancy of intercellular adhesion molecules (CAMs) could diminish the impact of any single CAM, consistent correlation between ALCAM expression and patient outcome for multiple cancers underscores its role in tumor progression. Unlike most oncogenes and tumor suppressors, ALCAM is neither mutated nor amplified or deleted. Experimental disruption of ALCAM-mediated adhesions implies that this IgSF member contributes to tumor progression through dynamic turnover of the protein at the cell surface. Since ALCAM is not frequently altered at the gene level, it appears to promote malignant behavior through regulation of its availability rather than its specific activity. These observations help explain its heterogeneous expression within malignant disease and the drastic changes in protein levels across tumor progression. To reveal how ALCAM contributes to tumor progression, we review regulation of its gene expression, alternative splicing, targeted proteolysis, binding partners, and surface shedding within the context of cancer. Studying ALCAM regulation has led to a novel understanding of the fine-tuning of cell adhesive state through the utilization of otherwise normal regulatory processes, which thereby enable tumor cell invasion and metastasis.
Keywords: ALCAM/CD166 regulation, ALCAM/CD166 alternative splicing, cell adhesion, dynamic cell adhesion, CAM regulation
Introduction:
The formation, organization, and maintenance of multi-cellular tissues is mediated in large part through cell adhesion molecules (CAMs). These molecules don’t necessarily provide the adhesive strength required for physical cohesion, but rather enable the cell-cell sensing needed to resolve tissue-specific patterning. Coordinating the dynamic function of CAMs is crucial for embryogenesis, the formation of differentiated tissues during morphogenesis, and wound healing. While the stimuli that initiate changes in cell adhesion range widely from mechanical disruption, to growth factor stimulation and intrinsic activity (such as cell division or apoptosis), the underlying dynamics that allow for the formation and dissociation of adhesive interactions are conserved. Both cell-cell and cell-matrix adhesions are subject to such dynamics and each adhesion of each cell is performed by members of four main CAM families: cadherins, IgSF, selectins, and integrins (Table 1). While each family contains multiple members, the IgSF is by far the largest and most diverse family with members characterized by a typical Ig-fold of sandwiched anti-parallel beta sheets. As such, IgSF members are not limited to CAMs, but rather extend across many subfamilies such as antigen receptors, growth factor receptors, costimulatory/inhibitory molecules, etc. This review details the contribution of Activated Leukocyte Cell Adhesion Molecule (ALCAM/CD166), an archetypal IgSF member, in the dynamic regulation of cell adhesion and provides insight into the broader significance of non-permanent dynamics of cell adhesion in cancer, its regulation, and the potential for leveraging this process in favor of reversing malignant behavior.
Table 1.
Summary of CAM family characteristics.
| CAM Family/Approximate Number of Genes: | Subgroups/Approximate Number of Genes: | Basic Structural Properties/Domain Organization: | General Biological Functions: | Refs: |
|---|---|---|---|---|
| Cadherin Family - 125 | • Major Cadherins- 32 • Protocadherins – 76 • Cadherin Related - 17 |
Characterized by the presence of at least two consecutive extracellular cadherin repeats with conserved calcium-binding pockets | Exhibit homotypic cell-cell adhesion, although heterotypic binding is possible, thus providing selectivity for cell sorting and transmitting cell-cell communication | 1, 5, 11, 12 |
| Integrin Family - 24 | • α-subunits - 18 • β-subunits – 8 |
Type I transmembrane proteins containing large extracellular domains and generally short cytoplasmic domains. Functional receptors are heterodimers of an α and βsubunit. | Engage extracellular matrix macromolecules and/or counter-receptors on adjacent cell surfaces producing extracellular adhesions and intracellular signaling cascades. | 2, 5, 9, 10 |
| Selectin Family - 3 | • E-selectin – 1 • P-selectin – 1 • L-selectin - 1 |
Single-chain transmembrane glycoproteins related to Ctype lectins through amino terminus similarities and calcium-dependent binding. | Mediates leukocyte cell tethering and rolling on endothelium through the recognition of sialyl-Lewis X tetrasaccharide carbohydrate moieties on glycoprotein ligands. | 3, 5, 7, 8 |
| Immunoglobulin Superfamily – 667+ | • C1-set domain containing −42 • C2-set domain containing - 40 • Ig-like domain containing −261 • I-set domain containing −161 • V-set domain containing −163 |
Possess at least one Igdomain or Ig-like domain characterized by a domain size of ~100 amino acids, the number of beta-strands, and general topology of the Igfold. | Members function as cell adhesion molecules and antigen/cytokine/growth factor receptors and coreceptors important for adhesion, binding, signaling, and recognition processes. | 4–6 |
Cell Adhesion Molecule Families:
The Cadherin Family:
Members of the cadherin family form relatively strong, calcium-dependent, homotypic adhesions between adjacent cells through a variable number of cadherin-like domains. These proteins facilitate mechanical cohesion as well as cell-cell recognition crucial for organogenesis and maintaining tissue organization [1].
The Integrin Family:
Integrin receptors are calcium-dependent heterodimers composed of non-covalently associated alpha and beta subunits. Members of the integrin family molecules link the extracellular matrix (ECM) and cytoskeleton. This family is capable of bi-directional signaling, which in turn allows for control not only over the physical mechanics of adhesion and migration through inside-out signaling, but also cellular polarity, cytoskeletal structure, and survival through outside-in signaling [2].
