

Abstract
Here we characterize a novel neuronal kinase, cyclin-dependent kinase 5 (cdk5)/p35-regulated kinase (cprk). Cprk is a member of a previously undescribed family of kinases that are predicted to contain two N-terminal membrane-spanning domains and a long C terminus, which harbors a dual-specificity serine/threonine/tyrosine kinase domain. Cprk was isolated in a yeast two-hybrid screen using the neuronal cdk5 activator p35 as “bait.” Cprk interacts with p35 in the yeast-two hybrid system, binds to p35 in glutathione S-transferase fusion pull-down assays, and colocalizes with p35 in cultured neurons and transfected cells. In these cells, cprk is present with p35 in the Golgi apparatus. Cprk is expressed in a number of tissues but is enriched in brain and muscle and within the brain is found in a wide range of neuronal populations. Cprk displays catalytic activity in in vitro kinase assays and is itself phosphorylated by cdk5/p35. Cdk5/p35 inhibits cprk activity. Cdk5/p35 may therefore regulate cprk function in the brain.
Keywords: cdk5, p35, tau, neurofilament, Alzheimer's disease, amyotrophic lateral sclerosis
Introduction
Cyclin-dependent kinase 5 (cdk5) is the only characterized cdk that is known to be active in postmitotic neurons. In common with other cdks, cdk5 activity requires binding to an activator subunit, and two mammalian cdk5 activators, p35 and p39, have now been described (Lee et al., 1997b; Dhavan and Tsai, 2001; Grant et al., 2001; Paglini and Cáceres, 2001).
Cdk5 activity is required for a number of functions, including neurite outgrowth and axon patterning, both of which are probably mediated by cdk5-induced alterations to the cytoskeleton (Nikolic et al., 1996; Pigino et al., 1997; Paglini et al., 1998; Connell-Crowley et al., 2000; Humbert et al., 2000). Indeed, cdk5 regulates actin dynamics and also phosphorylates NUDEL, a protein involved in the regulation of cytoplasmic dynein (Nikolic et al., 1998; Niethammer et al., 2000; Sasaki et al., 2000; Smith et al., 2000; Rashid et al., 2001). Probably linked to these functions, cdk5 is associated with β-catenin to regulate cell adhesion in the brain (Kwon et al., 2000; Kesavapany et al., 2001) and controls neuron migration during development (Ohshima et al., 1996; Chae et al., 1997; Gilmore et al., 1998; Ohshima et al., 1999; Gilmore and Herrup, 2001; Ko et al., 2001; Ohshima et al., 2001, 2002; Tanaka et al., 2001; Keshvara et al., 2002). Cdk5 is also involved in synaptic vesicle release, endocytosis, membrane trafficking (Fletcher et al., 1999; Floyd et al., 2001; Paglini et al., 2001; Tomizawa et al., 2002), dopamine signaling (Bibb et al., 1999; Nishi et al., 2002), neuregulin-induced expression of acetylcholine receptor expression at the neuromuscular junction (Fu et al., 2001), and adaptive changes in the brain induced by cocaine (Bibb et al., 2001a). Cdk5 has been linked to learning, memory, and synaptic plasticity, possibly via crosstalk between cdk5 and calcium/calmodulin-dependent protein kinase II (Dhavan et al., 2002; Fischer et al., 2002). Finally, cdk5 phosphorylates neurofilaments (Shetty et al., 1993; Guidato et al., 1996; Sun et al., 1996; Bajaj and Miller, 1997; Sharma et al., 1999), protein phosphatase inhibitor 1 (Agarwal-Mawal and Paudel, 2001; Bibb et al., 2001b), p53 (Zhang et al., 2002), presenilin 1 (Lau et al., 2002), the kinases mitogen-activated protein kinase/extracellular signal-regulated kinase kinase 1 (Sharma et al., 2002) and c-Jun N-terminal kinase 3 (Li et al., 2002), disabled (Keshvara et al., 2002), and the microtubule-associated protein tau (Baumann et al., 1993; Kobayashi et al., 1993; Paudel et al., 1993; Michel et al., 1998; Evans et al., 2000). although this latter function may require deregulation of cdk5/p35 activity (Patrick et al., 1999; Hashiguchi et al., 2002).
Aside from its normal functions, abnormal cdk5/p35 activity has been implicated in several neurodegenerative diseases (Baumann et al., 1993; Paudel, 1997; Bajaj et al., 1998; Patrick et al., 1999; Ahlijanian et al., 2000; Evans et al., 2000; Kusakawa et al., 2000; Lee et al., 2000; Sobue et al., 2000; Alvarez et al., 2001; Kesavapany et al., 2001; Nguyen et al., 2001; Bu et al., 2002; Lau et al., 2002).
There is evidence to suggest that p35 and p39 target the kinase to its substrates (Lee et al., 1997a; Nikolic et al., 1998; Qi et al., 1998; Kesavapany et al., 2001). To identify novel substrates for this cdk5/p35, we used the yeast two-hybrid system to screen for p35 binding partners. Here we report the characterization of a novel kinase, cdk5/p35-regulated kinase (cprk), which binds to p35 and whose activity is regulated by cdk5/p35.
Materials and Methods
Experiments involving the confirmation of yeast two-hybrid results, p35-cprk pull-down assays, localization of cprk by immunostaining, phosphorylation of cprk by cdk5/p35, and cprk autophosphorylation studies were all performed at least three times with similar results.
Yeast two-hybrid system. Yeast two-hybrid experiments were performed essentially as described previously (Guidato et al., 1998; Kesavapany et al., 2001). “Bait” plasmids for p35 and the p10 and p25 fragments of p35 in the bait vector pY2 were as described (Kesavapany et al., 2001). Library screens were performed by transforming yeast Y190 with the full-length p35 bait and a “prey” human brain cDNA library in pGAD10 (Clontech, Cambridge, UK). After selection, vigorously growing clones were subjected to freeze-fracture β-galactosidase assays. pGAD10 plasmids were rescued by transformation into Escherichia coli, and the inserts were sequenced.
Mammalian cell culture and transfection. Primary rat cortical neurons, Chinese hamster ovary (CHO) and COS cells were grown as previously described (Brownlees et al., 2000; Kesavapany et al., 2001). Briefly, cortical neurons were obtained from embryonic day 18 (E18) rat embryos and cultured in Neurobasal medium and B27 supplement (Invitrogen, San Diego, CA) containing 100 IU/ml penicillin, 100 μg/ml streptomycin, and 2 mm glutamine. Cells were cultured for 5 d and under these conditions were almost exclusively neurons as previously described by us and others (Nikolic et al., 1996; Ackerley et al., 2000).
CHO and COS cells were grown respectively in Ham's F-12 and DMEM (Invitrogen), both containing 10% (v/v) fetal bovine serum supplemented with 2 mm glutamine, 100 U/ml penicillin, and 100 μg/ml streptomycin. CHO and COS cells were transfected using LipofectAMINE (Invitrogen) according to the instructions of the manufacturer. For immunofluorescence studies, cells were cultured in 12-well plates (Falcon) on poly-d-lysine-coated glass coverslips. Neurons were cultured for 5 d before analyses.
Mammalian expression vectors for p35 and hemagluttinin-tagged cdk5 were as previously described (Tsai et al., 1994; Guidato et al., 1996). Cprk was expressed by cloning the full-length cDNA (KIAA1079) into pCIneo (Promega, Madison, WI) as an XhoI–NotI fragment.
Antibodies. Mouse and rabbit polyclonal antibodies to cprk were generated by immunization with a glutathione S-transferase (GST)-cprk fusion protein. To create the fusion protein, an EcoRI fragment encompassing residues 391–632 of cprk was cloned into the GST fusion vector pGEX-5x-1. GST-cprk 391–632 was expressed in E. coli DH5α and purified with glutathione-Sepharose beads according to the instructions of the manufacturer (Amersham Biosciences, Little Chalfont, UK).
p35 was detected using rabbit polyclonal antibody C19 (Santa Cruz Biotechnology, Santa Cruz, CA). Mouse monoclonal antibody to β-cop was obtained from Sigma (St. Louis, MO), and rabbit polyclonal antibody to calnexin was obtained from Stressgen. Active cdk5/p35 was immunoprecipitated from cdk5/p35-transfected CHO cells using antibody 12CA5 (Roche Molecular Biochemicals, Indianapolis, IN) to the hemagluttinin epitope tag on cdk5. Anti-phosphoserine, phosphothreonine and phosphotyrosine (4G10) monoclonal antibodies were obtained from Calbiochem (La Jolla, CA), Cell Signaling Technology, and Upstate Biotechnology (Lake Placid, NY), respectively. Guinea pig antibody to the glial glutamate transporter GLAST was obtained from Chemicon (Temecula, CA).
