

Abstract
Adaptation is a general property of sensory receptor neurons and has been extensively studied in isolated cell preparation of olfactory receptor neurons. In contrast, little is known about the conditions under which peripheral adaptation occurs in the CNS during odorant stimulation. Here, we used two-photon laser-scanning microscopy and targeted extracellular recording in freely breathing anesthetized rats to investigate the correlate of peripheral adaptation at the first synapse of the olfactory pathway in olfactory bulb glomeruli. We find that during sustained stimulation at high concentration, odorants can evoke local field potential (LFP) postsynaptic responses that rapidly adapt with time, some within two inhalations. Simultaneous measurements of LFP and calcium influx at olfactory receptor neuron terminals reveal that postsynaptic adaptation is associated with a decrease in odorant-evoked calcium response, suggesting that it results from a decrease in glutamate release. This glomerular adaptation was concentration-dependent and did not change the glomerular input–output curve. In addition, in situ application of antagonists of either ionotropic glutamate receptors or metabotropic GABAB receptors did not affect this adaptation, thus discarding the involvement of local presynaptic inhibition. Glomerular adaptation, therefore, reflects the response decline of olfactory receptor neurons to sustained odorant. We postulate that peripheral fast adaptation is a means by which glomerular output codes for high concentration of odor.
Introduction
In the vertebrate nasal epithelium, volatile odorants bind to odorant receptors (Buck and Axel, 1991) located in the cilia of olfactory receptor neurons (ORNs). There, they trigger a G-protein-coupled cascade (Jones and Reed, 1989; Bakalyar and Reed, 1990) that leads to the production of intraciliary cAMP and the opening of cyclic nucleotide-gated channels (CNG) (Nakamura and Gold, 1987) permeable to Na+, K+, and Ca2+ (for review, see Pifferi et al., 2006; Kleene, 2008). Ca2+ plays dual excitatory and inhibitory roles (Matthews and Reisert, 2003). It depolarizes ORNs and activates Ca2+-sensitive chloride channels which allow an efflux of chloride to further depolarize ORNs (Lowe and Gold, 1993). It also triggers several negative feedback processes that underlie adaptation to repeated or prolonged odor activation. Ca2+-binding proteins such as Ca2+/calmodulin decrease the sensitivity of CNG channels (Kurahashi and Menini, 1997) but also trigger cascades leading to the hydrolysis of cAMP (Borisy et al., 1992; Yan et al., 1995) or the inhibition of adenylyl cyclase (Wei et al., 1998; Leinders-Zufall et al., 1999). These processes have been extensively analyzed in vitro (for review, see Bradley et al., 2005; Kleene, 2008), whereas in vivo, only one group reported measurements of ORNs activity and adaptation of firing (Duchamp-Viret et al., 2000, 2003).
Furthermore, less is known about the consequences of peripheral adaptation in perceptual habituation or desensitization, an everyday experience in olfaction (for review, see Wilson and Linster, 2008). In the first and second relays of olfaction, i.e., in the olfactory bulb and the pyriform cortex, neurons do adapt (Wilson, 1998; Sobel et al., 2000; Schafer et al., 2005); however, the precise role of ORN adaptation versus local network processing in the decrement of central neuron responses is unclear. Here, we have simultaneously recorded the activation of the presynaptic and postsynaptic compartments of olfactory bulb glomeruli to investigate under which conditions peripheral adaptation can be detected and how it affects the spatial maps of activated glomeruli (for review, see Mori et al., 2006; Wachowiak and Shipley, 2006). Such maps vary with the odorant concentration and are temporally dynamic (Spors and Grinvald, 2002; Spors et al., 2006; Schaefer and Margrie, 2007). We report that under high odorant concentration, ORN adaptation participates to the spatial redistribution of active glomeruli.
Materials and Methods
Animals preparation, electrophysiological recordings, and two-photon imaging.
Wistar rats (29 postnatal day 30–70) were anesthetized with urethane (1.65 g/kg, i.p.) and held in a standard stereotaxic apparatus. Olfactory bulb surgery, local field potential recordings, and focal pressure applications of drugs were performed as described previously (Chaigneau et al., 2007). NBQX (2,3-dihydroxy-6-nitro-1,2,3,4-tetrahydrobenzo[f]quinoxaline-7-sulfonamide; 250 μm), D-APV [d-2-(−)amino-5-phosphonopentatoic acid; 500 μm], and CGP35348 [(3-aminopropyl(diethoxymethyl) phosphoninic acid; 1–10 mm] were obtained from Tocris Bioscience. The temperature of the animal was maintained at 37°C with a feedback-controlled heating blanket (Harvard Apparatus). For all experiments, breathing frequency was monitored through a pneumogram transducer (BIOPAC Systems). Olfactory nerve terminals were labeled with Ca2+ green 1 dextran, 10 kDa (Invitrogen), 2–8 d before experiments using the method developed by Wachowiak and Cohen (2001). Images from subregions of the field of view were acquired at rates up to 20 frames per second using a custom-built two-photon laser-scanning microscope (TPLSM).
Olfactometer.
