

Abstract
Background
Acute liver failure is an inflammation-mediated hepatocyte injury. Mesenchymal stem cell (MSC) transplantation is currently considered to be an effective treatment strategy for acute liver failure. Exosomes are an important paracrine factor that can be used as a direct therapeutic agent. However, the use of bone marrow mesenchymal stem cell-derived exosomes (BMSC-Exos) in the treatment of acute liver failure has not been reported.
Purpose
Here, we established a model of hepatocyte injury and apoptosis induced by D-galactosamine and lipopolysaccharide (D-GalN/LPS) to study the protective effect of BMSC-Exos on hepatocyte apoptosis, and further explored its protective mechanism.
Methods
BMSC-Exos was identified by transmission electron microscopy (TEM), nanoparticle tracking analysis (NTA) and Western blot. Laser confocal microscopy was used to observe the uptake of Dil-Exos by hepatocytes. D-GalN/LPS-induced primary hepatocytes were pretreated with BMSC-Exos in vitro, and then the cells were harvested. The apoptosis of hepatocytes was observed by TUNEL staining, flow cytometry and Western blot. Electron microscopy and mRFP-GFP-LC3 and Western blot was used to observe autophagy.
Results
BMSC-Exos increased the expression of autophagy marker proteins LC3 and Beclin-1 and promoted the formation of autophagosomes. After BMSC-Exos treatment, the expression levels of the proapoptotic proteins Bax and cleaved caspase-3 were significantly decreased, while the expression level of the anti-apoptotic protein Bcl-2 was upregulated. However, when the autophagy inhibitor 3MA was present, the effect of BMSC-Exos on inhibiting apoptosis was significantly reversed.
Conclusions
Our results showed for the first time that BMSC-Exos had the potential to reduce hepatocyte apoptosis after acute liver failure. In particular, we found that BMSC-Exos attenuated hepatocyte apoptosis by promoting autophagy.
Keywords: bone marrow mesenchymal stem cells, exosomes, D-GalN/LPS, apoptosis, autophagy, acute liver failure
Acute liver failure (ALF) is a clinical syndrome caused by inflammation-mediated hepatocyte injury accompanied by hepatocyte apoptosis and necrosis.1 It is caused by the hepatitis virus, hepatotoxic drugs and hepatic ischemia–reperfusion injury.2 Clinically, it is characterized by critical illness, rapid development, and high mortality. So far, there are no specific and effective therapeutic drugs or treatments other than liver transplantation. Hepatocyte apoptosis is an important pathological manifestation of early ALF. If hepatocyte apoptosis can be inhibited, it is expected to delay or even block the progression of ALF.3
Cell transplantation may be a promising strategy to improve hepatocyte apoptosis and liver function recovery after ALF. In recent years, with the development of regenerative medicine and stem cell technology, stem cell-based cell transplantation has brought a new dawn for patients with ALF.4 Bone marrow mesenchymal stem cells (BMSCs) are widely involved in immune regulation due to their wide range of sources, and have fewer associated ethical problems.5 Under certain conditions, they can differentiate into adipocytes, chondrocytes, bone cells, cardiomyocytes and nerve cells.5 Hepatocytes and the like are widely used in tissue engineering and regenerative medicine. Studies have shown that MSC transplantation has good efficacy and safety in the treatment of ALF.6,7 The beneficial effects of bone marrow mesenchymal stem cell transplantation are mainly mediated by hepatocyte differentiation or by hepatocyte fusion, paracrine effects, and immunomodulatory effects.8,9 However, direct transplantation of stem cells into target tissues remains challenging. For example, transplanted stem cells have been shown to have a lower survival rate in ischemic tissue.10 Other risks, such as cell dedifferentiation, immune rejection, and tumor formation, further limit the clinical application of direct stem cell transplantation for ALF.11
Recent studies have shown that transplanted stem cells play a therapeutic role mainly through a paracrine mechanism, and exosomes play an important role in this process.12 Exosomes are the smallest endocyte-derived membrane-bound nanovesicles involved in complex intercellular communication systems.13 They are released from different types of cells under normal or pathological conditions and affect the activity of the recipient cells by carrying an activity signal. Exosomes contain not only cellular proteins and lipids, but also host cell mRNA and miRNA.14 Since stem cell secretions seem to have greater benefits in tissue regeneration and repair than stem cells themselves, extracellular components, such as stem cell-derived exosomes, play an important role in their therapeutic effects and have begun to attract attention.15 Bone marrow mesenchymal stem cell-derived exosomes have been proven to be applicable to animal models of many diseases, such as reducing myocardial ischemia, promoting skin healing, promoting repair of damaged kidneys and nerves, inhibiting liver differentiation, etc.16–19 Therefore, we hypothesized that exosomes derived from bone marrow mesenchymal stem cells will also reduce hepatocyte apoptosis after ALF.