Selectin Family:
Members of the selectin family consist of an N-terminal calcium-dependent lectin type domain, an EGF-like domain, and variable numbers of short repeats homologous to complement-binding sequences. They exhibit transient cell-cell adhesions, through ligand interactions with sialyl-Lewis X tetrasaccharide carbohydrate moieties required for minimal recognition, on glycoproteins and glycolipids. This CAM family is well known for their role in leukocyte migration during inflammation. Reversible interactions between selectins on leukocytes and their ligands on activated endothelial cells facilitates leukocyte tethering which, if followed by the activation of integrins, potentiates rolling and firm cell adhesion to the vascular wall. This is a representative example of molecular synergy among CAMs from structurally and functionally distinct families [3].
Immunoglobulin Superfamily (IgSF):
IgSF proteins mediate calcium independent cell-cell adhesion through Ig-like domains and recognize both homophilic and heterophilic ligands. These interactions are not as strong as cadherin- or integrin-mediated adhesion, but members of the IgSF seem to contribute more to the fine-tuning, potentiation, and regulation of these adhesive interactions during tissue formation and maintenance [4].
Cell Adhesion and Cancer:
In normal tissues, adhesion complexes form in order to define specific tissue compartments, establish barrier functions, and enable cell migration. Importantly, adhesion mediated by CAMs is a tunable process in which adhesive state is dynamically regulated.
Dysregulation of CAMs contributes to cancer progression and metastasis through coordinated changes in cell adhesion to promote the motility of tumor cells and allow for migration into surrounding tissue. Alterations in the expression of CAMs are well documented in association with tumor progression, but the mechanisms controlling the tunable modulation of their activity remain poorly understood. A better understanding of what molecular changes occur and how they are altered is essential for understanding how cancer overcomes physiologically imposed restrictions on tissue aberration and better target potential sites for therapeutic intervention.
While there are no significant genomic alterations in ALCAM for any cancers, ALCAM expression has been reported to decrease during the formation and subsequent malignant progression of solid tumors [13–20]. In addition, changes in ALCAM mRNA expression do not always correlate with protein expression, suggesting there are multiple levels of regulation being employed [21]. Since ALCAM alterations have been used as prognostic markers for many solid tumors, this CAM has emerged as a functional contributor to cancer progression [13–20]. The main focus will be to illustrate direct regulatory mechanisms affecting ALCAM expression while emphasizing the functional outcome affecting cell adhesion.
Activated Leukocyte Cell Adhesion Molecule (ALCAM):
ALCAM is a member of a small subgroup within the IgSF characterized by transmembrane glycoproteins with five extracellular immunoglobulin (Ig)-like domains composed of two amino-terminal membrane distal variable-(V)-domains, three membrane proximal constant-(C2)-domains, one transmembrane region, and a short, carboxy-terminal cytoplasmic tail of varying length (Figure 1). These Ig-like domains mediate homophilic ALCAM-ALCAM interactions and heterophilic ALCAM-CD6 interactions. Cis-oligomerization occurs through lateral oligomerization of ALCAM molecules interacting through the three membrane proximal C-type domains, while trans ALCAM-ALCAM interactions involve the amino-terminal ligand binding modules (V-domains) [22].
Figure 1.
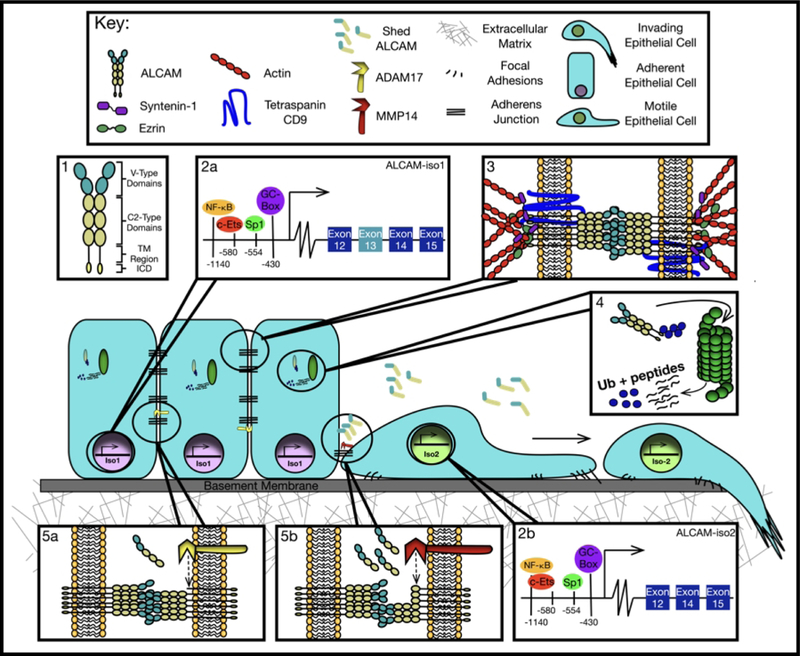
ALCAM regulation for tunable cell adhesion. (1) ALCAM is composed of five Ig-like domains, two V-type domains and three C2-type domains, a transmembrane (TM) region, and a small intracellular domain (ICD). ALCAM gene expression (2a/b) is regulated by the promoter elements NF-κB, Ets, Sp1, and a GC-box upstream of the translation start site (TSS). This molecule associates with the tetraspanin CD9 to enhance homotypic ALCAM-ALCAM interactions and facilitate clustering. It also associates with the actin cytoskeleton to strengthen cell adhesion through adaptors syntenin-1 and ezrin (3). ALCAM protein stability is regulated by CHIP mediated ubiquitination and subsequent proteasomal degradation (4). Alternative splicing affects the proteolytic susceptibility of ALCAM (4). ADAM17 proteolysis of iso1 promotes high cell-cell adhesion through low basal shedding (5a), while MMP14 proteolysis of iso2 enables cell motility through high basal shedding and disruption of ALCAM-ALCAM interactions (5b).
