

Abstract
Recalling happy memories elicits positive feelings and enhances one’s wellbeing, suggesting a potential adaptive function in using this strategy for coping with stress. In two studies, we explored whether recalling autobiographical memories that have a positive content – i.e., remembering the good times – can dampen the hypothalamic-pituitary-adrenal (HPA) axis stress response. Participants underwent an acute stressor or control task followed by autobiographical memory recollection (of only positive or neutral valence). Across both studies, recalling positive, but not neutral, memories resulted in a dampened cortisol rise and reduced negative affect. Further, individuals with greater self-reported resiliency showed enhanced mood, despite stress exposure. During positive reminiscence, we observed engagement of corticostriatal circuits previously implicated in reward-processing and emotion regulation, and stronger connectivity between ventrolateral and dorsolateral prefrontal cortex as a function of positivity. These findings highlight the restorative and protective function of self-generated positive emotions via memory recall in the face of stress.
Keywords: Positive Emotion, Autobiographical Memory, Stress, Cortisol, Wellbeing, Prefrontal Cortex, Striatum
Acute stress can leave us feeling anxious and distressed, with detrimental consequences to our physical and mental health1. We often use cognitive regulation strategies to suppress these unpleasant feelings altogether (e.g., suppression) or to reinterpret the negative situation into something less negative or neutral (e.g., cognitive reappraisal)2. Despite our best efforts, however, we are not always successful in diminishing unpleasant feelings when using cognitive strategies under stress3. This may not be surprising considering that stress is thought to compromise the exact neural circuitry that emotion regulation relies on4. Thus, a promising alternative may be to focus on increasing or sustaining positive feelings – a strategy that broadens one’s cognitive perspective5 – and may foster better coping with the stressor.
One way of bolstering positive emotions is to reminisce about past positive events. Indeed, autobiographical memories can bring back emotions tied to the original experience6. Retrieving positive memories in particular may be intrinsically valuable, by re-kindling pleasant feelings and engaging neural circuitry involved in reward-processing (e.g., striatum)7. Notably, such striatal activity correlates with self-reported resiliency and enhanced mood for some individuals, consistent with a role for corticostriatal circuits in sustaining positive mood8,9. Thus, savoring happy memories might be significant for one’s ability to cope with stress, potentially promoting better decision-making and wellbeing.
Yet, a critical question remains whether recalling the positive past can actually facilitate successful adaptation to stress (i.e., resiliency). Experiencing stress activates the hypothalamic-pituitary-adrenal (HPA) axis, which releases a cascade of hormones including the primary stress hormone cortisol10. Heightened cortisol response after stress has deleterious effects on affective and cognitive states, disrupting processes supported by the prefrontal cortex such as working memory11 and decision-making12,13. Acute stress is also thought to be a precursor to depressive episodes14 and may influence reward systems15. Individuals widely vary with respect to the psychological resources they have and the strategies they implement for coping with acute stressors16. Cognitive regulation strategies (e.g., reappraisal) that are typically effective for diminishing negative affect elicited by images17 or conditioned stimuli18 are often rendered ineffective after stress exposure3, highlighting a need for alternative ways to combat stress. The present study investigates one potential mechanism: remembering the good times. That is, can simply reminiscing about our own positive memories help diminish the physiological (e.g., reduce cortisol) and emotional consequences of stress exposure? Further, what are the neural mechanisms underlying the ability to buffer the effects of stress by recalling the positive past?
We explored this idea first behaviorally (N=134) and then using functional Magnetic Resonance Imaging (fMRI; N=43). In the behavioral study, we exposed participants to an acute stressor or control task prior to autobiographical memory recollection. Importantly, half of the sample reminisced about positive memories, whereas the other half reminisced about neutral memories, creating four experimental groups: Stress-Positive (N=33), Stress-Neutral (N=34), Control-Positive (N=33), and Control-Neutral (N=34; see Fig 1 for timeline). Stress participants underwent the Socially Evaluative Cold Pressor task (SECPT; immersed hand in ice cold water under social threat)19, which reliably activates the HPA axis, producing elevated cortisol levels about 15 min after the stressor10. To assess physiological changes to stress over time, salivary cortisol was collected at baseline (s1), after memory recollection when cortisol was expected to peak (s2, +20 min), and at the conclusion of the experiment when cortisol was expected to recover (s3, +50 min). We hypothesized that recalling positive memories, relative to neutral memories, would buffer the negative effects of stress by a) decreasing the cortisol response and b) having a positive effect on mood.
Figure 1. Schematic of Experimental Procedure and Timeline of Neuroendocrine Assessments for Day 2 (Stress Manipulation or Control Task).
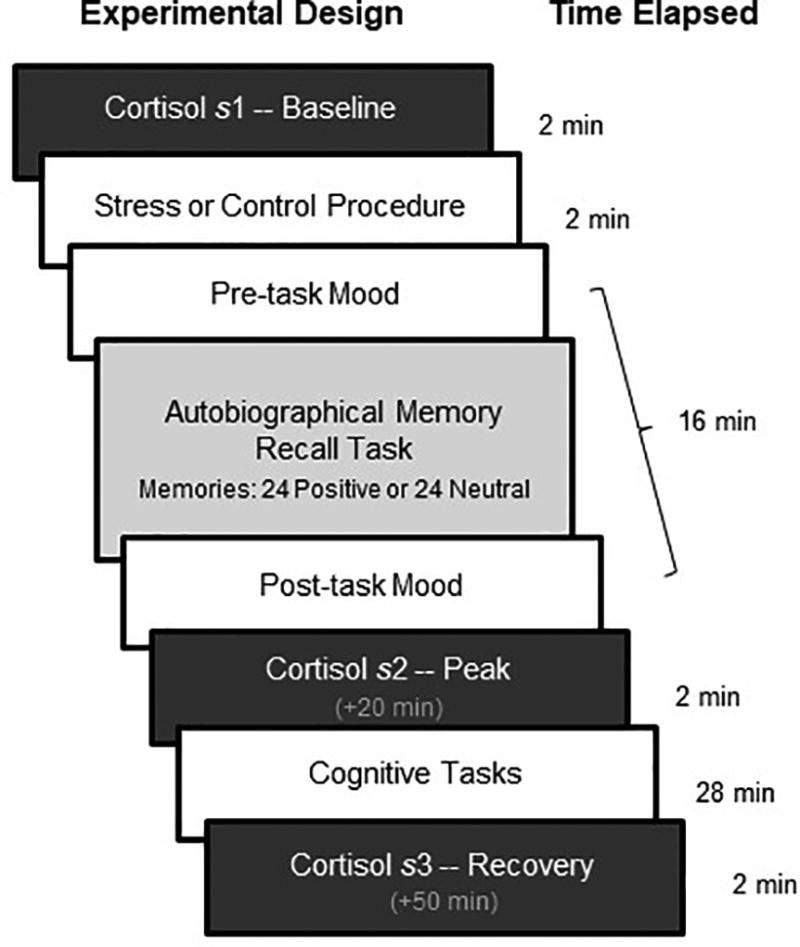
Salivary samples were collected immediately before the stress or control procedure (s1, baseline), after memory recollection when cortisol was expected to peak (s2, peak, +20 min), and at the conclusion of the experiment when cortisol was expected to recover (s3, recovery, +50 min).
We then investigated the neural correlates underlying the stress buffering effects of positive autobiographical memory retrieval. The fMRI study was nearly identical in terms of design to the behavioral study (see Methods), with the goal of comparing two groups exposed to stress that undergo different memory treatments (Stress-Positive=22; Stress-Neutral=21). We hypothesized that reminiscing about the positive past would recruit regions previously associated with positive emotion during memory recall (e.g., striatum7) and emotion regulation processes20 to overcome the detrimental consequences of acute stress.
Results
Behavioral Study
Autobiographical Memory Recollection
During the cued autobiographical memory recall task, 134 healthy adults (44 males; age= 20.8, SD= 4.2) reminisced about 24 real memories from their past prompted by common life event cues (e.g., Family Vacation)7. Event cues were selected to be either positive (e.g., Visiting Disneyland) or neutral in valence (e.g., Packing for a trip), depending on random group assignment, and were validated in a session three days prior. On each trial, participants had 14 sec to reminisce about the chosen memory and made button presses to indicate the onset and duration of recollection. For each memory, they gave subjective emotion ratings in terms of feeling (i.e., how did you feel when you recalled this memory) and emotional intensity (i.e., how intense was the particular memory).
