

Abstract
ATP-gated ionotropic receptors (P2X receptors) are distributed widely in the nervous system. For example, a hetero-oligomeric receptor containing both P2X2 and P2X3 subunits is involved in primary afferent sensation. Each subunit has two membrane-spanning domains. We have used disulfide bond formation between engineered cysteines to demonstrate close proximity between the outer ends of the first transmembrane domain of one subunit and the second transmembrane domain of another. After expression in HEK 293 cells of such modified P2X2 or P2X4 subunits, the disulfide bond formation is evident because an ATP-evoked channel opening requires previous reduction with dithiothreitol. In the hetero-oligomeric P2X2/3 receptor the coexpression of doubly substituted subunits with wild-type partners allows us to deduce that the hetero-oligomeric channel contains adjacent P2X3 subunits but does not contain adjacent P2X2 subunits. The results suggest a “head-to-tail” subunit arrangement in the quaternary structure of P2X receptors and show that a trimeric P2X2/3 receptor would have the composition P2X2(P2X3)2.
Keywords: P2X receptors, ATP-gated channels, subunit stoichiometry, disulfide bonds, ligand-gated channel, subunit order
Introduction
P2X receptors are membrane ion channels gated by extracellular ATP. They are involved in synaptic transmission in the peripheral and central nervous systems (Norenberg and Illes, 2000; Khakh, 2001; North, 2002) and play a role in initiating certain primary afferent signals (Bleehen and Keele, 1977; Ding et al., 2000; Hamilton et al., 2002). The receptors are oligomers, probably trimers (Bean, 1990; Nicke et al., 1998; Ding and Sachs, 1999; Stoop et al., 1999), and there are seven subunits in vertebrate species (P2X1-P2X7). The P2X2 and P2X4 subunits are distributed widely in nervous tissue. A particular role for the P2X3 subunit in the initiation of some forms of pain perception has been indicated recently by experiments with receptor antagonists (Honore et al., 2002a; Jarvis et al., 2002), antisense oligonucleotides (Honore et al., 2002b), and gene knock-outs (Cockayne et al., 2000; Souslova et al., 2000).
The response to ATP of some sensory neurons is not well reproduced by heterologous expression of any single P2X subunit, including P2X3. This led to the proposal (Lewis et al., 1995) that a hetero-oligomeric receptor containing both P2X2 and P2X3 subunits was expressed by these cells. This notion subsequently has found widespread support from biochemical (Radford et al., 1997) and functional approaches [trigeminal ganglion (Cook et al., 1997); nodose ganglion (Thomas et al., 1998); dorsal root ganglia (Burgard et al., 1999; Grubb and Evans, 1999; Ueno et al., 1999; Dunn et al., 2000; Lalo et al., 2001; Zhong et al., 2001; Petruska et al., 2002)]. Other examples of hetero-oligomeric P2X receptors also have been reported (for review, see North, 2002).
P2X receptor subunits are considered to have intracellular N and C termini and two membrane-spanning domains (TMs). The first TM is near the N terminus (residues 30-50 in the P2X2 subunit), and the second TM begins close to residue 330. Most of the protein is formed by the intervening ectodomain; this has five conserved intrinsic disulfide bonds (Clyne et al., 2002; Ennion and Evans, 2002) as well as charged residues close to TM1 and TM2 that play a role in ATP binding (Ennion et al., 2000; Jiang et al., 2000). There is evidence that TM2 contributes some residues to the ion permeation pathway (Rassendren et al., 1997; Egan et al., 1998), and both TMs are known to be major determinants of desensitization in P2X receptors (Werner et al., 1996); however, there is no information regarding the relative arrangement of the TMs within a subunit, and the quaternary organization of subunits within a receptor is unknown.
We investigated these questions by introducing cysteine residues that can form intersubunit disulfide bonds; when channels were expressed in human embryonic kidney (HEK) 293 cells, the reducing agents increased the ATP-evoked currents. From the pattern of the responsive substitutions we deduce that the homo-oligomeric P2X2 or P2X4 receptors have an ordered “head-to-tail” orientation of subunits (i.e., TM1 of one subunit is adjacent to TM2 of the next) and that the P2X2/3 hetero-oligomeric channel contains adjacent P2X3 subunits, but not adjacent P2X2 subunits.
Materials and Methods
Molecular and cell biology. The single and double cysteine mutant rat P2X2 receptors were made in our previous studies (Rassendren et al., 1997; Jiang et al., 2001; Spelta et al., 2003). The single and double cysteine mutant P2X3 receptors were made with the same protocols as for the P2X2 mutant receptors. The wild-type and mutant P2X2 and P2X3 subunits were transiently expressed in HEK 293 cells with Lipofectamine 2000 (Invitrogen, San Diego, CA) as described previously (Spelta et al., 2003). Subunits tagged at their C termini with the EMYPME epitope were used for Western blotting on SDS-PAGE as described (Kim et al., 2001). Samples that were applied were not denatured.
Electrophysiological recording. Whole-cell recordings were made at 20-22°C at 24-72 hr after transfection (Jiang et al., 2001). The holding potential was -60 mV. The extracellular solution contained (in mm): 147 NaCl, 2 KCl, 2 CaCl2, 1 MgCl2, 10 HEPES, and 13 glucose, and the intracellular (pipette) solution comprised (in mm): 147 NaCl or NaF, 10 HEPES, and 10 EGTA. All solutions were maintained at pH 7.3 and 300-315 mOsm/l. Chemicals were purchased from Sigma (Poole, UK). Agonists and dithiothreitol were applied by using an RSC 200 system (Biological Science Instruments, Grenoble, France).