This molecule’s expression is described in many cell types including hematopoietic stem cells, cancer stem cells, intestinal epithelial crypts, the central nervous system, microvascular endothelium, retinal vascular endothelial cells, and motor neurons [23–29]. Its broad tissue distribution in a wide variety of cells confers involvement in a vast scope of processes including epithelial migration, neuronal migration and differentiation, axon growth and pathfinding, and hematopoiesis [26–29]. The extensive tissue expression and functions of ALCAM speak to its ability to not only confer adhesion, but also act as a marker of cell identity. Being a junctional adhesion molecule, ALCAM is found at sites of cell-cell contact in epithelial, neuronal, stromal, and mesenchymal tissues.
Regulation of ALCAM:
ALCAM is regulated by transcription, alternative splicing, trafficking to the membrane, binding partners, and proteolysis. The protein’s adhesive ability is regulated through subcellular localization and availability of its extracellular domain and therefore is limited by intercellular binding partners and potential extracellular engagement of heterotypic and homotypic interactions. These are detailed below and summarized in Figure 1.
Gene Expression:
The ALCAM gene is located on the long arm of human chromosome 3 (3q13.11) with the most prominent transcription start site 349 bp upstream of the translation start site [30, 31]. The ALCAM promoter contains CpG rich sequences, a canonical Sp1 element, and a functional p65 NF-κB motif [31, 32]. Other putative transcription factor binding sites have also been identified by Tan, F. et al. These cis-acting elements contribute to ALCAM expression by regulating promoter activity.
The Sp1 site is located within the proximal region of the ALCAM promoter at −550 bp upstream of the translation start site and adjacent to an Ets site, which can establish cooperative transcriptional activation with Sp1. However, optimum ALCAM promoter activity requires only the Sp1 element [33, 34]. Changes in ALCAM expression have been seen upon Sp1 inhibition through siRNA mediated knockdown. In stem cells isolated from two colorectal cancer (CRC) cell lines, Sp1 knockdown suppressed ALCAM mRNA expression and proliferation while increasing apoptosis. Conversely, over-expression of Sp1 leads to an increase in ALCAM expression [35].
Upstream of the Sp1 element is the NF-κB consensus sequence at the putative site −1140 [31, 32]. Wang, J. et al. describes how the p50/p65 heterodimer increases ALCAM expression upon serum deprivation (SD) in melanoma cell lines, but not breast cancer cell lines. These conditions potentially model a poorly vascularized, thus nutrient poor, tumor microenvironment that forces cells to rely on survival mechanisms such as apoptosis and autophagy. This is interesting because translocation of p50/p65 to the ALCAM promoter appeared to have a time dependent effect on ALCAM protein and mRNA expression. While expression of both entities increased after 24 hours after SD, protein expression significantly decreased after 48 hours [36]. However, there are no studies elucidating other mechanisms of NF-kB induction on ALCAM transcription. This could be crucial in understanding the immunological aspect of ALCAM transcription regulation by connecting NF-kB pro-inflammatory signaling to cell adhesion.
Membrane Organization:
Cell-surface localization of ALCAM is controlled, in part, by trafficking to the membrane and association with the actin cytoskeleton and binding partner CD9 [37, 38]. ALCAM has 10 potential N-linked glycosylation sites further characterized in the melanoma cell line A375 as β 1–6 branched oligosaccharides by mass spectrometry [40] While no functional studies have been performed determining the role of these N-glycans on ALCAM post-golgi trafficking, they could contribute to sorting at apical membranes of polarized cells [41]. Based on co-immunoprecipitation and co-localization experiments, the tetraspanin CD9 interacts with ALCAM on leukocyte membranes [38]. Tetraspanins are a conserved family of protein organizers consisting of four transmembrane domains that form a small extracellular loop (SEL) and large extracellular loop (LEL). Tetraspanins typically interact with themselves and other proteins, both cytoplasmic and membrane bound, forming tetraspanin enriched microdomains (TEMs) [42]. These TEMs are endowed with distinct functional specialties ranging from modulation of growth factor signaling to integrin-dependent cell adhesion [43]. By using CD9-LEL mutants, optimal ALCAM interaction required the variable region within the LEL. CD9 therefore interacts with ALCAM through its LEL and was found to enhance homotypic adhesion by facilitating cis-clustering [38]. While ALCAM lacks a direct actin-binding site, it is anchored to the cytoskeleton through the scaffolding proteins syntenin-1 and ezrin, which interact with the cytoplasmic tail of ALCAM [44]. Membranous ALCAM is clearly linked to the actin cytoskeleton because lateral mobility is enabled upon inhibition of actin polymerization by cytochalasin D. This in turn allows for the formation of high avidity clusters ultimately affecting adhesion strength [37]. Altogether, ALCAM intracellular membrane distribution is regulated by the formation of CD9-associated ALCAM microdomains and subsequent redistribution upon ligand engagement whereby ALCAM is recruited to sites of trans-interaction. Thus, regulation of ALCAM availability through surface distribution is critical to its ability to contribute to cell-cell adhesion.