As expected, individuals who reminisced about positive memories experienced greater positive feeling (F1,130= 422.08, p< .001) and emotional intensity (F1,130= 202.39, p< .001) than individuals who reminisced about neutral memories, regardless of stress or control condition. Although individuals who recalled positive memories rated their memories as being higher in emotional intensity based on subjective ratings, there were no differences in skin conductance responses (SCRs) across the groups (all p> .32), suggesting that individuals had similar levels of sympathetic nervous system arousal during recollection, and group differences cannot merely be explained by arousal. There were also no differences in memory onset (all p> .17) or recall duration (all p> .31) across groups, indicating that neither the memory valence nor the condition (stress, control) influenced difficulty in recall.
Recalling the Positive Past Dampens Cortisol Response after Stress Exposure
We first confirmed that our stress manipulation (SECPT)19 was effective in inducing acute stress. In accordance, stressed participants had greater skin conductance levels (SCL) during the SECPT (t125= 2.06, p= .042), which is an indicator of sympathetic nervous system arousal, and reported greater subjective ratings of stress afterwards (t132= 13.70, p< .001) than control participants (Supplementary Fig 1).
We next observed changes in cortisol over time between stress and control conditions (Fig 2a; individual data points are reported for better visualization of the data). Specifically, we calculated the area under the curve with respect to increases from baseline (AUCI) for each participant using the trapezoidal method. We selected AUCI as our measure because it takes into account both time-related changes and overall intensity of the cortisol response21 (see Supplementary Results for cortisol analyses across individual timepoints). A Condition (Stress/Control) by Valence (Positive/Neutral) ANOVA examining AUCI revealed a main effect of Condition (F1,130= 5.74, p= .018), confirming that the stress procedure elevated cortisol levels whereas the control procedure did not (MStress= 0.76, SD= 5.65; MControl= −1.30, SD= 4.40), a main effect of Valence (F1,130= 7.66, p= .006), indicating dampened cortisol levels for positive relative to neutral recall regardless of condition (MPositive= −1.47, SD= 0.61 MNeutral= 0.90, SD= 0.60), and a non-significant interaction (F1,130= 0.26, p= .609).
Figure 2. Neuroendocrine Responses to Acute Stress in the Behavioral Study.
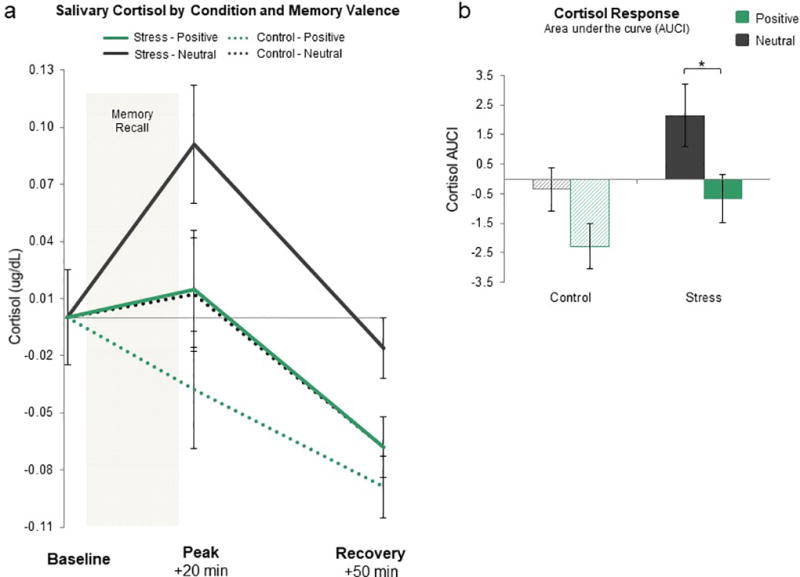
a) Baseline-corrected salivary cortisol measured in micrograms per deciliter at three time points across the experiment including immediately before the SECPT/control procedure (baseline), as well as 20 min (peak) and 50 min (recovery) after the SECPT/control procedure for all participants (N = 134). b) Cortisol response in terms of area under the curve with respect to increases from baseline (AUCI). The Stress-Neutral group has a significantly larger AUCI cortisol response than the other three groups. *p < .05; error bars denote SEM.
We then tested for group differences in AUCI to examine our specific hypothesis that recalling positive memories, but not neutral memories, would dampen the typical rise in cortisol after stress exposure. This analysis was necessary to confirm that our main effect of valence for AUCI was not simply driven by differences between Control-Positive and Control-Neutral groups. In line with our prediction, AUCI for the Stress-Positive group was significantly smaller than the Stress-Neutral group (MStress-Positive= −0.67, SD= 4.70; MStress-Neutral= 2.14, SD= 6.20; t65= −2.09, p= .041, d= 0.52, 95% confidence interval {0.1196 to 5.5004}; Fig 2b). This is particularly interesting when considering that before memory recollection the Stress-Positive group reported high subjective stress levels and had elevated SCLs just like the Stress-Neutral group (both p> .81), yet these individuals still exhibited lower cortisol levels after memory recollection. This suggests that internally generated positive emotion evoked by autobiographical recall may help dampen the heightened physiological response to acute stress.
Recalling the Positive Past Influences Mood after Stress Exposure
Given that individuals in the Stress-Positive group showed a dampened cortisol response, a critical question we sought to answer was whether recalling positive memories would also have a positive effect on mood, despite stress exposure. We assessed mood before and after memory recollection using the PANAS22.
For negative affect, we observed a significant Valence (positive/neutral) by Condition (stress/control) interaction for post-recall negative affect (F1,130= 5.53, p= .04). The Stress-Positive group reported lower negative affect after memory recollection than the Stress-Neutral group (MStress-Positive= 14.48; SD= 4.12; MStress-Neutral= 17.06; SD= 6.88; t65= −1.68, p= .069), which was trending but did not reach statistical significance.
Although there were no significant group differences for positive affect, we observed individual differences within the Stress-Positive group such that generating greater positive feelings during recollection was associated with enhanced mood (r32= 0.34, p= .05). In our prior study7, the strength of striatal activation while recalling positive memories was positively correlated with self-reported resiliency. This motivated the idea that resilient individuals who have greater adaptation to stress might be better able to utilize positive memories to increase positive feelings during recollection, in turn making them more successful in boosting their overall mood, even under stress. We explored the possibility that resiliency mediates the relation between positive emotion generated during recollection and enhanced mood after recollection using a mediation model (Fig 3).
Figure 3. Resiliency Mediates the Relationship between Emotion Ratings during Memory Recall and Subsequent Mood for Individuals in the Stress-Positive Group.
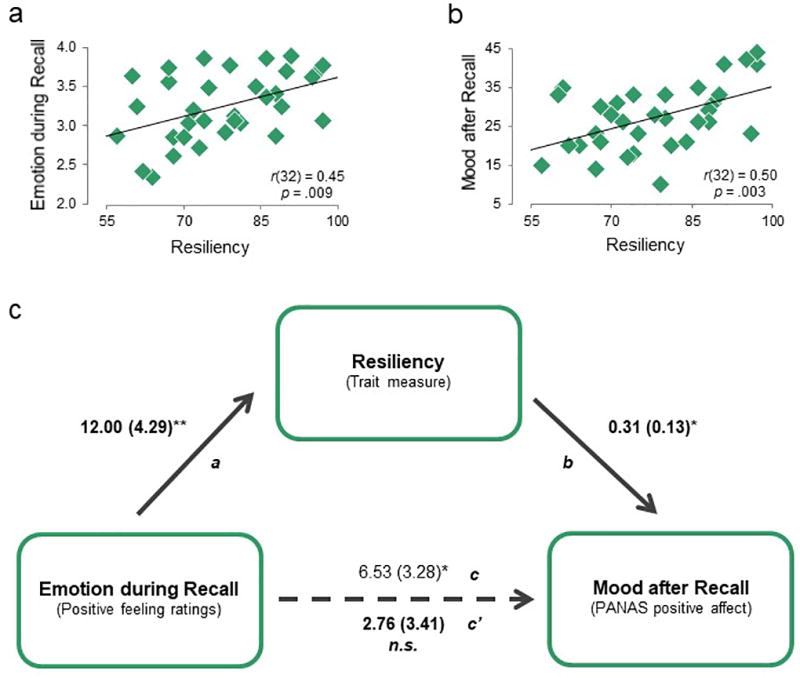
a) Greater resiliency (measured by the CD-RISC scale; N = 33) is associated with greater positive feelings during positive memory recollection. b) Greater resiliency is also associated with enhanced mood after positive memory recollection. c) Mediational model between emotion during recall, self-reported resiliency, and mood after recall. Path a: effect of emotion during recall on resiliency; path b: effect of resiliency on mood after recall, controlling for emotion during recall; path c: total effect of emotion during recall on mood after recall; and path c’: direct effect of emotion during recall on mood after recall, controlling for resilience. Path values represent unstandardized regression coefficients. *p < .05; **p < .01
For Stress-Positive individuals, greater self-reported resiliency was associated with both greater positive emotion during memory recollection (Fig 3a) and with enhanced positive mood afterwards (Fig 3b). To test whether self-reported resiliency was a mediator in this relationship, we conducted a bootstrapping analysis based on 5,000 bootstrapped samples23. The total effect of emotion during recall on mood after recall was significant (path c: B= 6.53, SE= 3.28, t32= 2.00, p= .05), but this relation was no longer significant when controlling for resiliency (direct effect, path c’: B= 2.76, SE= 3.41, t32= 0.81, p= .42: Fig 3c). Further, the indirect effect of emotion during recall (through resiliency) on subsequent mood was significant, indicating full mediation (B= 3.73, SE= 2.34, bias corrected bootstrapping 95% confidence interval {0.04 to 8.98}; see supplement for an alternative model). An important consideration is that baseline positive emotion did not differ between high and low resiliency individuals (MHigh-res= 30.4, SD= 7.43; MLow-res= 27.6, SD= 5.89; t20= 1.22, p= .233) and thus does not account for the finding. In sum, our results demonstrate that, for Stress-Positive individuals, positive emotion during memory recall related to greater resilience, which in turn related to better mood.
