

Abstract
Oxidative stress is associated with many diseases and has been found to induce DNA damage and cellular senescence. Numerous evidences support the detrimental effects of oxidative stress or cellular senescence on skeletal homeostasis. N-acetylcysteine (NAC) is a powerful antioxidant. However, it is unclear whether NAC can suppress orchiectomy (ORX)-induced osteoporosis by inhibiting oxidative stress and osteocyte senescence. In this study, ORX mice were supplemented with/without NAC, and were compared with each other and with sham-operated mice. Our results showed that NAC could prevent ORX-induced osteoporosis by inhibiting oxidative stress, DNA damage, osteocyte senescence and senescence-associated secretory phenotype (SASP), subsequently stimulating osteoblastic bone formation and inhibiting osteoclastic bone resorption. The results from this study suggest that NAC could be considered as a potential therapeutic agent for prevention and treatment of osteoporosis caused by testosterone deficiency.
Keywords: ORX-induced osteoporosis, N-acetylcysteine, oxidative stress, osteocyte senescence, SASP
Introduction
Osteoporosis is one of the most common clinical problems, characterized by decreased bone mass, worsening bone microarchitecture, increased risk of fracture and other complications that are associated with significant morbidity and mortality, affects both men and women [1]. Compared with women, male osteoporosis occurs later and is less prone to fractures [2,3]. However, men suffer from much higher morbidity and mortality post-fracture, and the secondary causes of osteoporosis in men are more common (about 50%) compared with osteoporosis in women. Excessive alcohol and caffeinated beverages, cigarette-smoking, excessive glucocorticoids and hypogonadism are the most frequent causes [4,5]. Therefore, it is still necessary to elucidate the pathogenesis of male osteoporosis.
Aging is closely related to osteoporosis. The loss of cancellous bone mass in human begins in the third decade, and cortical bone begins to decline after the age of 50 [6]. With aging, changes in sex steroid levels appear to be a major cause of osteoporosis in men. The amount of bone resorbed by the osteoclasts is not fully restored in the aging skeleton and the defective osteoblast number has been attributed to bone loss as a result of sex hormone suppression [7]. It has been reported that a decrease in bone mineral density is associated with a decrease in blood testosterone levels, and prostate cancer patients show a reduction in bone mass after receiving hormone deprivation therapy [8,9]. Although testosterone maintains bone mass in men, the use of testosterone-replacement therapy to prevent and treat male osteoporosis remains controversial [10]. Thus, an alternative treatment may be a viable option to prevent deterioration of male bone health.
Oxidative stress plays a role in a number of degenerative conditions including osteoporosis. Cumulating evidence from animal studies support the detrimental effect of oxidative stress on skeletal homeostasis. The production of oxidative stress in cultured bone is associated with the formation of new osteoclasts and enhanced bone resorption [11]. Increased ROS levels, decreased GSR activity were observed in 5-month-old females or males develop osteoporosis after gonadal resection [12]. Mice deficient in Cu,Zn-superoxide dismutase (SOD1) have elevated oxidative stress and decreased bone stiffness and strength compared to wild-type mice [13]. Consistently, in a number of animal studies, dietary antioxidant supplementation is an effective approach to mitigate and delay bone loss [14-17].
It is generally believed that excessive accumulation of ROS causes damage to the cell membrane and cytoplasm, this finally triggers DNA damage response and activation of cellular senescence [18]. Growing evidence for a critical role of cellular senescence in mediating osteoporosis were reported. Evidence in support of this was provided by a positive correlation between 8-hydroxy-2-deoxyguanosine, a widely used marker of DNA damage, and RANKL/OPG ratio in the osteoporotic patients [19]. Compared with 7-month-old mice, 21-month-old mice showed increased osteocyte expression of γH2AX, p16Ink4a and several SASP markers [20]. Consistent with the result of mice, the expression of p16Ink4a and SASP factors was significantly higher in old women (mean age, 78 years) than in young women (mean age, 27 years) [21].
N-acetylcysteine (NAC), as a scavenger of free radicals and as an antioxidant, has been commonly used to prevent and treat many diseases. It has been demonstrated that NAC was as effective as dihydrotestosterone in preventing the decrease of spinal BMD in ORX mice [12]. However, it is unclear whether reversed ORX-induced bone loss by antioxidant NAC treatment was associated with inhibition of DNA damage and cellular senescence. To answer this question, we established an orchiectomy (ORX) mouse model on both sides and then supplemented the ORX mice with NAC in drinking water. The control mice were sham operated and fed with normal water. We examined the alterations in bone microarchitecture, bone formation, bone turnover, oxidative stress, DNA damage, cell senescence, senescence-associated secretory phenotype (SASP), osteoblast and osteoclast activity of these mice to study whether NAC can inhibit the ORX-induced bone loss by inhibiting oxidative stress, DNA damage and cell senescence.
Materials and methods
Mice
Three-week-old male C57/BL6J mice were purchased from Vital River Laboratory Animal Technology Co. Ltd. (Beijing, China). They were maintained in a virus- and parasite-free barrier facility and exposed to a 12-h light, 12-h dark cycle. All the mice were randomly divided into three groups (n=10 per group): 1) sham surgery; 2) ORX group; 3) ORX group supplemented with 1 mg/ml N-acetylcysteine (NAC) in the drinking water from 3-week-old to 16-week-old. Mice were anesthetized with chloral hydrate and orchiectomized (ORX) at both sides when eight weeks old. Sham operations were carried out independently. All mice were sacrificed at 16 weeks of age, 8 weeks after surgery. This study was approved by the Institutional Animal Care and Use Committee of Nanjing Medical University.