Protein Degradation:
Post-translational turnover is especially important in the protein homeostasis of membrane proteins like ALCAM whose activity is primarily regulated by availability. The E3 ubiquitin ligase, Carboxy-terminus of Hsc70 Interacting Protein (CHIP), was found to increase ubiquitination and subsequent proteasomal degradation of ALCAM in head and neck cancer. Using multiple HNC cell lines, CHIP over-expression studies revealed increased ALCAM protein turnover, while mRNA levels remained unaffected. CHIP has also been shown to suppress tumor progression in breast cancer, which correlated with high expression in non-aggressive cell lines [45]. These observations demonstrate how protein stability regulates ALCAM function through membrane availability.
Proteolytic Shedding and the Consequences of Alternative Splicing:
ALCAM has nine recognized splice variants, four of which have open reading frames. Of these, two splice variants have been confirmed at both the mRNA and protein level. Canonical or full length ALCAM, from here on referred to as ALCAM-isoform1 (iso1), contains all 15 coding exons, while ALCAM-isoform2 (iso2), lacks exon 13, resulting in a 13 amino acid truncation of the stalk region. The gene structure analyses of these isoforms are described in detail by Hebron and colleagues [46]. Even though both isoforms contain the terminal Ig-domains responsible for adhesion, iso1 promotes cell-cell adhesion while iso2 enables single cell dispersion. These phenotypes are a result of differential proteolytic susceptibility of the two isoforms [46].
Like many receptors, ALCAM is proteolytically shed from the cell surface and released into interstitial fluid from which it enters circulation. Early studies identified the transmembrane protease a disintegrin and metalloprotease 17/tumor necrosis factor α converting enzyme (ADAM17/TACE) as being responsible for ALCAM shedding in ovarian cancer cells [47]. ADAM17-mediated shedding was verified in a metastatic prostate cancer cell line (PC3) and readily detected in the circulation of tumor-bearing mice as well as cancer patients [48]. ADAM17 cleaves its substrates in the stalk region of transmembrane proteins located on the extracellular membrane-proximal part of the molecule [49, 50]. The stalk region of ALCAM is encoded by exons 12–14 at amino acids 502–527. The presence of an alternative splice variant that lacked exon 13 (iso2) implied that a protease resistant ALCAM variant might exist. However, detailed protein-structure analysis studies revealed that the loss of exon 13 in iso2 enabled proteolytic cleavage of ALCAM by matrix metalloproteinase 14/matrix-type 1-metalloproteinase (MMP14/MT1-MMP) at a distal site in the fourth Ig-like domain [46]. These findings demonstrated that iso2 is shed at 10-times the rate of iso1 and explains why iso1 is able to maintain cohesion, despite being shed, while iso2 enables single-cell dispersion. While the specific need for two distinct mechanisms of shedding has yet to be investigated, we speculate that the shedding of iso1 by ADAM17 is a low shedding rate needed to turn-over adhesion between cells, while shedding of iso2 by MMP14 entails a high rate shedding that facilitates single-cell movement. This observation implies that proteolytic shedding of ALCAM is not merely regulated through availability or activity of the protease, but can be finely tuned by controlling the splicing of ALCAM post-transcriptionally to change the balance between a proteolytically-susceptible and proteolytically-resistant isoform (iso2 vs iso1 respectively). Therefore, there is an interdependency between isoform expression and protease activity that is leveraged depending on the state of tissue cohesion. The increase of iso2 expression in bladder cancer tissue compared to normal bladder is consistent with the idea that tumor malignancy favors a less cohesive and more readily dispersed behavior [46].
Proteolysis by both ADAM17 and MMP14 results in the release of soluble forms of ALCAM (sALCAM) containing most of the ectodomain. In general, these molecules are antagonistic to cell-surface ALCAM and disrupt ALCAM-mediated cell-cell adhesion which, in turn, promotes migration in both tumor and normal cells.
Considering that ALCAM is shed by cancer, where this shedding increases as a function of malignancy and metastasis [47–49], the detection of sALCAM could function as a biomarker of cancer progression. Several studies have evaluated ALCAM in ascites fluid, urine, and blood serum of cancer patients, and provide an increasingly robust indication for using sALCAM as a non-invasive means of disease monitoring [16, 39, 50]. Indeed, ALCAM shedding has prognostic implications across many types, including bladder, cervical, prostate, colorectal, and pancreatic cancers [16,17, 48, 51, 52]. The discovery that ALCAM shedding is regulated in part by its alternative splicing makes ALCAM shedding particularly intriguing as a biomarker of cancer malignancy because it implies that ALCAM shedding exists at the intersect between intrinsic regulation of gene expression and extrinsic regulation of protein activity. As such, it functions as an indicator of phenotype rather than genotype.
