

Abstract
In several species, including humans, the dentate granule cell layer (GCL) of the hippocampus exhibits neurogenesis throughout adult life. The ability to regulate adult neurogenesis pharmacologically may be of therapeutic value as a mechanism for replacing lost neurons. Insulin-like growth factor-I (IGF-I) is a growth-promoting peptide hormone that has been shown to have neurotrophic properties. The relationship between IGF-I and adult hippocampal neurogenesis is to date unknown. The aim of this study was to investigate the effect of the peripheral administration of IGF-I on cellular proliferation in the dentate subgranular proliferative zone, which contains neuronal progenitor cells, and on the subsequent migration and differentiation of progenitor cells within the GCL. Using bromodeoxyuridine (BrdU) labeling, we found a significant increase of BrdU-immunoreactive progenitors in the GCL after 6 d of peripheral IGF-I administration. To determine the cell fate in progenitor progeny, we characterized the colocalization of BrdU-immunolabeled cells with cell-specific markers. In animals treated with IGF-I for 20 d, BrdU-positive cells increased significantly. Furthermore, the fraction of newly generated neurons in the GCL increased, as evaluated by the neuronal markers Calbindin D28K, microtubule-associated protein-2, and NeuN. There was no difference in the fraction of newly generated astrocytes. Thus, our results show that peripheral infusion of IGF-I increases progenitor cell proliferation and selectively induces neurogenesis in the progeny of adult neural progenitor cells. This corresponds to a 78 ± 17% (p < 0.001) increase in the number of new neurons in IGF-I-treated animals compared with controls.
Keywords: brain, rat, IGF-I, insulin-like growth factor-I, stem cell, progenitor cell, hippocampus, neurogenesis, stereology
Most CNS neurons are terminally differentiated and are, thus, not replaced after neuronal cell death. However, self-renewing cells with multilineage potential, designated stem cells or progenitor cells, have been identified in the adult mammalian brain (Reynolds and Weiss, 1992; Richards et al., 1992;Craig et al., 1996; Suhonen et al., 1996; McKay, 1997; Temple and Alvarez-Buylla, 1999). Neural stem cells generate new neurons throughout life in the dentate gyrus of the hippocampus in several species, including humans (Altman and Das, 1965; Kuhn et al., 1996;Eriksson et al., 1998). Progenitor cells isolated from this region have demonstrated the capacity for self-renewal in vitro; when grafted back into the adult brain, they have the potential to develop into mature neurons with the phenotypic characteristics of surrounding neurons (Suhonen et al., 1996). Proliferation and differentiation of adult neural stem cells are affected by age, growth factors, excitatory input, adrenal steroids, seizures, and enrichment of the environment (Cameron and Gould, 1994; Palmer et al., 1995; Craig et al., 1996; Kuhn et al., 1996; Bengzon et al., 1997; Gould et al., 1997; Kempermann et al., 1997; McKay, 1997; Nilsson et al., 1999). However, there are to our knowledge no known exogenous substances that selectively induce neurogenesis in the adult mammalian CNS in vivo.
Insulin-like growth factor-I (IGF-I) is a growth-promoting peptide that is important during development of the brain (Sara and Carlsson-Skwirut, 1988; Bozyczko-Coyne et al., 1993; Giacobini et al., 1995; Dentremont et al., 1999). Overexpression of IGF-I, both peripherally and centrally, in transgenic mice results in an increase in brain size and myelin content (Carson et al., 1993). Conversely, the number of granule cells in the hippocampus, the oligodendrocyte and neuron density within the olfactory bulbs, and total brain size are reduced in knock-out mice (Beck et al., 1995; Cheng et al., 1998).
There are two potential pathways by which IGF-I may affect the hippocampus. IGF-I receptors expressed in the adult dentate gyrus (Lesniak et al., 1988) may be reached by IGF-I either from the periphery (by crossing the blood–brain barrier) (Duffy et al., 1988;Reinhardt and Bondy, 1994) or locally by paracrine IGF-I (Breese et al., 1996). In the present study we wanted to assess the impact of peripheral, normally liver-derived, IGF-I.
It is possible that the reported alterations in hippocampal granule cell number in animals with supranormal or subnormal IGF-I levels occur in part as a result of a perturbation in the proliferation of endogenous neural progenitor cells. This hypothesis led us to investigate the proliferation and differentiation of progenitor cells in the adult dentate gyrus in hypophysectomized (hx) rats treated with peripheral administration of IGF-I. The hx rat is widely used as a model of somatic IGF-I deficiency because hx animals have low circulating levels of IGF-I (Schoenle et al., 1982; Sjöberg et al., 1994). We used bromodeoxyuridine (BrdU) labeling to monitor proliferation and double or triple immunofluorescent labeling for BrdU and cell-specific markers to determine the phenotype of the progenitor progeny.
MATERIALS AND METHODS
Animals and hormonal treatment. We used hx rats, which have low circulating levels of IGF-I (Schoenle et al., 1982; Sjöberg et al., 1994). The effect of IGF-I treatment was shown by comparing the weight gain in treated versus untreated (control) hx rats (data not shown) (Schoenle et al., 1982; Fielder et al., 1996). Female Sprague Dawley rats (Møllegaard Breeding Center, Ejby, Denmark) were hypophysectomized at 50 d of age at the Møllegaard Breeding Center. Weight-matched female normal (non-hx) rats were also investigated. All rats were maintained under standard conditions of temperature (24–26°C) and humidity (50–60%) and with lights on between 05:00 and 19:00 hr. The rats had access to water and standard laboratory chow ad libitum. Hormonal treatment started 7–10 d after hypophysectomy. All hx rats received subcutaneous injections of cortisol phosphate (400 μg/kg per d; Solu-Cortef, Upjohn, Puurs, Belgium) andl-thyroxine (10 μg/kg per d; Nycomed Pharma, Oslo, Norway) diluted in saline (08:00 hr) (Sjöberg et al., 1994). l-Thyroxine and glucocorticoids were given because these hormones are known to be important for normal central neural plasticity and function (Sinha et al., 1994; McEwen, 1996; Reagan and McEwen, 1997). Control hx rats were given cortisol and l-thyroxine. Recombinant human IGF-I (Genentech, South San Francisco, CA) was diluted in saline and given as a continuous infusion using Alzet 2001 osmotic minipumps (Alza, Palo Alto, CA; 1.25 mg/kg per d) for short-term experiments and Alzet 2004 (0.9 mg/kg per d) for long-term experiments, resulting in comparable levels of serum IGF-I (see Results). The osmotic minipumps were implanted subcutaneously on the backs of the rats. Hormonal treatment continued for 6 d (short-term) or 20 d (long-term). Altogether three separate sets of animals were used (one short-term and two long-term). During the first 5 d of each treatment period, all animals received a daily intraperitoneal injection of bromodeoxyuridine (BrdU; 50 mg/kg of body weight; Boehringer Mannheim, Scandinavia AB, Bromma, Sweden). The animals were killed by decapitation at the end of each treatment period. The experimental methods were approved by the Board of Animal Ethics (Göteborg University, Göteborg, Sweden).