X-ray and Microcomputed tomography (Micro-CT)
After sacrifice, the right femurs were removed and fixed in PLP fixative (2% paraformaldehyde containing 0.075 M lysine and 0.01 M sodium periodate). These samples were then scanned for X-ray and micro-CT. Radiographs were performed. Same samples were then scanned on a micro-CT scanner (Sky Scan 1072 Scanner) using energy of 100 kV, and 98 µA intensity. Three-dimensional (3D) images were generated using the 3D Creator software supplied with the instrument as described previously [22].
Histology
The tibias were removed and dissected free of soft tissue, fixed with 2% PLP fixative at 4°C and processed histologically. Then the tibias were decalcified in 14% EDTA glycerol solution for 5 to 7 days at 4°C. The decalcified tibias were dehydrated and embedded in paraffin for paraffin sections, after which 5 μm sections were cut on a rotary microtome. The sections were stained with hematoxylin and eosin (H&E) or histochemically for total collagen with sirius red or alkaline phosphatase (ALP) activity, or tartrate resistant acid phosphatase (TRAP) activity as described previously [22] or immunohistochemically as described below.
Immunohistochemical staining
Immunohistochemical staining was carried out for type I collagen, phosphorylation of histone H2AX on Ser139 (γ-H2AX), β-galactosidase (β-gal), p16INK4a, Interleukin (IL)-1α, IL-1β and TNF-α, using the avidin-biotin-peroxidase complex technique with affinity-purified goat anti-rabbit type I collagen antibody (Southern Biotechnology Associates, Birmingham, AL, USA), γ-H2AX (Cell Signaling Technology, MA, USA), β-gal antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA), p16INK4a (Abcam, MA, USA), IL-1α (Abcam, MA, USA), IL-1β (Abcam, MA, USA) and TNF-α (Abcam, MA, USA), following previously-described methods [22].
Detection of ROS levels
The bone marrow cells from long bones were converted into single cell suspensions with syringe, then washed with cold PBS, and resuspended in binding buffer, then 106 cells per sample were incubated with 5 µL 2’,7’-dichlorofluorescein diacetate (DCFH-DA, Sigma-Aldrich, St. Louis, MO, USA) for 30 mins in the dark followed by incubation with 10% FBS for 20 mins at 37°C. ROS levels were calculated from mean fluorescence intensity (MFI) measured using a flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA).
Western blotting
Protein was extracted from the left femur of each group of mice. Immunoblotting was performed as previously described [23]. Primary antibodies against SOD1 (Abcam, MA, USA), SOD2 (Novus Biological, Centennial, CO, USA), Prdx 4 (Santa Cruz Biotechnology, Santa Cruz, CA, USA), γ-H2AX (Cell Signaling Technology, MA, USA) and p16INK4a (Abcam, MA, USA) were used. And β-actin (Bioworld Technology, St. Louis Park MN, USA) was used as a loading control. Immunoreactive bands were visualized with ECL chemiluminescence (Amersham Biosciences, Chalfont St. Giles, UK) and analysed by Scion Image Beta 4.02 (Scion, National Institutes of Health, Bethesda, MD, USA).
RNA isolation and quantitative real-time RT-PCR
RNA was isolated from left tibias with TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s protocol. Reverse transcription reactions were performed with the SuperScript First-Strand Synthesis System (Invitrogen) as previously described [22]. To determine the relative expression of genes of interest, quantitative real-time RT-PCR was carried out in an Applied Biosystems Cycler with a SYBR Green PCR reagent kit. The PCR primers were used as described previously [22]. GAPDH was used as the internal control for each reaction. All primers were tested for their specificity by conventional PCR before being used for quantitative analysis by real-time RT-PCR. All PCRs were performed in triplicate. Results were analysed with SDS 7300 software, and the relative amount of mRNA was calculated after normalization for GAPDH mRNA.
Computer-assisted image analysis
After histochemical or immunohistochemical staining of sections from mice of each group, images of micrographs from single sections were digitally recorded using a rectangular template, and recordings were processed and analysed using Northern Eclipse image analysis software.
Statistical analysis
All analyses were performed using SPSS software, version 16.0 (SPSS Inc., Chicago, IL, USA). Measured data are presented as mean ± SEM. Statistical comparisons were performed using a one-way ANOVA of qualitative data to compare differences between groups. P values < 0.05 were considered statistically significant.
Results
The effect of NAC on ORX-induced osteoporosis
To verify whether NAC supplementation can prevent male osteoporosis, we used orchiectomized (ORX) mice as an animal model. The mice were then fed with or without NAC for 8 weeks, and the bone phenotype from each group of mice was compared using X-ray, micro-CT and total collagen staining, respectively. The data showed a significant decrease in bone volume, bone mineral density (Figure 1A, 1B, 1D, 1G), total collagen positive area (Figure 1C and 1I), trabecular thickness (Figure 1E) and number (Figure 1F) and an increase in trabecular separation (Figure 1H) in ORX mice compared to sham-operated mice. However, these parameters were rescued in ORX mice by NAC supplementation. These findings indicate that supplementation with NAC can prevent ORX-induced osteoporosis.
Figure 1.
