

SUMMARY
Citrus tristeza virus (CTV) (genus Closterovirus, family Closteroviridae) is the causal agent of devastating epidemics that changed the course of the citrus industry. Adapted to replicate in phloem cells of a few species within the family Rutaceae and to transmission by a few aphid species, CTV and citrus probably coevolved for centuries at the site of origin of citrus plants. CTV dispersal to other regions and its interaction with new scion varieties and rootstock combinations resulted in three distinct syndromes named tristeza, stem pitting and seedling yellows. The first, inciting decline of varieties propagated on sour orange, has forced the rebuilding of many citrus industries using tristeza‐tolerant rootstocks. The second, inducing stunting, stem pitting and low bearing of some varieties, causes economic losses in an increasing number of countries. The third is usually observed by biological indexing, but rarely in the field. CTV polar virions are composed of two capsid proteins and a single‐stranded, positive‐sense genomic RNA (gRNA) of ~20 kb, containing 12 open reading frames (ORFs) and two untranslated regions (UTRs). ORFs 1a and 1b, encoding proteins of the replicase complex, are directly translated from the gRNA, and together with the 5′ and 3′UTRs are the only regions required for RNA replication. The remaining ORFs, expressed via 3′‐coterminal subgenomic RNAs, encode proteins required for virion assembly and movement (p6, p65, p61, p27 and p25), asymmetrical accumulation of positive and negative strands during RNA replication (p23), or suppression of post‐transcriptional gene silencing (p25, p20 and p23), with the role of proteins p33, p18 and p13 as yet unknown. Analysis of genetic variation in CTV isolates revealed (1) conservation of genomes in distant geographical regions, with a limited repertoire of genotypes, (2) uneven distribution of variation along the gRNA, (3) frequent recombination events and (4) different selection pressures shaping CTV populations. Measures to control CTV damage include quarantine and budwood certification programmes, elimination of infected trees, use of tristeza‐tolerant rootstocks, or cross protection with mild isolates, depending on CTV incidence and on the virus strains and host varieties predominant in each region. Incorporating resistance genes into commercial varieties by conventional breeding is presently unfeasible, whereas incorporation of pathogen‐derived resistance by plant transformation has yielded variable results, indicating that the CTV–citrus interaction may be more specific and complex than initially thought. A deep understanding of the interactions between viral proteins and host and vector factors will be necessary to develop reliable and sound control measures.
CTV–CITRUS PLANTS: FROM PACIFIC COEXISTENCE TO AN OPEN WAR?
Citrus tristeza virus (CTV), a member of genus Closterovirus, family Closteroviridae, is the causal agent of various diseases with dramatic effects on citrus crops. CTV dispersal to new regions occurs primarily via movement of infected plants or propagation of infected buds and then, at a local level, it can be spread by several aphid species (Bar‐Joseph et al., 1989; Duran‐Vila and Moreno, 2000; Timmer et al., 2000). Seed transmission has not been observed (McClean, 1957). In nature the ecological niche of this virus is restricted to phloem cells of some species of two genera within the family Rutaceae. Such a specific niche and the observation that most citrus species grown on their own roots are symptomless hosts for many CTV isolates suggest that CTV probably appeared many years ago in the areas of origin of citrus (South East Asia and the Malayan archipelago) and then coevolved with citrus hosts. For centuries citrus were grown from seeds and graft propagation became widely used only after commercial production was developed. Therefore, as seed transmission does not occur, we have to assume an early adaptation of CTV to aphid transmission as the only way to invade new hosts and survive, or that it evolved from an aphid‐transmissible ancestor.
The first movements of citrus from their site of origin to other regions were with fruits or seeds, thus making it unlikely that CTV was dispersed at that time (Zaragoza, 2007). More probably this occurred at the end of the 19th century, when important improvements in maritime transport enabled the movement of complete citrus plants and soil in terrariums. As these advances coincided with an increased botanical and commercial interest in citrus, thousands of plants were moved from Asia to other regions and numerous exotic citrus species were exchanged between collections (Roistacher, 1981). This probably represented the first large dispersal of CTV from its site of origin to new countries, where the virus interacted with new host varieties and combinations, under different climatic and environmental conditions. Coincidently, a significant change in citrus cultivation induced by a different pathogen also occurred in the 19th century. In 1836 a foot rot epidemic caused by oomycetes of the genus Phytophthora started in the Azores and later affected Mediterranean countries, decimating citrus plantings established from seeds or bud propagated on sweet orange [Citrus sinensis (L.) Osb.] seedlings or on rooted citron (C. medica L.) cuttings (Zaragoza, 2007). This disaster forced bud propagation of citrus varieties on sour orange (C. aurantium L.), a foot‐rot‐resistant rootstock, highly adaptable to all soil types, that induces good bearing and excellent fruit quality. Sour orange soon became almost the exclusive rootstock in the Mediterranean area and then in America.
The use of sour orange rootstock was critical for the development of commercial citriculture in many countries in the 20th century, but unbeknown to growers, it also created the conditions for a new disease epidemic. The new disease, called tristeza, was the direct result of the interaction of CTV with a sensitive new host created commercially after propagation of sweet orange, mandarin (C. reticulata Blanco) and grapefruit (C. paradisi Macf.) varieties on sour orange rootstock. The outcome of this interaction was dramatic for the citrus industry and included the loss of almost 100 million trees propagated on sour orange, and the need for tristeza‐tolerant rootstocks to rebuild a new citrus industry in affected countries. The most destructive epidemics of tristeza occurred in Argentina (1930), Brazil (1937), California (1939), Florida (1951), Spain (1957), Israel (1970) and Venezuela (1980), but important outbreaks have also been reported from Cyprus (1989), Cuba (1992), Mexico (1995), the Dominican Republic (1996) and Italy (2002) (Bar‐Joseph et al., 1989; Duran‐Vila and Moreno, 2000; Davino et al., 2003; Garnsey et al., 2000; Gottwald et al., 1998, 2002; Kyriakou et al., 1996; Timmer et al., 2000). In addition to tree losses, indirect damage associated with CTV epidemics are (1) the loss of sour orange rootstock, whose agronomic qualities are unmatched by any other rootstock; and (2) the appearance of new problems associated with the use of tristeza‐tolerant rootstocks, including damage from soil salinity or alkalinity, waterlogging in heavy soils, or sensitivity to soil fungi or graft‐transmissible pathogens. Examples of graft‐transmissible diseases associated with the use of these rootstocks are citrus blight, citrus sudden death or citrus tatter leaf (Duran‐Vila and Moreno, 2000; Román et al., 2004; Timmer et al., 2000). On the positive side, CTV epidemics and problems found with the new rootstocks forced intensive research on citrus pathology that led to (1) the discovery of new pathogens such as viroids, (2) the confirmation that most old‐clone citrus trees were symptomless carriers of these and other infectious agents, and (3) the setting up of citrus sanitation, quarantine and budwood certification procedures that have enabled development of a modern citrus industry in many countries (Duran‐Vila and Moreno, 2000; Navarro et al., 2002; Timmer et al., 2000).
In countries such as South Africa and Australia, early propagations on sour orange were unsuccessful and this rootstock never became widely used. At the time this was attributed to poor compatibility between sweet and sour orange, but it is now generally accepted that CTV was already present in these countries and the symptoms observed were probably due to tristeza disease (Webber, 1943). Early failures of trees on sour orange rootstock precluded its further use and avoided tristeza epidemics, but a different pathogenic interaction between CTV and citrus, called stem pitting (SP), was observed in those areas and later in different countries that had replanted citrus on tristeza‐tolerant rootstocks after previous tristeza epidemics (Hughes and Lister, 1949; McClean, 1956; Müller et al., 1968; Rocha‐Peña et al., 1995; Roistacher and Moreno, 1991; Wallace, 1978). Affected trees showed stunting, SP, low yield and poor fruit quality, regardless of the rootstock used.
CTV variants causing SP, initially restricted to regions of Asia, Australia, South Africa and South America, have been successively reported from other citrus areas, including California, Florida and the Mediterranean region (Ben Ze’ev et al., 1989; Kyriakou et al., 1996; Ruiz‐Ruiz et al., 2006; Z.‐N. Yang et al., 1999), albeit at lower frequency. In addition, other observations suggest that these variants often show higher fitness than mild variants and become predominant in co‐inoculated plants (Powell et al., 2003; Roistacher and Dodds, 1993; Sambade et al., 2007). Are these observations indicative of increasing CTV virulence? If so, is this trend due to population changes induced by constraints derived from the interaction of CTV with new citrus varieties and environmental conditions? If the answers to these questions are affirmative, it is tempting to speculate that CTV dispersal from its site of origin to new regions and varieties caused the CTV–citrus pathosystem to shift from pacific coexistence to an open war.
CTV‐INDUCED DISEASES AND HOST SPECIES AFFECTED
Natural CTV hosts essentially include species of the genera Citrus and Fortunella, but virus infection has been detected in experimentally inoculated citrus relatives of the genera Aegle, Aeglopsis, Afraegle, Atalantia, Citropsis, Clausena, Eremocitrus, Hesperthusa, Merrillia, Microcitrus, Pamburus, Pleiospermium and Swinglea (Duran‐Vila and Moreno, 2000; Timmer et al., 2000; Yoshida, 1996). Experimental infection was also achieved in the non‐citrus species Passiflora gracilis and Passiflora coerulea using aphid vectors (Price, 1966; Roistacher et al., 1988), and in Nicotiana benthamiana protoplasts or agroinfiltrated leaves (Gowda et al., 2005; Navas‐Castillo et al., 1997; our unpublished results). Resistance to most CTV isolates has been observed in Poncirus trifoliata (L.) Raf. (Mestre et al., 1997a; Rai, 2006) and resistance to specific CTV strains has been reported in Meiwa kumquat (Fortunella crassifolia Swing.) (Mestre et al., 1997b), some pummelos [C. grandis (L.) Osb.] (Fang and Roose, 1999; Garnsey et al., 1996) and sour orange (Asíns et al., 2004; our unpublished results). However, CTV replication has been observed in P. trifoliata, pummelo or sour orange protoplasts, suggesting that only CTV movement is impaired in these species (Albiach‐Martíet al., 2004; our unpublished data).
Depending on virus strains and on the species or scion–rootstock combinations, CTV may cause three distinct syndromes named tristeza, SP and seedling yellows (SY). Tristeza disease is a decline syndrome caused by CTV infection of different citrus species [sweet oranges, mandarins, grapefruits (Citrus paradisi Macf.), kumquats or limes (Citrus aurantifolia (Christm.) Swing.)] propagated on rootstock species such as sour orange or lemon [C. limon (L.) Burn. f.]. Its most dramatic expression is quick decline, a syndrome in which a tree with normal appearance starts showing wilt symptoms and completely collapses in a few weeks (Fig. 1A). Commonly, affected trees show dull green or yellow thin foliage, leaf shedding and twig dieback, small chlorotic leaves resembling the effects of nitrogen deficiency, and small pale‐coloured fruits that are unmarketable (Fig. 1B). CTV induces obliteration, collapse and necrosis of sieve tubes and companion cells close to the bud union, producing an excessive amount of non‐functional phloem (Schneider, 1959). This causes progressive reduction of the root system with deficient supply of water and minerals, which results in wilting, chlorosis and dieback symptoms. As this specific interaction does not occur with many other citrus species, the tristeza syndrome can be avoided using decline‐tolerant species as rootstocks.
Figure 1.

Tristeza decline, stem pitting and seedling yellows symptoms induced by Citrus tristeza virus (CTV) in different varieties and scion/rootstock combinations. (A) Quick decline syndrome in a sweet orange tree propagated on sour orange rootstock. (B) Different stages of decline of three trees of the same scion/rootstock combination, in comparison with non‐declined neighbour trees (dark green colour). (C) Severe stem pitting symptoms in the trunk of a grapefruit on Poncirus trifoliata rootstock (this grapefruit was from one of the first imports from South Africa into Argentina; Museum of the INTA experiment station at Concordia, Entre Rìos, Argentina). (D) Small‐sized fruits from a grapefruit tree on Poncirus trifoliata rootstock severely affected by stem pitting, in comparison with a normal grapefruit (left). (E) Stunted growth and small yellow leaves in a sour orange seedling inoculated with a seedling yellows isolate of CTV (left), in comparison with a similar plant inoculated with a mild non‐seedling yellows isolate (right). (F) Vein clearing in a young leaf of Citrus macrophylla (upper) and vein corking in a leaf of Mexican lime (lower), inoculated with a mild and a severe CTV isolate, respectively. (G) Stem pitting in the trunk of CTV‐infected Mexican lime seedlings grown in the greenhouse. (H) Transgenic Mexican lime leaf expressing p23 (left) and a similar leaf from a CTV‐infected non‐transgenic lime CTV (right).
SP disease is probably initiated by interruption of meristematic activity at limited areas of the cambium that results in irregular radial growth with local depression at the inactivated points (Schneider, 1959). Extensive pitting may limit radial growth and produce stunting, thin foliage with small yellow leaves, low bearing and small fruits with low juice content that are unmarketable (Fig. 1C,D). Citrus cultivars sensitive to SP are affected regardless of whether they are a seedling, or used in a grafted combination as a rootstock or a scion. Acid limes show the highest sensitivity, grapefruits and some sweet orange varieties intermediate sensitivity, and mandarins the highest tolerance (Duran‐Vila and Moreno, 2000; Timmer et al., 2000). Contrasting with tristeza, the SP syndrome usually does not cause tree death, but unthrifty growth and chronic yield reductions also cause high cumulative economic losses. Moreover, areas invaded by SP isolates may suffer permanent limitations to production by sensitive varieties.
The third CTV‐induced syndrome, SY, is characterized by stunting, production of small pale or yellow leaves, a reduced root system and sometimes a complete cessation of growth of sour orange, grapefruit or lemon seedlings (Fig. 1E) (Fraser, 1952; McClean, 1960). Sometimes plants showing SY recover and produce a new flush with normal leaves. Moreover, field isolates from grapefruit or lemon trees usually do not induce SY. These and other observations led to the interpretation that SY was caused by a component associated with CTV that eventually could be ‘filtered off’ by sour orange, grapefruit or lemon plants (McClean, 1963; Wallace and Drake, 1972). However, the finding that a clonal CTV population obtained from an infectious cDNA clone still produced SY similar to the original isolate indicated that this syndrome is induced by specific pathogenicity determinants in the CTV genome (Satyanarayana et al., 1999, 2001).