fMRI Study
Our behavioral findings highlight the restorative nature of positive autobiographical memory recollection under stress. We demonstrated that reminiscing about positive memories – but not neutral memories – led to a dampened rise in cortisol and lower levels of negative affect, instead of the heightened response characteristic of stress. As a next step, we sought to identify the neural mechanisms through which stress-buffering via positive memory recollection occurs. Given that recalling happy memories increases positive feelings and striatal activity7 and may serve emotion regulatory functions as per our behavioral study, we hypothesized such mechanisms to include corticostriatal systems involved in positive mood9 and emotion regulation24. We conducted an fMRI study that mirrored our behavioral design focused on the Stress-Positive and Stress-Neutral groups.
We asked a new cohort of participants (N=43) to undergo an acute stress procedure (SECPT) before fMRI scanning (see Supplementary Fig 2 for timeline). Afterwards, they reminisced about only 24 positive memories (N=22, 9 males, age= 22.4, SD= 3.3) or only 24 neutral memories (N=21, 10 males, age= 23.4, SD= 5.2) while undergoing fMRI scanning. Performance on the autobiographical memory task matched the behavioral sample. That is, the Stress-Positive group reported greater positive feeling (MStress-Positive= 2.90, SD= 0.33; MStress-Neutral= 2.06, SD= 0.52; t41= 6.41, p< .001) and emotional intensity (MStress-Positive= 2.42, SD= 0.41; MStress-Neutral= 1.76, SD= 0.47; t41= 4.89, p< .001) than the Stress-Neutral group, with no differences in memory onset or recall duration between groups (both p> .68).
Of particular significance, our cortisol results in the fMRI study replicated the behavioral study. Specifically, individuals who recalled positive memories had a smaller AUCI cortisol response than individuals who recalled neutral memories (MStress-Positive= 2.23, SD= 6.25; MStress-Neutral= 6.69, SD= 7.10; t41= −2.19, p= .035, d= 0.68, 95% confidence interval {0.3454 to 8.5746}; Fig 4a and b). Consistent with the behavioral study, this occurred even though the two stress groups did not differ in subjective ratings of stress or SCLs during the stress procedure (both p> .37). In the context of mood, Stress-Positive individuals reported less negative affect after memory recall than Stress-Neutral individuals (MStress-Positive= 11.91; SD= 2.11; MStress-Neutral= 14.90; SD= 6.01; t41= −2.20, p= .033).
Figure 4. Neuroendocrine Responses to Acute Stress in the fMRI Study.
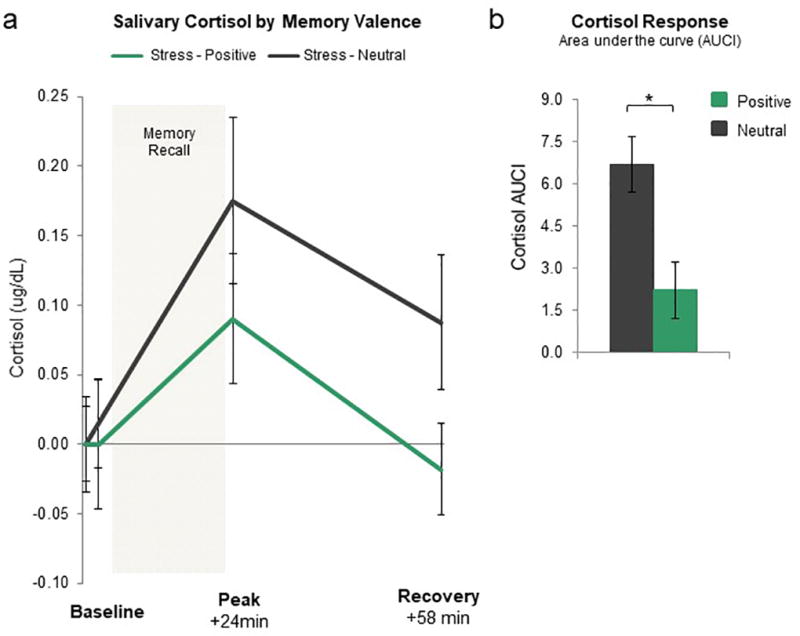
a) Baseline-corrected salivary cortisol measured in micrograms per deciliter at four time points across the experiment including immediately before the SECPT/control procedure (baseline), as well as 2 min, 24 min (peak) and 58 min (recovery) after the SECPT/control procedure for all participants (N = 43). b) Cortisol response in terms of area under the curve with respect to increases from baseline (AUCI). Stress-Neutral group has a significantly larger AUCI cortisol response than the Stress-Positive group. *p < .05; error bars denote SEM.
Reminiscing about Positive Memories to Combat Stress Recruits Regions Associated with Emotion Regulation
To examine the neural mechanisms associated with the dampening of the stress response via positive memory recollection, we conducted a random-effects whole-brain general linear model (GLM) that focused on the time of memory recall for Stress-Positive individuals. Importantly, because we hypothesized that enhancing positive emotion may be critical for reducing the stress response, we included trial-by-trial feeling ratings as a parametric modulator of memory recollection. We tested for regions whose activity increased linearly as feeling ratings increased, resulting in a statistical map set to an initial threshold of p< 0.001 (as suggested by Eklund and colleagues25) and corrected to a whole-brain cluster correction of p< 0.01 (using 216 mm3 as determined by BrainVoyager’s cluster-level threshold plugin26).
This parametric regression analysis of feeling identified regions being modulated by increases in subjective feeling ratings during memory recall for Stress-Positive individuals (Fig 5a, Supplementary Table 1). Notably, these included prefrontal regions previously implicated in cognitive control and emotion regulation such as bilateral ventrolateral prefrontal cortex17 and corticostriatal regions associated with reward-processing, such as the right ventral striatum and medial prefrontal cortex27,28. The same analysis in the Stress-Neutral group yielded no significant clusters. As a complementary analysis, we also examined feelings as a parametric modulator during memory recall contrasting the Stress-Positive relative to the Stress-Neutral groups, which revealed the right VLPFC (same peak coordinates as prior analysis) and the left DLPFC as being modulated by increases in subjective feeling ratings during recall for those who reminisced about positive, but not neutral, memories (Supplementary Table 2).
Figure 5. Neural Activity during the Recall of Autobiographical Memory under Acute Stress.