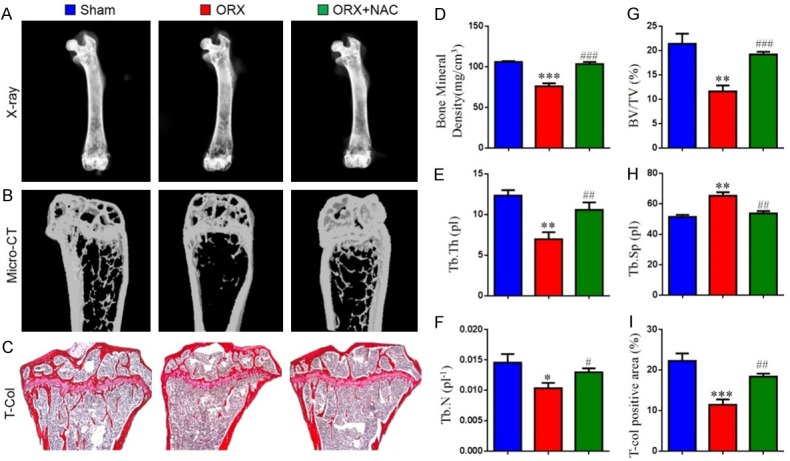
The effect of NAC on ORX-induced osteoporosis. Eight-week-old male C57/BL6J mice were randomly performed ORX or Sham surgery and euthanize after 8 weeks. (A) Representative radiographs of femurs. (B) Representative Micro-CT-scanned and 3D reconstructed sections along the longitudinal direction of femurs. (C) Representative micrographs of paraffin sections of tibias were stained histochemically for total collagen. Analysis of the distal femoral trabecular bone parameters by micro-CT, (D) Bone mineral density, (E) Thickness of trabecular bone (Tb.Th), (F) Trabecular number (Tb.N), (G) Trabecular bone volume relative to tissue volume (BV/TV, %), (H) Trabecular separation (Tb.Sp). (I) Total collagen positive area (%). Data are presented as the mean ± SEM of determinations, each data-point was the mean of five specimens. *P < 0.05, **P < 0.01, ***P < 0.001, versus sham mice; #P < 0.05, ##P < 0.01, ###P < 0.001 versus ORX mice. (C) Magnification, 50×.
The effect of NAC on the osteoblastic bone formation in ORX mice
To determine whether ORX-induced osteoporosis rescued by NAC was associated with increased osteoblastic bone formation, we examined the parameters of osteoblastic bone formation in paraffin-embedded sections of mice by staining with H&E, histochemically for ALP and immunohistochemically for type I collagen. The number of osteoblasts, ALP-positive surface and the type I collagen-positive area were determined by computer-assisted image analysis. We found that the number of osteoblasts (Figure 2A and 2D), the ALP-positive surface as a percentage of the bone perimeter (Figure 2B and 2E) and the type I collagen-positive area (Figure 2C and 2F) were reduced significantly in ORX mice compared with sham mice. However, these parameters were largely rescued in ORX mice with NAC supplementation. Therefore, NAC could increase osteoblastic bone formation in ORX mice.
Figure 2.

The effect of NAC on the osteoblastic bone formation in ORX mice. Representative photomicrographs of paraffin sections of tibias from 16-week-old mice of each group stained for (A) HE, histochemically for (B) ALP and immunohistochemically for (C) type I collagen. (D) The number of osteoblasts (N.Ob) per mm bone perimeter (B.Pm) was measured and presented. (E) ALP-positive surface as a percentage of the B.Pm in the bone (%). (F) The percentage of the Col I-immunopositive area was measured and presented by image analysis (%). (A, C) Magnification, 200×. (B) Magnification, 400×. Data are presented as the mean ± SEM of determinations, each data-point was the mean of five specimens. *P < 0.05, ***P < 0.001, versus sham mice; #P < 0.05, ##P < 0.01 versus ORX mice.
The effect of NAC on the osteoclastic bone resorption in ORX mice
To investigate whether ORX-induced osteoporosis rescued by NAC was associated with decreased osteoclastic bone resorption, the parameters of osteoclastic bone resorption were assessed by histochemical staining for TRAP, image analysis and real-time RT-PCR. The number of osteoclasts (Figure 3A and 3B), TRAP-positive osteoclast surface (Figure 3C), RANKL/OPG ratio (Figure 3E) and RANKL (Figure 3D) gene expression levels were increased in ORX mice compared with sham mice. However, NAC supplementation could normalize these changes. Our data demonstrated that NAC could inhibit osteoclastic bone resorption in ORX mice.
Figure 3.
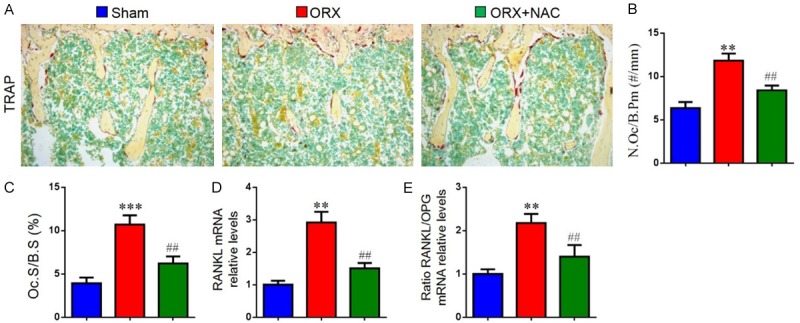
The effect of NAC on the osteoclastic bone resorption in ORX mice. (A) Representative photomicrographs of paraffin sections of tibias from 16-week-old mice of each group stained histochemically for TRAP. (B) The number of TRAP-positive osteoclasts (N.Oc) per mm bone perimeter (B.Pm) was measured and presented. (C) Osteoclast surface relative to bone surface (Oc.S/BS, %). Real-time RT-PCR was performed on bone tissue extracts from 16-week-old mice of each group. Gene expression of (D) RANKL, (E) RANKL/OPG ratio are shown. Messenger RNA expression, assessed by real-time RT-PCR analysis, was calculated as a ratio to the GAPDH mRNA level and expressed relative to levels in sham mice. Data are presented as the mean ± SEM of determinations, each data point is the mean of five specimens. **P < 0.01, ***P < 0.001, versus sham mice; ##P < 0.01 versus ORX mice. (A) Magnification, 200×.