TRANSMISSION AND EPIDEMIOLOGY
CTV dispersal occurs by propagation of virus‐infected buds and by vector transmission. The first is responsible for most CTV introductions into new areas, whereas the second is important for local spread. The virus has been also experimentally transmitted to healthy plants by dodder (Cuscuta subinclusa) (Weathers and Hartung, 1964) and by stem‐slash inoculation with partially purified extracts (Garnsey et al., 1977), but these procedures are epidemiologically unimportant.
CTV is vectored by several aphid species in a semipersistent mode with variable efficiency. The most efficient vector species, Toxoptera citricida (Kirkaldy), enables experimental CTV transmission using single aphids (Brlansky et al., 2003). T. citricida is well established in Asia, Australia, sub‐Saharan Africa, Central and South America, and different Caribbean countries (Costa and Grant, 1951; Halbert et al., 2004; McClean, 1975; Rocha‐Peña et al., 1995; Yokomi et al., 1994). Recently, it has been detected in isolated citrus trees in northern Spain and Portugal (Ilharco et al., 2005), far from the important citrus‐producing areas. Aphis gossypii (Glover), the second most efficient vector, is the main CTV vector in the Mediterranean basin and areas of North America (Cambra et al., 2000; Dickson et al., 1956; Hermoso de Mendoza et al., 1984; Marroquín et al., 2004; Raccah et al., 1976; Yokomi and Garnsey, 1987). Comparative transmission experiments of several CTV isolates with these two vector species showed that T. citricida can transmit CTV 6–25 times more efficiently than A. gossypii (Yokomi et al., 1994). Other aphid species such as A. spiraecola (Patch) (formerly A. citricola van der Goot) and T. aurantii (Boyer de Fonscolombe) have been reported as being less efficient CTV vectors than A. gossypii under experimental conditions (Hermoso de Mendoza et al., 1984; Norman and Grant, 1956; Yokomi and Garnsey, 1987); however, in some citrus areas A. spiraecola builds up larger populations than A. gossypii and its role in CTV dispersal could be important (Hermoso de Mendoza et al., 1984). Myzus persicae (Sulzer), A. craccivora (Koch) and Uroleucon jaceae (Linnaeus) have been reported as CTV vectors only in India (Varma et al., 1965). Viruliferous aphids can transmit CTV for at least 24 h but infectivity is lost within 48 h after acquisition (Raccah et al., 1976). The ratio of aphids carrying CTV in the field, as detected by nested RT‐PCR amplification of CTV RNA from individual aphids, ranged from 19 to 27% (Marroquín et al., 2004).
A number of factors influence the rate of CTV spread in a given area, including composition of the aphid fauna (Cambra et al., 2000; Gottwald et al., 1996b), aphid population density (Dickson et al., 1956), environmental conditions favouring new flush and aphid population build up (moderate temperatures, irrigation and fertilization) (Bar‐Joseph and Loebenstein, 1973), susceptibility of the predominant citrus varieties (Hermoso de Mendoza et al., 1988b; Marroquín et al., 2004; Roistacher and Bar‐Joseph, 1984) and transmissibility of the predominant CTV isolates (Bar‐Joseph and Loebenstein, 1973; Hermoso de Mendoza et al., 1984, 1988a; Yokomi and Garnsey, 1987). In this regard, it is worth mentioning that in many countries a lag period (sometimes more than 30 years) was observed between CTV introduction and noticeable field spread, suggesting a need for CTV isolates to become adapted to local aphid vector populations (Bar‐Joseph et al., 1989; Raccah et al., 1980; Roistacher et al., 1980). Failure to eradicate early CTV‐infected trees allowed further epidemics, a situation that could be repeated with other exotic pathogens.
The spatial and temporal spread of CTV has been studied in several countries using different tools for analysis (Gottwald et al., 1996a, 1998, 1999; Hughes and Gottwald, 1998). These studies showed that the spread patterns depended on whether T. citricida or A. gossypii was the predominant vector species. In locations where A. gossypii was predominant, CTV incidence increased from 5 to 95% infected trees in 8–15 years following a stair‐step line, infected trees showed limited aggregation and new infections were not closely associated with existing infected trees, but rather with trees several tree spaces distant. Contrastingly, in areas where T. citricida was predominant, the same disease increment often occurred in only 2–4 years with a rapid and essentially continuous increase, and aggregates of infected trees were common because spread to trees immediately adjacent to existing infections was frequent. The biology and feeding habits of both vector species might be the cause of these distinct spread patterns (Gottwald et al., 1996b).
CTV STRUCTURE, GENOME ORGANIZATION AND EXPRESSION
CTV was the first closterovirus to be biophysically characterized. Its virions are flexuous filaments of about 2000 × 11 nm, with about 6% RNA content, that are helically constructed with a basic pitch of about 3.7 nm (Bar‐Joseph et al., 1972). As with other members of the family Closteroviridae, CTV virions are bipolar and contain two coat proteins of 25 kDa (CP) and 27 kDa (CPm) that encapsidate about 97 and 3% of the virion length, respectively (Febres et al., 1996; Satyanarayana et al., 2004). The ~19.3‐kb genomic RNA (gRNA) of CTV is single‐stranded and positive sense, and its complete nucleotide sequence has been determined in at least nine distinct isolates (Albiach‐Martíet al., 2000c; Karasev et al., 1995; Mawassi et al., 1996; Pappu et al., 1994; Ruiz‐Ruiz et al., 2006; Suastika et al., 2001; Vives et al., 1999, 2005; Z.‐N. Yang et al., 1999; GenBank accession numbers AY340974 and DQ272579). The CTV gRNA is organized into 12 open reading frames (ORFs), potentially encoding at least 17 protein products, and two untranslated regions (UTRs) of about 107 and 273 nt at the 5′ and 3′ termini, respectively (Fig. 2A). ORFs 1a and 1b encode proteins of the replicase complex, whereas ORFs 2–11, spanning the 3′ terminal half of the gRNA, encode proteins p33, p6, p65, p61, p27, p25, p18, p13, p20 and p23. The genome regions important for replication and virion assembly have been mapped by engineering a cDNA clone of the complete CTV gRNA. RNA transcripts from this clone were infectious in N. benthamiana protoplasts, producing normal virions that induced in citrus plants the symptoms characteristic of the wild isolate (Satyanarayana et al., 1999, 2001).
Figure 2.
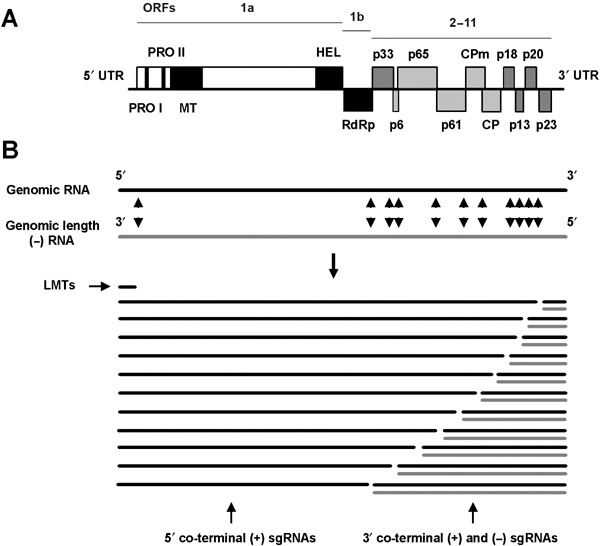
Organization and expression of the Citrus tristeza virus (CTV) genomic RNA. (A) Outline of the CTV genome. Boxes indicate ORFs, and proteins encoded by different ORFs are indicated. In ORFs 1a and 1b: PRO‐I and PRO‐II are papain‐like proteases; MT, methyl transferase; HEL, helicase; and Rdrp, RNA‐dependent RNA polymerase domains. CPm and CP are the minor and major coat proteins, and UTR indicates untranslated region. (B) Main RNA species produced for replication and expression of the genome. Black lines indicate positive stranded, and grey lines negative stranded, genomic or subgenomic (sg) RNAs. LMTs are low‐molecular‐weight tristeza sgRNAs. Arrowheads mark aproximate positions of the controller elements inducing formation of sgRNAs.
Sequences of the 5′‐UTR have been unequivocally classified into three types (I, II and III), but their predicted secondary structure of minimum energy always consisted of two stem‐loops separated by a short spacer region (López et al., 1998). Mutations disrupting secondary structure abolished replication whereas compensatory mutations allowed replication to resume, suggesting that the predicted secondary structure is more important for replication than primary structure. However, some sequences of the 5′‐UTR were necessary for both replication and virion assembly (Gowda et al., 2003b; Satyanarayana et al., 2004). The 3′‐UTR, a highly conserved region (López et al., 1998), is critical for recognition by the replicase complex. The secondary structure of minimum energy predicted for this region comprises a series of ten stem‐loops with variable ability to support mutations affecting the primary or the secondary structure. The core of the 3′ replication signal might be located in three of the central stem‐loops, as all mutations introduced in this region completely prevented replication (Satyanarayana et al., 2002a). A CCA triplet at the 3′ terminus seems necessary for efficient replication (López et al., 1998; Satyanarayana et al., 2002a).
The coding region of the gRNA comprises the replication module (ORFs 1a and 1b), a five‐gene module (p6, p65, p61, p27 and p25) encoding proteins involved in virion assembly and transport, which is conserved in all members of the family Closteroviridae, the p20 gene, a homologue of the p21 gene of Beet yellow virus (BYV), and four genes encoding proteins with no homologue in other closteroviruses (p33, p18, p13 and p23) (Dolja et al., 2006). ORF 1a encodes a ~349‐kDa polyprotein with two papain‐like protease domains, a type I methyltransferase‐like domain, and a helicase‐like domain bearing the motifs of the superfamily I helycase domain. ORF 1b, via a frameshift, extends the 1a proteins by 54 kDa, which contain the RNA‐dependent RNA polymerase (RdRp) domains (Karasev et al., 1995) (Fig. 2A). Proteins encoded by the five‐gene module include a transmembrane protein (p6), a homologue of the HSP70 plant heat‐shock proteins (p65), two diverged copies of the capsid protein (p25 and p27) (Febres et al., 1996; Pappu et al., 1994), and an additional protein (p61) also regarded as a diverged CP copy (Dolja et al., 2006). The p6 homologue in BYV has been shown to be a movement protein (Peremyslov et al., 2004). The coordinate action of p65 and p61, in addition to the CP and CPm coat proteins, is required for proper virion assembly (Satyanarayana et al., 2000). In normal CTV virions, CP coats most of the gRNA and CPm only ~630 nt at the 5′‐terminus, but in the absence of p65 or p61, CPm may coat larger segments or even the complete gRNA. These two proteins probably bind to the transition zone between CP and CPm and restrict CPm to the virion tail (Satyanarayana et al., 2004). In other closteroviruses the homologues of p65 and p61 are coordinately assembled with CPm in the virion structure and remain attached to the virions (Alzhanova et al., 2007; Tian et al., 1999). The p20 protein is a major component of CTV‐induced amorphous inclusion bodies (Gowda et al., 2000), and p23, an RNA binding protein with a Zn finger domain (López et al., 2000), regulates asymmetrical accumulation of the positive and negative strands during RNA replication (Satyanarayana et al., 2002b). Both p20 and p23, in addition to the CP, act as RNA silencing suppressors in N. benthamiana and N. tabacum plants, with p23 inhibiting intercellular silencing, CP intracellular silencing, and p20 both inter‐ and intracellular silencing (Lu et al., 2004). Deletion mutants lacking genes p33, p18 and p13 were capable of replication and assembly (Satyanarayana et al., 1999, 2000), indicating that they are not required for these functions. Their role in CTV biology remains unknown. By contrast, a construct carrying the green fluorescent protein (GFP) gene as an extra gene controlled either by a duplicated controller element (CE) of the CTV CP, or by a heterologous CE from BYV, stably replicated and moved in citrus plants and expressed GFP, suggesting that CTV can be reliably used as a virus vector for citrus (Folimonov et al., 2007).
Expression of the CTV genome includes at least three mechanisms widely used by positive‐stranded RNA viruses: proteolytic processing, ribosomal frameshifting and formation of a nested set of 3′‐coterminal subgenomic (sg) RNAs (Ayllón et al., 2003, 2004; Gowda et al., 2001, 2003a, 2003b; Hilf et al., 1995; Karasev et al., 1995, 1997; Satyanarayana et al., 2002a,b). Replication of the CTV gRNA involves synthesis of negative strands that serve as template for the generation of new positive strands, although the latter accumulate 10–20 times more than the negative strands (Satyanarayana et al., 2002b). ORFs 1a and 1b are directly translated from the gRNA. The first produces a ~349‐kDa polyprotein, whereas translation of ORF 1b occurs by occasional +1 ribosomal frameshifting to yield a ~400‐kDa polyprotein that is later proteolytically processed (Karasev et al., 1995). The ten genes in the 3′ half of the CTV gRNA are expressed by the synthesis of 3′ co‐terminal sgRNAs that act as messenger RNAs from which the 5′ proximal ORF is translated (Hilf et al., 1995). A similar set of negative‐stranded sgRNAs is also produced in infected cells, but these accumulate about 40–50 times less than positive‐stranded sgRNAs (Satyanarayana et al., 2002b). Expression of the ten 3′ proximal ORFs is regulated independently both in amount and in timing (Hilf et al., 1995; Navas‐Castillo et al., 1997) by individual CEs that might serve for internal promotion from the minus strand gRNA template, or for the synthesis of negative‐stranded sgRNAs by premature termination of some negative‐stranded gRNAs (Ayllón et al., 2003; Gowda et al., 2001). A set of less abundant 5′‐coterminal positive‐stranded sgRNAs is also generated, probably by premature termination of the gRNA at the CEs (Gowda et al., 2001). Finally, two abundant positive‐stranded 5′‐coterminal sgRNAs of about 800 nt (LMT1 and LMT2) are produced by premature termination of the gRNA synthesis at a completely different CE, thus totalling more than 30 sgRNAs in infected cells (Ayllón et al., 2004; Che et al., 2001; Mawassi et al., 1995b) (Fig. 2B). The viral function of this novel CE and of the two LMTs remains unknown (Gowda et al., 2003b).
