

Abstract
This study summarizes results of a DNA barcoding campaign on German Diptera, involving analysis of 45,040 specimens. The resultant DNA barcode library includes records for 2,453 named species comprising a total of 5,200 barcode index numbers (BINs), including 2,700 COI haplotype clusters without species‐level assignment, so called “dark taxa.” Overall, 88 out of 117 families (75%) recorded from Germany were covered, representing more than 50% of the 9,544 known species of German Diptera. Until now, most of these families, especially the most diverse, have been taxonomically inaccessible. By contrast, within a few years this study provided an intermediate taxonomic system for half of the German Dipteran fauna, which will provide a useful foundation for subsequent detailed, integrative taxonomic studies. Using DNA extracts derived from bulk collections made by Malaise traps, we further demonstrate that species delineation using BINs and operational taxonomic units (OTUs) constitutes an effective method for biodiversity studies using DNA metabarcoding. As the reference libraries continue to grow, and gaps in the species catalogue are filled, BIN lists assembled by metabarcoding will provide greater taxonomic resolution. The present study has three main goals: (a) to provide a DNA barcode library for 5,200 BINs of Diptera; (b) to demonstrate, based on the example of bulk extractions from a Malaise trap experiment, that DNA barcode clusters, labelled with globally unique identifiers (such as OTUs and/or BINs), provide a pragmatic, accurate solution to the “taxonomic impediment”; and (c) to demonstrate that interim names based on BINs and OTUs obtained through metabarcoding provide an effective method for studies on species‐rich groups that are usually neglected in biodiversity research projects because of their unresolved taxonomy.
Keywords: barcode library, biodiversity monitoring, CO1, cryptic diversity, Diptera, DNA barcoding, Germany, metabarcoding, mitochondrial DNA
1. INTRODUCTION
Recent evidence for major declines in insect populations has provoked intense public concern. Detailed research on economically important groups, such as pollinators, have linked declines in wild bees to pesticide contamination, climate change, habitat fragmentation and degradation (Potts et al., 2010; Vanbergen & the Insect Pollinators Initiative, 2013). Other studies using mass collecting methods suggest the declines may be general, as evidenced by reductions in the biomass of flying insects by 75% over a few decades (Hallmann et al., 2017; Sorg, Schwan, Stenmans, & Müller, 2013) or even within a few years (Lister & Garcia, 2018). However, the evidence for general declines has failed to ascertain if impacts span all insect groups and all size ranges. The failure to track the status of individual lineages reflects the fact that despite advances in taxonomic practices (e.g., integrative taxonomy), our knowledge of most insect species is limited (Brix, Leese, Riehl, & Kihara, 2015; Cruaud, Rasplus, Rodriguez, & Cruaud, 2017; Pante, Schoelinck, & Puillandre, 2014; Riedel, Sagata, Suhardjono, Tänzler, & Balke, 2013; Wheeler, Raven, & Wilson, 2004). Even in Germany, a country with more than 250 years of taxonomic and faunistic research activity, many groups remain poorly known. This gap, which hampers ecological baseline research, is particularly serious for the two hyperdiverse insect orders, the Diptera and Hymenoptera (Geiger, Moriniere, et al., 2016; Klausnitzer, 2006). With at least 9,500 (Schumann, Bährmann, & Stark, 1999; Schumann, Doczkal, & Ziegler, 2011) and 9,600 (Dathe & Blank, 2004) recorded species in Germany, respectively, these two groups comprise over half of its insect alpha diversity (Völkl, Blick, Kornacker, & Martens, 2004). Moreover, it is likely that the true diversity of these two groups is seriously underestimated, a conclusion reinforced by the extraordinarily high numbers of DNA barcode clusters retrieved by simultaneous analysis of arthropods using high‐throughput sequencing (HTS; metabarcoding) from insect collections at single monitoring sites (Morinière et al., 2016). As only about 1,000 (Santos, Sampronha, & Santos, 2017) new species of Diptera are described each year from the million or more species awaiting description, the taxonomic impediment in this group will not be resolved without the adoption of new approaches, such as modern molecular genetic methods and integrative taxonomy (Fujita, Leache, Burbrink, McGuire, & Moritz, 2012; Padial, Miralles, Riva, & Vences, 2010; Schlick‐Steiner, Arthofer, & Steiner, 2014; Schlick‐Steiner et al., 2010).
The known dipteran fauna of Germany includes roughly half of the almost 20,000 species recorded for Europe (as defined in Fauna Europaea, https://fauna-eu.org/; Pape, 2009). Although this is the highest number of Diptera species recorded from any European country, the inventory is certainly very incomplete. A recent checklist for the Empidoidea of Germany (Meyer & Stark, 2015) added 123 species new to Germany, an increase of 12.5%, Jaschhof (2009) added 34 species of Lestremiinae, an increase of 24.3%, and the collecting efforts for different barcoding campaigns resulted in more than 100 species from various families new to Germany among the identified material (Reimann & Rulik, 2015; Heller & Rulik, 2016; B. Rulik unpublished, D. Doczkal unpublished), with many more expected among the unidentified material. Rapid progress in inventorying is hampered by a lack of experts, also known as the taxonomic impediment (de Carvalho et al., 2007). For example, the German Dipterologist's working group (http://www.ak-diptera.de/index.htm, Accessed 18 December 2018) shows that experts were lacking for one‐third of the dipteran families, and that most other families had just one or two experts, often voluntary (i.e., unpaid) taxonomists (in the sense defined by Fontaine et al., 2012). A few families such as the Culicidae (https://mueckenatlas.com/), the Asilidae (Wolff, Gebel, & Geller‐Grimm, 2018) and the Syrphidae (Ssymank, Doczkal, Rennwald, & Dziock, 2011) are fairly well explored, but several of the species‐richest families (e.g., Cecidomyiidae, Ceratopogonidae, Phoridae, Chloropidae, Sphaeroceridae, Anthomyiidae) have received little attention. Malaise traps are widely used as method of choice to collect arthropods and especially flying insects for biodiversity assessments in terrestrial ecosystems, with Diptera being among the most commonly caught taxa (Doczkal, 2017; Hallmann et al., 2017; Hebert et al., 2016; Karlsson, Pape, Johansson, Liljeblad, & Ronquist, 2005; Matthews & Matthews, 1971; Ssymank et al., 2018). The analysis of specimens from two Malaise traps deployed for a single summer in Germany within the Global Malaise Trap Program (GMTP; http://biodiversitygenomics.net/projects/gmp/) revealed similar trends; here Diptera was the most diverse order being represented by 2,500 species, slightly more than half of all the species that were collected and 70.3% of all individuals (26,189) that were analysed (Geiger, Moriniere, et al., 2016).
Taxonomists working on Diptera have long been well aware of the immense number of undescribed species (Bickel et al., 2009) with estimates of global Diptera species diversity ranging from 400,000 to 800,000 species compared with ~160,000 named species (Borkent et al., 2018; Pape, Blagoderov, & Mostovski, 2011). Hebert et al. (2016), applying DNA barcoding to Canadian insects, proposed that the actual number of species could be much higher, suggesting the possible presence of 1.8 million species in just one family, the Cecidomyiidae (gall midges) alone. Although this estimate may be too high, it is very likely that this single family includes more species than are currently described for the order.
At a time when hundreds or possibly thousands of species become extinct each year (Chivian & Bernstein, 2008), a comprehensive species inventory based on accurately identified specimens represents the foundation for all conservation and biodiversity initiatives. However, the inventory of biodiversity cannot be completed through morphological approaches alone. Both the speed and costs associated with sequence characterization of a standardized DNA fragment can be improved using DNA barcoding. Usually DNA barcoding studies provide a basis for establishing the reference sequence libraries required to identify specimens of known species (Gwiazdowski, Foottit, Maw, & Hebert, 2015; Hebert, Cywinska, Ball, & Dewaard, 2003). Herein we additionally show that it is also an efficient method for registering unknown and taxonomically challenging species—so called “dark taxa” (Page, 2016). Sequenced taxa can subsequently be associated with established binomens by taxonomic specialists using a reverse taxonomy approach, based on accurately identified museum specimens (ideally type specimens) and expert knowledge. During this process, specimens that belong to unnamed molecular character‐based units (operational taxonomic units [OTUs] or barcode index numbers [BINs]) will either be referenced to known species or they may represent overlooked species that are new to science (Geiger, Moriniere, et al., 2016). A curated and comprehensive DNA barcode reference library enables fast and reliable species identifications in those many cases where time, personnel and taxonomic expertise are limited. Furthermore, such a library also supports large‐scale biodiversity monitoring that relies upon metabarcoding bulk samples (Hajibabaei, Shokralla, Zhou, Singer, & Baird, 2011; Hajibabaei, Spall, Shokralla, & Konynenburg, 2012; Shokralla, Spall, Gibson, & Hajibabaei, 2012), like those obtained from Malaise traps (Gibson et al., 2014; Leray & Knowlton, 2015; Morinière et al., 2016; Yu et al., 2012).
The results reported in this study derive from two major DNA barcoding campaigns: “Barcoding Fauna Bavarica” (BFB, http://www.faunabavarica.de; Haszprunar, 2009) and the “German Barcode of Life” project (GBOL, http://www.bolgermany.de; Geiger, Astrin, et al., 2016). Since 2009, DNA barcodes from over 23,000 German species of Metazoa have been assembled, reflecting the analysis of nearly 250,000 specimens that are curated at the SNSB‐Zoologische Staatssammlung München (ZSM, see http://www.barcoding-zsm.de) and ~180,000 specimens curated at the Zoologisches Forschungsmuseum Alexander Koenig Bonn (ZFMK). These records represent a major contribution to the global DNA barcode library that is maintained in the Barcode of Life Data System (BOLD, http://www.boldsystems.org; Ratnasingham & Hebert, 2007). Currently, the DNA barcode library created by the ZSM researchers represents the second‐most comprehensive library of any nation. Previous studies have reported on barcoding results for Coleoptera (Hendrich et al., 2015; Raupach, Hannig, Moriniere, & Hendrich, 2016; Raupach, Hannig, Morinière, & Hendrich, 2018; Rulik et al., 2017), Ephemeroptera, Plecoptera and Trichoptera (Morinière et al., 2017), Heteroptera (Havemann et al., 2018; Raupach et al., 2014), Hymenoptera (Schmid‐Egger et al., 2019; Schmidt, Schmid‐Egger, Morinière, Haszprunar, & Hebert, 2015; Schmidt et al., 2017), Lepidoptera (Hausmann, Haszprunar, & Hebert, 2011; Hausmann, Haszprunar, Segerer, et al., 2011), Neuroptera (Morinière et al., 2014), Orthoptera (Hawlitschek et al., 2017), Araneae and Opiliones (Astrin et al., 2016), and Myriapoda (Spelda, Reip, Oliveira Biener, & Melzer, 2011; Wesener et al., 2015). Concerning DNA barcoding studies performed for Diptera, no comprehensive study encompassing this entire highly diverse order has been published, but data have been used to revise smaller units thereof: for example, for Calliphoridae (Jordaens et al., 2013; Nelson, Wallman, & Dowton, 2007; Reibe, Schmitz, & Madea, 2009), Ceratopogonidae (Stur & Borkent, 2014), Chironomidae (Carew, Pettigrove, Cox, & Hoffmann, 2007; Carew, Pettigrove, & Hoffmann, 2005; Cranston et al., 2013; Ekrem, Stur, & Hebert, 2010; Ekrem, Willassen, & Stur, 2007; Montagna, Mereghetti, Lencioni, & Rossaro, 2016; Pfenninger, Nowak, Kley, Steinke, & Streit, 2007; Sinclair & Gresens, 2008; Stur & Ekrem, 2011), Culicidae (Ashfaq et al., 2014; Cywinska, Hunter, & Hebert, 2006; Kumar, Rajavel, Natarajan, & Jambulingam, 2007; Versteirt et al., 2015; Wang et al., 2012), Hybotidae (Nagy, Sonet, Mortelmans, Vandewynkel, & Grootaert, 2013), Muscidae (Renaud, Savage, & Adamowicz, 2012), Psychodidae (Gutiérrez, Vivero, Vélez, Porter, & Uribe, 2014; Krüger, Strüven, Post, & Faulde, 2011; Kumar, Srinivasan, & Jambulingam, 2012; Nzelu et al., 2015), Sciaridae (Eiseman, Heller, & Rulik, 2016; Heller, Köhler, Menzel, Olsen, & Gammelo, 2016; Heller & Rulik, 2016; Latibari, Moravvej, Heller, Rulik, & Namaghi, 2015; Ševčík, Kaspřák, & Rulik, 2016), Simuliidae (Rivera & Currie, 2009), Syrphidae (Jordaens et al., 2015) and Tachinidae (Pohjoismäki, Kahanpää, & Mutanen, 2016).
