

SUMMARY
Long considered intractable organisms by fungal genetic research standards, the oomycetes have recently moved to the centre stage of research on plant–microbe interactions. Recent work on oomycete effector evolution, trafficking and function has led to major conceptual advances in the science of plant pathology. In this review, we provide a historical perspective on oomycete genetic research and summarize the state of the art in effector biology of plant pathogenic oomycetes by describing what we consider to be the 10 most important concepts about oomycete effectors.
INTRODUCTION AND HISTORICAL PERSPECTIVE
Oomycetes, such as Phytophthora spp. and downy mildews, are a unique group of fungal‐like eukaryotic pathogens that cause some of the most destructive and economically significant plant diseases in the world (Erwin and Ribeiro, 1996; Kamoun, 2003; Lamour and Kamoun, 2009). Long considered intractable organisms by fungal genetic research standards, the oomycetes have recently emerged as key experimental systems in research on plant–microbe interactions. Experimental data on oomycete effector evolution, trafficking and function have led to major conceptual advances in the science of plant pathology (2009, 2008, 2006; Kamoun, 2006, 2007; Morgan and Kamoun, 2007; Tyler, 2009). However, historically, genetic research on oomycetes has faced its ups and downs. The low point was reached in the early 1980s when David Shaw, University of Wales, Bangor, UK, summed up the general feeling by declaring the oomycetes to be a ‘fungal geneticist's nightmare’ (Shaw, 1983).
In 1984, 1 year after David Shaw's gloomy quip, Brian Staskawicz, Doug Dahlbeck and Noel Keen reported the first cloning of a plant pathogen avirulence gene from the bacterium Pseudomonas syringae pv. glycinea (Staskawicz et al., 1984). This landmark event ushered in a golden age for research into plant–microbe interactions during which bacteria and a handful of fungi became the organisms of choice for molecular studies on host specificity and disease resistance (see other reviews in this issue). However, prior to this period, several leading laboratories used Phytophthora as a model system, resulting in significant conceptual developments that set the stage for the advances achieved later using molecular genetics tools. In the USA, the late Noel Keen at the University of California, Riverside, CA, developed the concept of host‐specific elicitors based on work with the soybean pathogen Phytophthora sojae (Collmer and Gold, 2007; Keen, 1975). Other influential researchers, such as Peter Albersheim, Noriyuke Doke and Klaus Hahlbrock, used Phytophthora to perform important experiments that further cemented the view that plant pathogens produce elicitor molecules that trigger plant defences (Albersheim and Valent, 1978; Ayers et al., 1976; Doke, 1983a, 1983b; Ebel et al., 1976; Hahlbrock et al., 1981; Lawton et al., 1983; 1984a, 1984b). Some of these elicitors displayed cultivar‐specific activities and were presumed to be avirulence factors.
The research related to the biochemical nature of the cultivar‐specific elicitors generated a notorious controversy, illustrating the importance of oomycete research in those days (Daly, 1984; Gracen, 1982). Conflicting results on whether the elusive avirulence factor was a proteinaceous or a carbohydrate preparation were obtained using races of P. sojae that differed in virulence on soybean plants carrying the Rps1a resistance gene (Keen, 1975; Wade and Albersheim, 1979). Interestingly, this issue can now be settled thanks to the recent cloning of P. sojae Avr1a in Mark Gijzen's laboratory at Agriculture and Agri‐Food Canada, London, ON (Qutob et al., 2009). Avr1a is a canonical 121‐amino‐acid RXLR effector that is translocated inside soybean cells, where it activates Rps1a and triggers cell death and resistance (Qutob et al., 2009) (see later for more on RXLR effectors).
Research on oomycetes waned and did not remain at the forefront of the field in most of the 1980s and 1990s, given the relative facility of cloning and manipulating avirulence genes from plant pathogenic bacteria and some phytopathogenic fungi. Even Noel Keen dropped P. sojae as an experimental system and shifted his focus entirely to bacterial plant pathogens starting from the early 1980s (Collmer and Gold, 2007). A resurgence of oomycete research occurred in the mid‐1990s and culminated in the emergence of genomics and the cloning of avirulence genes, as well as other classes of effectors (some of the history on the impact of genomics is summarized in Govers and Gijzen, 2006; Turner, 2008). Oomycetes moved on from being a ‘fungal geneticist's nightmare’ to becoming a ‘genomicist's dream’. The identification of conserved host translocation motifs in oomycete effectors was a defining breakthrough. It enabled the implementation of data mining strategies to develop genome‐wide effector catalogues (Haas et al., 2009; Jiang et al., 2008; Tyler et al., 2006; Win et al., 2007), a feat that remains problematic for most true fungi (Kamoun, 2007). Comparative transcriptomics and genomics of oomycetes added another dimension to our understanding of effector gene regulation, evolution and genome organization (Haas et al., 2009; Jiang et al., 2008; Tyler et al., 2006; Win et al., 2007), and the application of rapid cost‐effective sequencing and gene‐chip expression profiling technologies promises to greatly impact our understanding of the diversity of pathogenicity mechanisms in oomycetes (Lamour et al., 2007).
In this review, we outline the state of the art in effector biology of plant pathogenic oomycetes. We list and describe what we consider to be the 10 most important concepts about oomycete effectors. For more detailed and complementary perspectives on the topic, we refer readers to several informative reviews that have been published recently (2009, 2008, 2006; Kamoun, 2006, 2007; Morgan and Kamoun, 2007; Tyler, 2009).
1. OOMYCETE EFFECTORS LOCALIZE TO DIFFERENT SITES IN HOST TISSUE
Oomycete effector proteins target different sites in host plant tissue (Fig. 1) (Kamoun, 2006, 2007). Some effectors act in the extracellular space, where they interfere with apoplastic plant proteins involved in pathogen defence. These include inhibitors of plant hydrolases, such as glucanases, as well as serine and cysteine proteases (Damasceno et al., 2008; Rose et al., 2002; 2005, 2004, 2007). Other secreted proteins, such as a variety of small cysteine‐rich proteins, are also thought to function in the apoplast, but their effector activities remain largely unknown (Kamoun, 2006). Other oomycete effectors, namely members of the RXLR and Crinkler (CRN) families, translocate inside host cells (Kamoun, 2006, 2007). In fact, all known oomycete effectors with avirulence activities are host translocated (cytoplasmic) effectors of the RXLR class. They are recognized inside plant cells by so‐called resistance (R) proteins, intracellular immune receptors of the nucleotide‐binding leucine‐rich repeat (NB‐LRR) family, resulting in the induction of hypersensitive cell death and immunity.