Immunohistochemistry. After decapitation, the brains were removed and fixed in 4% p-formaldehyde for 24 hr and stored thereafter in a 30% sucrose solution. Coronal sections (40 μm) obtained by the use of a freezing microtome were stored in a cryoprotectant (25% ethylene glycol and 25% glycerin in a 0.05m phosphate buffer) at −20°C before immunohistochemistry or immunofluorescence. Staining was done on free-floating 40 μm sections pretreated with 0.6% H2O2 in Tris-buffered saline (TBS; 0.15 m NaCl and 0.1m Tris-HCl, pH 7.5) for 30 min to block endogenous peroxidase activity. To ensure the detection of BrdU-labeled nuclei, we denatured the DNA before incubation with mouse anti-BrdU antibody (1:400; Boehringer Mannheim). DNA denaturation was performed in the following manner: tissue was incubated in 50% formamide and 2× SSC (1× SSC, 0.3 m NaCl and 0.03m sodium citrate) for 2 hr at 65°C, rinsed for 15 min in 2× SSC, incubated again for 30 min in 2m HCl at 37°C, and rinsed again for 10 min in 0.1 m boric acid at pH 8.5. The tissue was then rinsed in TBS several times, followed by incubation in TBS, 0.25% Triton X-100, and 3% normal horse serum (TBS-TS) for 30 min and then with primary antibody in TBS-TS overnight at 4°C. The tissue sections were then incubated for 2 hr with biotinylated horse anti-mouse IgG (1:160) secondary antibodies (Vector Laboratories, Burlingame, CA) and rinsed in TBS. Avidin—biotin–peroxidase complex was applied for 1 hr before 5 min of peroxidase detection (using 0.25 mg/ml diaminobenzidine, 0.01% H2O2, and 0.04% NiCl).
Immunofluorescence. Sections were treated for DNA denaturation, as described above, and then incubated in TBS-TS for 30 min. Thereafter, they were incubated with rabbit anti-Calbindin D28K (1:500; SWant, Bellinzona, Switzerland), mouse NeuN (1:30; Chemicon, Temecula, CA), mouse microtubule-associated protein-2 (MAP2; 1:400; Boehringer Mannheim, Mannheim, Germany), or rabbit anti-GFAP (1:500; Dako, Glostrup, Denmark) and rat anti-BrdU antibody (1:200; Harlan, Loughborough, United Kingdom) overnight at 4°C. GFAP and Calbindin D28K were detected with Texas Red-conjugated anti-rabbit IgG (1:200 for GFAP and 1:100 for Calbindin D28K; Jackson ImmunoResearch, West Grove, PA), Calbindin D28K was also detected with Cy5-conjugated anti-rabbit IgG (1:150; Jackson ImmunoResearch), NeuN and MAP2 were detected with Texas Red-conjugated anti-mouse IgG (1:100; Jackson ImmunoResearch), and BrdU was labeled with an FITC-conjugated anti-rat IgG (1:150; Jackson ImmunoResearch) for 2 hr at 37°C. Immunofluorescence was detected and processed with a Nikon Diaphot fluorescence microscope and confocal laser-scanning microscopy using a Bio-Rad 1024 system (Hercules, CA).
Quantification. For each animal, the number of BrdU-positive cells in the granule cell layer (GCL; including the subgranular layer) and in the hilus and their corresponding sample volumes were determined in 12 immunoperoxidase-stained, 40-μm-thick coronal sections taken 240 μm apart. Area estimations were obtained by planimetry, using a Lucivid device (MicroBrightField, Colchester, VT) attached to a Leica microscope. The section thickness of 40 μm (microtome setting) was used in the dissector estimation of volume. The number of BrdU-positive cells was counted within the GCL and two cell diameters below the GCL, ignoring the cells in the uppermost focal plane and focusing through the thickness of the section to avoid errors caused by oversampling [for a discussion of the optical dissector principle, seeGundersen et al. (1988); West (1993); Coggeshall and Lekan (1996)]. Pictures were taken under a Nikon microscope equipped with a video camera. The results are expressed as BrdU-positive cells per sample volume per section. We determined the total number of BrdU-positive cells in the GCL and in the hilus. We also determined the colocalization of BrdU with cell-specific markers in the GCL in six to eight 40-μm-thick coronal sections taken 240 μm apart in each animal. For the neuronal marker Calbindin D28K, 457 BrdU-positive cells were analyzed respecting colocalization; for the neuronal marker NeuN, 735 BrdU-positive cells were analyzed; for the neuronal marker MAP2, 203 BrdU-positive cells were studied; and for the glial marker GFAP, 911 BrdU-positive cells were analyzed. All cell-counting procedures were blindly performed. The absolute volumes of the GCL and the hilus were determined in a Nissl-stained series of sections taken 480 μm apart throughout the hippocampus.
IGF-I measurement. Serum IGF-I concentrations were determined by a hydrochloric acid–ethanol extraction RIA, using human IGF-I for labeling (Nichols Institute Diagnostics, San Juan Capistrano, CA) (Sjöberg et al., 1994).
Statistical analysis. Values are expressed as the mean ± SEM. Comparisons between groups were made with one-way ANOVA throughout the study. Two-way ANOVA was applied to the results on colocalization of BrdU and Calbindin D28K/NeuN. Regression analysis with correlation analysis was also performed. Differences that are not statistically significant are followed by the abbreviation NS. p values < 0.05 were considered statistically significant.
RESULTS
IGF-I stimulates proliferation of neural progenitors in the adult rat dentate gyrus
Two IGF-I treatment protocols were used in the IGF-I-deficient hypophysectomized rats. The 6 d treatment was performed to evaluate the short-term effect of IGF-I on proliferation, and the 20 d treatment was used to evaluate effects on the cell fate of BrdU-immunoreactive progenitor progeny.
Blood samples were taken when the animals were killed; serum levels of IGF-I were measured to make sure that the miniosmotic delivery system was functioning properly. One day after the last injection of BrdU in animals treated for 6 d, the serum level was 148 ± 35 μg/dm3 compared with 47 ± 1.6 μg/dm3 in hx controls (p < 0.01). Fifteen days after the last injection of BrdU in animals treated for 20 d, the serum level was 222 ± 23 μg/dm3 in IGF-I-treated animals compared with 65 ± 7.0 μg/dm3 in hx controls (p < 0.001). Additionally, the weight gain of rats treated with IGF-I showed the expected response (data not shown). Together, these data show that the peripheral infusion of IGF-I reached the systemic circulation, thereby being able to affect the brain by passing the blood–brain barrier.
The number of newly generated cells in the adult dentate gyrus was determined by monitoring the incorporation and subsequent immunohistochemical detection of BrdU within the nuclei of dividing cells (Fig. 1b–e). BrdU-immunoreactive nuclei were located predominantly within the GCL or along the border between the GCL and the hilus. The nuclei were generally condensed and exhibited variable shapes. The number of BrdU-immunoreactive cells in IGF-I-treated animals was compared with the number of BrdU-immunoreactive cells in the hx controls. In animals that underwent short-term (6 d) IGF-I treatment [i.e., animals evaluated 1 d post-BrdU injection (p.i.)], the number of BrdU-immunoreactive cells in the GCL was 7291 ± 950 cells/mm3 compared with 4076 ± 731 cells/mm3 in hx controls, which corresponds to a 79 ± 29% increase (p = 0.015) (Fig. 2a). In the hilus, the number of BrdU-immunoreactive cells was 2765 ± 331 cells/mm3 in IGF-I-treated animals compared with 1808 ± 211 cells/mm3in hx controls (p < 0.05).