This publication presents the first results of the Diptera campaign and it provides coverage for 5,200 BINs (Ratnasingham & Hebert, 2013). It covers ~55% of the known Diptera fauna from Germany. According to the checklist of German Diptera (Schumann et al., 1999) and the three additions published so far (Schumann, 2002, 2004, 2010) 9,544 species of Diptera have been recorded from Germany. The Diptera library now includes a total of 2,453 reliable species identifications, and 2,700 BINs, which possess either interim species names or just higher‐level taxonomy (genus or family; “dark taxa”). Although it has been shown that BINs correspond closely to biological species of most insect orders (Hausmann et al., 2013), there are other studies reporting difficulties in determining species through DNA barcodes within Diptera. In particular, well‐studied groups such as the syrphids represent a problem, because here additional genes for a clear type assignment must be consulted in many genera (Mengual, Ståhls, Vujić, & Marcos‐Garcia, 2006; Rojo, Ståhls, Pérez‐Bañón, & Marcos‐García, 2006). Further examples of problems in species delineation due to barcode gaps, at least for some genera, are the Tachinidae and the Calliphoridae (Nelson et al., 2012; Pohjoismäki et al., 2016; Whitworth, Dawson, Magalon, & Baudry, 2007). In one of the few studies dealing with DNA barcoding in Diptera it was shown, that less than 70% of a composition of about 450 species covering 12 families of Diptera could be reliably identified by DNA barcoding, as there was wide overlap between intra‐ and interspecific genetic variability on the COI gene (Meier, Shiyang, Vaidya, & Ng, 2006). However we find that more than 88% of the studied species, identified based on morphology or BIN matches to the BOLD database, can be unambiguously identified using their DNA barcode sequences. BINs enable the creation of an interim taxonomic system in a structured, transparent and sustainable way and thus become a valuable foundation for subsequent detailed, integrative taxonomic studies. Furthermore, the BIN system enables analyses that are equivalent to studies based on named species, that is where the underlying specimens are identified by specialists using traditional methods (i.e., morphology). The latter will play a special role in the processing, classification and genetic inventorying of less‐explored “dark taxa,” which have been treated and processed with less priority by previous DNA barcoding activities. Moreover, this automated approach of delineating species is less affected by operational taxonomic biases, so it can provide more objective identifications than conventional approaches (Mutanen et al., 2016; Packer, Gibbs, Sheffield, & Hanner, 2009; Schmidt et al., 2015). Using DNA extracts derived from bulk collections made by Malaise traps, we further demonstrate that species delineation using interim names based on BINs and OTUs constitutes an effective method for biodiversity studies using DNA metabarcoding. As the reference libraries continue to grow and gaps in the species catalogue are subsequently filled, BIN lists assembled by metabarcoding will provide improved taxonomic resolution.
The present study has three main goals: (a) to provide a DNA barcode library for 5,200 BINs of Diptera; (b) to demonstrate, based on the example of bulk extractions from a Malaise trap experiment, that DNA barcode clusters, labelled with globally unique identifiers (such as OTUs and/or BINs), provide a pragmatic, accurate solution to the “taxonomic impediment”; and (c) to demonstrate that interim names based on BINs and OTUs obtained through metabarcoding is an effective method for studies on species‐rich groups that are usually neglected in biodiversity research projects because of their unresolved taxonomy.
2. MATERIALS AND METHODS
2.1. Fieldwork, specimens and taxonomy
A network of 130 (professional and voluntary) taxonomists and citizen scientists collected and contributed specimens to the DNA barcoding projects, primarily from various German states, but also from surrounding European countries (Austria, Belgium, Czech Republic, France, Italy). Most specimens (94.5%, 42,587 of 45,040 with COI sequences >500 bp) were collected by Malaise traps, which were deployed from 2009 to 2016. The study sites included more than 683 localities in state forests, public lands and protected areas such as the Nationalparks “Bayerischer Wald” and “Berchtesgadener Land,” the EU habitats directive site “Landskrone,” as well as alpine regions at altitudes up to 2,926 m (Zugspitze). Detailed information on collection sites and dates is available in Appendix S1. Since 2009, more than five million specimens of Diptera were collected by hand collecting, sweep netting, and by Malaise‐, window‐ and pitfall‐trapping. However, most voucher specimens have been extracted from Malaise trap samples. Twenty to 100 Malaise traps were deployed in each of seven years (2011–2017) mostly across habitats in Bavaria and Baden‐Wurttemberg; one trap was placed in Rhineland‐Palatinate. Samples were screened morphologically to maximize the diversity of species submitted for sequence characterization. Most vouchers were derived from Germany (44,511), but others were collected in France (222), Czech Republic (147), Belgium (106), Austria (70) and other Central European countries (18). All samples and specimens are now stored in the SNSB‐ZSM or ZFMK except for a few held in private collections. From the entire collection, ~3,000,000 specimens of potential interest, most of which derived from the huge Malaise trap experiments in the framework of the GMTP, were identified to family level mostly by D.D. and to a minor extent by B.R. and experienced specialists using appropriate literature (Oosterbroek, 2006 and references therein; Papp & Darvas, 1997, 1998, 2000a, 2000b, Schumann et al., 2011). From this material, 59,000 specimens were submitted for sequence analysis through the DNA barcoding pipeline (including sample preparation, high‐quality imaging and metadata acquisition for each specimen) established at the ZSM to support its involvement in national and international DNA barcoding projects. Most samples (>99%) were stored in 96% EtOH before DNA extraction. Specimen ages generally ranged from 1 to 5 years (43,112 specimens, 96%); only 4% were more than 5 years old. The number of specimens analysed per species ranged from one to 1,356 (i.e., Megaselia rufa) (Wood, 1908; see Appendix S1). When taxonomic expertise was available, specimens were sent to specialists to obtain as many species‐level identifications as possible.
2.2. Laboratory protocols
A tissue sample was removed from each specimen and transferred into 96‐well plates at the SNSB‐ZSM for subsequent DNA extraction. For specimens with a body length >2 mm a single leg or a leg segment was removed for DNA extraction. The whole voucher was used for some very small specimens (e.g., ≤1 mm, such as small Cecidomyiidae, Chironomidae and Sciaridae), but replacement vouchers from the same locality were retained. In other cases (vouchers from Malaise traps), DNA was extracted from the whole voucher at the CCDB (Guelph, Canada) using “voucher‐recovery” protocols (DeWaard et al., 2019) and the specimens were repatriated to the SNSB‐ZSM and ZFMK for identification and curation. DNA extraction plates with the tissue samples were sent to the Canadian Center for DNA Barcoding (CCDB) where they were processed using standard protocols. All protocols for DNA extraction, PCR amplifications and Sanger sequencing procedures are available online (ccdb.ca/resources/). All samples were PCR‐amplified with a cocktail of standard and modified Folmer primers CLepFolF (5′‐ATTCAACCAATCATAAAGATATTGG) and CLepFolR (5′TAAACTTCTGGATGTCCAAAAAATCA) for the barcode fragment (5′ COI; see Hernández‐Triana et al., 2014), and the same primers were employed for subsequent bidirectional Sanger sequencing reactions (see also Ivanova, Dewaard, & Hebert, 2006; deWaard, Ivanova, Hajibabaei, & Hebert, 2008, DeWaard et al., 2019). Voucher information such as locality data, habitat, altitude, collector, identifier, taxonomic classifications, habitus images, DNA barcode sequences, primer pairs and trace files for 40,753 specimens are publicly accessible in the “DS‐DIPBFGBL—A DNA Barcode reference library of German Diptera (BFB—Barcoding Fauna Bavarica & GBOL—German Barcode of Life” data set on BOLD (http://www.boldsystems.org – data set DOI: http://dx.doi.org/10.5883/DS-DIPBFGBL), whereas 4,420 specimen records will be stored in the private data set “DS‐DIPBFGBP—A DNA Barcode reference library of German Diptera (BFB—Barcoding Fauna Bavarica & GBOL—German Barcode of Life)—private records for future publication” for subsequent publication by the authors and associated taxonomists.
2.3. Data analysis
Sequence divergences for the COI‐5P barcode region (mean and maximum intraspecific variation and minimum genetic distance to the nearest‐neighbouring species) were calculated using the “Barcode Gap Analysis” tool on BOLD, employing the Kimura 2‐parameter (K2P) distance metric (Puillandre, Lambert, Brouillet, & Achaz, 2012). The program muscle was applied for sequence alignment restricting analysis to sequences with a minimum length of 500 bp. Neighbour‐joining (NJ) trees were calculated following alignment based on K2P distances. The “BIN Discordance” analysis on BOLD was used to reveal cases where species assigned to different species shared a BIN, and those cases where a particular species was assigned to two or more BINs. Sequences are grouped into clusters of closely similar COI barcode sequences, which are assigned a globally unique identifier, termed a “barcode index number” or BIN (Ratnasingham & Hebert, 2013). This system enables tentative species identifications when taxonomic information is lacking. The BIN system involves a three‐step online pipeline, which clusters similar barcode sequences algorithmically into OTUs being “named” by a number. For the majority of studied insect orders, specimens sharing a BIN very often represent a close species‐proxy as delineated by traditional taxonomy (e.g., for Lepidoptera, Hausmann et al., 2013). However, some genera or families throughout the insects exhibit problems with species delineation based on DNA barcodes, due to high intra‐ or low interspecific genetic distances (e.g., cryptic diversity, BIN sharing or the barcode gap; see Hubert & Hanner, 2015). Within the Diptera, this phenomenon has been well documented (Meier et al., 2006), at least in some families, such as calliphorid, syrphid and tachinid species (Mengual et al., 2006; Nelson et al., 2012; Pohjoismäki et al., 2016; Rojo et al., 2006; Whitworth et al., 2007), but may also occur in families of “dark taxa” as well.
Every other “disagreement/conflict” case is the starting point for re‐evaluation of both molecular and morphological data. We follow the concept of Integrative Taxonomy (Fujita et al., 2012; Padial et al., 2010; Schlick‐Steiner et al., 2014, 2010) to infer whether there are previously overlooked species (“cryptic taxa”) in the sample, or whether barcode divergence between species is too low or absent to allow valid species to be delineated using only COI characteristics.
2.4. Reverse‐taxonomy approach
When sequenced specimens could only be assigned to a category above the species level (family, subfamily or genus), we used interim species names (such as TachIntGen1 sp.BOLD:AAG2112) based on the corresponding BIN, so these specimens could be included in the “Barcode Gap Analysis” in order to provide more comprehensive estimates of the distribution of genetic divergences among both species assigned to Linnaean species and those with BIN assignments. This analysis was conducted on all specimens at the same time after updating the interim taxonomy where necessary. For specimen records, which lack lower taxonomy (e.g., those uploaded only as “Diptera”), we applied the highest “conflict‐free” taxonomy—for example the genus name, when other specimens within that BIN had the same identification—using a BIN match with the public data on BOLD (e.g., Melanagromyza sp. BOLD:ACP6151). All specimens, which could not be identified to species or genus level, and where the vouchers were in acceptable condition (e.g., unbroken antennae and/or legs after retrieval from Malaise trap), were selected using the corresponding BINs for identification by taxonomic specialists. Interim names were subsequently moved into the “Voucher status” field in the BOLD metadata tables after all analyses were performed.
2.5. Metabarcoding and bioinformatic data analysis
The potential utility of the DNA barcode library for biomonitoring Diptera was tested with field samples, focusing on an early warning system for pest and invasive species based on metabarcoding (L. A. Hardulak et al. in prep). In this study, nine Malaise traps were deployed in the Bayerischer Wald National Park and its surroundings during the vegetated period (May–September) in 2016. Trap bottles were changed twice monthly, producing a total of 90 bulk samples of macroinvertebrates. All specimens were dried and ground with a stainless steel pestle (no size‐sorting step), and tissue lysis of insect powder per trap sample was performed overnight, using a solution of 90% insect lysis buffer and 10% proteinase K. DNA extraction was performed with the DNEasy Blood & Tissue kit (Qiagen). A minibarcode region was amplified by PCR, using forward and reverse NGS primers (Leray et al., 2013) targeting a 313‐bp‐long coding region of mitochondrial COI. High‐throughput sequencing was performed on an Illumina MiSeq using version 2 (2 × 250 bp, 500 cycles, maximum of 20 million reads) chemistry at the Sequencing Service Unit of the Ludwig‐Maximilians University (LMU, Munich, Germany; see Appendix S5 for a more detailed metabarcoding protocol).
Sequence processing was performed with the vsearch version 2.4.3 suite (Rognes, Flouri, Nichols, Quince, & Mahé, 2016) and cutadapt version 1.14 (Martin, 2011). Forward and reverse reads in each sample were merged with the vsearch program “fastq_mergepairs” with a minimum overlap of 40 bp, yielding ~313‐bp sequences. Forward and reverse primers were removed with cutadapt, using the “discard_untrimmed” option to discard sequences for which primers were not detected at ≥90% identity. Quality filtering was done with the “fastq_filter” in vsearch, keeping sequences with zero expected errors (“fastq_maxee” 1). Sequences were dereplicated with “derep_fulllength,” first at the sample level, and then concatenated into a fasta file, which was then dereplicated. Chimeric sequences were removed from the fasta file using “uchime_denovo.” The remaining sequences were then clustered into OTUs at 97% identity employing “cluster_size,” a greedy, centroid‐based clustering program. OTUs were blasted against the Diptera database downloaded from BOLD including taxonomy and BIN information in geneious (version 9.1.7; Biomatters) following the methods described in Morinière et al. (2016). The resulting csv file, which included BIN, Hit‐%‐ID value, family, genus and species information for each out, was exported from Geneious and combined with the OTU table generated by the bioinformatic pipeline. The combined results table was then filtered by Hit‐%‐ID value and total read numbers per OTU. All entries with identifications below 97% and total read numbers below 0.01% of the summed reads per sample were removed from the analysis. OTUs were then assigned to the respective BIN (Appendix S2). Presence–absence overviews of selected Diptera taxa (BINs) within the metabarcoding study were created; one‐sided Pearson correlation coefficients were calculated to estimate the percentage of “dark taxa” with mid‐range body size versus the number of species reported in Germany, both with the inclusion and with the exclusion of families with 0% “dark taxa.” (r version 3.4.4 [2018–03‐15], R Core Team, 2018).