Figure 1.
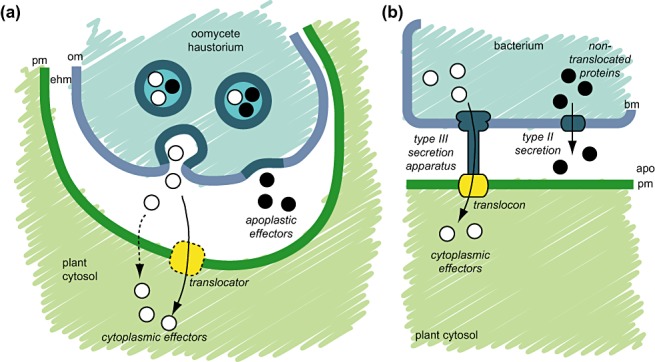
Oomycete and bacterial pathogen effector proteins target different sites in host plant tissue. Schematic view of effector secretion by an oomycete haustorium (a) and a bacterial plant pathogen (b). Apoplastic effectors (black circles) are secreted into the extracellular space, where they interfere with apoplastic plant defences. Cytoplasmic effectors (white circles) translocate inside host cells, thereby crossing two membranes, one from the pathogen and another from the host. The eukaryotic mechanism of host translocation remains unclear, but may include pathogen‐encoded translocator proteins (a). In bacteria, host translocation is achieved by the type III secretion system which involves dedicated translocon proteins (yellow), whereas type II secretion delivers proteins to the apoplast (b). apo, apoplast; bm, bacterial membrane; ehm, extrahaustorial matrix; om, oomycete membrane; pm, plasma membrane.
2. OOMYCETE EFFECTORS ARE MODULAR PROTEINS
Both apoplastic and cytoplasmic oomycete effectors are modular proteins (Fig. 2) (Kamoun, 2006, 2007). Apoplastic effectors contain N‐terminal signal peptides for secretion, followed by C‐terminal effector module(s), but are not known to carry additional host targeting signals (Damasceno et al., 2008; 2005, 2004, 2007). Several Kazal‐like serine protease inhibitors contain multiple inhibitor domains with different degrees of specificity towards serine proteases. In Phytophthora infestans EPI1, the first of two Kazal‐like domains is an efficient inhibitor of the tomato subtilisin‐like serine protease P69B (Tian and Kamoun, 2005), whereas, in EPI10, the second of three Kazal‐like domains is the P69B inhibitor (Tian et al., 2005). The targets of the other domains of EPI1 and EPI10 remain unknown (Tian and Kamoun, 2005). Cytoplasmic effectors consist of an N‐terminal region involved in secretion and translocation inside host cells and a C‐terminal domain carrying the biochemical effector activity (Kamoun, 2006, 2007; Morgan and Kamoun, 2007). These effectors are characterized by conserved motifs in the N‐terminal region following the signal peptide. The RXLR effectors are defined by the amino acid sequence Arg‐X‐Leu‐Arg (where X is any residue) that designates a domain involved in translocation inside host cells (Dou et al., 2008a; Whisson et al., 2007). This domain is functionally equivalent to the PEXEL host translocation domain of the malaria parasites Plasmodium spp. (Bhattacharjee et al., 2006; Grouffaud et al., 2008; Haldar et al., 2006). Higher levels of polymorphisms and signatures of positive (adaptive) selection have been observed in the C‐terminal half of RXLR effectors, consistent with the view that this region executes the effector activity inside plant cells and coevolves with host factors (Fig. 3) (2004, 2008; Rehmany et al., 2005; Win et al., 2007). The CRN effectors also contain N‐terminal sequence domains, including the conserved LXLFLAK motif, which are implicated in host translocation (Haas et al., 2009; Win et al., 2007) (E. Huitema et al., unpublished results). In addition, most CRN proteins carry a DWL domain after the LXLFLAK motif. This second N‐terminal domain ends with another conserved motif, the HVLVXXP motif. There is strong evidence of recombination among CRNs, particularly after the HVLVXXP motif, resulting in an extraordinary diversity of C‐terminal domains with 36 divergent amino acid sequences attached to the DWL domain in P. infestans (Haas et al., 2009). This suggests an evolutionary mechanism similar to the C‐terminal reassortment reported for type III‐secreted effectors of phytopathogenic bacteria (Stavrinides et al., 2006).
Figure 2.

Oomycete effectors are modular. All known effectors carry N‐terminal signal peptides for secretion (yellow). Cytoplasmic effectors, in addition, have conserved motifs in their N‐termini (RXLR or LXLFLAK) that are required for host translocation. The C‐terminal domain carries the module with biochemical effector activity.
Figure 3.
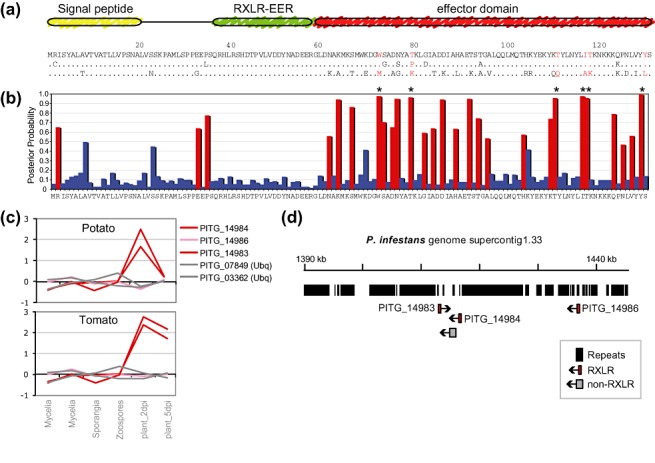
RXLR effector genes typically show adaptive selection in their C‐termini, are induced in plants and occur in clusters in the genome. The figure depicts the features of a representative RXLR gene cluster of Phytophthora infestans (Haas et al., 2009). (a) Domain structure and sequence variability of three paralogues of RXLR family 6 of P. infestans (PITG_14983, PITG_14984 and PITG_14986, top to bottom; based on Haas et al., 2009). Residues with evidence of positive selection (b) are highlighted in red. Dots in the alignment represent identical amino acid residues. (b) Positive selection analyses based on the methods described in Win et al. (2007). Posterior probabilities (blue, red) for the site class with expected ω value >1 (ω= 21.07706) and P= 0.16379 estimated under the model M8 in the paml program (http://abacus.gene.ucl.ac.uk/software/paml.html). Positively selected sites are shown in red. Asterisks label residues with P > 95%. (c) Relative oligonucleotide microarray expression levels at different developmental stages and during infection of potato and tomato plants 2 and 5 days post‐inoculation (dpi). Two RXLR genes are induced in plants (red lines) and one is not (pink line). Two constitutive ubiquitin genes (Ubq) are shown as controls (grey lines) (see Haas et al., 2009 for experimental details). (d) Genome browser view of ∼55‐kbp region of the P. infestans genome (supercontig 1.33) containing the cluster of related RXLR genes. The high content of repetitive sequences is evidenced by the black bars (repeats).