Concluding Remarks:
Cell adhesion is a crucial part of the formation and maintenance of tissue architecture. Molecular mechanisms of adhesion not only define the (spatial) organization and infrastructure of differentiated epithelial cell layers, but also enable the interpretation of external signals involved in cell-cell communication. While no adhesion molecule acts in isolation, we chose to focus this review on the regulation of the IgSF member ALCAM to show how dynamic regulation of this one molecule through distinct, but coordinated regulatory mechanisms, can enable fine-tuning of cell adhesion. The fact that this single molecule from a family of 600+ members can significantly impact cellular cohesion and demonstrate clinical relevance clarifies the enormous dynamic range in cellular adhesion that can be achieved with the IgSF members. While this has been recognized for some time in the field of developmental biology, the relevance of these processes in controlling the malignant phenotype independently from the underlying genotype has been under-appreciated in the area of cancer biology. Lessons learned from investigating ALCAM suggest that dynamic regulation of cell adhesion is essential for defining the differences between normal and malignant phenotype.
Understanding how the tuning of cell adhesion controls the malignant phenotype will have ramifications at both at the mechanistic and clinical level. For instance, dynamic turn-over of ALCAM-ALCAM adhesions not only disrupts the mechanics of cell-cell adhesion but also disables the intracellular association with syntenin-1 and ezrin which impacts not only this distribution of these scaffolding molecules, but all of the associated biological processes including actin organization, g-protein mediated signaling, and possibly vesicle biogenesis. Consequently, adhesion molecules such as ALCAM become much more than iterative additions to the cell adhesion milieu of normal and tumor tissue. The lack of any significant phenotype in the ALCAM knockout mouse initially diminished enthusiasm for finding a therapeutic intervention that could be effective. However, by gaining an understanding of the underlying regulatory dynamics, it has become possible to target its activity and direct the adhesive properties to benefit clinically. Published examples include targeting the alternative splicing to promote an adhesive phenotype [46] or treatment with an antagonistic molecule such as a competing peptide [53], sALCAM ectodomain [46], ALCAM blocking antibodies [54] or antibodies against the ALCAM ligand, CD6 [55].
While the focus of this review is limited to regulation of ALCAM-ALCAM interactions, ALCAM-CD6 interactions have emerged as a promising target in the cancer immunotherapy field. CD6 is a cognate ALCAM ligand expressed on T-cells and a small subset of B-cells that functions both as a costimulatory molecule for T-cell activation/proliferation and as an adhesion molecule to promote thymocyte-thymic epithelial cell interactions [56–60]. Previous studies primarily concentrate on ALCAM-CD6 in autoimmune diseases, such as multiple sclerosis, rheumatoid arthritis, and psoriasis, to inhibit leukocyte transendothelial migration to inflammatory sites [61–64]. There are very preliminary reports of ALCAM-independent CD6 targeting in hematological malignancies, however it is important to note that only less than five patients were evaluated in each study. Using the mouse anti-human CD6 mAb IOR-T1 in cutaneous T-cell lymphoma to topically treat resulting skin lesions induced clinical and histopathological regression based on CD6+ T-cell depleting properties [65]. Treatment with the fully humanized CD6 mAb T1h (Itolizumab) in patients with chronic lymphocytic leukemia also showed clinical improvement [66]. The first reports of targeting ALCAM/CD6 specific interactions in a solid tumor have been shown to yield promising results for the immunotherapy field by means of T-cell homing in brain cancer [67].
Identifying therapies that target the malignant phenotype without being restricted to the underlying genotype is increasingly important in a clinical setting that recognizes the heterogeneity of cancer as a disease and the need for individualized therapy. Consequently, we see the diversity of adhesion mechanisms not as a hindrance to our understanding of cancer progression and metastasis, but rather as a wealth of possible mechanisms by which we can re-direct and eliminate the malignant phenotype. ALCAM is an example of this approach, but numerous other mechanisms are likely to offer similar opportunities.
References:
- 1.Ivanov DB, Philippova MP, Tkachuk VA (2001) Structure and functions of classical cadherins. Biochemistry Mosc 66:1174–1186. [DOI] [PubMed] [Google Scholar]