SDS-PAGE and immunoblot analyses. Mouse brains were dissected from E15, E18, postnatal day 0 (P0), P10, P28, and adult (3-month-old) mice and homogenized as a 10% homogenate in ice-cold tissue homogenization buffer containing 125 mm Tris-Cl, pH 6.8, 5 mm EDTA, 5 mm EGTA, 5 μg/ml pepstatin A, 5 μg/ml leupeptin, 2 μg/ml aprotinin, and 1 mm PMSF using a Dounce homogenizer. Samples were prepared by addition of a quarter volume of 10% SDS sample buffer and heating in a boiling water bath for 10 min.
Cultured cells were processed for SDS-PAGE by washing twice with ice-cold PBS, scraping into ice-cold tissue homogenization buffer, addition of SDS PAGE sample buffer, and heating in a boiling water bath as above. Samples were separated on 8–10% (w/v) acrylamide gels and transferred to Protran nitrocellulose membranes (Schleicher & Schuell, Keene, NH) using a Bio-Rad (Hercules, CA) TransBlot system. The blots were probed with primary antibodies, washed in PBS, and incubated with horseradish peroxidase-conjugated anti-mouse or anti-rabbit Igs (Amersham Biosciences). Blots were developed using an enhanced chemiluminescence system (Amersham Biosciences) according to the instructions of the manufacturer.
Dephosphorylated samples were prepared essentially as described previously by treatment with λ-protein phosphatase (New England Biolabs, Beverly, MA) for 60 min at 30°C (Lau et al., 2002). After treatment, they were analyzed by SDS-PAGE by addition of sample buffer as above. Protein concentrations were determined using Bradford and Markwell assays.
Detection of p35-cprk391–632 interaction in GST fusion pull-down assays. To detect interactions between cprk 391–632 and p35, GST and the GST-cprk 391–632 fusion proteins were used as baits in pull-down experiments from nontransfected and p35- and p35/cdk5-transfected CHO cells and primary cortical neurons. Cells were lysed in ice-cold lysis buffer containing 50 mm Tris-HCl, 150 mm NaCl, 1 mm EDTA, 1% Triton X-100, 5 μg/ml leupeptin, 2 μg/ml aprotinin, 5 μg/ml pepstatin, and 0.25 mm PMSF and cleared by centrifugation at 13,000 × g for 15 min. Five hundred micrograms of the cell lysates were then incubated for 4 hr with the different baits at 4°C; the beads were pelleted by centrifugation at 500 × g and washed four times in lysis buffer; and the samples were prepared for SDS-PAGE by addition of SDS-PAGE sample buffer and heating in a boiling water bath for 10 min. After SDS-PAGE and transfer to nitrocellulose membranes, p35 was detected using antibody C19.
Northern analysis. A multiple human tissue Northern blot (Clontech) was probed with the 723 bp fragment of cprk isolated from the yeast two-hybrid library screen and purified as an EcoRI fragment. To demonstrate integrity of mRNAs, the blot was stripped and rehybridized with a β-actin probe supplied with the blot. Hybridizations and high-stringency washings were performed according to the instructions of the manufacturer using ExpressHyb hybridization solution (Clontech). The blot was stripped for rehybridization in 0.5% SDS at 90°C following the instructions of the manufacturer. Probes were labeled using a Prime-It II random primer labeling kit (Stratagene, La Jolla, CA).
Immunocytochemical analyses. Cortical neurons and transfected COS cells grown on coverslips were fixed in 4% (w/v) paraformaldehyde in PBS for 20 min, permeabilized in 0.1% (w/v) Triton X-100 in PBS for 10 min, blocked with 5% (v/v) fetal bovine serum and 0.1% (w/v) Tween 20 in PBS for 1 hr, and then probed with primary antibodies diluted in blocking solution. Primary antibodies were then localized using goat anti-mouse or goat anti-rabbit Igs coupled to Oregon green or Texas Red (Molecular Probes, Eugene, OR), and the samples were mounted in Vectashield (Vector Laboratories, Burlingame, CA).
For mapping cprk distribution in mouse brain, 3-month-old female mice were deeply anesthetized with pentobarbitone and transcardially perfused with PBS followed by a freshly made and filtered solution of 4% paraformaldehyde in PBS, pH 7.4. Brains were subsequently removed and postfixed for 6 hr at 4°C in the same fixative before cryoprotection at 4°C in a solution of 30% sucrose in Tris-buffered saline (TBS; 40 mm Tris and 0.7% NaCl in 10 mm phosphate buffer, pH 7.8) containing 0.05% sodium azide and bisected along the midline before freezing on dry ice and storage at -80°C. Forty micrometer frozen sections were cut coronally or sagittally from either hemisphere, collected in TBS-azide buffer, and stored at 4°C before immunohistochemical staining for cprk, which normally occurred on the same day as cutting. The optimal concentrations of primary and secondary antisera were determined by performing a checkerboard titration on control tissue. The specificity of the cprk immunoreactivity was tested by omission of this primary antiserum or by competition with the cprk antigen, both of which resulted in the complete absence of cprk immunoreactivity (data not shown).
Mapping of cprk distribution was performed as described by us for other brain proteins (Kesavapany et al., 2002). Briefly, every sixth coronal and sagittal section was selected for detection of cprk using the optimized staining protocol. This involved blocking endogenous peroxidase activity with 1% H2O2 in TBS for 15 min and then blocking with 15% normal goat serum (NGS). Sections were then incubated with rabbit cprk antibody in TBS-T (TBS containing 0.3% Triton X-100) and 10% NGS for 16 hr at 4°C, washed in TBS-T, and then incubated with biotinylated goat anti-rabbit Igs (Vector Laboratories) diluted in TBS-T and 10% NGS. After washing in TBS, sections were further incubated for 2 hr in avidin–biotin–peroxidase complex in TBS (Vectastain; Vector Laboratories), washed in TBS, and developed with 0.05% diaminobenzidine tetrahydrochloride and 0.001% H2O2 in TBS. Sections were mounted on gelatin-chrome alum-coated Superfrost slides, air-dried overnight, cleared in xylene, and coverslipped with DPX (BDH Chemicals, Poole, UK). For double-immunofluorescence labeling of sections, rabbit cprk antibody and guinea pig polyclonal antibody to GLAST were applied together and detected using fluorescently conjugated goat anti-rabbit and goat anti-guinea pig Igs essentially as described above for double-immunofluorescent labeling of cultured cells. Images were captured on a Zeiss (Welwyn Garden City, UK) Axioscope 2 MOT using an Axiocam (Zeiss). Brain sections from three mice were analyzed, and the relative intensity of cprk immunoreactivity in different CNS regions was scored in a qualitative manner ranging from +++ (darkly cprk immunoreactive) to -- (absence of cprk immunoreactivity). Similar results were obtained from all three mice, and representative sections have been used for documenting these data (see Fig. 5).
In vitro kinase assays. For analysis of cdk5/p35 phosphorylation of recombinant cprk, active cdk5/p35 was immunoprecipitated from transfected CHO cells using antibody 12CA5 to the hemagluttinin epitope tag on cdk5 as described (Brownlees et al., 2000). One microgram of either GST or GST-cprk 391–632 substrate was then incubated with immunoprecipitated kinase (prepared from 200 μg of precleared lysate) in kinase buffer containing 25 mm HEPES, pH 7.5, 20 mm MgCl2, 20 mm β-glycerophosphate, 20 mmp-nitrophenylphosphate, 0.1 mm sodium orthovanadate, 2 mm dithiothreitol, 0.185 MBq [γ-32P]ATP, and 20 μm ATP for 20 min at 30°C in a final volume of 40 μl. Reactions were stopped by addition of SDS-PAGE sample buffer and heating in a boiling water bath. Samples were separated by SDS-PAGE, and the gels were fixed and stained in Coomassie blue and then dried and subjected to autoradiography.
To assay autophosphorylation of cprk, cprk was immunoprecipitated from transfected CHO cells as described for cdk5/p35 above using rabbit anti-cprk antibody. Cprk was incubated in kinase buffer (see above) for 0–120 min at 30°C in a final volume of 40 μl. The reactions were stopped by addition of SDS sample buffer and heating in a boiling water bath, and the samples were then analyzed by SDS-PAGE and autoradiography to detect γ-32P incorporation as above. Such time course reactions revealed that cprk activity was in the linear range at 20 min, and all subsequent activity assays were performed at this time point. To detect immunoprecipitated cprk, samples were blotted and probed with the mouse cprk antibody.
Controls to demonstrate specificity of the cprk assay involved omission of cprk antibody or use of antibody that had been preabsorbed with antigen in the immunoprecipitations and also performing the reactions on samples of nontransfected CHO cells (CHO cells do not express cprk; see Figs. 2, 7, 8.