Studies have shown that autophagy is critical for protection after ALF.20 Autophagy is a self-feeding phenomenon in cells that removes damaged organelles or long-lived proteins mainly through the lysosomal pathway. It is a major intracellular degradation system that can recover and reuse damaged macromolecules and organelles. This degradation contributes to the normal renewal of intracellular components and organelles. It acts as a survival mechanism for cells and plays an important role in cell growth, survival, differentiation and homeostasis.21 Studies have shown that activation of autophagy plays an important role in reducing tissue damage, and induction of autophagy can effectively alleviate hepatocyte injury after ALF.22 However, it is unclear whether BMSC-Exos can promote autophagy activation.
In this study, we established a model of hepatocyte injury and apoptosis induced by D-galactosamine and lipopolysaccharide (D-GalN/LPS) to study the protective effect of BMSC-Exos on hepatocyte apoptosis, and further explored its protective mechanism against hepatocyte apoptosis. Here, we showed that BMSC-Exos had the potential to reduce hepatocyte apoptosis after ALF. In particular, we found that BMSC-Exos attenuated hepatocyte apoptosis by promoting autophagy.
Materials and methods
Isolation and purification of bone mesenchymal stem cells/exosomes
Exosomes were isolated from BMSC supernatant as previously described.23 The growth medium was collected and centrifuged at 300× g for 10 mins, followed by centrifugation at 2,000× g for 10 mins at 4 °C. After centrifugation, the cell supernatant was sterilized by filtration through a 0.22 µm filter to remove cell debris. After that, the upper layer of the supernatant was transferred to an Amicon Ultra-15 centrifugal filter (Millipore, Burlington, MA, USA) and centrifuge at 4,000× g at 4 °C until the volume of the upper chamber was reduced to nearly 200 µL. The ultrafiltrate was washed twice with phosphate-buffered saline (PBS) and ultrafiltered again to 200 µL. For exosome purification, the liquid was loaded onto a 30% sucrose/D2O pad in a sterile Ultra-Clear™ tube (Beckman Coulter, Brea, CA, USA) and centrifuged at 4 °C, 100,000× g for 60 min in a Sorvall Avanti J-26XP fixed angle rotor (Beckman Coulter). The supernatant partially containing BMSC-Exos was recovered using an 18g needle, diluted in PBS, and centrifuged at 4 °C, 4,000× g to centrifuge the filter unit until the final volume reached 200 µL. The solution was stored at −80 °C or used in another series of experiments. The BMSC-Exos protein content was determined by the double octanoic acid method (BCA; Thermo Fisher Scientific, Waltham, MA, USA). A microplate reader (ELx800; Bio-Tek Instruments, Inc., Winooski, VT, USA) was used to read the absorbance at 562 nm.
Identification of exosomes derived from bone mesenchymal stem cells
To analyze the characteristics of exosomes, a three-dimensional map of the particle size, solid shape and relative intensity of BMSC-Exos was tested using the NTA system. The morphology of the obtained exosomes was observed by transmission electron microscopy (TEM). Western blotting was used to detect specific exosomal surface markers, which were encapsulated in exosomes including CD9, CD63 and CD81.
BMSC-Exos uptake
For fluorescent labeling of BMSC-Exos, 4 mg/mL Dil solution (molecular probe) was added to PBS (1:200) and incubated according to the manufacturer’s instructions. Excess dye from labeled exosomes was removed by ultracentrifugation at 100,000× g for 1 hr at 4 °C and washed three times by resuspending the pellet in PBS. The final pellet was resuspended in PBS. Dil-Exos were co-cultured with hepatic cells for 12h or 24 h, and then the cells were washed with PBS and fixed in 4% paraformaldehyde. The uptake was then observed by laser confocal microscopy.
Isolation and culture of rat hepatocytes
The liver of the experimental rats was removed under aseptic conditions and transferred to a plate. After gently tearing off the liver capsule, the liver was rinsed with normal saline and cut into thin tissue blocks with small scissors. The liver tissue blocks were then transferred to 0.25% trypsin (in PBS) and cold digested at 4 °C for 10–12 h, then homogenized with a 200-mesh sieve, and a single cell suspension was generated with a certain amount of PBS. The cell suspension was then placed in a 25 mL centrifuge tube, centrifuged at 1000 rpm for 5 min, then the supernatant was discarded, and the centrifugation was repeated another three times. Liver parenchymal cells were collected and cultured in DMEM containing 10% fetal bovine serum. The density of isolated hepatocytes was adjusted to 1×105 mL, and the liver cell suspension was plated into 6-well (5 mL per well) and 24-well (1 mL per well) culture plates. After culture in a CO2 incubator at 37 °C for 12 h, the hepatocytes were all adherently grown for experimental use. The study was approved by the Ethics Committee of Medical college of Qingdao university. All procedures were conducted in accordance with the guidelines of the National Institutes of Health Laboratory Animal Care and Use Guidelines.