- 2.Barczyk M, Carracedo S, Gullberg D (2010) Integrins. Cell Tissue Res 339:269–280. doi: 10.1007/s00441-009-0834-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.McEver RP (2015) Selectins: initiators of leucocyte adhesion and signalling at the vascular wall. Cardiovasc Res 107:331–339. doi: 10.1093/cvr/cvv154 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wai Wong C, Dye DE, Coombe DR (2012) The role of immunoglobulin superfamily cell adhesion molecules in cancer metastasis. Int J Cell Biol 2012:340296. doi: 10.1155/2012/340296 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Yates B, Braschi B, Gray KA, et al. (2017) Genenames.org: the HGNC and VGNC resources in 2017. Nucleic Acids Res 45:D619–D625. doi: 10.1093/nar/gkw1033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Williams AF, Barclay AN (1988) The immunoglobulin superfamily--domains for cell surface recognition. Annu Rev Immunol 6:381–405. doi: 10.1146/annurev.iy.06.040188.002121 [DOI] [PubMed] [Google Scholar]
- 7.Vestweber D (2015) How leukocytes cross the vascular endothelium. Nat Rev Immunol 15:692–704. doi: 10.1038/nri3908 [DOI] [PubMed] [Google Scholar]
- 8.Ley K, Kansas GS (2004) Selectins in T-cell recruitment to non-lymphoid tissues and sites of inflammation. Nat Rev Immunol 4:325–335. [DOI] [PubMed] [Google Scholar]
- 9.Bökel C, Brown NH (2002) Integrins in Development: Moving on, Responding to, and Sticking to the Extracellular Matrix. Developmental Cell 3:311–321. [DOI] [PubMed] [Google Scholar]
- 10.Hynes RO (2002) Integrins: bidirectional, allosteric signaling machines. Cell 110:673–687. [DOI] [PubMed] [Google Scholar]
- 11.Gumbiner BM (2005) Regulation of cadherin-mediated adhesion in morphogenesis. Nature Reviews Molecular Cell Biology 6:622–634. doi: 10.1038/nrm1699 [DOI] [PubMed] [Google Scholar]
- 12.Harris TJC, Tepass U (2010) Adherens junctions: from molecules to morphogenesis. Nature Reviews Molecular Cell Biology 11:502–514. doi: 10.1038/nrm2927 [DOI] [PubMed] [Google Scholar]
- 13.Minner S, Kraetzig F, Tachezy M, et al. (2011) Low activated leukocyte cell adhesion molecule expression is associated with advanced tumor stage and early prostate-specific antigen relapse in prostate cancer. Hum Pathol 42:1946–1952. doi: 10.1016/j.humpath.2011.02.017 [DOI] [PubMed] [Google Scholar]
- 14.Burandt E, Bari Noubar T, Lebeau A, et al. (2014) Loss of ALCAM expression is linked to adverse phenotype and poor prognosis in breast cancer: a TMA-based immunohistochemical study on 2,197 breast cancer patients. Oncol Rep 32:2628–2634. doi: 10.3892/or.2014.3523 [DOI] [PubMed] [Google Scholar]
- 15.Lugli A, Iezzi G, Hostettler I, et al. (2010) Prognostic impact of the expression of putative cancer stem cell markers CD133, CD166, CD44s, EpCAM, and ALDH1 in colorectal cancer. Br J Cancer 103:382–390. doi: 10.1038/sj.bjc.6605762 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Arnold Egloff SA, Du L, Loomans HA, et al. (2017) Shed urinary ALCAM is an independent prognostic biomarker of three-year overall survival after cystectomy in patients with bladder cancer. Oncotarget 8:722–741. doi: 10.18632/oncotarget.13546 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ihnen M, Kress K, Kersten JF, et al. (2012) Relevance of activated leukocyte cell adhesion molecule (ALCAM) in tumor tissue and sera of cervical cancer patients. BMC Cancer 12:140. doi: 10.1186/1471-2407-12-140 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Fujiwara K, Ohuchida K, Sada M, et al. (2014) CD166/ALCAM expression is characteristic of tumorigenicity and invasive and migratory activities of pancreatic cancer cells. PLoS One 9:e107247. doi: 10.1371/journal.pone.0107247 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Donizy P, Zietek M, Halon A, et al. (2015) Prognostic significance of ALCAM (CD166/MEMD) expression in cutaneous melanoma patients. Diagn Pathol 10:86. doi: 10.1186/s13000-015-0331-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Clauditz TS, Rheinbaben von K, Lebok P, et al. (2014) Activated leukocyte cell adhesion molecule (ALCAM/CD166) expression in head and neck squamous cell carcinoma (HNSSC). Pathol Res Pract 210:649–655. doi: 10.1016/j.prp.2014.06.012 [DOI] [PubMed] [Google Scholar]
- 21.Ofori-Acquah SF, King JA (2008) Activated leukocyte cell adhesion molecule: a new paradox in cancer. Transl Res 151:122–128. doi: 10.1016/j.trsl.2007.09.006 [DOI] [PubMed] [Google Scholar]
- 22.van Kempen LC, Nelissen JM, Degen WG, et al. (2001) Molecular basis for the homophilic activated leukocyte cell adhesion molecule (ALCAM)-ALCAM interaction. J Biol Chem 276:25783–25790. doi: 10.1074/jbc.M011272200 [DOI] [PubMed] [Google Scholar]
- 23.Jeannet R, Cai Q, Liu H, et al. (2013) Alcam regulates long-term hematopoietic stem cell engraftment and self-renewal. Stem Cells 31:560–571. doi: 10.1002/stem.1309 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Manhas J, Bhattacharya A, Agrawal SK, et al. (2016) Characterization of cancer stem cells from different grades of human colorectal cancer. Tumour Biol 37:14069–14081. doi: 10.1007/s13277-016-5232-6 [DOI] [PubMed] [Google Scholar]
- 25.Wang F, Scoville D, He XC, et al. (2013) Isolation and characterization of intestinal stem cells based on surface marker combinations and colony-formation assay. Gastroenterology 145:383–95. 10.1053/j.gastro.2013.04.050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Smith NR, Davies PS, Levin TG, et al. (2017) Cell Adhesion Molecule CD166/ALCAM Functions Within the Crypt to Orchestrate Murine Intestinal Stem Cell Homeostasis. Cell Mol Gastroenterol Hepatol 3:389–409. doi: 10.1016/j.jcmgh.2016.12.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Cayrol R, Wosik K, Berard JL, et al. (2008) Activated leukocyte cell adhesion molecule promotes leukocyte trafficking into the central nervous system. Nat Immunol 9:137–145. doi: 10.1038/ni1551 [DOI] [PubMed] [Google Scholar]