To analyze the effect of cdk5/p35 on cprk activity, CHO cells were transfected with cprk either alone or with cdk5/p35, and cprk autophosphorylation was analyzed as above. Signals for the relative amounts of cprk and γ-32P incorporation into cprk were quantified by scanning of immunoblots and autorads and analyzing these by pixel densitometry using a Bio-Rad GS710 imaging densitometer and Quantity 1 software as described (Lau et al., 2002). Data were analyzed using the one-way ANOVA test.
Results
p35 interacts with cprk in yeast two-hybrid and GST pull-down assays
To identify binding partners and possible substrates for cdk5/p35, we screened a yeast GAL-4 two-hybrid library with p35 bait (Kesavapany et al., 2001). After selection on Trp-, Leu- and His- plates, we obtained several vigorously growing yeast colonies. After rescue of prey plasmids, one was found to encode a fragment of β-catenin (Kesavapany et al., 2001), and a further comprised a novel cDNA. Screening of databases revealed that this novel cDNA encoded in-frame residues 391–632 of KIAA1079 (GenBank accession number AB029002), a clone isolated as part of the Kazusa HUGE cDNA project (Kikuno et al., 2002). We term the full-length protein (see below) encoded by KIAA1079 cprk. Yeast harboring the p35 bait and cprk prey plasmids produced blue signals after ∼3–4 hr when assayed for β-galactosidase using filter lift freeze fracture assays; yeast carrying full-length GAL-4 or p53/SV40 large T antigen-positive control plasmids produce blue signals after ∼2 hr in our hands. Thus the interaction between the p35 bait and cprk prey plasmids is strong in the two-hybrid system. The cprk plasmid was not capable of transactivating the β-galactosidase reporter in yeast Y190 when transformed alone, with empty pY2 bait plasmid, or with pY2 containing laminin as a negative control.
p35 can be proteolytically cleaved within cells by calpain to produce N-terminal p10 and C-terminal p25 fragments (Lew et al., 1994; Tsai et al., 1994; Uchida et al., 1994; Patrick et al., 1999; Kusakawa et al., 2000; Lee et al., 2000; Nath et al., 2000). We therefore used both the p10 and p25 fragments of p35 as baits to test for interactions with cprk using the yeast two-hybrid system. Only a weak signal was obtained with the p25 fragment, and no signal was obtained with the p10 fragment. Thus, full-length p35 appears to be required for a strong interaction with cprk.
The sequence of KIAA1079 contains an open reading frame of 4365 bp that encodes a protein of 1455 amino acids. An in-frame stop codon is situated 69 bp upstream of the ATG start codon; therefore, clone KIAA1079 contains the full-length cDNA for cprk. Identification of domains and motifs present in crpk using simple modular architecture research tool (SMART) analyses (http://smart.embl-heidelberg.de/) predicts two N-terminally located membrane-spanning domains (residues 12–34 and 44–64) and a dual-specificity serine/threonine/tyrosine kinase (STYKc) domain (residues 137–407) (supplementary data 1, available at www.jneurosci.org). Basic local alignment search tool (BLAST) analyses (http://dove.embl-heidelberg.de/Blast2/) revealed significant structural homology between cprk and KIAA1883 (a further cDNA identified by the Kazusa HUGE cDNA project), apoptosis-associated tyrosine kinase (AATYK) and a mouse protein similar to apoptosis-associated tyrosine kinase (GenBank accession number XM_145594). Like cprk, all three of these proteins contain an STYKc domain and both KIAA1883 and XM_145594 also contain two N-terminal membrane-spanning domains (see supplementary data 1, available at www.jneurosci.org).
To confirm by another method that cprk interacts with p35, we prepared GST-cprk391–632 fusion proteins and used these as baits in pull-down experiments from nontransfected, p35-transfected, and p35/cdk5-transfected CHO cells. Cdk5 but not p35 is expressed in nontransfected CHO cells (Kesavapany et al., 2001). GST-cprk391–632 but not GST interacted with p35 from both p35-only and p35/cdk5-transfected CHO cells but not from nontransfected CHO cells (Fig. 1A). We also performed pull-down assays with GST and GST-cprk391–632 from 5-d-old cultures of cortical neurons. p35 and cdk5 are both expressed in 5-d-old cortical neurons (Nikolic et al., 1996). Again, GST-cprk391–632 but not GST alone interacted with p35 in these assays (Fig. 1B).
Figure 1.

Cprk binds to p35 in GST-pull-down assays. GST or GST-cprk 391–632 baits were used to capture p35 in pull-down assays from either nontransfected (NT), p35-transfected (p35), or cdk5/p35-transfected CHO cells (A) or from cortical neurons (B). Samples were then probed for p35 on immunoblots after SDS-PAGE. Pull-down assays are shown on the left, and samples of the various lysates are shown on the right. GST-cprk391–632 binds to p35 in p35- and cdk5/p35-transfected CHO cells but not nontransfected CHO cells (A); GST-cprk391–632 binds endogenous p35 in neurons (B). CHO cells do not express endogenous p35, but neurons express both cdk5 and p35. GST alone does not interact with p35 in any assay.
Cprk and p35 colocalize in neurons and in cprk/p35-transfected COS cells
All of the above biochemical evidence strongly suggests that p35 and cprk are complexed in cells. However, it is possible that p35 and cprk are present in different compartments of the cell and are therefore unable to interact in vivo. We therefore generated rabbit and mouse cprk polyclonal antibodies to study the subcellular distribution of cprk. These antisera all detect a major species of ∼220 kDa on immunoblots of mouse and rat brain and transfected but not nontransfected CHO cells (data not shown; for example of one antibody, see Fig. 2A). Previous incubation of antibody with antigen eliminated this signal (Fig. 2B).
Figure 2.

Specificity of rabbit cprk antibody. A, Immunoblot of nontransfected (NT) and cprk-transfected (cprk) CHO cells and mouse and rat brain. B, Identical immunoblot that had been probed with antibody that was preincubated with cprk antigen; labeling is abolished.
Cdk5 and p35 are both expressed in cultured rat cortical neurons, so we first analyzed expression of cprk in these cells by immunoblotting. Cprk was not detected in 3-d-old but was present in 5- and 7-d-old neurons (data not shown). p35 is also expressed in cultured cortical neurons after 3 d in culture (Nikolic et al., 1996). In these cells, p35 is present along neurites but is enriched in cell bodies and growth cones (Nikolic et al., 1996), which we confirmed by labeling cells with p35 antibody C19. Double labeling of the neurons revealed that cprk was also enriched in cell bodies and growth cones, and that its expression overlapped with p35 (Fig. 3A–C).
Figure 3.
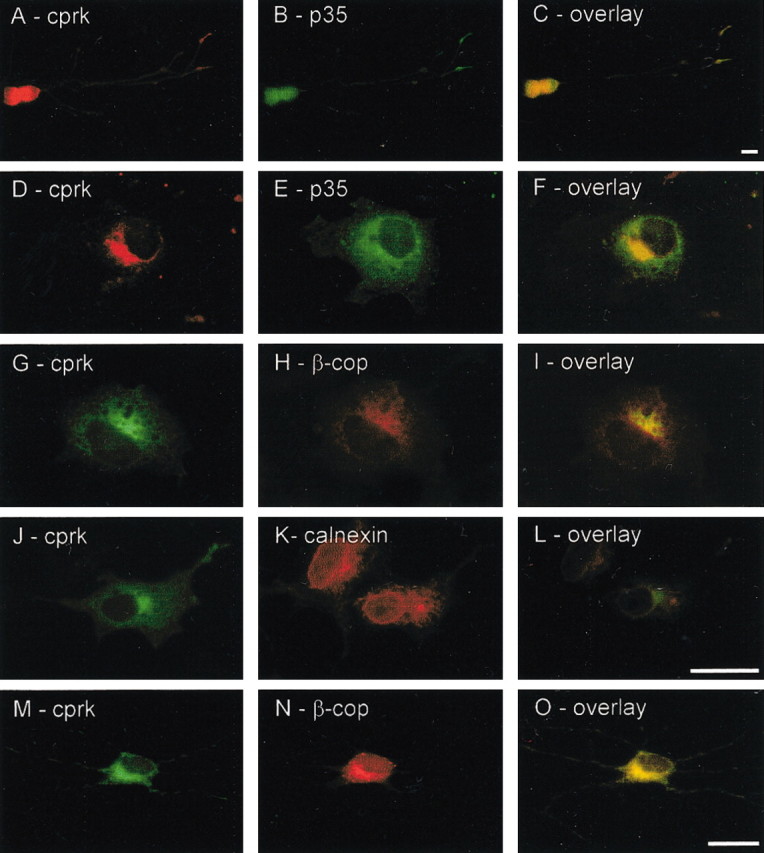
Cprk and p35 colocalize in cortical neurons and transfected COS cells, and cprk is present in the Golgi apparatus. A–C, Double immunostaining of 5-d-old cortical neurons for cprk (A) and p35 (B); an overlay is shown in C. D–F, Immunostaining for cprk (D) and p35 (E) in cprk/p35 cotransfected COS cells; an overlay is shown in F. G–L, Cprk andβ-cop (G–I) and cprk and calnexin (J–L) localization in cprk/p35 cotransfected COS cells; overlays are shown in I and L. M–O, Endogenous cprk and β-cop in cortical neurons; an overlay is shown in O. Scale bars, 20 μm.