Odorants were applied for 4 s with a custom-built olfactometer. The olfactometer delivered a constant flow of humidified air in a small teflon reservoir surrounding the rat snout. All tubes connecting the odorant reservoir to the snout were made of teflon to minimize odorant contamination. Odorants used were aldehydes and esters. Two flow meters (Aalborg), one in the clean air path and one before the odorant reservoir, allowed to control odorant concentration in a reproducible manner. To reveal adaptation, high odorant concentrations were used, i.e., from 3–100% when expressed in percentage of saturated air.
Intertrial alignments of Ca2+ or local field potential responses using respiratory movements.
Respiration monitoring revealed spontaneous variations of inhalation frequency. Although minor, they affected the intertrial averaged responses, with a progressive decrease of Ca2+ and local field potential (LFP) response amplitudes at each inhalation during sustained odorant stimulation (supplemental Fig. 1A, available at www.jneurosci.org as supplemental material). To take into account this respiration variability, we developed a new averaging algorithm using respiratory movements. For each individual trial, every breathing cycle period was detected and the position of the LFP response onset in the cycle period determined. An averaged breathing signal was then computed from several individuals, and each individual trial was stretched to fit with this average. Ca2+ and LFP responses were then extracted by averaging the trials. Supplemental Figure 1, available at www.jneurosci.org as supplemental material, illustrates that our algorithm allowed maintaining response amplitudes in the averages during prolonged stimulation. These alignments were used to illustrate LFP and Ca2+ responses (see Figs. 2, 3) but not to quantify individual LFP and Ca2+ peaks (see Fig. 2B,C).
Figure 2.
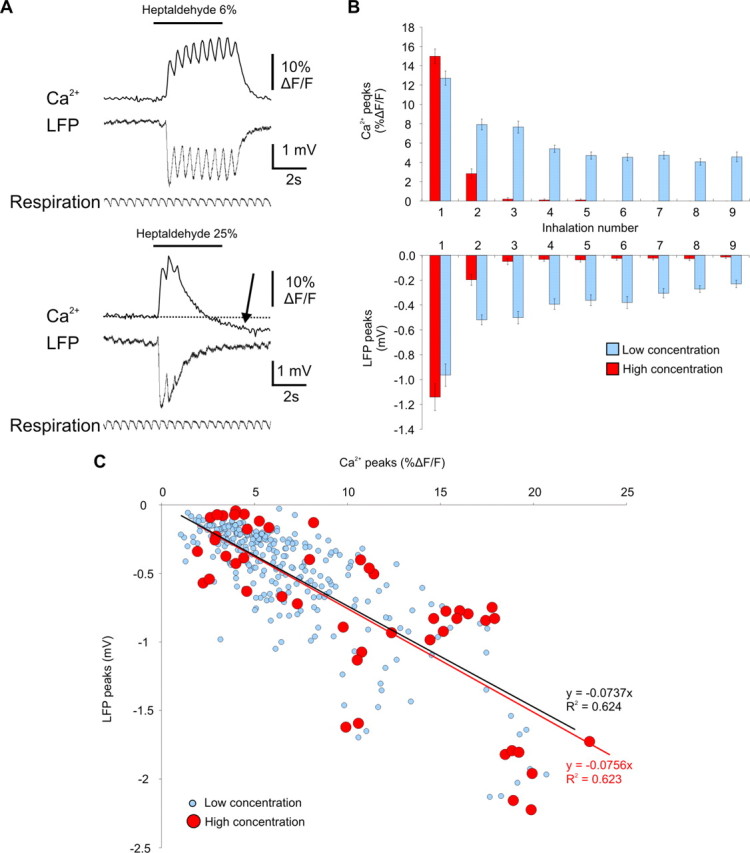
Both calcium (Ca2+) and LFP responses show adaptation. A, Presynaptic Ca2+ influx in ORNs and LFP were recorded in the same glomerulus. Increasing odorant concentration (bottom traces) strongly shortened both LFP and Ca2+ responses without affecting respiration. Note that in this example, Ca2+ level reversibly decreased below baseline during high odorant concentration (black arrow). B, Graphs of Ca2+ (ΔF/F, top traces) and LFP (ΔV, bottom traces) sniff evoked increments as a function of inhalation numbers, in the presence of an odorant (n = 8 animals). Ca2+ and LFP peak values are raw data (i.e., without averaging or stretching) measured for each inhalation. At low odorant concentration (blue bars), Ca2+ and LFP peak amplitudes were maintained, although they disappeared within two inhalations at high odorant concentration (red bars). C, The input (Ca2+ peaks)–output (LFP peaks) relationship was not affected by odorant concentration and adaptation, as shown by the perfect overlap of the two linear regression curves. Regression lines were drawn from all evoked responses in eight animals for low and high odorant concentrations.
Figure 3.