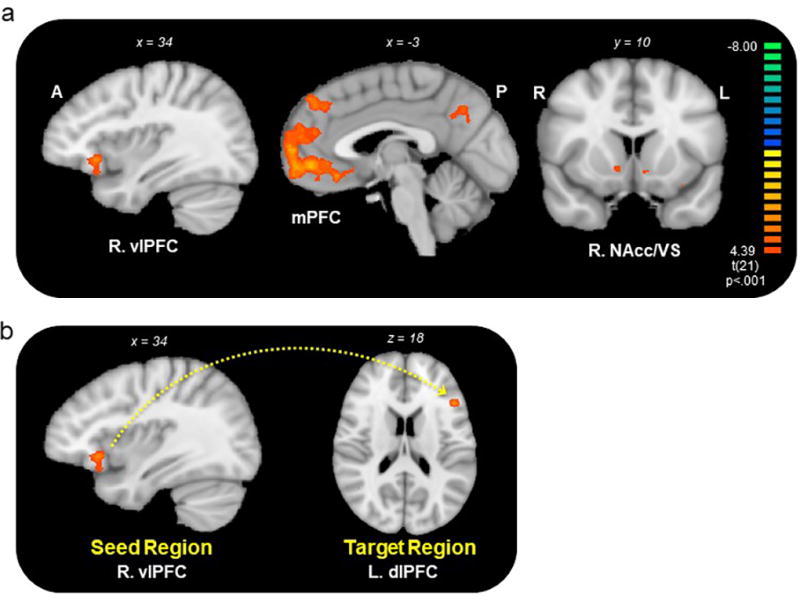
a) A parametric modulation of positive emotion ratings during memory recall in Stress-Positive individuals (N = 22) revealed activity in regions previously implicated in emotion regulation, such as the VLPFC, and in reward-processing, such as the MPFC and ventral striatum. p < 0.01, corrected. Warmer colors (yellow/orange) represent increases in activity, whereas cooler colors (green/blue) represent decreases in activity. See also Supplementary Table 1 and 2. b) A PPI analysis revealed greater connectivity between the R. VLPFC (seed region) and L. DLPFC (target region) as a function of increasing positive feeling ratings on a trial-by-trial basis for Stress-Positive individuals. p < 0.01, corrected. See also Supplementary Table 3.
VLPFC-DLPFC Connectivity Tracks Positive Feelings during Memory Recall
Our finding that recalling positive memories results in a dampened cortisol rise along with greater engagement of the VLPFC during positive recollection suggests that the ability to engage cortical regions involved in emotion regulation may be vital for combating stress. To explore this idea, we first conducted a psychophysiological interaction (PPI) analysis to identify neural regions that were functionally connected to the prefrontal cortex as a function of subjective feelings during positive memory recollection. We defined our VLPFC seed regions bilaterally based on our prior analyses, which showed this region being prominently activated in the Stress-Positive group during memory recall and in comparison to the Stress-Neutral group. For both seed regions, we performed a random-effects whole brain analysis for the parametric modulation of feeling ratings during recall for Stress-Positive individuals (using an initial threshold of p< 0.001 and the same cluster correction described above). Our PPI analysis with the right VLPFC seed (x,y,z: 35, 22, −3) revealed the left DLPFC (x,y,z: −46, 22, 18) as exhibiting greater connectivity (as a function of increasing feeling ratings) for the Stress-Positive group (Fig 5b, Supplementary Table 3). The left VLPFC seed region yielded no target regions that reached statistical significance. To examine the relationship between functional connectivity and the physiological stress response, we tested for correlation between our PPI parameter estimates (indexing the degree of connectivity between VLPFC and DLPFC as a function of feeling ratings) and cortisol in Stress-Positive individuals. Although approaching significance, the association between greater VLPFC-DLPFC connectivity and lower AUCI cortisol levels was not significant (r21= −0.35, p= .11). Taken together, our fMRI results provide converging evidence that engagement of cortical regions previously linked to emotion regulatory functions may be significant for enhancing or sustaining pleasant feelings during positive reminiscence, and thus dampening the physiological stress response.
Discussion
Acute stress elicits negative emotion1, lessens our ability to use cognitive emotion regulation3, diminishes responsiveness to rewards29, and is often a precursor to anxiety and depressive episodes14, making it imperative to identify effective strategies for reducing stress. Across two studies, our results showed that reminiscing about positive – but not neutral memories – buffered the physiological and emotional consequences of acute stress. Specifically, individuals who recalled positive memories showed a dampened rise in cortisol and reported lower levels of negative affect 20 min after stress exposure, resembling the non-stressed control groups. In contrast, recalling neutral memories under stress resulted in a heightened cortisol rise that is typical of the acute stress response10. Recalling positive memories also served to enhance mood despite stress exposure, but only for individuals with greater self-reported resiliency. For Stress-Positive individuals but not Stress-Neutral individuals, we observed greater activity in regions previously implicated in emotion regulation (e.g., VLPFC) and reward-processing (e.g., striatum) based on a parametric modulation of emotion ratings during memory recall. Further, we observed greater VLPFC-DLPFC connectivity as a function of increasing positive emotion. Our results underscore the restorative and protective function of self-generated positive emotions in the face of stress.
The finding that positive memory retrieval restored stress-induced deficits, such as alleviating negative affect and calming the physiological stress response (i.e., HPA-axis), might suggest a role for recalling positive (but not neutral) memories in motivating a more positive perspective that interrupts the ongoing experience of a stressful event. This possibility lends support to the idea that bolstering positive emotion broadens one’s cognitive perspective, in contrast to the narrowed perspective that occurs during negative affective states5. Indeed, the experience of positive emotion over time helps build psychological resources for adaptive coping, making it more likely to continue experiencing positive emotions in the future, and is perhaps a potential mechanism by which resiliency is built30. Notably, we observed that greater resiliency was associated with additional protective benefits for those who were given the opportunity to recall positive memories. This is consistent with research linking high resiliency to better adaptability to stress, such as faster cardiovascular recovery, more efficient and successful emotion regulation, and greater positive meaning finding30,31.
Our fMRI results highlight the significant relationship between experiencing positive emotion, greater engagement of prefrontal regions involved in emotion regulation, and lower cortisol after stress exposure. Acute stress is well known to compromise the prefrontal cortex in humans4, impairing self-regulation32, cognitive control, and task-relevant processing33, which diminishes our ability to adapt to the environment. Consistent with this, we did not observe prefrontal activity (VLPFC, DLPFC) in Stress-Neutral individuals (i.e., had higher cortisol responses). Although this null result should be treated with caution, it is noteworthy that these same regions were spared in Stress-Positive individuals (i.e., had lower cortisol responses), and the strength of their connectivity increased as a function of positivity. In light of this observation, we speculate that the effective use of memory recall to enhance positive emotion may serve emotion regulatory functions under stress. While it is possible that recruitment of prefrontal activity in the Stress-Positive group is due to more general cognitive control functioning, such as controlled memory retrieval34, both groups underwent a recall procedure, thus supporting a more emotionally driven explanation for prefrontal engagement and suggesting a potential mechanism by which positive memory recall may contribute to stress dampening.
The VLPFC and DLPFC are thought to play pivotal yet distinct roles in successful emotion regulation, particularly cognitive reappraisal17,20. For example, a recent meta-analysis revealed that both regions were significantly activated across 48 studies examining cognitive reappraisal of primarily negative emotional stimuli17. The DLPFC may aid such processes as working memory and cognitive flexibility, including manipulating mental representations of affective states to regulate emotion, whereas the VLPFC may serve response selection and inhibitory functions35 and is also linked to the cognitive control of memory34. Consistent with prior work36, one possibility in our study is that the VLPFC attempts to override negative appraisals during memory retrieval, while the DLPFC helps flexibly change one’s emotional state. Previous studies have also shown an association between greater VLPFC activity and either decreased amygdala activity or increased reward-related activity (e.g., striatum) to successfully regulate emotion24. Although we did not observe this in the present study, the link between different cortical regions and the striatum underlying regulation of positive mood9, particularly with respect to stress, is an important future inquiry.
It is worth considering how the strategy of savoring positive memories to combat stress relates to other emotion regulation strategies. Mindfulness-meditation has been shown to reduce stress and promote well-being through the nonjudgmental practice of self-awareness in the present moment37. Yet, physiological changes after mindfulness training are mixed as some studies show decreased cortisol whereas others show either an increase or no change (for review see38). Variability in type and length of mindfulness training and type of stressor may help explain mixed findings, although it is difficult to speculate given the paucity of studies thus far. The core idea of mindfulness – to focus on the present rather than the past or future – may seem at odds with the strategy we propose. However, savoring the past involves deliberate attention to an enjoyable experience with the aim of cultivating positive emotions, which is distinctly different than ruminating about past negative events – a characteristic of depression. Much like mindfulness, recalling positive memories motivates a broader perspective, and perhaps directs attention away from the current stressor in favor of something more positive or relaxing. This is in contrast to strategies that focus on reinterpreting the stressor as a way to diminish its meaning2, which may be less effective under stress3. Other strategies that may buffer stress via positive emotion include high self-esteem, receiving social support or positive feedback, and affirming personal values39. However, these strategies depend on situational or personality characteristics, leaving unclear their efficacy in alleviating stress in everyday life.