Isolation of current through P2X2/3hetero-oligomeric channels. When recordings are made from cells that have been transfected with both P2X2 and P2X3 subunit cDNAs, there is a response to ATP and αβmeATP that is not observed in cells transfected with either subunit alone [the current elicited by αβmeATP is sustained over several seconds (Lewis et al., 1995)]. However, the interpretation of such coexpression experiments is complicated because hetero-oligomeric channels are not formed exclusively; there is evidence for homo-oligomeric channels as well (Thomas et al., 1998; Virginio et al., 1998). We determined the appropriate ratio of plasmids for cotransfection experiments by testing a range of ratios and subsequently measuring the expression by Western blotting (Kim et al., 2001) and by recording membrane currents. The P2X2/3 receptor was activated by 10 μm αβmeATP, which does not activate the wild-type P2X2 receptor (Spelta et al., 2002) or the P2X2 mutants that have been used here (data not shown). We consider that homo-oligomeric P2X3 receptors did not contribute substantive current in cotransfection experiments because we did not observe any fast-desensitizing component to the current with 10 μm αβmeATP (see Fig. 1).
Figure 1.
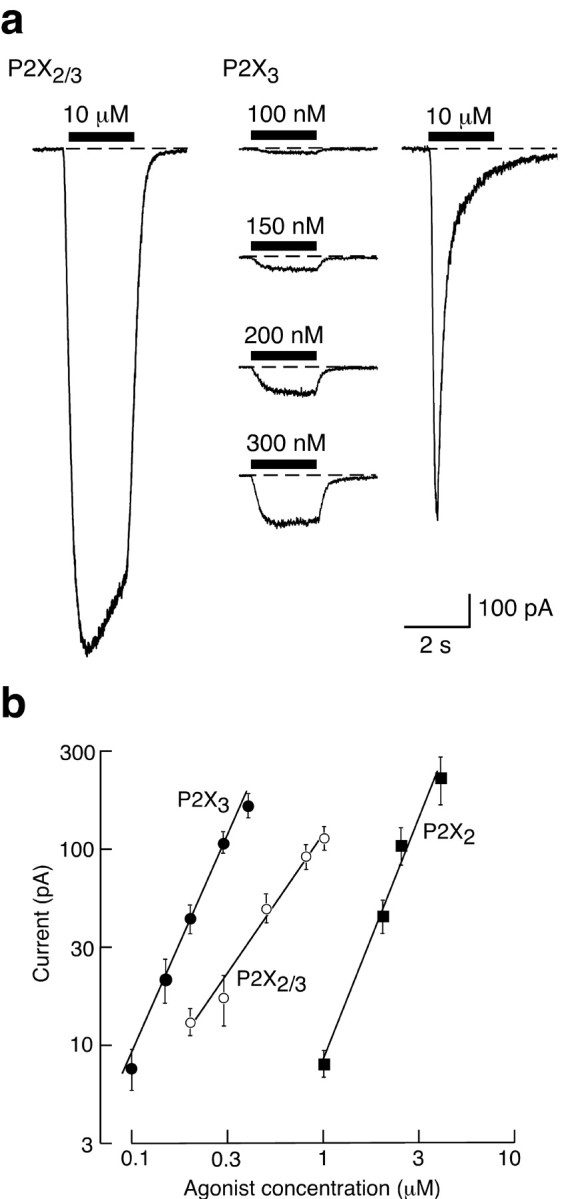
Distinct Hill slopes for activation of homo-oligomeric (P2X2 and P2X3) and hetero-oligomeric (P2X2/3) receptors. a, Left, Representative currents evoked by αβmeATP (10 μm) in HEK 293 cells transfected with P2X2 and P2X3 subunits (1:5 ratio; see Materials and Methods). This is the concentration used in subsequent experiments on the cysteine-substituted receptors. Right, Typical currents evoked by αβmeATP in HEK 293 cells transfected with P2X3 subunits. At 10 μm, αβmeATP evokes a rapidly rising and fast-desensitizing current that was not observed in the doubly transfected cells. At submicromolar concentrations the currents are sustained; such currents were used to estimate Hill coefficients. b, Currents were recorded from single cells held at -60 mV in response to increasing concentrations of agonists as indicated. ATP was used for P2X2 (filled squares) and αβmeATP for P2X2/3 (open circles) and P2X3 receptors (filled circles). The currents are presented as mean ± SEM (n = 4-6 cells for each point). The slopes of these averaged lines are P2X3 2.28, P2X2/3 1.46, and P2X2 2.52.
Data analysis. We applied four or five low agonist concentrations ([A]) noncumulatively to elicit small membrane currents (I) (typically 2-10% but always <20% of the peak current). When the current I is much less than its maximal value (Imax) and [A] is much less than that causing one-half of the maximal effect (K), then the Hill equation:
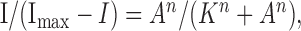 |
reduces to:
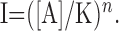 |
For each cell we estimated the Hill coefficient (n) by fitting a straight line by least squares to log(I) = n log[A]; we present the mean value of these estimates with the SEM. For illustration (see Fig. 1) we also averaged the value of the current in all cells that were tested with a given concentration. Other results are shown as mean ± SEM; tests for statistical significance were performed by using Student's unpaired t test or ANOVA.
Results
Hill slopes differ for homo-oligomeric and hetero-oligomeric channels
We used very low concentrations of agonists to estimate the initial slope of the concentration-response curve, because this estimate is less likely to be influenced by cooperative interactions among subunits and may indicate the number of ligand-binding sites (Fig. 1a). Figure 1b shows that this was clearly less for the hetero-oligomeric P2X2/3 than for the homo-oligomeric P2X2 and P2X3 receptors. The slopes were 2.7 ± 0.10 (n = 6), 2.5 ± 0.14 (n = 5), and 1.7 ± 0.14 (n = 5), respectively, for individual cells expressing P2X2 [agonist ATP; P2X2 receptors are not activated by αβmeATP at the concentrations used in this study (Evans et al., 1995; North, 2002; Spelta et al., 2002)], P2X3 (agonist αβmeATP), and P2X2/3 (agonist αβmeATP) receptors. The values for P2X2 and P2X3 homomers are not different, but both of these are different from the value for the P2X2/3 hetero-oligomer (ANOVA, p < 0.001). We cannot exclude the possibility that currents in cells expressing both P2X2 and P2X3 subunits pass through both homomeric P2X3 receptors and heteromeric P2X2/3 receptors. Nonetheless, the difference between the homo-oligomeric and hetero-oligomeric forms is striking and suggests that the hetero-oligomeric P2X2/3 receptor presents fewer αβmeATP binding sites than the homo-oligomeric P2X3 receptor.