Apoptosis assays by TUNEL/DAPI staining and flow cytometry: annexin V/PI double staining
After D-GaIN/LPS (44 μg/mL D-GalN and 100 ng/mL LPS) treatment, cultured hepatocytes with or without BMSC-Exos (100 ug/mL) for 24 h. TUNEL (Roche, Basel, Switzerland) was added according to the manufacturer’s instructions and incubated at 37 °C for 30 min in the dark. The cells were then incubated for 5 min with DAPI (Beyotime Institute of Biotechnology, Jiangsu China) and observed under an inverted fluorescence microscope. Randomly-selected fields of view and representative images were selected for apoptotic cell counts and total cell counts. The rate of TUNEL-positive cells in each zone was calculated.
Further verification of apoptosis was performed by flow cytometry. After hepatic cell treatment, cells were harvested by centrifugation at 1500 rpm for 5 min. The hepatic cells were washed twice with PBS, then resuspended in FITC-labeled Annexin V (5 µL; BD Biosciences, Franklin Lakes, NJ, USA) and PI (5 µL; BD Biosciences) and reacted in the dark for 5 mins. Apoptosis was detected by flow cytometry after washing three times with PBS.
Western blot analysis
Total protein was extracted from cells, and the protein concentration was measured using a BCA assay. The samples were subjected to sodium dodecyl sulfate gel electrophoresis, and the separated proteins were transferred onto a polyvinylidene difluoride membrane and blocked with 5% bovine serum albumin for 1 h at room temperature. The main antibodies used were cleaved caspase-3 (1:1,000, rabbit IgG; Cell Signal Technology, Danvers, MA, USA), Bcl-2 (1:1,000, rabbit IgG; Abcam, Cambridge, MA, USA), Bax (1:1,000, rabbit IgG; Abcam, USA), beclin-1 (1:1,000, rabbit IgG; Abcam), LC3B (1:1,000, mouse IgG1; Abcam), GAPDH (as a gel-loading control, 1:1,000 and rabbit IgG; Abcam). Membranes were then incubated with horseradish peroxidase-conjugated anti-rabbit IgG and anti-mouse IgG antibodies (1:2,000, Thermo Fisher Scientific) for 120 min, followed by visualization of the bands using an enhanced chemiluminescence reagent (Thermo Fisher Scientific). Protein expression levels were determined by densitometry using Image J software.
Double-labeled adenovirus mRFP-GFP-LC3 transfection and autophagy detection
The extracted hepatic cells were seeded and cultured in confocal dishes for 4 days, and then mRFP-GFP-LC3 lentivirus (Han Heng Biology, China) was transfected according to the manufacturer’s protocol. Then, the cells were divided into three groups: control group, D-GaIN/LPS group; pretreated BMSC-Exos; treated PBS-washed cells, then fixed with 4% paraformaldehyde, and fluorescence was observed using a laser scanning confocal microscope (LSM 510; Zeiss, Oberkochen, Germany). The number of yellow spots represents autophagic bodies and red spots represent autophagic lysosomes. The number of autophagosomes and autolysosomes were quantified using IN Cell Investigator software.
TEM assessment of autophagy
After the cells were treated, the adherent hepatocytes were digested with trypsin and then centrifuged. The supernatant was discarded and the cell pellet was fixed with a precooled 2% glutaraldehyde solution at 4 °C for 2 h. The cells were stained with 2% uranyl acetate solution for 2 h and then dehydrated in 50%, 70%, 90% and 100% acetone. The cells were embedded and ultrathin sections were observed under an electron microscope (FEI Tecnai, Hillsboro, OR, USA).
Statistical analysis
Data and images were processed and analyzed using IBM SPSS Statistics v17.0. Data are expressed as mean ± standard deviation of at least three independent experiments. Data were analyzed using Student’s t-tests. All tests were bilateral, with an asterisk (*) and a pound sign (#) indicating a p-value <0.05.
Results
Identification of BMSC-Exos
Exosomes were isolated from the BMSC culture supernatant by a combination of ultrafiltration centrifugation and ultracentrifugation. The purified BMSC nanoparticles were identified by TEM, nanoparticle tracking analysis (NTA) analysis and Western blotting. Typical exosomal structures were observed by TEM (Figure 1A). NTA analysis showed that these nanoparticles have a particle size distribution between 20 and 150 nm, similar to previous reports (Figure 1B). In addition, Western blotting results showed that the specific exosomal surface markers CD9, CD63, and CD81 in BMSC-Exos were positive, which further confirmed the existence of exosomes (Figure 1C). All of the above analyses indicated that exosomes extracted from BMSCs were successfully isolated and identified.
Figure 1.

Characterization of BMSC-derived exosomes (BMSC-Exos). (A) Exosome morphology revealed by transmission electron microscopy (TEM). (B) Particle size distribution measured by nanoparticle tracking analysis (NTA). (C) Western blot analysis of specific exosome surface markers.