Our findings have broad implications for better understanding the stress response in the context of mood disorders. For instance, individuals with depression not only have difficulty in retrieving positive memories40, but are also sensitive to the effects of stress. That is, they have difficulty regulating negative emotion41, report lower levels of resilience, and have higher cortisol levels during recovery from stress42, suggesting a critical need for understanding positive emotion deficits in depression. Thus unsurprisingly, one aim of behavioral activation therapy for depression is identifying and reengaging in positive activities that reinforce and enhance wellbeing, including positive reminiscence43,44. Consistent with this idea, a recent study showed that optogenetically reactivating neural circuits associated with positive experiences in rodents can lessen depression-like behaviors caused by stress45, providing complementary evidence that thinking about the past in a positive light can recruit reward-related neural circuits in humans7 and serve as a potentially effective way to reduce stress. In the present study, we also found that stressed individuals who recalled positive memories had a greater increase in ventral striatum activity as a function of increasing positive feelings (see Fig 5a), as well as for high relative to low feeling memories as compared to those who recalled neutral memories under stress (see Supplementary Fig 3 and Table 4). Thus, corticostriatal circuits recruited during reminiscing about positive memories are involved in increases in positive emotion that are linked to the coping of stress. This corroborates prior work showing that individuals with lower daily cortisol output tend to be happier46 and exhibit greater sustained neural activity to positive stimuli in the striatum47. Our approach extends these findings by demonstrating that we can use something we already do naturally – recalling the positive past – to buffer the detrimental effects of acute stress in the present moment.
There are some considerations about our study that warrant mention. First, our results showed dampening of the acute stress response, but cannot necessarily speak to the effectiveness of this strategy for different stress levels. Recurrent heightened levels of cortisol observed in chronic stress can be detrimental, for instance, by suppressing the immune system, increasing susceptibility to disease48, and resulting in atrophy and reduced neurogenesis in the hippocampus, a region where memory processing and storage occurs49. Thus, it may be useful to probe if increasing positive emotions can help build psychological resources to cope with chronic stress. Second, positive memory retrieval may not be effective for everyone. There may be individual differences leading some individuals to have fewer, less detailed or vivid positive memories or general difficulty in recalling their past (e.g., depression). This may be especially challenging for individuals who are more susceptible to stress (i.e., less resilient). Although the present study establishes positive recall as an effective stress-buffering strategy in a healthy population, it will be essential to test whether stress vulnerable individuals are able to self-generate positive memories after stress exposure and the efficacy of this strategy in sustaining reductions in negative affect and neuroendocrine responses, especially under less resilience.
It is also important to consider alternative accounts for our findings. One possibility is that greater interest, engagement or even distraction during positive relative to neutral recall may explain cortisol differences. Yet, we found this not to be the case (as measured by vividness ratings and recall duration; see Supplementary Results). There is also evidence to suggest that distraction alone does not lead to stress dampening, as stress participants who perform highly distracting working memory tasks still exhibit the typical cortisol rise11. Additionally, given that all participants recalled past experiences to elicit positive emotion, it is unclear whether other strategies aimed at increasing positive emotion would also be effective. For instance, positive feelings may be enhanced by engaging in positive mental time travel about the future50. Another such strategy is the use of positive imagery, which may be effective for mitigating pain51. However, in a prior study we found positive imagery to be less effective than positive memory retrieval in enhancing mood7, which may be vital for reducing stress. Finally, while our focus was to examine the stress-buffering nature of positive reminiscence, it is also important to note that stress can be beneficial in certain contexts. Specifically, stress hormones (e.g., cortisol) exhibit an inverted U-shaped response curve, such that extreme levels (both low and high), but not moderate levels, impair cognitive performance including memory52,53. Such cortisol effects on performance can further depend on age54,55, exposure to novelty56, and the presence of other hormones, such as oxytocin. Oxytocin, for instance, may have been a positive mediator in the present study given its role in modulating fear and anxiety57. While beyond the scope of the current study, future work could explore the role of varying levels of cortisol and other stress-modulating hormones on the emotion regulatory function of positive recall.
Although stress can be adaptive for learning and cognitive performance58, sometimes our stress response (e.g., panic attack) is out of proportion to the stressor (e.g., studying for a test), compromising our ability to use cognitive emotion regulation skills we already know3. Our results highlight a more proactive way to alleviate stress. Rather than attempting to decrease negative feelings or deliberately reinterpret the meaning of a stressful experience, which may be effective in only certain contexts or for particular individuals59, one might focus on increasing positive feelings instead. We demonstrate that this can even be done with a strategy – recalling pleasant memories – that is unrelated to the stressor. When uncontrolled, psychological stress can drive us far from a desirable state. Enhancing positive feelings when reminiscing about the past may be one way to bring us back.
Methods
Behavioral Study
Participants
Healthy undergraduate students (N=149) at Rutgers University completed Day 1 and returned for Day 2 only if they met inclusion criteria (see Day 1 procedures; 139 met criteria). Additional exclusions included not following directions (<50% responses; N=2) and extreme cortisol responses (>3 SD from mean; N=3). Participants from the final sample (N=134; 44 males; mean age= 20.8, SD= 4.2) were randomly assigned to four experimental groups: Stress-Neutral (N=34), Control-Neutral (N=34), Stress-Positive (N=33), and Control-Positive (N=33). We chose thirty-five participants per group as our target sample size to attain an effect size comparable to prior stress studies, which typically have a between-subjects design with 30–35 participants per group3. Participants gave informed consent in accordance with the Rutgers University Institutional Review Board for the Protection of Human Subjects in Research and received course credit for their participation.
Experimental Design
Day 1: Autobiographical memory questionnaire
Participants were presented with 84 common life event cues (e.g., Family Vacation). For each cue, participants selected a real memory in which they had been personally involved and had occurred at a specific place and time. Participants then reported a description, location, date, and gave subjective ratings for valence (positive or neutral), emotional intensity (i.e., how intense was the memory; 1–4: 1= not intense, 4= very intense), and feeling (i.e., how did you feel when you recalled this memory; 1–4: 1= neither good nor bad, 4= very good). Importantly, memories were positive (e.g. visiting Disneyland) or neutral (e.g. packing for a trip), but not negative memories (e.g., lost luggage).
Only participants who reported at least 24 positive or 24 neutral memories (depending on random assignment) returned for Day 2. For each participant, the 24 most positive (or neutral) cues were used in the memory recall task on Day 2. Participants also completed the Connor-Davidson Resiliency scale60 and the Beck Depression Inventory61.
Day 2: Stress induction and memory recall
The second session (three days later) was run between 1:00pm and 5:30pm to account for diurnal variations in cortisol levels10. Participants were informed of the stress procedure and notified that they could withdraw from the experiment at any time. Day 2 included: 1) salivary cortisol collection s1, 2) stress induction or control procedure, 3) pre-task mood assessment, 4) cued recall autobiographical memory task of either only positive memories or of only neutral memories, 5) post-task mood assessment, 6) salivary cortisol collection s2, 7) cognitive tasks, 8) salivary cortisol collection s3 (Fig 1).
Stress Induction
We used the Socially Evaluative Cold Pressor task (SECPT)19 for induction of acute stress. Stress participants were videotaped while immersing their hand into ice water (1–3°C) for 2 min. The experimenter dressed in a white lab coat recorded the participant and acted neutral. Participants were told that the recording would be used for further analysis after the session. The control task was identical except that participants immersed their hand in warm water (23–25°C), there was no video camera, and no lab coat. Afterwards, participants rated how unpleasant, stressful, and painful it was ranging from 0 (not at all) to 100 (very much). The sum of these three ratings created a subjective stress rating.
Neuroendocrine Measurement and Analysis
Salivary samples were collected to assess cortisol concentrations, via swab placed under the tongue for 2 min. Swabs were kept in cold storage at −10°C until sent to Salimetrics Laboratory for duplicate biochemical assay analysis. To assess cortisol change across time, we calculated the area under the curve with respect to increases from baseline (AUCI) for each participant using the trapezoidal method.
To assess sympathetic nervous system arousal, we measured skin conductance via electrodes placed on the participant’s fingers, sampled at 200 Hz using an MP100 Data Acquisition Module (Biopac Systems). During the 2 min SECPT/control procedure, skin conductance levels (SCL) were measured as the mean tonic activity. During the memory task, skin conductance responses (SCR) were assessed via the sum of trough-to-peak waveform amplitude responses (in microsiemens, µS) across all trials (0.5s to 14.5s window; square-root transformed). Responses lower than 0.02 µS were recorded as zero. Data were preprocessed by low-pass filtering (25 Hz cut-off) and mean-value smoothing using a three-sample window.