The effect of NAC on redox balance in ORX mice
To investigate whether the effect of NAC on ORX-induced osteoporosis was associated with regulation of oxidative stress, the ROS levels and the protein expression of antioxidant enzymes in long bone were measured. Results revealed that ROS levels (Figure 4A and 4B) were increased markedly, whereas the protein expression levels (Figure 4C-F) of superoxide dismutase 1 (SOD1), superoxide dismutase 2 (SOD2) and peroxiredoxin 4 (Prdx4) were reduced significantly in ORX mice compared with sham mice. However, these parameters were largely rescued in ORX mice with NAC supplementation. These results indicated that NAC may prevent ORX-induced osteoporosis by inhibiting oxidative stress.
Figure 4.
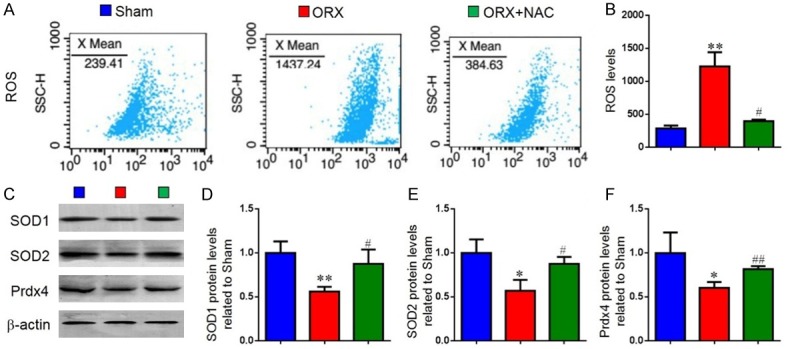
The effect of NAC on redox balance in ORX mice. (A) Representative flow cytometric analysis of ROS levels of bone marrow cells from sham, ORX and ORX+NAC mice. (B) Relative fluorescence intensity (RFI) of ROS was calculated and expressed relative to the sham mice. (C) Representative western blots of bone tissue extracts showing expression of superoxide dismutase1 (SOD1), superoxide dismutase2 (SOD2) and peroxiredoxin 4 (Prdx4); β-actin was used as loading control for the western blots in the sham, ORX and ORX+NAC groups. (D) SOD1, (E) SOD2, (F) Prdx4 protein levels relative to β-actin protein levels were assessed by densitometric analysis and expressed relative to levels in sham-operated mice. Data are presented as the mean ± SEM of determinations, each data-point was the mean of five specimens. *P < 0.05, **P < 0.01 versus sham mice. #P < 0.05, ##P < 0.01 versus ORX mice.
The effect of NAC on DNA damage and osteocyte senescence in ORX mice
To determine the effect of NAC on DNA damage and osteocyte senescence in ORX mice, the DNA damage markers, γ-H2AX, and the cellular senescence markers, β-galactosidase(β-gal) and p16INK4a, were examined by Western blot or immunohistochemical staining in paraffin-embedded sections of long bone. We found that the percentages of osteocytes positive for γ-H2AX (Figure 5A and 5B), p16 (Figure 5D and 5E), β-gal (Figure 5G and 5H) and the protein expression levels of γ-H2AX, p16 (Figure 5C, 5F, 5I) were elevated in ORX mice compared with sham-operated mice. However, these parameters were down-regulated dramatically in NAC-treated ORX mice. These data showed that testosterone deficiency could induce DNA damage and osteocyte senescence whereas NAC supplementation could inhibit DNA damage and osteocyte senescence in ORX mice.
Figure 5.
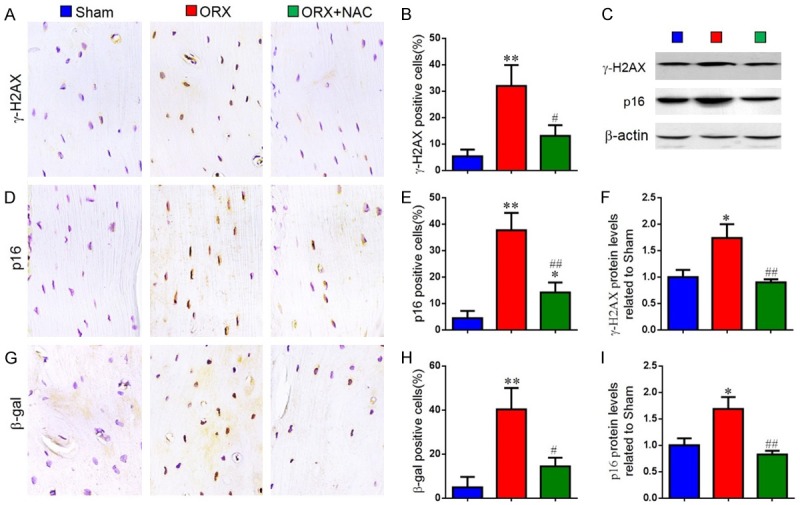
The effect of NAC on DNA damage and osteocyte senescence in ORX mice. Representative micrographs of paraffin sections of tibias from 16-week-old mice of each group stained immunohistochemically for (A) γ-H2AX, (D) p16INK4a and (G) β-gal. The percentages of (B) γ-H2AX-positive, (E) p16INK4a-positive and (H) β-gal-positive cells were determined by image analysis. (C) Representative western blots of bone tissue extracts showing expression of γ-H2AX and p16INK4a; β-actin was used as loading control for the western blots in the sham, ORX and ORX+NAC groups. (F) γ-H2AX and (I) p16INK4a protein levels relative to β-actin protein levels were assessed by densitometric analysis and expressed relative to levels in sham-operated mice. Data are presented as the mean ± SEM of determinations, each data-point was the mean of five specimens. *P < 0.05, **P < 0.01 versus sham mice. #P < 0.05, ##P < 0.01 versus ORX mice. (A, D, G) Magnification, 400×.
