

Abstract
The recent shift in sociopolitical debates and growing liberalization of cannabis use across the globe has raised concern regarding its impact on vulnerable populations, such as pregnant women and adolescents. Epidemiological studies have long demonstrated a relationship between developmental cannabis exposure and later mental health symptoms. This relationship is especially strong in people with particular genetic polymorphisms, suggesting that cannabis use interacts with genotype to increase mental health risk. Seminal animal research directly linked prenatal and adolescent exposure to delta-9-tetrahydrocannabinol, the major psychoactive component of cannabis, with protracted effects on adult neural systems relevant to psychiatric and substance use disorders. In this article, we discuss some recent advances in understanding the long-term molecular, epigenetic, electrophysiological, and behavioral consequences of prenatal, perinatal, and adolescent exposure to cannabis/delta-9-tetrahydrocannabinol. Insights are provided from both animal and human studies, including in vivo neuroimaging strategies.
Keywords: cannabis, adolescence, perinatal, reward, cognition, psychiatric disorders
Introduction
The legalization and decriminalization of cannabis are rapidly expanding worldwide; this has important societal and medical implications in relation to the developing brain. Cannabis is frequently used by pregnant women (Ebrahim and Gfroerer, 2003; Brown et al., 2017; Young-Wolff et al., 2017; Agrawal et al., 2019), due to its anecdotal use as a treatment for morning sickness and its widespread acceptance as a harmless drug (Dickson et al., 2018). Moreover, nursing mothers often use cannabis without realizing that delta-9-tetrahydrocannabinol (THC; the psychoactive cannabinoid) is transferred into breast milk and therefore poses risks to infants (Baker et al., 2018). Indeed, exposure to cannabis during critical windows of neurodevelopment has the potential to alter the endogenous cannabinoid (endocannabinoid [eCB]) system that is critical for hardwiring the developing brain (Harkany et al., 2007; Wu et al., 2011; Torii et al., 2017). This can predispose children to psychiatric disorders, many of which have a developmental etiology. Such risks are not limited to the prenatal and perinatal periods: the onset of cannabis use normally occurs during adolescence, a period of continued brain maturation. Epidemiological data also suggest a greater frequency of use during the adolescent period is associated with psychiatric and substance use vulnerability (Silins et al., 2014; Leadbeater et al., 2019).
An important aspect of the developmental effects of cannabis is that cannabinoid 1 receptors (CB1Rs), which mediate the actions of THC, are predominantly expressed in mesocorticolimbic brain structures during prenatal development as compared to wide expression in the adult brain (Wang et al., 2004, 2006). CB1Rs are targeted by endocannabinoid (eCB) ligands, anandamide (AEA) and 2-arachidonoylglycerol (2-AG), which are synthesized by N-acyl-phosphatidylethanolamine-hydrolyzing phospholipase D (NAPE-PLD) and sn-1-specific diacylglycerol lipase-α (DGLα) or DGLβ and are metabolized by fatty acid amide hydrolase 1 (FAAH) and monoacylglycerol lipase (MGL), respectively (Mechoulam et al., 2014). The eCB ligands have been shown to be synthesized “on demand” in the postsynaptic terminal and transferred in a retrograde manner across the synaptic cleft to directly bind to CB1R at presynaptic terminal where they play a critical role in regulating neurotransmitter release and synaptic plasticity (Katona and Freund, 2008). CB1Rs are expressed at glutamatergic, GABAergic, and dopaminergic terminals, which are highly implicated in various addiction and psychiatric disorders. However, the eCB ligands are also critical for neurodevelopment programming, contributing to, for example, cell fate determination, neuronal proliferation and migration, neurite outgrowth, and synaptogenesis (Keimpema et al., 2013; Maccarrone et al., 2014). Therefore, the direct and indirect regulation of the eCB system by exposure to cannabinoids during development can have important effects on brain and behavior later in life.
The following sections highlight some of our collective research in humans and other animals that, together with yet unpublished data, provide insight about specific molecular, epigenetic, neurophysiological, and neural circuits affected by exposure to cannabis and THC during different developmental stages: in utero, nursing, and adolescence. We particularly focus on effects on adult brain and behavior relevant to psychiatric risk.
Maternal THC biases offspring dopamine system toward behavioral metaplasticity
Maternal cannabis use likely affects fetal development of numerous circuits, but effects on the mesocorticolimbic system may have particularly strong impacts on future psychiatric health of the offspring. The functioning of this system is regulated by dopamine, a neurotransmitter highly implicated in various psychiatric and substance use disorders, and its receptors are involved in developmental differentiation and circuit formation of the forebrain (Frederick and Stanwood, 2009; Conio et al., 2019). Human fetal studies have documented alterations in dopamine D2 receptor (DRD2) gene expression in mesocorticolimbic brain structures after in utero cannabis exposure (Wang et al., 2004, 2006). Animal models have substantiated dopamine receptor alterations in forebrain regions similar to those observed in humans (Jutras-Aswad et al., 2009; DiNieri et al., 2011; Morris et al., 2011; Szutorisz and Hurd, 2018) (Fig. 1). These brain regions play key roles in motivation, emotional regulation, reward, and cognition. Therefore, cannabis-induced perturbation of the dopaminergic system during early development may influence the onset and trajectory of psychiatric disorders (Shrivastava et al., 2014; Bolhuis et al., 2018; Fine et al., 2019).
Figure 1.
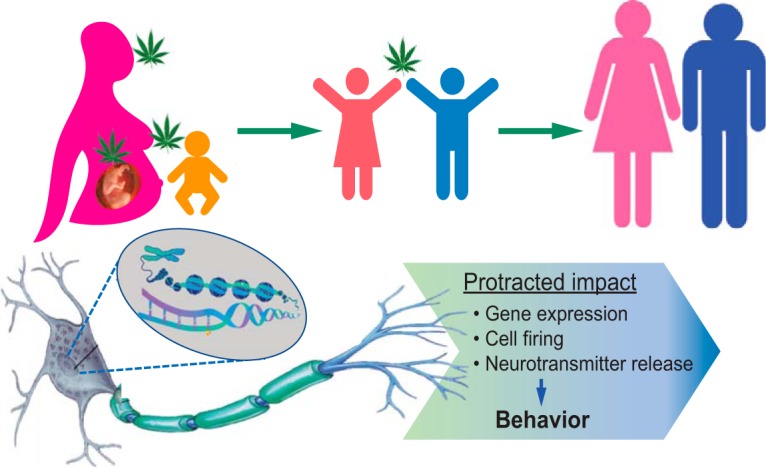
Cannabis exposure during prenatal, perinatal, and adolescent periods of development exerts protracted effects on adult neural processes that underlie behaviors relevant to psychiatric vulnerability. Modified with permission from Szutorisz and Hurd (2018).
In contrast to forebrain alterations, limited information is known about the effects of maternal cannabis/THC exposure on the ventral tegmental area (VTA) from which mesocorticolimbic dopamine forebrain projections arise. But Fernandez-Ruiz et al. (1998) hypothesized that cannabis exposure during perinatal development might confer permanent changes in the function of dopaminergic neurons, making individuals more susceptible to the effects of dopaminergic drugs in adulthood. Recent evidence has verified that juvenile male rats exposed in utero to THC exhibit increased spontaneous activity of dopamine neurons. In addition, in utero THC-exposed rats manifest an imbalance in the ratio between excitatory and inhibitory inputs (E/I ratio) onto dopamine neurons, which might contribute to the enhanced excitability (Frau et al., 2019). The latter might be secondary to changes in eCB signaling: indeed, inhibitory afferents impinging upon dopamine neuron dendrites undergo adaptive changes in CB1R function (Frau et al., 2019). Particularly, THC exposure in utero also induces a marked change in the presynaptic nanoarchitecture of the active zone at inhibitory synapses onto dopamine neurons, which might increase molecular crowding at vesicle release sites and limit GABA diffusion into the synaptic cleft (Glebov et al., 2017), thus contributing to the observed reduced probability of GABA release on dopamine neurons. Indeed, nanoscale superresolution data indicate that the ratio of the presynaptic CB1R and their molecular effectors is shifted in THC-exposed male offspring, thereby contributing to an increased effect of cannabinoids on the probability of GABA release (Frau et al., 2019). These phenomena might account for the E/I ratio imbalance and might depend upon homeostatic regulation of eCB signaling (i.e., prior, tonic, and persistent engagement of the different molecular components of eCB signaling) (Melis et al., 2014) through which the VTA circuit maladaptively changes. Aberrant behavior related to maladaptive alterations of VTA dopamine system function subsequent to in utero THC are manifest in male, but not female, rats as an increased locomotor activity (a sign of psychomotor agitation) and deficits in sensorimotor gating function upon a challenge with THC (Frau et al., 2019).
The results described above suggest that in utero THC exposure increases the general responsiveness of the mesolimbic dopamine system. A hyperdopaminergic phenotype is also suggested by decreased Drd2 expression in VTA targets. This occurs through epigenetic mechanisms in the amygdala, nucleus accumbens (Wang et al., 2004), and ventral striatum (DiNieri et al., 2011) of humans exposed to cannabis in utero. Accordingly, sensitized responses in extracellular dopamine levels in nucleus accumbens (shell subregion), a region central to reward, occur in response to a single exposure to THC in male rat offspring that were exposed to THC prenatally (Frau et al., 2019). These enhanced THC-induced increases in dopamine levels might result from maladaptive changes in the response of VTA dopamine neurons to THC (Frau et al., 2019) and/or from deficits in the homeostatic control of synaptic plasticity by eCB signaling in the nucleus accumbens (Mato et al., 2004). Given the important role played by dopamine D2 receptors and the eCB system in vulnerability to substance use disorders, such changes might contribute to the observation that children/adolescents exposed to THC in the pre/perinatal period are vulnerable to substance use later in life (Porath and Fried, 2005; Day et al., 2006; Sonon et al., 2015). Accordingly, prenatal THC exposure enhances heroin-seeking profiles in adult rat offspring (Spano et al., 2007) and alters the expression of CB1R, dopamine, and glutamate receptor genes in the striatum, resulting in impaired striatal synaptic plasticity (Tortoriello et al., 2014).