Fig. 1.

BrdU immunohistochemistry in two experiments in which IGF-I was given for 6 d (corresponding to 1 d p.i. of BrdU) or 20 d (corresponding to 15 d p.i. of BrdU). a, The hippocampal region of the adult rat brain immunoperoxidase-stained for the neuronal marker Calbindin D28K. b–e, Differential interference contrast photomicrographs of BrdU-immunopositive cells in the hippocampus. A comparison of BrdU labeling in controls (b, d) and IGF-I-treated animals (c,e) at 1 and 15 d p.i. of BrdU is shown. (For quantification, see Fig. 2.) Scale bar, 100 μm.
Fig. 2.
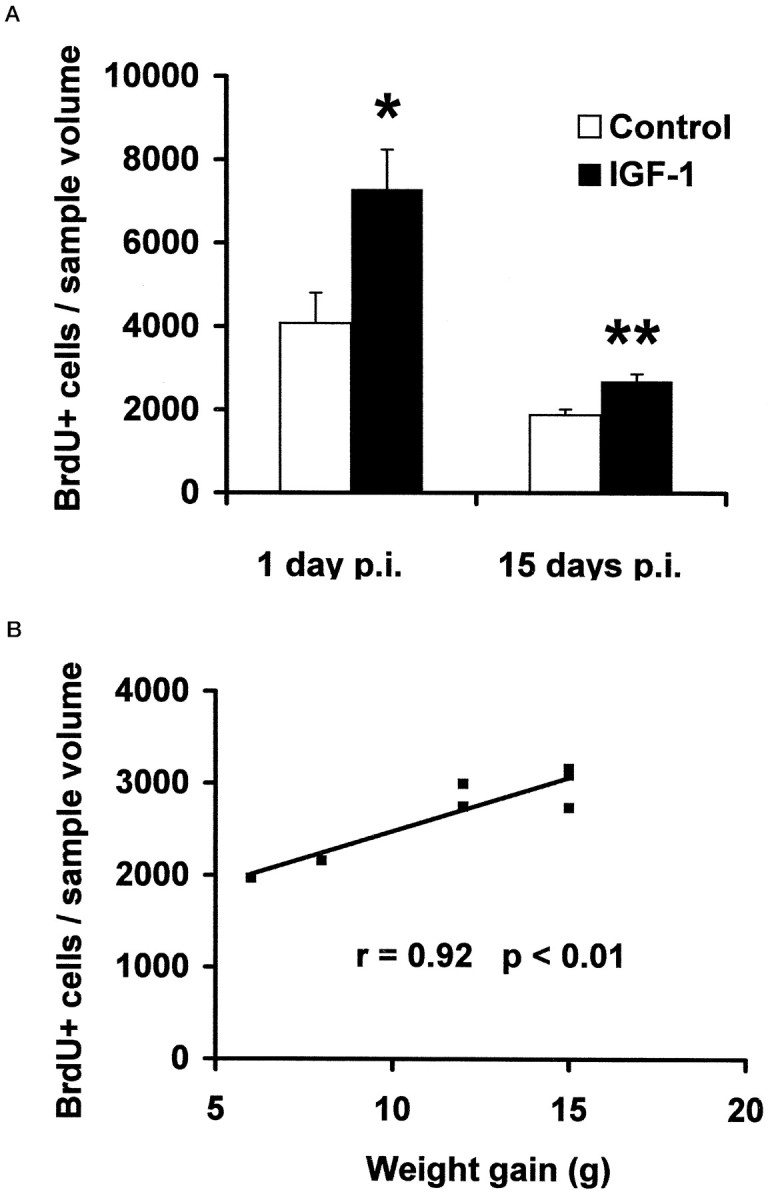
Quantification of BrdU-positive cells in the adult rat hippocampus. A, The density of BrdU-positive cells (cells per cubic millimeter of sample volume) in the GCL 1 and 15 d p.i. was determined stereologically. IGF-I-treated animals (n = 4 at 1 d p.i.; n = 7 at 15 d p.i.) are hx rats given l-thyroxine, cortisol, and IGF-I, as described in Materials and Methods. Controls (n = 5 at 1 d p.i.; n = 8 at 15 d p.i.) are hx rats given l-thyroxine and cortisol only. Means ± SEM are given. *p < 0.05; **p < 0.01. B, Weight gain and the number of BrdU-labeled cells in the GCL at the individual level are compared. There is a correlation between S-IGF-I and BrdU-positive cells after 20 d of IGF-I treatment (15 d p.i.); animals with a relatively low serum IGF-I have relatively low numbers of BrdU-positive cells, and vice versa.
In animals that underwent long-term (20 d) IGF-I treatment (i.e., those evaluated 15 d p.i.), the number of BrdU-immunoreactive cells in the GCL was 2695 ± 175 cells/mm3 in IGF-I-treated animals compared with 1883 ± 138 cells/mm3 in hx controls, which corresponds to a 43 ± 12% increase (p < 0.01) (Fig. 2A). In the hilus, the number of BrdU-immunoreactive cells was 640 ± 56 cells/mm3 in IGF-I-treated animals compared with 306 ± 62 cells/mm3 in hx controls (p < 0.01).
In normal (nonhx) rats, the number of BrdU-immunoreactive cells detected 15 d p.i. was 2416 ± 283 cells/mm3 in the GCL (n = 8), which is close to that in the IGF-I-treated hx rats. This observation supports a role for IGF-I in the regulation of neurogenesis in the GCL. Furthermore, a statistically significant positive correlation between weight gain and the number of BrdU-positive cells in the GCL was observed (r = 0.92; p < 0.01) (Fig. 2B). This observation demonstrates that a stronger biological activity of IGF-I in the periphery also has a greater effect on hippocampal neurogenesis.
The absolute volume of the GCL and of the hilus and the ratio between these volumes were determined in animals that underwent long-term IGF-I treatment. There were no significant differences in the volume of the GCL and of the hilus or between the ratios of these volumes in IGF-I-treated animals versus animals in the hx control group (data not shown).