In vivo blockade of postsynaptic activity reveals peripheral ORN adaptation. A, B, Simultaneous recordings of presynaptic and postsynaptic activity in control condition and after local NBQX/D-APV (250 μm/500 μm) application. Single (A) and averaged (B) responses (n = 8 animals) revealed that glutamate antagonists reversibly blocked LFP responses and increased Ca2+ peak amplitudes. The first Ca2+ peak was also increased, indicating tonic presynaptic inhibition. NBQX/D-APV barely affected adaptation of Ca2+ responses at high odorant concentration. C, Presynaptic activity in control condition and after local application of the GABAB antagonist CGP35348 (1–10 mm). Single (top) and averaged (bottom) responses (n = 7) confirmed that tonic presynaptic inhibition is mediated through GABAB receptors but not responsible of adaptation at high odorant concentration. D, Summary graphs showing the effects of NBQX/D-APV and CGP35348 on Ca2+ responses.
Data analysis.
All data were analyzed through a homemade MATLAB program (MathWorks). In the entire study, average values are expressed as mean ± SEM.
Results
High concentration of odorant elicits two types of LFP responses in glomeruli
To trigger ORN adaptation, odorants were applied at high concentrations for 4 s. LFP responses were recorded with fluorescent extracellular glass pipettes targeted in labeled glomeruli with TPLSM. Odorant evoked complex LFP responses with respiration-locked rapid negativities (Fig. 1). We have previously shown that these LFP rapid negativities reflect neuronal postsynaptic activation restricted to a single glomerulus: they are odorant and glomerular specific; they display sharp transitions in space (laterally and in depth), they are blocked by intraglomerular application of ionotropic glutamate antagonists; they are correlated to mitral cell EPSPs (Chaigneau et al., 2007), indicating that mitral cell dendrites participate to their generation. During sustained stimulation at high odorant concentration, we observed two types of LFP responses that differed on their level of adaptation. Note that although the odorant was continuously applied, ORN stimulation was discontinuous because stimulation occurred only at each inhalation every ∼0.5 s. Figure 1A shows a “weakly adapting response”: when the odorant concentration increased, the first negativity, i.e., the initial field EPSP increased and saturated. The following negativities (or peaks) were either maintained or showed a slight decrease in amplitude as the response developed in time. In contrast, some responses strongly adapted with high concentration (from 10 to 100% of saturated air) (Fig. 1B). These “strongly adapting responses” were characterized by a large decrease or even an abolition of all the peaks after the initial one. In addition, the slow negative envelop which was crowned by the peaks at low odorant concentration also disappeared. Occasionally, even the first peak amplitude slightly decreased (2 of 8 animals). Differences in adaptation level (weak vs strong) occurred between neighboring glomeruli for a given odorant (see below) or in a single glomerulus for two different odorants. Several mechanisms could underlie strongly adapting responses during sustained odorant, e.g., a postsynaptic inhibition of mitral cells involving the glomerular network or a progressive decrease in ORN terminals activation, i.e., a decrease in glutamate release.
Figure 1.

High odorant concentration elicits two types of LFP responses. LFP responses were recorded in different glomeruli. Odorants elicited strong respiration-locked LFP negativities. Increasing odorant concentration revealed two types of responses that weakly (A) or strongly (B) adapted. Arrow shows strong adaptation.
Adaptation of Ca2+ influx in olfactory nerve terminals during odor
To analyze the activation of ORN terminals, we imaged Ca2+ influx in terminals loaded with organic Ca2+ sensors (see Materials and Methods). Labeled terminals outlined glomerular boundaries and allowed to assess the kinetics of odorant-evoked Ca2+ signals (Wachowiak and Cohen, 2001). In single glomeruli (n = 8 animals), we simultaneously recorded presynaptic Ca2+ and LFP responses. We therefore investigated the relation between presynaptic and postsynaptic compartments during odorant responses. Figure 2A shows a typical example in which presynaptic Ca2+ responses were surprisingly similar to LFP responses. At low odorant concentration, the initial Ca2+ and LFP peaks were similarly larger than the following ones which maintained their amplitude during the stimulus duration. Note that all peaks were phase-locked to respiration. At high odorant concentration, both Ca2+ and LFP responses strongly adapted, the third inhalation peaks being barely detectable. In half of the cases (4 of 8 animals), Ca2+ concentration decreased below baseline (Fig. 2, arrow). It was not attributable to photo-bleaching as no rundown was visible before odor and as no decay with time was observable during control experiments (no odorant application; data not shown). It also did not involve changes in respiration frequency. To quantify adaptation at both presynaptic and postsynaptic levels, we measured on each individual trace (i.e., without averaging or stretching; see Materials and Methods) the peak amplitude of Ca2+ (ΔF/F increment) (Fig. 2B, top) and LFP (ΔV increment) (Fig. 2B, bottom) signals at each inhalation. At low concentration, Ca2+ and LFP peaks decreased after the initial one but then remained similarly constant during most of the odorant application (4 s). At high concentration, Ca2+ and LFP peaks disappeared after the second inhalation but in one case (out of 8) persisted during five inhalations. The average delay between the first peak and the first inhalation without any response was 1.04 ± 0.14 s (n = 8), indicating that after only two inhalations, little glomerular excitatory output was sent to mitral cell somata. Because LFP responses mirrored adapting Ca2+ signals and, in some cases (see Fig. 4A), adaptation already occurred on the initial Ca2+ peak, dye saturation could not account for adaptation of fluorescence signals. Overall, these results suggest that glomerular LFP adaptation is associated with a concentration- and time-dependent adaptation of Ca2+ influx in ORN terminals and most probably from a decrease in glutamate release.