Frequently, CTV‐infected tissues also accumulate large amounts of positive‐ and negative‐stranded defective RNAs (D‐RNAs) that contain the 3′ and 5′ termini of the gRNA but lack variable portions of the central region. These viral RNAs, which are easily observed by electrophoretic analysis of plant extracts enriched in double‐stranded RNA (dsRNA), are very common in field isolates (Dodds et al., 1987; Guerri et al., 1991; Mawassi et al., 1995a,b; Moreno et al., 1990). Most D‐RNAs are 2.0–5.0 kb in size, but large D‐RNAs comprising ORFs 1a and 1b in their 5′ proximal moiety, or ORFs 2–11 in their 3′ terminal moiety, have been reported (Che et al., 2002, 2003). Those large D‐RNAs resemble RNAs 1 and 2 of criniviruses (family Closteroviridae). Sequence analysis of the junction site and flanking regions suggest that most D‐RNAs must be generated by a template‐switching mechanism induced by different factors (Ayllón et al., 1999a; Yang et al., 1997). The minimal replication signals required for D‐RNA replication in trans are located in the 5′ proximal 1 kb and at the 3′‐UTR of the D‐RNA sequence (Mawassi et al., 2000a,b). Although the biological role of D‐RNAs is presently unknown, their wide occurrence in CTV isolates suggests that they may provide some advantage. At least in one case, the presence of a D‐RNA was reported to modulate CTV symptom expression (G. Yang et al., 1999).
The interaction of CTV with host factors is largely unknown, but analysis of the Mexican lime transcriptome using a citrus microarray showed altered expression of 334 genes after infection with a severe CTV isolate, about half of them without significant similarity to other known proteins (Gandía et al., 2007).
CTV DIAGNOSIS, STRAIN CHARACTERIZATION AND GENETIC VARIATION
Diagnosis of CTV infection was performed for years by biological indexing on sensitive indicator hosts, mainly Mexican lime or C. macrophylla seedlings that upon CTV infection show vein clearing in young leaves, leaf cupping, short internodes and stem pitting (Fig. 1F,G) (Roistacher, 1991). After CTV purification different antisera and monoclonal antibodies to the coat protein were obtained that allowed routine CTV detection by immunoenzymatic procedures (Bar‐Joseph et al., 1979; Cambra et al., 1991; Garnsey et al., 1993; Gonsalves et al., 1978; Nikolaeva et al., 1998; Permar et al., 1990; Vela et al., 1986). Availability of ELISA detection was a critical tool to expand research in areas such as CTV epidemiology, virus movement or isolate characterization, and to improve efficiency of eradication programmes (Bar‐Joseph et al., 1989; Ben Ze’ev et al., 1989; Gottwald et al., 1996a,1996b, 1998, 2002; Kyriakou et al., 1996). ELISA detection has probably been used with CTV more than with any other plant virus. Finally, after the complete nucleotide sequence of the CTV gRNA was available, a variety of diagnostic procedures based on specific detection of viral RNA were developed, including molecular hybridization with cDNA or cRNA probes (Barbarossa and Savino, 2006; Narváez et al., 2000; Rosner and Bar‐Joseph, 1984) and several RT‐PCR amplification‐based methods (Nolasco et al., 1993; Olmos et al., 1999). Real‐time RT‐PCR protocols have greatly improved sensitivity of detection and allowed quantification of genomic RNA copies in infected citrus tissues or in viruliferous aphids (Bertolini et al., 2007; Ruiz‐Ruiz et al., 2007; Saponari et al., 2007).
Variations in biological characteristics of CTV isolates, including the type and intensity of symptoms induced or aphid transmissibility, have been observed since the first epidemics. Indeed the presence of several components associated with the different types of symptoms (McClean, 1963) was a working hypothesis until CTV was characterized and shown to be the only causal agent of those symptoms (Garnsey et al., 1977; Satyanarayana et al., 1999, 2001). Evidence for the presence of different variants in CTV isolates, which could be separated after aphid transmission or host change (Hermoso de Mendoza et al., 1988a; Grant and Higgins, 1957; Raccah et al., 1980), was also obtained before the concept of RNA viruses as a population of genetic variants was established (Domingo and Holland, 1994). However, the genetic basis for this biological variability is still largely unknown.
Comparison of CTV isolates to provide a pathogenicity profile has been and still is done by indexing on different indicator plants to define biogroups (Ballester‐Olmos et al., 1993; Garnsey et al., 1991). After purification and molecular characterization of CTV, rapid procedures were devised to study CTV variation, to identify specific isolates or to try to associate pathogenicity characteristics with different molecular markers (Moreno and Guerri, 1997). Some of these procedures detected differences in the CP using a variety of monoclonal and polyclonal antibodies (Nikolaeva et al., 1998; Permar et al., 1990; Vela et al., 1986) or by analysis of peptide maps generated with endoproteases (Albiach‐Martíet al., 2000a). The most widely used monoclonal antibody for isolate discrimination is MCA13 (Permar et al., 1990), which recognizes an epitope largely conserved in severe isolates in Florida, but lacking in mild isolates (Pappu et al., 1993). Other procedures detected variations in viral RNA, including: (1) differences in dsRNA pattern between isolates (Dodds et al., 1987; Guerri et al., 1991; Moreno et al., 1990, 1993), later shown to be due to the presence of D‐RNAs in CTV isolates; (2) hybridization pattern with cDNA or cRNA probes of several gRNA regions (Albiach‐Martíet al., 2000b; Narváez et al., 2000; Rosner and Bar‐Joseph, 1984; Rosner et al., 1986); (3) restriction fragment length polymorphism (RFLP) analysis of the CP gene (Gillings et al., 1993; Roy et al., 2003); (4) RT‐PCR amplification patterns with primer sets specific for several CTV genotypes (Hilf et al., 1999, 2005), for 5′‐UTR sequence types I, II and III (Ayllón et al., 2001; Ruiz‐Ruiz et al., 2006), or for three groups of isolates differing by their p23 sequence (Sambade et al., 2003); and (5) single‐strand conformation polymorphism (SSCP) analysis of different gRNA regions (Ayllón et al., 1999b; d’Urso et al., 2000, 2003; Rubio et al., 1996; Sambade et al., 2002, 2007). This last technique has been used to characterize the population structure of CTV isolates and select specific variants for sequencing, thus allowing estimates of the genetic diversity within and between isolates (Ayllón et al., 2006; Rubio et al., 2001; Sambade et al., 2003). The values obtained were in the range of those calculated for other plant viruses (García‐Arenal et al., 2001), but higher than those estimated for other members of the family Closteroviridae (Rubio et al., 1999, 2001).
Phylogenetic analysis of the complete sequences reported for nine CTV isolates revealed three main clusters that included (1) the severe SP isolates T318A from Spain (Ruiz‐Ruiz et al., 2006), SY568R from California (Vives et al., 2005; Z.‐N. Yang et al., 1999), NuagA from Japan (Suastika et al., 2001) and VT from Israel (Mawassi et al., 1996); (2) the mild isolates T30 from Florida (Albiach‐Martíet al., 2000c) and T385 from Spain (Vives et al., 1999); and (3) isolates T36 from Florida (Karasev et al., 1995; Pappu et al., 1994), Qaha from Egypt (AY340974) and a Mexican isolate (DQ272579) (Fig. 3). Within‐group nucleotide identities were over 97.5%, whereas the lowest identity (75.6%) was between VT and Qaha.
Figure 3.
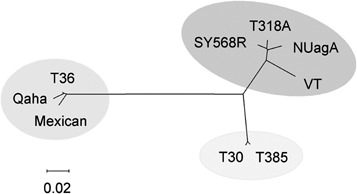
Neighbour‐joining tree calculated with the complete nucleotide sequence of the genomic RNA from nine CTV isolates, using 1000 bootstrap replicates. Branch length indicates genetic distance. The three clusters obtained include severe isolates inducing seedling yellows (SY) and stem pitting in sweet orange and/or grapefruit (SP) (T318A, SY568R, NuagA and VT), mild non‐SY, non‐SP isolates (T30 and T385) and a group of isolates with intermediate characteristics (T36, Qaha and Mexican).
Genetic variation is unevenly distributed along the CTV gRNA, the most conserved region being the 3′‐UTR, with over 95% identity between isolates, and the most variable being the 5′‐UTR, with identity values as low as 44–45% between sequence types I and III (López et al., 1998). Uneven distribution of variation is also observed in coding regions and even within genes, probably reflecting different selective pressures along the gRNA. For example, genetic diversity of a group of Californian and Spanish isolates was 0.038 for the CP gene, but 0.137 for an ORF 1a region (Rubio et al., 2001). Overall, sequence comparisons between CTV isolates of different geographical origin and pathogenicity characteristics show (1) a high degree of conservation between CTV genomes separated in time and space, with a limited repertoire of genotypes (Albiach‐Martíet al., 2000c; Ruiz‐Ruiz et al., 2006; Vives et al., 1999; our unpublished data), and (2) a population structure variable between isolates, with some consisting of a predominant sequence and some closely related variants, and others having a complex structure with highly divergent sequence variants (Ayllón et al., 2006; Vives et al., 2005).
Factors shaping CTV populations in the field include mutation, recombination events between diverged sequence variants, selection, genetic drift and gene flow due to repeated inoculation of field trees and movement of infected buds between regions. Mutation due the error‐prone nature of RNA‐dependent RNA polymerases is the basic mechanism generating diversity (Domingo and Holland, 1994). The contribution of mutants to CTV variation was assessed using a clonal CTV isolate derived from an infectious cDNA clone (Satyanarayana et al., 1999, 2001) serially propagated in plants of four different citrus species. Analysis of the resulting populations showed high genetic stability in some hosts, as reported for other plant viruses (García‐Arenal et al., 2001), but rapid diversification in others, suggesting that this second host type might have contributed to CTV variation after the virus was moved from its areas of origin and came into contact with new host varieties (P. Moya et al., unpublished results).
Homologous and non‐homologous recombination might be a fast means for CTV evolution. Non‐homologous recombination giving rise to multiple D‐RNAs have been widely documented (Ayllón et al., 1999a; Guerri et al., 1991; Mawassi et al., 1995a,b; Yang et al., 1997). Homologous recombination was first observed when comparing the gRNA sequence of isolates T36 or SY568 with that of other isolates (Mawassi et al., 1996; Vives et al., 1999, 2005) and later confirmed by additional sequence comparisons. Incongruent phylogenetic relationships between CTV isolates in different gRNA regions indicated that homologous recombination between diverged sequence variants may be a frequent phenomenon in CTV isolates (Sambade et al., 2003; our unpublished data). Recombination could repair in some cases lethal or biologically disadvantageous mutations, providing opportunities for rapid evolution.
Nucleotide diversity values higher than 0.13 among CTV isolates have been estimated in some coding regions, but most of the nucleotide substitutions are transitions usually affecting the third codon position. The ratio between non‐synonymous and synonymous substitutions (dN/dS) estimated for all gRNA regions was below 1 (Rubio et al., 2001; A. Sambade et al., unpublished data), and in the range of values calculated for other plant viruses (García‐Arenal et al., 2001) and for most protein coding sequences (Nei, 1987), suggesting selective pressure for amino acid conservation. More detailed analyses showed purifying as well as balancing selection depending on isolates and gRNA regions (Sentandreu et al., 2006). In addition, sequence differences compatible with genetic drift, probably associated with reduced effective size of viral populations affected by transmission bottlenecks, have been shown after aphid transmission or host change (Albiach‐Martíet al., 2000b; Ayllón et al., 2006; Sentandreu et al., 2006). These changes were sometimes accompanied by a different pathogenic behaviour of isolates (Albiach‐Martíet al., 2000b; Brlansky et al., 2003; Hermoso de Mendoza et al., 1988a).
Finally, CTV populations are also affected by natural and cultural dispersal of the virus. Citrus are long‐lived trees that can be repeatedly inoculated by aphids with divergent CTV variants, thus increasing within‐isolate genetic diversity (Ayllón et al., 2006; Kong et al., 2000; Rubio et al., 2001). Uneven distribution of sequence variants within infected plants and/or random selection of some of them during aphid acquisition or transmission may be additional factors contributing to CTV population changes in the field (d’Urso et al., 2000, 2003; Sambade et al., 2007). The presence of divergent sequence variants in a CTV isolate also increases the possibility of genetic variation via recombination (Vives et al., 2005). By contrast, the movement of CTV‐infected buds tends to reduce genetic diversity between regions creating a unique CTV population. For example, CTV isolates from California and Spain were shown to be genetically the same population (Rubio et al., 2001).
In summary, although CTV seems to be genetically stable in some hosts, human‐driven interactions with different scion/rootstock combinations under different environmental conditions might have generated genetic variability, later modulated by other factors such as recombination, selection, genetic drift or gene flow between regions.
CONTROL OF CTV DISEASES
The ability to control disease damage depend to a large extent on the CTV incidence and on the virus strains and citrus varieties predominant in each region (Garnsey et al., 1998). Effective quarantine and budwood certification programmes are the best measures to avoid CTV introduction or dispersal in CTV‐free areas where citrus are still propagated on sour orange rootstock (Navarro et al., 2002). Elimination of infected trees is the best means to avoid or delay an epidemic, if CTV is at low incidence and infected trees are grouped in a limited number of foci (Bar‐Joseph et al., 1989; Gottwald et al., 2002; Kyriakou et al., 1996). When eradication becomes unfeasible due to efficient CTV dispersal by vectors, propagation of citrus on tristeza‐tolerant rootstocks is the only viable option to manage CTV without tristeza decline. P. trifoliata and its hybrids Carrizo and Troyer citrange (sweet orange ×Poncirus trifoliata) and Swingle citrumelo (grapefruit ×P. trifoliata) and Rangpur lime (C. limonia Osb.) are among the most widely used. Quarantine and certification programmes are still necessary to avoid importation and propagation of severe strains causing SP in commercial varieties. If severe SP isolates are widespread, using constitutive resistance to the virus or cross protecting sensitive varieties with appropriate mild isolates becomes necessary. However, technical challenges posed by these options need to be solved.