Autobiographical Recall Task
Participants first reported their current mood state via the PANAS22. Then, they completed a cued recall autobiographical memory task where they reminisced about 24 positive memories (Positive groups) or 24 neutral memories (Neutral groups) triggered by event cues from their Day 1 questionnaire. Each trial included one written event cue displayed for 14s. Participants recalled the same memory from Day 1 and elaborated on it until the 14s were up. Participants made button presses to indicate the ‘‘beginning’’ (i.e., when it began to form) and ‘‘end’’ of their memory (if they finished elaborating before time was up). After a delay of 2–4s, participants rated the memory for emotional intensity and feeling (3.5s each). The length of one trial was 24 sec, with a delay of 6–10s separating one trial from the next. Afterwards, participants rated their post-recall mood state via the PANAS. We did not assess mood before the stressor, because we assumed baseline mood levels would be similar across groups given random assignment.
Behavioral Analysis
Group differences in subjective stress ratings, SCL/SCR, AUCI cortisol response, autobiographical memory task performance, and mood were tested using Condition (stress/control) by Valence (positive/neutral) ANOVAs. We included demographic variables as covariates in our analyses such as age, depressive symptoms (BDI), resiliency (CD-RISC), gender, and menstrual cycle phase (collected for 64 out of 90 female participants; luteal phase= 40; follicular phase= 24). None of these factors significantly impacted the results.
fMRI Study
Participants
Fifty-two healthy adults participated. Exclusions included not following directions (<50% responses, N=1), extreme cortisol responses (N=7), and claustrophobia (N=1). Participants from the final sample (N=43) were randomly assigned to two experimental groups: Stress-Positive (N=22, 9 males, age= 22.4, SD= 3.3) or Stress-Neutral (N=21, 10 males, age= 23.4, SD= 5.2). Participants gave informed consent in accordance with the Rutgers University Institutional Review Board for the Protection of Human Subjects in Research and received compensation for their participation.
Experimental Design
Day 1: Autobiographical memory questionnaire
This session was identical to the behavioral study.
Day 2: Stress induction and fMRI scanning
Participants returned for the second session two to four days later. Day 2 included: 1) salivary cortisol collection s1 (baseline), 2) stress induction via the SECPT in the scanning environment, 3) salivary cortisol collection s2, 4) Set-up in the scanner, 5) pre-task mood assessment, 6) cued recall autobiographical memory task of either only positive memories or of only neutral memories, 7) post-task mood assessment, 8) salivary cortisol collection s3 (+24 min, peak), 9) reward task, 10) salivary cortisol collection s4 (+58 min, recovery; Supplementary Fig 2).
We used the same memory recall task as described previously for the behavioral design (24 positive or 24 neutral memories depending on group assignment). Cortisol collection and SECPT administration were identical to the behavioral study with minor changes to be compatible with the fMRI scanner environment. For instance, all cortisol samples were collected while the participant was in the scanner room. To allow for the participant to acclimate to the scanner environment and for salivary cortisol to stabilize, the baseline sample (s1) was collected 30 min after the participant arrived and 10 min after being in the scanner room. The peak cortisol response (s3) was the only sample collected while the participant was in the scanner because it occurred between runs of the memory task and the reward task. While the fMRI design had an additional cortisol sample (as described above), which was included in this analysis, it is important to note that this sample did not differ from the baseline cortisol measurement (taken 2 min earlier; p= .525) given the slow nature of cortisol release after stress exposure (10–15 min)10. For our stress protocol (SECPT), participants immersed their hand in ice cold water (1–3°C) for 2 min, while sitting in the scanner room. The experimenter dressed in a white lab coat videotaped the participant from the doorway. Consistent with the behavioral study, we collected SCR during the SECPT and subjective stress ratings after.
We also asked participants to perform a surprise monetary reward task (the card-guessing game adapted from62) while still in the scanner. This task was a surprise as to not influence the prior memory task and mood ratings. The purpose was to identify reward-related regions of interest (ROIs) to serve as independent ROIs to test with high and low feeling memory regressors across groups. In each trial of the card task, participants saw a card with a question mark inside for 2s. They guessed whether the card’s value was higher or lower than the number 5 via button press. After a 2–4s delay, the card and monetary outcome were displayed for 2s. A correct response earned a green checkmark signifying a gain of $1.00 whereas an incorrect response earned a red X signifying a loss of $0.50. Unbeknownst to participants, outcomes were predetermined to control schedule of reinforcement and number of gain and loss trials (20 each for a total of 40 trials). A trial lasted 9s, with a delay of 4–6s separating one trial from the next.
At the conclusion of the scanning session, participants were debriefed and compensated for their time in the scanner and bonus money earned in the card game.
fMRI Data Acquisition
A 3T Siemens Magnetom Trio scanner was used for acquisition of T2-weighted MPRAGE structural images (256 × 256 matrix, FOV= 256 mm, 176 1-mm sagittal slices). Functional images were taken in 35 contiguous oblique-axial slices (3×3×3 mm voxels) prescribed parallel to the AC-PC plane with a single shot gradient echo EPI sequence (TR= 2s, TE= 25ms, FOV= 192, flip angle 90, bandwidth= 2232 Hz/Px, echo spacing= 0.51). Data were preprocessed and analyzed using BrainVoyager QX (v2.8, Brain Innovation). Functional images were motion-corrected (six parameters), slice-timing corrected using a cubic spline interpolation, and spatially smoothed using a Gaussian kernel of 4mm FWHM. Further, the data were temporally smoothed with voxelwise linear detrending and high-pass filtering of frequencies (three cycles per time course). The images were spatially normalized to the Talairach stereotaxic space63.
fMRI Data Analysis
Functional data were analyzed using a whole brain random-effects general linear model (GLM). The memory task was modeled using a regressor for memory recall, a parametric regressor for subjective feeling ratings during memory recall (orthogonalized with respect to the memory regressor), and a regressor representing missed trials (i.e., no valence rating given for the memory, 1.6% missed trials). The memory regressor and feeling parametric regressor began at memory formation and ended after elaboration, with this period defined by participants’ own button presses in each trial (for onset and conclusion of memory recall). We performed three analyses. We first examined the parametric modulation of feeling in each group separately, and then a contrast of Stress-Positive > Stress-Neutral to examine group differences in neural activity as a function of subjective feeling ratings during memory recollection. This allowed us to test for regions whose activity increased linearly as feeling ratings increased on a trial-by-trial basis for each of these analyses.
The monetary reward task was modeled using two regressors representing gain and loss trials during the 2s outcome phase along with a regressor representing missed trials (no response). We conducted a contrast of gain and loss outcomes to identify reward-related ROIs. Using the functionally defined reward ROIs in the striatum, we then ran a GLM using high (rating of 3 or 4) and low (rating of 1 or 2) feeling memory regressors. The goal of this analysis was to confirm that a “reward-related” functionally defined ROI would show an independent effect of high versus low feeling during memory recall for the Stress-Positive relative to the Stress-Neutral group.
For both the memory and monetary reward tasks, regressors were convolved with a canonical double-gamma hemodynamic response function and six regressors for motion parameters were included in the model. To correct for multiple comparisons, we used the cluster level statistical threshold plugin in Brain Voyager26. This plugin employs Monte Carlo simulations to determine the likelihood that observed clusters of activation are significant and not false positives (over 1000 iterations), resulting in a whole brain corrected threshold of p< .01. After correction, the map automatically applies the minimum cluster size threshold that produces the desired cluster-level false-positive alpha rate (1% was chosen). For the memory task, we applied a voxel cluster threshold of 8 contiguous voxels (216mm3 as determined by the plugin) defined at a threshold of p< 0.001 to obtain a corrected alpha< 0.01. Because our goal for the monetary reward task was to simply identify independent ROIs, we applied a more stringent initial threshold of p< 0.000001 (which required a voxel cluster threshold of 1 voxel, 27mm3, to obtain a corrected alpha< 0.01).