Cannabinoid exposure during pregnancy or lactation disrupts the developmental trajectory and cognitive and synaptic function in adult offspring
Longitudinal studies have documented that people exposed perinatally to cannabis have cognitive impairments and deficits in executive functions as adults (Fried and Watkinson, 1990; Day et al., 1994; Fried and Smith, 2001; Smith et al., 2004, 2006). Consistent with this, a wide range of behavioral deficits are evident during adolescence and adulthood in rats exposed to cannabinoids during prenatal or perinatal periods (Mereu et al., 2003; Antonelli et al., 2005; O'Shea et al., 2006; Campolongo et al., 2007; Silva et al., 2012). Many of these cognitive deficits are associated with alterations in hippocampal and cortical excitatory neurotransmission (Mereu et al., 2003; Antonelli et al., 2004; Campolongo et al., 2007; Castaldo et al., 2007, 2010; Ferraro et al., 2009).
In addition to deficits in executive function, animal models of perinatal cannabinoid exposure (PCE; here, reflecting both prenatal and early postnatal exposure) also affects emotional behaviors in rodents. For example, in rats exposed perinatally to cannabinoids, ultrasonic vocalizations are modified in early life (Antonelli et al., 2005; Trezza et al., 2008). At later ages (adolescent and adult), both rats and mice exposed to cannabinoids perinatally show specific deficits in sociobehavioral repertoire (O'Shea et al., 2006; Trezza et al., 2008; Vargish et al., 2017; Bara et al., 2018). Notably, perinatal cannabinoid exposure affects the social repertoire of offspring in a sex-dependent manner: adult male but not female rats exhibit social deficits.
The long-term effects of perinatal cannabinoid exposure might stem from abnormal development of the prefrontal cortex, which mediates cognitive function. The eCB system is a key mediator for the proper maturation of the PFC (Harkany et al., 2007; Wu et al., 2011; Dow-Edwards and Silva, 2017), a cognitive hub whose neurodevelopmental perturbation has been linked to significant long-term behavioral deficits (Goldstein and Volkow, 2011; Schubert et al., 2015; Scheyer et al., 2019). In male PCE offspring, eCB-mediated long-term depression (LTD) in the PFC is disrupted and PFC pyramidal neurons' excitability is heightened (Bara et al., 2018). Interestingly, eCB-LTD is preserved in the nucleus accumbens in both sexes, indicating a particular sensitivity of the PFC to prenatal cannabis insult. Despite sexually divergent outcomes, expression levels of key components of the eCB system are altered in both sexes. Specifically, mRNA levels of the perisynaptic mGlu5 glutamate receptor are decreased in male and female offspring. In female progeny, however, mRNA expression of TRPV1, a synaptic receptor activated by the eCB ligand AEA, and DAGL-α, the synthetizing enzyme of 2-AG, is decreased. Confirming the importance of the eCB system in synaptic and behavioral dysregulation in relation to PCE, positive allosteric modulation of mGlu5 receptors and pharmacological enhancement of AEA levels restore LTD and social interaction in cannabis-exposed males while having no effects in normal animals (Bara et al., 2018). These findings bolster previous results indicating sexual divergence in the long-term functional and behavioral consequences of PCE (e.g., drug self-administration) (Vela et al., 1998; B. González et al., 2003; Economidou et al., 2007; Spano et al., 2007).
Breastfeeding significantly prolongs the potential period of direct transfer of cannabinoids consumed by the mother to the developing offspring during critical stages of neurodevelopment. In rodents, the first 10 postnatal days are equivalent to the third trimester of human pregnancy (Spear and File, 1996). Thus, the transfer of cannabinoids to the developing offspring through breast milk during the preweaning period serves as a model of both lactation-mediated cannabis transfer and cannabinoid exposure during late-gestation neurodevelopment in humans.
Cannabinoid exposure impacts multiple transmitter systems that play important roles during development, including GABAergic transmission. Whereas GABA is the primary inhibitory neurotransmitter in the adult brain, GABAergic transmission is excitatory during early development. GABA switches from being excitatory to inhibitory during the postnatal period as a result of reciprocal changes in expression of the K+/Cl− cotransporters KCC2 and NKCC1. The timing of this shift is a decisive moment in the neurodevelopmental trajectory, and perturbations during this critical period are linked with numerous disorders (Kaila et al., 2014). Exposure to THC or a synthetic cannabinoid during early lactation (postnatal day 1–10) has recently been shown to retard transcriptional upregulation and expression of KCC2 in the PFC, thereby delaying the GABA switch in pups of both sexes via a CB1R-dependent mechanism (Scheyer et al., 2019). The perturbed trajectory was corrected by the NKCC1 antagonist bumetanide and accompanied by alterations in ultrasonic vocalization. These results indicate that the developmental trajectory of GABA in PFC neurons is significantly altered by perinatal lactation exposure to cannabinoids that can impact emotional behavior of the offspring. Thus, while it is challenging to distinguish the effects of in utero exposure from those of breastmilk exposure in most human developmental studies, the results from animal studies provide evidence that even lactation cannabinoid exposure impacts brain and behavioral outcomes later in life.
Role of the endocannabinoid anandamide on adolescent brain development and vulnerability to drug addiction
In addition to the prenatal and perinatal periods, adolescence is also a critical neurodevelopmental period characterized by dynamic changes in the mesolimbic dopamine pathway (Gee et al., 2018) that is fine-tuned by the eCB system (Parsons and Hurd, 2015) to regulate reward-associated behaviors (Solinas et al., 2007). The eCB system reaches peak expression and activity throughout the brain during adolescence (Meyer et al., 2018). This positions the eCB system as a key modulator of adolescent developmental processes involving the mesolimbic reward circuitry and vulnerability to drug addiction (Hurd et al., 2014). Use of cannabis during this period can therefore disrupt normal development, and thus increase vulnerability to drug addiction.
Contributing to the heightened activity of the eCB system during adolescence are peaks in expression of both CB1R and the eCB ligand AEA. AEA activity appears to represent a tonic signal that gates and maintains steady-state (homeostatic) conditions (Hill and Tasker, 2012). In rodents, the highest expression of CB1R is observed at the onset of adolescence, particularly in PFC and striatum, with a subsequent decline approaching adulthood (Rodríguez de Fonseca et al., 1993; Berrendero et al., 1998; Ellgren et al., 2008; Heng et al., 2011). AEA levels in the nucleus accumbens and striatum appear to exhibit unique developmental trajectories, gradually increasing during early life and fluctuating during adolescence (Ellgren et al., 2008). The levels of the eCB AEA are tightly regulated by the catabolic enzyme FAAH, which is highly expressed in brain regions implicated in reward and addiction and exerts widespread modulatory influences on molecular and behavioral responses to drugs of abuse (Parsons and Hurd, 2015). In rodents, complementary changes in FAAH activity occur during adolescence, which may contribute to the observed fluctuations in AEA levels during this time period (Lee and Hill, 2013).
In humans, a common single nucleotide polymorphism (SNP) in the FAAH gene (C385A; rs324420), carried by 38% of individuals of European descent (Genomes Project Consortium et al., 2012), results in the substitution of a proline at position 129 with a threonine residue, which in turn renders the FAAH protein more vulnerable to proteolytic degradation and thus results in increased AEA levels (Chiang et al., 2004). In addition, humans carrying this FAAH SNP display enhanced functional connectivity between PFC and amygdala, as well as changes in fear-related behaviors compared with humans without the SNP. This difference emerges during adolescence (Dincheva et al., 2015; Gee et al., 2016). Human studies demonstrate that, compared with individuals without the SNP, FAAH C385A carriers also display increased striatal activity and increased impulsivity during a reward behavioral task (Hariri et al., 2009), an endophenotype associated with addiction disorders. Moreover, many studies have also linked this FAAH SNP to problem drug use (Chiang et al., 2004; López-Moreno et al., 2012; Parsons and Hurd, 2015). Although this SNP has been associated with increased likelihood to try cannabis (Tyndale et al., 2007), the influence on the progression to cannabis dependence remains unclear because of inconclusive results (Parsons and Hurd, 2015).
Recently, the Lee group has developed a knock-in mouse that biologically recapitulates the FAAH C385A polymorphism and is thus characterized by decreased brain levels of FAAH protein and increased levels of AEA (Dincheva et al., 2015). Furthermore, like humans carrying the FAAH C385A SNP, these mice display enhanced functional connectivity between PFC and amygdala and changes in fear-related behaviors compared with WT mice (Dincheva et al., 2015; Gee et al., 2016). FAAH SNP mice also exhibit greater alcohol intake (Zhou et al., 2016); however, the contribution of this FAAH SNP to other types of addiction, such as cannabis dependence, remains unknown. In ongoing investigations, this humanized eCB SNP mouse model is being used to determine whether genetic variation in FAAH can alter the structure and function of reward pathways during adolescence and in relation to THC exposure. In addition, the effect of altered mesolimbic circuit activity on THC reward-related behaviors is being examined. These animal studies will allow causal and mechanistic studies to determine the contribution of the gene–environment relationship to cannabis sensitivity on discrete neural systems and behavior relevant to the human condition.