Identification of an intersubunit disulfide bond
Cysteine-scanning mutagenesis in and around the two TMs of the P2X2 receptor (Rassendren et al., 1997; Jiang et al., 2001; Spelta et al., 2003) has identified two positions that, when both are substituted by cysteine, could form a disulfide bond. After expression in HEK 293 cells the channels had very small responses to ATP, but this increased more than fivefold after dithiothreitol (DTT) or bis(2-mercaptoethyl)sulfone; the currents declined again when the reducing agent was removed (Jiang et al., 2001), suggesting that the two thiols spontaneously reoxidized. These two residues (Val48 and Ile328) are situated near the extracellular ends of TM1 and TM2, respectively (Newbolt et al., 1998). None of 10 further pairs that were tested appeared to form similar disulfides (Spelta et al., 2003).
Our initial interpretation of these results was that a disulfide bond formed between cysteines provided by the two TMs of the same subunit (i.e., intrasubunit), but it is also possible that the disulfide forms between different P2X2 subunits (i.e., intersubunit), implying that gating involves movements of the subunits relative to each other. We distinguished these possibilities by solubilizing the epitope-tagged P2X2 receptor after expression in HEK 293 cells. The wild-type subunits, and subunits containing either V48C or I328C substitutions alone, migrated on SDS-PAGE predominantly at the position expected for the monomeric subunit (∼62 kDa; arrowhead in Fig. 2a). A smaller amount of higher molecular mass form also was observed. In contrast, no monomeric form was observed for the subunit containing cysteines at both positions (P2X2[V48C/I328C]). The higher molecular mass protein that was observed presumably represents a disulfided dimer, because DTT reduced this to a monomeric size (Fig. 2a). DTT had no effect on the wild-type subunit or subunits with single cysteine substitutions (Fig. 2a), suggesting that the disulfide bond is predominantly intersubunit rather than intrasubunit. Moreover, in parallel experiments with P2X2[V48C/T336C], P2X2[Q37C/L338C], and P2X2[F44C/L338C] (Jiang et al., 2001; Spelta et al., 2003) a band corresponding to the monomer (60-70 kDa) was observed clearly both in the absence or the presence of DTT (data not shown). We previously reported (Jiang et al., 2001) that ATP-evoked currents were unaffected by DTT in the case of P2X2[V48C] and P2X2[I328C] expressed alone, so these experiments all indicate that in the intact channel the disulfide forms between the TM1 of one subunit and the TM2 of another (Fig. 2b).
Figure 2.

Intersubunit disulfide bond formation between V48C and I328C in the homo-oligomeric P2X2 receptor. a, Wild-type P2X2, single cysteine mutants P2X2[V48C] and P2X2[I328C], and double mutant P2X2[V48C/I328C] subunits (each carrying a C-terminal EE epitope) were transiently expressed in HEK 293 cells. Cells were lysed in buffers with or without dithiothreitol as indicated. Protein samples were separated on SDS-PAGE gels and detected by Western blotting via an anti-EE antibody. The band indicated by an arrowhead corresponds to the expected size of the monomeric P2X2 subunit; protein molecular weight markers are indicated on right. These results were observed in at least three independent experiments for each receptor. b, Schematic illustrations of the disulfide formation in homo-oligomeric P2X2 receptors, assuming that the channel is a trimer. TM1 and TM2 of each subunit are indicated. Arrows indicate the positions of disulfide formation.
We have found similar results for the P2X4 receptor. ATP evoked only very small currents in HEK 293 cells expressing the double cysteine-substituted P2X4 receptor (P2X4[V47C/I333C]). These currents (59 ± 16 pA; n = 7) were increased 12-fold after a 4 min application of DTT (10 mm; to 721 ± 175 pA; n = 7). However, receptors with either cysteine substituted singly gave currents that were not different from those in cells expressing wild-type P2X4 receptors (1589 ± 812 pA; n = 4), and dithiothreitol had no effect (P2X4[V47C]: 1989 ± 457 pA, n = 4; P2X4[I333C]: 2053 ± 427 pA, n = 3).
Head-to-tail disulfide formation between TMs in a hetero-oligomeric P2X2/3 receptor
In experiments on coexpression of P2X2 and P2X3 subunits we first determined the appropriate plasmid ratio for transfection that would produce approximately equal membrane expression. Both densitometry of membrane protein (Fig. 3a) and functional experiments comparing ATP and αβmeATP (Fig. 3b) indicated that the appropriate ratio was 1:5 or 1:10; we used a ratio of 1:5 in subsequent experiments.
Figure 3.