BMSC-Exos localization in cells
To further investigate whether BMSC-Exos can be taken up by hepatocytes, Dil-Exos were incubated with hepatocytes in vitro and the uptake was observed by fluorescence microscopy. After 12h or 24 h of incubation, it was observed that Dil-Exos had been taken up by the liver cells and transferred to the cytoplasm (Figure 2A, B). Taken together, these data indicated that BMSC-Exos can be taken up by hepatocytes.
Figure 2.

DiI-labeled BMSC-Exos uptake. (A) Uptake of DiI-labeled BMSC-Exos by hepatic cells after 12 h. (B) Uptake of DiI-labeled BMSC-Exos by hepatic cells after 24 h. BMSC-Exos, Bone marrow mesenchymal stem cell-derived exosomes.
BMSC-Exos inhibit D-GaIN/LPS-induced hepatocyte apoptosis
Studies have shown that D-GaIN/LPS can cause large-scale death of liver cells after intraperitoneal injection of mice. Therefore, we used D-GaIN/LPS to induce hepatocyte apoptosis to mimic hepatocyte injury after ALF in vivo. We used TUNEL to study hepatocyte apoptosis. As shown, after administration of D-GaIN/LPS, hepatocytes exhibited a relative apoptotic ratio. We then evaluated the protective effect of BMSC-Exos in the D-GaIN/LPS-induced hepatocyte apoptosis model. The results showed that after application of BMSC-Exos (100 µg/mL), the number of TUNEL-positive neurons decreased significantly, indicating that BMSC-Exos had a protective effect on D-GaIN/LPS-induced hepatocyte apoptosis (Figure 3A, B).
Figure 3.

BMSC-Exos pretreatment attenuates D-GaIN/LPS-induced hepatocyte apoptosis. (A) TUNEL-staining (red) for detection of apoptosis in hepatocytes. Cell nuclei were counterstained with DAPI (blue). (B) Quantitative estimation of the proportion of apoptotic cells in each experimental group: control, D-GaIN/LPS, and BMSC-Exos pretreatment followed by D-GaIN/LPS as indicated (BMSC-Exos+D-GaIN/LPS). Pretreatment with BMSC-Exos substantially reduced D-GaIN/LPS-induced apoptosis. (C) Annexin V/FITC/PI double staining flow cytometry was also used to detect hepatocyte apoptosis induced by D-GaIN/LPS with or without BMSC-Exos pretreatment. (D) Quantitative results of flow cytometry confirming that BMSC-Exos pretreatment reduced D-GaIN/LPS-induced apoptosis of hepatic cells. (E) Western blot analysis of hepatocyte apoptosis-related proteins. (F) Relative expression levels of apoptosis-related proteins normalized to GAPDH. Pretreatment with BMSC-Exos upregulated anti-apoptotic Bcl-2 and downregulated pro-apoptotic Bax and cleaved caspase-3. *p<0.05 compared to the Control group, #p<0.05 compared to the D-GaIN/LPS group.
Abbreviations: BMSC-Exos, Bone marrow mesenchymal stem cell-derived exosomes; D-GaIN/LPS, D-galactosamine and lipopolysaccharide; Bax, Bcl-2-associated X protein; Bcl-2, B cell lymphoma 2; FITC, fluorescein isothiocyanate; PI, propidium iodide; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; TUNEL, terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling assay.
Next, we further evaluated apoptosis using Annexin V-FITC/PI double staining flow cytometry to determine the early and late apoptotic rate of cells. As shown in Figure 3C, exposure to D-GaIN/LPS significantly increased apoptosis compared with the control group, while administration of BMSC-Exos significantly attenuated the apoptosis of D-GaIN/LPS in hepatocytes (Figure 3C, D).
Western blotting results showed that after exposure to D-GaIN/LPS for 24 h, the proapoptotic proteins Bax and cleaved caspase-3 were decreased after co-treatment with BMSC-Exos, while when Bcl-2 and other anti-apoptotic proteins were co-incubated with BMSC-Exos, the level of apoptotic proteins was elevated (Figure 3E, F).
These data indicated that BMSC-Exos protected hepatocytes from D-GaIN/LPS-induced apoptosis.
BMSC-Exos induce autophagy activation
Because autophagy plays an important role in protection from liver injury, we examined the effect of BMSC-Exos on autophagy activation. D-GaIN/LPS induces autophagy in hepatic cells.24 As in the pretreatment method, hepatocytes were treated with D-GaIN/LPS and continued to be cultured with or without BMSC-Exos, and autophagy-related indicators were detected. TEM showed that a certain number of autophagosomes appeared in hepatocytes after D-GaIN/LPS treatment. Compared with the D-GaIN/LPS group and the control group, the number of autophagosomes was higher in BMSC-Exo-treated cells (Figure 4A, B). To better illustrate autophagy, we transfected hepatocytes with the mRFP-GFP-LC3 virus and observed autophagy flux using a laser confocal microscope. In this assay, autophagosomes were labeled with red and green fluorescence (yellow spots); autophagic lysosomes were labeled with red fluorescence (red spots). The results showed that the BMSC-Exos group had more yellow and red spots than the D-GaIN/LPS group (Figure 4C, D). Subsequently, we used Western blotting to further detect autophagy-related proteins. The results showed that treatment with BMSC-Exos increased the expression of the autophagy-related proteins LC3II and Beclin-1 (Figure 4E, F). These data indicated that BMSC-Exos activated hepatocyte autophagy.