We also analyzed the distribution of cprk and p35 in transfected COS cells. COS cells express endogenous cdk5 but not p35 (Guidato et al., 1996). In p35/cprk-transfected cells, both proteins were present in perinuclear areas where they colocalized (Fig. 3D–F). p35 contains an N-terminal myristoylation signal motif and is associated with membranes at the cell periphery and in perinuclear regions including Golgi (Patrick et al., 1999; Paglini et al., 2001). We therefore analyzed cprk localization using the Golgi and endoplasmic reticulum markers β-cop and calnexin; these studies revealed that cprk was also present in Golgi (Fig. 3G–L). Finally, we confirmed that a proportion of endogenous cprk is present in the Golgi in cortical neurons by double staining for cprk and β-cop. These experiments revealed overlapping staining patterns for cprk and β-cop within cell bodies of the neurons (Fig. 3M–O).
Thus, p35 and cprk interact in the yeast two-hybrid system and in GST pull-down assays and display overlapping subcellular distribution patterns in neurons and in transfected cells, where a proportion of cprk is present in the Golgi.
Cprk is expressed in multiple tissues and displays developmental and cell type-specific expression in brain
To characterize expression of cprk, we first performed Northern analyses to detect cprk mRNA in human tissues. Cprk mRNA was present in all tissues examined but was enriched in brain, heart, and skeletal muscle (Fig. 4A). We also analyzed developmental expression of cprk protein in brain by immunoblotting. Cprk was first detected at P0, after which expression levels increased markedly (Fig. 4B).
Figure 4.
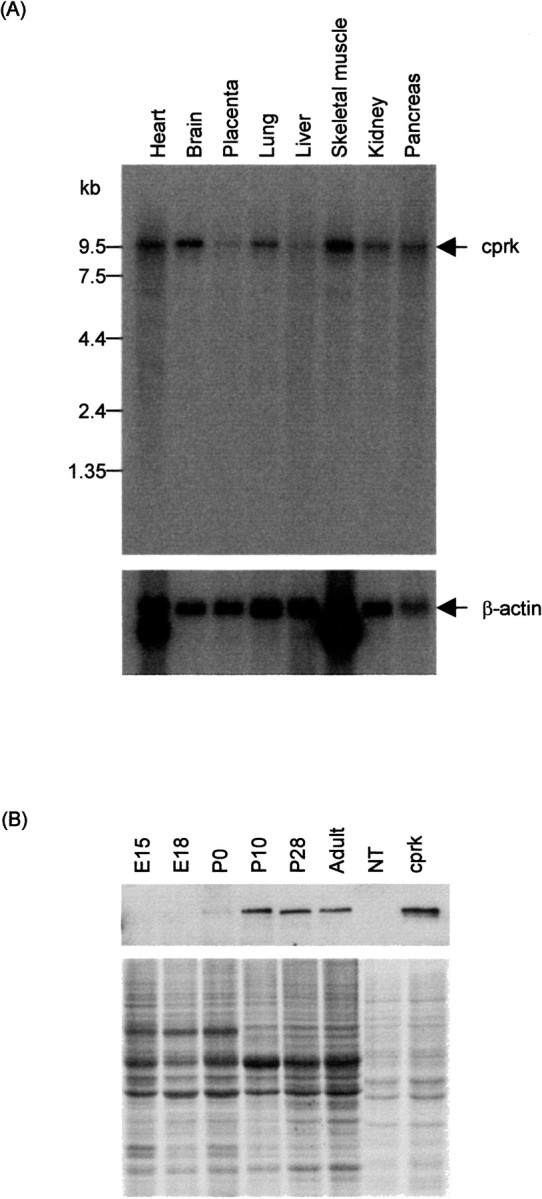
Expression of cprk. A, Northern blot showing cprk expression in human tissues; top, cprk; bottom,β-actin. B, Developmental expression of cprk in mouse brain; top, immunoblot of cprk; bottom, identically loaded Coomassie blue-stained gel showing equal protein loading of brain samples. NT, cprk, Samples of nontransfected and cprk-transfected CHO cells as controls.
We also mapped expression of cprk in mouse brain by immunostaining of tissue sections. Cprk was expressed in a variety of brain regions with a widespread distribution of immunoreactivity in a diverse range of neuronal subtypes (Fig. 5, supplementary data 2, available at www.jneurosci.org). However, cprk expression was not universal, with staining absent or not detectable above background in many neuronal populations. Cprk-positive neurons showed varying intensities of staining; for example, dense cprk immunoreactivity was present in the deep cerebellar nuclei but was virtually absent in neurons of the granule cell layer (Fig. 5E–G). Particularly intense cprk immunoreactivity was present in neurons of the reticular thalamic nucleus and both medial and lateral ventroposterior thalamic nuclei, whereas more medial and intralaminar nuclei and more ventral nuclei such as the zona incerta were virtually unstained (Fig. 5K–M).
Figure 5.
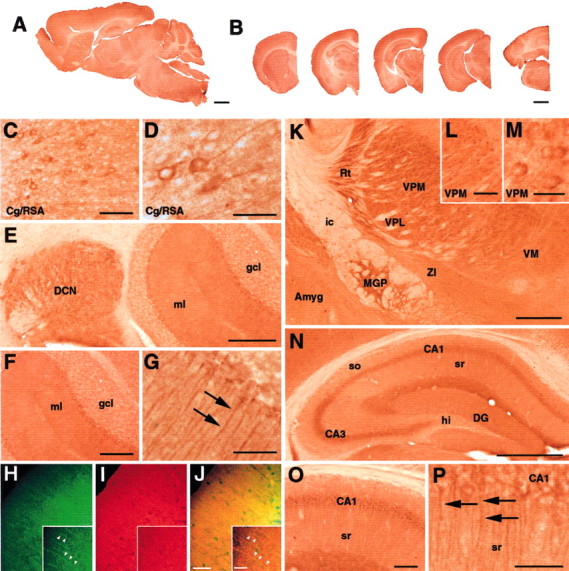
Localization of cprk in adult mouse brain. Immunohistochemical staining reveals widespread cprk expression within the CNS that varies in intensity between different brain regions, as demonstrated in low-power views of sagittal (A) and coronal (B) sections. Intense cprk immunoreactivity was observed in neurons in the cortical mantle (C) with well defined staining of apical dendrites (D). Neurons of the deep cerebellar nuclei (E) were also strongly labeled, which is in contrast to neurons of the granule cell layer (gcl), which exhibit minimal cprk immunoreactivity (E, F). The molecular layer (ml) contains numerous thin crpk-positive fibers (E,F, arrows in G) As revealed by dual-channel immunofluorescence, these cprk-positive fibers (green,arrowheads in H), are also labeled by GLAST (I, red), revealing processes immunoreactive for both antigens (yellow, arrowheads in J). The thalamus exhibits intense immunoreactivity in the medial globus pallidus (MGP), reticular (Rt), ventromedial posterior (VPM) and ventrolateral posterior (VPL) nuclei compared with the zona incerta (ZI), ventromedial nucleus (VM), or amygdala (Amyg; K–M). White matter such as the internal capsule (ic) is unstained (K). The hippocampal formation exhibits varied levels of cprk within areas CA3, CA1, and the hilus (hi; N). Neurons within CA1 exhibit particularly well defined cprk staining of apical dendrites extending into the stratum radiatum (O, arrows in P). The stratum oriens (so) and stratum radiatum (sr) contain numerous intensely stained interneurons (N). Scale bars: A, 1 mm; B, 1.25 mm; C, L, O, 100 μm; D, G, P,50 μm; E, 250 μm; H–J, 50 and 15 μm (inset); F, 200 μm; K, N, 500 μm; M, 25 μm.
Dense cprk immunoreactivity was also seen in specific structures within the olfactory bulb, basal ganglia, hypothalamus nucleus, midbrain, pons, and medulla, whereas only low cprk immunoreactivity was found in the septum (see supplementary data 2, available at www.jneurosci.org). Within the hippocampus, dense cprk immunoreactivity labeled neurons with interneuron morphology and distribution within the stratum oriens and stratum radiatum. Cprk staining in the principal hippocampal cell layers was more prominent in CA3 than CA1 but was completely absent in the hilus (Fig. 5N–P). Cprk immunoreactivity exhibited a laminar specific distribution within the neocortex, with most intense staining of neurons within lamina IV (Fig. 5C,D).