Figure 4.
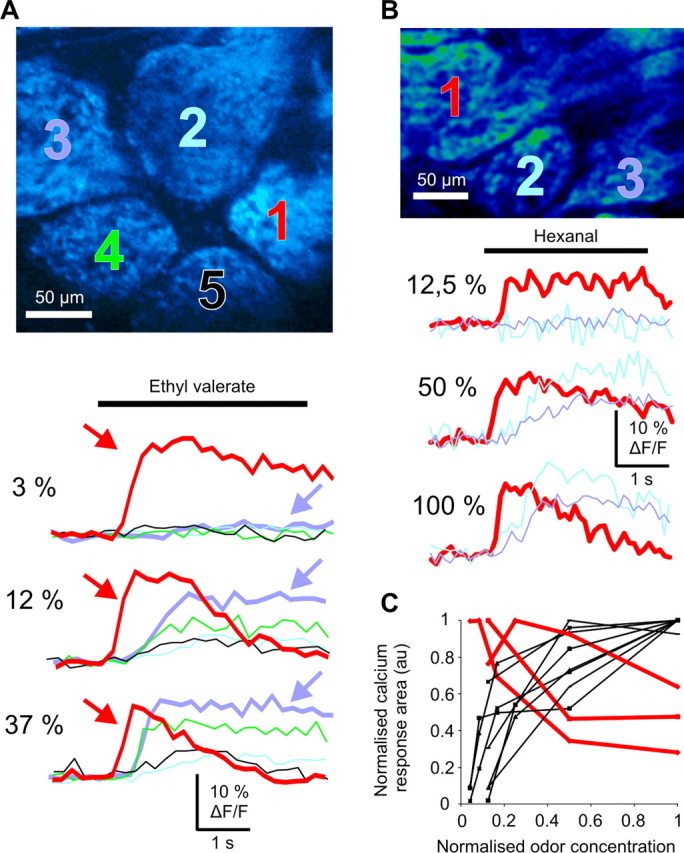
Peripheral adaptation causes a spatial redistribution of activated glomeruli. A, B, Top, Two-photon imaging of two large fields of view comprising five (A) and three (B) glomeruli. ORN terminals were labeled with Ca2+ green dextran (10 kDa). A, Bottom, Glomeruli presented two types of Ca2+ responses according to the odorant concentration. At low concentration, ethyl valerate evoked a rapid Ca2+ plateau (red arrow) in glomerulus 1, although it did not affect neighboring glomeruli (2–5). Raising the odorant concentration shortened the Ca2+ response of glomerulus 1, although it revealed those of glomeruli 2–5. Note the appearance of a Ca2+ plateau in glomerulus 2 (blue arrow). B, Bottom, Another case where the most sensitive glomerulus (glomerulus 1) shows a Ca2+ response adaptation. C, Quantification of Ca2+ signal adaptation. The overall Ca2+ responses (area) were normalized and plotted as a function of the odorant concentration (normalized to its maximum value) for three groups of glomeruli (11 glomeruli, three animals). Peripheral adaptation greatly reduced Ca2+ responses of the three most sensitive glomeruli (red traces), whereas responses of the neighboring glomeruli increased (black traces).
The glomerular input–output curve
The two graphs in Figure 2B indicate that presynaptic and postsynaptic responses were highly correlated. To determine whether this correlation was sensitive to odorant concentration, we plotted LFP peaks as a function of presynaptic Ca2+ peaks (Fig. 2C), i.e., the input–output curve of all recorded glomeruli. At both low and high odorant concentration, the linear regression curves surprisingly overlapped, indicating that Ca2+ peaks are excellent markers of LFP peaks, which are themselves highly correlated to EPSPs recorded in mitral cells (Chaigneau et al., 2007). The perfect overlap of the two linear regression curves also indicates that LFP adaptation does not involve a postsynaptic mechanism. It involves either a presynaptic inhibition of ORN terminals or an ORN adaptation at the nasal epithelium level. Presynaptic inhibition regulates glutamate release at ORN terminals (Hsia et al., 1999; Wachowiak and Cohen, 1999; Aroniadou-Anderjaska et al., 2000; Ennis et al., 2001; Olsen and Wilson, 2008) by decreasing Ca2+ influx in the terminals (Wachowiak and Cohen, 1999; McGann et al., 2005; Vucinić et al., 2006; Pírez and Wachowiak, 2008). To investigate the role of presynaptic inhibition in glomerular adaptation, we tested the effects of local application of antagonists of ionotropic glutamate receptors and GABAB receptors on ORN terminals Ca2+ responses.