Breeding to incorporate resistance genes in commercial cultivars is considered the best approach to avoid crop losses due to pathogens. However, different features of citrus biology, particularly their complex genetics and reproductive biology, together with their large plant size have greatly impaired genetic improvement through conventional breeding. Most citrus genotypes are facultatively apomictic, and adventitious embryos usually preclude development of the less vigorous zygotic embryos, thus limiting the recovery of sexual progeny. Partial or total pollen and/or ovule sterility of important genotypes, and frequent cross‐ and self‐incompatibility, also limit the options for potential parents. The long juvenile period (at least 5 years to start blooming and even more to bear mature true‐to‐type fruits) makes conventional breeding unusually slow. Finally, their high levels of heterozygosity, a lack of knowledge on how the most important horticultural traits are inherited and finding quantitative inheritance for some of them have further complicated citrus breeding programmes.
Although strain‐specific resistance to CTV has been observed in some citrus species (Asíns et al., 2004; Fang and Roose, 1999; Garnsey et al., 1987; our unpublished results), resistance is more common in citrus relatives of other genera within the subfamily Aurantioideae, including some species of Fortunella, P. trifoliata, Severinia buxifolia and Swinglea glutinosa (Garnsey et al., 1987; Mestre et al., 1997a,b; Yoshida, 1996). CTV accumulation in S. buxifolia, Sw. glutinosa and P. trifoliata (cultivars carrying two, one or no Ctv alleles) protoplasts was as high as in Itaborai sweet orange protoplasts, indicating that none of these Citrus relatives was resistant at the cellular level (Albiach‐Martíet al., 2004). Particular attention has been devoted to P. trifoliata, which shows resistance to most CTV strains and is sexually compatible with Citrus. This resistance is associated with a single dominant locus (Ctv) that has been thoroughly characterized and mapped (Deng et al., 2001; Fang et al., 1998; Gmitter et al., 1996; Mestre et al., 1997a; Yang et al., 2003; Yoshida, 1985, 1993). The Ctv locus has been restricted to a 121‐kb segment in the Poncirus genome comprising ten genes (Rai, 2006). Transgenic grapefruit lines carrying any one of the ten candidate genes were susceptible to CTV infection, suggesting that more than one gene in the locus may be involved in resistance to CTV or that the role of other loci was overlooked (E. Mirkov, personal communication).
At present, the only means to protect commercial varieties sensitive to SP is classical cross protection with mild CTV strains. This approach has prevented low yield and small‐sized fruits of important varieties such as Pera sweet orange in Sao Paulo, Brazil (Costa and Müller, 1980), and Marsh grapefruit in South Africa (Van Vuuren et al., 1993). Protected isolates were selected from field trees of the same cultivar that had grown for years displaying mild or no symptoms at all. It was assumed that these plants were protected against infection with the most aggressive variants and they were used as budwood sources to propagate new plants. Pera cross protection has been critical for the success of the Brazilian citriculture, which has grown more than 90 million protected trees in the last 30 years with little damage. However, the same protection strategy had limited success in other areas or with other varieties (Broadbent et al., 1991; Ieki and Yamaguchi, 1988; Müller et al., 1988), indicating that cross protection probably depends on the varieties, CTV strains and environmental conditions prevalent in each region. Whether cross protection is coat protein‐ or RNA‐mediated has been a matter of discussion for years (Fraser, 1998). Although the role of the coat protein cannot be ruled out, post‐transcriptional gene silencing (PTGS), a dsRNA‐induced process, has recently arisen as a more likely mechanism to explain plant defence against viral infections and cross protection between closely related virus strains (Covey et al., 1997; Hammond et al., 2000; Ratcliff et al., 1997, 1999). CTV replication involves production of dsRNA intermediates that may trigger plant defence via PTGS. Indeed, Fagoaga et al. (2006) have observed high accumulation of viral‐specific small‐interfering (si)‐RNAs in CTV‐inoculated Mexican lime plants, indicating a strong natural PTGS‐mediated antiviral response in this host. On this basis, it is tempting to speculate that in cross protected plants the siRNAs generated by the mild strain could prevent subsequent infection by aggressive variants targeting for degradation their genomic and subgenomic RNAs. However, observation that mild CTV isolates often fail to protect or they afford only temporal protection to pre‐inoculated plants (Roistacher and Dodds, 1993; Sambade et al., 2002, 2007) strongly suggests that CTV is able to overcome PTGS or that other factors may be important for durable cross protection.
Pathogen‐derived resistance (PDR) in transgenic plants, also based on PTGS, could represent a better and more predictable strategy to achieve durable resistance to CTV in Citrus. The first demonstration of PDR to CTV came from the incorporation of the CP gene from a severe or a mild CTV strain in Mexican lime (Domínguez et al., 2002c). CTV inoculation of transgenic lines yielded two types of response: (1) most lines developed the same symptoms as the non‐transgenic controls, and (2) some lines exhibited a resistance behaviour that consisted of a fraction of plants (10–33%) being CTV immune, and the others showing a significant delay in virus accumulation and symptom onset. In some resistant lines CP accumulated at high levels, suggesting that a protein‐mediated resistance mechanism was operating in these lines, whereas in others CP was barely detectable. The same strategy has been used to transform other Citrus hosts, but results from challenge experiments were unclear or not reported (Febres et al., 2003; Ghorbel et al., 2000; Gutiérrez et al., 1997). Lime and grapefruit plants transformed with untranslatable versions of the CP gene to achieve RNA‐mediated resistance did not prevent CTV infection and the protection afforded was rapidly overcome by the challenging virus (Domínguez et al., 2002a,b; Febres et al., 2003; Herron et al., 2002). For the same purpose, constructs derived from the CTV 3′‐UTR were used to transform sweet orange protoplasts and grapefruit plants but conclusive results on protection at the whole plant level were not reported (Febres et al., 2003; Olivares‐Fuster et al., 2003). Ghorbel et al. (2001) explored over‐expression of the p23 protein in transgenic plants as a potential method to interfere in the normal CTV infection process. Unexpectedly, constitutive expression of p23 induced symptoms resembling those incited by CTV in non‐transgenic lime plants (Fig. 1H), whereas transgenic limes carrying a p23‐truncated version were normal. The intensity of symptoms paralleled p23 accumulation in transgenic plants (Ghorbel et al., 2001). Transgenic limes expressing the p23 gene from a severe or from a mild CTV isolate displayed leaf symptoms of similar intensity, and again symptom intensity was associated with the accumulation level of the p23 protein (Fagoaga et al., 2005). Transformation of other CTV‐susceptible or CTV‐resistant species, including sweet and sour orange and P. trifoliata, with the p23 gene also led to CTV‐like symptoms that did not appear in similar plants transformed with a truncated p23 version. The intensity of symptoms in Citrus species and relatives other than Mexican lime was associated with their level of p23 transcripts, but the p23 protein was barely detectable in these hosts. Higher accumulation of p23 in Mexican lime in comparison with sweet and sour orange was also observed in non‐transgenic plants inoculated with CTV, suggesting that even minimal amounts of p23 may cause deleterious effects in the latter two species. By contrast, transgenic expression of p23 in CTV non‐host N. tabacum and N. benthamiana species led to consistent accumulation of p23 without phenotypic aberrations, even if p23 acts as an RNA silencing suppressor in these species (Lu et al., 2004), suggesting that p23 might interact with miRNA‐regulated plant development processes in Citrus and Poncirus but not in Nicotiana non‐hosts. There is increasing evidence indicating that disease induction in the host may not be a side‐effect of silencing suppression but a consequence of disruption of the miRNA metabolism (Lewsey et al., 2007; Voinnet, 2005). Altogether, these results indicate that p23 is an important CTV pathogenicity determinant that interferes with plant development specifically in Citrus species and relatives (Fagoaga et al., 2005).
Some transgenic limes carrying the p23 gene had the characteristics typical of PTGS (multiple copies of the transgene and methylation of the silenced transgene, low levels of p23 mRNA, and accumulation of p23‐specific siRNAs) and showed normal aspect and development. Propagations of these lines graft‐ or aphid‐inoculated with CTV exhibited the resistance pattern previously observed: some plants were immune, others showed delayed and attenuated symptoms, and others were fully susceptible and showed normal symptoms and high virus titre. The third group accumulated more p23‐specific siRNAs than immune or non‐inoculated propagations and showed transgene demethylation. This variable response among clonal propagations of the same transgenic line indicates that factors other than the genetic background of the transgenic plant may play a key role in PTGS‐mediated resistance (Fagoaga et al., 2006).
Batuman et al. (2006) transformed N. benthamiana and C. macrophylla plants with a hairpin‐like construct comprising the p23 gene and 3′‐UTR in sense and antisense, separated by an intron, to yield a dsRNA structure upon expression in transgenic cells. Only a fraction of the transgenic C. macrophylla lines showed delayed CTV infection and none afforded durable protection. However, transgenic plants were propagated on susceptible rootstocks that after CTV inoculation would provide a permanent virus supply to the scion. We transformed Mexican lime plants with a similar construct and inoculated with CTV selected lines that carried a single transgene copy and accumulated high levels of siRNAs. Again, a variable number of propagations from some lines were fully resistant to CTV challenge (our unpublished results). Roy et al. (2006) found that transgenic N. benthamiana plants carrying CTV‐genome‐based constructs p25‐p20‐3′UTR or p25‐p23‐3′UTR showed resistance against Potato virus X (PVX)‐based vectors containing some homologous CTV sequences, but all transformants carrying p25 were susceptible to PVX. The reasons behind this unexpected result are not understood, but they may be related with the fact that N. benthamiana is not a natural CTV host.
In summary, the available data demonstrate that PDR is feasible with CTV in its natural hosts, but factors other than the genetic background of the transgenic plant may affect the resistance phenotype displayed by transgenic plant propagations. Whether transgenic citrus plants expressing CTV‐derived sequences could be an efficient alternative to cross protection for controlling SP damage in the field, or whether resistance to tristeza decline could be incorporated into the sour orange rootstock, remain to be tested.
GENERAL CONCLUSIONS AND FUTURE PROSPECTS
Seventy‐five years of tristeza epidemics have resulted in huge economic losses for the most important citrus industries propagated on sour orange rootstock. Moreover, CTV dispersal is an ongoing process and future losses from tristeza decline may exceed those that have occurred to date. Dispersal of T. citricida in Mexico and the Mediterranean basin, the two regions accumulating the largest citrus areas still grown on sour orange rootstock, could rapidly increase the rate of tristeza spread. The presence of this vector in some Mexican states and in localities of northern Spain and Portugal makes this scenario credible. With this perspective, it seems that CTV dispersal to new areas dramatically changed the course of the citrus industry.
While replacement of declining trees with new trees on tristeza‐tolerant rootstocks will eventually restore production, CTV‐induced SP will remain a more enduring problem. Indeed, detection of SP isolates in new regions has been periodically reported in the last 30 years. International movement of new varieties has unknowingly spread severe SP isolates and it is likely to continue in spite of quarantines established by many citrus‐producing countries. SP variants may also spread from current, unrecognized foci of infection, such as SP‐tolerant cultivars, and display symptoms only upon infection of an SP reactive host.
In the last 15 years, our understanding of the CTV molecular characteristics, expression strategies, genetic variability and epidemiology has improved significantly, and new highly sensitive and specific diagnostic procedures have been developed. However, permanent solutions to CTV‐induced diseases have remained elusive, mainly due to the lack of knowledge on the interactions between CTV, host plants and vectors. A deeper insight into these aspects is needed before sound control measures based on the use of resistance genes, PDR or cross protection can be implemented to avoid SP effects. Effective genetic systems based on agroinoculation techniques need to be fully developed to test the effects that genetic modifications engineered in different gRNA regions have on biological characteristics, such as host range, systemic spread and virus accumulation in the plant, symptom expression or aphid transmission.
Identification of the pathogenicity determinants for different disease syndromes should markedly improve our ability to identify specific pathotypes and limit dispersal of those that are potentially dangerous via nursery propagation of infected buds. Knowledge on the host factors interacting with CTV proteins will be necessary to understand the pathosystem and also the variable behaviour observed in transgenic plants expressing CTV sequences, a critical step in developing PDR. Understanding virus–host interactions is also a necessary step to develop reliable cross protection strategies, particularly considering the extensive genetic variability detected in CTV isolates. In this regard, the finding that CTV has three potential silencing suppressors and that half of the genes induced by CTV infection in Mexican lime do not have obvious similarities with other plant proteins suggests that the CTV–citrus interaction may be unusually complex and specific. Characterization of the CTV–vector interaction, including identification of potential helper factors, will provide new insights on the virus epidemiology and perhaps open new control possibilities. In summary, recent advances have raised new important questions on virus interactions with host plant and vector that need to be answered before a pacific coexistance between CTV and citrus can be re‐established.
ACKNOWLEDGEMENTS
S.A. and M.R.A.M. were recipients of postdoctoral contracts from the ‘Ramón y Cajal’ programme and from the INIA, respectively, Ministerio de Educación y Ciencia (MEC), and presently they have a research contract from Agroalimed (‘Fundación de la Comunidad Valenciana para la Investigación Agroalimentaria’). This work was supported by grants AGL2004‐05099/AGR and AGL2007‐61885/AGR from the MEC.