Psychophysiological Interaction Analysis
To identify neural regions that were functionally connected to cortical regions identified in our main contrast of memory recall as a function of subject feeling ratings during positive memory recollection, we conducted an exploratory psychophysiological interaction (PPI) analysis. We chose our prefrontal cortex seed regions based on our parametric modulation of feeling during memory recollection (e.g., right and left VLPFC). The regressor of interest – the PPI interaction term – was created by calculating the element-by-element product of the seed region time series (physiological factor) and trial-by-trial subjective feeling ratings (psychological factor) during the memory recollection task. Each PPI model included regressors for the interaction term (psychophysiological factor), the time series of the seed region (physiological factor), and trial-by-trial subjective feeling ratings (psychological factor). For each subject, we extracted volumes of interest to use as seeds in single-subject whole-brain PPI analyses. These were then combined into a group level model for performing a random-effects whole brain analysis to identify regions exhibiting connectivity with the seed region. To correct for multiple comparisons, we set an initial threshold of p< 0.001 and applied a cluster correction of 8 contiguous voxels (216mm3) to obtain a corrected alpha< 0.01.
Supplementary Material
Acknowledgments
This research was supported by the National Institute on Drug Abuse (DA027764). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. The authors thank Jamil Bhanji for helpful comments and discussion, and Heena Manglani and Eunbin Kim for assistance with data collection.
Footnotes
Data Availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Author Contributions
M.E.S. and M.R.D. designed the experiments. M.E.S performed the experiments and analyzed data. M.E.S. and M.R.D. wrote the manuscript and approved of the final version for publication.
Competing Interests
The authors declare no competing interests.
References
- 1.Lazarus RS, Folkman S. Stress, appraisal and coping. Springer; 1984. [Google Scholar]
- 2.Gross JJ. Emotion regulation: Affective, cognitive, and social consequences. Psychophysiology. 2002;39:281–291. doi: 10.1017/s0048577201393198. [DOI] [PubMed] [Google Scholar]
- 3.Raio CM, Orederu TA, Palazzolo L, Shurick AA, Phelps EA. Cognitive emotion regulation fails the stress test. Proc. Natl. Acad. Sci. U. S. A. 2013;110:15139–44. doi: 10.1073/pnas.1305706110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Arnsten AFT. Stress signaling pathways that impair prefrontal cortex structure and function. Nat. Rev. Neurosci. 2009;10:410–422. doi: 10.1038/nrn2648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Catalino LI, Fredrickson BL. A Tuesday in the life of a flourisher: The role of positive emotional reactivity in optimal mental health. Emotion. 2011;11:938–50. doi: 10.1037/a0024889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bower GH. Mood and memory. Am. Psychol. 1981;36:129–48. doi: 10.1037//0003-066x.36.2.129. [DOI] [PubMed] [Google Scholar]
- 7.Speer ME, Bhanji JP, Delgado MR. Savoring the past: Positive memories evoke value representations in the striatum. Neuron. 2014;84:847–856. doi: 10.1016/j.neuron.2014.09.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Heller AS, et al. Reduced capacity to sustain positive emotion in major depression reflects diminished maintenance of fronto-striatal brain activation. Proc. Natl. Acad. Sci. U. S. A. 2009;106:22445–22450. doi: 10.1073/pnas.0910651106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Admon R, Pizzagalli DA. Corticostriatal pathways contribute to the natural time course of positive mood. Nat. Commun. 2015;6:10065. doi: 10.1038/ncomms10065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kirschbaum C, Hellhammer DH. Salivary cortisol in psychoneuroendocrine research: Recent developments and applications. Psychoneuroendocrinology. 1994;19:313–333. doi: 10.1016/0306-4530(94)90013-2. [DOI] [PubMed] [Google Scholar]
- 11.Schoofs D, Wolf OT, Smeets T. Cold pressor stress impairs performance on working memory tasks requiring executive functions in healthy young men. Behav. Neurosci. 2009;123:1066–1075. doi: 10.1037/a0016980. [DOI] [PubMed] [Google Scholar]
- 12.Porcelli AJ, Delgado MR. Acute stress modulates risk taking in financial decision making. Psychol. Sci. 2009;20:278–83. doi: 10.1111/j.1467-9280.2009.02288.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Porcelli AJ, Delgado MR. Stress and decision making: effects on valuation, learning, and risk-taking. Curr. Opin. Behav. Sci. 2017;14:33–39. doi: 10.1016/j.cobeha.2016.11.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kendler KS, Karkowski LM, Prescott CA. Causal relationship between stressful life events and the onset of major depression. Am. J. Psychiatry. 1999;156:837–841. doi: 10.1176/ajp.156.6.837. [DOI] [PubMed] [Google Scholar]
- 15.Porcelli AJ, Lewis AH, Delgado MR. Acute stress influences neural circuits of reward processing. Front. Neurosci. 2012;6:1–9. doi: 10.3389/fnins.2012.00157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Folkman S, Lazarus RS, Gruen RJ, DeLongis A. Appraisal, coping, health status, and psychological symptoms. J. Pers. Soc. Psychol. 1986;50:571–579. doi: 10.1037//0022-3514.50.3.571. [DOI] [PubMed] [Google Scholar]
- 17.Buhle JT, et al. Cognitive reappraisal of emotion: A meta-analysis of human neuroimaging studies. Cereb. Cortex. 2014;24:2981–90. doi: 10.1093/cercor/bht154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Delgado MR, Nearing KI, Ledoux JE, Phelps Ea. Neural circuitry underlying the regulation of conditioned fear and its relation to extinction. Neuron. 2008;59:829–38. doi: 10.1016/j.neuron.2008.06.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Schwabe L, Haddad L, Schachinger H. HPA axis activation by a socially evaluated cold-pressor test. Psychoneuroendocrinology. 2008;33:890–5. doi: 10.1016/j.psyneuen.2008.03.001. [DOI] [PubMed] [Google Scholar]
- 20.Morawetz C, Bode S, Derntl B, Heekeren HR. The effect of strategies, goals and stimulus material on the neural mechanisms of emotion regulation: A meta-analysis of fMRI studies. Neurosci. Biobehav. Rev. 2017;72:111–128. doi: 10.1016/j.neubiorev.2016.11.014. [DOI] [PubMed] [Google Scholar]
- 21.Pruessner JC, Kirschbaum C, Meinlschmid G, Hellhammer DH. Two formulas for computation of the area under the curve represent measures of total hormone concentration versus time-dependent change. Psychoneuroendocrinology. 2003;28:916–931. doi: 10.1016/s0306-4530(02)00108-7. [DOI] [PubMed] [Google Scholar]
- 22.Watson D, Clark LA, Tellegen A. Development and validation of brief measures of positive and negative affect: the PANAS scales. J. Pers. Soc. Psychol. 1988;54:1063–70. doi: 10.1037//0022-3514.54.6.1063. [DOI] [PubMed] [Google Scholar]
- 23.Preacher KJ, Hayes AF. SPSS and SAS procedures for estimating indirect effects in simple mediation models. Behav. Res. Methods, Instruments, Comput. 2004;36:717–731. doi: 10.3758/bf03206553. [DOI] [PubMed] [Google Scholar]
- 24.Wager TD, Davidson ML, Hughes BL, Lindquist MA, Ochsner KN. Prefrontal-subcortical pathways mediating successful emotion regulation. Neuron. 2008;59:1037. doi: 10.1016/j.neuron.2008.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Eklund A, Nichols TE, Knutsson H. Cluster failure: Why fMRI inferences for spatial extent have inflated false-positive rates. Proc. Natl. Acad. Sci. 2016;113:7900–5. doi: 10.1073/pnas.1602413113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Goebel R, Esposito F, Formisano E. Analysis of functional image analysis contest (FIAC) data with brainvoyager QX: From single-subject to cortically aligned group general linear model analysis and self-organizing group independent component analysis. Hum. Brain Mapp. 2006;27:392–401. doi: 10.1002/hbm.20249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.O’Doherty JP. Beyond simple reinforcement learning: the computational neurobiology of reward-learning and valuation. Eur. J. Neurosci. 2012;35:987–90. doi: 10.1111/j.1460-9568.2012.08074.x. [DOI] [PubMed] [Google Scholar]