3. RESULTS
3.1. DNA barcoding/developing a reference library
From the 59,102 specimens submitted for Sanger sequencing, 50,963 COI‐5P sequences (86.23%) were recovered. Length of the recovered sequence varied with the sequencing protocol; 12.54% (7,410 specimens) were bidirectionally sequenced and yielded a full‐length (658 bp) barcode while the rest (43,533) were unidirectionally sequenced yielding 69.95% (41,339) with sequences <658 to >500 bp and 3.75% (2,214 specimens) with sequences <500 bp. No sequence information was recovered from 13.77% (8,139) of the specimens. Barcode recovery was most successful for EtOH‐preserved specimens less than 10 years old. For the subsequent analyses we selected 45,040 specimens with high‐quality DNA barcode sequences (≥500 bp), which fulfilled the requirements for being assigned to a BIN. This data set included ~5,200 BINs (2,500 were assigned a total of 2,453 Linnean species while 2,700 lacked a species designation, 52.4% of the data set). These BINs included one or more representatives from 88 of the 117 (75%) dipteran families known from Germany (Figure 1, Table 1; Appendix S3, Krona graph in Figure S2). More than one‐third (1,829) of the BINs were new to BOLD.
Figure 1.
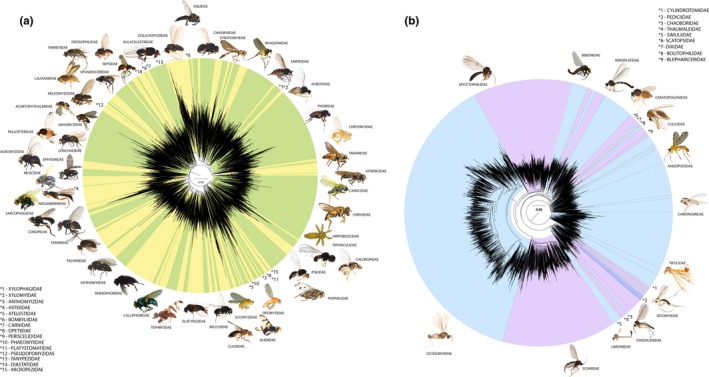
Illustrative circular neighbour‐joining (NJ) trees for (a) all Brachycera and (b) all Nemtatocera within the Diptera barcode library; each line in the trees corresponds to one barcode index number (BIN). NJ tree calculations were performed on the BOLD database. A more detailed observation of the BIN diversity for each family can be studied within the Krona graph within the supporting information (Figure S2) [Colour figure can be viewed at http://www.wileyonlinelibrary.com]
Table 1.
Families of Diptera reported in Germany. Information on BIN count, and on the numbers of named and unnamed species within the reference database
| Infraorder | Family | Species reported in Germany | BINs | Ratio barcoded/species (%) | Size (mm) | Total number of taxa/with barcode | Unnamed/with barcode | % of dark taxa |
|---|---|---|---|---|---|---|---|---|
| Brachycera | Acartophthalmidae | 2 | 1 | 50 | 1.0–2.5 | 2 | 0 | 0 |
| Brachycera | Acroceridae | 11 | 0 | 0 | 2.5–20.0 | N/A | N/A | N/A |
| Brachycera | Agromyzidae | 552 | 218 | 39 | 1.0–6.0 | 214 | 149 | 70 |
| Nematocera | Anisopodidae (& Mycetobiidae) | 8 | 7 | 88 | 4.0–12.0 | 7 | 2 | 29 |
| Brachycera | Anthomyiidae | 227 | 188 | 83 | 4.0–12.0 | 178 | 64 | 36 |
| Brachycera | Anthomyzidae | 14 | 5 | 36 | 1.3–4.5 | 5 | 0 | 0 |
| Brachycera | Asilidae | 81 | 18 | 22 | 8.0–20.0 | 18 | 6 | 33 |
| Brachycera | Asteiidae | 7 | 3 | 43 | 1.0–3.0 | 3 | 0 | 0 |
| Brachycera | Atelestidae | 3 | 3 | 100 | 1.5–3.5 | 3 | 0 | 0 |
| Brachycera | Athericidae | 5 | 3 | 60 | 7.5–10.0 | 3 | 1 | 33 |
| Brachycera | Aulacigastridae | 1 | 0 | 0 | 2.0–5.0 | 0 | 0 | N/A |
| Nematocera | Bibionidae (& Pleciidae) | 21 | 12 | 57 | 2.0–15.0 | 10 | 2 | 20 |
| Nematocera | Blephariceridae | 7 | 2 | 29 | 3.0–15.0 | 2 | 1 | 50 |
| Nematocera | Bolitophilidae | 22 | 14 | 64 | 4.0–7.0 | 13 | 7 | 54 |
| Brachycera | Bombyliidae | 40 | 6 | 15 | 1.0–20.0 | 6 | 1 | 17 |
| Brachycera | Braulidae | 1 | 0 | 0 | 1.2–2.5 | 2 | 0 | 0 |
| Brachycera | Calliphoridae | 62 | 35 | 56 | 4.0–16.0 | 39 | 6 | 15 |
| Brachycera | Camillidae | 4 | 0 | 0 | 2.0–3.5 | N/A | N/A | N/A |
| Brachycera | Campichoetidae | 3 | 0 | 0 | 2.5–4.0 | N/A | N/A | N/A |
| Brachycera | Canacidae | 2 | 9 | 450 | 1.6–5.0 | 9 | 1 | 11 |
| Nematocera | Canthyloscelidae | 1 | 0 | 0 | 2.5–9.0 | N/A | N/A | N/A |
| Brachycera | Carnidae | 11 | 7 | 64 | 1.0–2.5 | 7 | 7 | 100 |
| Nematocera | Cecidomyiidae | 836 | 927 | 111 | 0.5–3.0 | 926 | 882 | 95 |
| Nematocera | Ceratopogonidae | 332 | 131 | 39 | 1.0–5.0 | 128 | 97 | 76 |
| Brachycera | Chamaemyiidae | 29 | 17 | 59 | 1.0–5.0 | 17 | 13 | 76 |
| Nematocera | Chaoboridae | 7 | 2 | 29 | 2.0–10.0 | 2 | 0 | 0 |
| Nematocera | Chironomidae | 696 | 455 | 65 | 1.0–10.0 | 438 | 286 | 65 |
| Brachycera | Chloropidae | 198 | 101 | 51 | 1.0–5.0 | 101 | 59 | 58 |
| Brachycera | Chyromyidae | 5 | 2 | 40 | 0.5–8.0 | 2 | 0 | 0 |
| Brachycera | Clusiidae | 9 | 6 | 67 | 1.5–8.0 | 7 | 3 | 43 |
| Brachycera | Coelopidae | 2 | 0 | 0 | 2.5–9.0 | N/A | N/A | N/A |
| Brachycera | Coenomyiidae | 1 | 0 | 0 | 14.0–20.0 | N/A | N/A | N/A |
| Brachycera | Conopidae | 52 | 9 | 17 | 5.0–15.0 | 9 | 0 | 0 |
| Brachycera | Cremifaniidae | 1 | 0 | 0 | 1.5–2.6 | N/A | N/A | N/A |
| Brachycera | Cryptochetidae | 1 | 0 | 0 | 2.0–4.0 | N/A | N/A | N/A |
| Nematocera | Culicidae | 46 | 8 | 17 | 3.0–9.0 | 7 | 0 | 0 |
| Nematocera | Cylindrotomidae | 4 | 1 | 25 | 11.0–16.0 | 1 | 0 | 0 |
| Nematocera | Diadocidiidae | 4 | 3 | 75 | 3–4.5.0 | 3 | 0 | 0 |
| Brachycera | Diastatidae | 6 | 8 | 133 | 2.5–4.0 | 8 | 2 | 25 |
| Nematocera | Ditomyiidae | 4 | 1 | 25 | 6.0–8.0 | 1 | 0 | 0 |
| Nematocera | Dixidae | 16 | 4 | 25 | 3.0–5.5 | 4 | 1 | 25 |
| Brachycera | Dolichopodidae | 356 | 112 | 31 | 1.0–9.0 | 112 | 58 | 52 |
| Brachycera | Drosophilidae | 59 | 28 | 47 | 1.5–7.0 | 28 | 5 | 18 |
| Brachycera | Dryomyzidae | 3 | 2 | 67 | 5.0–18.0 | 2 | 1 | 50 |
| Brachycera | Eginiidae | 1 | 0 | 0 | 2.0–18.0 | N/A | N/A | N/A |
| Brachycera | Empididae (& Brachystomatidae) | 383 | 161 | 42 | 1.0–12.0 | 161 | 107 | 66 |
| Brachycera | Ephydridae | 177 | 130 | 73 | 1.0–11.0 | 132 | 16 | 12 |
| Brachycera | Fanniidae | 56 | 46 | 82 | 2.0–5.0 | 44 | 13 | 30 |
| Brachycera | Gasterophilidae | 4 | 0 | 0 | 9.0–16.0 | N/A | N/A | N/A |
| Brachycera | Helcomyzidae | 3 | 0 | 0 | 6.0–11.0 | N/A | N/A | N/A |
| Brachycera | Heleomyzidae (& Heteromyzidae) | 74 | 58 | 78 | 1.2–12.0 | 55 | 26 | 47 |
| Nematocera | Hesperinidae | 1 | 0 | 0 | 4.0–6.0 | N/A | N/A | N/A |
| Brachycera | Hilarimorphidae | 2 | 0 | 0 | 2.0–7.0 | N/A | N/A | N/A |
| Brachycera | Hippoboscidae | 12 | 7 | 58 | 2.5–10.0 | 7 | 1 | 14 |
| Brachycera | Hybotidae | 229 | 140 | 61 | 1.0–6.0 | 139 | 83 | 60 |
| Brachycera | Hypodermatidae | 5 | 0 | 0 | 10.0–22 | N/A | N/A | N/A |
| Nematocera | Keroplatidae | 60 | 30 | 50 | 4.0–15.0 | 30 | 12 | 40 |
| Brachycera | Lauxaniidae | 67 | 25 | 37 | 2.0–7.0 | 25 | 11 | 44 |
| Nematocera | Limoniidae | 280 | 96 | 34 | 2.0–11.0 | 91 | 50 | 55 |
| Brachycera | Lonchaeidae | 47 | 16 | 34 | 3.0–6.0 | 16 | 9 | 56 |
| Brachycera | Lonchopteridae | 9 | 5 | 56 | 2.0–5.0 | 6 | 0 | 0 |
| Brachycera | Megamerinidae | 1 | 1 | 100 | 6.0–9.0 | 1 | 0 | 0 |
| Brachycera | Micropezidae | 13 | 5 | 38 | 3.0–16.0 | 4 | 1 | 25 |
| Brachycera | Microphoridae | 6 | 0 | 0 | 1.5–3.0 | N/A | N/A | N/A |
| Brachycera | Milichiidae | 13 | 17 | 131 | 1.0–6.0 | 16 | 9 | 56 |
| Brachycera | Muscidae | 317 | 174 | 55 | 2.0–18.0 | 167 | 66 | 40 |
| Nematocera | Mycetophilidae | 573 | 306 | 53 | 2.0–15.0 | 301 | 89 | 30 |
| Brachycera | Neottiophilidae | 1 | 0 | 0 | 1.5–7.0 | N/A | N/A | N/A |
| Brachycera | Nycteribiidae | 8 | 0 | 0 | 1.5–5.0 | N/A | N/A | N/A |
| Brachycera | Odiniidae | 9 | 0 | 0 | 2.0–5.0 | N/A | N/A | N/A |
| Brachycera | Oestridae | 6 | 0 | 0 | 9.0–18.0 | N/A | N/A | N/A |
| Brachycera | Opetiidae | 1 | 1 | 100 | 2.0–5.0 | 1 | 0 | 0 |
| Brachycera | Opomyzidae | 15 | 4 | 27 | 2.0–5.0 | 4 | 1 | 25 |
| Brachycera | Otitidae | 26 | 0 | 0 | 2.5–11.0 | N/A | N/A | N/A |
| Brachycera | Pallopteridae | 16 | 8 | 50 | 2.5–7.0 | 7 | 0 | 0 |
| Nematocera | Pediciidae | 36 | 13 | 36 | 5.0–35.0 | 13 | 3 | 23 |
| Brachycera | Periscelididae | 6 | 1 | 17 | 1.0–5.0 | 1 | 0 | 0 |
| Brachycera | Phaeomyiidae | 3 | 2 | 67 | 3.0–11.0 | 2 | 0 | 0 |
| Brachycera | Phoridae | 364 | 289 | 79 | 0.5–6.0 | 276 | 166 | 60 |
| Brachycera | Piophilidae | 12 | 12 | 100 | 1.5–7.0 | 12 | 4 | 33 |
| Brachycera | Pipunculidae | 111 | 42 | 38 | 2.0–12.0 | 40 | 7 | 18 |
| Brachycera | Platypezidae | 23 | 4 | 17 | 1.5–6.0 | 4 | 0 | 0 |
| Brachycera | Platystomatidae | 3 | 2 | 67 | 3.0–11.0 | 2 | 0 | 0 |
| Brachycera | Pseudopomyzidae | 1 | 1 | 100 | 1.7–2.5 | 1 | 0 | 0 |
| Brachycera | Psilidae | 30 | 12 | 40 | 2.5–10.0 | 12 | 8 | 67 |
| Nematocera | Psychodidae | 143 | 51 | 36 | 2.0–6.0 | 50 | 25 | 50 |
| Nematocera | Ptychopteridae | 8 | 0 | 0 | 7.0–15.0 | N/A | N/A | N/A |
| Brachycera | Pyrgotidae | 1 | 0 | 0 | 8.0–9.0 | N/A | N/A | N/A |
| Brachycera | Rhagionidae | 35 | 20 | 57 | 2.0–20.0 | 20 | 10 | 50 |
| Brachycera | Rhinophoridae | 10 | 9 | 90 | 2.0–11.0 | 7 | 1 | 14 |
| Brachycera | Sarcophagidae | 130 | 49 | 38 | 3.0–22.0 | 49 | 17 | 35 |
| Brachycera | Scatophagidae | 57 | 0 | 0 | 3.0–12.0 | 0 | 0 | N/A |
| Nematocera | Scatopsidae | 47 | 30 | 64 | 0.5–4.0 | 30 | 24 | 80 |
| Brachycera | Scenopinidae | 3 | 0 | 0 | 2.0–7.0 | N/A | N/A | N/A |
| Nematocera | Sciaridae | 342 | 310 | 91 | 1.0–6.0 | 284 | 81 | 29 |
| Brachycera | Sciomyzidae | 78 | 19 | 24 | 2.0–14.0 | 18 | 4 | 22 |
| Brachycera | Sepsidae | 31 | 15 | 48 | 2.0–6.0 | 13 | 1 | 8 |
| Nematocera | Simuliidae | 50 | 19 | 38 | 1.2–6.0 | 18 | 9 | 50 |
| Brachycera | Sphaeroceridae | 137 | 79 | 58 | 0.7–5.5 | 77 | 31 | 40 |
| Brachycera | Stratiomyidae | 66 | 21 | 32 | 2.0–25.0 | 22 | 6 | 27 |
| Brachycera | Strongylophthalmyiidae | 1 | 0 | 0 | 3.0–5.5 | N/A | N/A | N/A |
| Brachycera | Syrphidae | 440 | 242 | 55 | 3.5–35.0 | 297 | 24 | 8 |
| Brachycera | Tabanidae | 58 | 46 | 79 | 6.0–30.0 | 45 | 3 | 7 |
| Brachycera | Tachinidae | 494 | 214 | 43 | 2.0–20.0 | 211 | 76 | 36 |
| Brachycera | Tanypezidae | 1 | 1 | 100 | 5.0–8.0 | 1 | 0 | 0 |
| Brachycera | Tephritidae | 110 | 28 | 25 | 2.5–10.0 | 27 | 5 | 19 |
| Brachycera | Tethinidae | 10 | 0 | 0 | 1.5–3.5 | N/A | N/A | N/A |
| Nematocera | Thaumaleidae | 15 | 13 | 87 | 3.0–5.0 | 13 | 1 | 8 |
| Brachycera | Therevidae | 32 | 4 | 13 | 2.5–15.0 | 4 | 1 | 25 |
| Brachycera | Thyreophoridae | 2 | 0 | 0 | 1.5–7.0 | N/A | N/A | N/A |
| Nematocera | Tipulidae | 123 | 46 | 37 | 7.0–35.0 | 46 | 15 | 33 |
| Nematocera | Trichoceridae | 18 | 24 | 133 | 3.0–9.0 | 24 | 17 | 71 |
| Brachycera | Trixoscelididae | 4 | 0 | 0 | 2.0–4.0 | N/A | N/A | N/A |
| Brachycera | Ulidiidae | 4 | 9 | 225 | 2.5–11.0 | 9 | 4 | 44 |
| Brachycera | Xylomyidae | 3 | 1 | 33 | 6.0–20.0 | 1 | 0 | 0 |
| Brachycera | Xylophagidae | 4 | 1 | 25 | 5.0–11.0 | 1 | 0 | 0 |
Additional information on the average body size of the specimens in each family is included.