Figure 4.

BMSC-Exos pretreatment induces autophagy in hepatocytes. (A, B) Transmission electron micrographs of autophagosomes in BMSC-Exos-pretreated hepatocytes. (C, D) Autophagic flux of mRFP-GFP-LC3 transfected hepatocytes revealed by laser confocal microscopy. Autophagosomes are labeled by red and green fluorescence (yellow spots), while autophagic lysosomes are labeled by red fluorescence (red spots). The BMSC-Exos+D-GaIN/LPS group demonstrated a large number of yellow and red spots than the D-GaIN/LPS-only group. (E) Western blot detection of hepatocyte autophagy markers LC3B and Beclin-1. (F) Semi-quantitative analysis showing enhanced expression of autophagy-related proteins in BMSC-Exos-pretreated hepatocytes compared to controls. *p<0.05 compared to the D-GaIN/LPS group.
Abbreviations: BMSC-Exos, Bone marrow mesenchymal stem cell-derived exosomes; D-GaIN/LPS, D-galactosamine and lipopolysaccharide.
BMSC-Exos reduce hepatocyte apoptosis by activating autophagy
Considering the key role of autophagy in hepatocyte apoptosis, we wondered whether BMSC-Exos rely on autophagy to reduce apoptosis after liver injury. Hepatocytes were treated with BMSC-Exos in the presence or absence of the autophagy inhibitor 3MA by exposure of hepatocytes to D-GaIN/LPS-established injury and apoptosis models. The results were consistent with the above; after the application of BMSC-Exos, the number of TUNEL-positive hepatocytes was significantly reduced (Figure 5A, B). However, when the autophagy inhibitor 3MA was present, the inhibition of apoptosis by BMSC-Exos was significantly reversed (Figure 5A, B). Annexin V-FITC/PI double staining flow cytometry was consistent with TUNEL results (Figure 5C, D). Next, we went on to use Western blotting to further verify that 3MA inhibited the increase of BMSC-Exo-mediated the autophagy marker proteins LC3II and the anti-apoptotic protein BCL-2, and reversed the decline of pro-apoptotic protein cleaved caspase-3 (Figure 5E-H). Taken together, these results indicated that activation of autophagy by BMSC-Exos helped to reduce hepatocyte apoptosis.
Figure 5.
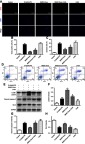
BMSC-Exos reduces hepatocyte apoptosis by activating autophagy. (A) TUNEL staining was used to detect hepatocyte apoptosis induced by D-GaIN/LPS with or without BMSC-Exos and 3MA pretreatment. (B) TUNEL-positive hepatocyte numbers. The reduction in TUNEL-positive hepatocytes observed in the D-GaIN/LPS+BMSC-Exos group compared to the D-GaIN/LPS group was reversed by co-treatment with 3MA. (C, D) Hepatocytes apoptosis was detected by Annexin V/FITC/PI double staining and flow cytometry. Results were consistent with TUNEL staining. (E) Western blot was used to detect the expression of autophagy- and apoptosis-related proteins. 3MA pretreatment inhibited BMSC-Exos-induced elevations in autophagy marker proteins LC3B and the anti-apoptotic protein Bcl-2, and reversed the BMSC-Exos-induced decrease in proapoptotic cleaved caspase-3 expression. (F-H) Semi-quantification of autophagy and expression levels of apoptosis-related proteins. *p<0.05 compared to the D-GaIN/LPS group, #p<0.05 compared to the BMSC-Exos group.
Abbreviations: BMSC-Exos, Bone marrow mesenchymal stem cell-derived exosomes; D-GaIN/LPS, D-galactosamine and lipopolysaccharide; TUNEL, terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling assay; 3MA, 3-Methyladenine; FITC, fluorescein isothiocyanate; PI, propidium iodide; Bax, Bcl-2-associated X protein; Bcl-2, B cell lymphoma 2.
Discussion
The pathological manifestation of ALF is that a large number of hepatocytes are severely damaged and undergo apoptosis or necrosis, hepatocyte proliferation is inhibited, liver function is severely impaired, and multiple organ failure can result.25 D-GalN/LPS is widely used to establish a mouse ALF model and to evaluate the protective effect of drugs on ALF.26 This study demonstrated for the first time that BMSC-Exos could attenuate D-GalN/LPS-induced hepatocyte injury and apoptosis by inducing autophagy activation.