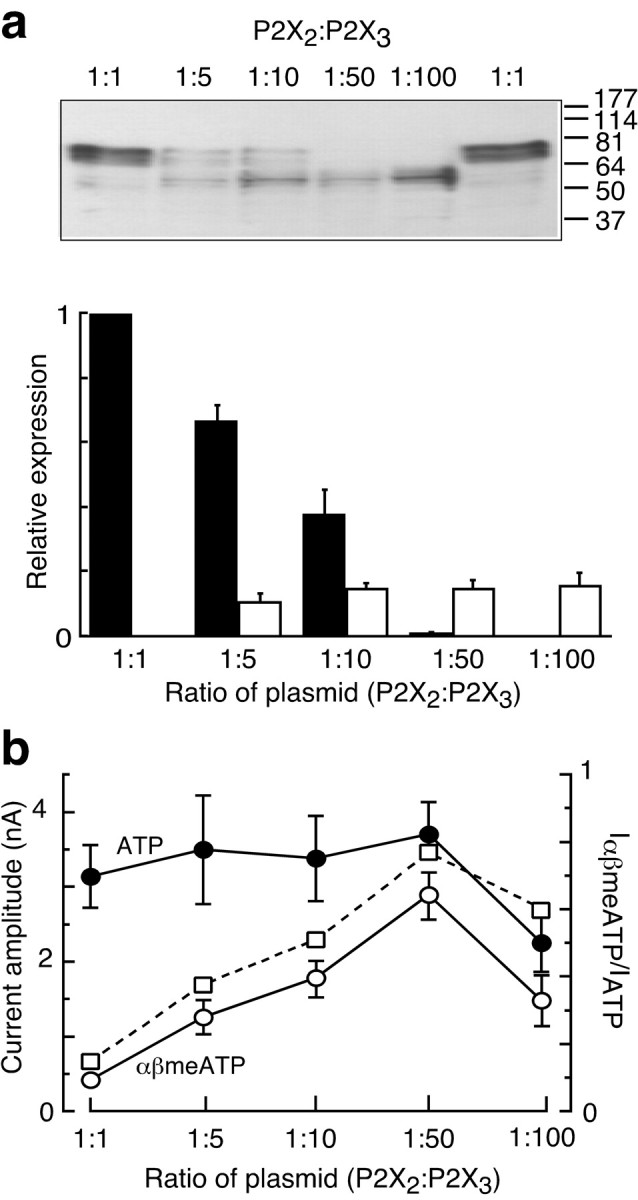
Coexpression of P2X2 and P2X3 subunits. a, Western blots on SDS-PAGE gel show the relative expression of the two subunits expressed in HEK 293 cells, each detected with the same C-terminal EE epitope antibody. The bottom panel shows the quantitation for such experiments from x-ray film densitometric analysis (n = 4). b, The currents elicited by ATP and αβmeATP (left axis) and their ratio (right axis) are shown in HEK 293 cells transfected with various proportions of P2X2 and P2X3 plasmids (n = 4-7). In all experiments 1 μg of P2X3 plasmid was used, with 200, 100, 20, or 10 ng of P2X2 plasmid; error bars indicate SEM.
Figure 4 shows that disulfides also can form between cysteines at the extracellular end of TM1 in the P2X2 subunit (P2X2[V48C]) and the outer end of TM2 in the P2X3 subunit (P2X3[I319C]). Coexpression of these two forms resulted in currents evoked by αβmeATP (10 μm; 191 ± 32 pA; n = 9) that were much smaller than those seen with the wild-type hetero-oligomer, and these currents were increased threefold by dithiothreitol (Fig. 4b). The increase by dithiothreitol reversed on washout and could be repeated with a second application (Fig. 4b). A clear effect of dithiothreitol also was observed with coexpression of the two subunits with cysteines at the outer end of TM2 of the P2X2 subunit (P2X2[I328C]) and the outer end of TM1 of the P2X3 subunit (P2X3[V42C]). In this case the initial current was 432 ± 122 pA (n = 3), and the increase in dithiothreitol was to 145 ± 15% of this control value. In contrast, there was no effect of dithiothreitol on the currents evoked by αβmeATP in cells expressing subunits with cysteines in both TM1s (i.e., of P2X2 and P2X3) or both TM2s or in cells coexpressing a wild-type subunit with a single cysteine-substituted subunit (Fig. 4). The effects of dithiothreitol on the nine pairs of subunits that were tested are summarized in Figure 5a, and the results are illustrated schematically in Figure 5b. They indicate that disulfide bridges can form only between the first TM of one subunit and the second TM domain of another. We cannot exclude the possibility that disulfide bonds can form with some of the other combinations that were tested, but we can say that, if they do, there is no effect on binding or gating of the channel at the macroscopic level.
Figure 4.

Formation of a disulfide bond between P2X2[V48C] and P2X3[I319C] in the hetero-oligomeric P2X2/3 receptor. a, Representative current recordings from cells expressing wild-type P2X2 and wild-type P2X3 (left top), P2X2[V48C] and wild-type P2X3 (left middle), wild-type P2X2 and P2X3[I319C] (left bottom), and P2X2[V48C] and P2X3[I319C] (right). Cells were voltage-clamped at -60 mV, and currents were evoked by αβmeATP (10 μm, 2 sec) at 2 min intervals. Cells were perfused with dithiothreitol (10 mm) at the times indicated. b, Summary of time course of effects by dithiothreitol as described in a. Currents from each cell were normalized to that measured at time 0 (n = 3-9 for each combination). Dithiothreitol had no effect in the case of P2X2 and P2X3 (open squares), P2X2[V48C] and P2X3 (filled triangles), and P2X2 and P2X3[I319C] (filled diamonds); error bars indicate SEM.
Figure 5.

a, Summary of effects of DTT (10 mm, 12 min) on αβmeATP-evoked currents recorded from cells coexpressing different combinations of wild-type or single mutant P2X2 and P2X3 receptors (n = 3-9 cells for each combination). Filled bars represent forms with cysteines in both subunits. Actual currents in those seven combinations in which DTT had no effect included the following (pA ± SEM for the number of cells in parentheses): P2X2[V48C] with P2X3[V42C], 1244 ± 358 (4); P2X2[I328C] with P2X3[I319C], 404 ± 61 (4); P2X2[V48C] with wild-type P2X3, 471 ± 122 (4); P2X2[I328C] with wild-type P2X3, 840 ± 215 (4); wild-type P2X2 with P2X3[V42C], 1399 ± 468 (3); wild-type P2X2 with P2X3[I319C], 692 ± 276 (4); and wild-type P2X2 with wild-type P2X3, 924 ± 258 (4). b, Schematic representation of subunit arrangements for forms with cysteines in both subunits, assuming that the channel is a trimer. Left, Channels containing one P2X3 subunit (black). Right, Channels containing one P2X2 subunit (gray). TM1 and TM2 of each subunit are indicated; cysteines that have been introduced are depicted by small filled circles. The arrows indicate the disulfides that have formed, based on an effect of dithiothreitol on the expressed currents.
Hetero-oligomeric channels have adjacent P2X3subunits, but not adjacent P2X2subunits The specificity of the “crossed” disulfide formation with respect to TM1 and TM2, together with our observation that disulfides could form in the double cysteine homo-oligomeric P2X2 receptor (Jiang et al., 2001) (Fig. 2b) and P2X4 receptor, led us to hypothesize that disulfide bonds would form between two adjacent copies of the same subunit within a hetero-oligomeric channel. We coexpressed wild-type P2X3 subunits with the double cysteine P2X2 subunit (P2X2[V48C/I328C]). The currents (772 ± 132 pA; n = 14 with 10 μm αβmeATP) were not different from those of wild-type P2X2/3 hetero-oligomeric channels, and DTT had no effect (Fig. 6a). Thus although the double cysteine P2X2 subunits can form DTT-sensitive disulfide bonds readily in the homo-oligomeric receptor (Jiang et al., 2001), they are unable to do so in the hetero-oligomeric complex. This implies either that there is only a single P2X2 subunit or that multiple P2X2 subunits are nonadjacent.