Inspection of the COI sequence clusters using NJ trees (created with analytical tools on BOLD) and using the TaxCl‐approach for detecting taxonomic incongruences (Rulik et al., 2017) revealed high congruence with morphology‐based identifications. Among the 2,453 taxa assigned a Linnean binomen based on morphological identifications and “conflict‐free” BIN matches, 88.67% (2,138) were unambiguously discriminated by their COI sequences. Another 122 species (4.97%), representing 8.7% of all studied specimens (3,951 individuals), were assigned to more than one BIN, resulting in a total of 255 BINs (Table 1; Appendix S3). For purposes of re‐identification, the species in this subset can also be unambiguously assigned to a current species. For 34 of these taxa, the maximum intraspecific variation (maxISP) was <3% (range: 1.1%–3.0%), cases which may reflect either young sibling species or high intraspecific variation arising from secondary contact between phylogeographical lineages. Another 88 species showed considerably higher divergences with maxISP ranging from 3% to 6% in 48 species and from 6% to 12% in another 40 species, cases that are strong candidates for overlooked cryptic diversity. Most of these cases involved species whose members were assigned to two BINs (112 species), but specimens of nine species were assigned to three BINs and those of one other to four BINs. Another 156 species (6.56%), representing 2.9% of all specimens (1,316 specimens), involved two or more named species that shared a BIN (Table 2). Ten of these species pairs possessed shallow but consistent divergences within the BIN, meaning that COI sequences enabled species identification (e.g., Chrysotoxum bicinctum Linnaeus, 1758 and Chrysotoxum festivum Linnaeus, 1758; Sericomyia lappona Linnaeus, 1758 and Sericomyia silentis Harris 1776; Paragus majoranae Rondani, 1857 and Paragus pecchiolii, Rondani, 1857). Interestingly, almost two‐thirds (105/156) of the species exhibiting BIN sharing (168) were hoverflies (Syrphidae), a family that has seen intensive taxonomic study.
Table 2.
All cases of high intraspecific sequence variation at COI; cases of multiple BINs and/or cryptic diversity candidates (CDC)
| Family | Species | CDC rank | Mean intraspecific variation | Max. intraspecific variation | BIN |
|---|---|---|---|---|---|
| Agromyzidae | Napomyza cichorii | CDC (2) | 2.47 | 3.71 | BOLD:AAP2990 |
| BOLD:AAX3741 | |||||
| Phytomyza continua | CDC (2) | 2.84 | 5.44 | BOLD:AAM6330 | |
| BOLD:AAY2701 | |||||
| Phytomyza ranunculi | CDC (2) | 3.26 | 6.43 | BOLD:AAY3895 | |
| BOLD:ACL2003 | |||||
| Anthomyiidae | Anthomyia liturata | CDC (2) | 0.87 | 1.98 | BOLD:ACE4539 |
| BOLD:ACE4540 | |||||
| Delia nuda | CDC (2) | 1.06 | 1.87 | BOLD:ACJ0544 | |
| BOLD:ACJ0545 | |||||
| Hydrophoria lancifer | CDC (2) | 0.61 | 3.04 | BOLD:AAG2460 | |
| BOLD:ADC1814 | |||||
| Pegomya flavifrons | CDC (2) | 2.5 | 8.83 | BOLD:AAG2479 | |
| BOLD:AAG6754 | |||||
| Pegomya solennis | CDC (2) | 0.85 | 2.67 | BOLD:ACD8686 | |
| BOLD:ACM6225 | |||||
| Pegomya winthemi | CDC (2) | 0.54 | 5.53 | BOLD:AAG1783 | |
| BOLD:ABA6845 | |||||
| Bibionidae | Bibio clavipes | CDC (2) | 1.2 | 2.46 | BOLD:ACC6151 |
| BOLD:ACR0881 | |||||
| Bibio nigriventris | CDC (2) | 1 | 3.13 | BOLD:ABX1732 | |
| BOLD:ACU5368 | |||||
| Bolitophilidae | Bolitophila austriaca | CDC (2) | 1.27 | 2.18 | BOLD:AAG4863 |
| BOLD:ACI5612 | |||||
| Ceratopogonidae | Brachypogon sociabilis | CDC (2) | 1.24 | 2.31 | BOLD:ABW3958 |
| BOLD:ACE8195 | |||||
| Ceratopogon grandiforceps | CDC (2) | 2.63 | 3.94 | BOLD:ABW3984 | |
| BOLD:ACP4327 | |||||
| Forcipomyia sp. 4ES | CDC (2) | 2.18 | 5.98 | BOLD:AAM6200 | |
| BOLD:ACQ8860 | |||||
| Chironomidae | Brillia bifida | CDC (2) | 2.31 | 6.93 | BOLD:AAD7726 |
| BOLD:ADI4999 | |||||
| Cricotopus bicinctus | CDC (2) | 1.86 | 3.2 | BOLD:AAI6018 | |
| BOLD:AAT9677 | |||||
| Gymnometriocnemus brumalis | CDC (2) | 0.5 | 2.41 | BOLD:ACD4501 | |
| BOLD:ACU9207 | |||||
| Limnophyes natalensis | CDC (2) | 1.51 | 2.89 | BOLD:AAB7361 | |
| BOLD:ACT1270 | |||||
| Limnophyes sp. 4SW | CDC (2) | 1.49 | 4.03 | BOLD:ACR9428 | |
| BOLD:ACU4225 | |||||
| Mesosmittia flexuella | CDC (2) | 0.79 | 2.02 | BOLD:ADE7569 | |
| BOLD:ACU4856 | |||||
| Orthocladius fuscimanus | CDC (2) | 2 | 2.66 | BOLD:AAV5075 | |
| BOLD:ACX3046 | |||||
| Parametriocnemus stylatus | CDC (2) | 0.76 | 2.03 | BOLD:AAI2687 | |
| BOLD:ACT9205 | |||||
| Paraphaenocladius exagitans | CDC (3) | 2.54 | 5.88 | BOLD:AAE3719 | |
| BOLD:ACQ4724 | |||||
| BOLD:ACT8523 | |||||
| Paraphaenocladius impensus | CDC (4) | 6.85 | 11.99 | BOLD:AAC4200 | |
| BOLD:ACT2714 | |||||
| BOLD:ACT5784 | |||||
| BOLD:ACU4175 | |||||
| Paratanytarsus laccophilus | CDC (2) | 2.09 | 3.14 | BOLD:AAC8842 | |
| BOLD:ACF2457 | |||||
| Polypedilum convictum | CDC (2) | 2.45 | 4.61 | BOLD:AAW4661 | |
| BOLD:ACT9278 | |||||
| Smittia reissi | CDC (2) | 1.72 | 3.47 | BOLD:ACS9748 | |
| BOLD:ACU4112 | |||||
| Conopidae | Myopa testacea | CDC (2) | 3.72 | 3.72 | BOLD:AAK8836 |
| BOLD:AAK8838 | |||||
| Dolichopodidae | Microphor anomalus | CDC (2) | 5.47 | 11 | BOLD:ACH9042 |
| BOLD:ACH9043 | |||||
| Microphor holosericeus | CDC (2) | 4.06 | 12.7 | BOLD:ACB6469 | |
| BOLD:ACH6989 | |||||
| Empididae | Hemerodromia adulatoria | CDC (2) | 8.52 | 8.52 | BOLD:ACJ6728 |
| BOLD:ACJ6729 | |||||
| Kowarzia barbatula | CDC (2) | 7.21 | 10.71 | BOLD:ACJ6935 | |
| BOLD:ACJ7236 | |||||
| Kowarzia tenella | CDC (2) | 5.39 | 10.8 | BOLD:ACJ6935 | |
| BOLD:ACJ7236 | |||||
| Ephydridae | Allotrichoma laterale | CDC (2) | 6.44 | 6.44 | BOLD:ABA8753 |
| BOLD:ACF1575 | |||||
| Ditrichophora fuscella | CDC (2) | 3.81 | 7.62 | BOLD:ABA8605 | |
| BOLD:ABA8606 | |||||
| Ditrichophora palliditarsis | CDC (2) | 3.87 | 6.57 | BOLD:AAX8675 | |
| BOLD:ABA8748 | |||||
| Halmopota salinarius | CDC (2) | 2.43 | 3.81 | BOLD:ABA7826 | |
| BOLD:ABA7827 | |||||
| Hydrellia flaviceps | CDC (2) | 4.22 | 6.33 | BOLD:ABA8652 | |
| BOLD:ABV8173 | |||||
| Philygria flavipes | CDC (2) | 1.19 | 2.03 | BOLD:ABA8663 | |
| BOLD:ACK3229 | |||||
| Polytrichophora duplosetosa | CDC (2) | 2.05 | 4.11 | BOLD:ABA8627 | |
| BOLD:ABA8628 | |||||
| Scatella obsoleta | CDC (2) | 1.25 | 2.5 | BOLD:ABA7493 | |
| BOLD:ABA7494 | |||||
| Scatophila signata | CDC (2) | 3.3 | 3.3 | BOLD:ABA7651 | |
| BOLD:ABA7652 | |||||
| Fanniidae | Fannia postica | CDC (2) | 2.35 | 7.03 | BOLD:ABW2012 |
| BOLD:ACG3518 | |||||
| Heleomyzidae | Heleomyza serrata | CDC (2) | 0.37 | 3.54 | BOLD:ABX8716 |
| BOLD:ACV1127 | |||||
| Lauxaniidae | Minettia longipennis | CDC (2) | 0.96 | 1.45 | BOLD:ACR0546 |
| BOLD:ACR0548 | |||||
| Limoniidae | Chionea lutescens | CDC (2) | 1.1 | 1.1 | BOLD:ABV5195 |
| BOLD:ADD1050 | |||||
| Euphylidorea meigenii | CDC (2) | 1.91 | 4.88 | BOLD:ABV4905 | |
| BOLD:ACU9122 | |||||
| Milichiidae | Phyllomyza equitans | CDC (2) | 1.39 | 4.05 | BOLD:ACB3455 |
| BOLD:ACD3072 | |||||
| Muscidae | Helina evecta | CDC (3) | 1.83 | 4.27 | BOLD:AAE3133 |
| BOLD:ACB3279 | |||||
| BOLD:ADB5997 | |||||
| Mydaea humeralis | CDC (2) | 1.95 | 5.84 | BOLD:AAE0058 | |
| BOLD:ACD1934 | |||||
| Mycetophilidae | Boletina dispecta | CDC (3) | 9.01 | 11.2 | BOLD:AAY5579 |
| BOLD:AAY5580 | |||||
| BOLD:AAY5581 | |||||
| Brevicornu griseicolle | CDC(2) | 9.06 | 13.6 | BOLD:ACU9474 | |
| BOLD:ABA1563 | |||||
| Brevicornu sericoma | CDC (2) | 1.99 | 4.58 | BOLD:AAY6368 | |
| BOLD:ABA1564 | |||||
| Phronia obtusa | CDC (2) | 0.83 | 1.18 | BOLD:AAY8505 | |
| BOLD:ACJ2989 | |||||
| Stigmatomeria crassicornis | CDC (2) | 0.56 | 1.86 | BOLD:AAY6370 | |
| BOLD:ACU7541 | |||||
| Zygomyia angusta | CDC (3) | 3.29 | 14.88 | BOLD:AAY5526 | |