IGF-I selectively induces neurogenesis
Colocalization of BrdU immunoreactivity with immunoreactivity of the granule cell marker Calbindin D28K (Fig.3a–h) and the astrocyte marker GFAP (Fig. 3i) was investigated to determine the phenotype of progenitor cell progeny in the dentate gyrus after long-term IGF-I therapy (i.e., in animals evaluated 15 d p.i.) compared with that of hx controls. Using confocal microscopy, we were able to detect colocalization of BrdU with either Calbindin D28K or GFAP in the GCL. In IGF-I-treated hx animals, we found an increase in the Calbindin D28K- and BrdU-immunoreactive cell fraction (from 43 ± 1.5 to 54 ± 3.0%) compared with that of hx controls; this corresponds to a 26 ± 9% (p < 0.01) increase (Fig.4). In addition, 17 ± 1.1% of BrdU-positive cells were also GFAP positive compared with 20 ± 1.3% in hx controls (NS). In the hilus, no colocalization of BrdU and Calbindin D28K was found in IGF-I-treated or control animals, whereas BrdU colabeled with GFAP in 27 ± 1.4% of BrdU-positive cells in IGF-I-treated animals compared with 23 ± 2.5% in controls (NS). To confirm that the increase of BrdU-and Calbindin D28K-immunolabeled cells represented new neurons, we performed additional double- and triple-immunolabeling characterizations with other neuronal markers, namely, MAP2 and NeuN. We found that all (100%) of the newly generated BrdU- and Calbindin D28K-positive neurons are MAP2 positive. We also found an increase in the fraction of BrdU- and NeuN-labeled cells (from 54 ± 5.0 to 65 ± 4.0%) in IGF-I-treated animals compared with controls. A two-way ANOVA of these data, using treatment and type of antibodies as factors, shows that IGF-I treatment results in an increase in the fraction of newly generated neurons (p < 0.01). Together, these data show that long-term treatment with IGF-I affected differentiation into neurons but not into astrocytes.
Fig. 3.
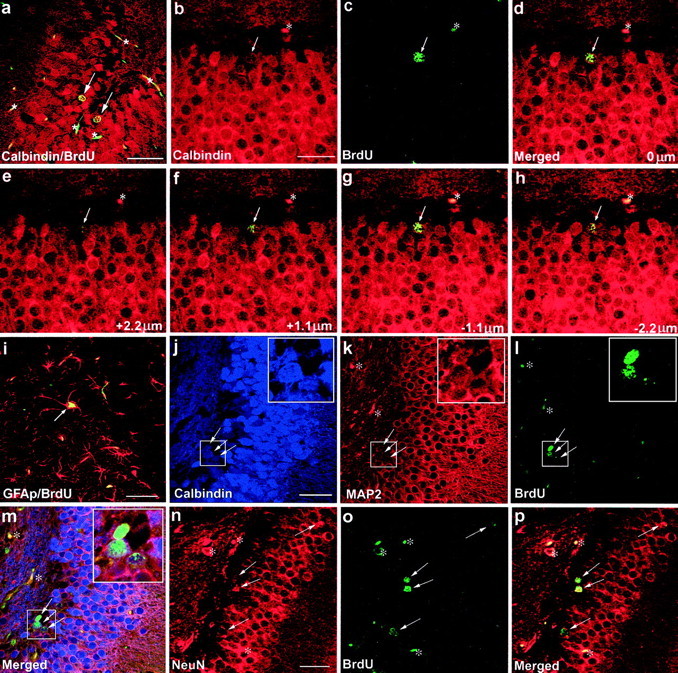
Colocalization of BrdU immunoreactivity (green–yellow in a, c–i, l, m, o, p) with immunoreactivity of the granule cell marker Calbindin D28K (red in a–h;blue in j,m), the astrocyte marker GFAP (red in i), the neuronal marker MAP2 (red in k,m), or the neuronal marker NeuN (red inn,p) (arrows indicate colocalization in a–p). Red blood cells and endothelial cells in several small blood vessels also emit nonspecificgreen and red fluorescence (asterisks in a–p). The specificity of BrdU and Calbindin D28K coexpression in three dimensions is demonstrated by a Z-series of focal planes above (e,f) and below (g,h) the focal plane shown in d(arrows in e–h indicate the same cell as in d). The merged image of b andc is shown in d, the merged image ofj, k, and l is shown inm, and the merged image of n and ois shown in p. Insets of eachboxed area in j–m are magnified 2.5×. Scale bars: a, i, 50 μm; b—h,j—m, n—p, 25 μm.
Fig. 4.

Percentage of the surviving BrdU-positive cells after 20 d of IGF-I treatment that differentiated into either neurons or astrocytes. Colocalization of BrdU immunoreactivity with cell-specific markers, either the granule cell marker Calbindin D28K or the astrocyte marker GFAP, was monitored to determine the phenotype of newborn cells after treatment with IGF-I, when compared with controls in the GCL (n = 5 for controls; n = 7 for IGF-I-treated animals). Means ± SEM are given. **p < 0.01.
To determine the total number of newly generated neurons, we determined the absolute volume of the GCL (data not shown) and multiplied this by the number of cells per sample volume after 20 d of IGF-I treatment (Fig. 2A) and the ratio of Calbindin D28K-/BrdU-positive cells (Fig. 4) for each respective group. We found that there were a total of 1971 ± 154 newly generated neurons in IGF-I-treated hx animals compared with 1106 ± 107 in hx controls. This corresponds to a 78 ± 17% (p < 0.001) increase in the number of newly generated neurons in IGF-I-treated animals.
DISCUSSION
We investigated the effects of IGF-I on the proliferation and differentiation of neural progenitor cells in the adult rat hippocampus. A substantial increase in the total number of BrdU-immunoreactive cells was observed after 6 d of IGF-I administration, demonstrating the proliferative effect of IGF-I. Furthermore, an analysis of the phenotypic distribution after 20 d of IGF-I treatment indicated that the number of newly generated neurons in the GCL increased significantly. To our knowledge, this is the first study showing a selective induction of neurogenesis in the adult mammalian CNS.
It has been demonstrated that the incorporation of BrdU into hippocampal granule cells reflects the number of proliferating progenitor cells. The thymidine analog BrdU is incorporated into DNA and can be detected immunohistochemically in cell progeny (del Rio and Soriano, 1989; Kuhn et al., 1996). Bioavailability of BrdU after injection has been estimated to last ∼2 hr, and BrdU labels DNA only during the S phase, which has been estimated to last ∼8 hr (Nowakowski et al., 1989; Takahashi et al., 1992). Thus, the regimen used in this study is not likely to cause an overestimation of the number of proliferating cells, because one injection cannot label all dividing cells during a 24 hr period. The resulting BrdU-positive cells also include the progeny of labeled cells that divided again after BrdU was discontinued. Although these cells still represent newborn cells, the exact time of cell division is unknown; therefore, injections of BrdU over a 5 d period should reduce the proportion of BrdU-positive cells that divide again compared with a single dose of BrdU. The 5 d treatment period should also reduce the significance of any treatment-induced alterations in the length of any phase of the cell cycle.
Peripheral-derived IGF-I stimulates neurogenesis in the dentate gyrus
The main regulators of circulating levels of IGF-I are nutritional factors and growth hormone (GH) (Humbel, 1990; Thissen et al., 1994). IGF-I is expressed in most tissues in the body, including large organs like skeletal muscle (Isgaard et al., 1988), adipose tissue (Vikman et al., 1991), and liver (Schwander et al., 1983;D'Ercole et al., 1984; Humbel, 1990). However, the liver-derived IGF-I is the principal source of circulating IGF-I that is able to affect the brain (Schwander et al., 1983; Sjögren et al., 1999).