Cell type-specific mechanisms of genetic vulnerability to adverse cognitive effects of adolescent cannabis exposure
As noted above, cannabis use during adolescence has been considered a significant environmental factor that might contribute to the development of psychotic disorders and cognitive impairment (Saito et al., 2013). However, as with other environmental adversities, such as infection or social stress, epidemiological data for cannabis are inconclusive, as not all teenagers who smoked cannabis go on to manifest psychoses or exhibit cognitive dysfunction. Individual vulnerability to adolescent cannabis exposure might explain heterogeneity of behavioral and cognitive outcomes in different people. Unfortunately, little is known about the mechanisms of individual susceptibility to adverse effects of cannabis, the so-called gene–environment interaction. Because the molecular mechanisms of gene–environment interactions might differ across cell types, recent studies have begun to identify molecular cascades activated by an environmental stimulus in a cell type-specific manner.
The expression of the CNR1 gene in different brain cells suggests that effects of THC might initiate diverse cell type-specific responses and variable behavioral effects. Indeed, a critical role of CNR1 in astrocytes in mediating cognitive effects of THC has been reported (Han et al., 2012). THC stimulation of astrocyte CNR1 activates inflammatory signaling, glutamate release, abnormal neuronal activities, and poor memory (Chen et al., 2013). These findings indicate that genetic risk factors associated with major psychiatric disorders might differentially influence the cognitive effects of THC depending on whether genetic variants are expressed in astrocytes or neurons. Indeed, such an effect was shown for a rare, highly penetrant mutation, a dominant-negative form of Disrupted in Schizophrenia 1 (DN-DISC1) (Ballinger et al., 2015; Jouroukhin et al., 2018).
Mutation of the DISC1 gene, initially identified in a Scottish family, is produced by a chromosomal abnormality that leads to the protein being truncated. The mutation was associated with several major psychiatric disorders in the family (Millar et al., 2000). Although the DISC1 locus did not meet significance in recent genomewide association studies (Schizophrenia Working Group of the Psychiatric Genomics, 2014), rare mutations of large effect can contribute to behavioral and cognitive abnormalities (Farrell et al., 2015) and can have important roles in mechanistic studies (Geschwind and Flint, 2015). Therefore, DN-DISC1 was selectively expressed in astrocytes or neurons of mice exposed to adolescent THC to determine putative cell-type specific mechanisms of gene–environment interactions that shape individual vulnerability to adverse cognitive effects of cannabis. The results demonstrated that perturbation of expression of Disc1 in astrocytes, but not neurons, exacerbated the effects of adolescent THC exposure on recognition memory assessed in adult mice. The findings further suggested that DN-DISC1 and THC converged and caused synergistic activation of the NF-κB-COX-2 pathway in astrocytes. Activation of this proinflammatory pathway leads to secretion of glutamate by astrocytes and dysfunction of GABAergic neurons in the hippocampus (Jouroukhin et al., 2018). The neuronal and synaptic changes can at least in part explain deficient recognition memory observed in DN-DISC1 mice after adolescent THC. Critically, in contrast to mice that express DN-DISC1 in astrocytes, mice that expressed DN-DISC1 in forebrain neurons and received THC did not demonstrate altered recognition memory but exhibited attenuated fear conditioning (Ballinger et al., 2015). These cell type-specific cognitive outcomes suggest that expression of the same risk factor in different brain cells leads to differential neurobehavioral alterations in mice treated with THC.
Importantly, it was possible to prevent the development of cognitive impairments in DN-DISC1 mice by blocking activation of the proinflammatory pathway identified as a convergent target in this model. These findings demonstrate that astrocyte genetic risk factors can exacerbate cognitive effects of adolescent cannabis use and indicate a putative target for preventive treatment (Jouroukhin et al., 2018).
Neurodevelopmental THC impacts the epigenetic landscape relevant to psychiatric vulnerability
As previously mentioned, the PFC continues to develop throughout adolescence and dynamic fluctuations in components of the eCB system occur throughout adolescent development (Ellgren et al., 2008; Heng et al., 2011; Lee et al., 2013). The critical role of the eCB system in neuronal development and synaptic plasticity (Mackie, 2008) suggests that this eCB-controlled regulation relates to the fine-tuning of PFC circuits established during adolescence.
The prelimbic subregion of the rodent ventromedial PFC mediates cognitive function, decision-making, and emotional regulation, making it a central component of mesolimbic and cortical circuitries (Hempel et al., 2000; Zhang, 2004; Hoover and Vertes, 2007; MacAskill et al., 2012). Disruption of these circuits is implicated in the etiology of multiple psychiatric illnesses in humans. Pyramidal neurons in the prelimbic area, particularly in layer III (Bourgeois et al., 1994), exhibit the most pronounced developmental pruning and the highest rate of spine turnover of PFC subdivisions during adolescence (Pattwell et al., 2016). Research has shown that THC exposure during adolescence in male rats disrupts normal cortical development with structural remodeling of prelimbic pyramidal neurons reflected in the premature pruning of spines and protracted atrophy of distal apical trees (Miller et al., 2019). These findings suggest that adolescent THC exposure reduces the complexity of pyramidal neurons, which might prematurely attenuate the capacity for plasticity in neural circuits central for normal adult behavior. Similar findings were observed in adult female rats: adolescent THC exposure resulted in premature pruning in the PFC as well as impairment of eCB-mediated LTD associated with altered synaptic markers (Rubino et al., 2015). The alterations included glutamatergic receptor remodeling, resulting in a less functional adult PFC that was also accompanied by reduced spatial working memory.
The long-term impact of prenatal and adolescent THC exposure on adult brain and behavior strongly suggests a contribution of epigenetic processes. These processes modulate gene expression without altering the genetic code and are critical aspects of molecular memory. Epigenetic modifications include DNA methylation, post-translational modifications of nucleosomal histones, changes in the structural accessibility of specific chromatin regions, and nucleosomal repositioning, histone replacement, and small RNA molecules that influence protein production (Maze et al., 2014; Cholewa-Waclaw et al., 2016). An important issue germane to epigenetics, the transcriptome, and behavior relates to the discussions above regarding heterogeneity of the brain in regard to cell types, neurochemical phenotype, and anatomical connectivity. Recent research strategies have enabled physical isolation of specific cells to characterize the transcriptome of discrete cell populations. For example, laser capture microdissection in combination with next-generation RNA sequencing was used to isolate mRNA from discrete cortical cellular populations in a region- and layer-specific manner to determine how THC exposure affects the pyramidal cell transcriptome (Miller et al., 2019). This strategy revealed that pyramidal neurons of adult rats with adolescent THC exposure have marked reprogramming of the transcriptome consistent with the morphological disturbances evident in these cells. Surprisingly, only ∼5% of all differentially expressed genes in adulthood overlapped between THC-exposed and normal animals emphasizing that cannabinoid exposure greatly perturbs the developmental trajectory of prelimbic transcriptional maturation. The specific gene networks altered by adolescent THC involved regulation of actin dynamics at excitatory synapses and dendritic spines, which recapitulates on a molecular level the morphological alterations (Miller et al., 2019). Notable genes with enhanced THC sensitivity included pyramidal-enriched Pacsin1, as well as Clu and Snap25, which are implicated in psychiatric diseases, such as schizophrenia and mood disorders (Wang et al., 2015; Pouget et al., 2016; Mahadevan et al., 2017). Intriguingly, the coordinated expression of genes altered by THC across development overlap with PFC gene expression networks dysregulated in humans with schizophrenia (Miller et al., 2019).
In addition to the dysregulation of genes central to cytoskeletal organization and synaptic function, adolescent THC exposure induced a profound reorganization of the epigenetic landscape with developmentally dynamic networks of histone and chromatin modifications (Miller et al., 2019). The strongest functional alterations were associated with Kmt2a (Lysine methyltransferase 2A; also termed mixed lineage leukemia, Mll1) and H3K4me3. This finding is intriguing given that MLL1, an H3K4 methyltransferase, is essential for neurogenesis (Yu et al., 1998; Lim et al., 2009) and PFC synaptic plasticity (Jakovcevski et al., 2015). Moreover, animal models have established an essential role of Kmt2a/Mll1 in cognition and emotion relevant to cortical disturbances in schizophrenia (Huang et al., 2007).
Ongoing studies focused on identifying epigenetic modifications in adults with prenatal THC exposure suggest similar alteration of epigenetic and transcription impairments related to cytoskeletal architecture and synaptic plasticity.