Figure 6.

Subunit arrangement of hetero-oligomeric P2X2/3 receptor. Shown are representative current recordings from cells coexpressing P2X2[V48C/I328C] with wild-type P2X3 subunits (a) or wild-type P2X2 with P2X3[V42C/I319C] subunits (b). Currents were evoked by αβmeATP (10 μm, 2 sec), and DTT (10 mm) was applied as indicated. c, Summary of time course of effect by DTT as described in a and b; n = 6 cells for mutant P2X2[V48C/I328C] and wild-type P2X3; n = 9 cells for wild-type P2X2 and P2X3[V42C/I319C]. d, Schematic representation of subunit arrangements in a hetero-oligomeric P2X2/3 receptors, assuming the channel is a trimer. Left, Channels containing one P2X3 subunit (black). Right, Channels containing one P2X2 subunit (gray). The arrow indicates the disulfide that has formed, based on an effect of DTT on the expressed currents. TM1 and TM2 of each subunit are indicated; cysteines that have been introduced are depicted by small filled circles.
In marked contrast was the result of the converse experiment in which we coexpressed the wild-type P2X2 subunit with the P2X3 subunit containing two cysteines. The currents evoked by αβmeATP were only ∼10% of the control amplitude (123 ± 26 pA; n = 11), and DTT evoked a threefold increase (Fig. 6b). The increase was sustained, reversed when the DTT was washed out, and was reproducible (Fig. 6). This result indicates that P2X3 subunits are sufficiently close in the hetero-oligomeric complex for disulfide formation and that this disulfide impedes channel gating. In other words, the channel contains at least two P2X3 subunits, and these have a head-to-tail arrangement (i.e., TM1 of one P2X3 subunit is adjacent to TM2 of another).
Discussion
Estimates of the number of subunits in a P2X receptor have been provided by biochemical approaches. Nicke et al. (1998) concluded that a trimer was a fundamental constituent form of the P2X1 (and P2X3) channels heterologously expressed in Xenopus oocytes on the basis of both cross-linking and nondenaturing gel electrophoresis. Functional approaches have provided estimates of the number of agonist binding sites. In his recordings from dorsal root ganglion cells, Bean (1990) examined the effects of different ATP concentrations and suggested that three molecules of ATP might be required to activate the channel. P2X2 receptors expressed in oocytes exhibit single channel kinetics that are best fit by the assumption of three ATP molecules binding (Ding and Sachs, 1999), and experiments with concatenated P2X2 subunit cDNAs carrying functional reporter mutations are also consistent with a trimeric channel (Stoop et al., 1999). The simplest interpretation of our present results with low agonist concentrations is that the homo-oligomeric channel has three identical, independent binding sites. This would give an initial slope of three for the log-log plot, which is close to what we observed for both the homo-oligomeric P2X2 and P2X3 receptors (Fig. 1). The value for the slope that was observed (1.7) for the hetero-oligomeric P2X2/3 receptor must be interpreted in the context that at least two sets of channels might be present (homomeric P2X3 and heteromeric P2X2/3); in this case the slope might represent a composite value, but any heteromeric channel or channels would have fewer than three binding sites for αβmeATP.
The P2X2 subunit containing two cysteine substitutions (P2X2[V48C/I328C]) behaved very differently on SDS-PAGE from the subunit containing one or neither (Fig. 2). Along with the striking effect of DTT, we interpret this to indicate that these cysteines can participate in intersubunit disulfide bond formation. The proximity of these two cysteines in the functioning channel is indicated by the finding that DTT causes a fivefold increase in the ATP-evoked currents in cells expressing P2X2[V48C/I328C] subunits but has no effect on channels formed from P2X2[V48C], P2X2[I328C], or wild-type P2X2 subunits (Jiang et al., 2001). This indicates that in the functioning channels these residues at the outer ends of TM1 and TM2 are close to each other; the distance from Cα to Cα calculated from bond lengths is 0.84 nm. The finding that no disulfides were observed either biochemically (Fig. 2) or functionally (Jiang et al., 2001) with P2X2[V48C] or P2X2[I328C] further suggests that the subunits must be arranged in a head-to-tail manner in which contacts are made between the TM1 segment of one subunit and the TM2 segment of another. The findings on the P2X4 subunit suggest that the principle of this organization can be generalized.
These observations set the stage for our experiments on the P2X2/3 hetero-oligomer. Our functional expression showed that DTT increased the membrane current so long as the cysteines were introduced into TM1 of P2X2 and TM2 of P2X3 or vice versa. The effect was smaller in the latter case (P2X2[I328C] + P2X3[V42C]), and this might indicate that disulfide formation was incomplete, occurring in only a fraction of channels. In contrast, we found no evidence for interaction between P2X2 and P2X3 subunits when each had cysteines introduced at the outer end of TM1 (P2X2[V48C] + P2X3[V42C]); current amplitudes were normal, and DTT had no effect. When both outer ends of TM2 were substituted (P2X2[I328C] + P2X3[I319C]), there was also no effect of DTT, although initial αβmeATP currents were small. Each of these substitutions was well tolerated when the subunit was expressed singly (normal current amplitudes; data not shown) (see also Rassendren et al., 1997). These results are consistent with a head-to-tail arrangement of P2X2 and P2X3 subunits in the channel (Fig. 5b), as proposed above for the homo-oligomeric P2X2 receptor. The difference in the effectiveness of DTT between P2X2[V48C] + P2X3[I319C] (300% increase) and P2X2[I328C] + P2X3[V42C] (50% increase) is not unexpected, given that the nearby amino acids in these regions of the two subunits are not identical. The positions of the two reacting cysteines at the outer ends of the transmembrane domains are compatible with earlier studies with immunoprecipitation of chimeric P2X receptor subunits, which indicated that these domains were required for hetero-oligomer formation (Torres et al., 1999b). Such coimmunoprecipitation experiments have indicated that various other pairs of P2X subunits can associate with each other (Torres et al., 1999a), and it will be important to determine whether our approach with cysteine substitutions at the ends of TM1 and TM2 can be generalized to other pairs of different subunits.