The administration of IGF-I results in weight gain in hypophysectomized animals, and this fact has been used in previous studies to verify the bioactivity of exogenous IGF-I (Schoenle et al., 1982; Fielder et al., 1996). The data showing a correlation between weight gain and the proliferation rate of neural progenitors are noteworthy, because they support the existence of a relationship between the effect of IGF-I on peripheral tissue and that on the CNS and thus suggest that circulating IGF-I is neurotrophic.
Possible mechanisms of IGF-I-induced neurogenesis in the adult dentate gyrus
We show that peripheral administration of IGF-I induces neurogenesis in the adult hippocampus. In our study, the number of newly generated neurons represents a combination of the proliferative rate, the differentiation rate, and net cell survival/apoptosis. This assumption is supported by other studies, in which IGF-I increased the proliferation of embryonic CNS precursors in vitro(DiCicco-Bloom and Black, 1988; Drago et al., 1991; Zackenfels et al., 1995) and in vivo (Ye et al., 1996), and conversely, the number of hippocampal granule neurons was reduced in studies of IGF-I knock-out mice (Beck et al., 1995). These results are consistent with our data on the adult rat, which demonstrate a pronounced increase in the number of BrdU-positive cells after 6 d of IGF-I treatment and suggest that a considerable proportion of the newborn neurons detected after 20 d of IGF-I treatment is attributable to stimulation of progenitor cell proliferation.
We further show that differentiation is also affected by IGF-I, because an increase was seen not only in the absolute number of cells but also in the fraction of newly generated neurons. Similarly, IGF-I stimulates mouse embryonic CNS neural precursors to differentiate into non-GABAergic neurons in vitro (Arsenijevic and Weiss, 1998). In vivo, IGF-I increases neuronal recruitment from the adult songbird subependymal zone (Jiang et al., 1998). Together, our observations and the observations of others support the hypothesis that IGF-I increases neuronal differentiation in both the developing and adult CNS. However, a third possibility—an effect on cell survival/apoptosis—could also be responsible for the increase in neurons observed after IGF-I treatment, because IGF-I supports the survival of neurons in the embryo (Bozyczko-Coyne et al., 1993; Hughes et al., 1993; Lindholm et al., 1996; Blair et al., 1999). The knowledge is still limited about the role of apoptosis in the regulation of adult neurogenesis. Because apoptotic nuclei have been described in the subgranular zone, cell elimination by programmed cell death could explain the decrease in BrdU-positive cells observed 15 d p.i. compared with 1 d p.i. (Bengzon et al., 1997). Quantitatively, the loss of BrdU-positive cells might reflect an overestimation of apoptosis, because BrdU itself could damage some labeled cells. The fact that IGF-I has been shown to attenuate apoptosis in hippocampal neurons after ischemia (Tagami et al., 1997) suggests that the observed increase in the fraction of newborn neurons after IGF-I treatment is in part an effect on cell survival.
Potential of induced neurogenesis in the dentate gyrus
It has been suggested that an increase in the expression of the IGF-I gene or peripherally administered IGF-I plays an important role in the neurotrophic response after injury such as epilepsy or ischemia in the adult rat brain (Gluckman et al., 1992; Guan et al., 1993; Breese et al., 1996; Saatman et al., 1997; Tagami et al., 1997;Liu et al., 1998; Hughes et al., 1999). Several disorders are associated with GCL pathology. Epilepsy is associated with granule cell pathophysiology, including changes in hippocampal calbindin immunoreactivity in rats (Köhr et al., 1991). Similarly, temporal lobe seizures and memory dysfunction are associated with abnormalities in the GCL and hilus (Lowenstein et al., 1992; Beach et al., 1995).
Also, ischemic injury causes neuronal cell loss in the hippocampus and subsequent memory impairment in animals and humans (Zola Morgan et al., 1992; Squire and Zola, 1996). Considering that dentate neurogenesis is increased with enriched environment and learning (Kempermann et al., 1997, 1998; Gould et al., 1999; Nilsson et al., 1999), it is plausible that new neurons actually participate in and mediate this process. The opposite order of events is also very exciting and may also hold true, i.e., improved ability of memory formation after for example drug-induced neurogenesis. This however is a very complex process and remains to be proven. The relationship between memory and adult neurogenesis is supported further by the fact that radiation, which is known to impair memory (Leibel and Sheline, 1987), also inhibits GCL neurogenesis (Parent et al., 1999). Probably IGF-I-stimulated proliferation and neurogenesis in neural progenitors play a role in reducing neuronal loss after hippocampal damage as occurs with epilepsy and ischemia. This issue, however, warrants further investigation.
In conclusion, our in vivo data of selective induction of neurogenesis by peripheral IGF-I on hippocampal progenitors suggest a role for this substance in the generation of new neurons in the adult hippocampus. Additional studies of the effects of IGF-I may therefore yield novel ideas with possible implications for future therapeutic approaches.
Footnotes
This study was supported by grants from the Swedish Medical Research Council (project no. 12X-12535), the Faculty of Medicine of the University of Göteborg, the John and Brit Wennerströms Foundation for Neurological Research, the Rune and Ulla Amlövs Foundation for Neurological and Rheumatological Research, Stiftelsen Göteborgs MS förenings forsknings och byggnadsfond, Stiftelsen Handlanden Hjalmar Svenssons Forskningsfond, the Swedish Society of Medicine, Göteborgs Läkaresällskap, the Edit Jacobssons Foundation, Stiftelsen Thure Carlssons Minne, Stiftelsen Lars Hiertas Minne, the Novo Nordisk Foundation, the Åke Wiberg Foundation, the Magnus Bergvalls Foundation, Axel Linders stiftelse, and the Tore Nilssons Foundation. We appreciate the technical advice of Ann-Marie Alborn and the assistance of Barbro Eriksson with coding slides. We also want to thank Maria Wågberg and Knut Pettersson at Astra Hässle Gothenburg for kindly letting us use their microscope. Recombinant human IGF-I was a kind gift from Genentech (South San Francisco, CA). We are grateful to Dr. Tommy Johnson at the Department of Statistics (Göteborg University, Göteborg, Sweden) for the statistical advice and to Proper English (Alfta, Sweden) for proofreading of this manuscript in English.
Correspondence should be addressed to Dr. Peter S. Eriksson, Institute of Clinical Neuroscience, Göteborg University, Blå Stråket 7, SE-413 45 Göteborg, Sweden. E-mail: per@neuro.gu.se.