With the rapid development of cell transplantation therapy, bone marrow mesenchymal stem cells have the advantages of strong self-renewal ability, strong differentiation ability and low immunogenicity, and have therefore attracted wide attention.27 Studies have shown that bone marrow mesenchymal stem cells have unique liver function protection activity, which can promote liver function recovery after ALF.28,29 However, it has also been suggested that only 1% of transplanted bone marrow mesenchymal stem cells can be successfully transferred to the target tissue where they are needed. A large number of transplanted bone marrow mesenchymal stem cells are trapped in the lungs after intravenous injection.30 Therefore, although bone marrow mesenchymal stem cell therapy has achieved some success in various animal disease models, due to the huge physiological differences between humans and animals, many problems remain to be solved before being approved for clinical applications.
Exosomes are small vesicles released by cells whose surface antigens are characterized by cell origin. These vesicles may play a key role in cellular communication by transmitting RNA, proteins and bioactive lipids.31 A large number of studies have confirmed that different types of MSC-Exos have functions of angiogenesis, cell protection, inflammation regulation and anti-apoptosis.32,33 MSC-Exos have been used in many fields such as cardiovascular disease, kidney damage, immune diseases, tumors, and nervous system diseases.17–19 However, there are few research reports concerning the treatment of liver disease. In regenerative medicine, exosomes may be more advantageous than stem cells because they avoid all the limitations of direct stem cell transplantation.34 Stem cell transplantation therapy may work through a paracrine mechanism.35 Therefore, we hypothesized that direct administration of bone marrow mesenchymal stem cell exosomes could overcome the limitations and challenges of direct administration of stem cells. We conducted a series of experiments in vitro to prove our hypothesis. First, we successfully extracted bone marrow mesenchymal stem cells and isolated high concentrations of exosomes from their supernatant. Exosomes with a diameter of 20–150 nm were then identified using NTA and further verified by TEM and by analysis of the specific exosomal surface markers CD9, CD63, and CD81.
The pathophysiological mechanism of ALF is complex, and hepatocyte apoptosis is an important pathological manifestation of early ALF.36 After hepatocyte injury, the expression of the proapoptotic proteins Bax and cleaved caspase 3 was upregulated, while the expression of the anti-apoptotic protein Bcl-2 was downregulated. The study revealed that D-GaIN/LPS can cause large-scale death of liver cells after intraperitoneal injection of mice.37 We extracted rat liver cells as research materials and further tested BMSC-Exos on D-GaIN/LPS-induced hepatocyte apoptosis. In vitro TUNEL and flow cytometry results showed that co-incubation with BMSC-Exos reduced D-GaIN/LPS-induced hepatocyte apoptosis, as evidenced by our Western blot results, which showed down-regulation of proapoptotic-related markers in injured cells and up-regulation of anti-apoptotic markers. These results showed that BMSC-Exos inhibited D-GaIN/LPS-induced hepatocyte apoptosis.
The role of autophagy as a degradation pathway is critical in regenerative medicine. Many reports indicate that basal or physiological autophagy contributes to the maintenance of cell homeostasis and the quality control of proteins and subcellular organelles.38 Pathological conditions or cellular stress can induce autophagy as a cell’s adaptive and protective mechanism to promote survival.39 After autophagy, the expression of the autophagy-related proteins LC3II and beclin-1 was up-regulated.40,41 Studies have shown that in a rat model of ALF, an increase in autophagy can reduce damage to cells by treating the components of the lesion.22 It is worth noting that autophagy induction as a self-protection mechanism has been confirmed in an experimental model of cell damage induced by ALF.22,42 Baixauli et al noted that exosomes have a novel function that synergizes with autophagy-lysosomal pathways to alleviate intracellular stress conditions and are critical for maintaining intracellular protein and RNA homeostasis.43 However, whether BMSC-Exos can activate autophagy of target cells to prevent tissue damage has not been reported. Here, our in vitro electron microscopy and mRFP-GFP-LC3 lentiviral transfection results showed that co-incubation with BMSC-Exos increased D-GaIN/LPS-induced autophagy, as evidenced by our Western blot results. Results showed that BMSC-Exos can induce autophagy in hepatocytes. To demonstrate whether BMSC-Exo-mediated autophagy is critical for inhibition of apoptosis, we pretreated hepatic cells with the autophagy-specific inhibitor 3MA. The results indicated that autophagy inhibition reduced the anti-apoptotic effect of BMSC-Exos. In general, these data indicated that BMSC-Exos inhibited apoptosis by inducing autophagy after hepatocyte injury.
In summary, this study demonstrated for the first time that BMSC-Exos could effectively reduce D-GaIN/LPS-induced hepatocyte apoptosis. In particular, we found that BMSC-Exos could induce autophagy and protect hepatic cells from damage caused by various stresses by mediating autophagy. Therefore, we believe that BMSC-Exos may inhibit the development of ALF by preventing the apoptosis of hepatic cells by autophagy, and our findings lay a solid theoretical foundation and realistic basis for the future clinical research of BMSC-Exos as a new biological treatment for ALF.