3. SOME RXLR EFFECTOR PROTEINS ARE ABLE TO ENTER PLANT CELLS IN THE ABSENCE OF THE PATHOGEN
How the N‐termini of RXLR effectors enable translocation inside host cells is a challenging and important question. Machinery for the transport of RXLR effectors into host cells could be encoded by the pathogen, the host or both (Birch et al., 2008; Morgan and Kamoun, 2007). Dou et al. (2008b) reported that RXLR effectors of P. sojae can enter plant cells in the absence of the pathogen. First, they showed that the RXLR motif of P. sojae Avr1b is required for cell death induction when a full‐length construct with the signal peptide is expressed in soybean cells, presumably to enable re‐entry of the protein following secretion. Second, recombinant fusion proteins of the RXLR domain and green fluorescent protein can directly enter soybean root cells. These experiments suggest that oomycete effectors do not require a pathogen‐derived machinery to traffic inside host cells and exclusively exploit host‐derived molecules. However, these findings require further support, given that some of the results obtained by Dou et al. (2008b) deviate from results obtained with other experimental systems (Bos et al., 2006; Oh et al., 2009). In addition, a parasite‐derived translocon mediating the host translocation of PEXEL effectors was reported in Plasmodium, suggesting an active parasite machinery for host targeting (de Koning‐Ward et al., 2009). The PEXEL motif is similar in sequence, position and function to the oomycete RXLR motif (Bhattacharjee et al., 2006; Haldar et al., 2006; Grouffaud et al., 2008).
4. HUNDREDS OF EFFECTOR GENES OCCUR IN OOMYCETE GENOMES
The occurrence of conserved domains and motifs within the N‐termini of RXLR and CRN effectors enabled the implementation of computational approaches to generate genome‐wide catalogues of effector genes from oomycete species, such as Hyaloperonospora arabidopsidis (previously H. parasitica), P. infestans, P. ramorum and P. sojae (Haas et al., 2009; Jiang et al., 2008; Tyler et al., 2006; Win et al., 2007). The effector secretome of these pathogens is highly complex, with several hundred proteins predicted to function as effectors (Haas et al., 2009; Jiang et al., 2008; Tyler et al., 2006; Win et al., 2007). With 563 predicted genes, the number of RXLR genes in P. infestans is almost double that in P. sojae (335) and P. ramorum (309) (Haas et al., 2009). The CRN‐like genes form the most expanded family of putative effectors in P. infestans (196 genes and 255 pseudogenes) relative to P. sojae (100 and 102) and P. ramorum (19 and 42) (Haas et al., 2009). This dramatic expansion of the number of RXLR and CRN genes in P. infestans correlates with an expansion of their flanking intergenic regions, which averages 4.1, 4.06 and 16.7 kb for P. ramorum, P. sojae and P. infestans RXLR genes, respectively (Haas et al., 2009). RXLR and CRN genes are often found organized in clusters in the Phytophthora genomes (Haas et al., 2009; Jiang et al., 2008; Tyler et al., 2006; Win et al., 2007). Therefore, expansion of the effector gene families may have been driven by nonallelic homologous recombination and tandem gene duplication, favoured by their localization in dynamic regions of the genome that are rich in repeats and transposable elements (Haas et al., 2009).
5. PHYTOPHTHORA SPECIES CARRY DISTINCT REPERTOIRES OF EFFECTOR GENES
Evolutionary analyses of effector genes in oomycete genomes have revealed that they are undergoing accelerated patterns of birth and death evolution, with evidence of extensive gene duplication and gene loss in the genomes of P. sojae, P. ramorum and P. infestans (Haas et al., 2009; Jiang et al., 2008; Tyler et al., 2006; Win et al., 2007). For instance, only 16 of the 563 predicted P. infestans RXLR genes belong to the set of core orthologues with 1 : 1 : 1 orthology relationships between P. infestans, P. ramorum and P. sojae (Haas et al., 2009). Segmental duplication and intraspecific copy number variation are prevalent among the RXLR genes of P. sojae (Qutob et al., 2009). Copy number variation of P. sojae effector genes, such as Avr1a and Avr3a, has an impact on transcript accumulation and virulence phenotypes, highlighting an unusual mechanism of genetic adaptation to host resistance (Qutob et al., 2009). Another P. sojae gene, Avr3c, occurs in a three‐copy segmental duplication, but there is no evidence for copy number variation of this segment among analysed P. sojae strains (Dong et al., 2009). Another effector class, the cystatin protease inhibitor family (EPIC1–4), also exhibits a dynamic evolutionary history. EPIC1 and EPIC2 bind and inhibit several tomato apoplastic proteases (Song et al., 2009; Tian et al., 2007). epiC1 and epiC2 genes are up‐regulated in planta and lack orthologues in P. sojae and P. ramorum, unlike epiC3 and epiC4 genes of P. infestans, which have orthologues in P. sojae and P. ramorum and are not known to encode proteins with inhibitory activities against plant proteases (Song et al., 2009; Tian et al., 2007). epiC1 and epiC2 are part of a cluster of four cystatin‐like paralogues in a region of 70 kb that apparently emerged in the P. infestans lineage by duplication of the epiC3 gene, and subsequently evolved to inhibit tomato defence proteases (Song et al., 2009; Tian et al., 2007).
6. EFFECTORS TYPICALLY SHOW HIGH RATES OF AMINO ACID POLYMORPHISMS AND EVIDENCE OF POSITIVE SELECTION
Because effectors are major virulence determinants that can be recognized by plant R proteins, effector repertoires are thought to be shaped through coevolutionary arms races (Hogenhout et al., 2009; Kamoun, 2007). Consistent with this view, effector genes undergo rapid sequence diversification and their products typically show high rates of amino acid polymorphisms, particularly nonsynonymous substitutions with signatures of positive selection (Fig. 3) (Allen et al., 2004; Liu et al., 2005; Rehmany et al., 2005; Win et al., 2007). Indeed, positive selection is a useful criterion for the identification of candidate effector genes from genome sequences (Liu et al., 2005). In contrast with relaxed selection, positive selection does not typically act uniformly across the length of the protein. The characterization of individual RXLR effector genes, as well as genome‐wide analyses, has shown that positive selection has, for the most part, targeted the C‐terminal effector domain rather than the N‐terminal secretion and host translocation region, confirming the modular structure of these proteins (Allen et al., 2004; Liu et al., 2005; Rehmany et al., 2005; Win et al., 2007).