REFERENCES
- Albiach‐Martí, M.R. , Guerri, J. , Cambra, M. , Garnsey, S.M. and Moreno, P. (2000a) Differentiation of citrus tristeza virus isolates by serological analysis of the p25 coat protein peptide maps. J. Virol. Methods, 88, 25–34. [DOI] [PubMed] [Google Scholar]
- Albiach‐Martí, M.R. , Guerri, J. , Hermoso de Mendoza, A. , Laigret, F. , Ballester‐Olmos, J.F. and Moreno, P. (2000b) Aphid transmission alters the genomic and defective RNA populations of citrus tristeza virus. Phytopathology, 90, 134–138. [DOI] [PubMed] [Google Scholar]
- Albiach‐Martí, M.R. , Mawassi, M. , Gowda, S. , Satyanayanana, T. , Hilf, M.E. , Shanker, S. , Almira, E.C. , Vives, M.C. , López, C. , Guerri, J. , Flores, R. , Moreno, P. , Garnsey, S.M. and Dawson, W.O. (2000c) Sequences of Citrus tristeza virus separated in time and space are essentially identical. J. Virol. 74, 6856–6865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albiach‐Martí, M.R. , Grosser, J.W. , Gowda, S. , Mawassi, M. , Satyanarayana, T. , Garnsey, S.M. and Dawson, W.O. (2004) Citrus tristeza virus replicates and forms infectious virions in protoplast of resistant citrus relatives. Mol. Breeding, 14, 117–128. [Google Scholar]
- Alzhanova, D.V. , Prokhnevsky, A.I. , Peremyslov, V.V. and Dolja, V.V. (2007) Virion tails of beet yellows virus: coordinated assembly by three structural proteins. Virology, 359, 220–226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asíns, M.J. , Bernet, G.P. , Ruiz, C. , Cambra, M. , Guerri, J. and Carbonell, E. (2004) QTL analysis of citrus tristeza virus–citradia interaction. Theor. Appl. Genet. 97, 1145–1154. [DOI] [PubMed] [Google Scholar]
- Ayllón, M.A. , López, C. , Navas‐Castillo, J. , Mawassi, M. and Dawson, W.O. (1999a) New defective RNAs from citrus tristeza virus: evidence for a replicase driven template switching mechanism in their generation. J. Gen. Virol. 80, 817–821. [DOI] [PubMed] [Google Scholar]
- Ayllón, M.A. , Rubio, L. , Moya, A. , Guerri, J. and Moreno, P. (1999b) The haplotype distribution of two genes of Citrus tristeza virus is altered after host change or aphid transmission. Virology, 255, 32–39. [DOI] [PubMed] [Google Scholar]
- Ayllón, M.A. , López, C. , Navas‐Castillo, J. , Garnsey, S.M. , Guerri, J. , Flores, R. and Moreno, P. (2001) Polymorphism of the 5’ terminal region of Citrus tristeza virus (CTV) RNA: incidence of three sequence types in isolates of different origin and pathogenicity. Arch. Virol. 146, 27–40. [DOI] [PubMed] [Google Scholar]
- Ayllón, M.A. , Gowda, S. , Satyanayanana, T. , Karasev, A.V. , Adkins, S. , Mawassi, M. , Guerri, J. , Moreno, P. and Dawson, W.O. (2003) Effects of modification of the transcription initiation site context on Citrus tristeza virus subgenomic RNA synthesis. J. Virol. 77, 9232–9243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ayllón, M.A. , Gowda, S. , Satyanarayana, T. and Dawson, W.O. (2004) Cis‐acting elements at opposite ends of the Citrus tristeza virus genome differ in initiation and termination of subgenomic RNAs. Virology, 322, 41–50. [DOI] [PubMed] [Google Scholar]
- Ayllón, M.A. , Rubio, L. , Sentandreu, V. , Moya, A. , Guerri, J. and Moreno, P. (2006) Variations in two gene sequences of citrus tristeza virus after host passage. Virus Genes, 32, 119–128. [DOI] [PubMed] [Google Scholar]
- Ballester‐Olmos, J.F. , Pina, J.A. , Carbonell, E. , Moreno, P. , Hermoso de Mendoza, A. , Cambra, M. and Navarro, L. (1993) Biological diversity of citrus tristeza virus (CTV) isolates in Spain. Plant Pathol. 42, 219–229. [Google Scholar]
- Barbarossa, L. and Savino, V. (2006) Sensitive and specific digoxigenin‐labelled RNA probes for routine detection of Citrus tristeza virus by dot‐blot hybridization. J. Phytopathol. 154, 329–335. [Google Scholar]
- Bar‐Joseph, M. and Loebenstein, G. (1973) Effects of strain, source plant, and temperature on transmissibility of citrus tristeza virus by melon aphid. Phytopathology, 63, 716–720. [Google Scholar]
- Bar‐Joseph, M. , Loebenstein, G. and Cohen, J. (1972) Further purification and characterization of threadlike particles associated with the citrus tristeza disease. Virology, 50, 821–828. [DOI] [PubMed] [Google Scholar]
- Bar‐Joseph, M. , Garnsey, S.M. , Gonsalves, D. , Moscovitz, M. , Purcifull, D.E. , Clark, M.F. and Loebenstein, G. (1979) The use of enzyme‐linked immunosorbent assay for detection of citrus tristeza virus. Phytopathology, 69, 190–194. [Google Scholar]
- Bar‐Joseph, M. , Marcus, R. and Lee. R.F. (1989) The continuous challenge of citrus tristeza virus control. Annu. Rev. Phytopathol. 27, 291–316. [Google Scholar]
- Batuman, O. , Mawassi, M. and Bar‐Joseph, M. (2006) Transgenes consisting of a dsRNA of an RNAi suppressor plus the 3′UTR provide resistance to Citrus tristeza virus sequences in Nicotiana benthamiana but not in citrus. Virus Genes, 33, 319–327. [DOI] [PubMed] [Google Scholar]
- Ben Ze’ev, I.S. , Bar‐Joseph, M. , Nitzan, Y. and Marcus, R. (1989) A severe citrus tristeza virus isolate causing collapse of trees of sour orange before virus is detectable throughout the canopy. Ann. Appl. Biol. 114, 293–300. [Google Scholar]
- Bertolini, E. , Moreno, A. , Capote, N. , Olmos, A. , De Luis, A. , Vidal, E. , Pérez‐Panadés, J. and Cambra, M. (2007) Quantitative detection of Citrus tristeza virus in plant tissues and single aphids by real‐time RT‐PCR. Eur. J. Plant Pathol. doi 10.10071s10658-077-9206-9. [Google Scholar]
- Brlansky, R.H. , Damsteegt, V.D. , Howd, D.S. and Roy, A. (2003) Molecular analyses of citrus tristeza virus subisolates separated by aphid transmission. Plant Dis. 87, 397–401. [DOI] [PubMed] [Google Scholar]
- Broadbent, P. , Bevington, K.B. and Coote, B.G. (1991) Control of stem pitting of grapefruit in Australia by mild strain cross protection In: Proceedings of the 11th Conference of the International Organization of Citrus Virologists (Brlansky R.H., Lee R.F. and Timmer L.W., eds), pp. 64–70. Riverside, CA: IOCV. [Google Scholar]
- Cambra, M. , Camarasa, E. , Gorris, M.T. , Garnsey, S.M. and Carbonell, E. (1991) Comparison of different immunosorbent assays for citrus tristeza virus (CTV) using CTV‐specific monoclonal and polyclonal antibodies In: Proceedings of the 11th Conference of the International Organization of Citrus Virologists (Brlansky R.H., Lee R.F. and Timmer L.W., eds), pp. 38–45. Riverside, CA: IOCV. [Google Scholar]
- Cambra, M. , Gorris, M.T. , Marroquín, C. , Román, M.P. , Olmos, A. , Martínez, P.C. , Hermoso de Mendoza, A.H. , López, A. and Navarro, L. (2000) Incidence and epidemiology of citrus tristeza virus in the Valencian Community of Spain. Virus Res. 71, 85–95. [DOI] [PubMed] [Google Scholar]
- Che, X. , Piestum, D. , Mawassi, M. , Satyanayanana, T. , Gowda, S. , Dawson W.O. and Bar‐Joseph, M. (2001) 5′ coterminal subgenonic RNAs in citrus tristeza virus‐infected cells. Virology, 283, 374–381. [DOI] [PubMed] [Google Scholar]
- Che, X. , Mawassi, M. and Bar‐Joseph, M. (2002) A novel class of large and infectious defective RNAs of Citrus tristeza virus . Virology, 298, 133–145. [DOI] [PubMed] [Google Scholar]
- Che, X. , Dawson, W.O. and Bar‐Joseph, M. (2003) Defective RNAs of Citrus tristeza virus analogous to Crinivirus genomic RNAs. Virology, 310, 298–309. [DOI] [PubMed] [Google Scholar]
- Costa, A.S. and Grant, T.J. (1951) Studies on transmission of the tristeza virus by the vector Aphis citricidus. Phytopathology, 41, 103–115. [Google Scholar]
- Costa, A.S. and Müller, G.W. (1980) Tristeza control by cross protection: a US–Brazil cooperative success. Plant Dis. 64, 538–541. [Google Scholar]
- Covey, S.N. , Alkaff, N.S. , Langara, A. and Turner, D.S. (1997) Plants combat infection by gene silencing. Nature, 385, 781–782. [Google Scholar]
- D’Urso, F. , Ayllón, M.A. , Rubio, L. , Sambade, A. , Hermoso de Mendoza, A. , Guerri, J. and Moreno, P. (2000) Contribution of uneven distribution of genomic RNA variants of citrus tristeza virus (CTV) within the plant to changes in the viral population following aphid transmission. Plant Pathol. 49, 288–294. [Google Scholar]
- D’Urso, F. , Sambade, A. , Moya, A. , Guerri, J. and Moreno, P. (2003) Variation of haplotype distributions of two genomic regions of Citrus tristeza virus populations from eastern Spain. Mol. Ecol. 12, 517–526. [DOI] [PubMed] [Google Scholar]
- Davino, S. , Davino, M. , Sambade, A. , Guardo, M. and Caruso, A. (2003) The first Citrus tristeza virus outbreak found in a relevant citrus producing area of Sicily, Italy. Plant Dis. 87, 314. [DOI] [PubMed] [Google Scholar]
- Deng, Z. , Huang, S. , Ling, P. , Yu, C. , Tao, Q. , Chen, C. , Wendell, M.K. , Zhang, H.‐B. and Gmitter, F.G. Jr. (2001) Fine genetic mapping and BAC contig development for the citrus tristeza virus resistance gene locus in Poncirus trifoliata (Raf.). Mol. Genet. Genomics, 265, 739–747. [DOI] [PubMed] [Google Scholar]
- Dickson, R.C. , Johnson, M.M. , Flock, R.A. and Laird, E.F. (1956) Flying aphid populations in Southern California citrus groves and their relation to the transmission of the tristeza virus. Phytopathology, 46, 204–210. [Google Scholar]
- Dodds, J.A. , Jarupat, T. , Lee, J.G. and Roistacher, C.N. (1987) Effects of strain, host, time of harvest, and virus concentration on double‐stranded RNA analysis of citrus tristeza virus. Phytopathology, 77, 442–447. [Google Scholar]
- Dolja, V.V. , Kreuze, J.F. and Valkonen, J.P.T. (2006) Comparative and functional genomics of closteroviruses. Virus Research, 117, 38–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Domingo, E. and Holland, J.J. (1994) Mutation rates and rapid evolution of RNA viruses In: The Evolutionary Biology of Viruses (Morse S.S., ed.), pp. 161–184. New York: Raven Press. [Google Scholar]
- Domínguez, A. , Fagoaga, C. , Navarro, L. , Moreno, P. and Peña, L. (2002a) Regeneration of transgenic citrus plants under non selective conditions results in high frequency recovery of plants with silenced transgenes. Mol. Genet. Genomics, 267, 544–556. [DOI] [PubMed] [Google Scholar]
- Domínguez, A. , Fagoaga, C. , Navarro, L. , Moreno, P. and Peña, L. (2002b) Constitutive expression of untranslatable versions of the p25 coat protein gene in Mexican lime (Citrus aurantifolia (Christm.) Swing.) transgenic plants does not confer resistance to citrus tristeza virus (CTV) In: Proceedings of the 15th Conference of the International Organization of Citrus Virologists (Durán‐Vila N., Milne R.G. and Da Graça J.V., eds), pp. 341–344. Riverside, CA: IOCV. [Google Scholar]
- Domínguez, A. , Hermoso de Mendoza, A. , Guerri, J. , Cambra, M. , Navarro, L. , Moreno, P. and Peña, L. (2002c) Pathogen‐derived resistance to Citrus tristeza virus (CTV) in transgenic Mexican lime (Citrus aurantifolia (Christm.) Swing.) plants expressing its p25 coat protein gene. Mol. Breeding, 10, 1–10. [Google Scholar]
- Duran‐Vila, N. and Moreno, P. (2000) Enfermedades de los Cítricos. Madrid: Ediciones Mundi‐Prensa. [Google Scholar]
- Fagoaga, C. , López, C. , Moreno, P. , Navarro, L. , Flores, R. and Peña, L. (2005) Viral‐like symptoms induced by the ectopic expresión of the p23 of Citrus tristeza virus are citrus specific and do not correlate with the patogenicity of the virus strain. Mol. Plant–Microbe Interact. 18, 435–445. [DOI] [PubMed] [Google Scholar]
- Fagoaga, C. , López, C. , Hermoso de Mendoza, A.H. , Moreno, P. , Navarro, L. , Flores, R. and Peña, L. (2006) Post‐transcriptional gene silencing of the p23 silencing suppressor of Citrus tristeza virus confers resistance to the virus in transgenic Mexican lime. Plant Mol. Biol. 66, 153–165. [DOI] [PubMed] [Google Scholar]
- Fang, D.Q. and Roose, M.L. (1999) A novel gene conferring Citrus tristeza virus resistance in Citrus maxima (Burm.) Merrill. HortScience, 34, 334–335. [Google Scholar]
- Fang, D.Q. , Federici, C.T. and Roose, M.L. (1998) A high‐resolution linkage map of the citrus tristeza virus resistance gene region in Poncirus trifoliata (L.) Raf. Genetics, 150, 883–890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Febres, V.J. , Ashoulin, L. , Mawassi, M. , Frank, A. , Bar‐Joseph, M. , Manjunath, K.L. , Lee, R.F. and Niblett, C.L. (1996) The p27 protein is present at one end of citrus tristeza virus particles. Phytopathology, 86, 1331–1335. [Google Scholar]