The coexpression of P2X2 and P2X3 subunits each carrying a single cysteine substitution cannot distinguish between trimers containing one P2X2 subunit and trimers containing two (Fig. 5b). We therefore introduced both cysteines into the same subunit to test whether it was present in the channel adjacent to another copy of itself. We found that P2X2 subunits containing both cysteines (P2X2[V48C/I328C]) cannot make intersubunit disulfides in the P2X2/3 hetero-oligomer, although this readily occurs in the homo-oligomeric channel (Jiang et al., 2001). Thus coexpression of P2X2[V48C/I328C] with wild-type P2X3 subunits gave essentially normal P2X2/3 currents, and these were unaffected by DTT (Fig. 6a). This experiment indicates that the hetero-oligomeric channel does not contain adjacent P2X2 subunits. It also renders unlikely the alternative interpretation that the disulfide formation is interchannel, as distinct from intersubunit within the channel oligomer.
Coexpression of the double cysteine-substituted P2X3 subunit with wild-type P2X2 subunits (P2X3[V42C/I319C] + P2X2) provided oligomeric channel currents that were much smaller than control currents, and these were augmented greatly by DTT (Fig. 6b). This experimental result is explained most readily by the assumption that the channel activated by αβmeATP contains adjacent head-to-tail P2X3 subunits. In other words, both of these two independent experiments indicate that, if the channel is a trimer, then its composition must be P2X2(P2X3)2. If the channel is a tetramer with four subunits arranged head-to-tail around a central pore, then we can conclude that it does not contain adjacent P2X2 subunits, but it does contain adjacent P2X3 subunits; in other words, only the asymmetrical tetramer (P2X2-P2X3-P2X3-P2X3) is allowed. Such an asymmetric arrangement has been proposed recently for the cyclic nucleotide-gated channels (Zheng et al., 2002; Zhong et al., 2002). A hexameric channel would be consistent with our results, but the requirement for adjacent P2X3 subunits and nonadjacent P2X2 subunits would seem to eliminate the symmetrical hexamer (P2X2-P2X3-P2X2-P2X3-P2X2-P2X3).
It is easiest to interpret the effects of DTT as indicating that movement of one subunit with respect to another is necessary for channel opening. We have shown previously that a cysteine substituted for Val48 becomes more accessible to methanethiosulfonate when the channel is opened, suggesting an outward movement of TM1 (Jiang et al., 2001). We cannot exclude the possibility, however, that the intersubunit disulfide formation primarily impedes ligand binding. State-dependent disulfide formation has been observed in other channels, both for engineered and naturally occurring cysteines. For example, voltage-gated Shaker potassium channels undergo an outward movement of the S4 segment during channel activation (Larsson et al., 1996; Yusaf et al., 1996) (see also Jiang et al., 2003); depolarization can drive disulfide formation in channels in which a cysteine is placed at the outer end of this segment (L361C) (Aziz et al., 2002). Disulfide formation between endogenous cysteines (Cys481) in the C-linker region of subunits of the cyclic nucleotide-gated channel (CNGA1) favors channel opening; these also can form between adjacent, but not opposite, subunits of the tetramer (Rosenbaum and Gordon, 2002). The other channel family that shares with P2X subunits the overall topology of two TMs and a cysteine-rich ectodomain is the epithelial sodium channel/acid-sensing ion channel (ENaC/ASIC) family, some members of which are widely expressed in neurons. The present approach might be applicable also to determine the arrangement of subunits in those channels. On the other hand, it is important not to over-interpret the present data in any structural sense. Our schematics (Figs. 2, 5, 6) depict an arrangement in which three subunits surround a central pore, and it is known that single cysteines introduced at these positions are accessible to water-soluble methane thiosulfonates (Rassendren et al., 1997; Jiang et al., 2001). However, there is no direct evidence that positions these residues on the permeation pathway, and there remains nothing known about the proximity or otherwise of the inner aspects of the membrane-spanning domains (Spelta et al., 2003).
In summary, our results suggest that movement of one P2X subunit relative to another is a key part of gating in both homo- and hetero-oligomeric channels. They provide a means to detect adjacent subunits in the receptor oligomer; in the case of the P2X2/3 receptor the channel activated by αβmeATP has adjacent P2X3 subunits, but not adjacent P2X2 subunits. These findings contribute to our understanding of the molecular operation of P2X receptors and will inform our view of the subunit composition and arrangement of the receptors as they are found in nerve cells, including sensory neurons.
Footnotes
This work was supported by the Wellcome Trust. We thank D. Estoppey and V. Porteous for cell culture.
Correspondence should be addressed to R. A. North, Institute of Molecular Physiology, University of Sheffield, Sheffield S10 2TN, UK. E-mail: r.a.north@shef.ac.uk.
M. Kim's present address: Liver Research Center, Brown University School of Medicine, 55 Claverick Street, Providence, RI 02903.
V. Spelta's present address: Vollum Institute, Oregon Health and Science University, Portland, OR 97201.
X. Bo's present address: Academic Department of Neurosurgery, Medical Science Building, Queen Mary, University of London, Mile End Road, London E1 4NS, UK.