Effects of adolescent onset of regular cannabis use on in vivo brain function and chemistry
In addition to neurobiological insights gleaned from postmortem human fetal brain studies and animal models of perinatal and adolescent cannabis/THC exposure, significant knowledge has been obtained regarding the impact of cannabis use on function and neurochemistry in the living human brain using in vivo neuroimaging. Systematic reviews of the effects of regular cannabis use on human brain function using neuroimaging approaches have not observed consistent patterns of brain function alterations (Quickfall and Crockford, 2006; Chang and Chronicle, 2007; R. González, 2007; Martín-Santos et al., 2010; Bhattacharyya and Sendt, 2012; Bhattacharyya et al., 2012a; Batalla et al., 2013). Methodological factors, such as heterogeneity in study populations, the duration and extent of exposure to cannabis, and the neuroimaging techniques used (e.g., fMRI, PET, single-photon emission CT), may underlie the inconsistent pattern of results. Additionally, inconsistencies between studies may relate to whether neuroimaging data were acquired while participants were performing cognitive tasks or were at rest (Martín-Santos et al., 2010; Batalla et al., 2013). Brain function alterations associated with cannabis use independent of specific cognitive processes of interest can be investigated with a meta-analytic approach to quantitatively synthesize whole-brain fMRI data from studies using a diverse set of cognitive activation paradigms. This strategy revealed a pattern of greater activation in adolescent cannabis users compared with controls in two clusters of brain regions: one with a peak in the right inferior parietal gyrus extending to the superior parietal and angular gyri and a second with peak in the right putamen extending to the striatum and insula (Blest-Hopley et al., 2018). Some of these regions are part of large-scale brain functional networks, such as the default mode and salience networks (Sridharan et al., 2008; Menon, 2011; Andrews-Hanna et al., 2014) that have been implicated in psychotic disorders, such as schizophrenia and in addiction (Radua et al., 2015; Luijten et al., 2017; O'Neill et al., 2018). The angular gyrus in particular is involved in a number of cognitive processes and is thought to serve as a cross-modal hub involved in integrating information from multiple modalities, reorienting attention, and retrieving stored information while giving meaning to new experiences and problem-solving (Seghier, 2013).
While the alteration of brain function observed in adolescent cannabis users may reflect the wide range of cognitive activation paradigms used in various studies, it may also reflect an alteration in the attribution of meaning or significance to internal or external experiences. Alteration in the attribution of meaning or salience has been well recognized with cannabis use and following experimental THC administration (Bhattacharyya et al., 2012b, 2015; Wijayendran et al., 2018). Misattribution of significance or salience to everyday experiences or stimuli has also been suggested to underlie the development of symptoms, such as delusions characteristic of psychotic disorders (Kapur, 2003). The insula, a core node within the salience network, is thought to play a key role in switching cognitive resources between the central executive and the default mode networks, thereby allowing the detection of a salient event (Sridharan et al., 2008; Menon, 2011). Therefore, the meta-analysis results suggest that cannabis use in adolescence may lead to altered functioning of the network-switching process that normally allows efficient allocation of cognitive resources.
Studies have shown that cognitive performance improves in cannabis users following a period of abstinence (Hanson et al., 2010) to levels similar to those of nonusing controls (Schulte et al., 2014), with cognitive deficits possibly only detectable within the first 25 d of abstinence. Nonetheless, meta-analysis of adolescent cannabis users reveals significantly greater activation in cannabis users compared with nonusers in the dorsolateral and ventrolateral prefrontal and posterior parietal cortices, consistent with results from a study in adolescent cannabis users following an abstinence period of ∼5 weeks (Jager et al., 2010). These regions are part of the central executive network known to be involved in higher-order cognitive processes, such as attentional control, executive function, and working memory (Seeley et al., 2007; Sridharan et al., 2008). Furthermore, abstinent adolescent cannabis users display greater activation compared with nonusing adolescents in the cuneus, inferior parietal cortex, and angular gyrus, as well as the visual cortex, all of which are related to the default mode network (Buckner et al., 2008). These results underscore the particular vulnerability of the adolescent brain to the residual effects of long-term cannabis use, even after the drug and its metabolites have been excreted from the body. Ongoing studies in the S.B. group are investigating neurochemical underpinnings of these effects as well as their relationship with functional alterations, especially in light of emerging evidence of acute regionally specific neurochemical modulation of brain glutamate levels by psychoactive cannabinoids, such as THC (Colizzi et al., 2019).
In conclusion, common themes have emerged from diverse seminal papers and a growing number of new research studies providing significant evidence that prenatal, perinatal, and adolescent cannabis exposure can induce a wide array of brain and behavioral alterations in adulthood. This occurs through interfering with multiple neurobiological systems in brain regions involved in psychotic/affective disorders. Whether such risk truly results in psychiatric and substance use disorders will depend on various factors, such as genetics, sex, and environmental conditions that will be better understood as research continues to expand in this field. Nevertheless, governmental and medical policies need to leverage the data already accrued to date to educate the public about the potential health risk to offspring and the long-term effects on adult mental health.
Footnotes
This work was supported by Institut National de la Santé et de la Recherche Médicale Agence National de la Recherche to O.J.M.; National Institutes of Health, National Institute of Drug Abuse DA030359 to Y.L.H., DA044925 to M.M., DA043982 to O.J.M., DA046196 to O.J.M., and DA041208 to M.V.P.; National Institutes of Health, National Institute of Mental Health MH094268 and MH083728 to M.V.P.; National Institutes of Health NS052819 to F.S.L.; National Institute for Health Research 16/126/53 to S.B.; Medical Research Council to S.B.; National Institutes of Health, Mental Health Biomedical Research Centre at South London, and Maudsley National Health Service Foundation Trust and King's College London to S.B.
The authors declare no competing financial interests.
References
- Agrawal A, Grucza RA, Rogers CE (2019) Public health implications of rising marijuana use in pregnancy in an age of increasing legalization: reply. JAMA Pediatr 173:607. 10.1001/jamapediatrics.2019.0618 [DOI] [PubMed] [Google Scholar]
- Andrews-Hanna JR, Smallwood J, Spreng RN (2014) The default network and self-generated thought: component processes, dynamic control, and clinical relevance. Ann N Y Acad Sci 1316:29–52. 10.1111/nyas.12360 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Antonelli T, Tanganelli S, Tomasini MC, Finetti S, Trabace L, Steardo L, Sabino V, Carratu MR, Cuomo V, Ferraro L (2004) Long-term effects on cortical glutamate release induced by prenatal exposure to the cannabinoid receptor agonist (R)-(+)-[2,3-dihydro-5-methyl-3-(4-morpholinyl-methyl)pyrrolo[1,2,3-de]-1,4-benzo xazin-6-yl]-1-naphthalenylmethanone: an in vivo microdialysis study in the awake rat. Neuroscience 124:367–375. 10.1016/j.neuroscience.2003.10.034 [DOI] [PubMed] [Google Scholar]
- Antonelli T, Tomasini MC, Tattoli M, Cassano T, Tanganelli S, Finetti S, Mazzoni E, Trabace L, Steardo L, Cuomo V, Ferraro L (2005) Prenatal exposure to the CB1 receptor agonist WIN 55,212–2 causes learning disruption associated with impaired cortical NMDA receptor function and emotional reactivity changes in rat offspring. Cereb Cortex 15:2013–2020. 10.1093/cercor/bhi076 [DOI] [PubMed] [Google Scholar]
- Baker T, Datta P, Rewers-Felkins K, Thompson H, Kallem RR, Hale TW (2018) Transfer of inhaled cannabis into human breast milk. Obstet Gynecol 131:783–788. 10.1097/AOG.0000000000002575 [DOI] [PubMed] [Google Scholar]