- 28.Smith JR, Chipps TJ, Ilias H, et al. (2012) Expression and regulation of activated leukocyte cell adhesion molecule in human retinal vascular endothelial cells. Exp Eye Res 104:89–93. doi: 10.1016/j.exer.2012.08.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Karagogeos D, Pourquie C, Kyriakopoulou K, et al. (1997) Expression of the cell adhesion proteins BEN/SC1/DM-GRASP and TAG-1 defines early steps of axonogenesis in the human spinal cord. J Comp Neurol 379:415–427. doi: [DOI] [PubMed] [Google Scholar]
- 30.Bowen MA, Patel DD, Li X, et al. (1995) Cloning, mapping, and characterization of activated leukocyte-cell adhesion molecule (ALCAM), a CD6 ligand. J Exp Med 181:2213–2220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.King JA, Tan F, Mbeunkui F, et al. (2010) Mechanisms of transcriptional regulation and prognostic significance of activated leukocyte cell adhesion molecule in cancer. Mol Cancer 9:266. doi: 10.1186/1476-4598-9-266 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Tan F, Mbunkui F, Ofori-Acquah SF (2012) Cloning of the human activated leukocyte cell adhesion molecule promoter and identification of its tissue-independent transcriptional activation by Sp1. Cell Mol Biol Lett 17:571–585. doi: 10.2478/s11658-012-0028-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kavurma MM, Bobryshev Y, Khachigian LM (2002) Ets-1 positively regulates Fas ligand transcription via cooperative interactions with Sp1. J Biol Chem 277:36244–36252. [DOI] [PubMed] [Google Scholar]
- 34.Shirasaki F, Makhluf HA, LeRoy C, et al. (1999) Ets transcription factors cooperate with Sp1 to activate the human tenascin-C promoter. Oncogene 18:7755–7764. doi: 10.1038/sj.onc.1203360 [DOI] [PubMed] [Google Scholar]
- 35.Zhao Y, Zhang W, Guo Z, et al. (2013) Inhibition of the transcription factor Sp1 suppresses colon cancer stem cell growth and induces apoptosis in vitro and in nude mouse xenografts. Oncol Rep 30:1782–1792. doi: 10.3892/or.2013.2627 [DOI] [PubMed] [Google Scholar]
- 36.Wang J, Gu Z, Ni P, et al. (2011) NF-kappaB P50/P65 hetero-dimer mediates differential regulation of CD166/ALCAM expression via interaction with micoRNA-9 after serum deprivation, providing evidence for a novel negative auto-regulatory loop. Nucleic Acids Res 39:6440–6455. doi: 10.1093/nar/gkr302 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Nelissen JM, Peters IM, de Grooth BG, et al. (2000) Dynamic regulation of activated leukocyte cell adhesion molecule-mediated homotypic cell adhesion through the actin cytoskeleton. Mol Biol Cell 11:2057–2068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Gilsanz A, Sanchez-Martin L, Gutierrez-Lopez MD, et al. (2013) ALCAM/CD166 adhesive function is regulated by the tetraspanin CD9. Cell Mol Life Sci 70:475–493. doi: 10.1007/s00018-012-1132-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Weidle UH, Eggle D, Klostermann S, Swart GWM (2010) ALCAM/CD166: cancer-related issues. Cancer Genomics Proteomics 7:231–243. [PubMed] [Google Scholar]
- 40.Ochwat D, Hoja-Lukowicz D, Litynska A (2004) N-glycoproteins bearing beta1–6 branched oligosaccharides from the A375 human melanoma cell line analysed by tandem mass spectrometry. Melanoma Res 14:479–485. [DOI] [PubMed] [Google Scholar]
- 41.Yu M-J, Pisitkun T, Wang G, et al. (2006) LC-MS/MS analysis of apical and basolateral plasma membranes of rat renal collecting duct cells. Mol Cell Proteomics 5:2131–2145. doi: 10.1074/mcp.M600177-MCP200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kovalenko OV, Metcalf DG, DeGrado WF, Hemler ME (2005) Structural organization and interactions of transmembrane domains in tetraspanin proteins. BMC Struct Biol 5:11. doi: 10.1186/1472-6807-5-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Hemler ME (2003) Tetraspanin proteins mediate cellular penetration, invasion, and fusion events and define a novel type of membrane microdomain. Annu Rev Cell Dev Biol 19:397–422. doi: 10.1146/annurev.cellbio.19.111301.153609 [DOI] [PubMed] [Google Scholar]
- 44.Tudor C, Riet te J, Eich C, et al. (2014) Syntenin-1 and ezrin proteins link activated leukocyte cell adhesion molecule to the actin cytoskeleton. J Biol Chem 289:13445–13460. doi: 10.1074/jbc.M113.546754 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Xiao M, Yan M, Zhang J, et al. (2017) Cancer stem-like cell related protein CD166 degrades through E3 ubiquitin ligase CHIP in head and neck cancer. Exp Cell Res 353:46–53. doi: 10.1016/j.yexcr.2017.03.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Hebron KE, Li EY, Arnold Egloff SA, et al. (2018) Alternative splicing of ALCAM enables tunable regulation of cell-cell adhesion through differential proteolysis. Sci Rep 8:3208. doi: 10.1038/s41598-018-21467-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Rosso O, Piazza T, Bongarzone I, et al. (2007) The ALCAM shedding by the metalloprotease ADAM17/TACE is involved in motility of ovarian carcinoma cells. Mol Cancer Res 5:1246–1253. doi: 10.1158/1541-7786.MCR-07-0060 [DOI] [PubMed] [Google Scholar]