- Febres, V.J. , Niblett, C.L. , Lee, R.F. and Moore, G.A. (2003) Characterization of grapefruit plants (Citrus paradisi Macf.) transformed with citrus tristeza closterovirus genes. Plant Cell Rep. 21, 421–428. [DOI] [PubMed] [Google Scholar]
- Folimonov, A.S. , Folimonova, S.Y. , Bar‐Joseph, M. and Dawson, W.O. (2007) A stable RNA virus‐based vector for citrus trees. Virology, 368, 205–216. [DOI] [PubMed] [Google Scholar]
- Fraser, L. (1952) Seedling yellows, an unreported virus disease of citrus. Agr. Gaz. N.S. Wales, 63, 125–131. [Google Scholar]
- Fraser, R.S.S. (1998) Introduction to classical cross protection In: Methods in Molecular Biology volume 81, Plant Virus Protocols (Foster D. and Taylor S.J., eds), pp. 13–24. Totowa, NJ: Humana Press. [DOI] [PubMed] [Google Scholar]
- Gandía, M. , Conesa, A. , Ancillo, G. , Gadea, J. , Forment, J. , Pallás, V. , Flores, R. , Duran‐Vila, N. , Moreno, P. and Guerri, J. (2007) Transcriptional response of Citrus aurantifolia to infection by Citrus tristeza virus . Virology, 367, 298–306. [DOI] [PubMed] [Google Scholar]
- García‐Arenal, F. , Fraile, A. and Malpica, J.M. (2001) Variability and genetic structure of plant virus populations. Annu. Rev. Phytopathol. 39, 157–186. [DOI] [PubMed] [Google Scholar]
- Garnsey, S.M. , Barrett, H.C. and Hutchinson, D.J. (1987) Identification of citrus tristeza virus resistance in citrus relatives and its potential applications. Phytophylactica, 19, 187–191. [Google Scholar]
- Garnsey, S.M. , Civerolo, E.L. , Gumpf, D.J. , Yokomi, R.K. and Lee, R.F. (1991) Development of a worldwide collection of citrus tristeza virus isolates In: Proceedings of the 11th Conference of the International Organization of Citrus Virologists (Brlansky R.H., Lee R.F. and Timmer L.W., eds), pp. 113–120. Riverside, CA: IOCV. [Google Scholar]
- Garnsey, S.M , Permar, T.A. , Cambra, M. and Henderson, C.T. (1993) Direct tissue blot immunoassay (DTBIA) for detection of citrus tristeza virus (CTV) In: Proceedings of the 12th Conference of the International Organization of Citrus Virologists (Moreno P., Da Graça J. and Timmer L.W., eds), pp. 39–50. Riverside, CA: IOCV. [Google Scholar]
- Garnsey, S.M. , Gonsalves, D. and Purcifull, D.E. (1977) Mechanical transmission of citrus tristeza virus. Phytopathology, 67, 965–968. [Google Scholar]
- Garnsey, S.M. , Gottwald, T.R. and Yokomi, R.K. (1998) Control strategies for citrus tristeza virus In: Plant Virus Disease Control (Hadidi A., Khetarpal R. and Koganezawa H., eds), pp. 639–658. St. Paul, MN: APS Press. [Google Scholar]
- Garnsey, S.M. , Gottwald, T.R. , Hilf, M.E. , Matos, L. and Borbón, J. (2000) Emergence and spread of severe strains of Citrus tristeza virus isolates in the Dominican Republic In: Proceedings of the 14th Conference of the International Organization of Citrus Virologists (Da Graça J.V., Lee R.F. and Yokomi R. K., eds), pp. 57–68. Riverside, CA: IOCV. [Google Scholar]
- Garnsey, S.M. , Su, H.J. and Tsai, M.C. (1996) Differential susceptibility of pummelo and Swingle citrumelo to isolates of Citrus tristeza virus. In: Proceedings of the 13th Conference of the International Organization of Citrus Virologists (Da Graça J.V., Moreno P. and Yokomi. R.K., eds), pp. 138–146. Riverside, CA: IOCV. [Google Scholar]
- Ghorbel, R. , Domínguez, A. , Navarro, L. and Peña, L. (2000) High efficiency genetic transformation of sour orange (Citrus aurantium L.) and production of transgenic trees containing the coat protein gene of Citrus tristeza virus . Tree Physiol. 20, 1183–1189. [DOI] [PubMed] [Google Scholar]
- Ghorbel, R. , López, C. , Moreno, P. , Navarro, L. , Flores, R. and Peña, L. (2001) Transgenic citrus plants expressing the Citrus tristeza virus p23 protein exhibit viral‐like symptoms. Mol. Plant Pathol. 2, 27–36. [DOI] [PubMed] [Google Scholar]
- Gillings, M. , Broadbent, P. , Indsto, J. and Lee, R. (1993) Characterization of isolates and strains of citrus tristeza closterovirus using restriction analysis of the coat protein gene amplified by the polymerase chain reaction. J. Virol. Methods, 44, 305–317. [DOI] [PubMed] [Google Scholar]
- Gmitter, F.G. Jr , Xiao, S.Y. , Huang, S. and Hu, X.L. (1996) A localized linkage map of the citrus tristeza virus resistance gene region. Theor. Appl. Genet. 92, 688–695. [DOI] [PubMed] [Google Scholar]
- Gonsalves, D. , Purcifull, D.E. and Garnsey, S.M. (1978) Purification and serology of citrus tristeza virus. Phytopathology, 68, 553–559. [Google Scholar]
- Gottwald, T.R. , Cambra, M. , Moreno, P. , Camarasa, E. and Piquer, J. (1996a) Spatial and temporal analyses of citrus tristeza virus in Eastern Spain. Phytopathology, 86, 45–55. [Google Scholar]
- Gottwald, T.R. , Garnsey, S.M. , Cambra, M. , Moreno, P. , Irey, M. and Borbón, J. (1996b) Differential effects of Toxoptera citricida vs. Aphis gossypii on temporal increase and spatial patterns of spread of citrus tristeza In: Proceedings of the 13th Conference of the International Organization of Citrus Virologists (Da Graça J.V., Moreno P. and Yokomi R.K., eds), pp. 120–129. Riverside, CA: IOCV. [Google Scholar]
- Gottwald, T.R. , Garnsey, S.M. and Borbón, J. (1998) Increase and patterns of spread of citrus tristeza virus infections in Costa Rica and the Dominican Republic in the presence of the brown citrus aphid, Toxoptera citricida . Phytopathology, 88, 621–636. [DOI] [PubMed] [Google Scholar]
- Gottwald, T.R. , Gibson, G.J. , Garnsey, S.M. and Irey, M. (1999) Examination of the effect of aphid vector population composition on the spatial dynamics of citrus tristeza virus spread by stochastic modeling. Phytopathology, 89, 603–608. [DOI] [PubMed] [Google Scholar]
- Gottwald, T.R. , Polek, M. and Riley, K. (2002) History, present incidence, and spatial distribution of Citrus tristeza virus in the California Central Valley. In Proceedings of the 15th Conference of the International Organization of Citrus Virologists (Duran‐Vila N., Milne R.G. and Da Graça J.V., eds), pp. 83–94. Riverside, CA: IOCV. [Google Scholar]
- Gowda, S. , Satyanayanana, T. , Davis, C.L. , Navas‐Castillo, J. , Albiach‐Martí, M.R. , Mawassi, M. , Valkov, N. , Bar‐Joseph, M. , Moreno, P. and Dawson, W.O. (2000) The p20 gene product of Citrus tristeza virus accumulates in the amorphous inclusion bodies. Virology, 274, 246–254. [DOI] [PubMed] [Google Scholar]
- Gowda, S. , Satyanayanana, T. , Ayllón, M.A. , Albiach‐Martí, M.R. , Mawassi, M. , Rabindran, S. and Dawson, W.O. (2001) Characterization of the cis‐acting elements controlling subgenomic mRNAs of Citrus tristeza virus: production of positive‐and negative‐stranded 3′‐terminal and positive‐stranded 5′ terminal RNAs. Virology, 286, 134–151. [DOI] [PubMed] [Google Scholar]
- Gowda, S. , Ayllón, M.A. , Satyanayanana, T. , Bar‐Joseph, M. and Dawson, W.O. (2003a) Transcription strategy in a Closterovirus: a novel 5′‐proximal controler element of Citrus tristeza virus produces 5′‐ and 3′‐terminal subgenomic RNAs and differs from 3′ open reading frame controler elements. J. Virol. 77, 340–352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gowda, S. , Satyanayanana, T. , Ayllón, M.A. , Moreno, P. , Flores, R. and Dawson, W.O. (2003b) The conserved structures of the 5′ nontranslated region of Citrus tristeza virus are involved in replication and virion assembly. Virology, 317, 50–64. [DOI] [PubMed] [Google Scholar]
- Gowda, S. , Satyanarayana, T. , Robertson, C.J. , Garnsey, S.M. and Dawson, W.O. (2005) Infection of citrus plants with virions generated in Nicotiana benthamiana plants agroinfiltrated with binary vector based Citrus tristeza virus. In: Proceedings of the 16th Conference of the International Organization of Citrus Virologists (Hilf M.E., Duran‐Vila N. and Rocha‐Peña M.A., eds), pp. 23–33. Riverside, CA: IOCV. [Google Scholar]
- Grant, T.J. and Higgins, R.P. (1957) Occurrence of mixtures of tristeza virus strains in citrus. Phytopathology, 47, 272–276. [Google Scholar]
- Guerri, J. , Moreno, P. , Muñoz, N. and Martínez, M.E. (1991) Variability among Spanish citrus tristeza virus isolates revealed by double‐stranded RNA analysis. Plant Pathol. 40, 38–44. [Google Scholar]
- Gutiérrez, M.A. , Luth, D.E. and Moore, G.A. (1997) Factors affecting Agrobacterium‐mediated transformation in Citrus and production of sour orange (Citrus aurantium L.) plants expressing the coat protein gene of Citrus tristeza virus . Plant Cell Rep. 16, 745–753. [DOI] [PubMed] [Google Scholar]
- Halbert, S.E. , Genc, H. , Çevik, B. , Brown, L.G. , Rosales, I.M. , Manjunath, K.L. , Pomerinke, M. , Davison, D.A. , Lee, R.F. and Niblett, C.L. (2004) Distribution and characterization of Citrus tristeza virus in South Florida following establishment of Toxoptera citricida . Plant Dis. 88, 935–941. [DOI] [PubMed] [Google Scholar]
- Hammond, S.M. , Bernstein, E. , Beach, D. and Hannon, G.J. (2000) An RNA‐directed nuclease mediates post‐transcriptional gene silencing in Drosophila cells. Nature, 404, 293–296. [DOI] [PubMed] [Google Scholar]
- Hermoso de Mendoza, A. , Ballester‐Olmos, J.F. and Pina, J.A. (1984) Transmission of citrus tristeza virus by aphids (Homoptera, Aphididae) in Spain In: Proceedings of the 9th Conference of the International Organization of Citrus Virologists (Garnsey S.M., Timmer L.W. and Dodds J.A., eds), pp. 23–27. Riverside, CA: IOCV. [Google Scholar]
- Hermoso de Mendoza, A. , Ballester‐Olmos, J.F. and Pina, J.A. (1988a) Comparative aphid transmission of a common citrus tristeza virus isolate and a seedling yellows isolate recently introduced into Spain In: Proceedings of the 10th Conference of the International Organization of Citrus Virologists (Timmer L.W., Garnsey S.M. and Navarro L., eds), pp. 68–70. Riverside, CA: IOCV. [Google Scholar]
- Hermoso de Mendoza, A. , Ballester‐Olmos, J.F. , Pina, J.A. , Serra, J.A. and Fuertes, C. (1988b) Difference in transmission efficiency of citrus tristeza virus by Aphis gossypii using sweet orange, mandarin or lemon trees as donor or receptor host plant In: Proceedings of the 10th Conference of the International Organization of Citrus Virologists (Timmer L.W., Garnsey S.M. and Navarro L., eds), pp. 62–64. Riverside, CA: IOCV. [Google Scholar]
- Herron, C.M. , Yang, Z.‐N. , Molina, J.J. , da Graça, J.V. , Van Vuuren, S.P. and Mirkov, T.E. (2002) Preliminary evaluation of uncp transgenic Rio Red grapefruit scions for resistance to Citrus tristeza virus In: Proceedings of the 15th Conference of the International Organization of Citrus Virologists (Durán‐Vila N., Milne R.G. and Da Graça J.V., eds), p. 434 Riverside, CA: IOCV. [Google Scholar]
- Hilf, M.E. , Karasev, A.V. , Pappu, H.R. , Gumpf, D.J. , Niblett, C.L. and Garnsey, S.M. (1995) Characterization of citrus tristeza virus subgenomic RNAs in infected tissue. Virology, 208, 576–582. [DOI] [PubMed] [Google Scholar]
- Hilf, M.E. , Karasev, A.V. , Albiach‐Martí, M.R. , Dawson, W.O. and Garnsey, S.M. (1999) Two paths of sequence divergence in the citrus tristeza virus complex. Phytopathology, 89, 336–342. [DOI] [PubMed] [Google Scholar]
- Hilf, M.E. , Mavrodieva, V.A. and Garnsey, S.M. (2005) Genetic marker analysis of a global collection of isolates of Citrus tristeza virus: Characterization and distribution of CTV genotypes and association with symptoms. Phytopathology, 95, 909–917. [DOI] [PubMed] [Google Scholar]
- Hughes, G. and Gottwald, T.R. (1998) Survey methods for assessment of citrus tristeza virus incidence. Phytopathology, 88, 715–723. [DOI] [PubMed] [Google Scholar]
- Hughes, W.A. and Lister, C.A. (1949) Lime disease in the Gold Coast. Nature, 164, 880. [DOI] [PubMed] [Google Scholar]
- Ieki, H. and Yamaguchi, A. (1988) Protective interference of mild strains of citrus tristeza virus against a severe strain in Morita Navel orange. In: Proceedings of the 10th Conference of the International Organization of Citrus Virologists (Timmer L.W., Garnsey S.M. and Navarro L., eds), pp. 86–90. Riverside, CA: IOCV. [Google Scholar]
- Ilharco, F.A. , Sousa‐Silva, C.R. and Alvarez‐Alvarez, A. (2005) First report on Toxoptera citricidus (Kirkaldy) in Spain and continental Portugal. Agron. Lusit. 51, 19–21. [Google Scholar]
- Karasev, A.V. , Boyko, V.P. , Gowda, S. , Nikolaeva, O.V. , Hilf, M.E. , Koonin, E.V. , Niblett, C.L. Cline, K. , Gumpf, D.J. , Lee, R.F. , Garnsey, S.M. , Lewandowski, D.J. and Dawson, W.O. (1995) Complete sequence of the citrus tristeza virus RNA genome. Virology, 208: 511–520. [DOI] [PubMed] [Google Scholar]