Fast peripheral adaptation, and not presynaptic inhibition, underlies glomerular adaptation
We have shown previously that odorant-evoked local LFP responses are abolished by focal applications of NBQX and D-APV, antagonists of glutamate AMPA and NMDA receptors, respectively (Chaigneau et al., 2007). Here, we coapplied Alexa Fluor 594 and glutamate antagonists (NBQX, 250 μm; D-APV, 500 μm) through the LFP recording pipette and maintained the pipette pressure until the drug diffused into the entire glomerular and periglomerular boundaries, ensuring that we blocked all intraglomerular and interglomerular glutamatergic synaptic inputs. Glutamate antagonists reversibly blocked the local LFP response, i.e., the LFP peaks and part of the slow component (Fig. 3A,B). Note that the remaining slow component was not locally generated because it persists in the external plexiform layer (Chaigneau et al., 2007) and reverses its polarity in the granule cell layer (E. Chaigneau and S. Charpak, unpublished data), thus reflecting the activation of granule cells. In contrast, NBQX/D-APV reversibly enhanced presynaptic Ca2+ responses evoked by both low and high odorant concentrations. Glutamate antagonists increased the initial Ca2+ peak to 126% of control [Ca2+ peak (percentage of ΔF/F]: control, 15.30 ± 3.15; NBQX/D-APV, 19.39 ± 3.33; recovery, 15.24 ± 3.26; n = 8; p = 0.013) as well as the overall Ca2+ signal (area) to 119% of control [Ca2+ area (arbitrary units, a.u.)]: control, 37.34 ± 5.69; NBQX/D-APV, 44.58 ± 6.45; recovery, 36.11 ± 5.95; n = 8; p = 0.009). These results confirm, although under different anesthesia conditions (Pírez and Wachowiak, 2008), that tonic presynaptic inhibition modulates odorant-evoked Ca2+ signals in ORN terminals and as a consequence the glomerular output. Note that the increase of presynaptic Ca2+ (area) by NBQX/D-APV was similar for odorant stimulations at low and high concentration (117 ± 7% and 125 ± 9%, respectively; p = 0.54). This further suggests that glomerular adaptation results from tonic inhibition rather than from odorant-evoked presynaptic inhibition. Similarly, local application of the GABAB receptor antagonist CGP35348 (1–10 mm) (Fig. 3C) also increased the overall signal [Ca2+ area (a.u.)]: control, 22.86 ± 2.15; CGP35348, 27.97 ± 2.20; n = 8; p = 0.01), indicating that presynaptic tonic inhibition involved GABAB receptors (Pírez and Wachowiak, 2008). In contrast to presynaptic Ca2+ signals, LFP responses were either maintained or slightly decreased (data not shown) on CGP35348, suggesting that GABAB receptors are involved at both presynaptic and postsynaptic sites, and their activation induces a complex modulation of the glomerular network activity. More importantly, blockade of presynaptic inhibition did not significantly affect the adaptation of Ca2+ influx. Ca2+ responses to low odorant concentration in control conditions continued to be significantly larger than responses to high odorant concentration in the absence of presynaptic inhibition. The general shape of Ca2+ responses was not modified by glutamate or GABAB receptor antagonists, implying that in urethane anesthetized animals, peripheral adaptation of ORNs underlies the glomerular adaptation that occurs during high odorant concentration.
Peripheral adaptation modulates glomerular maps
What are the consequences of peripheral adaptation on glomerular maps? TPLSM imaging of large fields of view comprising several glomeruli, with a lower temporal resolution, allowed assessing the spread of odor-evoked Ca2+ signals with odorant concentration. Figure 4, A and B, illustrate two experiments where several neighboring glomeruli were activated by an odorant. At low concentration, odor evoked a rapid Ca2+ response in a single glomerulus (Fig. 4A,B, traces), confirming the glomerular specificity of odorant stimulation at low concentration. The Ca2+ response had a sustained plateau-like shape, showing little attenuation during the 4 s stimulation. Note that the absence of Ca2+ peaks resulted from the low temporal resolution, each image lasting ∼200 ms. In each “odorant specific” glomerulus, increasing the odor concentration drastically shortened the plateau-like response observed at low concentration (Fig. 4A,B, traces in red) and even decreased, in some cases, the initial amplitude (Fig. 4A). In contrast, high odorant concentration recruited neighboring glomeruli, which showed little adaptation of Ca2+ responses. Such spatial redistribution of activated glomeruli was observed in all tested animals (n = 5). The graph in Figure 4C quantifies this effect on three groups of glomeruli (n = 11 glomeruli), which were tested for at least 3–4 odorant concentrations. In each group, one glomeruli showed a marked Ca2+ response decrease at high odorant concentration (red traces). These results show that because of peripheral adaptation, the relative glomerular output of a “specific” glomerulus (responsive to low odorant concentration) versus its neighbors changes drastically at high odor concentration.