- Ballinger MD, Saito A, Abazyan B, Taniguchi Y, Huang CH, Ito K, Zhu X, Segal H, Jaaro-Peled H, Sawa A, Mackie K, Pletnikov MV, Kamiya A (2015) Adolescent cannabis exposure interacts with mutant DISC1 to produce impaired adult emotional memory. Neurobiol Dis 82:176–184. 10.1016/j.nbd.2015.06.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bara A, Manduca A, Bernabeu A, Borsoi M, Serviado M, Lassalle O, Murphy M, Wager-Miller J, Mackie K, Pelissier-Alicot AL, Trezza V, Manzoni OJ (2018) Sex-dependent effects of in utero cannabinoid exposure on cortical function. Elife 7:e36234. 10.7554/eLife.36234 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Batalla A, Bhattacharyya S, Yücel M, Fusar-Poli P, Crippa JA, Nogué S, Torrens M, Pujol J, Farré M, Martín-Santos R (2013) Structural and functional imaging studies in chronic cannabis users: a systematic review of adolescent and adult findings. PLoS One 8:e55821. 10.1371/journal.pone.0055821 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berrendero F, García-Gil L, Hernández ML, Romero J, Cebeira M, de Miguel R, Ramos JA, Fernández-Ruiz JJ (1998) Localization of mRNA expression and activation of signal transduction mechanisms for cannabinoid receptor in rat brain during fetal development. Development 125:3179–3188. [DOI] [PubMed] [Google Scholar]
- Bhattacharyya S, Sendt KV (2012) Neuroimaging evidence for cannabinoid modulation of cognition and affect in man. Front Behav Neurosci 6:22. 10.3389/fnbeh.2012.00022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhattacharyya S, Atakan Z, Martín-Santos R, Crippa JA, McGuire PK (2012a) Neural mechanisms for the cannabinoid modulation of cognition and affect in man: a critical review of neuroimaging studies. Curr Pharm Des 18:5045–5054. 10.2174/138161212802884636 [DOI] [PubMed] [Google Scholar]
- Bhattacharyya S, Crippa JA, Allen P, Martín-Santos R, Borgwardt S, Fusar-Poli P, Rubia K, Kambeitz J, O'Carroll C, Seal ML, Giampietro V, Brammer M, Zuardi AW, Atakan Z, McGuire PK (2012b) Induction of psychosis by Δ9-tetrahydrocannabinol reflects modulation of prefrontal and striatal function during attentional salience processing. Arch Gen Psychiatry 69:27–36. 10.1001/archgenpsychiatry.2011.161 [DOI] [PubMed] [Google Scholar]
- Bhattacharyya S, Atakan Z, Martín-Santos R, Crippa JA, Kambeitz J, Malhi S, Giampietro V, Williams S, Brammer M, Rubia K, Collier DA, McGuire PK (2015) Impairment of inhibitory control processing related to acute psychotomimetic effects of cannabis. Eur Neuropsychopharmacol 25:26–37. 10.1016/j.euroneuro.2014.11.018 [DOI] [PubMed] [Google Scholar]
- Blest-Hopley G, Giampietro V, Bhattacharyya S (2018) Residual effects of cannabis use in adolescent and adult brains: a meta-analysis of fMRI studies. Neurosci Biobehav Rev 88:26–41. 10.1016/j.neubiorev.2018.03.008 [DOI] [PubMed] [Google Scholar]
- Bolhuis K, Kushner SA, Yalniz S, Hillegers MH, Jaddoe VW, Tiemeier H, El Marroun H (2018) Maternal and paternal cannabis use during pregnancy and the risk of psychotic-like experiences in the offspring. Schizophr Res 202:322–327. 10.1016/j.schres.2018.06.067 [DOI] [PubMed] [Google Scholar]
- Bourgeois JP, Goldman-Rakic PS, Rakic P (1994) Synaptogenesis in the prefrontal cortex of rhesus monkeys. Cereb Cortex 4:78–96. 10.1093/cercor/4.1.78 [DOI] [PubMed] [Google Scholar]
- Brown QL, Sarvet AL, Shmulewitz D, Martins SS, Wall MM, Hasin DS (2017) Trends in marijuana use among pregnant and nonpregnant reproductive-aged women, 2002–2014. JAMA 317:207–209. 10.1001/jama.2016.17383 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buckner RL, Andrews-Hanna JR, Schacter DL (2008) The brain's default network: anatomy, function, and relevance to disease. Ann N Y Acad Sci 1124:1–38. 10.1196/annals.1440.011 [DOI] [PubMed] [Google Scholar]
- Campolongo P, Trezza V, Cassano T, Gaetani S, Morgese MG, Ubaldi M, Soverchia L, Antonelli T, Ferraro L, Massi M, Ciccocioppo R, Cuomo V (2007) Perinatal exposure to delta-9-tetrahydrocannabinol causes enduring cognitive deficits associated with alteration of cortical gene expression and neurotransmission in rats. Addict Biol 12:485–495. 10.1111/j.1369-1600.2007.00074.x [DOI] [PubMed] [Google Scholar]
- Castaldo P, Magi S, Gaetani S, Cassano T, Ferraro L, Antonelli T, Amoroso S, Cuomo V (2007) Prenatal exposure to the cannabinoid receptor agonist WIN 55,212–2 increases glutamate uptake through overexpression of GLT1 and EAAC1 glutamate transporter subtypes in rat frontal cerebral cortex. Neuropharmacology 53:369–378. 10.1016/j.neuropharm.2007.05.019 [DOI] [PubMed] [Google Scholar]
- Castaldo P, Magi S, Cataldi M, Arcangeli S, Lariccia V, Nasti AA, Ferraro L, Tomasini MC, Antonelli T, Cassano T, Cuomo V, Amoroso S (2010) Altered regulation of glutamate release and decreased functional activity and expression of GLT1 and GLAST glutamate transporters in the hippocampus of adolescent rats perinatally exposed to Delta(9)-THC. Pharmacol Res 61:334–341. 10.1016/j.phrs.2009.11.008 [DOI] [PubMed] [Google Scholar]
- Chang L, Chronicle EP (2007) Functional imaging studies in cannabis users. Neuroscientist 13:422–432. 10.1177/1073858406296601 [DOI] [PubMed] [Google Scholar]
- Chen R, Zhang J, Fan N, Teng ZQ, Wu Y, Yang H, Tang YP, Sun H, Song Y, Chen C (2013) Delta9-THC-caused synaptic and memory impairments are mediated through COX-2 signaling. Cell 155:1154–1165. 10.1016/j.cell.2013.10.042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiang KP, Gerber AL, Sipe JC, Cravatt BF (2004) Reduced cellular expression and activity of the P129T mutant of human fatty acid amide hydrolase: evidence for a link between defects in the endocannabinoid system and problem drug use. Hum Mol Genet 13:2113–2119. 10.1093/hmg/ddh216 [DOI] [PubMed] [Google Scholar]
- Cholewa-Waclaw J, Bird A, von Schimmelmann M, Schaefer A, Yu H, Song H, Madabhushi R, Tsai LH (2016) The role of epigenetic mechanisms in the regulation of gene expression in the nervous system. J Neurosci 36:11427–11434. 10.1523/JNEUROSCI.2492-16.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colizzi M, Weltens N, McGuire P, Lythgoe D, Williams S, Van Oudenhove L, Bhattacharyya S (2019) Delta-9-tetrahydrocannabinol increases striatal glutamate levels in healthy individuals: implications for psychosis. Mol Psychiatry. Advance online publication. Retrieved Feb 15, 2019. doi: 10.1038/s41380-019-0374-8. 10.1038/s41380-019-0374-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conio B, Martino M, Magioncalda P, Escelsior A, Inglese M, Amore M, Northoff G (2019) Opposite effects of dopamine and serotonin on resting-state networks: review and implications for psychiatric disorders. Mol Psychiatry. Advance online publication. Retrieved Apr 5, 2019. doi: 10.1038/s41380-019-0406-4. 10.1038/s41380-019-0406-4 [DOI] [PubMed] [Google Scholar]
- Day NL, Richardson GA, Goldschmidt L, Robles N, Taylor PM, Stoffer DS, Cornelius MD, Geva D (1994) Effect of prenatal marijuana exposure on the cognitive development of offspring at age three. Neurotoxicol Teratol 16:169–175. 10.1016/0892-0362(94)90114-7 [DOI] [PubMed] [Google Scholar]
- Day NL, Goldschmidt L, Thomas CA (2006) Prenatal marijuana exposure contributes to the prediction of marijuana use at age 14. Addiction 101:1313–1322. 10.1111/j.1360-0443.2006.01523.x [DOI] [PubMed] [Google Scholar]
- Dickson B, Mansfield C, Guiahi M, Allshouse AA, Borgelt LM, Sheeder J, Silver RM, Metz TD (2018) Recommendations from cannabis dispensaries about first-trimester cannabis use. Obstet Gynecol 131:1031–1038. 10.1097/AOG.0000000000002619 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dincheva I, Drysdale AT, Hartley CA, Johnson DC, Jing D, King EC, Ra S, Gray JM, Yang R, DeGruccio AM, Huang C, Cravatt BF, Glatt CE, Hill MN, Casey BJ, Lee FS (2015) FAAH genetic variation enhances fronto-amygdala function in mouse and human. Nat Commun 6:6395. 10.1038/ncomms7395 [DOI] [PMC free article] [PubMed] [Google Scholar]
- DiNieri JA, Wang X, Szutorisz H, Spano SM, Kaur J, Casaccia P, Dow-Edwards D, Hurd YL (2011) Maternal cannabis use alters ventral striatal dopamine D2 gene regulation in the offspring. Biol Psychiatry 70:763–769. 10.1016/j.biopsych.2011.06.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dow-Edwards D, Silva L (2017) Endocannabinoids in brain plasticity: cortical maturation, HPA axis function and behavior. Brain Res 1654:157–164. 10.1016/j.brainres.2016.08.037 [DOI] [PubMed] [Google Scholar]
- Ebrahim SH, Gfroerer J (2003) Pregnancy-related substance use in the united states during 1996–1998. Obstet Gynecol 101:374–379. 10.1016/S0029-7844(02)02588-7 [DOI] [PubMed] [Google Scholar]
- Economidou D, Mattioli L, Ubaldi M, Lourdusamy A, Soverchia L, Hardiman G, Campolongo P, Cuomo V, Ciccocioppo R (2007) Role of cannabinoidergic mechanisms in ethanol self-administration and ethanol seeking in rat adult offspring following perinatal exposure to Delta9-tetrahydrocannabinol. Toxicol Appl Pharmacol 223:73–85. 10.1016/j.taap.2007.05.008 [DOI] [PubMed] [Google Scholar]