| BOLD:AAY5527 | |||||
| BOLD:ABW0168 | |||||
| Zygomyia valida | CDC (2) | 9.51 | 14.5 | BOLD:AAY5526 | |
| BOLD:ABW0168 | |||||
| Pallopteridae | Toxoneura aff. modesta | CDC (2) | 3.41 | 5.13 | BOLD:ACB4053 |
| BOLD:ACV1580 | |||||
| Phoridae | Megaselia consetigera | CDC (2) | 0.65 | 2.63 | BOLD:ACG2938 |
| BOLD:ACX1476 | |||||
| Megaselia glabrifrons | CDC (2) | 0.66 | 1.78 | BOLD:ACG3433 | |
| BOLD:ACI6910 | |||||
| Megaselia longicostalis | CDC (3) | 1.32 | 5.72 | BOLD:AAG3263 | |
| BOLD:ADA4916 | |||||
| BOLD:AAG7025 | |||||
| Megaselia lutea | CDC (2) | 2.14 | 6.46 | BOLD:AAG3351 | |
| BOLD:ACG3608 | |||||
| Megaselia nigriceps | CDC (3) | 0.76 | 7.16 | BOLD:AAG7022 | |
| BOLD:AAY6384 | |||||
| BOLD:ACF7950 | |||||
| Megaselia pulicaria complex | CDC (3) | 5.85 | 11.96 | BOLD:AAL9073 | |
| BOLD:AAP4698 | |||||
| BOLD:AAU8534 | |||||
| Megaselia rufa | CDC (2) | 1.83 | 8.31 | BOLD:ACD9573 | |
| BOLD:ACD9606 | |||||
| Megaselia ruficornis | CDC (2) | 5.46 | 17.53 | BOLD:ACF7708 | |
| BOLD:ACG4585 | |||||
| Megaselia sepulchralis | CDC (2) | 2.27 | 4.27 | BOLD:ACF7622 | |
| BOLD:ACZ9853 | |||||
| Megaselia subpalpalis | CDC (2) | 1.05 | 2.17 | BOLD:AAL9083 | |
| BOLD:ACZ7449 | |||||
| Megaselia tarsella | CDC (3) | 0.45 | 5.61 | BOLD:ACE0332 | |
| BOLD:ACF7226 | |||||
| Psychodidae | Psychoda nr. albipennis | CDC (2) | 1.55 | 3.45 | BOLD:ABA0876 |
| BOLD:ACN5049 | |||||
| Rhinophoridae | Rhinomorinia sarcophagina | CDC (2) | 0.75 | 1.78 | BOLD:ACD9526 |
| BOLD:ACG3259 | |||||
| Sciaridae | Bradysia brevispina | CDC (2) | 2.86 | 8.4 | BOLD:ACE4845 |
| BOLD:ACI5443 | |||||
| Bradysia inusitata | CDC (2) | 6.61 | 6.61 | BOLD:ACE7273 | |
| BOLD:ACH4332 | |||||
| Bradysia praecox | CDC (2) | 1.09 | 2.35 | BOLD:ACF3561 | |
| BOLD:ACU9870 | |||||
| Bradysia regularis | CDC (2) | 0.1 | 1.67 | BOLD:ACC1391 | |
| BOLD:ACQ7807 | |||||
| Bradysia tilicola | CDC (2) | 2.87 | 6.03 | BOLD:AAN6444 | |
| BOLD:ACP0919 | |||||
| Bradysia trivittata | CDC (2) | 0.57 | 3.57 | BOLD:AAH3947 | |
| BOLD:ACB1143 | |||||
| Bradysiopsis vittata | CDC (2) | 2.24 | 4.62 | BOLD:ACC1999 | |
| BOLD:ACR0949 | |||||
| Corynoptera grothae | CDC (2) | 4.75 | 9.36 | BOLD:ACK0158 | |
| BOLD:ACO7236 | |||||
| Corynoptera luteofusca | CDC (2) | 8.16 | 11.8 | BOLD:ACJ1951 | |
| BOLD:ACQ8494 | |||||
| Corynoptera polana | CDC (2) | 1.95 | 3.81 | BOLD:ACF6941 | |
| BOLD:ACF7764 | |||||
| Corynoptera subtilis | CDC (2) | 2.91 | 6.26 | BOLD:ACD5314 | |
| BOLD:ACT9420 | |||||
| Corynoptera tetrachaeta | CDC (2) | 4.16 | 4.16 | BOLD:ACG5327 | |
| BOLD:ACL4032 | |||||
| Corynoptera tridentata | CDC (2) | 9.95 | 9.95 | BOLD:ACJ1561 | |
| BOLD:ACJ9791 | |||||
| Epidapus atomarius | CDC (2) | 0.07 | 3.98 | BOLD:ACD4767 | |
| BOLD:ACX3063 | |||||
| Leptosciarella fuscipalpa | CDC (2) | 5.24 | 9.24 | BOLD:ACE2641 | |
| BOLD:ACQ8733 | |||||
| Leptosciarella scutellata | CDC (3) | 4.84 | 7.98 | BOLD:ACD6061 | |
| BOLD:ACG4078 | |||||
| BOLD:ACI9623 | |||||
| Pnyxiopsis degener | CDC (2) | 1.83 | 5.17 | BOLD:ACE2293 | |
| BOLD:ACF9729 | |||||
| Scatopsciara neglecta | CDC (2) | 0.53 | 1.78 | BOLD:ACC7986 | |
| BOLD:ACQ2637 | |||||
| Scatopsciara subciliata | CDC (2) | 1.93 | 4.32 | BOLD:AAH4004 | |
| BOLD:ACA8369 | |||||
| Sciara hemerobioides | CDC (2) | 4.1 | 4.1 | BOLD:ACQ8933 | |
| BOLD:ACR4627 | |||||
| Trichosia morio | CDC (2) | 0.78 | 3.99 | BOLD:ACD5342 | |
| BOLD:ACO9950 | |||||
| Simuliidae | Simulium cryophilum | CDC (2) | 1.49 | 3.14 | BOLD:ACU9243 |
| BOLD:AAU1818 | |||||
| Sphaeroceridae | Opacifrons coxata | CDC (2) | 6.41 | 14 | BOLD:ACP2618 |
| BOLD:ACP5793 | |||||
| Spelobia clunipes | CDC (2) | 2.89 | 6.93 | BOLD:AAG7312 | |
| BOLD:ACF9400 | |||||
| Syrphidae | Cheilosia albipila | CDC (2) | 2.51 | 6.88 | BOLD:AAW3610 |
| BOLD:AAZ1026 | |||||
| Cheilosia chrysocoma | CDC (2) | 3.69 | 3.69 | BOLD:ABY6892 | |
| BOLD:ACJ5068 | |||||
| Cheilosia derasa | CDC (2) | 0.58 | 3.47 | BOLD:AAY9044 | |
| BOLD:AAW3649 | |||||
| Cheilosia flavipes | CDC (2) | 8.79 | 8.79 | BOLD:AAW3610 | |
| BOLD:AAY9045 | |||||
| Cheilosia impressa | CDC (2) | 1.95 | 5.74 | BOLD:AAW3651 | |
| BOLD:AAW3615 | |||||
| Cheilosia lenis | CDC (2) | 3.85 | 7.86 | BOLD:AAY8876 | |
| BOLD:AAY8875 | |||||
| Cheilosia mutabilis | CDC (2) | 1.94 | 2.74 | BOLD:AAY9746 | |
| BOLD:AAY9747 | |||||
| Cheilosia personata | CDC (2) | 1.35 | 1.88 | BOLD:ACH1700 | |
| BOLD:ACX0819 | |||||
| Cheilosia proxima | CDC (3) | 3.28 | 6.91 | BOLD:AAW3607 | |
| BOLD:AAW3651 | |||||
| BOLD:ABY8734 | |||||
| Cheilosia vernalis‐agg. | CDC (2) | 2.07 | 3.84 | BOLD:ACF0974 | |
| BOLD:ACJ5218 | |||||
| Eupeodes nitens | CDC (2) | 3.97 | 3.97 | BOLD:AAB2384 | |
| BOLD:ACH1529 | |||||
| Melanogaster nuda | CDC (2) | 0.81 | 2.44 | BOLD:AAY8880 | |
| BOLD:ACH5745 | |||||
| Merodon rufus | CDC (2) | 0.68 | 1.09 | BOLD:ADI8358 | |
| BOLD:AAQ1380 | |||||
| Paragus pecchiolii | CDC (2) | 0.96 | 4.86 | BOLD:ABA3664 | |
| BOLD:ACG8255 | |||||
| Parasyrphus punctulatus | CDC (2) | 1.11 | 2.65 | BOLD:AAZ4514 | |
| BOLD:ACG4772 | |||||
| Pipiza noctiluca | CDC (2) | 1.54 | 3.92 | BOLD:AAL4100 | |
| BOLD:ACG4983 | |||||
| Platycheirus albimanus | CDC (2) | 0.37 | 3.01 | BOLD:AAL7898 | |
| BOLD:ACJ4919 | |||||
| Sericomyia lappona | CDC (2) | 2.06 | 3.9 | BOLD:AAB1553 | |
| BOLD:ACH1641 | |||||
| Tabanidae | Tabanus bromius | CDC (2) | 2.04 | 2.93 | BOLD:AAF3864 |
| BOLD:ACJ5745 | |||||
| Tabanus glaucopis | 3.27 | 4.43 | BOLD:AAF3858 | ||
| BOLD:AAF3859 | |||||
| Tachinidae | Actia dubitata | CDC (2) | 2.36 | 2.36 | BOLD:ACP3766 |
| BOLD:ACH1972 | |||||
| Bessa selecta | CDC (2) | 1.45 | 2.38 | BOLD:ADK1760 | |
| BOLD:AAW3422 | |||||
| Cyzenis albicans | CDC (2) | 1.18 | 2.18 | BOLD:ACB0896 | |
| BOLD:ACM9631 | |||||
| Kirbya moerens | CDC (2) | 1.22 | 1.86 | BOLD:ACJ2730 | |
| BOLD:ACB0261 | |||||
| Peribaea fissicornis | CDC (2) | 2.22 | 8.17 | BOLD:ACH1961 | |
| BOLD:ACJ2910 | |||||
| Phorinia aurifrons | CDC (2) | 3.76 | 11.2 | BOLD:ADK4076 | |
| BOLD:ACB0795 |
Appendix S1 provides species names, sample IDs, BIN assignment and collection information. All project data are available under the publicly accessible DOI: http://dx.doi.org/10.5883/DS-DIPBFGBL.
3.2. Performance of the reference library for metabarcoding of Malaise trap samples
Among the 90 Malaise trap samples from the Bavarian Forest National Park (L. A. Hardulak et al. in prep.), metabarcoding revealed 1,735 dipteran OTUs, comprising 536,376 reads: 5,960 average reads per sample, matching at 97% or higher to a taxon in the DNA barcode library downloaded from BOLD (average read count was 6,928 per sample with a total of 2,809 OTUs matched to the Diptera database with ≥90%). Multiple OTU matches to a single BIN were merged. Using the Diptera data, we identified a total of 1,403 BINs including representatives of 71 families (1,385 species) within the metabarcoding data set (Appendix S2). Almost one‐third (498/1403) of these BINs belonged to “dark taxa.” Figure 2 illustrates examples of presence/absence overviews for the families Muscidae, Cecidomyiidae, Chironomidae and Syrphidae for selected Malaise trap sites.
Figure 2.

Examples from the metabarcoding results. Presence–absence overviews for three sample sites (Jos, T1‐63B and SAL) and illustrative examples for the families Cecidomyiidae, Chironomidae, Muscidae and Syrphidae [Colour figure can be viewed at http://www.wileyonlinelibrary.com]
Among families containing “dark taxa,” the percentage of unnamed taxa was inversely correlated with body size (r = −0.41, p = 0.0004) and positively with numbers of species reported from Germany (r = 0.33, p = 0.0037) (Figure 3; Appendix S4).