The effect of NAC on senescence-associated secretory phenotype(SASP) in ORX mice
Senescent cells can secrete multiple cytokines after loss of proliferative capacity, that is senescence-associated secretory phenotype (SASP), including IL-1α, IL-1β, IL-6, IL-8, Mmp-3, Mmp-13, TNF-α and TGF-β. To verify if NAC supplementation could prevent SASP production in ORX mice, we examined the alterations of SASP by immunohistochemistry and real-time RT-PCR. Compared with the sham-operated mice, the percentage of osteocytes positive for IL-1α (Figure 6A and 6B), IL-1β (Figure 6D and 6E), TNF-α (Figure 6G and 6H) and the gene expression levels of IL-1α (Figure 6C), IL-1β (Figure 6F), IL-6 (Figure 6I) were all upregulated dramatically in ORX mice. However, they were markedly rescued in ORX mice by NAC supplementation. These results indicate that testosterone deficiency could increase the production of SASP, while NAC supplementation could inhibit SASP production.
Figure 6.
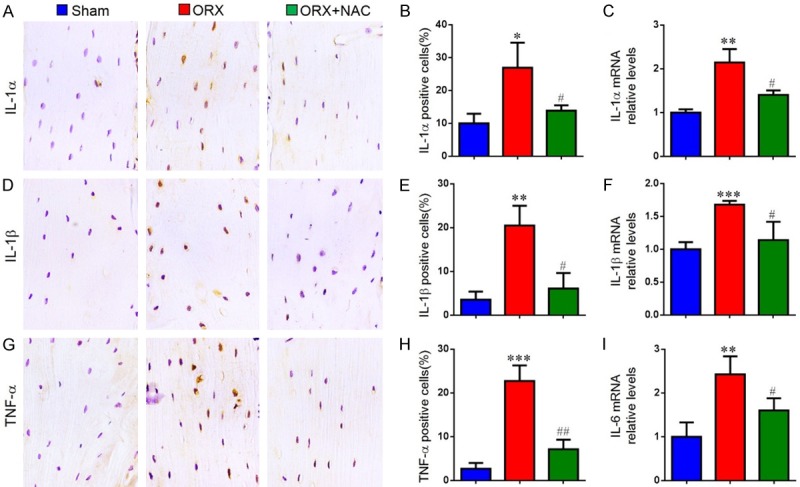
The effect of NAC on senescence-associated secretory phenotype(SASP) in ORX mice. Representative micrographs of paraffin sections of tibias from 16-week-old mice of each group stained immunohistochemically for (A) IL-1α, (D) IL-1β and (G) TNF-α. The percentages of (B) IL-1α-positive, (E) IL-1β-positive and (H) TNF-α-positive cells were determined by image analysis. Real-time RT-PCR was performed on extracts of tibias from 16-week-old mice of each group to investigate gene expression of (C) IL-1α, (F) IL-1β and (I) IL-6. Messenger RNA expression, assessed by real-time RT-PCR analysis, was calculated as a ratio to the GAPDH mRNA level and shown relative to levels in sham mice. Data are presented as the mean ± SEM of determinations, each data-point is the mean of five specimens. *P < 0.05, **P < 0.01, ***P < 0.001 versus sham mice. #P < 0.05, ##P < 0.01 versus ORX mice. (A, D, G) Magnification, 400×.
Discussion
Male osteoporosis is now increasingly viewed as an important health care problem to society because of higher morbidity and mortality rate after fracture than women. As aging, changes in sex steroid levels appear to be the leading cause of osteoporosis in men. But there is only limited preventive and therapeutic drug for male osteoporosis [24,25]. Therefore, elucidating the osteoporotic mechanism and finding effective therapies for male osteoporosis is especially essential. In our present study, we used the ORX mouse model to study the possible mechanism of male osteoporosis and the effect of NAC on the male osteoporosis. Firstly, in order to track changes of bone mass in ORX mice, we confirmed that ORX resulted in bone loss by X-ray, Micro-CT scanning, three-dimensional (3D) reconstruction and total collagen staining, which is consistent with previous studies [12,16]. Simultaneously, we demonstrated that NAC supplementation rescued bone loss induced by ORX.
Studies have shown that rapid bone loss in men is more common in the absence of testosterone or estradiol [26]. Androgen receptors have been found to be expressed in osteoblasts, osteoclasts, osteocytes and bone marrow mesenchymal cells. Androgen may directly affect the function of all these bone-related cells [27,28]. It has also been found that the number of osteoclasts was increased in the lumbar spine of androgen receptor knockout mouse model [29]. Unlike women who lose trabecular bone with age, men show more trabecular thinning, which is thought to be secondary to reduced bone formation and correlated with indices of reduced bone formation [30,31]. Thus, we then examined alterations of osteoblastic bone formation and osteoclastic bone resorption in ORX mice. Our results demonstrated that ORX not only reduced osteoblastic bone formation but also increased osteoclastic bone resorption. However, the bone formation of osteoblasts and bone resorption of osteoclasts were normalized in ORX mice by the supplementation of NAC. Our results suggest that NAC could prevent ORX-induced osteoporosis by inhibiting osteoclastic bone resorption and stimulating osteoblastic bone formation.