Discussion
Peripheral adaptation depends on Ca2+ influx in ORNs and as a consequence on odorant concentration. Spatial maps of glomerular activity vary with odorant concentration; however, most of the variability is assigned to the low specificity of odorant receptors. Here, we investigated under which conditions odor could modulate glomerular activation through a mechanism involving peripheral adaptation. We specifically analyzed postsynaptic glomerular responses that expressed strong adaptation under high odorant concentration. We found that glomerular adaptation resulted from a decrease in ORN terminals activation, i.e., in glutamate release, which did not involve local presynaptic inhibition. Our study, thus, indirectly demonstrates that peripheral adaptation of ORNs underlies glomerular adaptation, without hypothesizing which peripheral adaptation processes are involved. Direct demonstration would require firing measurements of targeted axons converging in an adapting glomerulus. Such tour de force is not necessary because reported ORN properties fully account for glomerular adaptation. In addition, our pharmacological experiments confirm the existence of a tonic presynaptic inhibition of ORN terminals (Petzold et al., 2008; Pírez and Wachowiak, 2008). They also show that at high odorant concentration, peripheral adaptation widely dominates evoked-presynaptic inhibition.
We believe that the observed adaptation is a general mechanism that simply depends on the level of ORN activation. As a consequence, we postulate that odorant stimulation of increasing intensity will first increase and then progressively shorten specific glomerular responses such that at high concentration, only the first few inhalations will transmit information to second-order neurons. In contrast, the surrounding less specific glomeruli will become more active and relay peripheral information. Although LFP negativities strictly indicate what glomerular output (EPSPs) is transmitted to mitral/tufted cell somata, and gives no quantitative indication on the spiking output to the cortex, we hypothesize that peripheral adaptation is a major modulatory determinant of mitral/tufted cell firing during strong odorant stimulation. The validation of our hypothesis will require challenging experiments to measure mitral/tufted cell activity, first none invasively (in cell attached mode) and to extract in a second step (in whole-cell mode) the respective weights of peripheral adaptation and lateral inhibition in shaping the spiking output. It is also important to stress that the persistence of an odor response during the first inhalation remains compatible with the ability to discriminate odors. Indeed, recent studies have demonstrated that discrimination tasks can be achieved within few hundred milliseconds (Uchida and Mainen, 2003; Abraham et al., 2004; Rinberg et al., 2006; Slotnick, 2007). In fact, it has been shown that successful odor discrimination occurs before full development of the first inhalation glomerular map (Wesson et al., 2008). In addition to coding of high odor concentration, the fast peripheral adaptation underlying glomerular adaptation could increase the dynamic range of glomerular activation. Such a role had been proposed but not observed for evoked-presynaptic inhibition (Pírez and Wachowiak, 2008) and is classically reported at the level of ORN transduction (for review, see Kleene, 2008).
Footnotes
This work was supported by the Institut National de la Santé et de la Recherche Médicale, the Ministère de l'Education Nationale, de la Recherche et de la Technologie (Grant NIC0025), the Centre National de la Recherche Scientifique, the Fondation pour la Recherche Médicale (Grant ICP20001222128), the Région Ile de France (Sesame Program), the Fondation Bettancourt Schueller, and the Human Frontier Science Program Organization. J.L. was supported by a fellowship from the Fondation pour la Recherche Médicale (Grant FDT20070910324), and P.T. was supported by a fellowship from the Association Française contre les Myopathies (Grant 12411). We thank Jonathan Bradley for his critical comments.
References
- Abraham et al., 2004.Abraham NM, Spors H, Carleton A, Margrie TW, Kuner T, Schaefer AT. Maintaining accuracy at the expense of speed: stimulus similarity defines odor discrimination time in mice. Neuron. 2004;44:865–876. doi: 10.1016/j.neuron.2004.11.017. [DOI] [PubMed] [Google Scholar]
- Aroniadou-Anderjaska et al., 2000.Aroniadou-Anderjaska V, Zhou FM, Priest CA, Ennis M, Shipley MT. Tonic and synaptically evoked presynaptic inhibition of sensory input to the rat olfactory bulb via GABA(B) heteroreceptors. J Neurophysiol. 2000;84:1194–1203. doi: 10.1152/jn.2000.84.3.1194. [DOI] [PubMed] [Google Scholar]