Figure 3.
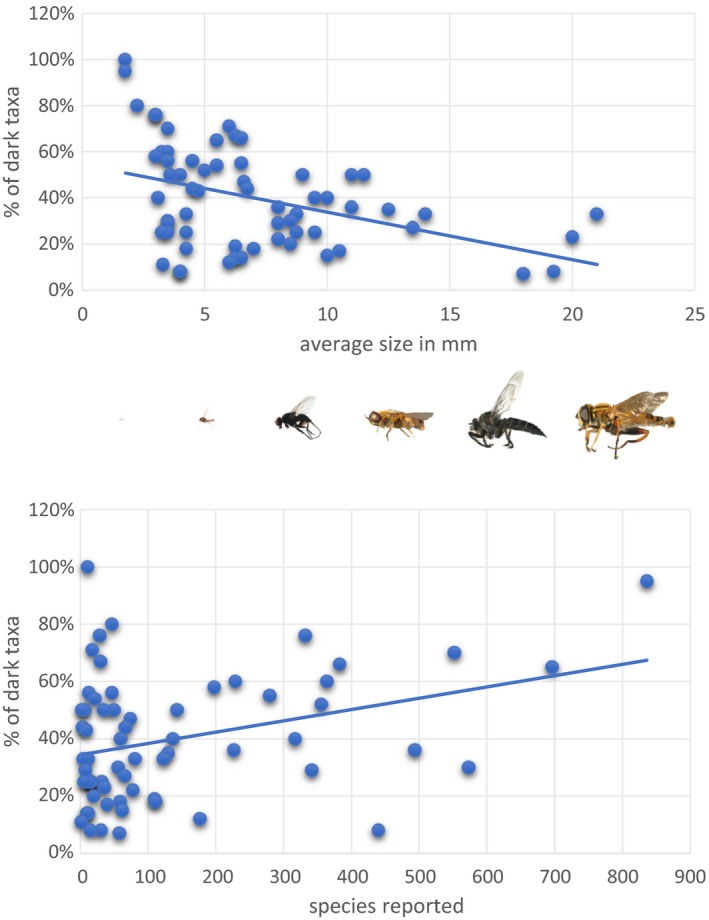
Illustration of the relationship between the percentage of “dark taxa” and average body size (mm), and in number of species reported for a family [Colour figure can be viewed at http://www.wileyonlinelibrary.com]
4. DISCUSSION
This study summarizes the results of a DNA barcoding campaign on German Diptera, work based on the characterization of 45,040 specimens. The resultant DNA barcode reference library included records for 5,200 BINs (2,453 named species comprising 2,500 BINs plus 2,700 unnamed BINs) belonging to 88 families, covering ~ 50% of the Diptera fauna reported for Germany (Schumann, 2002, 2004, 2010; Schumann et al., 1999). Until now, most of these families, especially some of the most diverse, have been taxonomically inaccessible because of the lack of specialists. By contrast, within just a few years, this study provided an interim taxonomic identification system for half of the German Diptera fauna. Although half these species still lack a Linnean name, their BIN assignments are useful “taxonomic handles” for work in ecology, conservation biology and other biodiversity research (see Geiger, Moriniere, et al., 2016). The study demonstrates the efficiency of DNA barcoding in the identification of Central European Diptera, reinforcing the results of earlier studies. DNA barcode coverage was nearly complete for many species‐poor families (e.g., Megamerinidae, Opetiidae, Phaeomyiidae) known from Germany and the incidence of “dark taxa” in these families was low. Overall, there was a strong inverse relationship between the number of “dark taxa” and average body size: the smaller the average body size of a family, the higher the ratio of “dark taxa” (Figure 3). Among families with the smallest body sizes, our results suggest a higher incidence of cryptic diversity and overlooked species, indicating the number of dipteran species in Germany is likely to be much higher than previously recognized. Among families, such as the “Iteaphila group” (Empidoidea; see Meyer & Stark, 2015), Milichiidae and Trichoceridae, DNA barcoding indicates unexpectedly high levels of diversity as their BIN count is substantially higher than the number of species known from Germany (Schumann et al., 1999). The Cecidomyiidae represent the most impressive example, as we encountered 930 BINs while only 886 species are known from Germany (Table 1; Jaschhof, 2009; Schumann et al., 1999). As such, they represent by far the largest family of Diptera in the studied area. When compared with the other families in Figure 2b, it is clear that the Cecidomyiidae show a lower average interspecific variation, indicating an increased evolutionary rate. As already proposed by Hebert et al. (2016), the extraordinary species—or BIN number—might be linked to their unusual mode of reproduction, namely haplodiploidy. Here, paternally inherited genomes of diploid males are inactivated during embryogenesis (Normark, 2003). The phenomenon of haplodiploidy is known from Hymenoptera (Branstetter et al., 2018; Hansson & Schmidt, 2018) another group known to be rate accelerated, but it is largely unstudied throughout Diptera. Despite the need for more study, we conclude the true diversity of Diptera in Germany, Europe and the world has been seriously underestimated, a conclusion reached in several other studies (Erwin, 1982; Hebert et al., 2016; May, 1988; Ødegaard, 2000).
Within the metabarcoded Malaise trap samples collected over just one season in one region of Germany, we identified 1,735 OTUs with a sequence identity higher than 97% to a dipteran record. This result indicates that metabarcode analysis of bulk samples will be a valuable approach for assessing the diversity of Diptera in Germany (Appendix S2). Variation in overall biodiversity between sampling sites as well as annual phenologies of certain taxa can easily be visualized using presence–absence maps (Figure 2). This will be a useful feature for comparison of large data sets and for monitoring beneficial or pest insects (L. A. Hardulak et al. in preparation). Although a third of the OTUs within the metabarcoding data set could not be assigned to a Linnean species, interim names, such as BIN assignments, make it possible to compare sampling sites. OTUs with lower sequence similarities (<97%) to known taxa can be used to track “dark taxa,” those species missing from the reference sequence library. Although such taxa may only be assigned to a family or genus, their records are still valuable for evaluating differences between samples from various environments or sites. At present, dipteran species, although overall present in very high numbers, are extremely under‐represented within environmental assessments in Germany: ~2,000 species from 11 families (Asilidae, Atelestidae, Ceratopogonidae, Chaoboridae, Dixidae, Dolichopodidae, Empididae, Hybotidae, Psychodidae, Syrphidae, Thaumaleidae) are included in the German red list (Gruttke et al., 2016), but not a single dipteran species is listed among the ~1,000 species being protected according to the European Flora‐Fauna‐Habitat directive (Council Directive 92/43/EEC on the Conservation of natural habitats and of wild fauna and flora, 1992), which ensures the conservation of a wide range of rare, threatened or endemic animal and plant species in Europe. The present study is a first step to permit the proper evaluation of the status of dipterans and the potential designation of some species as targets for conservation action.
Previous studies have shown the great potential of metabarcoding for biotic assessments in various contexts, including Malaise trap surveys (Morinière et al., 2016), biosurveillance of invasive and pest species (Ashfaq & Hebert, 2016; L. A. Hardulak et al. in prep), macrozoobenthos sampling for assessing water and stream health (Elbrecht & Leese, 2015; Serrana, Miyake, Gamboa, & Watanabe, 2018), faeces analyses for dietary inference (De Barba et al., 2014; Hawlitschek, Fernández‐González, Balmori‐de la Puente, & Castresana, 2018), species identification for forensic entomology (Chimeno et al., 2018) and for soil biology (Oliverio, Gan, Wickings, & Fierer, 2018). This approach combines the advantages of DNA barcoding, namely the capacity to identify any life stage, body fragment or even trace DNA in the environment, with the ability of high‐throughput sequencers to analyse millions of DNA fragments and thousands of specimens at a time. The application of this technology to biodiversity assessments will certainly enable species surveys at larger scales, shorter time and lower costs compared with classical morphological approaches (Douglas et al., 2012; Hajibabaei et al., 2011; Ji et al., 2013; Taberlet, Coissac, Pompanon, Brochmann, & Willerslev, 2012). The ability to upscale biomonitoring projects is crucial, as is the need to generate biodiversity data fast and with less dependence on often unavailable taxonomic experts. Additionally, data generated by ongoing metabarcoding studies, such as from annual national biomonitoring projects, can be combined and reanalysed, producing recursively more comprehensive species lists, when new reference sequences become available or when taxonomic annotations have been improved. While biomonitoring studies have traditionally employed small subsets of indicator species, metabarcoding will enable comprehensive assessments of biodiversity because even “dark taxa” can be tracked. Furthermore, metabarcoding can enhance the ability to rapidly assess biodiversity patterns to identify regions that are of most significance for conservation.
Although this project aimed to develop a comprehensive DNA barcode library, resource constraints meant that only half the specimens sorted to a family or better taxonomy could be analysed. It is certain that many species and genera currently absent from the reference library remain within this sorted material, making the remaining samples a valuable resource for future extension of the reference library. Our work has also highlighted the potential of DNA barcoding and metabarcoding to aid efforts to conserve the world's fauna. Because these technologies greatly enhance our ability to identify, and thus conserve, biodiversity, they should be pursued—vigorously. As our study has provided several thousands of voucher‐based DNA barcode records, we invite the global community of dipteran taxonomists to improve identifications for the many “dark taxa” encountered in our study by identifying these vouchers using reverse taxonomic approaches.
The present study represents an important component of a decade of work directed toward creating a comprehensive DNA barcode library for German animal species. Because Diptera represents the largest and taxonomically most challenging insect order, they have received less attention than other orders (e.g., Lepidoptera, Coleoptera, freshwater orders) with lower species richness and more taxonomic expertise. Our work on Diptera has not only confirmed that this order is extremely species‐rich, but also that several of its most diverse families include a large proportion of “dark taxa.” The present study represents a cornerstone for subsequent research on these unexplored groups of Diptera. This paper presents the results of one of the most comprehensive studies on DNA barcoding of Diptera, with a coverage of over 80% of German families. Due to the general lack of taxonomy in many groups of Diptera, only a fraction of the specimens could be identified to species level. Most specimens for the study were obtained from just three Malaise traps deployed as a component of the Global Malaise programme (see http://biodiversitygenomics.net/projects/gmp/). Voucher specimens are still being identified by external specialists, a process that is labour intensive and time consuming, especially for taxonomically challenging taxa.
Our study presents results from one of the most comprehensive DNA barcoding projects on Diptera, a megadiverse, and, almost certainly, most diverse insect order. Our results strongly support the conclusion that DNA barcoding will enable the discovery and identification of most dipteran taxa. Some cases of low interspecific variation were observed in the Syrphidae, Tachinidae and Calliphoridae where additional markers may be needed for species identification (Haarto & Ståhls, 2014; Nelson et al., 2012; Pohjoismäki et al., 2016; Whitworth et al., 2007). However, in most cases, there was congruence between BINs and species defined by traditional morphological methods, supporting the use of DNA barcoding as a species identification tool for Diptera. This conclusion and the finding that many of the species we encountered represent “dark taxa” indicates that DNA barcoding will speed the discovery of genetic entities that will eventually gain recognition as biological species. Our data release aims at making these results accessible to the scientific community through a public data portal so they will be available for taxonomic research, biodiversity studies and barcoding initiatives at national and international levels.
In summary, the application of DNA barcoding enabled a comprehensive assessment of German Diptera, including several highly diverse families, which would otherwise have been excluded due to a lack of taxonomic expertise. By selecting morphospecies from the pool of specimens collected by the year‐long deployment of Malaise traps in ecosystems ranging from alpine to lowland settings, we constructed a reference library for most dipteran families known from Germany. Due to the diversity of sampling sites, we encountered a wide range of taxa from microendemics to wide‐ranging generalists with varied seasonal phenologies. We emphasize that DNA barcoding and the resultant barcode reference libraries provide an easy, intuitive introduction to molecular genetics, an approach accessible to undergraduate students in a way that genome sequencing is not. Because DNA barcoding workflows have been implemented in many laboratories around the world and because current primer sets reliably generate amplicons, this method is ideal for educational purposes. Democratization of the method, the analytical tools and data through the BOLD database (Ratnasingham & Hebert, 2007) further facilitates its use in real world situations. The approach has the additional advantage of allowing students to not only work with “real organisms,” but also to solve long‐standing taxonomic puzzles. The latter work leads students to probe the historical literature, to regale in past expeditions in search of type locations or type material, and potentially to end the chase by describing a new species. However, it is critical that senior taxonomists and professors need to recognize these possibilities and encourage their students to embrace this approach as it offers such a clear solution to the taxonomic impediment.
Germany has a tradition of more than 250 years of entomological research, and the number of Diptera species recorded is the highest for any European country comprising almost half of the European fauna. Despite this long effort, knowledge of its Diptera fauna must be regarded as fragmentary. In accordance with the species accumulation curve presented by Pape (2009) for the British Isles, additional species were revealed from current collecting efforts for practically every species‐rich family. Recording “new” species is slowed by the lack of experts for many of these families as well as by the lack of up‐to‐date identification keys. A particularly important result of our study is that the estimated number of dipteran species in Germany is certainly much higher than formerly thought. High proportions of unrecorded species were evident for the Agromyzidae, Anthomyiidae, Cecidomyiidae, Ceratopogonidae, Chironomidae, Chloropidae, Phoridae, Sciaridae and Sphaeroceridae, and to a lesser extent for the Empidoidea, Limoniidae, Mycetophilidae and others. Further studies point to an enormous under‐estimation of the species diversity in the Cecidomyiidae (Borkent et al., 2018; Hebert et al., 2016). Although our data do not allow for an accurate projection for the size of the total species numbers, it seems quite likely that this single family contains thousands of unrecorded species in Germany.