- Karasev, A.V. , Hilf, M.E. , Garnsey, S.M. and Dawson, W.O. (1997) Transcriptional strategy of closteroviruses: maping the 5′ termini of the citrus tristeza virus subgenomic RNAs. J. Virol. 71, 6233–6236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kong, P. , Rubio, L. Polek, M. and Falk, B.W. (2000) Population structure and genetic diversity within California Citrus tristeza virus (CTV) field isolates. Virus Genes, 21, 139–145. [DOI] [PubMed] [Google Scholar]
- Kyriakou, A. , Ioannou, N. , Gavriel, J. , Bar‐Joseph, M. , Papayiannis, Chr. , Kapar‐Isaia, Th. and Savva, G. (1996) Management of citrus tristeza virus in Cyprus In: Proceedings of the 13th Conference of the International Organization of Citrus Virologists (Da Graça J.V., Moreno P. and Yokomi R.K., eds), pp. 172–178. Riverside, CA: IOCV. [Google Scholar]
- Lewsey, M. , Robertson, F.C. , Canto, T. , Palukaitis, P. and Carr, J.P. (2007) Selective targeting of miRNA‐regulated plant development by a viral counter‐silencing protein. Plant J. 50, 240–252. [DOI] [PubMed] [Google Scholar]
- López, C. , Ayllón, M.A. , Navas‐Castillo, J. , Guerri, J. , Moreno, P. and Flores, R. (1998) Molecular variability of the 5′ and 3′ terminal regions of citrus tristeza virus RNA. Phytopathology, 88, 685–691. [DOI] [PubMed] [Google Scholar]
- López, C. , Navas‐Castillo, J. , Gowda, S. , Moreno, P. and Flores, R. (2000) The 23‐kDa protein coded by the 3′‐terminal gene of citrus tristeza virus is an RNA‐binding protein. Virology, 269, 462–470. [DOI] [PubMed] [Google Scholar]
- Lu, R. , Folimonov, A. , Shintaku, M. , Li, W.X. , Falk, B.W. , Dawson, W.O. and Ding, S.W. (2004) Three distinct suppressors of RNA silencing encoded by a 20‐kb viral RNA genome. Proc. Natl Acad. Sci. USA, 101, 15742–15747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marroquín, C. , Olmos, A. , Gorris, M.T. , Bertolini, E. , Martínez, M.C. , Carbonell, E.A. , Hermoso de Mendoza, A.H. and Cambra, M. (2004) Estimation of the number of aphids carrying Citrus tristeza virus that visit adult citrus trees. Virus Res. 100, 101–108. [DOI] [PubMed] [Google Scholar]
- Mawassi, M. , Karasev, A.V. , Mietkiewska, E. , Gafny, R. , Lee, R.F. , Dawson, W.O. and Bar‐Joseph, M. (1995a) Defective RNA molecules associated with citrus tristeza virus. Virology, 208, 383–387. [DOI] [PubMed] [Google Scholar]
- Mawassi, M. , Mietkiewska, E. , Hilf, M.E. , Ashoulin, L. , Karasev, A.V. , Gafny, A.V. , Lee, R.F. , Garnsey, S.M. , Dawson, W.O. and Bar‐Joseph, M. (1995b) Multiple species of defective RNAs in plants infected with citrus tristeza virus. Virology, 214, 264–268. [DOI] [PubMed] [Google Scholar]
- Mawassi, M. , Mietkiewska, E. , Gofman, R. , Yang, G. and Bar‐Joseph, M. (1996) Unusual sequence relationships between two isolates of citrus tristeza virus. J. Gen.Virol. 77, 2359–2364. [DOI] [PubMed] [Google Scholar]
- Mawassi, M. , Satyanayanana, T. , Albiach‐Martí, M.R. , Gowda, S. , Ayllón, M.A , Robertson, C. and Dawson, W.O. (2000a) The fitness of Citrus tristeza virus defective RNA is affected by the length of their 5′ and 3′ termini and by coding capacity. Virology, 275, 42–56. [DOI] [PubMed] [Google Scholar]
- Mawassi, M. , Satyanayanana, T. , Gowda, S. , Albiach‐Martí, M.R. , Robertson, C. and Dawson, W.O. (2000b) Replication of heterologous combinations of helper and defective RNA of Citrus tristeza virus . Virology, 267, 360–369. [DOI] [PubMed] [Google Scholar]
- McClean, A.P.D. (1956) Tristeza and stem pitting diseases of citrus in South Africa. FAO Plant Protect. Bull. 4, 88–94. [Google Scholar]
- McClean, A.P.D. (1957) Tristeza virus of citrus: evidence for absence of seed transmission. Plant Dis. Reptr. 41, 821. [Google Scholar]
- McClean, A.P.D. (1960) Seedling‐yellows in South African citrus trees. S. Afr. J. Agric. Sci. 3, 259–279. [Google Scholar]
- McClean, A.P.D. (1963) The tristeza complex: Its variability in field‐grown citrus in South Africa. S. Afr. J. Agric. Sci. 6, 303–332. [Google Scholar]
- McClean, A.P.D. (1975) Tristeza virus complex: its transmission by aphid Toxoptera citricidus . Phytophylactica, 7, 109–113 [Google Scholar]
- Mestre, P.F. , Asins, M.J. , Pina, J.A. , Carbonell, E.A. and Navarro, L. (1997a) Molecular markers flanking citrus tristeza virus resistance gene from Poncirus trifoliata (L.) Raf. Theor. Appl. Genetics, 94, 458–464. [Google Scholar]
- Mestre, P.F. , Asins, M.J. , Pina, J.A. and Navarro , L (1997b) Efficient search for new resistant genotypes to the citrus tristeza closterovirus in the orange subfamily Aurantioideae . Theor. Appl. Genetics, 95, 1282–1288. [Google Scholar]
- Moreno, P. and Guerri, J. (1997) Variability of citrus tristeza closterovirus (CTV): methods for differentiating isolates In: Filamentous Viruses of Woody Plants (Monette P., ed.), pp. 97–107. Trivandrum, India: Research Signpost. [Google Scholar]
- Moreno, P. , Guerri, J. and Muñoz, N. (1990) Identification of Spanish strains of citrus tristeza virus (CTV) by analysis of double‐stranded RNAs (dsRNA). Phytopathology, 80, 477–482. [Google Scholar]
- Moreno, P. , Guerri, J. , Ballester‐Olmos, J.F. , Albiach, R. and Martínez, M.E. (1993) Separation and interference of strains from a citrus tristeza virus isolate evidenced by biological activity and double‐stranded RNA (dsRNA) analysis. Plant Pathol. 42, 35–41. [Google Scholar]
- Müller, G.W. , Costa, A.S. , Castro, J.L. and Guirado, N. (1988) Results from preimmunization tests to control the Capao Bonito strain of tristeza In: Proceedings of the 10th Conference of the International Organization of Citrus Virologists (Timmer L.W., Garnsey S.M. and Navarro L., eds), pp. 82–85. Riverside, CA: IOCV. [Google Scholar]
- Müller, G.W. , Rodriguez, O. and Costa, A.S. (1968) A tristeza virus complex severe to sweet orange varieties. In: Proceedings of the 4th Conference of the International Organization of Citrus Virologists (Childs J.F.L., ed.), pp. 64–71. Gainesville, FL: University of Florida Press. [Google Scholar]
- Narváez, G. , Skander, B.S. , Ayllón, M.A. , Rubio, L. , Guerri, J. and Moreno, P. (2000) A new procedure to differentiate citrus tristeza virus isolates by hybridisation with digoxigenin‐labelled cDNA probes. J. Virol. Methods, 85, 83–92. [DOI] [PubMed] [Google Scholar]
- Navarro, L. , Pina, J.A. , Juárez, J. , Ballester‐Olmos, J.F. , Arregui, J.M. , Ortega, C. , Navarro, A. , Duran‐Vila, N. , Guerri, J. , Moreno, P. , Cambra, M. , Medina, A. and Zaragoza, S. (2002) The Citrus Variety Improvement program in Spain in the period 1975–2001 In: Proceedings of the 15th Conference of the International Organization of Citrus Virologists (Duran‐Vila N., Milne R.G. and Da Graça J.V., eds), pp. 306–316. Riverside, CA: IOCV. [Google Scholar]
- Navas‐Castillo, J. , Albiach‐Martí, M.R. , Gowda, S. , Hilf, M.E. , Garnsey, S.M. and Dawson, W.O. (1997) Kinetics of accumulation of Citrus tristeza virus RNAs. Virology, 228, 92–97. [DOI] [PubMed] [Google Scholar]
- Nei, M. (1987) Molecular Evolutionary Genetics. New York: Columbia University Press. [Google Scholar]
- Nikolaeva, O.V. , Karasev, A.V. , Garnsey, S.M. and Lee, R.F. (1998) Serological differentiation of Citrus tristeza virus isolates causing stem pitting in sweet orange. Plant Dis. 82, 1276–1280. [DOI] [PubMed] [Google Scholar]
- Nolasco, G. , Deblas, C. , Torres, V. and Ponz, F. (1993) A method combining immunocapture and PCR amplification in a microtiter plate for the detection of plant‐viruses and subviral pathogens. J. Virol. Methods, 45, 201–218. [DOI] [PubMed] [Google Scholar]
- Norman, P.A. and Grant, T.J. (1956) Transmission of tristeza virus by aphids. Proc. Fla. Hort. Soc. 69, 38–42. [Google Scholar]
- Olivares‐Fuster, O. , Fleming, G.H. , Albiach‐Martí, M.R. , Gowda, S. , Dawson, W.O. and Grosser, J.W. (2003) Citrus tristeza virus (CTV) resistance in transgenic citrus based on virus challenge of protoplasts. In Vitro Cell. Dev. Biol.—Plant, 39, 567–572. [Google Scholar]
- Olmos, A. , Cambra, M. , Esteban, O. , Gorris, M.T. and Terrada, E. (1999) New device and method for capture, reverse transcription and nested PCR in a single closed tube. Nucleic Acids Res. 27, 1564–1565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pappu, H.R. , Karasev, A.V. , Anderson, E.J. , Pappu, S.S. , Hilf, M.E. , Febres, V.J. , Eckloff, R.M.G. , McCaffery, M. , Boyko, V. , Gowda, S. , Dolja, V.V. , Koonin, E.V. , Gumpf, D.J. , Cline, K. , Garnsey, S.M. , Dawson, W.O. , Lee, R.F. and Niblett, C.L. (1994) Nucleotide sequence and organization of eight 3′ open reading frames of citrus tristeza closterovirus genome. Virology, 199, 35–46. [DOI] [PubMed] [Google Scholar]
- Pappu, H.R. , Manjunath, K.L. , Lee, R.F. and Niblett, C.L. (1993). Molecular characterization of a structural epitope that is largely conserved among severe isolates of a plant virus. Proc. Natl Acad. Sci. USA, 90, 3641–3644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peremyslov, V.V. , Pan, Y.W. and Dolja, V.V. (2004) Movement protein of a closterovirus is a type III integral transmembrane protein localized to the endoplasmic reticulum. J. Virol. 78, 3704–3709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Permar, T.A. , Garnsey, S.M. , Gumpf, D.J. and Lee, R.F. (1990) A monoclonal antibody which discriminates strains of citrus tristeza virus. Phytopathology, 80, 224–228. [Google Scholar]
- Powell, C.A. , Pelosi, R.R. , Rundell, P.A. and Cohen, M. (2003) Breakdown of cross‐protection of grapefruit from decline‐inducing isolates of Citrus tristeza virus following introduction of the brown citrus aphid. Plant Dis. 87, 1116–1118. [DOI] [PubMed] [Google Scholar]
- Price, W.C. (1966) Flexuous rods in phloem cells of lime plants infected with citrus tristeza virus. Virology, 29, 285–294. [DOI] [PubMed] [Google Scholar]
- Raccah, B. , Loebenstein, G. and Bar‐Joseph, M. (1976) Transmission of citrus tristeza virus by melon aphid. Phytopathology, 66, 1102–1104. [Google Scholar]
- Raccah, B. , Loebenstein, G. and Singer, S. (1980) Aphid‐transmissibility variants of citrus tristeza virus in infected citrus trees. Phytopathology, 70, 89–93. [Google Scholar]
- Rai, M. (2006) Refinement of the Citrus tristeza virus resistance gene (Ctv) positional map in Poncirus trifoliata and generation of transgenic grapefruit (Citrus paradisi) plant lines with candidate resistance genes in this region. Plant Mol. Biol. 61, 399–414. [DOI] [PubMed] [Google Scholar]
- Ratcliff, F. , Harrison, B.D. and Baulcombe, D.C. (1997) A similarity between viral defense and gene silencing in plants. Science, 276, 1558–1560. [DOI] [PubMed] [Google Scholar]
- Ratcliff, F. , MacFarlane, S. and Baulcombe, D.C. (1999) Gene silencing without DNA. RNA‐mediated cross‐protection between viruses. Plant Cell, 11, 1207–1216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rocha‐Peña, M.A. , Lee, R.F. , Lastra, R. , Niblet, C.L. , Ochoa‐Corona, F.M. , Garnsey, S.M. and Yokomi, R.K. (1995) Citrus tristeza virus and its vector Toxoptera citricida . Plant Dis. 79, 437–445. [Google Scholar]
- Roistacher, C.N. (1981) A blueprint for disaster. I. The history of seedling yellows disease. Citrograph, 67, 4–24. [Google Scholar]
- Roistacher, C.N. (1991) Graft‐Transmissible Diseases of Citrus. Handbook for Detection and Diagnosis. Rome: FAO. [Google Scholar]
- Roistacher, C.N. and Bar‐Joseph, M. (1984) Transmission of tristeza and seedling yellows tristeza virus by Aphis gossypii from sweet orange, grapefruit and lemon to Mexican lime, grapefruit and lemon In: Proceedings of the 9th Conference of the International Organization of Citrus Virologists (Garnsey S.M., Timmer L.W. and Dodds J.A., eds), pp. 9–18. Riverside, CA: IOCV. [Google Scholar]
- Roistacher, C.N. and Dodds, J. A. (1993) Failure of 100 mild Citrus tristeza virus isolates from California to cross protect against a challenge by severe sweet orange stem pitting isolates In: Proceedings of the 12th Conference of the International Organization of Citrus Virologists (Moreno P., Da Graça J. and Timmer L.W., eds), pp. 100–107. Riverside, CA: IOCV. [Google Scholar]
- Roistacher, C.N. and Moreno, P. (1991) The worlwide threat from destructive isolates of citrus tristeza virus. A review In: Proceedings of the 11th Conference of the International Organization of Citrus Virologists (Brlansky R.H., Lee R.F. and Timmer L.W., eds), pp. 7–19. Riverside, CA: IOCV. [Google Scholar]