- 28.Delgado MR. Reward-related responses in the human striatum. Ann. N. Y. Acad. Sci. 2007;1104:70–88. doi: 10.1196/annals.1390.002. [DOI] [PubMed] [Google Scholar]
- 29.Bogdan R, Pizzagalli DA. Acute stress reduces reward responsiveness: Implications for depression. Biol. Psychiatry. 2006;60:1147–54. doi: 10.1016/j.biopsych.2006.03.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ong AD, Bergeman CS, Bisconti TL, Wallace KA. Psychological resilience, positive emotions, and successful adaptation to stress in later life. J. Pers. Soc. Psychol. 2006;91:730–49. doi: 10.1037/0022-3514.91.4.730. [DOI] [PubMed] [Google Scholar]
- 31.Tugade MM, Fredrickson BL. Resilient individuals use positive emotions to bounce back from negative emotional experiences. J. Pers. Soc. Psychol. 2004;86:320–33. doi: 10.1037/0022-3514.86.2.320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Heatherton TF, Wagner DD. Cognitive neuroscience of self-regulation failure. Trends Cogn. Sci. 2011;15:132–139. doi: 10.1016/j.tics.2010.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Liston C, McEwen BS, Casey BJ. Psychosocial stress reversibly disrupts prefrontal processing and attentional control. Proc. Natl. Acad. Sci. U. S. A. 2009;106:912–917. doi: 10.1073/pnas.0807041106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Badre D, Poldrack Ra, Paré-Blagoev EJ, Insler RZ, Wagner AD. Dissociable Controlled Retrieval and Generalized Selection Mechanisms in Ventrolateral Prefrontal Cortex. Neuron. 2005;47:907–918. doi: 10.1016/j.neuron.2005.07.023. [DOI] [PubMed] [Google Scholar]
- 35.Robbins T. Shifting and stopping: fronto-striatal substrates, neurochemical modulation and clinical implications. Philos. Trans. R. Soc. B Biol. Sci. 2007;362:917–932. doi: 10.1098/rstb.2007.2097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Morawetz C, Bode S, Baudewig J, Kirilina E, Heekeren HR. Changes in Effective Connectivity Between Dorsal and Ventral Prefrontal Regions Moderate Emotion Regulation. Cereb. Cortex. 2016;26:1923–1937. doi: 10.1093/cercor/bhv005. [DOI] [PubMed] [Google Scholar]
- 37.Baer RA. Mindfulness training as a clinical intervention: A conceptual and empirical review. Clin. Psychol. Sci. Pract. 2003;10:125–143. [Google Scholar]
- 38.Tang Y-Y, Hölzel BK, Posner MI. The neuroscience of mindfulness meditation. Nat. Rev. Neurosci. 2015;16:1–13. doi: 10.1038/nrn3916. [DOI] [PubMed] [Google Scholar]
- 39.Creswell JD, et al. Affirmation of personal values buffers neuroendocrine and psychological stress responses. Psychol. Sci. 2005;16:846–851. doi: 10.1111/j.1467-9280.2005.01624.x. [DOI] [PubMed] [Google Scholar]
- 40.Young KD, Bellgowan PSF, Bodurka J, Drevets WC. Behavioral and neurophysiological correlates of autobiographical memory deficits in patients with depression and individuals at high risk for depression. JAMA Psychiatry. 2013:1–10. doi: 10.1001/jamapsychiatry.2013.1189. [DOI] [PubMed] [Google Scholar]
- 41.Greening SG, Osuch EA, Williamson PC, Mitchell DGV. The neural correlates of regulating positive and negative emotions in medication-free major depression. Soc. Cogn. Affect. Neurosci. 2014;9:628–637. doi: 10.1093/scan/nst027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Burke HM, Davis MC, Otte C, Mohr DC. Depression and cortisol responses to psychological stress: A meta-analysis. Psychoneuroendocrinology. 2005;30:846–56. doi: 10.1016/j.psyneuen.2005.02.010. [DOI] [PubMed] [Google Scholar]
- 43.Bryant FB, Smart CM, King SP. Using the past to enhance the present: Boosting happiness through positive reminiscence. J. Happiness Stud. 2005;6:227–260. [Google Scholar]
- 44.Mazzucchelli TG, Kane RT, Rees CS. Behavioral activation interventions for well-being: A meta-analysis. J. Posit. Psychol. 2010;5:105–121. doi: 10.1080/17439760903569154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Ramirez S, et al. Activating positive memory engrams suppresses depression-like behaviour. Nature. 2015;522:335–339. doi: 10.1038/nature14514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Steptoe A, Wardle J, Marmot M. Positive affect and health-related neuroendocrine, cardiovascular, and inflammatory processes. Proc. Natl. Acad. Sci. U. S. A. 2005;102:6508–6512. doi: 10.1073/pnas.0409174102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Heller AS, et al. Sustained striatal activity predicts eudaimonic well-being and cortisol output. Psychol. Sci. 2013;24:2191–2200. doi: 10.1177/0956797613490744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.McEwen BS. Physiology and neurobiology of stress and adaptation: Central role of the brain. Physiol. Rev. 2007;87:873–904. doi: 10.1152/physrev.00041.2006. [DOI] [PubMed] [Google Scholar]
- 49.Sheline YI. Neuroimaging studies of mood disorder effects on the brain. Biol. Psychiatry. 2003;54:338–352. doi: 10.1016/s0006-3223(03)00347-0. [DOI] [PubMed] [Google Scholar]
- 50.Quoidbach J, Wood AM, Hansenne M. Back to the future: The effect of daily practice of mental time travel into the future on happiness and anxiety. J. Posit. Psychol. 2009;4:349–355. [Google Scholar]
- 51.Fardo F, Allen M, Jegindø E-ME, Angrilli A, Roepstorff A. Neurocognitive evidence for mental imagery-driven hypoalgesic and hyperalgesic pain regulation. Neuroimage. 2015;120:350–361. doi: 10.1016/j.neuroimage.2015.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Diamond DM, Bennett MC, Fleshner M, Rose GM. Inverted-U relationship between the level of peripheral corticosterone and the magnitude of hippocampal primed burst potentiation. Hippocampus. 1992;2:421–430. doi: 10.1002/hipo.450020409. [DOI] [PubMed] [Google Scholar]
- 53.Joels M. Corticosteroid effects in the brain: U-shape it. Trends Pharmacol. Sci. 2006;27:244–250. doi: 10.1016/j.tips.2006.03.007. [DOI] [PubMed] [Google Scholar]
- 54.Lupien SJ, et al. The modulatory effects of corticosteroids on cognition: studies in young human populations. Psychoneuroendocrinology. 2002;27:401–416. doi: 10.1016/s0306-4530(01)00061-0. [DOI] [PubMed] [Google Scholar]
- 55.Lupien SJ, et al. Acute Modulation of Aged Human Memory by Pharmacological Manipulation of Glucocorticoids. J. Clin. Endocrinol. Metab. 2002;87:3798–3807. doi: 10.1210/jcem.87.8.8760. [DOI] [PubMed] [Google Scholar]
- 56.Okuda S, Roozendaal B, McGaugh JL. Glucocorticoid effects on object recognition memory require training-associated emotional arousal. Proc. Natl. Acad. Sci. U. S. A. 2004;101:853–8. doi: 10.1073/pnas.0307803100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Leuner B, Caponiti JM, Gould E. Oxytocin stimulates adult neurogenesis even under conditions of stress and elevated glucocorticoids. Hippocampus. 2012;22:861–868. doi: 10.1002/hipo.20947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Lupien SJ, Maheu F, Tu M, Fiocco A, Schramek TE. The effects of stress and stress hormones on human cognition: Implications for the field of brain and cognition. Brain Cogn. 2007;65:209–237. doi: 10.1016/j.bandc.2007.02.007. [DOI] [PubMed] [Google Scholar]
- 59.Troy AS, Wilhelm FH, Shallcross AJ, Mauss IB. Seeing the silver lining: Cognitive reappraisal ability moderates the relationship between stress and depressive symptoms. Emotion. 2010;10:783–795. doi: 10.1037/a0020262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Connor KM, Davidson JRT. Development of a new resilience scale: The Connor-Davidson Resilience Scale (CD-RISC) Depress. Anxiety. 2003;18:76–82. doi: 10.1002/da.10113. [DOI] [PubMed] [Google Scholar]
- 61.Beck A, Ward C, Mendelson M, Mock J, Erbaugh J. An inventory for measuring depression. Arch Gen Psychiatry. 1961;4:561–571. doi: 10.1001/archpsyc.1961.01710120031004. [DOI] [PubMed] [Google Scholar]
- 62.Delgado MR, Nystrom LE, Fissell C, Noll DC, Fiez JA. Tracking the hemodynamic responses to reward and punishment in the striatum. J. Neurophysiol. 2000;84:3072–7. doi: 10.1152/jn.2000.84.6.3072. [DOI] [PubMed] [Google Scholar]
- 63.Talairach J, Tournoux P. Co-Planar Stereotaxic Atlas of the Human Brain. Thieme Medical Publishers; 1988. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.