7. OOMYCETE EFFECTOR GENES SHOW DISTINCT PATTERNS OF EXPRESSION DURING COLONIZATION OF PLANTS
Phytophthora effector genes show diverse patterns of temporal expression during the infection of host plants. Several P. infestans RXLR effectors are transcriptionally induced during pre‐infection stages (germinated cysts) and early stages of infection of potato (Whisson et al., 2007). At least 79 of the predicted P. infestans RXLR effectors, including RXLR effectors with known avirulence activity, such as Avr3a, Avr4 and AVRblb1/ipiO, show a distinct expression pattern with a sharp peak of transcriptional up‐regulation during the biotrophic phase of the infection (until 2–3 days post‐inoculation) (Haas et al., 2009) (Fig. 3c) (see also van West et al., 1998 for a detailed analysis of the expression of ipiO, which was later shown to be AVRblb1 by Vleeshouwers et al., 2008). Some effectors, such as the apoplastic enzyme inhibitors, show the same distinct transient pattern of induction, whereas others, such as the toxin NPP1, are expressed later during the necrotrophic phase (Kanneganti et al., 2006; Qutob et al., 2002). Tight temporal regulation of these effector genes results in distinct stage‐specific expression patterns that reflect the intricate processes of cellular control exhorted by Phytophthora as effectors are deployed during host colonization.
8. SEVERAL EFFECTORS SUPPRESS PLANT IMMUNITY
The virulence activities of oomycete effectors remain largely unknown, but some have been shown to suppress plant immunity. The protease and glucanase inhibitors target apoplastic plant hydrolases that have been previously implicated in immunity (Damasceno et al., 2008; Rose et al., 2002). Recently, a loss‐of‐function mutant of the tomato papain‐like cysteine protease Rcr3, which is inhibited by P. infestans EPIC1 and EPIC2, has been shown to display enhanced susceptibility to P. infestans, suggesting a role in defence (Song et al., 2009). In addition to apoplastic effectors, host‐translocated effectors are also thought to suppress immunity. Phytophthora infestans Avr3a suppresses hypersensitive cell death induced by another P. infestans protein INF1 elicitin (Fig. 4) (Armstrong et al., 2005; 2006, 2009). INF1 is an abundantly secreted P. infestans protein with features of pathogen‐associated molecular patterns (Kawamura et al., 2009; Vleeshouwers et al., 2006). Overexpression of the P. sojae RXLR effector Avr1b confers increased pathogen virulence on compatible soybean plants (Dou et al., 2008a) and suppresses cell death induced by the proapoptotic BAX protein in yeast and plants (Dou et al., 2008a). Hyaloperonospora arabidopsidis ATR13 suppresses callose deposition triggered by Ps. syringae, suggesting that it targets basal defences against diverse pathogens (Sohn et al., 2007). In addition, Sohn et al. (2007) have shown that type III delivery of H. arabidopsidis ATR1 and ATR13 enhances the virulence of the bacterial pathogen Ps. syringae.
Figure 4.
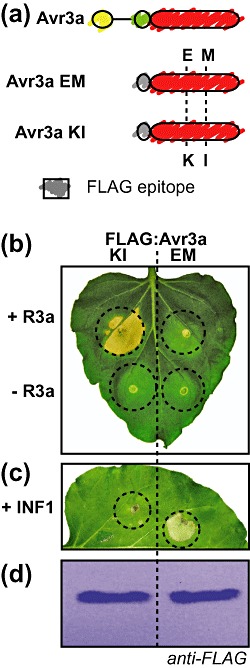
Some effectors suppress plant immunity. The figure describes the RXLR effector AVR3a of Phytophthora infestans as an example of an effector with dual activities: activation of R gene‐specific hypersensitive cell death and suppression of plant immunity (Armstrong et al., 2005, 2006, 2009). Allelic variants of AVR3a, AVR3aKI (containing amino acids K80 and I103) and AVR3aEM (E80 and M103) are shown in (a). The experiments involve the expression of FLAG epitope‐tagged versions of both variants (a) in leaves of Nicotiana benthamiana. The two variants have distinct activities in plants. AVR3aKI but not AVR3aEM triggers hypersensitive cell death in the presence of the resistance gene R3a (b), and suppresses the hypersensitive cell death induced by another P. infestans protein, INF1 elicitin, a secreted P. infestans protein with features of pathogen‐associated molecular patterns (c). (d) FLAG antisera Western blot of leaf extracts, showing that the two variants are equally stable in planta.
9. EFFECTORS ARE USEFUL IN BREEDING, CLONING AND DEPLOYMENT OF DISEASE RESISTANCE GENES
Effectors can be used to identify R proteins and classify their activities into discrete recognition specificities (Ellis et al., 2009; Oh et al., 2009; Vleeshouwers et al., 2008). Effector assays may accelerate the cloning of R genes and help to avoid redundant cloning efforts. In the first study of this kind, RXLR effector genes mined from the genome sequence of P. infestans were employed in high‐throughput in planta expression assays on wild potato germplasms to uncover avirulence activities and accelerate the cloning of their cognate R genes (Vleeshouwers et al., 2008). This approach resulted in the rapid identification and cloning of closely related orthologues of the Solanum bulbocastanum R gene Rpiblb1 from S. stoloniferum and S. papita. More recently, Lokossou et al.( 2009) have reported the cloning of four R genes, Rpi‐blb3, Rpi‐abpt, R2 and R2‐like, from different Solanum germplasms that map to a major late blight resistance locus on potato chromosome IV. All four gene products recognize the RXLR effector PiAvr2 (Lokossou et al., 2009). The availability of effectors that match particular R genes should also prove to be useful in assisting with the deployment of R genes in agriculture. The evaluation of the distribution of effector variants across pathogen populations, coupled with activity assays, can be used to monitor pathogen populations for the potential occurrence or emergence of races that overcome the R genes.