There is growing evidence that increased reactive oxygen species (ROS) and decreased antioxidant defence mechanisms lead to estrogen deficiency-induced bone loss [32-34]. Then, we asked whether osteoporosis caused by ORX was associated with increased oxidative stress. Recent studies showed that oxidative stress induced by testosterone deficiency is associated with decreased levels of antioxidant enzymes and activity in mouse vertebral tissue, which leads to decreased osteoblast formation and increased osteoclast resorption [12,16]. In agreement with previous findings, our current study confirmed that ORX could induce oxidative stress and reduced antioxidants levels which support that decreased osteoblast formation and increased osteoclast resorption by ORX may be partly related to oxidative stress. Other researchers have shown that inhibition of H2O2 by antioxidants, NAC, ascorbic acid, pegylated catalase or PQQ can prevent bone loss caused by acute loss of estrogen [14,35-37]. Then, we asked whether NAC could exert an anti-osteoporosis role in ORX mice through inhibiting oxidative stress. Thus, we gave the ORX mice with the antioxidant NAC in the drinking water for 8 weeks and demonstrated that NAC supplementation effectively prevented ORX-induced bone loss by defending against oxidative stress.
Age-related oxidative stress is associated with many diseases, and oxidative stress has been found to induce DNA damage and cellular senescence [38]. Numerous studies have shown that inhibition of cellular senescence may be an effective therapy for bone loss. In vitro studies have shown that senescent-cell conditioned medium impairs osteoblast mineralization and enhances osteoclast progenitor cell survival, resulting in increased osteoclastogenesis [39]. It has been proposed to target senescent cells pharmacologically in vivo to eliminate them and thus delay the emergence of osteoporosis [40]. In the current study, the DNA damage markers, γ-H2AX, and the cellular senescence markers, β-gal and p16INK4a, were increased in the osteocytes of ORX mice. These results suggested that increased oxidative stress in the bone of ORX mice can trigger DNA damage and osteocyte senescence.
Senescent cells can exhibit a complex pro-inflammatory response called the senescence-associated secretory phenotype (SASP), which allows them to communicate with other cells and the microenvironment, stimulating the senescence of adjacent cells [41]. The evidence from recent study showed that clearance of the senescent cells in aged (20 to 22 months old) mice with established bone loss inhibited the production of the proinflammatory secretome of senescent cells, while also resulting in higher bone mass, strength and better bone microarchitecture [42]. Therefore, we asked if testosterone deficiency-induced osteoporosis prevented by supplementation of NAC is associated with reduced DNA damage, osteocyte senescence and SASP induced by ORX. To answer this question, we compared the alterations of DNA damage, osteocyte senescence and SASP between ORX mice and ORX mice supplemented with NAC. Our results found that supplementation of NAC could prevent DNA damage, osteocyte senescence and SASP induced by ORX.
In conclusion, our present study exhibited that testosterone deficiency-induced osteoporosis was not only related with reduced osteoblastic bone formation, increased osteoclastic bone resorption and oxidative stress, but also related with increased DNA damage, osteocyte senescence and SASP. Supplementation of NAC played a preventive and protective role in ORX-induced osteoporosis by inhibiting osteoclastic bone resorption and oxidative stress, preventing DNA damage, osteocyte senescence and SASP production, and promoting osteoblastic bone formation. Therefore, our findings from this research provided experimental evidence for the clinical application of NAC, indicating that NAC could be used to prevent and treat osteoporosis in men.
Acknowledgements
This work was supported by grants from the National Natural Science Foundation of China (81500682 to L.C.), from the Postdoctoral Science Foundation of China (2018M642279 to L.C.) and from Jiangsu Planned Projects for Postdoctoral Research Funds.
Disclosure of conflict of interest
None.
References
- 1.Vidal M, Thibodaux RJ, Neira LFV, Messina OD. Osteoporosis: a clinical and pharmacological update. Clinical Rheumatology. 2019;38:385–395. doi: 10.1007/s10067-018-4370-1. [DOI] [PubMed] [Google Scholar]
- 2.Cawthon PM. Gender differences in osteoporosis and fractures. Clin Orthop Relat Res. 2011;469:1900–5. doi: 10.1007/s11999-011-1780-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Jung Y, Ko Y, Kim HY, Ha YC, Lee YK, Kim TY, Choo DS, Jang S. Gender differences in anti-osteoporosis drug treatment after osteoporotic fractures. J Bone Miner Metab. 2019;37:134–141. doi: 10.1007/s00774-018-0904-5. [DOI] [PubMed] [Google Scholar]
- 4.Chatterton BD, Moores TS, Ahmad S, Cattell A, Roberts PJ. Cause of death and factors associated with early in-hospital mortality after hip fracture. Bone Joint J. 2015;97-B:246–51. doi: 10.1302/0301-620X.97B2.35248. [DOI] [PubMed] [Google Scholar]