In the majority of areas, cprk immunoreactivity was primarily confined to the neuronal soma, often with fine granular immunoreactive perinuclear deposits on a background of paler staining of the cytoplasm, and a complete absence of nuclear staining. However, cprk immunoreactivity was also present in dendritic processes of several neuronal populations, most notably in the cortex (Fig. 5D) and CA1 region of the hippocampus (Fig. 5N–P). Cprk staining of the neuropil was evident in some brain regions but was almost entirely absent in the white matter. Although almost exclusively confined to neurons, cprk immunoreactivity was also clearly present in thin fibrous processes extending across the width of the cerebellar molecular layer, suggestive of Bergmann radial glial cell morphology (Fig. 5G). To investigate this further, sections were double immunfluorescently labeled with the rabbit cprk antibody and a guinea pig antibody to the glial glutamate transporter GLAST. GLAST is highly expressed in Bergmann glia (Rothstein et al., 1994; Schmitt et al., 1997). These dual-channel immunofluorescence studies revealed that the cprk-positive fibers were also immunoreactive for GLAST (Fig. 5H–J). Purkinje cell soma and dendrites did not exhibit cprk immunoreactivity in either coronal or sagittal sections.
In summary, cprk exhibits widespread but not universal expression within the CNS, although this expression was not entirely confined to neuronal populations. Cdk5, p35, and p39 are likewise broadly expressed in the nervous system (Zheng et al., 1998; Honjyo et al., 1999).
Cprk is phosphorylated by cdk5/p35
p35 has been shown to target the cdk5/p35 complex to its substrates (Lee et al., 1997a; Nikolic et al., 1998; Qi et al., 1998; Kesavapany et al., 2001) and as such, the interaction between p35 and cprk suggests that cprk is a substrate for cdk5/p35. Favored targets for cdk5/p35 are serines and threonines preceding a proline, and cprk contains several of these motifs (see supplementary data 1, available at www.jneurosci.org). We therefore performed in vitro kinase assays with recombinant GST-cprk391–632. Cdk5/p35 isolated by immunoprecipitation from transfected cells phosphorylated GST-cprk391–632 but not GST alone (Fig. 6).
Figure 6.

In vitro phosphorylation of cprk by cdk5/p35. GST-cprk391–632 and GST alone were phosphorylated by cdk5/p35 in the presence of [γ-32P]ATP. A, Cprk phosphorylation. B, Control reactions with GST alone. RM, Reaction mix only with no substrate; cprk, reaction mix with GST-cprk391–632 substrate; GST, reaction mix with GST substrate. +, -, Presence and absence of antibody 12CA5 to the hemagluttinin epitope tag on cdk5 in the cdk5/p35 kinase immunoprecipitations. Top, Autoradiographs; bottom, corresponding Coomassie blue-stained gels.
We also tested whether cprk is a cellular substrate for cdk5/p35. To do so, we immunoprecipitated cprk from CHO cells transfected with cprk alone or in combination with cdk5 and p35 and then analyzed cprk phosphorylation using antibodies that specifically detect phosphoserine, phosphothreonine, and phosphotyrosine residues. Cdk5/p35 is a serine/threonine kinase but does not phosphorylate tyrosines. As detailed above, CHO cells express endogenous cdk5 but not p35 (Kesavapany et al., 2001) and do not express endogenous cprk (see Figs. 2, 7, 8). In these experiments, transfections were balanced by including a vector containing the E. coli chloramphenicol acetyltransferase gene as a control so that all cells received the same amount of plasmids. These studies revealed that cprk labeling by the phosphoserine and phosphothreonine antibodies was markedly increased in cdk5/p35-cotransfected cells (Fig. 7). However, labeling of cprk by the phosphotyrosine antibody was diminished in cdk5/p35-cotransfected cells (Fig. 7). Treatment of the samples with λ-protein phosphatase before electrophoresis eliminated labeling by the phospho-dependent antibodies (Fig. 7), and this demonstrates the specificity of these reagents for phosphorylated residues. Thus, cprk is phosphorylated by cdk5/p35 in vitro, and cotransfection of cprk with cdk5/p35 increases cprk phosphorylation on serine and threonine but not tyrosine residues. Together, these studies demonstrate that cprk is a substrate for cdk5/p35.
Figure 7.

Cellular phosphorylation of cprk by cdk5/p35 on serine and threonine but not tyrosine residues in transfected CHO cells. Cells were transfected with cprk alone or with cprk and cdk5/p35. Cprk was immunoprecipitated with rabbit cprk antibody and then detected on immunoblots with mouse cprk antibody to demonstrate equal amounts of immunoprecipitated cprk (anti-cprk). Identically loaded blots were then probed with phosphoserine, phosphothreonine, and phosphotyrosine antibodies as indicated (anti-phospho-serine, -threonine, -tyrosine). For these latter immunoblots, samples were either untreated or treated with λ-protein phosphatase (λppase) before analyses. IPs, Immunoprecipitations; Lysates, samples of the lysates including nontransfected CHO cells (NT). -, +, Absence and presence of immunoprecipitating antibody to cprk in the pull-down assays.
Figure 8.

Cdk5/p35 inhibits cprk activity. Cprk autophosphorylation assays were performed on cprk immunoprecipitated from nontransfected (NT), cprk-transfected, or cprk- and cdk5/p35-cotransfected CHO cells. Top, Immunoblots probed with cprk antibody to demonstrate equal amounts of immunoprecipitated cprk; bottom, autoradiographs of γ-32P incorporation into cprk. -, +, Absence and presence of immunoprecipitating antibody to cprk in the pull-down assays. Two separate transfection experiments are shown for each treatment (expt 1, expt 2). The experiments were performed two additional times, and the relative cprk activities in nontransfected, cprk-transfected, and cprk- and cdk5/p35-transfected cells were then analyzed as shown in the bottom graph. Cdk5/p35 significantly inhibits cprk activity (p < 0.001, one-way ANOVA). Error bars indicate SD.
Cprk activity is regulated by cdk5/p35
The sequence of cprk predicts it to be a kinase. To formally test this possibility, we performed in vitro kinase assays with cprk immunoprecipitated from transfected CHO cells. Many kinases display autocatalytic activity (e.g., receptor-tyrosine kinases, protein kinase N/protein kinase C-related kinase, and cdk5/p35; Palmer and Parker, 1995; Patrick et al., 1998) and in the absence of known cprk substrates, we therefore studied whether cprk is capable of autophosphorylation. In these assays, cprk incorporated isotopic phosphate, but no such incorporation was observed in samples from nontransfected cells or in assays in which immunoprecipitating cprk antibody was omitted or in which the antibody was preabsorbed with cprk antigen (data not shown; Fig. 8). For these studies, the reactions were analyzed after 20 min, because pilot studies (involving halting the reactions at 0, 10, 20, 40, 60, and 120 min) revealed that at this time point, cprk activity was in the linear range (data not shown).
We next investigated whether cdk5/p35 influenced cprk activity in any way. To do so, cprk activity was assayed as above in cells transfected with cprk or cprk and cdk5/p35. These studies revealed that cdk5/p35 inhibited cprk activity (Fig. 8). This finding is consistent with the results shown in Figure 7, which show a diminution of phosphotyrosine labeling in cdk5/p35-cotransfected cells and which may well be attributable to decreased autophosphorylation of cprk on tyrosine residues. Thus, cprk is a kinase, and its activity is inhibited by cdk5/p35.
Discussion
In this report, we characterize a novel binding partner for the cdk5 activator p35, which we term cprk. Our analyses reveal that cprk is expressed in a number of tissues but is enriched in brain, muscle, and heart; whether the lower levels of expression in other tissues are attributable to innervation with cprk being present only in neurons is not clear at this stage. Cdk5/p35 and cdk5/p39 activities are restricted to the nervous system and muscle (Dhavan and Tsai, 2001). Cprk is expressed in a diverse range of neuronal cell types within the brain and is present in the Golgi apparatus. Interestingly, p35 is also present within membranous compartments, including the Golgi (Patrick et al., 1999; Paglini et al., 2001), and we show that cprk and p35 colocalize in these organelles.
The sequence of cprk predicts it to be a dual-specificity serine/threonine/tyrosine kinase. It contains two N-terminal membrane-spanning domains followed by a long C-terminal domain that contains the STYKc domain. In vitro kinase assays with immunoprecipitated cprk provide formal evidence that cprk is indeed a kinase. We further demonstrate that cprk is a substrate for cdk5/p35 and that cdk5/p35 inhibits cprk activity.