- Ellgren M, Artmann A, Tkalych O, Gupta A, Hansen HS, Hansen SH, Devi LA, Hurd YL (2008) Dynamic changes of the endogenous cannabinoid and opioid mesocorticolimbic systems during adolescence: THC effects. Eur Neuropsychopharmacol 18:826–834. 10.1016/j.euroneuro.2008.06.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farrell MS, Werge T, Sklar P, Owen MJ, Ophoff RA, O'Donovan MC, Corvin A, Cichon S, Sullivan PF (2015) Evaluating historical candidate genes for schizophrenia. Mol Psychiatry 20:555–562. 10.1038/mp.2015.16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandez-Ruiz J, Bonnin A, De Miguel R, Castro JG, Ramos JA (1998) Perinatal exposure to marihuana or its main psychoactive constituent, delta9-tetrahydrocannabinol, affects the development of brain dopaminergic neurons. Arq Med 12:67–77. [Google Scholar]
- Ferraro L, Tomasini MC, Beggiato S, Gaetani S, Cassano T, Cuomo V, Amoroso S, Tanganelli S, Antonelli T (2009) Short- and long-term consequences of prenatal exposure to the cannabinoid agonist WIN55,212–2 on rat glutamate transmission and cognitive functions. J Neural Transm (Vienna) 116:1017–1027. 10.1007/s00702-009-0230-0 [DOI] [PubMed] [Google Scholar]
- Fine JD, Moreau AL, Karcher NR, Agrawal A, Rogers CE, Barch DM, Bogdan R (2019) Association of prenatal cannabis exposure with psychosis proneness among children in the adolescent brain cognitive development (ABCD) Study. JAMA Psychiatry 76:762–764. 10.1001/jamapsychiatry.2019.0076 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frau R, Miczán V, Traccis F, Aroni S, Pongor CI, Saba P, Serra V, Sagheddu C, Fanni S, Congiu M, Devoto P, Cheer JF, Katona I, Melis M (2019) Prenatal THC produces a hyperdopaminergic phenotype accompanied by maladaptive behavioral susceptibility. Nat Neurosci, in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frederick AL, Stanwood GD (2009) Drugs, biogenic amine targets and the developing brain. Dev Neurosci 31:7–22. 10.1159/000207490 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fried PA, Smith AM (2001) A literature review of the consequences of prenatal marihuana exposure: an emerging theme of a deficiency in aspects of executive function. Neurotoxicol Teratol 23:1–11. 10.1016/S0892-0362(00)00119-7 [DOI] [PubMed] [Google Scholar]
- Fried PA, Watkinson B (1990) 36- and 48-month neurobehavioral follow-up of children prenatally exposed to marijuana, cigarettes, and alcohol. J Dev Behav Pediatr 11:49–58. [PubMed] [Google Scholar]
- Gee DG, Fetcho RN, Jing D, Li A, Glatt CE, Drysdale AT, Cohen AO, Dellarco DV, Yang RR, Dale AM, Jernigan TL, Lee FS, Casey BJ, Consortium P (2016) Individual differences in frontolimbic circuitry and anxiety emerge with adolescent changes in endocannabinoid signaling across species. Proc Natl Acad Sci U S A 113:4500–4505. 10.1073/pnas.1600013113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gee DG, Bath KG, Johnson CM, Meyer HC, Murty VP, van den Bos W, Hartley CA (2018) Neurocognitive development of motivated behavior: dynamic changes across childhood and adolescence. J Neurosci 38:9433–9445. 10.1523/JNEUROSCI.1674-18.2018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Genomes Project Consortium, Abecasis GR, Auton A, Brooks LD, DePristo MA, Durbin RM, Handsaker RE, Kang HM, Marth GT, McVean GA (2012) An integrated map of genetic variation from 1,092 human genomes. Nature 491:56–65. 10.1038/nature11632 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geschwind DH, Flint J (2015) Genetics and genomics of psychiatric disease. Science 349:1489–1494. 10.1126/science.aaa8954 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glebov OO, Jackson RE, Winterflood CM, Owen DM, Barker EA, Doherty P, Ewers H, Burrone J (2017) Nanoscale structural plasticity of the active zone matrix modulates presynaptic function. Cell Rep 18:2715–2728. 10.1016/j.celrep.2017.02.064 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldstein RZ, Volkow ND (2011) Dysfunction of the prefrontal cortex in addiction: neuroimaging findings and clinical implications. Nat Rev Neurosci 12:652–669. 10.1038/nrn3119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- González B, de Miguel R, Martín S, Pérez-Rosado A, Romero J, García-Lecumberri C, Fernández-Ruiz J, Ramos JA, Ambrosio E (2003) Effects of perinatal exposure to delta 9-tetrahydrocannabinol on operant morphine-reinforced behavior. Pharmacol Biochem Behav 75:577–584. 10.1016/S0091-3057(03)00115-1 [DOI] [PubMed] [Google Scholar]
- González R. (2007) Acute and non-acute effects of cannabis on brain functioning and neuropsychological performance. Neuropsychol Rev 17:347–361. 10.1007/s11065-007-9036-8 [DOI] [PubMed] [Google Scholar]
- Han J, Kesner P, Metna-Laurent M, Duan T, Xu L, Georges F, Koehl M, Abrous DN, Mendizabal-Zubiaga J, Grandes P, Liu Q, Bai G, Wang W, Xiong L, Ren W, Marsicano G, Zhang X (2012) Acute cannabinoids impair working memory through astroglial CB1 receptor modulation of hippocampal LTD. Cell 148:1039–1050. 10.1016/j.cell.2012.01.037 [DOI] [PubMed] [Google Scholar]
- Hanson KL, Winward JL, Schweinsburg AD, Medina KL, Brown SA, Tapert SF (2010) Longitudinal study of cognition among adolescent marijuana users over three weeks of abstinence. Addict Behav 35:970–976. 10.1016/j.addbeh.2010.06.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hariri AR, Gorka A, Hyde LW, Kimak M, Halder I, Ducci F, Ferrell RE, Goldman D, Manuck SB (2009) Divergent effects of genetic variation in endocannabinoid signaling on human threat- and reward-related brain function. Biol Psychiatry 66:9–16. 10.1016/j.biopsych.2008.10.047 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harkany T, Guzmán M, Galve-Roperh I, Berghuis P, Devi LA, Mackie K (2007) The emerging functions of endocannabinoid signaling during CNS development. Trends Pharmacol Sci 28:83–92. 10.1016/j.tips.2006.12.004 [DOI] [PubMed] [Google Scholar]
- Hempel CM, Hartman KH, Wang XJ, Turrigiano GG, Nelson SB (2000) Multiple forms of short-term plasticity at excitatory synapses in rat medial prefrontal cortex. J Neurophysiol 83:3031–3041. 10.1152/jn.2000.83.5.3031 [DOI] [PubMed] [Google Scholar]
- Heng L, Beverley JA, Steiner H, Tseng KY (2011) Differential developmental trajectories for CB1 cannabinoid receptor expression in limbic/associative and sensorimotor cortical areas. Synapse 65:278–286. 10.1002/syn.20844 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill MN, Tasker JG (2012) Endocannabinoid signaling, glucocorticoid-mediated negative feedback, and regulation of the hypothalamic-pituitary-adrenal axis. Neuroscience 204:5–16. 10.1016/j.neuroscience.2011.12.030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoover WB, Vertes RP (2007) Anatomical analysis of afferent projections to the medial prefrontal cortex in the rat. Brain Struct Funct 212:149–179. 10.1007/s00429-007-0150-4 [DOI] [PubMed] [Google Scholar]
- Huang HS, Matevossian A, Whittle C, Kim SY, Schumacher A, Baker SP, Akbarian S (2007) Prefrontal dysfunction in schizophrenia involves mixed-lineage leukemia 1-regulated histone methylation at GABAergic gene promoters. J Neurosci 27:11254–11262. 10.1523/JNEUROSCI.3272-07.2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hurd YL, Michaelides M, Miller ML, Jutras-Aswad D (2014) Trajectory of adolescent cannabis use on addiction vulnerability. Neuropharmacology 76:416–424. 10.1016/j.neuropharm.2013.07.028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jager G, Block RI, Luijten M, Ramsey NF (2010) Cannabis use and memory brain function in adolescent boys: a cross-sectional multicenter functional magnetic resonance imaging study. J Am Acad Child Adolesc Psychiatry 49:561–572.e1–3. 10.1016/j.jaac.2010.02.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jakovcevski M, Ruan H, Shen EY, Dincer A, Javidfar B, Ma Q, Peter CJ, Cheung I, Mitchell AC, Jiang Y, Lin CL, Pothula V, Stewart AF, Ernst P, Yao WD, Akbarian S (2015) Neuronal Kmt2a/Mll1 histone methyltransferase is essential for prefrontal synaptic plasticity and working memory. J Neurosci 35:5097–5108. 10.1523/JNEUROSCI.3004-14.2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jouroukhin Y, Kageyama Y, Misheneva V, Shevelkin A, Andrabi S, Prandovszky E, Yolken RH, Dawson VL, Dawson TM, Aja S, Sesaki H, Pletnikov MV (2018) DISC1 regulates lactate metabolism in astrocytes: implications for psychiatric disorders. Transl Psychiatry 8:76. 10.1038/s41398-018-0123-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jutras-Aswad D, DiNieri JA, Harkany T, Hurd YL (2009) Neurobiological consequences of maternal cannabis on human fetal development and its neuropsychiatric outcome. Eur Arch Psychiatry Clin Neurosci 259:395–412. 10.1007/s00406-009-0027-z [DOI] [PubMed] [Google Scholar]
- Kaila K, Price TJ, Payne JA, Puskarjov M, Voipio J (2014) Cation-chloride cotransporters in neuronal development, plasticity and disease. Nat Rev Neurosci 15:637–654. 10.1038/nrn3819 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kapur S. (2003) Psychosis as a state of aberrant salience: a framework linking biology, phenomenology, and pharmacology in schizophrenia. Am J Psychiatry 160:13–23. 10.1176/appi.ajp.160.1.13 [DOI] [PubMed] [Google Scholar]