- 5.Willson T, Nelson SD, Newbold J, Nelson RE, LaFleur J. The clinical epidemiology of male osteoporosis: a review of the recent literature. Clin Epidemiol. 2015;7:65–76. doi: 10.2147/CLEP.S40966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Riggs BL, Melton LJ, Robb RA, Camp JJ, Atkinson EJ, McDaniel L, Amin S, Rouleau PA, Khosla S. A population-based assessment of rates of bone loss at multiple skeletal sites: evidence for substantial trabecular bone loss in young adult women and men. J Bone Miner Res. 2008;23:205–14. doi: 10.1359/JBMR.071020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Walsh JS, Eastell R. Osteoporosis in men. Nat Rev Endocrinol. 2013;9:637–45. doi: 10.1038/nrendo.2013.171. [DOI] [PubMed] [Google Scholar]
- 8.Stathopoulos IS, Ballas EG, Lampropoulou-Adamidou K, Trovas G. A review on osteoporosis in men. Hormones. 2014;13:441–457. doi: 10.14310/horm.2002.1550. [DOI] [PubMed] [Google Scholar]
- 9.Lassemillante AC, Doi SA, Hooper JD, Prins JB, Wright OR. Prevalence of osteoporosis in prostate cancer survivors: a meta-analysis. Endocrine. 2014;45:370–381. doi: 10.1007/s12020-013-0083-z. [DOI] [PubMed] [Google Scholar]
- 10.Kamel HK. Male osteoporosis: new trends in diagnosis and therapy. Drugs Aging. 2005;22:741–8. doi: 10.2165/00002512-200522090-00003. [DOI] [PubMed] [Google Scholar]
- 11.Garrett IR, Boyce BF, Oreffo RO, Bonewald L, Poser J, Mundy GR. Oxygen-derived free radicals stimulate osteoclastic bone resorption in rodent bone in vitro and in vivo. J Clin Invest. 1990;85:632–639. doi: 10.1172/JCI114485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Almeida M, Han L, Martin-Millan M, Plotkin LI, Stewart SA, Roberson PK, Kousteni S, O’Brien CA, Bellido T, Parfitt AM, Weinstein RS, Jilka RL, Manolagas SC. Skeletal involution by age-associated oxidative stress and its acceleration by loss of sex steroids. J Biol Chem. 2007;282:27285–27297. doi: 10.1074/jbc.M702810200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Smietana MJ, Arruda EM, Faulkner JA, Brooks SV, Larkin LM. Reactive oxygen species on bone mineral density and mechanics in Cu,Zn superoxide dismutase (Sod1) knockout mice. Biochem Biophys Res Commun. 2010;403:149–153. doi: 10.1016/j.bbrc.2010.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Geng Q, Gao H, Yang R, Guo K, Miao D. Pyrroloquinoline quinone prevents estrogen deficiency-induced osteoporosis by inhibiting oxidative stress and osteocyte senescence. Int J Biol Sci. 2019;15:58–68. doi: 10.7150/ijbs.25783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hwang YH, Kim KJ, Kim SJ, Mun SK, Hong SG, Son YJ, Yee ST. Suppression effect of astaxanthin on osteoclast formation in vitro and bone loss in vivo. Int J Mol Sci. 2018;19 doi: 10.3390/ijms19030912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wu X, Li J, Zhang H, Wang H, Yin G, Miao D. Pyrroloquinoline quinone prevents testosterone deficiency-induced osteoporosis by stimulating osteoblastic bone formation and inhibiting osteoclastic bone resorption. Am J Transl Res. 2017;9:1230–1242. [PMC free article] [PubMed] [Google Scholar]
- 17.Deng L, Ding Y, Peng Y, Wu Y, Fan J, Li W, Yang R, Yang M, Fu Q. gamma-Tocotrienol protects against ovariectomy-induced bone loss via mevalonate pathway as HMG-CoA reductase inhibitor. Bone. 2014;67:200–207. doi: 10.1016/j.bone.2014.07.006. [DOI] [PubMed] [Google Scholar]
- 18.Droge W. Free radicals in the physiological control of cell function. Physiol Rev. 2002;82:47–95. doi: 10.1152/physrev.00018.2001. [DOI] [PubMed] [Google Scholar]
- 19.Cervellati C, Romani A, Cremonini E, Bergamini CM, Fila E, Squerzanti M, Greco P, Massari L, Bonaccorsi G. Higher urinary levels of 8-hydroxy-2’-deoxyguanosine are associated with a worse RANKL/OPG ratio in postmenopausal women with osteopenia. Oxid Med Cell Longev. 2016;2016:6038798. doi: 10.1155/2016/6038798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Piemontese M, Almeida M, Robling AG, Kim HN, Xiong J, Thostenson JD, Weinstein RS, Manolagas SC, O’Brien CA, Jilka RL. Old age causes de novo intracortical bone remodeling and porosity in mice. JCI Insight. 2017;2 doi: 10.1172/jci.insight.93771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Farr JN, Fraser DG, Wang H, Jaehn K, Ogrodnik MB, Weivoda MM, Drake MT, Tchkonia T, LeBrasseur NK, Kirkland JL, Bonewald LF, Pignolo RJ, Monroe DG, Khosla S. Identification of senescent cells in the bone microenvironment. J Bone Miner Res. 2016;31:1920–1929. doi: 10.1002/jbmr.2892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Qin R, Sun J, Wu J, Chen L. Pyrroloquinoline quinone prevents knee osteoarthritis by inhibiting oxidative stress and chondrocyte senescence. Am J Transl Res. 2019;11:1460–1472. [PMC free article] [PubMed] [Google Scholar]
- 23.Chen L, Yang R, Qiao W, Zhang W, Chen J, Mao L, Goltzman D, Miao D. 1,25-Dihydroxyvitamin D exerts an antiaging role by activation of Nrf2-antioxidant signaling and inactivation of p16/p53-senescence signaling. Aging Cell. 2019;18:e12951. doi: 10.1111/acel.12951. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 24.Farahmand P, Spiegel R, Ringe JD. Male osteoporosis. Z Rheumatol. 2016;75:459–465. doi: 10.1007/s00393-016-0078-2. [DOI] [PubMed] [Google Scholar]