BLAST analyses revealed homology between the cprk STYKc domain and a number of other kinases, including receptor kinases. However, cprk displayed significant structural homology to KIAA1883 (a further cDNA identified by the Kazusa HUGE cDNA project) and a mouse protein similar to apoptosis-associated tyrosine kinase (GenBank accession number XM_145594). Like cprk, both of these proteins also contain two N-terminal membrane-spanning domains and an STYKc domain. Homology among cprk, KIAA1883, and XM_145594 was most notable within their N termini, which contain the two membrane-spanning domains and an STYKc domain. However, the sequences diverged toward their C termini apart from two short domains (residues 460–481 and 536–556 of cprk). These domains show no homology to other proteins. Thus there appears to be a family of cprk-related, membrane-associated dual-specificity kinases that all contain two N-terminal membrane-spanning domains (for alignment of cprk with other cprk family members, see supplementary data 1, available at www.jneurosci.org). However, no functions for any of these cprk-related kinases have so far been described. Our findings reported here are therefore the first detailed descriptions of any cprk family member.
AATYK, which displays significant homology to cprk over its kinase domain and within its C-terminal region but which does not contain N-terminal membrane-spanning domains, is implicated in programmed cell death (Gaozza et al., 1997). AATYK expression is induced during growth arrest and apoptosis in myeloid cells, and a spliced version of AATYK is expressed in the nervous system (Gaozza et al., 1997; Baker et al., 2001). AATYK may therefore function in cell growth and apoptosis within the nervous system, although whether cprk fulfils similar functions remains to be determined. However, it is interesting to note that cdk5/p35 has also been implicated in such functions (Lee et al., 1997a; Dhavan and Tsai, 2001).
Modulating cprk expression in cortical neurons and nonneuronal cells by transfection did not alter cell morphology in any gross detectable manner (our unpublished results). A proper understanding of cprk function will therefore require identification of its cellular substrates and any signaling pathways that regulate cprk activity in future studies. However, we show here that cdk5/p35 can regulate cprk autocatalytic activity, so one such pathway probably involves cdk5/p35.
Thus, we have identified a new binding partner for p35, cprk, which is a member of a novel family of membrane-associated kinases and which is present in Golgi. Cdk5/p35 phosphorylates cprk and regulates its activity, so signaling pathways that modulate cdk5/p35 activity may well also regulate cprk function.
Footnotes
This work was supported by grants from the Wellcome Trust, Medical Research Council, UK Motor Neurone Disease Association, including the Jim Tew Memorial Studentship, Tim Perkins Fund, Natalie Fund, and Croucher Foundation. We thank the Kazusa DNA Research Institute for clone KIAA1079 and Li-Huei Tsai (Harvard University, Cambridge, MA) for cdk5 and p35 cDNAs.
Correspondence should be addressed to Christopher C. J. Miller, Department of Neuroscience, The Institute of Psychiatry, P.O. Box P037, De Crespigny Park, Denmark Hill, London SE5 8AF, UK. E-mail: chris.miller@iop.kcl.ac.uk.
S. Kesavapany's present address: National Institute of Neurological Disorders and Stroke, National Institutes of Health, Building 36, 4D-28, Bethesda, MD 20892.
Copyright © 2003 Society for Neuroscience 0270-6474/03/234975-09$15.00/0
S.K. and K.-F.L. contributed equally to this work.
References
- Ackerley S, Grierson AJ, Brownlees J, Thornhill P, Anderton BH, Leigh PN, Shaw CE, Miller CCJ ( 2000) Glutamate slows axonal transport of neurofilaments in transfected neurons. J Cell Biol 150: 165–175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Agarwal-Mawal A, Paudel HK ( 2001) Neuronal Cdc2-like protein kinase (Cdk5/p25) is associated with protein phosphatase 1 and phosphorylates inhibitor-2. J Biol Chem 24: 24. [DOI] [PubMed] [Google Scholar]
- Ahlijanian MK, Barrezueta NX, Williams RD, Jakowski A, Kowsz KP, McCarthy S, Coskran T, Carlo A, Seymour PA, Burkhardt JE, Nelson RB, McNeish JD ( 2000) Hyperphosphorylated tau and neurofilament and cytoskeletal disruptions in mice overexpressing human p25, an activator of cdk5. Proc Natl Acad Sci USA 87: 2910–2915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alvarez A, Muñoz JP, Maccioni RB ( 2001) A cdk5–p35 stable complex is involved in the β-amyloid-induced deregulation of cdk5 activity in hippocampal neurons. Exp Cell Res 264: 266–274. [DOI] [PubMed] [Google Scholar]
- Bajaj NP, Miller CCJ ( 1997) Phosphorylation of neurofilament heavy-chain side-arm fragments by cyclin-dependent kinase-5 and glycogen synthase kinase-3α in transfected cells. J Neurochem 69: 737–743. [DOI] [PubMed] [Google Scholar]
- Bajaj NPS, Al-Sarraj S-T, Anderson V, Kibble M, Leigh N, Miller CCJ ( 1998) Cyclin-dependent kinase-5 is associated with lipofuscin in motor neurons in amyotrophic lateral sclerosis. Neurosci Lett 245: 45–48. [DOI] [PubMed] [Google Scholar]
- Baker SJ, Sumerson R, Reddy CD, Berrebi AS, Flynn DC, Reddy EP ( 2001) Characterization of an alternatively spliced AATYK mRNA: expression pattern of AATYK in the brain and neuronal cells. Oncogene 20: 1015–1021. [DOI] [PubMed] [Google Scholar]
- Baumann K, Mandelkow E-M, Biernat J, Piwnica-Worms H, Mandelkow E ( 1993) Abnormal Alzheimer-like phosphorylation of tau-protein by cyclin dependent kinases cdk2 and cdk5. FEBS Lett 336: 417–424. [DOI] [PubMed] [Google Scholar]
- Bibb JA, Snyder GL, Nishi A, Yan Z, Meijer L, Fienberg AA, Tsai L-H, Kwon YT, Girault J-A, Czernik AJ, Huganir RL, Hemmings HCJ, Nairn AG, Greengard P ( 1999) Phosphorylation of DARPP-32 by cdk5 modulates dopamine signalling in neurons. Nature 402: 669–671. [DOI] [PubMed] [Google Scholar]
- Bibb JA, Chen JS, Taylor JR, Svenningsson P, Nishi A, Snyder GL, Yan Z, Sagawa ZK, Ouimet CC, Nairn AC, Nestler EJ, Greengard P ( 2001a) Effects of chronic exposure to cocaine are regulated by the neuronal protein Cdk5. Nature 410: 376–380. [DOI] [PubMed] [Google Scholar]
- Bibb JA, Nishi A, O'Callaghan JP, Ule J, Lan M, Snyder GL, Horiuchi A, Saito T, Hisanaga S, Czernik AJ, Nairn AC, Greengard P ( 2001b) Phosphorylation of protein phosphatase inhibitor-1 by Cdk5. J Biol Chem 276: 14490–14497. [DOI] [PubMed] [Google Scholar]
- Brownlees J, Yates A, Bajaj NP, Davis D, Anderton BH, Leigh PN, Shaw CE, Miller CCJ ( 2000) Phosphorylation of neurofilament heavy chain sidearms by stress activated protein kinase-1b/Jun N-terminal kinase-3. J Cell Sci 113: 401–407. [DOI] [PubMed] [Google Scholar]
- Bu B, Li J, Davies P, Vincent I ( 2002) Deregulation of cdk5, hyperphosphorylation, and cytoskeletal pathology in the Niemann-Pick type C murine model. J Neurosci 22: 6515–6525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chae T, Kwon YT, Bronson R, Dikkes P, Li E, Tsai LH ( 1997) Mice lacking p35, a neuronal specific activator of Cdk5, display cortical lamination defects, seizures, and adult lethality. Neuron 18: 29–42. [DOI] [PubMed] [Google Scholar]
- Connell-Crowley L, Le Gall M, Vo DJ, Giniger E ( 2000) The cyclin-dependent kinase cdk5 controls multiple aspects of axon patterning in vivo. Curr Biol 10: 599–602. [DOI] [PubMed] [Google Scholar]
- Dhavan R, Tsai L-H ( 2001) A decade of cdk5. Nature Rev Mol Cell Biol 2: 749–759. [DOI] [PubMed] [Google Scholar]