- Katona I, Freund TF (2008) Endocannabinoid signaling as a synaptic circuit breaker in neurological disease. Nat Med 14:923–930. 10.1038/nm.f.1869 [DOI] [PubMed] [Google Scholar]
- Keimpema E, Calvigioni D, Harkany T (2013) Endocannabinoid signals in the developmental programming of delayed-onset neuropsychiatric and metabolic illnesses. Biochem Soc Trans 41:1569–1576. 10.1042/BST20130117 [DOI] [PubMed] [Google Scholar]
- Leadbeater BJ, Ames ME, Linden-Carmichael AN (2019) Age-varying effects of cannabis use frequency and disorder on symptoms of psychosis, depression and anxiety in adolescents and adults. Addiction 114:278–293. 10.1111/add.14459 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee TT, Hill MN (2013) Age of stress exposure modulates the immediate and sustained effects of repeated stress on corticolimbic cannabinoid CB(1) receptor binding in male rats. Neuroscience 249:106–114. 10.1016/j.neuroscience.2012.11.017 [DOI] [PubMed] [Google Scholar]
- Lee TT, Hill MN, Hillard CJ, Gorzalka BB (2013) Temporal changes in N-acylethanolamine content and metabolism throughout the peri-adolescent period. Synapse 67:4–10. 10.1002/syn.21609 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim DA, Huang YC, Swigut T, Mirick AL, Garcia-Verdugo JM, Wysocka J, Ernst P, Alvarez-Buylla A (2009) Chromatin remodelling factor Mll1 is essential for neurogenesis from postnatal neural stem cells. Nature 458:529–533. 10.1038/nature07726 [DOI] [PMC free article] [PubMed] [Google Scholar]
- López-Moreno JA, Echeverry-Alzate V, Bühler KM (2012) The genetic basis of the endocannabinoid system and drug addiction in humans. J Psychopharmacol 26:133–143. 10.1177/0269881111416689 [DOI] [PubMed] [Google Scholar]
- Luijten M, Schellekens AF, Kühn S, Machielse MW, Sescousse G (2017) Disruption of reward processing in addiction: an image-based meta-analysis of functional magnetic resonance imaging studies. JAMA Psychiatry 74:387–398. 10.1001/jamapsychiatry.2016.3084 [DOI] [PubMed] [Google Scholar]
- MacAskill AF, Little JP, Cassel JM, Carter AG (2012) Subcellular connectivity underlies pathway-specific signaling in the nucleus accumbens. Nat Neurosci 15:1624–1626. 10.1038/nn.3254 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maccarrone M, Guzmán M, Mackie K, Doherty P, Harkany T (2014) Programming of neural cells by (endo)cannabinoids: from physiological rules to emerging therapies. Nat Rev Neurosci 15:786–801. 10.1038/nrn3846 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackie K. (2008) Signaling via CNS cannabinoid receptors. Mol Cell Endocrinol 286:S60–S65. 10.1016/j.mce.2008.01.022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mahadevan V, Khademullah CS, Dargaei Z, Chevrier J, Uvarov P, Kwan J, Bagshaw RD, Pawson T, Emili A, De Koninck Y, Anggono V, Airaksinen M, Woodin MA (2017) Native KCC2 interactome reveals PACSIN1 as a critical regulator of synaptic inhibition. Elife 6:e2827. 10.7554/eLife.28270 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martín-Santos R, Fagundo AB, Crippa JA, Atakan Z, Bhattacharyya S, Allen P, Fusar-Poli P, Borgwardt S, Seal M, Busatto GF, McGuire P (2010) Neuroimaging in cannabis use: a systematic review of the literature. Psychol Med 40:383–398. 10.1017/S0033291709990729 [DOI] [PubMed] [Google Scholar]
- Mato S, Chevaleyre V, Robbe D, Pazos A, Castillo PE, Manzoni OJ (2004) A single in-vivo exposure to delta 9THC blocks endocannabinoid-mediated synaptic plasticity. Nat Neurosci 7:585–586. 10.1038/nn1251 [DOI] [PubMed] [Google Scholar]
- Maze I, Noh KM, Soshnev AA, Allis CD (2014) Every amino acid matters: essential contributions of histone variants to mammalian development and disease. Nat Rev Genet 15:259–271. 10.1038/nrg3673 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mechoulam R, Hanus LO, Pertwee R, Howlett AC (2014) Early phytocannabinoid chemistry to endocannabinoids and beyond. Nat Rev Neurosci 15:757–764. 10.1038/nrn3811 [DOI] [PubMed] [Google Scholar]
- Melis M, Greco B, Tonini R (2014) Interplay between synaptic endocannabinoid signaling and metaplasticity in neuronal circuit function and dysfunction. Eur J Neurosci 39:1189–1201. 10.1111/ejn.12501 [DOI] [PubMed] [Google Scholar]
- Menon V. (2011) Large-scale brain networks and psychopathology: a unifying triple network model. Trends Cogn Sci 15:483–506. 10.1016/j.tics.2011.08.003 [DOI] [PubMed] [Google Scholar]
- Mereu G, Fà M, Ferraro L, Cagiano R, Antonelli T, Tattoli M, Ghiglieri V, Tanganelli S, Gessa GL, Cuomo V (2003) Prenatal exposure to a cannabinoid agonist produces memory deficits linked to dysfunction in hippocampal long-term potentiation and glutamate release. Proc Natl Acad Sci U S A 100:4915–4920. 10.1073/pnas.0537849100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer HC, Lee FS, Gee DG (2018) The role of the endocannabinoid system and genetic variation in adolescent brain development. Neuropsychopharmacology 43:21–33. 10.1038/npp.2017.143 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Millar JK, Wilson-Annan JC, Anderson S, Christie S, Taylor MS, Semple CA, Devon RS, St Clair DM, Muir WJ, Blackwood DH, Porteous DJ (2000) Disruption of two novel genes by a translocation co-segregating with schizophrenia. Hum Mol Genet 9:1415–1423. 10.1093/hmg/9.9.1415 [DOI] [PubMed] [Google Scholar]
- Miller ML, Chadwick B, Dickstein DL, Purushothaman I, Egervari G, Rahman T, Tessereau C, Hof PR, Roussos P, Shen L, Baxter MG, Hurd YL (2019) Adolescent exposure to Delta(9)-tetrahydrocannabinol alters the transcriptional trajectory and dendritic architecture of prefrontal pyramidal neurons. Mol Psychiatry 24:588–600. 10.1038/s41380-018-0243-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morris CV, DiNieri JA, Szutorisz H, Hurd YL (2011) Molecular mechanisms of maternal cannabis and cigarette use on human neurodevelopment. Eur J Neurosci 34:1574–1583. 10.1111/j.1460-9568.2011.07884.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Neill A, Mechelli A, Bhattacharyya S (2018) Dysconnectivity of large-scale functional networks in early psychosis: a meta-analysis. Schizophr Bull. Advance online publication. Retrieved Jul 3, 2019. doi: 10.1093/schbul/sby094. 10.1093/schbul/sby094 [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Shea M, McGregor IS, Mallet PE (2006) Repeated cannabinoid exposure during perinatal, adolescent or early adult ages produces similar longlasting deficits in object recognition and reduced social interaction in rats. J Psychopharmacol 20:611–621. 10.1177/0269881106065188 [DOI] [PubMed] [Google Scholar]
- Parsons LH, Hurd YL (2015) Endocannabinoid signalling in reward and addiction. Nat Rev Neurosci 16:579–594. 10.1038/nrn4004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pattwell SS, Liston C, Jing D, Ninan I, Yang RR, Witztum J, Murdock MH, Dincheva I, Bath KG, Casey BJ, Deisseroth K, Lee FS (2016) Dynamic changes in neural circuitry during adolescence are associated with persistent attenuation of fear memories. Nat Commun 7:11475. 10.1038/ncomms11475 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porath AJ, Fried PA (2005) Effects of prenatal cigarette and marijuana exposure on drug use among offspring. Neurotoxicol Teratol 27:267–277. 10.1016/j.ntt.2004.12.003 [DOI] [PubMed] [Google Scholar]
- Pouget JG, Gonçalves VF, Schizophrenia Working Group of the Psychiatric Genomics C, Spain SL, Finucane HK, Raychaudhuri S, Kennedy JL, Knight J (2016) Genome-wide association studies suggest limited immune gene enrichment in schizophrenia compared to 5 autoimmune diseases. Schizophr Bull 42:1176–1184. 10.1093/schbul/sbw059 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quickfall J, Crockford D (2006) Brain neuroimaging in cannabis use: a review. J Neuropsychiatry Clin Neurosci 18:318–332. 10.1176/jnp.2006.18.3.318 [DOI] [PubMed] [Google Scholar]
- Radua J, Schmidt A, Borgwardt S, Heinz A, Schlagenhauf F, McGuire P, Fusar-Poli P (2015) Ventral striatal activation during reward processing in psychosis: a neurofunctional meta-analysis. JAMA Psychiatry 72:1243–1251. 10.1001/jamapsychiatry.2015.2196 [DOI] [PubMed] [Google Scholar]
- Rodríguez de Fonseca F, Ramos JA, Bonnin A, Fernández-Ruiz JJ (1993) Presence of cannabinoid binding sites in the brain from early postnatal ages. Neuroreport 4:135–138. 10.1097/00001756-199302000-00005 [DOI] [PubMed] [Google Scholar]
- Rubino T, Prini P, Piscitelli F, Zamberletti E, Trusel M, Melis M, Sagheddu C, Ligresti A, Tonini R, Di Marzo V, Parolaro D (2015) Adolescent exposure to THC in female rats disrupts developmental changes in the prefrontal cortex. Neurobiol Dis 73:60–69. 10.1016/j.nbd.2014.09.015 [DOI] [PubMed] [Google Scholar]