- Roistacher, C.N. , Nauer, E.M. , Kishaba, A. and Calavan, E.C. (1980) Transmission of citrus tristeza virus by Aphis gossypii reflecting changes in virus transmissibility in California In: Proceedings of the 8th Conference of the International Organization of Citrus Virologists (Calavan E.C., Garnsey S.M. and Timmer L.W., eds), pp. 76–82. Riverside, CA: IOCV. [Google Scholar]
- Roistacher, C.N. , Dodds, J.A. and Bash, J.A. (1988) Cross protection against citrus tristeza seedling yellows and stem pitting viruses by protective isolates developed in greenhouse plants In: Proceedings of the 10th Conference of the International Organization of Citrus Virologists (Timmer L.W., Garnsey S.M. and Navarro L., eds), pp. 91–100. Riverside, CA: IOCV. [Google Scholar]
- Román, M.P. , Cambra, M. , Juárez, J. , Moreno, P. , Duran‐Vila, N. , Tanaka, F.A.O. , Alves, E. , Kitajima, E.W. , Yamamoto, P.T. , Bassanezi, R.B. , Teixeira, D.C. , Junior, W.C.J. , Ayres, A.J. , Gimenes‐Fernandes, N. , Rabenstein, F. , Girotto, L.F. and Bové, J.M. (2004) Sudden death of citrus in Brazil: a graft‐transmissible bud union disease. Plant Dis. 88, 453–467. [DOI] [PubMed] [Google Scholar]
- Rosner, A. and Bar‐Joseph, M. (1984) Diversity of citrus tristeza virus strains indicated by hybridization with cloned cDNA sequences. Virology, 139, 189–193. [DOI] [PubMed] [Google Scholar]
- Rosner, A. , Lee, R.F. and Bar‐Joseph, M. (1986) Differential hybridization with cloned cDNA sequences for detecting a specific isolate of citrus tristeza virus. Phytopathology, 76, 820–824. [Google Scholar]
- Roy, A., Ramachandran, P. and Brlansky, R.H. (2003) Grouping and comparison of Indian citrus tristeza virus isolates based on coat protein gene sequences and restriction analysis patterns. Arch. Virol. 148, 707–722. [DOI] [PubMed] [Google Scholar]
- Roy, G. , Sudarshana, M.R. , Ullman, D.E. , Ding, S.‐W. , Dandekar, A.M. and Falk, B.W. (2006) Chimeric cDNA sequences from Citrus tristeza virus confer RNA silencing‐mediated resistance in transgenic Nicotiana benthamiana plants. Phytopathology, 96, 819–827. [DOI] [PubMed] [Google Scholar]
- Rubio, L. , Ayllón, M.A. , Guerri, J. , Pappu, H. , Niblett, C. and Moreno, P. (1996) Differentiation of citrus tristeza closterovirus (CTV) isolates by single‐strand conformation polymorphism analysis of the coat protein gene. Ann. Appl. Biol. 129, 479–489. [Google Scholar]
- Rubio, L. , Soong, J. , Kao, J. and Falk, B.W. (1999) Geographic distribution and molecular variation of isolates of three whitefly‐borne closteroviruses of cucurbits: lettuce infectious yellows virus, cucurbit yellow stunting disorder virus, and beet pseudo‐yellows virus. Phytopathology, 89, 707–711. [DOI] [PubMed] [Google Scholar]
- Rubio, L. , Ayllón, M.A. , Kong, P. , Fernández, A. , Polek, M.L. , Guerri, J. , Moreno, P. and Falk, B.W. (2001) Genetic variation of Citrus tristeza virus isolates from California and Spain: evidence for mixed infections and recombination. J. Virol. 75, 8054–8062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruiz‐Ruiz, S. , Moreno, P. , Guerri, J. and Ambrós, S. (2006) The complete nucleotide sequence of a severe stem pitting isolate of Citrus tristeza virus from Spain: comparison with isolates from different origins. Arch. Virol. 151, 387–398. [DOI] [PubMed] [Google Scholar]
- Ruiz‐Ruiz, S. , Moreno, P. , Guerri, J. and Ambrós, S. (2007) A real‐time RT‐PCR assay for detection and absolute quantitation of Citrus tristeza virus in different plant tissues. J. Virol. Methods 145, 96–105. [DOI] [PubMed] [Google Scholar]
- Sambade, A. , Ambrós, S. , López, C. , Ruiz‐Ruiz, S. , Hermoso de Mendoza, A. , Flores, R. , Guerri, J. and Moreno, P. (2007) Preferential accumulation of severe variants of Citrus tristeza virus in plants co‐inoculated with mild and severe variants. Arch. Virol. 152, 1115–1126. [DOI] [PubMed] [Google Scholar]
- Sambade, A. , Rubio, L. , Garnsey, S.M. , Costa, N. , Müller, G.W. , Peyrou, M. , Guerri, J. and Moreno, P. (2002) Comparison of the viral RNA populations of pathogenically distinct isolates of Citrus tristeza virus: application to monitoring cross protection. Plant Pathol. 51, 257–265. [Google Scholar]
- Sambade, A. , López, C. , Rubio, L. , Flores, R. , Guerri, J. and Moreno, P. (2003) Polymorphism of a specific region in the gene p23 of Citrus tristeza virus allows differentiation between mild and severe isolates. Arch. Virol. 148, 2281–2291. [DOI] [PubMed] [Google Scholar]
- Saponari, M. , Manjunath, K. and Yokomi, R.K. (2007) Quantitative detection of Citrus tristeza virus in citrus and aphids by real‐time reverse transcription‐PCR (TaqMan®). J. Virol. Methods, doi: 10.1016/j.jviromet.2007.07.026.. [DOI] [PubMed] [Google Scholar]
- Satyanarayana, T. , Gowda, S. , Ayllón, M.A. , Albiach‐Martí, M.R. and Dawson, W.O. (2002a) Mutational analysis of the replication signals in the 3′‐non translated region of Citrus tristeza virus. Virology, 300, 140–152. [DOI] [PubMed] [Google Scholar]
- Satyanarayana, T. , Gowda, S. , Ayllón, M.A. , Albiach‐Martí, M.R. , Rabindram, R. and Dawson, W.O. (2002b) The p23 protein of Citrus tristeza virus controls asymmetrical RNA accumulation. J. Virol. 76, 473–483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Satyanayanana, T. , Gowda, S. , Boyko, V.P. , Albiach‐Martí, M.R. , Mawassi, M. , Navas‐Castillo, J. , Karasev, A.V. , Dolja, V. , Hilf, M.E. , Lewandowsky, D.J. , Moreno, P. , Bar‐Joseph, M. , Garnsey, S.M. and Dawson, W.O. (1999) An engineered closterovirus RNA replicon and analysis of heterologous terminal sequences for replication. Proc. Natl Acad. Sci. USA, 96, 7433–7438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Satyanayanana, T. , Gowda, S. , Mawassi, M. , Albiach‐Martí, M.R. , Ayllón, M.A. , Robertson, C. , Garnsey, S.M. and Dawson, W.O. (2000) Closterovirus encoded HSP70 homolog and p61 in addition to both coat proteins function in efficient virion assembly. Virology, 278, 253–265. [DOI] [PubMed] [Google Scholar]
- Satyanayanana, T. , Bar‐Joseph, M. , Mawassi, M. , Albiach‐Martí, M.R. , Ayllón, M.A. , Gowda, S. , Hilf, M.E. , Moreno, P. , Garnsey, S.M. and Dawson, W.O. (2001) Amplification of Citrus tristeza virus from a cDNA clone and infection of citrus trees. Virology, 280, 87–96. [DOI] [PubMed] [Google Scholar]
- Satyanayanana, T. , Gowda, S. , Ayllón, M.A. and Dawson, W.O. (2004) Closterovirus bipolar virion: evidence for initiation of assembly by minor coat protein and its restriction to the genomic RNA 5′ region. Proc. Natl Acad. Sci. USA, 101, 799–804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider, H. (1959) The anatomy of tristeza‐virus‐infected citrus In: Citrus Virus Diseases (Wallace J.M., ed.), pp. 73–84. Berkeley, CA: University of California Division of Agricultural Sciences. [Google Scholar]
- Sentandreu, V. , Castro, J.A. , Ayllón, M.A. , Rubio, L. , Guerri, J. , González‐Candelas, F. , Moreno, P. and Moya, A. (2006) Evolutionary analysis of genetic variation observed in Citrus tristeza virus (CTV) after host passage. Arch. Virol. 151, 875–894. [DOI] [PubMed] [Google Scholar]
- Suastika, G. , Natsuaki, T. , Terui, H. , Kano, T. Ieki, H. , and Okuda, S. (2001) Nucleotide sequence of Citrus tristeza virus seeding yellows isolate. J. Gen. Plant Pathol. 67, 73–77 [Google Scholar]
- Tian, T. , Rubio, L. , Yeh, H.H. , Crawford, B. and Falk, B.W. (1999) Lettuce infectious yellows virus: In vitro acquisition analysis using partially purified virions and the whitefly, Bemisia tabaci . J. Gen. Virol. 80, 1111–1117. [DOI] [PubMed] [Google Scholar]
- Timmer, L.W. , Garnsey, S.M. and Graham, J.H. (2000) Compendium of Citrus Diseases. St Paul, MN: APS Press. [Google Scholar]
- Van Vuuren, S.P. , Collins, R.P. and da Graça, J.V. (1993) Evaluation of citrus tristeza virus isolates for cross protection of grapefruit in South Africa. Plant Dis. 77, 24–28. [Google Scholar]
- Varma, P.M. , Rao, D.G. and Capoor, S.P. (1965) Transmission of tristeza virus by Aphis craccivora Koch and Dactynotus jaceae (L.). Indian J. Entomol. 27, 67–71 [Google Scholar]
- Vela, C. , Cambra, M. , Cortés, E. , Moreno, P. , Miguet, S.G. , Pérez de San Román, C. and Sanz, A. (1986) Production and characterisation of monoclonal antibodies specific for citrus tristeza virus and their use for diagnosis. J. Gen. Virol. 67, 91–96. [Google Scholar]
- Vives, M.C. , Rubio, L. , López, C. , Navas‐Castillo, J. , Albiach‐Martí, M.R. , Dawson, W.O. , Guerri, J. , Flores, R. and Moreno, P. (1999) The complete genome sequence of the major component of a mild citrus tristeza virus isolate. J. Gen. Virol. 80, 811–816. [DOI] [PubMed] [Google Scholar]
- Vives, M.C. , Rubio, L. , Sambade, A. , Mirkov , Moreno, P. and Guerri, J. (2005) Evidence of multiple recombination events between two RNA sequence variants within a Citrus tristeza virus isolate. Virology, 331, 232–237. [DOI] [PubMed] [Google Scholar]
- Voinnet, O. (2005) Induction and suppression of RNA silencing: insights from viral infections. Nature Gen. Rev. 6, 206–220. [DOI] [PubMed] [Google Scholar]
- Wallace, J.M. (1978) Virus and viruslike diseases In: The Citrus Industry (Reuther W., Calavan E.C. and Carman G.E., eds), pp. 67–184. Berkeley, CA: University of California, Division of Agricultural Sciences. [Google Scholar]
- Wallace, J.M. and Drake, R.J. (1972) Studies on recovery of citrus plants from seedling yellows and the resulting protection against reinfection In: Proceedings of the 5th Conference of the International Organization of Citrus Virologists (Price W.C., ed.), pp. 127–136. Gainesville, FL: University of Florida Press. [Google Scholar]
- Weathers, L.G. and Hartung, M.K. (1964) Transmission of citrus viruses by dodder, Cuscuta subinclusa . Plant Dis. Reptr. 48, 102–103. [Google Scholar]
- Webber, H.J. (1943) The ‘Tristeza’ disease of sour‐orange rootstock. Amer. Soc. Hort. Sci. 43, 160–168. [Google Scholar]
- Yang, G. , Mawassi, M. , Gofman, R. , Gafny, R. and Bar‐Joseph, M. (1997) Involvement of a subgenomic mRNA in the generation of a variable population of defective citrus tristeza virus molecules. J. Virol. 71, 9800–9802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang, G. , Che, X. , Gofman, R. , Ben Shalom, Y. , Piestun, D. , Gafny, R. , Mawassi, M. and Bar‐Joseph, M. (1999) D‐RNA molecules associated with subisolates of the VT strain of citrus tristeza virus which induce different seedling‐yellows reactions. Virus Genes, 19, 5–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang, Z.‐N. , Mathews, D.M. , Dodds, J.A. and Mirkov, T.E. (1999) Molecular characterization of an isolate of citrus tristeza virus that causes severe symptoms in sweet orange. Virus Genes, 19, 11–142. [DOI] [PubMed] [Google Scholar]
- Yang, Z.‐N. , Ye, X.‐R. , Molina, J. , Roose, M.L. and Mirkov, T.E. (2003) Sequence analysis of a 282‐kilobase region surrounding the Citrus tristeza virus resistance gene (ctv) locus in Poncirus trifoliata (L.) Raf. Plant Physiol. 131, 482–492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yokomi, R.K. and Garnsey, S.M. (1987) Transmision of citrus tristeza virus by A. gossypii and A. citricola in Florida. Phytophylactica, 19, 169–172. [Google Scholar]
- Yokomi, R.K. , Lastra, R. , Stoetzel, M.B. , Damsteegt, V.D. , Lee, R.F. , Garnsey, S.M. , Gottwald, T.R. , Rocha‐Peña, M.A. and Niblett, C.L. (1994) Establishment of the brown citrus aphid (Homoptera, Aphididae) in Central‐America and the Caribbean Basin and transmission of citrus tristeza virus. J. Econ. Entomol. 87, 1078–1085. [Google Scholar]
- Yoshida, T. (1985) Inheritance of susceptibility to citrus tristeza virus in trifoliate orange (Poncirus trifoliata Raf.). Bull. Fruit Tree Res. Sta. 12, 17–25. [Google Scholar]
- Yoshida, T. (1993) Inheritance of immunity to citrus tristeza virus of trifoliate orange in some citrus intergeneric hybrids. Bull. Fruit Tree Res. Sta. 25, 33–43 [Google Scholar]
- Yoshida, T. (1996) Graft compatibility of Citrus with plants in the Aurantoidea and their susceptibility to citrus tristeza virus. Plant Dis. 80, 414–417. [Google Scholar]
- Zaragoza, S. (2007) Aproximación a la historia de los cítricos. Origen, dispersión y evolución de su uso y cultivo. PhD thesis, Universidad Politécnica de Valencia. [Google Scholar]