REFERENCES
- 1.Altman J, Das GD. Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J Comp Neurol. 1965;124:319–336. doi: 10.1002/cne.901240303. [DOI] [PubMed] [Google Scholar]
- 2.Arsenijevic Y, Weiss S. Insulin-like growth factor-I is a differentiation factor for postmitotic CNS stem cell-derived neuronal precursors: distinct actions from those of brain-derived neurotrophic factor. J Neurosci. 1998;18:2118–2128. doi: 10.1523/JNEUROSCI.18-06-02118.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Beach TG, Woodhurst WB, MacDonald DB, Jones MW. Reactive microglia in hippocampal sclerosis associated with human temporal lobe epilepsy. Neurosci Lett. 1995;191:27–30. doi: 10.1016/0304-3940(94)11548-1. [DOI] [PubMed] [Google Scholar]
- 4.Beck KD, Powell-Braxton L, Widmer HR, Valverde J, Hefti F. IGF-I gene disruption results in reduced brain size, CNS hypomyelination, and loss of hippocampal granule and striatal parvalbumin-containing neurons. Neuron. 1995;14:717–730. doi: 10.1016/0896-6273(95)90216-3. [DOI] [PubMed] [Google Scholar]
- 5.Bengzon J, Kokaia Z, Elmer E, Nanobashvili A, Kokaia M, Lindvall O. Apoptosis and proliferation of dentate gyrus neurons after single and intermittent limbic seizures. Proc Natl Acad Sci USA. 1997;94:10432–10437. doi: 10.1073/pnas.94.19.10432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Blair LAC, Bence Hanulec KK, Mehta S, Franke T, Kaplan D, Marshall J. Akt-dependent potentiation of L channels by insulin-like growth factor-I is required for neuronal survival. J Neurosci. 1999;19:1940–1951. doi: 10.1523/JNEUROSCI.19-06-01940.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bozyczko-Coyne D, Glicksman MA, Prantner JE, McKenna B, Connors T, Friedman C, Dasgupta M, Neff T. IGF-I supports the survival and/or differentiation of multiple types of central nervous system neurons. Ann NY Acad Sci. 1993;692:311–313. doi: 10.1111/j.1749-6632.1993.tb26244.x. [DOI] [PubMed] [Google Scholar]
- 8.Breese CR, D'Costa A, Rollins YD, Adams C, Booze RM, Sonntag WE, Leonard S. Expression of insulin-like growth factor-I (IGF-I) and IGF-binding protein 2 (IGF-BP2) in the hippocampus following cytotoxic lesion of the dentate gyrus. J Comp Neurol. 1996;369:388–404. doi: 10.1002/(SICI)1096-9861(19960603)369:3<388::AID-CNE5>3.0.CO;2-1. [DOI] [PubMed] [Google Scholar]
- 9.Cameron HA, Gould E. Adult neurogenesis is regulated by adrenal steroids in the dentate gyrus. Neuroscience. 1994;61:203–209. doi: 10.1016/0306-4522(94)90224-0. [DOI] [PubMed] [Google Scholar]
- 10.Carson MJ, Behringer RR, Brinster RL, McMorris FA. Insulin-like growth factor-I increases brain growth and central nervous system myelination in transgenic mice. Neuron. 1993;10:729–740. doi: 10.1016/0896-6273(93)90173-o. [DOI] [PubMed] [Google Scholar]
- 11.Cheng CM, Joncas G, Reinhart RR, Farrer R, Quarles R, Janssen J, McDonald MP, Crawley JN, Powell-Braxton L, Bondy CA. Biochemical and morphometric analyses show that myelination in the insulin-like growth factor-I null brain is proportionate to its neuronal composition. J Neurosci. 1998;18:5673–5681. doi: 10.1523/JNEUROSCI.18-15-05673.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Coggeshall RE, Lekan HA. Methods for determining numbers of cells and synapses: a case for more uniform standards of review. J Comp Neurol. 1996;364:6–15. doi: 10.1002/(SICI)1096-9861(19960101)364:1<6::AID-CNE2>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- 13.Craig CG, Tropepe V, Morshead CM, Reynolds BA, Weiss S, Van der Kooy D. In vivo growth factor expansion of endogenous subependymal neural precursor cell populations in the adult mouse brain. J Neurosci. 1996;16:2649–2658. doi: 10.1523/JNEUROSCI.16-08-02649.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.del Rio JA, Soriano E. Immunocytochemical detection of 5′-bromodeoxyuridine incorporation in the central nervous system of the mouse. Brain Res Dev Brain Res. 1989;49:311–317. doi: 10.1016/0165-3806(89)90033-3. [DOI] [PubMed] [Google Scholar]
- 15.Dentremont KD, Ye P, D'Ercole AJ, O' Kusky JR. Increased insulin-like growth factor-I (IGF-I) expression during early postnatal development differentially increases neuron number and growth in medullary nuclei of the mouse. Brain Res Dev Brain Res. 1999;114:135–141. doi: 10.1016/s0165-3806(99)00024-3. [DOI] [PubMed] [Google Scholar]
- 16.D'Ercole AJ, Stiles AD, Underwood LE. Tissue concentrations of somatomedin C: further evidence for multiple sites of synthesis and paracrine or autocrine mechanisms of action. Proc Natl Acad Sci USA. 1984;81:935–939. doi: 10.1073/pnas.81.3.935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.DiCicco-Bloom E, Black IB. Insulin growth factors regulate the mitotic cycle in cultured rat sympathetic neuroblasts. Proc Natl Acad Sci USA. 1988;85:4066–4070. doi: 10.1073/pnas.85.11.4066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Drago J, Murphy M, Carroll SM, Harvey RP, Bartlett PF. Fibroblast growth factor-mediated proliferation of central nervous system precursors depends on endogenous production of insulin-like growth factor-I. Proc Natl Acad Sci USA. 1991;88:2199–2203. doi: 10.1073/pnas.88.6.2199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Duffy KR, Pardridge WM, Rosenfeld RG. Human blood-brain barrier insulin-like growth factor receptor. Metabolism. 1988;37:136–140. doi: 10.1016/s0026-0495(98)90007-5. [DOI] [PubMed] [Google Scholar]
- 20.Eriksson PS, Perfilieva E, Björk-Eriksson T, Alborn A-M, Nordborg C, Peterson DA, Gage FH. Neurogenesis in the adult human hippocampus. Nat Med. 1998;4:1313–1317. doi: 10.1038/3305. [DOI] [PubMed] [Google Scholar]
- 21.Fielder PJ, Mortensen DL, Mallet P, Carlsson B, Baxter RC, Clark RG. Differential long-term effects of insulin-like growth factor-I (IGF-I), growth hormone (GH), and IGF-I plus GH on body growth and IGF binding proteins in hypophysectomized rats. Endocrinology. 1996;137:1913–1920. doi: 10.1210/endo.137.5.8612531. [DOI] [PubMed] [Google Scholar]