- Saito A, Ballinger MD, Pletnikov MV, Wong DF, Kamiya A (2013) Endocannabinoid system: potential novel targets for treatment of schizophrenia. Neurobiol Dis 53:10–17. 10.1016/j.nbd.2012.11.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheyer A, Wager-Miller J, Pelissier-Alicot A, Murphy M, Mackie K, Manzoni OJ (2019) Maternal cannabinoid exposure during lactation alters the developmental trajectory of prefrontal cortex GABA-currents in offspring. Biol Psychiatry, in press. [Google Scholar]
- Schizophrenia Working Group of the Psychiatric Genomics C (2014) Biological insights from 108 schizophrenia-associated genetic loci. Nature 511:421–427. 10.1038/nature13595 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schubert D, Martens GJ, Kolk SM (2015) Molecular underpinnings of prefrontal cortex development in rodents provide insights into the etiology of neurodevelopmental disorders. Mol Psychiatry 20:795–809. 10.1038/mp.2014.147 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schulte MH, Cousijn J, den Uyl TE, Goudriaan AE, van den Brink W, Veltman DJ, Schilt T, Wiers RW (2014) Recovery of neurocognitive functions following sustained abstinence after substance dependence and implications for treatment. Clin Psychol Rev 34:531–550. 10.1016/j.cpr.2014.08.002 [DOI] [PubMed] [Google Scholar]
- Seeley WW, Menon V, Schatzberg AF, Keller J, Glover GH, Kenna H, Reiss AL, Greicius MD (2007) Dissociable intrinsic connectivity networks for salience processing and executive control. J Neurosci 27:2349–2356. 10.1523/JNEUROSCI.5587-06.2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seghier ML. (2013) The angular gyrus: multiple functions and multiple subdivisions. Neuroscientist 19:43–61. 10.1177/1073858412440596 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shrivastava A, Johnston M, Terpstra K, Bureau Y (2014) Cannabis and psychosis: neurobiology. Indian J Psychiatry 56:8–16. 10.4103/0019-5545.124708 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silins E, Horwood LJ, Patton GC, Fergusson DM, Olsson CA, Hutchinson DM, Spry E, Toumbourou JW, Degenhardt L, Swift W, Coffey C, Tait RJ, Letcher P, Copeland J, Mattick RP, Cannabis Cohorts Research Consortium (2014) Young adult sequelae of adolescent cannabis use: an integrative analysis. Lancet Psychiatry 1:286–293. 10.1016/S2215-0366(14)70307-4 [DOI] [PubMed] [Google Scholar]
- Silva L, Zhao N, Popp S, Dow-Edwards D (2012) Prenatal tetrahydrocannabinol (THC) alters cognitive function and amphetamine response from weaning to adulthood in the rat. Neurotoxicol Teratol 34:63–71. 10.1016/j.ntt.2011.10.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith AM, Fried PA, Hogan MJ, Cameron I (2004) Effects of prenatal marijuana on response inhibition: an fMRI study of young adults. Neurotoxicol Teratol 26:533–542. 10.1016/j.ntt.2004.04.004 [DOI] [PubMed] [Google Scholar]
- Smith AM, Fried PA, Hogan MJ, Cameron I (2006) Effects of prenatal marijuana on visuospatial working memory: an fMRI study in young adults. Neurotoxicol Teratol 28:286–295. 10.1016/j.ntt.2005.12.008 [DOI] [PubMed] [Google Scholar]
- Solinas M, Yasar S, Goldberg SR (2007) Endocannabinoid system involvement in brain reward processes related to drug abuse. Pharmacol Res 56:393–405. 10.1016/j.phrs.2007.09.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sonon KE, Richardson GA, Cornelius JR, Kim KH, Day NL (2015) Prenatal marijuana exposure predicts marijuana use in young adulthood. Neurotoxicol Teratol 47:10–15. 10.1016/j.ntt.2014.11.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spano MS, Ellgren M, Wang X, Hurd YL (2007) Prenatal cannabis exposure increases heroin seeking with allostatic changes in limbic enkephalin systems in adulthood. Biol Psychiatry 61:554–563. 10.1016/j.biopsych.2006.03.073 [DOI] [PubMed] [Google Scholar]
- Spear LP, File SE (1996) Methodological considerations in neurobehavioral teratology. Pharmacol Biochem Behav 55:455–457. 10.1016/S0091-3057(96)00272-9 [DOI] [PubMed] [Google Scholar]
- Sridharan D, Levitin DJ, Menon V (2008) A critical role for the right fronto-insular cortex in switching between central-executive and default-mode networks. Proc Natl Acad Sci U S A 105:12569–12574. 10.1073/pnas.0800005105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szutorisz H, Hurd YL (2018) High times for cannabis: epigenetic imprint and its legacy on brain and behavior. Neurosci Biobehav Rev 85:93–101. 10.1016/j.neubiorev.2017.05.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torii M, Sasaki M, Chang YW, Ishii S, Waxman SG, Kocsis JD, Rakic P, Hashimoto-Torii K (2017) Detection of vulnerable neurons damaged by environmental insults in utero. Proc Natl Acad Sci U S A 114:2367–2372. 10.1073/pnas.1620641114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tortoriello G, Morris CV, Alpar A, Fuzik J, Shirran SL, Calvigioni D, Keimpema E, Botting CH, Reinecke K, Herdegen T, Courtney M, Hurd YL, Harkany T (2014) Miswiring the brain: Delta9-tetrahydrocannabinol disrupts cortical development by inducing an SCG10/stathmin-2 degradation pathway. EMBO J 33:668–685. 10.1002/embj.201386035 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trezza V, Campolongo P, Cassano T, Macheda T, Dipasquale P, Carratu MR, Gaetani S, Cuomo V (2008) Effects of perinatal exposure to delta-9-tetrahydrocannabinol on the emotional reactivity of the offspring: a longitudinal behavioral study in Wistar rats. Psychopharmacology (Berl) 198:529–537. 10.1007/s00213-008-1162-3 [DOI] [PubMed] [Google Scholar]
- Tyndale RF, Payne JI, Gerber AL, Sipe JC (2007) The fatty acid amide hydrolase C385A (P129T) missense variant in cannabis users: studies of drug use and dependence in Caucasians. Am J Med Genet B Neuropsychiatr Genet 144B:660–666. 10.1002/ajmg.b.30491 [DOI] [PubMed] [Google Scholar]
- Vargish GA, Pelkey KA, Yuan X, Chittajallu R, Collins D, Fang C, McBain CJ (2017) Persistent inhibitory circuit defects and disrupted social behaviour following in utero exogenous cannabinoid exposure. Mol Psychiatry 22:56–67. 10.1038/mp.2016.17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vela G, Martín S, García-Gil L, Crespo JA, Ruiz-Gayo M, Fernández-Ruiz JJ, Garcia-Lecumberri C, Pélaprat D, Fuentes JA, Ramos JA, Ambrosio E (1998) Maternal exposure to delta9-tetrahydrocannabinol facilitates morphine self-administration behavior and changes regional binding to central mu opioid receptors in adult offspring female rats. Brain Res 807:101–109. 10.1016/S0006-8993(98)00766-5 [DOI] [PubMed] [Google Scholar]
- Wang Q, Wang Y, Ji W, Zhou G, He K, Li Z, Chen J, Li W, Wen Z, Shen J, Qiang Y, Ji J, Wang Y, Shi Y, Yi Q, Wang Y (2015) SNAP25 is associated with schizophrenia and major depressive disorder in the Han Chinese population. J Clin Psychiatry 76:e76–82. 10.4088/JCP.13m08962 [DOI] [PubMed] [Google Scholar]
- Wang X, Dow-Edwards D, Anderson V, Minkoff H, Hurd YL (2004) In utero marijuana exposure associated with abnormal amygdala dopamine D2 gene expression in the human fetus. Biol Psychiatry 56:909–915. 10.1016/j.biopsych.2004.10.015 [DOI] [PubMed] [Google Scholar]
- Wang X, Dow-Edwards D, Anderson V, Minkoff H, Hurd YL (2006) Discrete opioid gene expression impairment in the human fetal brain associated with maternal marijuana use. Pharmacogenomics J 6:255–264. 10.1038/sj.tpj.6500375 [DOI] [PubMed] [Google Scholar]
- Wijayendran SB, O'Neill A, Bhattacharyya S (2018) The effects of cannabis use on salience attribution: a systematic review. Acta Neuropsychiatr 30:43–57. 10.1017/neu.2016.58 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu CS, Jew CP, Lu HC (2011) Lasting impacts of prenatal cannabis exposure and the role of endogenous cannabinoids in the developing brain. Future Neurol 6:459–480. 10.2217/fnl.11.27 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young-Wolff KC, Tucker LY, Alexeeff S, Armstrong MA, Conway A, Weisner C, Goler N (2017) Trends in self-reported and biochemically tested marijuana use among pregnant females in California from 2009–2016. JAMA 318:2490–2491. 10.1001/jama.2017.17225 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu BD, Hanson RD, Hess JL, Horning SE, Korsmeyer SJ (1998) MLL, a mammalian trithorax-group gene, functions as a transcriptional maintenance factor in morphogenesis. Proc Natl Acad Sci U S A 95:10632–10636. 10.1073/pnas.95.18.10632 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang ZW. (2004) Maturation of layer V pyramidal neurons in the rat prefrontal cortex: intrinsic properties and synaptic function. J Neurophysiol 91:1171–1182. 10.1152/jn.00855.2003 [DOI] [PubMed] [Google Scholar]
- Zhou Y, Huang T, Lee F, Kreek MJ (2016) Involvement of endocannabinoids in alcohol “binge” drinking: studies of mice with human fatty acid amide hydrolase genetic variation and after CB1 receptor antagonists. Alcohol Clin Exp Res 40:467–473. 10.1111/acer.12989 [DOI] [PMC free article] [PubMed] [Google Scholar]