- Bakalyar and Reed, 1990.Bakalyar HA, Reed RR. Identification of a specialized adenylyl cyclase that may mediate odorant detection. Science. 1990;250:1403–1406. doi: 10.1126/science.2255909. [DOI] [PubMed] [Google Scholar]
- Borisy et al., 1992.Borisy FF, Ronnett GV, Cunningham AM, Juilfs D, Beavo J, Snyder SH. Calcium/calmodulin-activated phosphodiesterase expressed in olfactory receptor neurons. J Neurosci. 1992;12:915–923. doi: 10.1523/JNEUROSCI.12-03-00915.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradley et al., 2005.Bradley J, Reisert J, Frings S. Regulation of cyclic nucleotide-gated channels. Curr Opin Neurobiol. 2005;15:343–349. doi: 10.1016/j.conb.2005.05.014. [DOI] [PubMed] [Google Scholar]
- Buck and Axel, 1991.Buck L, Axel R. A novel multigene family may encode odorant receptors: a molecular basis for odor recognition. Cell. 1991;65:175–187. doi: 10.1016/0092-8674(91)90418-x. [DOI] [PubMed] [Google Scholar]
- Chaigneau et al., 2007.Chaigneau E, Tiret P, Lecoq J, Ducros M, Knöpfel T, Charpak S. The relationship between blood flow and neuronal activity in the rodent olfactory bulb. J Neurosci. 2007;27:6452–6460. doi: 10.1523/JNEUROSCI.3141-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duchamp-Viret et al., 2000.Duchamp-Viret P, Duchamp A, Chaput MA. Peripheral odor coding in the rat and frog: quality and intensity specification. J Neurosci. 2000;20:2383–2390. doi: 10.1523/JNEUROSCI.20-06-02383.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duchamp-Viret et al., 2003.Duchamp-Viret P, Duchamp A, Chaput MA. Single olfactory sensory neurons simultaneously integrate the components of an odour mixture. Eur J Neurosci. 2003;18:2690–2696. doi: 10.1111/j.1460-9568.2003.03001.x. [DOI] [PubMed] [Google Scholar]
- Ennis et al., 2001.Ennis M, Zhou FM, Ciombor KJ, Aroniadou-Anderjaska V, Hayar A, Borrelli E, Zimmer LA, Margolis F, Shipley MT. Dopamine D2 receptor-mediated presynaptic inhibition of olfactory nerve terminals. J Neurophysiol. 2001;86:2986–2997. doi: 10.1152/jn.2001.86.6.2986. [DOI] [PubMed] [Google Scholar]
- Hsia et al., 1999.Hsia AY, Vincent JD, Lledo PM. Dopamine depresses synaptic inputs into the olfactory bulb. J Neurophysiol. 1999;82:1082–1085. doi: 10.1152/jn.1999.82.2.1082. [DOI] [PubMed] [Google Scholar]
- Jones and Reed, 1989.Jones DT, Reed RR. Golf: an olfactory neuron specific-G protein involved in odorant signal transduction. Science. 1989;244:790–795. doi: 10.1126/science.2499043. [DOI] [PubMed] [Google Scholar]
- Kleene, 2008.Kleene SJ. The electrochemical basis of odor transduction in vertebrate olfactory cilia. Chem Senses. 2008;33:839–859. doi: 10.1093/chemse/bjn048. [DOI] [PubMed] [Google Scholar]
- Kurahashi and Menini, 1997.Kurahashi T, Menini A. Mechanism of odorant adaptation in the olfactory receptor cell. Nature. 1997;385:725–729. doi: 10.1038/385725a0. [DOI] [PubMed] [Google Scholar]
- Leinders-Zufall et al., 1999.Leinders-Zufall T, Ma M, Zufall F. Impaired odor adaptation in olfactory receptor neurons after inhibition of Ca2+/calmodulin kinase II. J Neurosci. 1999;19:RC19. doi: 10.1523/JNEUROSCI.19-14-j0005.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lowe and Gold, 1993.Lowe G, Gold GH. Nonlinear amplification by calcium-dependent chloride channels in olfactory receptor cells. Nature. 1993;366:283–286. doi: 10.1038/366283a0. [DOI] [PubMed] [Google Scholar]
- Matthews and Reisert, 2003.Matthews HR, Reisert J. Calcium, the two-faced messenger of olfactory transduction and adaptation. Curr Opin Neurobiol. 2003;13:469–475. doi: 10.1016/s0959-4388(03)00097-7. [DOI] [PubMed] [Google Scholar]
- McGann et al., 2005.McGann JP, Pírez N, Gainey MA, Muratore C, Elias AS, Wachowiak M. Odorant representations are modulated by intra- but not interglomerular presynaptic inhibition of olfactory sensory neurons. Neuron. 2005;48:1039–1053. doi: 10.1016/j.neuron.2005.10.031. [DOI] [PubMed] [Google Scholar]
- Mori et al., 2006.Mori K, Takahashi YK, Igarashi KM, Yamaguchi M. Maps of odorant molecular features in the mammalian olfactory bulb. Physiol Rev. 2006;86:409–433. doi: 10.1152/physrev.00021.2005. [DOI] [PubMed] [Google Scholar]
- Nakamura and Gold, 1987.Nakamura T, Gold GH. A cyclic nucleotide-gated conductance in olfactory receptor cilia. Nature. 1987;325:442–444. doi: 10.1038/325442a0. [DOI] [PubMed] [Google Scholar]