AUTHOR CONTRIBUTIONS
Obtained funding: G.H., W.W., A.H., P.D.N.H. Collected the samples: D.D., B.R. Conceived and designed the experiments: J.M., L.A.H., M.G., B.R. Analysed the data: J.M., L.A.H., M.F.G., L.R., B.R. Wrote the paper: J.M., L.A.H., S.S., M.B., D.D. Contributed (additions/corrections) to the manuscript: P.D.N.H., A.H., M.F.G., L.H., G.H.
5.
Table 3.
All cases of low intraspecific sequence variation at COI; cases of BIN sharing (BS)
| Family | Species | BS rank | Mean intraspecific variation | Max intraspecific | BIN |
|---|---|---|---|---|---|
| Anthomyiidae | Hylemya nigrimana | BS (2) | 0.34 | 0.52 | BOLD:ABA6492 |
| Hylemya vagans | 0.37 | 1.58 | |||
| Calliphoridae | Calliphora loewi | BS (2) | 1.07 | 1.07 | BOLD:AAB6579 |
| Calliphora vicina | 0.84 | 2.59 | |||
| Lucilia caesar | BS (3) | 0.95 | 3.07 | BOLD:AAA7470 | |
| Lucilia caesarillustris | 0.7 | 2.43 | |||
| Lucilia illustris | N/A | 0 | |||
| Dolichopodidae | Medetera petrophiloides | BS (2) | 0.35 | 1.22 | BOLD:ACA1124 |
| Medetera truncorum | N/A | 0 | |||
| Sphyrotarsus argyrostomus | BS (2) | 0.91 | 1.37 | BOLD:ADB6106 | |
| Sphyrotarsus hygrophilus | N/A | 0 | |||
| Empididae | Kowarzia madicola | BS (2) | 0 | 0 | BOLD:ACJ7236 |
| Kowarzia tenella | 5.39 | 10.8 | |||
| Kowarzia barbatula | BS (2) | 4.8 | 11.3 | BOLD:ACJ6935 | |
| Kowarzia tenella | 5.39 | 10.8 | |||
| Ephydridae | Allotrichoma bezzii | BS (4) | 0.13 | 0.31 | BOLD:ACF1575 |
| Allotrichoma filiforme | 0.08 | 0.15 | |||
| Allotrichoma laterale | 6.44 | 6.44 | |||
| Allotrichoma schumanni | 0 | 0 | |||
| Ephydra macellaria | BS (3) | N/A | 0 | BOLD:AAG2729 | |
| Ephydra murina | N/A | 0 | |||
| Ephydra riparia | 2.83 | 2.83 | |||
| Hydrellia nigricans | BS (2) | 0.23 | 0.31 | BOLD:ABA8624 | |
| Hydrellia subalbiceps | 0.31 | 0.46 | |||
| Notiphila cinerea | BS (2) | 0.26 | 0.46 | BOLD:ABA7513 | |
| Notiphila graecula | 0 | 0 | |||
| Notiphila riparia | BS (2) | 0.16 | 0.35 | BOLD:AAX5585 | |
| Notiphila subnigra | 0.41 | 0.62 | |||
| Philygria flavipes | BS (2) | 1.19 | 2.03 | BOLD:ACK3229 | |
| Philygria punctatonervosa | 0.15 | 0.15 | |||
| Psilopa compta | BS (2) | 0.08 | 0.16 | BOLD:AAG6948 | |
| Psilopa nitidula | 0.38 | 0.77 | |||
| Iteaphila‐group | Anthepiscopus indet. | BS (2) | 0.14 | 0.48 | BOLD:ACD9492 |
| Anthepiscopus sp. 1 | 11.3 | 11.3 | |||
| Anthepiscopus sp. 1 | BS (2) | 11.3 | 11.3 | BOLD:ACJ7111 | |
| Anthepiscopus sp. 4 | 0.91 | 1.58 | |||
| Iteaphila sp. 1 | BS (2) | 0.07 | 0.15 | BOLD:ACD3033 | |
| Iteaphila sp. 2 | 4.49 | 9.77 | |||
| Lonchopteridae | Lonchoptera lutea | BS (2) | 0.39 | 1.09 | BOLD:ABX0277 |
| Lonchoptera nitidifrons | N/A | 0 | |||
| Muscidae | Hydrotaea dentipes | BS (2) | 2.15 | 9.78 | BOLD:AAZ9882 |
| Hydrotaea similis | 0 | 0 | |||
| Mycetophilidae | Boletina gripha | BS (2) | 0.52 | 0.9 | BOLD:AAF6783 |
| Boletina groenlandica | N/A | 0 | |||
| Mycetophila distigma | BS (2) | N/A | 0 | BOLD:AAY8340 | |
| Mycetophila flava | 0.19 | 0.19 | |||
| Zygomyia angusta | BS (2) | 4.6 | 15.4 | BOLD:AAY5526 | |
| Zygomyia valida | 14.5 | 14.5 | |||
| Zygomyia angusta | BS (2) | 4.6 | 15.4 | BOLD:ABW0168 | |
| Zygomyia valida | 14.5 | 14.5 | |||
| Phoridae | Triphleba bicornuta | BS (2) | N/A | 0 | BOLD:ACF0365 |
| Triphleba sp. BOLD:ACF0365 | 0.66 | 1.22 | |||
| Sarcophagidae | Sarcophaga depressifrons | BS (2) | 0 | 0 | BOLD:ABV4597 |
| Sarcophaga haemorrhoa | 0.47 | 0.7 | |||
| Simuliidae | Simulium balcanicum | BS (2) | N/A | 0 | BOLD:AAM4036 |
| Simulium equinum | 1.59 | 2.66 | |||
| Syrphidae | Baccha elongata | BS (6) | N/A | 0 | BOLD:ABA3006 |
| Baccha elongata s.s. | 0 | 0 | |||
| Baccha obscuripennis | 1.23 | 2.02 | |||
| Baccha sp. BOLDABA3006 | N/A | 0 | |||
| Brachypalpus laphriformis | 0.56 | 1.54 | BOLD:AAY9039 | ||
| Brachypalpus valgus | N/A | 0 | |||
| Cheilosia albipila | BS (2) | 2.51 | 6.88 | BOLD:AAW3610 | |
| Cheilosia flavipes | 8.79 | 8.79 | |||
| Cheilosia barbata | BS (3) | 0.1 | 0.3 | BOLD:AAW3615 | |
| Cheilosia impressa | 1.95 | 5.74 | |||
| Cheilosia sp. BOLDAAW3615 | 0 | 0 | |||
| Cheilosia chloris | BS (8) | 0.57 | 1.42 | BOLD:ACF0974 | |
| Cheilosia chlorus | 0.12 | 0.18 | |||
| Cheilosia chlorus‐group | N/A | 0 | |||
| Cheilosia fraterna | 0.55 | 0.87 | |||
| Cheilosia melanura | 0.06 | 0.2 | |||
| Cheilosia ruficollis | N/A | 0 | |||
| Cheilosia sp. BOLDACF0974 | 0.47 | 0.71 | |||
| Cheilosia vernalis‐agg. | 2.07 | 3.84 | |||
| Cheilosia crassiseta | BS (6) | N/A | 0 | BOLD:AAW3647 | |
| Cheilosia impudens | N/A | 0 | |||
| Cheilosia nigripes | N/A | 0 | |||
| Cheilosia sp. BIOUG17085‐G07 | 0.75 | 1.94 | |||
| Cheilosia aff. grisella | N/A | 0 | |||
| Cheilosia antiqua | N/A | 0 | |||
| Cheilosia faucis | BS (2) | 0.7 | 0.88 | BOLD:AAY8874 | |
| Cheilosia nivalis | 0 | 0 | |||
| Cheilosia grisella | BS (2) | 0.18 | 0.18 | BOLD:AAW3619 | |
| Cheilosia pubera | 0.49 | 0.87 | |||
| Cheilosia canicularis | BS (2) | 0.08 | 0.38 | BOLD:ACI2500 | |
| Cheilosia montana | N/A | 0 | |||
| Cheilosia carbonaria | BS (2) | 0.37 | 0.37 | BOLD:AAY8876 | |
| Cheilosia lenis | 3.85 | 7.86 | |||
| Chrysotoxum bicinctum | BS (2) | 0.86 | 2 | BOLD:AAJ0967 | |
| Chrysotoxum festivum | 0 | 0 | |||
| Dasysyrphus hilaris | BS (3) | 0.35 | 0.52 | BOLD:AAA7375 | |
| Dasysyrphus laskai | 0.3 | 0.3 | |||
| Dasysyrphus venustus | N/A | 0 | |||
| Dasysyrphus lenensis | BS (3) | 0.58 | 0.58 | BOLD:AAB2865 | |
| Dasysyrphus pinastri | 1.25 | 2.1 | |||
| Dasysyrphus sp. BOLDAAB2865 | 0.12 | 0.17 | |||
| Eupeodes bucculatus | BS (5) | 1.14 | 3.13 | BOLD:AAB2384 | |
| Eupeodes nielseni | 0.15 | 0.37 | |||
| Eupeodes nitens | 3.97 | 3.97 | |||
| Eupeodes sp. BOLDAAB2384 | 0.39 | 1.03 | |||
| Eupeodes luniger | 0.53 | 1.05 | |||
| Melanogaster aerosa | BS (2) | N/A | 0 | BOLD:AAQ4015 | |
| Melanogaster hirtella | 0.26 | 0.7 | |||
| Melanostoma dubium | BS (7) | 0 | 0 | BOLD:AAB2866 | |
| Melanostoma mellinum | 0.58 | 1.21 | |||
| Melanostoma mellinum‐agg. | N/A | 0 | |||
| Melanostoma scalare | 0.49 | 1.3 | |||
| Melanostoma sp. A | 0 | 0 | |||
| Melanostoma sp. B | 0.11 | 0.16 | |||
| Melanostoma sp. BOLDAAB2866 | 0.63 | 2.69 | |||
| Merodon avidus | BS (2) | N/A | 0 | BOLD:AAQ1379 | |
| Merodon avidus B | 0.55 | 1.03 | |||
| Paragus aff. haemorrhous | BS (5) | N/A | 0 | BOLD:ABZ4619 | |
| Paragus constrictus | N/A | 0 | |||
| Paragus haemorrhous | 0.26 | 0.87 | |||
| Paragus sp. BOLDABZ4619 | 0.07 | 0.37 | |||
| Paragus tibialis | N/A | 0 | |||
| Paragus majoranae | BS (2) | 0.87 | 0.87 | BOLD:ABA3664 | |
| Paragus pecchiolii | 0.96 | 4.86 | |||
| Parasyrphus lineola | BS (2) | 0.19 | 0.39 | BOLD:ACE7140 | |
| Parasyrphus vittiger | 0.63 | 1.44 | |||
| Pipiza bimaculata | BS (4) | N/A | 0 | BOLD:AAL4100 | |
| Pipiza nocticula | N/A | 0 | |||
| Pipiza noctiluca‐agg. | N/A | 0 | |||
| Pipiza sp. BOLDAAL4100 | 0.55 | 1.65 | |||
| Platycheirus angustatus | BS (3) | 0.84 | 2.02 | BOLD:ACF4733 | |
| Platycheirus europaeus | 1.95 | 1.95 | |||
| Platycheirus sp. BOLDACF4733 | 0.21 | 1.15 | |||
| Platycheirus clypeatus | BS (5) | 0.38 | 0.88 | BOLD:AAA9506 | |
| Platycheirus fulviventris | 1.04 | 1.04 | |||
| Platycheirus occultus | 0.51 | 1.04 | |||
| Platycheirus perpallidus | N/A | 0 | |||
| Platycheirus sp. BOLDAAA9506 | 0.9 | 2.03 | |||
| Platycheirus melanopsis | BS (2) | 0.25 | 0.62 | BOLD:AAP0412 | |
| Platycheirus tatricus | N/A | 0 | |||
| Platycheirus nielseni | BS (3) | 0 | 0 | BOLD:AAC6630 | |
| Platycheirus peltatus | 0.24 | 0.72 | |||
| Platycheirus peltatus‐group | N/A | 0 | |||
| Platycheirus scutatus | BS (3) | 0.05 | 0.19 | BOLD:AAG4665 | |
| Platycheirus scutatus‐group | 0.44 | 0.71 | |||
| Platycheirus splendidus | N/A | 0 | |||
| Scaeva dignota | BS (2) | N/A | 0 | BOLD:AAF2374 | |
| Scaeva pyrastri | 0.25 | 0.91 | |||
| Scaeva pyrastri | BS (2) | 0.25 | 0.91 | BOLD:AAF2374 | |
| Scaeva dignota | N/A | 0 | |||
| Sericomyia lappona | BS (2) | 2.06 | 3.9 | BOLD:AAB1553 | |
| Sericomyia silentis | 0.05 | 0.24 | |||
| Sphaerophoria bankowskae | BS (9) | N/A | 0 | BOLD:AAA7374 | |
| Sphaerophoria infuscata | 0.24 | 0.38 | |||
| Sphaerophoria interrupta | 0 | 0 | |||
| Sphaerophoria interrupta‐group | 0.49 | 0.75 | |||
| Sphaerophoria philanthus | N/A | 0 | |||
| Sphaerophoria rueppellii | N/A | 0 | |||
| Sphaerophoria sp. BOLDAAA7374 | 0.31 | 6.54 | |||
| Sphaerophoria taeniata | N/A | 0 | |||
| Sphaerophoria virgata | N/A | 0 | |||
| Sphegina montana | BS (2) | N/A | 0 | BOLD:ABX4867 | |
| Sphegina sibirica | 0.4 | 0.41 | |||
| Temnostoma apiforme | BS (2) | 0.52 | 0.52 | BOLD:AAV6543 | |
| Temnostoma meridionale | 0.35 | 0.52 | |||
| Stratiomyidae | Beris geniculata | BS (2) | N/A | 0 | BOLD:AAW3384 |
| Beris morrisii | 0.48 | 1.47 | |||
| Tachinidae | Lydella stabulans | BS (2) | 0.12 | 0.44 | BOLD:AAP8653 |
| Lydella thompsoni | 0.68 | 1.31 | |||
| Medina luctuosa | BS (3) | 1.35 | 1.35 | BOLD:AAG6902 | |
| Medina melania |
DATA AVAILABILITY
All specimen data have been made publicly available within the BOLD workbench ‐ a DOI for the dataset has been added.