Copyright © 2003 Society for Neuroscience 0270-6474/03/238903-08$15.00/0
References
- Aziz QH, Partridge CJ, Munsey TS, Sivaprasadarao A ( 2002) Depolarization induces intersubunit cross-linking in a S4 cysteine mutant of the Shaker potassium channel. J Biol Chem 277: 42719-42725. [DOI] [PubMed] [Google Scholar]
- Bean BP ( 1990) ATP-activated channels in rat and bullfrog sensory neurons: concentration dependence and kinetics. J Neurosci 10: 1-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bleehen T, Keele CA ( 1977) Observations on the algogenic actions of adenosine compounds on the human blister base preparation. Pain 3: 367-377. [DOI] [PubMed] [Google Scholar]
- Burgard EC, Niforatos W, van Biesen T, Lynch KJ, Touma E, Metzger RE, Kowaluk EA, Jarvis MF ( 1999) P2X receptor-mediated ionic currents in dorsal root ganglion neurons. J Neurophysiol 82: 1590-1598. [DOI] [PubMed] [Google Scholar]
- Clyne JD, Wang LF, Hume RI ( 2002) Mutational analysis of the conserved cysteines of the rat P2X2 purinoceptor. J Neurosci 22: 3873-3880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cockayne DA, Hamilton SG, Zhu QM, Dunn PM, Zhong Y, Novakovic S, Malmberg AB, Cain G, Berson A, Kassotakis L, Hedley L, Lachnit WG, Burnstock G, McMahon SB, Ford AP ( 2000) Urinary bladder hyporeflexia and reduced pain-related behaviour in P2X3-deficient mice. Nature 407: 1011-1015. [DOI] [PubMed] [Google Scholar]
- Cook SP, Vulchanova L, Hargreaves KM, Elde R, McCleskey EW ( 1997) Distinct ATP receptors on pain-sensing and stretch-sensing neurons. Nature 387: 505-508. [DOI] [PubMed] [Google Scholar]
- Ding S, Sachs F ( 1999) Single channel properties of P2X2 purinoceptors. J Gen Physiol 113: 695-720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ding Y, Cesare P, Nikitaki D, Wood JN ( 2000) ATP, P2X receptors and pain pathways. J Auton Nerv Syst 81: 289-294. [DOI] [PubMed] [Google Scholar]
- Dunn PM, Liu M, Zhong Y, King BF, Burnstock G ( 2000) Diinosine pentaphosphate: an antagonist which discriminates between recombinant P2X3 and P2X2/3 receptors and between two P2X receptors in rat sensory neurones. Br J Pharmacol 130: 1378-1384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Egan TM, Haines WR, Voigt MM ( 1998) A domain contributing to the ion channel of ATP-gated P2X2 receptors identified by the substituted cysteine accessibility method. J Neurosci 18: 2350-2359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ennion SJ, Evans RJ ( 2002) Conserved cysteine residues in the extracellular loop of the human P2X1 receptor form disulfide bonds and are involved in receptor trafficking to the cell surface. Mol Pharmacol 61: 303-311. [DOI] [PubMed] [Google Scholar]
- Ennion SJ, Hagan S, Evans RJ ( 2000) The role of positively charged amino acids in ATP recognition by human P2X1 receptors. J Biol Chem 275: 29361-29367. [DOI] [PubMed] [Google Scholar]
- Evans RJ, Lewis C, Buell G, Valera S, North RA, Surprenant A ( 1995) Pharmacological characterization of heterologously expressed ATP-gated cation channels (P2X purinoceptors). Mol Pharmacol 48: 178-183. [PubMed] [Google Scholar]
- Grubb BD, Evans RJ ( 1999) Characterization of cultured dorsal root ganglion neuron P2X receptors. Eur J Neurosci 11: 149-154. [DOI] [PubMed] [Google Scholar]
- Hamilton SG, Warburton J, Bhattacharjee A, Ward J, McMahon SB ( 2002) ATP in human skin elicits a dose-related pain response which is potentiated under conditions of hyperalgesia. Brain 123: 1238-1246. [DOI] [PubMed] [Google Scholar]
- Honore P, Mikusa J, Bianchi B, McDonald H, Cartmell J, Faltynek C, Jarvis MF ( 2002a) TNP-ATP, a potent P2X3 receptor antagonist, blocks acetic acid-induced abdominal constriction in mice: comparison with reference analgesics. Pain 96: 99-105. [DOI] [PubMed] [Google Scholar]
- Honore P, Kage K, Mikusa J, Watt AT, Johnston JF, Wyatt JR, Faltynek CR, Jarvis MF, Lynch K ( 2002b) Analgesic profile of intrathecal P2X3 antisense oligonucleotide treatment in chronic inflammatory and neuropathic pain states in rats. Pain 99: 11-19. [DOI] [PubMed] [Google Scholar]
- Jarvis MF, Burgard EC, McGaraughty S, Honore P, Lynch K, Brennan TJ, Subieta A, Van Biesen T, Cartmell J, Bianchi B, Niforatos W, Kage K, Yu H, Mikusa J, Wismer CT, Zhu CZ, Chu K, Lee CH, Stewart AO, Polakowski J, Cox BF, Kowaluk E, Williams M, Sullivan J, Faltynek C ( 2002) A-317491, a novel potent and selective non-nucleotide antagonist of P2X3 and P2X2/3 receptors, reduces chronic inflammatory and neuropathic pain in the rat. Proc Natl Acad Sci USA 99: 17179-17184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang L-H, Rassendren F, Surprenant A, North RA ( 2000) Identification of amino acid residues contributing to the ATP-binding site of a purinergic P2X receptor. J Biol Chem 275: 34190-34196. [DOI] [PubMed] [Google Scholar]
- Jiang L-H, Rassendren F, Spelta V, Surprenant A, North RA ( 2001) Amino acid residues involved in gating identified in the first membrane-spanning domain of the rat P2X2 receptor. J Biol Chem 276: 14902-14908. [DOI] [PubMed] [Google Scholar]
- Jiang Y, Lee A, Chen J, Ruta V, Cadene M, Chait BT, MacKinnon R ( 2003) X-ray structure of a voltage-dependent K+ channel. Nature 423: 33-41. [DOI] [PubMed] [Google Scholar]
- Khakh BS ( 2001) Molecular physiology of P2X receptors and ATP signaling at synapses. Nat Rev Neurosci 2: 165-174. [DOI] [PubMed] [Google Scholar]