- 22.Giacobini MM, Zetterström RH, Young D, Hoffer B, Sara V, Olson L. IGF-I influences olfactory bulb maturation. Evidence from anti-IGF-I antibody treatment of developing grafts in oculo. Brain Res Dev Brain Res. 1995;84:67–76. doi: 10.1016/0165-3806(94)00154-r. [DOI] [PubMed] [Google Scholar]
- 23.Gluckman P, Klempt N, Guan J, Mallard C, Sirimanne E, Dragunow M, Klempt M, Singh K, Williams C, Nikolics K. A role for IGF-I in the rescue of CNS neurons following hypoxic-ischemic injury. Biochem Biophys Res Commun. 1992;182:593–599. doi: 10.1016/0006-291x(92)91774-k. [DOI] [PubMed] [Google Scholar]
- 24.Gould E, McEwen BS, Tanapat P, Galea LA, Fuchs E. Neurogenesis in the dentate gyrus of the adult tree shrew is regulated by psychosocial stress and NMDA receptor activation. J Neurosci. 1997;17:2492–2498. doi: 10.1523/JNEUROSCI.17-07-02492.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gould E, Beylin A, Tanapat P, Reeves A, Shors TJ. Learning enhances adult neurogenesis in the hippocampal formation. Nat Neurosci. 1999;2:260–265. doi: 10.1038/6365. [DOI] [PubMed] [Google Scholar]
- 26.Guan J, Williams C, Gunning M, Mallard C, Gluckman P. The effects of IGF-I treatment after hypoxic-ischemic brain injury in adult rats. J Cereb Blood Flow Metab. 1993;13:609–616. doi: 10.1038/jcbfm.1993.79. [DOI] [PubMed] [Google Scholar]
- 27.Gundersen HJ, Bagger P, Bendtsen TF, Evans SM, Korbo L, Marcussen N, Mρller A, Nielsen K, Nyengaard JR, Pakkenberg B, Sρrensen FB, Versterby A, West MJ. The new stereological tools: disector, fractionator, nucleator and point sampled intercepts and their use in pathological research and diagnosis. APMIS. 1988;96:857–881. doi: 10.1111/j.1699-0463.1988.tb00954.x. [DOI] [PubMed] [Google Scholar]
- 28.Hughes PE, Alexi T, Walton M, Williams CE, Dragunow M, Clark RG, Gluckman PD. Activity and injury-dependent expression of inducible transcription factors, growth factors and apoptosis-related genes within the central nervous system. Prog Neurobiol. 1999;57:421–450. doi: 10.1016/s0301-0082(98)00057-4. [DOI] [PubMed] [Google Scholar]
- 29.Hughes RA, Sendtner M, Thoenen H. Members of several gene families influence survival of rat motoneurons in vitro and in vivo. J Neurosci Res. 1993;36:663–671. doi: 10.1002/jnr.490360607. [DOI] [PubMed] [Google Scholar]
- 30.Humbel RE. Insulin-like growth factors I and II. Eur J Biochem. 1990;190:445–462. doi: 10.1111/j.1432-1033.1990.tb15595.x. [DOI] [PubMed] [Google Scholar]
- 31.Isgaard J, Carlsson L, Isaksson OG, Jansson JO. Pulsatile intravenous growth hormone (GH) infusion to hypophysectomized rats increases insulin-like growth factor-I messenger ribonucleic acid in skeletal tissues more effectively than continuous GH infusion. Endocrinology. 1988;123:2605–2610. doi: 10.1210/endo-123-6-2605. [DOI] [PubMed] [Google Scholar]
- 32.Jiang J, McMurtry J, Niedzwiecki D, Goldman SA. Insulin-like growth factor-I is a radial cell-associated neurotrophin that promotes neuronal recruitment from the adult songbird ependyma/subependyma. J Neurobiol. 1998;36:1–15. doi: 10.1002/(sici)1097-4695(199807)36:1<1::aid-neu1>3.0.co;2-6. [DOI] [PubMed] [Google Scholar]
- 33.Kempermann G, Kuhn HG, Gage FH. More hippocampal neurons in adult mice living in an enriched environment. Nature. 1997;386:493–495. doi: 10.1038/386493a0. [DOI] [PubMed] [Google Scholar]
- 34.Kempermann G, Kuhn HG, Gage FH. Experience-induced neurogenesis in the senescent dentate gyrus. J Neurosci. 1998;18:3206–3212. doi: 10.1523/JNEUROSCI.18-09-03206.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Köhr G, Lambert CE, Mody I. Calbindin-D28K (CaBP) levels and calcium currents in acutely dissociated epileptic neurons. Exp Brain Res. 1991;85:543–551. doi: 10.1007/BF00231738. [DOI] [PubMed] [Google Scholar]
- 36.Kuhn HG, Dickinson-Anson H, Gage FH. Neurogenesis in the dentate gyrus of the adult rat: age-related decrease of neuronal progenitor proliferation. J Neurosci. 1996;16:2027–2033. doi: 10.1523/JNEUROSCI.16-06-02027.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Leibel SA, Sheline GE. Radiation therapy for neoplasms of the brain. J Neurosurg. 1987;66:1–22. doi: 10.3171/jns.1987.66.1.0001. [DOI] [PubMed] [Google Scholar]
- 38.Lesniak MA, Hill JM, Kiess W, Rojeski M, Pert CB, Roth J. Receptors for insulin-like growth factors I and II: autoradiographic localization in rat brain and comparison to receptors for insulin. Endocrinology. 1988;123:2089–2099. doi: 10.1210/endo-123-4-2089. [DOI] [PubMed] [Google Scholar]
- 39.Lindholm D, Carroll P, Tzimagiogis G, Thoenen H. Autocrine-paracrine regulation of hippocampal neuron survival by IGF-I and the neurotrophins BDNF, NT-3 and NT-4. Eur J Neurosci. 1996;8:1452–1460. doi: 10.1111/j.1460-9568.1996.tb01607.x. [DOI] [PubMed] [Google Scholar]
- 40.Liu JL, Solway K, Messing RO, Sharp FR. Increased neurogenesis in the dentate gyrus after transient global ischemia in gerbils. J Neurosci. 1998;18:7768–7778. doi: 10.1523/JNEUROSCI.18-19-07768.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lowenstein DH, Thomas MJ, Smith DH, McIntosh TK. Selective vulnerability of dentate hilar neurons following traumatic brain injury: a potential mechanistic link between head trauma and disorders of the hippocampus. J Neurosci. 1992;12:4846–4853. doi: 10.1523/JNEUROSCI.12-12-04846.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.McEwen BS. Gonadal and adrenal steroids regulate neurochemical and structural plasticity of the hippocampus via cellular mechanisms involving NMDA receptors. Mol Neurobiol. 1996;16:103–116. doi: 10.1007/BF02088170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.McKay R. Stem cells in the central nervous system. Science. 1997;276:66–71. doi: 10.1126/science.276.5309.66. [DOI] [PubMed] [Google Scholar]
- 44.Nilsson M, Perfilieva E, Johansson U, Orwar O, Eriksson P. Enriched environment increases neurogenesis in the adult rat dentate gyrus and improves spatial memory. J Neurobiol. 1999;39:569–578. doi: 10.1002/(sici)1097-4695(19990615)39:4<569::aid-neu10>3.0.co;2-f. [DOI] [PubMed] [Google Scholar]
- 45.Nowakowski RS, Lewin SB, Miller MW. Bromodeoxyuridine immunohistochemical determination of the lengths of the cell cycle and the DNA-synthetic phase for an anatomically defined population. J Neurocytol. 1989;18:311–318. doi: 10.1007/BF01190834. [DOI] [PubMed] [Google Scholar]