- 48.Hansen AG, Arnold SA, Jiang M, et al. (2014) ALCAM/CD166 is a TGF-beta-responsive marker and functional regulator of prostate cancer metastasis to bone. Cancer Res 74:1404–1415. doi: 10.1158/0008-5472.CAN-13-1296 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Hinkle CL, Sunnarborg SW, Loiselle D, et al. (2004) Selective roles for tumor necrosis factor alpha-converting enzyme/ADAM17 in the shedding of the epidermal growth factor receptor ligand family: the juxtamembrane stalk determines cleavage efficiency. J Biol Chem 279:24179–24188. [DOI] [PubMed] [Google Scholar]
- 50.Carbotti G, Orengo AM, Mezzanzanica D, et al. (2013) Activated leukocyte cell adhesion molecule soluble form: a potential biomarker of epithelial ovarian cancer is increased in type II tumors. Int J Cancer 132:2597–2605. doi: 10.1002/ijc.27948 [DOI] [PubMed] [Google Scholar]
- 51.Hansen AG, Freeman TJ, Arnold SA, et al. (2013) Elevated ALCAM shedding in colorectal cancer correlates with poor patient outcome. Cancer Res 73:2955–2964. doi: 10.1158/0008-5472.CAN-12-2052 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Tachezy M, Zander H, Marx AH, et al. (2012) ALCAM (CD166) expression and serum levels in pancreatic cancer. PLoS One 7:e39018. doi: 10.1371/journal.pone.0039018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Wierzbicki A, Gil M, Ciesielski M, et al. (2008) Immunization with a mimotope of GD2 ganglioside induces CD8+ T cells that recognize cell adhesion molecules on tumor cells. J Immunol 181:6644–6653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Soto MS, Serres S, Anthony DC, Sibson NR (2013) Functional role of endothelial adhesion molecules in the early stages of brain metastasis. Neuro Oncol 16:540–551. doi: 10.1093/neuonc/not222 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Bughani U, Saha A, Kuriakose A, et al. (2017) T cell activation and differentiation is modulated by a CD6 domain 1 antibody Itolizumab. PLoS One 12:e0180088. doi: 10.1371/journal.pone.0180088 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hassan NJ, Barclay AN, Brown MH (2004) Frontline: Optimal T cell activation requires the engagement of CD6 and CD166. Eur J Immunol 34:930–940. doi: 10.1002/eji.200424856 [DOI] [PubMed] [Google Scholar]
- 57.Zimmerman AW, Joosten B, Torensma R, et al. (2006) Long-term engagement of CD6 and ALCAM is essential for T-cell proliferation induced by dendritic cells. Blood 107:3212–3220. doi: 10.1182/blood-2005-09-3881 [DOI] [PubMed] [Google Scholar]
- 58.Oliveira MI, Goncalves CM, Pinto M, et al. (2012) CD6 attenuates early and late signaling events, setting thresholds for T-cell activation. Eur J Immunol 42:195–205. doi: 10.1002/eji.201040528 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Orta-Mascaro M, Consuegra-Fernandez M, Carreras E, et al. (2016) CD6 modulates thymocyte selection and peripheral T cell homeostasis. J Exp Med 213:1387–1397. doi: 10.1084/jem.20151785 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Patel DD, Wee SF, Whichard LP, et al. (1995) Identification and characterization of a 100-kD ligand for CD6 on human thymic epithelial cells. J Exp Med 181:1563–1568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Li Y, Singer NG, Whitbred J, et al. (2017) CD6 as a potential target for treating multiple sclerosis. Proc Natl Acad Sci U S A 114:2687–2692. doi: 10.1073/pnas.1615253114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Rodriguez PC, Torres-Moya R, Reyes G, et al. (2012) A clinical exploratory study with itolizumab, an anti-CD6 monoclonal antibody, in patients with rheumatoid arthritis. Results Immunol 2:204–211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Consuegra-Fernandez M, Julia M, Martinez-Florensa M, et al. (2018) Genetic and experimental evidence for the involvement of the CD6 lymphocyte receptor in psoriasis. Cell Mol Immunol 15:898–906. doi: 10.1038/cmi.2017.119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Cayrol R, Wosik K, Berard JL, et al. (2008) Activated leukocyte cell adhesion molecule promotes leukocyte trafficking into the central nervous system. Nat Immunol 9:137–145. doi: 10.1038/ni1551 [DOI] [PubMed] [Google Scholar]
- 65.Tormo BR, García CA, Chong A, Ochoa C, Faxas ME, Sagaró B, et al. (1994) Immunohistopatology of cutaneous T-cell lymphomas treated with topic ior t1 (anti CD6) monoclonal antibody Biotecnol Apl, 11 pp. 20–23 [Google Scholar]
- 66.Izquierdo-Cano L, Espinosa-Estrada E, Hernández-Padrón C, et al. (2014) Anticuerpo monoclonal humanizado itolizumab (anti-cd6) en síndromes linfoproliferativos cd 6+. Experiencia preliminar. [Humanized monoclonal antibody Itolizumab (anti-cd6) in patients with lymphoproliferative disorders cd6 +. Preliminary experience]. Revista Cubana de Hematología, Inmunología y Hemoterapia 30(3):0–0. Spanish. [Google Scholar]
- 67.Samaha H, Pignata A, Fousek K, et al. (2018) A homing system targets therapeutic T cells to brain cancer. Nature 561:331–337 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]