- Kim M, Spelta V, Sim J, North RA, Surprenant A ( 2001) Differential assembly of rat purinergic P2X7 receptor in immune cells of the brain and periphery. J Biol Chem 276: 23262-23267. [DOI] [PubMed] [Google Scholar]
- Lalo UV, Pankratov YV, Arndts D, Krishtal OA ( 2001) Omega-conotoxin GVIA potently inhibits the currents mediated by P2X receptors in rat DRG neurons. Brain Res Bull 54: 507-512. [DOI] [PubMed] [Google Scholar]
- Larsson HP, Baker OS, Dhillon DS, Isacoff EY ( 1996) Transmembrane movement of the Shaker K+ channel S4. Neuron 16: 387-397. [DOI] [PubMed] [Google Scholar]
- Lewis C, Neidhart S, Holy C, North RA, Buell G, Surprenant A ( 1995) Coexpression of P2X2 and P2X3 receptor subunits can account for ATP-gated currents in sensory neurons. Nature 377: 432-435. [DOI] [PubMed] [Google Scholar]
- Newbolt A, Stoop R, Virginio C, Surprenant A, North RA, Buell G, Rassendren F ( 1998) Membrane topology of an ATP-gated ion channel (P2X receptor). J Biol Chem 273: 15177-15182. [DOI] [PubMed] [Google Scholar]
- Nicke A, Baumert HG, Rettinger J, Eichele A, Lambrecht G, Mutschler E, Schmalzing G ( 1998) P2X1 receptors form stable trimers: a novel structural motif of ligand-gated ion channels. EMBO J 17: 3016-3028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Norenberg W, Illes P ( 2000) Neuronal P2X receptors: localisation and functional properties. Naunyn Schmiedebergs Arch Pharmacol 362: 324-339. [DOI] [PubMed] [Google Scholar]
- North RA ( 2002) Molecular physiology of P2X receptors. Physiol Rev 82: 1013-1067. [DOI] [PubMed] [Google Scholar]
- Petruska JC, Napaporn J, Johnson RD, Cooper BY ( 2002) Chemical responsiveness and histochemical phenotype of electrophysiologically classified cells of the adult rat dorsal root ganglion. Neuroscience 115: 15-30. [DOI] [PubMed] [Google Scholar]
- Radford KM, Virginio C, Surprenant A, North RA, Kawashima E ( 1997) Baculovirus expression provides direct evidence for heteromeric assembly of P2X2 and P2X3 receptors. J Neurosci 17: 6529-6533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rassendren F, Buell G, Newbolt A, North RA, Surprenant A ( 1997) Identification of amino acid residues contributing to the pore of a P2X receptor. EMBO J 16: 3446-3454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenbaum T, Gordon SE ( 2002) Dissecting intersubunit contacts in cyclic nucleotide-gated ion channels. Neuron 33: 703-713. [DOI] [PubMed] [Google Scholar]
- Souslova V, Cesare P, Ding Y, Akopian AN, Stanfa L, Suzuki R, Carpenter K, Dickenson A, Boyce S, Hill R, Nebenuis-Oosthuizen D, Smith AJ, Kidd EJ, Wood JN ( 2000) Warm-coding deficits and aberrant inflammatory pain in mice lacking P2X3 receptors. Nature 407: 1015-1017. [DOI] [PubMed] [Google Scholar]
- Spelta V, Jiang LH, Surprenant A, North RA ( 2002) Kinetics of antagonist actions at rat P2X2/3 heteromeric receptors. Br J Pharmacol 135: 1524-1530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spelta V, Jiang LH, Bailey RJ, Surprenant A, North RA ( 2003) Interaction between cysteines introduced into each transmembrane domain of the rat P2X2 receptor. Br J Pharmacol 138: 131-136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stoop R, Thomas S, Rassendren F, Kawashima E, Buell G, Surprenant A, North RA ( 1999) Contribution of individual subunits to the multimeric P2X2 receptor: estimates based on methanethiosulfonate block at T336C. Mol Pharmacol 56: 973-981. [DOI] [PubMed] [Google Scholar]
- Thomas S, Virginio C, North RA, Surprenant A ( 1998) The antagonist trinitrophenyl-ATP reveals coexistence of distinct P2X receptor channels in rat nodose neurones. J Physiol (Lond) 509: 411-417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torres GE, Egan TM, Voigt MM ( 1999a) Hetero-oligomeric assembly of P2X receptor subunits. Specificities exist with regard to possible partners. J Biol Chem 274: 6653-6659. [DOI] [PubMed] [Google Scholar]
- Torres GE, Egan TM, Voigt MM ( 1999b) Identification of a domain involved in ATP-gated ionotropic receptor subunit assembly. J Biol Chem 274: 22359-22365. [DOI] [PubMed] [Google Scholar]
- Ueno S, Tsuda M, Iwanaga T, Inoue K ( 1999) Cell type-specific ATP-activated responses in rat dorsal root ganglion neurons. Br J Pharmacol 126: 429-436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Virginio C, Robertson G, Surprenant A, North RA ( 1998) Trinitrophenyl-substituted nucleotides are potent antagonists selective for P2X1, P2X3, and heteromeric P2X2/3 receptors. Mol Pharmacol 53: 969-973. [PubMed] [Google Scholar]
- Werner P, Seward EP, Buell GN, North RA ( 1996) Domains of P2X receptors involved in desensitization. Proc Natl Acad Sci USA 93: 15485-15490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yusaf SP, Wray D, Sivaprasadarao A ( 1996) Measurement of the movement of the S4 segment during the activation of a voltage-gated potassium channel. Pflügers Arch 433: 91-97. [DOI] [PubMed] [Google Scholar]
- Zheng J, Trudeau MC, Zagotta WN ( 2002) Rod cyclic nucleotide-gated channels have a stoichiometry of three CNGA1 subunits and one CNGB1 subunit. Neuron 36: 891-896. [DOI] [PubMed] [Google Scholar]
- Zhong H, Molday LL, Molday RS, Yau KW ( 2002) The heteromeric cyclic nucleotide-gated channel adopts a 3A:1B stoichiometry. Nature 420: 193-198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhong Y, Dunn PM, Bardini M, Ford AP, Cockayne DA, Burnstock G ( 2001) Changes in P2X receptor responses of sensory neurons from P2X3-deficient mice. Eur J Neurosci 14: 1784-1792. [DOI] [PubMed] [Google Scholar]