10. THERE IS MUCH THAT WE STILL DO NOT KNOW ABOUT OOMYCETE EFFECTORS
Despite the substantial progress in recent years, there is much that we still do not know about oomycete effectors. What will this top 10 list look like in a few years? We can already note that the oomycete community is moving from an emphasis on linking sequences to phenotypes (Huitema et al., 2004; Oh et al., 2009) to research activities linking the discovered phenotypes to biochemical functions. A mechanistic understanding of how oomycete effectors traffic inside host cells, modify host targets and perturb plant processes is certain to continue to have an impact on the field. The identification of effector molecules from eukaryotic pathogens enables us to draw conceptual parallels between prokaryotic and eukaryotic pathogens and to investigate the extent to which diverse pathogens share virulence strategies and target similar pathways of plant immunity.
ACKNOWLEDGEMENTS
We thank Mark Gijzen for suggestions and for providing background information. Our laboratory is funded by the Gatsby Charitable Foundation, the Biotechnology and Biological Sciences Research Council (BBSRC) and BASF Plant Sciences. SS is supported by a Deutsche Forschungsgemeinschaft (DFG) fellowship, TOB is supported by a Marie Curie Intra‐European fellowship, and MT is supported by a fellowship from the Max Planck Society, the Elite Program for Postdocs of the Landesstiftung Baden‐Württemberg, and the German Science foundation (DFG).
REFERENCES
- Albersheim, P. and Valent, B. (1978) Host–pathogen interactions in plants: plants, when exposed to oligosaccharides of fungal origin, defend themselves by accumulating antibiotics. J. Cell. Biol. 78, 627–643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allen, R.L. , Bittner‐Eddy, P.D. , Grenville‐Briggs, L.J. , Meitz, J.C. , Rehmany, A.P. , Rose, L.E. and Beynon, J.L. (2004) Host–parasite coevolutionary conflict between Arabidopsis and downy mildew. Science, 306, 1957–1960. [DOI] [PubMed] [Google Scholar]
- Allen, R.L. , Meitz, J.C. , Baumber, R.E. , Hall, S.A. , Lee, S.C. , Rose, L.E. and Beynon, J.L. (2008) Natural variation reveals key amino acids in a downy mildew effector that alters recognition specificity by an Arabidopsis resistance gene. Mol. Plant Pathol. 9, 511–523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Armstrong, M.R. , Whisson, S.C. , Pritchard, L. , Bos, J.I. , Venter, E. , Avrova, A.O. , Rehmany, A.P. , Bohme, U. , Brooks, K. , Cherevach, I. , Hamlin, N. , White, B. , Fraser, A. , Lord, A. , Quail, M.A. , Churcher, C. , Hall, N. , Berriman, M. , Huang, S. , Kamoun, S. , Beynon, J.L. and Birch, P.R. (2005) An ancestral oomycete locus contains late blight avirulence gene Avr3a, encoding a protein that is recognized in the host cytoplasm. Proc. Natl. Acad. Sci. USA, 102, 7766–7771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ayers, A.R. , Ebel, J. , Finelli, F. , Berger, N. and Albersheim, P. (1976) Host–pathogen interactions IX. Quantitative assays of elicitor activity and characterization of the elicitor present in the extracellular medium of cultures of Phytophthora megasperma var. sojae . Plant Physiol. 57, 751–759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhattacharjee, S. , Hiller, N.L. , Liolios, K. , Win, J. , Kanneganti, T.D. , Young, C. , Kamoun, S. and Haldar, K. (2006) The malarial host‐targeting signal is conserved in the Irish potato famine pathogen. PLoS Pathog. 2, e50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Birch, P.R. , Armstrong, M. , Bos, J. , Boevink, P. , Gilroy, E.M. , Taylor, R.M. , Wawra, S. , Pritchard, L. , Conti, L. , Ewan, R. , Whisson, S.C. , Van West, P. , Sadanandom, A. and Kamoun, S. (2009) Towards understanding the virulence functions of RXLR effectors of the oomycete plant pathogen Phytophthora infestans . J. Exp. Bot. 60, 1133–1140. [DOI] [PubMed] [Google Scholar]
- Birch, P.R. , Boevink, P.C. , Gilroy, E.M. , Hein, I. , Pritchard, L. and Whisson, S.C. (2008) Oomycete RXLR effectors: delivery, functional redundancy and durable disease resistance. Curr. Opin. Plant Biol. 11, 373–379. [DOI] [PubMed] [Google Scholar]
- Birch, P.R. , Rehmany, A.P. , Pritchard, L. , Kamoun, S. and Beynon, J.L. (2006) Trafficking arms: oomycete effectors enter host plant cells. Trends Microbiol. 14, 8–11. [DOI] [PubMed] [Google Scholar]
- Bos, J.I. , Kanneganti, T.D. , Young, C. , Cakir, C. , Huitema, E. , Win, J. , Armstrong, M.R. , Birch, P.R. and Kamoun, S. (2006) The C‐terminal half of Phytophthora infestans RXLR effector AVR3a is sufficient to trigger R3a‐mediated hypersensitivity and suppress INF1‐induced cell death in Nicotiana benthamiana . Plant J. 48, 165–176. [DOI] [PubMed] [Google Scholar]
- Bos, J.I.B. , Chaparro‐Garcia, A. , Quesada‐Ocampo, L.M. , McSpadden‐Gardener, B.B. and Kamoun, S. (2009) Distinct amino acids of the Phytophthora infestans effector AVR3a condition activation of R3a hypersensitivity and suppression of cell death. Mol. Plant–Microbe Interact. 22, 269–281. [DOI] [PubMed] [Google Scholar]
- Collmer, A. and Gold, S. (2007) Noel T. Keen: Pioneer leader in molecular plant pathology. Annu. Rev. Phytopathol. 45, 25–42. [DOI] [PubMed] [Google Scholar]
- Daly, J.M. (1984) The role of recognition in plant disease. Annu. Rev. Phytopathol. 22, 273–307. [Google Scholar]
- Damasceno, C.M. , Bishop, J.G. , Ripoll, D.R. , Win, J. , Kamoun, S. and Rose, J.K. (2008) Structure of the glucanase inhibitor protein (GIP) family from Phytophthora species suggests coevolution with plant endo‐beta‐1,3‐glucanases. Mol. Plant–Microbe Interact. 21, 820–830. [DOI] [PubMed] [Google Scholar]