- Olsen and Wilson, 2008.Olsen SR, Wilson RI. Lateral presynaptic inhibition mediates gain control in an olfactory circuit. Nature. 2008;452:956–960. doi: 10.1038/nature06864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petzold et al., 2008.Petzold GC, Albeanu DF, Sato TF, Murthy VN. Coupling of neural activity to blood flow in olfactory glomeruli is mediated by astrocytic pathways. Neuron. 2008;58:897–910. doi: 10.1016/j.neuron.2008.04.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pifferi et al., 2006.Pifferi S, Boccaccio A, Menini A. Cyclic nucleotide-gated ion channels in sensory transduction. FEBS Lett. 2006;580:2853–2859. doi: 10.1016/j.febslet.2006.03.086. [DOI] [PubMed] [Google Scholar]
- Pírez and Wachowiak, 2008.Pírez N, Wachowiak M. In vivo modulation of sensory input to the olfactory bulb by tonic and activity-dependent presynaptic inhibition of receptor neurons. J Neurosci. 2008;28:6360–6371. doi: 10.1523/JNEUROSCI.0793-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rinberg et al., 2006.Rinberg D, Koulakov A, Gelperin A. Speed-accuracy tradeoff in olfaction. Neuron. 2006;51:351–358. doi: 10.1016/j.neuron.2006.07.013. [DOI] [PubMed] [Google Scholar]
- Schaefer and Margrie, 2007.Schaefer AT, Margrie TW. Spatiotemporal representations in the olfactory system. Trends Neurosci. 2007;30:92–100. doi: 10.1016/j.tins.2007.01.001. [DOI] [PubMed] [Google Scholar]
- Schafer et al., 2005.Schafer JR, Kida I, Rothman DL, Hyder F, Xu F. Adaptation in the rodent olfactory bulb measured by fMRI. Magn Reson Med. 2005;54:443–448. doi: 10.1002/mrm.20588. [DOI] [PubMed] [Google Scholar]
- Slotnick, 2007.Slotnick B. Odor-sampling time of mice under different conditions. Chem Senses. 2007;32:445–454. doi: 10.1093/chemse/bjm013. [DOI] [PubMed] [Google Scholar]
- Sobel et al., 2000.Sobel N, Khan RM, Hartley CA, Sullivan EV, Gabrieli JD. Sniffing longer rather than stronger to maintain olfactory detection threshold. Chem Senses. 2000;25:1–8. doi: 10.1093/chemse/25.1.1. [DOI] [PubMed] [Google Scholar]
- Spors and Grinvald, 2002.Spors H, Grinvald A. Spatio-temporal dynamics of odor representations in the mammalian olfactory bulb. Neuron. 2002;34:301–315. doi: 10.1016/s0896-6273(02)00644-x. [DOI] [PubMed] [Google Scholar]
- Spors et al., 2006.Spors H, Wachowiak M, Cohen LB, Friedrich RW. Temporal dynamics and latency patterns of receptor neuron input to the olfactory bulb. J Neurosci. 2006;26:1247–1259. doi: 10.1523/JNEUROSCI.3100-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uchida and Mainen, 2003.Uchida N, Mainen ZF. Speed and accuracy of olfactory discrimination in the rat. Nat Neurosci. 2003;6:1224–1229. doi: 10.1038/nn1142. [DOI] [PubMed] [Google Scholar]
- Vucinić et al., 2006.Vucinić D, Cohen LB, Kosmidis EK. Interglomerular center-surround inhibition shapes odorant-evoked input to the mouse olfactory bulb in vivo. J Neurophysiol. 2006;95:1881–1887. doi: 10.1152/jn.00918.2005. [DOI] [PubMed] [Google Scholar]
- Wachowiak and Cohen, 1999.Wachowiak M, Cohen LB. Presynaptic inhibition of primary olfactory afferents mediated by different mechanisms in lobster and turtle. J Neurosci. 1999;19:8808–8817. doi: 10.1523/JNEUROSCI.19-20-08808.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wachowiak and Cohen, 2001.Wachowiak M, Cohen LB. Representation of odorants by receptor neuron input to the mouse olfactory bulb. Neuron. 2001;32:723–735. doi: 10.1016/s0896-6273(01)00506-2. [DOI] [PubMed] [Google Scholar]
- Wachowiak and Shipley, 2006.Wachowiak M, Shipley MT. Coding and synaptic processing of sensory information in the glomerular layer of the olfactory bulb. Semin Cell Dev Biol. 2006;17:411–423. doi: 10.1016/j.semcdb.2006.04.007. [DOI] [PubMed] [Google Scholar]
- Wei et al., 1998.Wei J, Zhao AZ, Chan GC, Baker LP, Impey S, Beavo JA, Storm DR. Phosphorylation and inhibition of olfactory adenylyl cyclase by CaM kinase II in neurons: a mechanism for attenuation of olfactory signals. Neuron. 1998;21:495–504. doi: 10.1016/s0896-6273(00)80561-9. [DOI] [PubMed] [Google Scholar]
- Wesson et al., 2008.Wesson DW, Carey RM, Verhagen JV, Wachowiak M. Rapid encoding and perception of novel odors in the rat. PLoS Biol. 2008;6:e82. doi: 10.1371/journal.pbio.0060082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson, 1998.Wilson DA. Synaptic correlates of odor habituation in the rat anterior piriform cortex. J Neurophysiol. 1998;80:998–1001. doi: 10.1152/jn.1998.80.2.998. [DOI] [PubMed] [Google Scholar]
- Wilson and Linster, 2008.Wilson DA, Linster C. Neurobiology of a simple memory. J Neurophysiol. 2008;100:2–7. doi: 10.1152/jn.90479.2008. [DOI] [PubMed] [Google Scholar]
- Yan et al., 1995.Yan C, Zhao AZ, Bentley JK, Loughney K, Ferguson K, Beavo JA. Molecular cloning and characterization of a calmodulin-dependent phosphodiesterase enriched in olfactory sensory neurons. Proc Natl Acad Sci U S A. 1995;92:9677–9681. doi: 10.1073/pnas.92.21.9677. [DOI] [PMC free article] [PubMed] [Google Scholar]