- Dhavan R, Greer PL, Morabito MA, Orlando LR, Tsai LH ( 2002) The cyclin-dependent kinase 5 activators p35 and p39 interact with the α subunit of Ca2+/calmodulin-dependent protein kinase II and α-actinin-1 in a calcium-dependent manner. J Neurosci 22: 7879–7891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans DB, Rank KB, Bhattacharya K, Thomsen DR, Gurney ME, Sharma SK ( 2000) Tau phosphorylation at serine 396 and serine 404 by human recombinant tau protein kinase II inhibits tau's ability to promote microtubule assembly. J Biol Chem 275: 24977–24983. [DOI] [PubMed] [Google Scholar]
- Fischer A, Sananbenesi F, Schrick C, Spiess J, Radulovic J ( 2002) Cyclin-dependent kinase 5 is required for associative learning. J Neurosci 22: 3700–3707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fletcher AI, Shuang R, Giovannucci DR, Zhang L, Bittner MA, Stuenkel EL ( 1999) Regulation of exocytosis by cyclin-dependent kinase 5 via phosphorylation of Munc18. J Biol Chem 274: 4027–4935. [DOI] [PubMed] [Google Scholar]
- Floyd SR, Porro EB, Slepnev VI, Ochoa GC, Tsai LH, De Camilli P ( 2001) Amphiphysin 1 binds the cyclin-dependent kinase (cdk) 5 regulatory subunit p35 and is phosphorylated by cdk5 and cdc2. J Biol Chem 276: 8104–8110. [DOI] [PubMed] [Google Scholar]
- Fu AK, Fu WY, Cheung J, Tsim KW, Ip FC, Wang JH, Ip NY ( 2001) Cdk5 is involved in neuregulin-induced AChR expression at the neuromuscular junction. Nat Neurosci 4: 374–381. [DOI] [PubMed] [Google Scholar]
- Gaozza E, Baker SJ, Vora RK, Reddy EP ( 1997) AATYK: a novel tyrosine kinase induced during growth arrest and apoptosis of myeloid cells. Oncogene 15: 3127–3135. [DOI] [PubMed] [Google Scholar]
- Gilmore EC, Herrup K ( 2001) Neocortical cell migration: GABAergic neurons and cells in layers I and VI move in a cyclin-dependent kinase 5-independent manner. J Neurosci 21: 9690–9700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilmore EC, Ohshima T, Goffinet AM, Kulkarni AB, Herrup K ( 1998) Cyclin-dependent kinase 5-deficient mice demonstrate novel developmental arrest in cerebral cortex. J Neurosci 18: 6370–6377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grant P, Sharma P, Pant HC ( 2001) Cyclin-dependent protein kinase 5 (Cdk5) and the regulation of neurofilament metabolism. Eur J Biochem 268: 1534–1546. [PubMed] [Google Scholar]
- Guidato S, Tsai L-H, Woodgett J, Miller CCJ ( 1996) Differential cellular phosphorylation of neurofilament heavy side-arms by glycogen synthase kinase-3 and cyclin-dependent kinase-5. J Neurochem 66: 1698–1706. [DOI] [PubMed] [Google Scholar]
- Guidato S, McLoughlin D, Grierson AJ, Miller CCJ ( 1998) Cyclin D2 interacts with cdk-5 and modulates cellular cdk-5/p35 activity. J Neurochem 70: 492–500. [DOI] [PubMed] [Google Scholar]
- Hashiguchi M, Saito T, Hisanaga SI, Hashiguchi T ( 2002) Truncation of CDK5 activator p35 induces intensive phosphorylation of Ser202/Thr205 of human tau. J Biol Chem 277: 44525–44530. [DOI] [PubMed] [Google Scholar]
- Honjyo Y, Kawamoto Y, Nakamura S, Nakano S, Akiguchi I ( 1999) Immunohistochemical localization of CDK5 activator p39 in the rat brain. NeuroReport 10: 3375–3379. [DOI] [PubMed] [Google Scholar]
- Humbert S, Dhavan R, Tsai L ( 2000) p39 activates cdk5 in neurons, and is associated with the actin cytoskeleton. J Cell Sci 113: 975–983. [DOI] [PubMed] [Google Scholar]
- Kesavapany S, Lau K-F, McLoughlin DM, Brownlees J, Ackerley S, Leigh PN, Shaw CE, Miller CCJ ( 2001) p35/cdk5 binds and phosphorylates beta-catenin and regulates beta-catenin/presenilin-1 interaction. Eur J Neurosci 13: 241–247. [PubMed] [Google Scholar]
- Kesavapany S, Banner SJ, Lau KF, Shaw CE, Miller CC, Cooper JD, McLoughlin DM ( 2002) Expression of the Fe65 adapter protein in adult and developing mouse brain. Neuroscience 115: 951–960. [DOI] [PubMed] [Google Scholar]
- Keshvara L, Magdaleno S, Benhayon D, Curran T ( 2002) Cyclin-dependent kinase 5 phosphorylates disabled 1 independently of Reelin signaling. J Neurosci 22: 4869–4877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kikuno R, Nagase T, Waki M, Ohara O ( 2002) HUGE: a database for human large proteins identified in the Kazusa cDNA sequencing project. Nucleic Acids Res 30: 166–168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ko J, Humbert S, Bronson RT, Takahashi S, Kulkarni AB, Li E, Tsai LH ( 2001) p35 and p39 are essential for cyclin-dependent kinase 5 function during neurodevelopment. J Neurosci 21: 6758–6771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kobayashi S, Ishiguro K, Omori A, Takamatsu M, Arioka M, Imahori K, Uchida T ( 1993) A cdc2-related kinase PSSALRE/cdk5 is homologous with the 30 kDa subunit of tau protein kinase II, a proline-directed protein kinase associated with microtubule. FEBS Lett 335: 171–175. [DOI] [PubMed] [Google Scholar]
- Kusakawa G-I, Saito T, Onuki R, Ishiguro K, Kishimoto T, Hisanaga S-I ( 2000) Calpain-dependent proteolytic cleavage of the p35 cyclin-dependent kinase 5 activator to p25. J Biol Chem 275: 17166–17172. [DOI] [PubMed] [Google Scholar]
- Kwon YT, Gupta A, Zhou Y, Nikolic M, Tsai L-H ( 2000) Regulation of N-cadherin-mediated adhesion by the p35-Cdk5 kinase. Curr Biol 10: 363–372. [DOI] [PubMed] [Google Scholar]
- Lau KF, Howlett DR, Kesavapany S, Standen CL, Dingwall C, McLoughlin DM, Miller CCJ ( 2002) Cyclin-dependent kinase-5/p35 phosphorylates presenilin 1 to regulate carboxy-terminal fragment stability. Mol Cell Neurosci 20: 13–20. [DOI] [PubMed] [Google Scholar]
- Lee KY, Helbing CC, Choi K-S, Johnston RN, Wang JH ( 1997a) Neuronal Cdc2-like kinase (Nclk) binds and phosphorylates the retinoblastoma protein. J Biol Chem 272: 5622–5626. [DOI] [PubMed] [Google Scholar]
- Lee KY, Qi Z, Yu YP, Wang JH ( 1997b) Neuronal Cdc2-like kinases: neuron-specific forms of Cdk5. Int J Biochem Cell Biol 29: 951–958. [DOI] [PubMed] [Google Scholar]
- Lee MS, Kwon YT, Li M, Peng J, Friedlander RM, Tsai LH ( 2000) Neurotoxicity induces cleavage of p35 to p25 by calpain. Nature 405: 360–364. [DOI] [PubMed] [Google Scholar]
- Lew J, Huang Q-Q, Zhong Q, Winkfein RJ, Aebersold R, Hunt T, Wang JH ( 1994) A brain-specific activator of cyclin-dependent kinase 5. Nature 371: 423–426. [DOI] [PubMed] [Google Scholar]
- Li BS, Zhang L, Takahashi S, Ma W, Jaffe H, Kulkarni AB, Pant HC ( 2002) Cyclin-dependent kinase 5 prevents neuronal apoptosis by negative regulation of c-Jun N-terminal kinase 3. EMBO J 21: 324–333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michel G, Mercken M, Murayama M, Noguchi K, Ishiguro K, Imahori K, Takashima A ( 1998) Characterization of tau phosphorylation in glycogen synthase kinase-3β and cyclin dependent kinase-5 activator (p23) transfected cells. Biochim Biophys Acta 1380: 177–182. [DOI] [PubMed] [Google Scholar]
- Nath R, Davis M, Probert AW, Kupina NC, Ren X, Schielke GP, Wang KK ( 2000) Processing of cdk5 activator p35 to its truncated form (p25) by calpain in acutely injured neuronal cells. Biochem Biophys Res Commun 274: 16–21. [DOI] [PubMed] [Google Scholar]
- Nguyen MD, Larivière RC, Julien JP ( 2001) Deregulation of Cdk5 in a mouse model of ALS: toxicity alleviated by perikaryal neurofilament inclusions. Neuron 30: 135–147. [DOI] [PubMed] [Google Scholar]
- Niethammer M, Smith DS, Ayala R, Peng JM, Ko J, Lee MS, Morabito M, Tsai LH ( 2000) NUDEL is a novel Cdk5 substrate that associates with LIS1 and cytoplasmic dynein. Neuron 28: 697–711. [DOI] [PubMed] [Google Scholar]
- Nikolic M, Dudek H, Kwon YT, Ramos YFM, Tsai LH ( 1996) The cdk5/p35 kinase is essential for neurite outgrowth during neuronal differentiation. Genes Dev 10: 816–825. [DOI] [PubMed] [Google Scholar]