- Doke, N. (1983a) Generation of superoxide anion by potato tuber protoplasts during the hypersensitive response to hyphal wall components of Phytophthora infestans and specific inhibition of the reaction by suppressors of hypersensitivity. Physiol. Plant Pathol. 23, 359–367. [Google Scholar]
- Doke, N. (1983b) Involvement of superoxide anion generation in the hypersensitive response of potato tuber tissues to infection with an incompatible race of Phytophthora infestans and to the hyphal wall components. Physiol. Plant Pathol. 23, 345–357. [Google Scholar]
- Dong, S. , Qutob, D. , Tedman‐Jones, J. , Kuflu, K. , Wang, Y. , Tyler, B.M. and Gijzen, M. (2009) The Phytophthora sojae avirulence locus Avr3c encodes a multi‐copy RXLR effector with sequence polymorphisms among pathogen strains. PLoS ONE, 4, e5556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dou, D. , Kale, S.D. , Wang, X. , Chen, Y. , Wang, Q. , Jiang, R.H. , Arredondo, F.D. , Anderson, R.G. , Thakur, P.B. , McDowell, J.M. , Wang, Y. and Tyler, B.M. (2008a) Conserved C‐terminal motifs required for avirulence and suppression of cell death by Phytophthora sojae effector Avr1b. Plant Cell, 20, 1118–1133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dou, D. , Kale, S.D. , Wang, X. , Jiang, R.H. , Bruce, N.A. , Arredondo, F.D. , Zhang, X. and Tyler, B.M. (2008b)RXLR‐mediated entry of Phytophthora sojae effector Avr1b into soybean cells does not require pathogen‐encoded machinery. Plant Cell,20,1930–1947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ebel, J. , Ayers, A.R. and Albersheim, P. (1976) Host–pathogen interactions XII: response of suspension‐cultured soybean cells to the elicitor isolated from Phytophthora megasperma var. sojae, a fungal pathogen of soybeans. Plant Physiol. 57, 775–779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellis, J.G. , Rafiqi, M. , Gan, P. , Chakrabarti, A. and Dodds, P.N. (2009) Recent progress in discovery and functional analysis of effector proteins of fungal and oomycete plant pathogens. Curr. Opin. Plant Biol. 4, 399–405. [DOI] [PubMed] [Google Scholar]
- Erwin, D.C. and Ribeiro, O.K. (1996) Phytophthora Diseases Worldwide. St. Paul, MN: APS Press. [Google Scholar]
- Govers, F. and Gijzen, M. (2006) Phytophthora genomics: the plant destroyers' genome decoded. Mol. Plant–Microbe Interact. 19, 1295–1301. [DOI] [PubMed] [Google Scholar]
- Gracen, V.E. (1982) Role of genetics in etiological phytopathology. Annu. Rev. Phytopathol. 20, 219–233. [Google Scholar]
- Grouffaud, S. , Van West, P. , Avrova, A.O. , Birch, P.R. and Whisson, S.C. (2008) Plasmodium falciparum and Hyaloperonospora parasitica effector translocation motifs are functional in Phytophthora infestans . Microbiology, 154, 3743–3751. [DOI] [PubMed] [Google Scholar]
- Haas, B.J. , Kamoun, S. , Zody, M.C. , Jiang, R.H. , Handsaker, R.E. , Cano, L.M. , Grabherr, M. , Kodira, C.D. , Raffaele, S. , Torto‐Alalibo, T. , Bozkurt, T.O. , Ah‐Fong, A.M. , Alvarado, L. , Anderson, V.L. , Armstrong, M.R. , Avrova, A. , Baxter, L. , Beynon, J. , Boevink, P.C. , Bollmann, S.R. , Bos, J.I. , Bulone, V. , Cai, G. , Cakir, C. , Carrington, J.C. , Chawner, M. , Conti, L. , Costanzo, S. , Ewan, R. , Fahlgren, N. , Fischbach, M.A. , Fugelstad, J. , Gilroy, E.M. , Gnerre, S. , Green, P.J. , Grenville‐Briggs, L.J. , Griffith, J. , Grunwald, N.J. , Horn, K. , Horner, N.R. , Hu, C.H. , Huitema, E. , Jeong, D.H. , Jones, A.M. , Jones, J.D. , Jones, R.W. , Karlsson, E.K. , Kunjeti, S.G. , Lamour, K. , Liu, Z. , Ma, L. , Maclean, D. , Chibucos, M.C. , McDonald, H. , McWalters, J. , Meijer, H.J. , Morgan, W. , Morris, P.F. , Munro, C.A. , O'Neill, K. , Ospina‐Giraldo, M. , Pinzon, A. , Pritchard, L. , Ramsahoye, B. , Ren, Q. , Restrepo, S. , Roy, S. , Sadanandom, A. , Savidor, A. , Schornack, S. , Schwartz, D.C. , Schumann, U.D. , Schwessinger, B. , Seyer, L. , Sharpe, T. , Silvar, C. , Song, J. , Studholme, D.J. , Sykes, S. , Thines, M. , Van De Vondervoort, P.J. , Phuntumart, V. , Wawra, S. , Weide, R. , Win, J. , Young, C. , Zhou, S. , Fry, W. , Meyers, B.C. , Van West, P. , Ristaino, J. , Govers, F. , Birch, P.R. , Whisson, S.C. , Judelson, H.S. and Nusbaum, C. (2009) Genome sequence and comparative analysis of the Irish potato famine pathogen Phytophthora infestans . Nature, 461, 393–398. [DOI] [PubMed] [Google Scholar]
- Hahlbrock, K. , Lamb, C.J. , Purwin, C. , Ebel, J. , Fautz, E. and Schafer, E. (1981) Rapid response of suspension‐cultured parsley cells to the elicitor from Phytophthora megasperma var. sojae—induction of the enzymes of general phenylpropanoid metabolism. Plant Physiol. 67, 768–773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haldar, K. , Kamoun, S. , Hiller, N.L. , Bhattacharje, S. and Van Ooij, C. (2006) Common infection strategies of pathogenic eukaryotes. Nat. Rev. Microbiol. 4, 922–931. [DOI] [PubMed] [Google Scholar]
- Hogenhout, S.A. , Van der Hoorn, R.A. , Terauchi, R. and Kamoun, S. (2009) Emerging concepts in effector biology of plant‐associated organisms. Mol. Plant–Microbe Interact. 22, 115–122. [DOI] [PubMed] [Google Scholar]
- Huitema, E. , Bos, J.I.B. , Tian, M. , Win, J. , Waugh, M.E. and Kamoun, S. (2004) Linking sequence to phenotype in Phytophthora–plant interactions. Trends Microbiol. 12, 193–200. [DOI] [PubMed] [Google Scholar]