- 46.Palmer TD, Ray J, Gage FH. FGF-2-responsive neuronal progenitors reside in proliferative and quiescent regions of the adult rodent brain. Mol Cell Neurosci. 1995;6:474–486. doi: 10.1006/mcne.1995.1035. [DOI] [PubMed] [Google Scholar]
- 47.Parent JM, Tada E, Fike JR, Lowenstein DH. Inhibition of dentate granule cell neurogenesis with brain irradiation does not prevent seizure-induced mossy fiber synaptic reorganization in the rat. J Neurosci. 1999;19:4508–4519. doi: 10.1523/JNEUROSCI.19-11-04508.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Reagan LP, McEwen BS. Controversies surrounding glucocorticoid-mediated cell death in the hippocampus. J Chem Neuroanat. 1997;13:149–167. doi: 10.1016/s0891-0618(97)00031-8. [DOI] [PubMed] [Google Scholar]
- 49.Reinhardt RR, Bondy CA. Insulin-like growth factors cross the blood-brain barrier. Endocrinology. 1994;135:1753–1761. doi: 10.1210/endo.135.5.7525251. [DOI] [PubMed] [Google Scholar]
- 50.Reynolds BA, Weiss S. Generation of neurons and astrocytes from isolated cells of the adult mammalian central nervous system. Science. 1992;255:1707–1710. doi: 10.1126/science.1553558. [DOI] [PubMed] [Google Scholar]
- 51.Richards LJ, Kilpatrick TJ, Bartlett PF. De novo generation of neuronal cells from the adult mouse brain. Proc Natl Acad Sci USA. 1992;89:8591–8595. doi: 10.1073/pnas.89.18.8591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Saatman KE, Contreras PC, Smith DH, Raghupathi R, McDermott KL, Fernandez SC, Sanderson KL, Voddi M, McIntosh TK. Insulin-like growth factor-I (IGF-I) improves both neurological motor and cognitive outcome following experimental brain injury. Exp Neurol. 1997;147:418–427. doi: 10.1006/exnr.1997.6629. [DOI] [PubMed] [Google Scholar]
- 53.Sara VR, Carlsson-Skwirut C. The role of the insulin-like growth factors in the regulation of brain development. Prog Brain Res. 1988;73:87–99. doi: 10.1016/S0079-6123(08)60499-9. [DOI] [PubMed] [Google Scholar]
- 54.Schoenle E, Zapf J, Humbel RE, Froesch ER. Insulin-like growth factor-I stimulates growth in hypophysectomized rats. Nature. 1982;296:252–253. doi: 10.1038/296252a0. [DOI] [PubMed] [Google Scholar]
- 55.Schwander JC, Hauri C, Zapf J, Froesch ER. Synthesis and secretion of insulin-like growth factor and its binding protein by the perfused rat liver: dependence on growth hormone status. Endocrinology. 1983;113:297–305. doi: 10.1210/endo-113-1-297. [DOI] [PubMed] [Google Scholar]
- 56.Sinha AK, Pickard MR, Kim KD, Ahmed MT, al Yatama F, Evans IM, Elkins RP. Perturbation of thyroid hormone homeostasis in the adult and brain function. Acta Med Austriaca. 1994;21:35–43. [PubMed] [Google Scholar]
- 57.Sjöberg A, Oscarsson J, Olofsson SO, Edén S. Insulin-like growth factor-I and growth hormone have different effects on serum lipoproteins and secretion of lipoproteins from cultured rat hepatocytes. Endocrinology. 1994;135:1415–1421. doi: 10.1210/endo.135.4.7925103. [DOI] [PubMed] [Google Scholar]
- 58.Sjögren K, Liu JL, Blad K, Skrtic S, Vidal O, Wallenius V, LeRoith D, Törnell J, Isaksson OG, Jansson JO, Ohlsson C. Liver-derived insulin-like growth factor-I (IGF-I) is the principal source of IGF-I in blood but is not required for postnatal body growth in mice. Proc Natl Acad Sci USA. 1999;96:7088–7092. doi: 10.1073/pnas.96.12.7088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Squire LR, Zola SM. Ischemic brain damage and memory impairment: a commentary. Hippocampus. 1996;6:546–552. doi: 10.1002/(SICI)1098-1063(1996)6:5<546::AID-HIPO7>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]
- 60.Suhonen JO, Peterson DA, Ray J, Gage FH. Differentiation of adult hippocampus-derived progenitors into olfactory neurons in vivo. Nature. 1996;383:624–627. doi: 10.1038/383624a0. [DOI] [PubMed] [Google Scholar]
- 61.Tagami M, Ikeda K, Nara Y, Fujino H, Kubota A, Numano F, Yamori Y. Insulin-like growth factor-I attenuates apoptosis in hippocampal neurons caused by cerebral ischemia and reperfusion in stroke-prone spontaneously hypertensive rats. Lab Invest. 1997;76:613–617. [PubMed] [Google Scholar]
- 62.Takahashi T, Nowakowski RS, Caviness VS., Jr BrdU as an S-phase marker for quantitative studies of cytokinetic behaviour in the murine cerebral ventricular zone. J Neurocytol. 1992;21:185–197. doi: 10.1007/BF01194977. [DOI] [PubMed] [Google Scholar]
- 63.Temple S, Alvarez-Buylla A. Stem cells in the adult mammalian central nervous system. Curr Opin Neurobiol. 1999;9:135–141. doi: 10.1016/s0959-4388(99)80017-8. [DOI] [PubMed] [Google Scholar]
- 64.Thissen JP, Ketelslegers JM, Underwood LE. Nutritional regulation of the insulin-like growth factors. Endocr Rev. 1994;15:80–101. doi: 10.1210/edrv-15-1-80. [DOI] [PubMed] [Google Scholar]
- 65.Vikman K, Isgaard J, Edén S. Growth hormone regulation of insulin-like growth factor-I mRNA in rat adipose tissue and isolated rat adipocytes. J Endocrinol. 1991;131:139–145. doi: 10.1677/joe.0.1310139. [DOI] [PubMed] [Google Scholar]
- 66.West MJ. New stereological methods for counting neurons. Neurobiol Aging. 1993;14:275–285. doi: 10.1016/0197-4580(93)90112-o. [DOI] [PubMed] [Google Scholar]
- 67.Ye P, Xing Y, Dai Z, D'Ercole AJ. In vivo actions of insulin-like growth factor-I (IGF-I) on cerebellum development in transgenic mice: evidence that IGF-I increases proliferation of granule cell progenitors. Brain Res Dev Brain Res. 1996;95:44–54. doi: 10.1016/0165-3806(96)00492-0. [DOI] [PubMed] [Google Scholar]
- 68.Zackenfels K, Oppenheim RW, Rohrer H. Evidence for an important role of IGF-I and IGF-II for the early development of chick sympathetic neurons. Neuron. 1995;14:731–741. doi: 10.1016/0896-6273(95)90217-1. [DOI] [PubMed] [Google Scholar]
- 69.Zola Morgan S, Squire LR, Rempel NL, Clower RP, Amaral DG. Enduring memory impairment in monkeys after ischemic damage to the hippocampus. J Neurosci. 1992;12:2582–2596. doi: 10.1523/JNEUROSCI.12-07-02582.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]