Disclosure
The authors report no conflicts of interest in this work.
References
- 1.Jedicke N, Struever N, Aggrawal N, et al. alpha-1-antitrypsin inhibits acute liver failure in mice. Hepatology. 2014;59:2299–2308. [DOI] [PubMed] [Google Scholar]
- 2.Lee WM, Squires RH Jr., Nyberg SL, Doo E, Hoofnagle JH. Acute liver failure: summary of a workshop. Hepatology. 2008;47:1401–1415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Josephs MD, Bahjat FR, Fukuzuka K, et al. Lipopolysaccharide and D-galactosamine-induced hepatic injury is mediated by TNF-alpha and not by Fas ligand. Am J Physiol Regul Integr Comp Physiol. 2000;278:R1196–1201. [DOI] [PubMed] [Google Scholar]
- 4.Wang Y, Yu X, Chen E, Li L. Liver-derived human mesenchymal stem cells: a novel therapeutic source for liver diseases. Stem Cell Res Ther. 2016;7:71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Esrefoglu M. Role of stem cells in repair of liver injury: experimental and clinical benefit of transferred stem cells on liver failure. World J Gastroenterol. 2013;19:6757–6773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Tan CY, Lai RC, Wong W, Dan YY, Lim SK, Ho HK. Mesenchymal stem cell-derived exosomes promote hepatic regeneration in drug-induced liver injury models. Stem Cell Res Ther. 2014;5:76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Volarevic V, Nurkovic J, Arsenijevic N, Stojkovic M. Concise review: therapeutic potential of mesenchymal stem cells for the treatment of acute liver failure and cirrhosis. Stem Cells. 2014;32:2818–2823. [DOI] [PubMed] [Google Scholar]
- 8.Manzini BM, da Silva Santos Duarte A, Sankaramanivel S, et al. Useful properties of undifferentiated mesenchymal stromal cells and adipose tissue as the source in liver-regenerative therapy studied in an animal model of severe acute fulminant hepatitis. Cytotherapy. 2015;17:1052–1065. [DOI] [PubMed] [Google Scholar]
- 9.Lotfinia M, Kadivar M, Piryaei A, et al. Effect of secreted molecules of human embryonic stem cell-derived mesenchymal stem cells on acute hepatic failure model. Stem Cells Dev. 2016;25:1898–1908. doi: 10.1089/scd.2016.0177 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Balsam LB, Wagers AJ, Christensen JL, Kofidis T, Weissman IL, Robbins RC. Haematopoietic stem cells adopt mature haematopoietic fates in ischaemic myocardium. Nature. 2004;428:668–673. [DOI] [PubMed] [Google Scholar]
- 11.Jeong JO, Han JW, Kim JM, et al. Malignant tumor formation after transplantation of short-term cultured bone marrow mesenchymal stem cells in experimental myocardial infarction and diabetic neuropathy. Circ Res. 2011;108:1340–1347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ratajczak MZ, Jadczyk T, Pedziwiatr D, Wojakowski W. New advances in stem cell research: practical implications for regenerative medicine. Pol Arch Med Wewn. 2014;124:417–426. [DOI] [PubMed] [Google Scholar]
- 13.Thery C, Witwer KW. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the international society for extracellular vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018;7:1535750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Barteneva NS, Maltsev N, Vorobjev IA. Microvesicles and intercellular communication in the context of parasitism. Front Cell Infect Microbiol. 2013;3:49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Teixeira FG, Carvalho MM, Sousa N, Salgado AJ. Mesenchymal stem cells secretome: a new paradigm for central nervous system regeneration? Cell Mol Life Sci. 2013;70:3871–3882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wang B, Jia H, Zhang B, et al. Pre-incubation with hucMSC-exosomes prevents cisplatin-induced nephrotoxicity by activating autophagy. Stem Cell Res Ther. 2017;8:75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zhao J, Li X, Hu J et al. Mesenchymal stromal cell-derived exosomes attenuate myocardial ischaemia-reperfusion injury through miR-182-regulated macrophage polarization. Cardiovascular research. 2019; 115:1205-1216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Rong Y, Liu W, Wang J, et al. Neural stem cell-derived small extracellular vesicles attenuate apoptosis and neuroinflammation after traumatic spinal cord injury by activating autophagy. Cell Death Dis. 2019;10:340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wang C, Wang M, Xu T, et al. Engineering bioactive self-healing antibacterial exosomes hydrogel for promoting chronic diabetic wound healing and complete skin regeneration. Theranostics. 2019;9:65–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Allaire M, Rautou PE, Codogno P, Lotersztajn S. Autophagy in liver diseases: time for translation? J Hepatol. 2019;70:985–998. [DOI] [PubMed] [Google Scholar]
- 21.Mizushima N, Komatsu M. Autophagy: renovation of cells and tissues. Cell. 2011;147:728–741. [DOI] [PubMed] [Google Scholar]