- Jiang, R.H. , Tripathy, S. , Govers, F. and Tyler, B.M. (2008) RXLR effector reservoir in two Phytophthora species is dominated by a single rapidly evolving superfamily with more than 700 members. Proc. Natl. Acad. Sci. USA, 105, 4874–4879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamoun, S. (2003) Molecular genetics of pathogenic oomycetes. Eukaryot. Cell, 2, 191–199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamoun, S. (2006) A catalogue of the effector secretome of plant pathogenic oomycetes. Annu. Rev. Phytopathol. 44, 41–60. [DOI] [PubMed] [Google Scholar]
- Kamoun, S. (2007) Groovy times: filamentous pathogen effectors revealed. Curr. Opin. Plant Biol. 10, 358–365. [DOI] [PubMed] [Google Scholar]
- Kanneganti, T.D. , Huitema, E. , Cakir, C. and Kamoun, S. (2006) Synergistic interactions of the plant cell death pathways induced by Phytophthora infestans Nep1‐like protein PiNPP1.1 and INF1 elicitin. Mol. Plant–Microbe Interact. 19, 854–863. [DOI] [PubMed] [Google Scholar]
- Kawamura, Y. , Hase, S. , Takenaka, S. , Kanayama, Y. , Yoshioka, H. , Kamoun, S. and Takahashi, H. (2009) INF1 elicitin activates jasmonic acid‐ and ethylene‐mediated signalling pathways and induces resistance to bacterial wilt disease in tomato. J. Phytopathol. 157, 287–297. [Google Scholar]
- Keen, N.T. (1975) Specific elicitors of plant phytoalexin production: determinants of race specificity in pathogens? Science, 187, 74–75. [DOI] [PubMed] [Google Scholar]
- De Koning‐Ward, T.F. , Gilson, P.R. , Boddey, J.A. , Rug, M. , Smith, B.J. , Papenfuss, A.T. , Sanders, P.R. , Lundie, R.J. , Maier, A.G. , Cowman, A.F. and Crabb, B.S. (2009) A newly discovered protein export machine in malaria parasites. Nature, 459, 945–949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamour, K. and Kamoun, S. (2009) Oomycete Genetics and Genomics: Diversity, Interactions and Research Tools. Hoboken, NJ: Wiley‐Blackwell. [Google Scholar]
- Lamour, K.H. , Win, J. and Kamoun, S. (2007) Oomycete genomics: new insights and future directions. FEMS Microbiol. Lett. 274, 1–8. [DOI] [PubMed] [Google Scholar]
- Lawton, M.A. , Dixon, R.A. , Hahlbrock, K. and Lamb, C.J. (1983) Elicitor induction of mRNA activity. Rapid effects of elicitor on phenylalanine ammonia‐lyase and chalcone synthase mRNA activities in bean cells. Eur. J. Biochem. 130, 131–139. [PubMed] [Google Scholar]
- Liu, Z. , Bos, J.I.B. , Armstrong, M. , Whisson, S.C. , Da Cunha, L. , Torto‐Alalibo, T. , Win, J. , Avrova, A.O. , Wright, F. , P.R.J., B. and Kamoun, S. (2005) Patterns of diversifying selection in the phytotoxin‐like scr74 gene family of Phytophthora infestans . Mol. Biol. Evol. 22, 659–672. [DOI] [PubMed] [Google Scholar]
- Lokossou, A.A. , Park, T.H. , Van Arkel, G. , Arens, M. , Ruyter‐Spira, C. , Morales, J. , Whisson, S.C. , Birch, P.R. , Visser, R.G. , Jacobsen, E. and Van Der Vossen, E.A. (2009) Exploiting knowledge of R/Avr genes to rapidly clone a new LZ‐NBS‐LRR family of late blight resistance genes from potato linkage group IV. Mol. Plant–Microbe Interact. 22, 630–641. [DOI] [PubMed] [Google Scholar]
- Morgan, W. and Kamoun, S. (2007) RXLR effectors of plant pathogenic oomycetes. Curr. Opin. Microbiol. 10, 332–338. [DOI] [PubMed] [Google Scholar]
- Oh, S.K. , Young, C. , Lee, M. , Oliva, R. , Bozkurt, T. , Cano, L.M. , Win, J. , Bos, J.I.B. , Liu, H.‐Y. , Van Damme, M. , Morgan, W. , Choi, D. , Van Der Vossen, E.A.G. , Vleeshouwers, V. and Kamoun, S. (2009) In planta expression screens of Phytophthora infestans RXLR effectors reveal diverse phenotypes including activation of the Solanum bulbocastanum disease resistance protein Rpi‐blb2. Plant Cell, In press. [DOI] [PMC free article] [PubMed]
- Qutob, D. , Kamoun, S. and Gijzen, M. (2002) Expression of a Phytophthora sojae necrosis‐inducing protein occurs during transition from biotrophy to necrotrophy. Plant J. 32, 361–373. [DOI] [PubMed] [Google Scholar]
- Qutob, D. , Tedman‐Jones, J. , Dong, S. , Kuflu, K. , Pham, H. , Wang, Y. , Dou, D. , Kale, S.D. , Arredondo, F.D. , Tyler, B.M. and Gijzen, M. (2009) Copy number variation and transcriptional polymorphisms of Phytophthora sojae RXLR effector genes Avr1a and Avr3a . PLoS ONE, 4, e5066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rehmany, A.P. , Gordon, A. , Rose, L.E. , Allen, R.L. , Armstrong, M.R. , Whisson, S.C. , Kamoun, S. , Tyler, B.M. , Birch, P.R. and Beynon, J.L. (2005) Differential recognition of highly divergent downy mildew avirulence gene alleles by RPP1 resistance genes from two Arabidopsis lines. Plant Cell, 17, 1839–1850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rose, J.K. , Ham, K.S. , Darvill, A.G. and Albersheim, P. (2002) Molecular cloning and characterization of glucanase inhibitor proteins: coevolution of a counterdefense mechanism by plant pathogens. Plant Cell, 14, 1329–1345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharp, J.K. , Albersheim, P. , Ossowski, P. , Pilotti, A. , Garegg, P.J. and Lindberg, B. (1984a) Comparison of the structures and elicitor activities of a synthetic and a mycelial‐wall‐derived hexa(β‐D‐gluco‐pyranosyl)‐D‐glucitol. J. Biol. Chem. 259, 11341–11345. [PubMed] [Google Scholar]
- Sharp, J.K. , McNeil, M. and Albersheim, P. (1984b) The primary structures of one elicitor‐active and seven elicitor‐inactive hexa(β‐D‐gluco‐pyranosyl)‐D‐glucitols isolated from the mycelial wall of Phytophthora megasperma f. sp. glycinea . J. Biol. Chem. 259, 11321–11336. [PubMed] [Google Scholar]