- Nikolic M, Chou MM, Lu W, Mayer BJ, Tsai L-H ( 1998) The p35/cdk5 kinase is a neuron-specific Rac effector that inhibits Pak1 activity. Nature 395: 194–198. [DOI] [PubMed] [Google Scholar]
- Nishi A, Bibb JA, Matsuyama S, Hamada M, Higashi H, Nairn AC, Greengard P ( 2002) Regulation of DARPP-32 dephosphorylation at PKA- and Cdk5-sites by NMDA and AMPA receptors: distinct roles of calcineurin and protein phosphatase-2A. J Neurochem 81: 832–841. [DOI] [PubMed] [Google Scholar]
- Ohshima T, Ward JM, Huh C-G, Longenecker G, Veeranna., Pant HC, Brady RO, Martin LJ, Kulkarni AB ( 1996) Targeted disruption of the cyclin-dependent kinase 5 gene results in abnormal corticogenesis, neuronal pathology and perinatal death. Proc Natl Acad Sci USA 93: 11173–11178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohshima T, Gilmore EC, Longenecker G, Jacobowitz DM, Brady RO, Herrup K, Kulkarni AB ( 1999) Migration defects of cdk5(-/-) neurons in the developing cerebellum is cell autonomous. J Neurosci 19: 6017–6026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohshima T, Ogawa M, Veeranna, Hirasawa M, Longenecker G, Ishiguro K, Pant HC, Brady RO, Kulkarni AB, Mikoshiba K ( 2001) Synergistic contributions of cyclin-dependant kinase 5/p35 and Reelin/Dab1 to the positioning of cortical neurons in the developing mouse brain. Proc Natl Acad Sci USA 98: 2764–2769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohshima T, Ogawa M, Takeuchi K, Takahashi S, Kulkarni AB, Mikoshiba K ( 2002) Cyclin-dependent kinase 5/p35 contributes synergistically with Reelin/Dab1 to the positioning of facial branchiomotor and inferior olive neurons in the developing mouse hindbrain. J Neurosci 22: 4036–4044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paglini G, Cáceres A ( 2001) The role of the Cdk5–p35 kinase in neuronal development. Eur J Biochem 268: 1528–1533. [PubMed] [Google Scholar]
- Paglini G, Pigino G, Kunda P, Morfini G, Maccioni R, Quiroga S, Ferreira A, Cáceres A ( 1998) Evidence for the participation of the neuron-specific CDK5 activator p35 during laminin-enhanced axonal growth. J Neurosci 18: 9858–9869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paglini G, Peris L, Diez-Guerra J, Quiroga S, Caceres A ( 2001) The Cdk5–p35 kinase associates with the Golgi apparatus and regulates membrane traffic. EMBO Rep 2: 1139–1144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palmer RH, Parker PJ ( 1995) Expression, purification and characterization of the ubiquitous protein kinase C-related kinase 1. Biochem J 309: 315–320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patrick G, Zukerberg L, Nikolic M, de la Monte S, Dikkes P, Tsai L-H ( 1999) Conversion of p35 to p25 deregulates Cdk5 activity and promotes neurodegeneration. Nature 402: 615–622. [DOI] [PubMed] [Google Scholar]
- Patrick GN, Zhou PB, Kwon YT, Howley PM, Tsai LH ( 1998) p35, the neuronal-specific activator of cyclin-dependent kinase 5 (Cdk5) is degraded by the ubiquitin-proteasome pathway. J Biol Chem 273: 24057–24064. [DOI] [PubMed] [Google Scholar]
- Paudel HK ( 1997) Phosphorylation by neuronal cdc2-like protein kinase promotes dimerization of tau protein in vitro. J Biol Chem 272: 28328–28334. [DOI] [PubMed] [Google Scholar]
- Paudel HK, Lew J, Ali Z, Wang JH ( 1993) Brain proline-directed protein kinase phosphorylates tau on sites that are abnormally phosphorylated in tau associated with Alzheimer's paired helical filaments. J Biol Chem 268: 23512–23518. [PubMed] [Google Scholar]
- Pigino G, Paglini G, Ulloa L, Avila J, Cáceres A ( 1997) Analysis of the expression, distribution and function of cyclin dependent kinase 5 (cdk5) in developing cerebellar macroneurons. J Cell Sci 110: 257–270. [DOI] [PubMed] [Google Scholar]
- Qi Z, Tang DM, Zhu XD, Fujita DJ, Wang JH ( 1998) Association of neurofilament proteins with neuronal Cdk5 activator. J Biol Chem 273: 2329–2335. [DOI] [PubMed] [Google Scholar]
- Rashid T, Banerjee M, Nikolic M ( 2001) Phosphorylation of Pak1 by the p35/Cdk5 kinase affects neuronal morphology. J Biol Chem 276: 49043–49052. [DOI] [PubMed] [Google Scholar]
- Rothstein JD, Martin L, Levey AI, Dykes-Hoberg M, Jin L, Wu D, Nash N, Kuncl RW ( 1994) Localisation of glial and neuronal glutamate transporters. Neuron 13: 713–725. [DOI] [PubMed] [Google Scholar]
- Sasaki S, Shionoya A, Ishida M, Gambello MJ, Yingling J, Wynshaw-Boris A, Hirotsune S ( 2000) A LIS1/NUDEL/cytoplasmic dynein heavy chain complex in the developing and adult nervous system. Neuron 28: 681–696. [DOI] [PubMed] [Google Scholar]
- Schmitt A, Asan E, Puschel B, Kugler P ( 1997) Cellular and regional distribution of the glutamate transporter GLAST in the CNS of rats: nonradioactive in situ hybridization and comparative immunocytochemistry. J Neurosci 17: 1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma M, Sharma P, Pant HC ( 1999) CDK-5-mediated neurofilament phosphorylation in SHSY5Y human neuroblastoma cells. J Neurochem 73: 79–86. [DOI] [PubMed] [Google Scholar]
- Sharma P, Veeranna, Sharma M, Amin ND, Sihag RK, Grant P, Ahn N, Kulkarni AB, Pant HC ( 2002) Phosphorylation of MEK1 by cdk5/p35 down-regulates the mitogen-activated protein kinase pathway. J Biol Chem 277: 528–534. [DOI] [PubMed] [Google Scholar]
- Shetty KT, Link WT, Pant HC ( 1993) cdc2-like kinase from rat spinal cord specifically phosphorylates KSPXK motifs in neurofilament proteins: isolation and characterization. Proc Natl Acad Sci USA 90: 6844–6848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith DS, Niethammer M, Ayala R, Zhou Y, Gambello MJ, Wynshaw-Boris A, Tsai LH ( 2000) Regulation of cytoplasmic dynein behaviour and microtubule organization by mammalian Lis1. Nat Cell Biol 2: 767–775. [DOI] [PubMed] [Google Scholar]
- Sobue K, Agarwal-Mawal A, Li W, Sun W, Miura Y, Paudel HK ( 2000) Interaction of neuronal Cdc2-like protein kinase with microtubule-associated protein tau. J Biol Chem 275: 16673–16680. [DOI] [PubMed] [Google Scholar]
- Sun D, Leung CL, Liem RKH ( 1996) Phosphorylation of the high molecular weight neurofilament protein (NF-H) by cdk-5 and p35. J Biol Chem 271: 14245–14251. [DOI] [PubMed] [Google Scholar]
- Tanaka T, Veeranna, Ohshima T, Rajan P, Amin N, Cho A, Sreenath T, Pant H, Brady R, Kulkarni A ( 2001) Neuronal cyclin-dependent kinase 5 activity is critical for survival. J Neurosci 21: 550–558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomizawa K, Ohta J, Matsushita M, Moriwaki A, Li ST, Takei K, Matsui H ( 2002) Cdk5/p35 regulates neurotransmitter release through phosphorylation and downregulation of P/Q-type voltage-dependent calcium channel activity. J Neurosci 22: 2590–2597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsai L-H, Delalle I, Caviness JVS, Chae T, Harlow E ( 1994) p35 is a neural-specific regulatory subunit of cyclin-dependent kinase 5. Nature 371: 419–423. [DOI] [PubMed] [Google Scholar]
- Uchida T, Ishiguro K, Ohnuma J, Takamatsu M, Yonekura S, Imahori K ( 1994) Precursor of cdk5 activator, the 23 kDa subunit of tau protein kinase II: its sequence and developmental change in brain. FEBS Lett 355: 35–40. [DOI] [PubMed] [Google Scholar]
- Zhang J, Krishnamurthy PK, Johnson GV ( 2002) Cdk5 phosphorylates p53 and regulates its activity. J Neurochem 81: 307–313. [DOI] [PubMed] [Google Scholar]
- Zheng M, Leung CL, Liem RKH ( 1998) Region-specific expression of cyclin-dependent kinase 5 (cdk5) and its activators, p35 and p39, in the developing and adult rat central nervous system. J Neurobiol 35: 141–159. [DOI] [PubMed] [Google Scholar]