- 25.Giusti A, Bianchi G. Treatment of primary osteoporosis in men. Clin Interv Aging. 2015;10:105–115. doi: 10.2147/CIA.S44057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Fink HA, Ewing SK, Ensrud KE, Barrett-Connor E, Taylor BC, Cauley JA, Orwoll ES. Association of testosterone and estradiol deficiency with osteoporosis and rapid bone loss in older men. J Clin Endocrinol Metab. 2006;91:3908–3915. doi: 10.1210/jc.2006-0173. [DOI] [PubMed] [Google Scholar]
- 27.Seeman E. Estrogen, androgen, and the pathogenesis of bone fragility in women and men. Curr Osteoporos Rep. 2004;2:90–96. doi: 10.1007/s11914-004-0016-0. [DOI] [PubMed] [Google Scholar]
- 28.Noble B, Routledge J, Stevens H, Hughes I, Jacobson W. Androgen receptors in bone-forming tissue. Horm Res. 1999;51:31–36. doi: 10.1159/000023310. [DOI] [PubMed] [Google Scholar]
- 29.Pansini F, Mollica G, Bergamini CM. Management of the menopausal disturbances and oxidative stress. Curr Pharm Des. 2005;11:2063–2073. doi: 10.2174/1381612054065819. [DOI] [PubMed] [Google Scholar]
- 30.Sinnesael M, Claessens F, Boonen S, Vanderschueren D. Novel insights in the regulation and mechanism of androgen action on bone. Curr Opin Endocrinol Diabetes Obes. 2013;20:240–244. doi: 10.1097/MED.0b013e32835f7d04. [DOI] [PubMed] [Google Scholar]
- 31.Khosla S, Riggs BL, Atkinson EJ, Oberg AL, McDaniel LJ, Holets M, Peterson JM, Melton LJ 3rd. Effects of sex and age on bone microstructure at the ultradistal radius: a population-based noninvasive in vivo assessment. J Bone Miner Res. 2006;21:124–31. doi: 10.1359/JBMR.050916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Rana K, Davey RA, Zajac JD. Human androgen deficiency: insights gained from androgen receptor knockout mouse models. Asian J Androl. 2014;16:169–177. doi: 10.4103/1008-682X.122590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Cervellati C, Bonaccorsi G, Cremonini E, Bergamini CM, Patella A, Castaldini C, Ferrazzini S, Capatti A, Picarelli V, Pansini FS, Massari L. Bone mass density selectively correlates with serum markers of oxidative damage in post-menopausal women. Clin Chem Lab Med. 2013;51:333–338. doi: 10.1515/cclm-2012-0095. [DOI] [PubMed] [Google Scholar]
- 34.Cervellati C, Bergamini CM. Oxidative damage and the pathogenesis of menopause related disturbances and diseases. Clin Chem Lab Med. 2016;54:739–753. doi: 10.1515/cclm-2015-0807. [DOI] [PubMed] [Google Scholar]
- 35.Lean JM, Davies JT, Fuller K, Jagger CJ, Kirstein B, Partington GA, Urry ZL, Chambers TJ. A crucial role for thiol antioxidants in estrogen-deficiency bone loss. J Clin Invest. 2003;112:915–923. doi: 10.1172/JCI18859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Jagger CJ, Lean JM, Davies JT, Chambers TJ. Tumor necrosis factor-alpha mediates osteopenia caused by depletion of antioxidants. Endocrinology. 2005;146:113–118. doi: 10.1210/en.2004-1058. [DOI] [PubMed] [Google Scholar]
- 37.Huang Y, Chen N, Miao D. Effect and mechanism of pyrroloquinoline quinone on anti-osteoporosis in Bmi-1 knockout mice-Anti-oxidant effect of pyrroloquinoline quinone. Am J Transl Res. 2017;9:4361–4374. [PMC free article] [PubMed] [Google Scholar]
- 38.Rai P, Onder TT, Young JJ, McFaline JL, Pang B, Dedon PC, Weinberg RA. Continuous elimination of oxidized nucleotides is necessary to prevent rapid onset of cellular senescence. Proc Natl Acad Sci U S A. 2009;106:169–174. doi: 10.1073/pnas.0809834106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Farr JN, Xu M, Weivoda MM, Monroe DG, Fraser DG, Onken JL, Negley BA, Sfeir JG, Ogrodnik MB, Hachfeld CM, LeBrasseur NK, Drake MT, Pignolo RJ, Pirtskhalava T, Tchkonia T, Oursler MJ, Kirkland JL, Khosla S. Targeting cellular senescence prevents age-related bone loss in mice. Nat Med. 2017;23:1072–1079. doi: 10.1038/nm.4385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tachikart Y, Malaise O, Constantinides M, Jorgensen C, Brondello JM. Senescent cells: a target in osteo-articular diseases. Med Sci (Paris) 2018;34:547–553. doi: 10.1051/medsci/20183406014. [DOI] [PubMed] [Google Scholar]
- 41.Acosta JC, Banito A, Wuestefeld T, Georgilis A, Janich P, Morton JP, Athineos D, Kang TW, Lasitschka F, Andrulis M, Pascual G, Morris KJ, Khan S, Jin H, Dharmalingam G, Snijders AP, Carroll T, Capper D, Pritchard C, Inman GJ, Longerich T, Sansom OJ, Benitah SA, Zender L, Gil J. A complex secretory program orchestrated by the inflammasome controls paracrine senescence. Nat Cell Biol. 2013;15:978–990. doi: 10.1038/ncb2784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Farr JN, Xu M, Weivoda MM, Monroe DG, Fraser DG, Onken JL, Negley BA, Sfeir JG, Ogrodnik MB, Hachfeld CM, LeBrasseur NK, Drake MT, Pignolo RJ, Pirtskhalava T, Tchkonia T, Oursler MJ, Kirkland JL, Khosla S. Corrigendum: Targeting cellular senescence prevents age-related bone loss in mice. Nat Med. 2017;23:1384. doi: 10.1038/nm1117-1384c. [DOI] [PubMed] [Google Scholar]