- Shaw, D.S. (1983) The perenosporales: a fungal geneticist's nightmare In: Zoosporic Plant Pathogens (Buczacki S.T., ed.), pp. 85–121. London: Academic Press. [Google Scholar]
- Sohn, K.H. , Lei, R. , Nemri, A. and Jones, J.D. (2007) The downy mildew effector proteins ATR1 and ATR13 promote disease susceptibility in Arabidopsis thaliana . Plant Cell, 19, 4077–4090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song, J. , Win, J. , Tian, M. , Schornack, S. , Kaschani, F. , Ilyas, M. , Van Der Hoorn, R.A. and Kamoun, S. (2009) Apoplastic effectors secreted by two unrelated eukaryotic plant pathogens target the tomato defense protease Rcr3. Proc. Natl. Acad. Sci. USA, 106, 1654–1659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Staskawicz, B.J. , Dahlbeck, D. and Keen, N.T. (1984) Cloned avirulence gene of Pseudomonas syringae pv. glycinea determines race‐specific incompatibility on Glycine max (L.) Merr. Proc. Natl. Acad. Sci. USA, 81, 6024–6028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stavrinides, J. , Ma, W. and Guttman, D.S. (2006) Terminal reassortment drives the quantum evolution of type III effectors in bacterial pathogens. PLoS Pathog. 2, e104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tian, M. , Benedetti, B. and Kamoun, S. (2005) A second Kazal‐like protease inhibitor from Phytophthora infestans inhibits and interacts with the apoplastic pathogenesis‐related protease P69B of tomato. Plant Physiol. 138, 1785–1793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tian, M. , Huitema, E. , Cunha, L. , Torto‐Alalibo, T. and Kamoun, S. (2004) A Kazal‐like extracellular serine protease inhibitor from Phytophthora infestans targets the tomato pathogenesis‐related protease P69B. J. Biol. Chem. 279, 26370–26377. [DOI] [PubMed] [Google Scholar]
- Tian, M. and Kamoun, S. (2005) A two disulfide bridge Kazal domain from Phytophthora exhibits stable inhibitory activity against serine proteases of the subtilisin family. BMC Biochem. 6, 15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tian, M. , Win, J. , Song, J. , Van Der Hoorn, R. , Van Der Knaap, E. and Kamoun, S. (2007) A Phytophthora infestans cystatin‐like protein targets a novel tomato papain‐like apoplastic protease. Plant Physiol. 143, 364–377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turner, S.R. (2008) Potato agriculture, late blight science, and the molecularization of plant pathology. Hist. Stud. Nat. Sci. 38, 223–257. [DOI] [PubMed] [Google Scholar]
- Tyler, B.M. (2009) Entering and breaking: virulence effector proteins of oomycete plant pathogens. Cell. Microbiol. 11, 13–20. [DOI] [PubMed] [Google Scholar]
- Tyler, B.M. , Tripathy, S. , Zhang, X. , Dehal, P. , Jiang, R.H. , Aerts, A. , Arredondo, F.D. , Baxter, L. , Bensasson, D. , Beynon, J.L. , Chapman, J. , Damasceno, C.M. , Dorrance, A.E. , Dou, D. , Dickerman, A.W. , Dubchak, I.L. , Garbelotto, M. , Gijzen, M. , Gordon, S.G. , Govers, F. , Grunwald, N.J. , Huang, W. , Ivors, K.L. , Jones, R.W. , Kamoun, S. , Krampis, K. , Lamour, K.H. , Lee, M.K. , McDonald, W.H. , Medina, M. , Meijer, H.J. , Nordberg, E.K. , Maclean, D.J. , Ospina‐Giraldo, M.D. , Morris, P.F. , Phuntumart, V. , Putnam, N.H. , Rash, S. , Rose, J.K. , Sakihama, Y. , Salamov, A.A. , Savidor, A. , Scheuring, C.F. , Smith, B.M. , Sobral, B.W. , Terry, A. , Torto‐Alalibo, T.A. , Win, J. , Xu, Z. , Zhang, H. , Grigoriev, I.V. , Rokhsar, D.S. and Boore, J.L. (2006) Phytophthora genome sequences uncover evolutionary origins and mechanisms of pathogenesis. Science, 313, 1261–1266. [DOI] [PubMed] [Google Scholar]
- Van West, P. , De Jong, A.J. , Judelson, H.S. , Emons, A.M.C. and Govers, F. (1998) The ipiO gene of Phytophthora infestans is highly expressed in invading hyphae during infection. Funct. Gen. Biol. 23, 126–138. [DOI] [PubMed] [Google Scholar]
- Vleeshouwers, V.G. , Rietman, H. , Krenek, P. , Champouret, N. , Young, C. , Oh, S.K. , Wang, M. , Bouwmeester, K. , Vosman, B. , Visser, R.G. , Jacobsen, E. , Govers, F. , Kamoun, S. and Van der Vossen, E.A. (2008) Effector genomics accelerates discovery and functional profiling of potato disease resistance and Phytophthora infestans avirulence genes. PLoS ONE, 3, e2875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vleeshouwers, V.G.A.A. , Driesprong, J.‐D. , Kamphuis, L.G. , Torto‐Alalibo, T. , Van‘t Slot, K.A.E. , Govers, F. , Visser, R.G.F. , Jacobsen, E. and Kamoun, S. (2006) Agroinfection‐based high throughput screening reveals specific recognition of INF elicitins in Solanum . Mol. Plant Pathol. 7, 499–510. [DOI] [PubMed] [Google Scholar]
- Wade, M.A. and Albersheim, P. (1979) Race‐specific molecules that protect soybeans from Phytophthora megasperma var. sojae . Proc. Natl. Acad. Sci. USA, 187, 74–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whisson, S.C. , Boevink, P.C. , Moleleki, L. , Avrova, A.O. , Morales, J.G. , Gilroy, E.M. , Armstrong, M.R. , Grouffaud, S. , Van West, P. , Chapman, S. , Hein, I. , Toth, I.K. , Pritchard, L. and Birch, P.R. (2007) A translocation signal for delivery of oomycete effector proteins into host plant cells. Nature, 450, 115–118. [DOI] [PubMed] [Google Scholar]
- Win, J. , Morgan, W. , Bos, J. , Krasileva, K.V. , Cano, L.M. , Chaparro‐Garcia, A. , Ammar, R. , Staskawicz, B.J. and Kamoun, S. (2007) Adaptive evolution has targeted the C‐terminal domain of the RXLR effectors of plant pathogenic oomycetes. Plant Cell, 19, 2349–2369. [DOI] [PMC free article] [PubMed] [Google Scholar]