- 22.Ni HM, Bockus A, Boggess N, Jaeschke H, Ding WX. Activation of autophagy protects against acetaminophen-induced hepatotoxicity. Hepatology. 2012;55:222–232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Montecalvo A, Larregina AT, Morelli AE. Methods of analysis of dendritic cell-derived exosome-shuttle microRNA and its horizontal propagation between dendritic cells. Methods Mol Biol. 2013;1024:19–40. [DOI] [PubMed] [Google Scholar]
- 24.Wang K, Damjanov I, Wan YJ. The protective role of pregnane X receptor in lipopolysaccharide/D-galactosamine-induced acute liver injury. Lab Invest. 2010;90:257–265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Tunon MJ, Alvarez M, Culebras JM, Gonzalez-Gallego J. An overview of animal models for investigating the pathogenesis and therapeutic strategies in acute hepatic failure. World J Gastroenterol. 2009;15:3086–3098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Liu YM, Lv J, Zeng QL, et al. AMPK activation ameliorates D-GalN/LPS-induced acute liver failure by upregulating Foxo3A to induce autophagy. Exp Cell Res. 2017;358:335–342. [DOI] [PubMed] [Google Scholar]
- 27.Abbasi-Malati Z, Roushandeh AM, Kuwahara Y, Roudkenar MH. Mesenchymal stem cells on horizon: a new arsenal of therapeutic agents. Stem Cell Rev. 2018;14:484–499. [DOI] [PubMed] [Google Scholar]
- 28.Wang K, Li Y, Zhu T, et al. Overexpression of c-Met in bone marrow mesenchymal stem cells improves their effectiveness in homing and repair of acute liver failure. Stem Cell Res Ther. 2017;8:162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zare H, Jamshidi S, Dehghan MM, Saheli M, Piryaei A. Bone marrow or adipose tissue mesenchymal stem cells: comparison of the therapeutic potentials in mice model of acute liver failure. J Cell Biochem. 2018;119:5834–5842. [DOI] [PubMed] [Google Scholar]
- 30.Phinney DG, Prockop DJ. Concise review: mesenchymal stem/multipotent stromal cells: the state of transdifferentiation and modes of tissue repair–current views. Stem Cells. 2007;25:2896–2902. [DOI] [PubMed] [Google Scholar]
- 31.Morel O, Toti F, Hugel B, Freyssinet JM. Cellular microparticles: a disseminated storage pool of bioactive vascular effectors. Curr Opin Hematol. 2004;11:156–164. [DOI] [PubMed] [Google Scholar]
- 32.Katsuda T, Tsuchiya R, Kosaka N, et al. Human adipose tissue-derived mesenchymal stem cells secrete functional neprilysin-bound exosomes. Sci Rep. 2013;3:1197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Xin H, Li Y, Cui Y, Yang JJ, Zhang ZG, Chopp M. Systemic administration of exosomes released from mesenchymal stromal cells promote functional recovery and neurovascular plasticity after stroke in rats. J Cereb Blood Flow Metab. 2013;33:1711–1715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Baglio SR, Pegtel DM, Baldini N. Mesenchymal stem cell secreted vesicles provide novel opportunities in (stem) cell-free therapy. Front Physiol. 2012;3:359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Katsuda T, Kosaka N, Takeshita F, Ochiya T. The therapeutic potential of mesenchymal stem cell-derived extracellular vesicles. Proteomics. 2013;13:1637–1653. [DOI] [PubMed] [Google Scholar]
- 36.Wang Y, Yang F, Jiao FZ. Modulations of histone deacetylase 2 offer a protective effect through the mitochondrial apoptosis pathway in acute liver failure. Oxid Med Cell Longev. 2019;2019:8173016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Ahmad A, Raish M, Ganaie MA, et al. Hepatoprotective effect of commiphora myrrha against d-GalN/LPS-induced hepatic injury in a rat model through attenuation of pro inflammatory cytokines and related genes. Pharm Biol. 2015;53:1759–1767. [DOI] [PubMed] [Google Scholar]
- 38.Singh R, Cuervo AM. Autophagy in the cellular energetic balance. Cell Metab. 2011;13:495–504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Levine B, Kroemer G. Autophagy in the pathogenesis of disease. Cell. 2008;132:27–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kang R, Zeh HJ, Lotze MT, Tang D. The Beclin 1 network regulates autophagy and apoptosis. Cell Death Differ. 2011;18:571–580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Huang R, Liu W. Identifying an essential role of nuclear LC3 for autophagy. Autophagy. 2015;11:852–853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Ni HM, Williams JA, Jaeschke H, Ding WX. Zonated induction of autophagy and mitochondrial spheroids limits acetaminophen-induced necrosis in the liver. Redox Biol. 2013;1:427–432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Baixauli F, Lopez-Otin C, Mittelbrunn M. Exosomes and autophagy: coordinated mechanisms for the maintenance of cellular fitness. Front Immunol. 2014;5:403. [DOI] [PMC free article] [PubMed] [Google Scholar]