Supporting information
ACKNOWLEDGEMENTS
We express our extreme gratitude to the taxonomists, citizen scientists and nature enthusiasts who supported this campaign by collecting thousands of dipteran species. The realization of this mammoth task would not have been possible without the help of Adaschkiewitz, W., Assum, Babiy, P. P., Bährmann, R., Baranov, V., Beermann, A., Behounek, G., Belleuvre, N., Blick, T., Bolz, R., Brandt‐Floren, C., Brenzinger, S., Brown, A., Burmeister E. G., Charabidze, D., Chimeno, C., Claussen, C., Dettinger‐Klemm, A., Diller, E., Drozd, P., Dunz, A., Duschl Miesbach, M., Dworschak, W., Eckert, I., Esser, J., Fahldieck, M., Fiedler, Fittkau, E. J. (+), Flügel, H. J.,Forster, W., Forstner, P., Fünfstück, J., Fütterer, S., Fuhrmann, S., Gabriel, I., Gammelmo, O., Gerecke, R., Glaw, F., Guggemoos, T., , Haberberger, S., Hable, J., Haeselbarth, E., Hansen, L. O., Hartop, E., Hawlitschek, O., Heller, K., Hessing, R., Hierlmeier, V., Hilbig, D., Höglund, J. R., Höhne, F., Honold, D., Jaschof, M., Jon, T., Kamin, J., Kappert, J., Kehlmaier, C., Kilian, D., Kirsch, H., Kjaerandsen, J., Kleiner, M., Koehler, F., Koehler, J., Kölbl, N., Koenig, T., Kolbeck, H., Kraus, G., Kraus, W., Kuehbandner, M., Kuehlhorn, F., Kuhlmann, M., Kusdas, K., Kvifte, G. M., Lindner, S., Loennve, O. J., Lucas, W., Mair, K., Mandery, K., Mengual, X., Merkel‐Wallner, G., Mortelmans, J., Müller, Mueller, R., Mueller‐Kroehling, S., Neumann, C., Olberg, S., Olsen, K. M., Pavlova, A., Pötter, L., Steven, M., Plassmann, E., Podhorna, J., Prescher, S., Prozeller, M., Pushkar, V., Reckel, F., Rehm, T., Reiff, N., Reimann, A., Reiso, S., Rennwald, K., Richter, B., Riedel, G., Rohrmoser, S., Rozo, P., Rudzinksi, H. G., Ruf, T., Salomon, C., Schacht, W. (+), Schäfer, A., Scheingraber, M., Scheler, Schmieder, F., Schödl, M., Schoenitzer, K., Schrott, S., Schubart, C., Schubert, C., Schubert, W., Schwarz, K., Schwemmer, R., Sedlak, G., Segerer, A., Sellmayer, G., Spelda, J., Spies, M., Ssymank, A., Steigemann, U., Stenhouse, G., Stoecklein, F., Stuke, J. H., Stur, E., Tänzler, K., Tänzler, R., Telfer, A., Toussaint, C., Toussaint, E., Treiber, R., Troester, M., v. Tschirnhaus, M., v. d. Dunk, K., , Vallenduuk, H., van Ess, L., Velterop, J., Voith, J., Volf, M., Wachtel, F., Wagner, R., Warncke, K., Weber, D., Weiffenbach, H., Weigand, A. M., Weixler, K., Wiedenbrug, S., Windmaisser, T., Winqvist, K., Woodley, N., E., Zahn, A. The project was funded by grants from the Bavarian State Ministry of Science and the Arts (2009‐2018: Barcoding Fauna Bavarica, BFB) and the German Federal Ministry of Education and Research (German Barcode of Life: 2012‐2019, BMBF FKZ 01LI1101 and 01LI1501). We are grateful to the team at the Centre for Biodiversity Genomics in Guelph (Ontario, Canada) for their great support and help and particularly to Sujeevan Ratnasingham for developing the BOLD database (BOLD; http://www.boldsystems.org) infrastructure and the BIN management tools. The sequencing work was supported, in part, by funding from the Government of Canada to Genome Canada through the Ontario Genomics Institute, while the Ontario Ministry of Research and Innovation and NSERC supported development of the BOLD informatics platform. We also thank all the students who assisted in the ZSM‐barcoding projects (barcoding‐zsm.de) for picking countless legs and photographing countless specimens. We would like to express our thanks to Dr Vedran Bozicevic (AIM GmbH, Munich, Germany) for assisting with the KRONA file to enable inspection of BIN diversity. Fieldwork permits were issued by the responsible state environmental offices of Bavaria (Bayerisches Staatsministerium für Umwelt und Gesundheit, Munich, Germany, project: “Barcoding Fauna Bavarica”; confirmed by the regional governments “Bezirksregierungen”) and Rhineland‐Palatinate (“Struktur‐ und Genehmigungsdirektion Nord”, Axel Schmidt [Koblenz, Germany]).
Morinière J, Balke M, Doczkal D, et al. A DNA barcode library for 5,200 German flies and midges (Insecta: Diptera) and its implications for metabarcoding‐based biomonitoring. Mol Ecol Resour. 2019;19:900–928. 10.1111/1755-0998.13022
Data Availability Statement: All specimen data have been made publicly available within the BOLD workbench ‐ a DOI for the dataset has been added.
Contributor Information
Jérôme Morinière, Email: moriniere@snsb.de.
Dieter Doczkal, Email: doczkal@snsb.de.
REFERENCES
- Ashfaq, M. , & Hebert, P. D. N. (2016). DNA barcodes for bio‐surveillance: Regulated and economically important arthropod plant pests. Genome, 59(11), 933–945. 10.1139/gen-2016-0024 [DOI] [PubMed] [Google Scholar]
- Ashfaq, M. , Hebert, P. D. N. , Mirza, J. H. , Khan, A. M. , Zafar, Y. , & Mirza, M. S. (2014). Analyzing mosquito (Diptera: Culicidae) diversity in Pakistan by DNA barcoding. PLoS ONE, 9(5), e97268 10.1371/journal.pone.0097268 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Astrin, J. J. , Höfer, H. , Spelda, J. , Holstein, J. , Bayer, S. , Hendrich, L. , … Muster, C. (2016). Towards a DNA barcode reference database for spiders and harvestmen of Germany. PLoS ONE, 11(9), e0162624 (24 pp, supplements). 10.1371/journal.pone.0162624 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bickel D., Pape T., & Meier R. (Eds.) (2009). Diptera diversity: status, challenges and tools (pp. 459). Leiden, Netherlands: Brill. [Google Scholar]
- Borkent, A. , Brown, B. , Adler, P. H. , Amorim, D. D. S. , Barber, K. , Bickel, D. , … Capellari, R. S. (2018). Remarkable fly (Diptera) diversity in a patch of Costa Rican cloud forest. Zootaxa, 4402(1), 53–90. [DOI] [PubMed] [Google Scholar]
- Branstetter, M. G. , Childers, A. K. , Cox‐Foster, D. , Hopper, K. R. , Kapheim, K. M. , Toth, A. L. , & Worley, K. C. (2018). Genomes of the Hymenoptera. Current Opinion in Insect Science, 25, 65–75. 10.1016/j.cois.2017.11.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brix, S. , Leese, F. , Riehl, T. , & Kihara, T. C. (2015). A new genus and new species of Desmosomatidae Sars, 1897 (Isopoda) from the eastern South Atlantic abyss described by means of integrative taxonomy. Marine Biodiversity, 45(1), 7–61. 10.1007/s12526-014-0218-3 [DOI] [Google Scholar]
- Carew, M. E. , Pettigrove, V. , Cox, R. L. , & Hoffmann, A. A. (2007). DNA identification of urban Tanytarsini chironomids (Diptera: Chironomidae). Journal of the North American Benthological Society, 26(4), 587–600. 10.1899/06-120.1 [DOI] [Google Scholar]
- Carew, M. E. , Pettigrove, V. , & Hoffmann, A. A. (2005). The utility of DNA markers in classical taxonomy: Using cytochrome oxidase I markers to differentiate Australian Cladopelma (Diptera: Chironomidae) midges. Annals of the Entomological Society of America, 98(4), 587–594. [Google Scholar]
- Chimeno, C. , Morinière, J. , Podhorna, J. , Hardulak, L. , Hausmann, A. , Reckel, F. , … Haszprunar, G. (2018). DNA barcoding in forensic entomology – Establishing a DNA reference library of potentially forensic relevant arthropod species. Journal of Forensic Sciences, 64(2), 593–601. 10.1111/1556-4029.13869 [DOI] [PubMed] [Google Scholar]
- Chivian E., & Bernstein A. (Eds.) (2008). Sustaining life: How human health depends on biodiversity. Oxford, UK: Oxford University Press. [Google Scholar]
- Cranston, P. S. , Ang, Y. C. , Heyzer, A. , Lim, R. B. H. , Wong, W. H. , Woodford, J. M. , & Meier, R. (2013). The nuisance midges (Diptera: Chironomidae) of Singapore's Pandan and Bedok reservoirs. Raffles Bulletin of Zoology, 61(2), 779–793. [Google Scholar]
- Cruaud, P. , Rasplus, J. Y. , Rodriguez, L. J. , & Cruaud, A. (2017). High‐throughput sequencing of multiple amplicons for barcoding and integrative taxonomy. Scientific Reports, 7, 41948 10.1038/srep41948 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cywinska, A. , Hunter, F. F. , & Hebert, P. D. (2006). Identifying Canadian mosquito species through DNA barcodes. Medical and Veterinary Entomology, 20(4), 413–424. 10.1111/j.1365-2915.2006.00653.x [DOI] [PubMed] [Google Scholar]
- Dathe, H. H. , & Blank, S. M. . (2004). Nachträge zum Verzeichnis der Hautflügler Deutschlands, Entomofauna Germanica Band 4 (Hymenoptera). (1). Entomologische Nachrichten Und Berichte, 48(3–4), 179–182. [Google Scholar]
- De Barba, M. , Miquel, C. , Boyer, F. , Mercier, C. , Rioux, D. , Coissac, E. , & Taberlet, P. (2014). DNA metabarcoding multiplexing and validation of data accuracy for diet assessment: Application to omnivorous diet. Molecular Ecology Resources, 14(2), 306–323. 10.1111/1755-0998.12188 [DOI] [PubMed] [Google Scholar]
- de Carvalho, M. R. , Bockmann, F. A. , Amorim, D. S. , Brandão, C. R. F. , de Vivo, M. , de Figueiredo, J. L. , … Nelson, G. J. (2007). Taxonomic impediment or impediment to taxonomy? A commentary on systematics and the cybertaxonomic‐automation paradigm. Evolutionary Biology, 34(3), 140–143. 10.1007/s11692-007-9011-6 [DOI] [Google Scholar]
- DeWaard, J. R. , Ivanova, N. V. , Hajibabaei, M. , & Hebert, P. D. N. (2008). Assembling DNA barcodes. Environmental Genomics, 410, 275–294. [DOI] [PubMed] [Google Scholar]
- DeWaard, J. R. , Levesque‐Beaudin, V. , deWaard, S. L. , Ivanova, N. V. , McKeown, J. T. A. , Miskie, R. , … Hebert, P. D. N. (2019). Expedited assessment of terrestrial arthropod diversity by coupling Malaise traps with DNA barcoding. Genome, 62(3), 85–95. 10.1139/gen-2018-0093 [DOI] [PubMed] [Google Scholar]
- Doczkal, D. (2017). Vorsortierung der Proben und Vollständigkeit der Erfassung In Ssymank A., & Doczkal D. (Eds.), Biodiversität des südwestlichen Dinkelbergrandes und des Rheintals bei Grenzach‐Whylen, eine Bestandsaufnahme im südwestlichen Einfallstor Deutschlands für neue Arten in der Folge des Klimawandels. Mauritiana (Altenburg) 34, 900–910. [Google Scholar]
- Douglas, W. Y. , Ji, Y. , Emerson, B. C. , Wang, X. , Ye, C. , Yang, C. , & Ding, Z. (2012). Biodiversity soup: Metabarcoding of arthropods for rapid biodiversity assessment and biomonitoring. Methods in Ecology and Evolution, 3(4), 613–623. 10.1111/j.2041-210X.2012.00198.x [DOI] [Google Scholar]
- Eiseman, C. S. , Heller, K. , & Rulik, B. (2016). A new leaf‐mining dark‐winged fungus gnat (Diptera: Sciaridae), with notes on other insect associates of marsh marigold (Ranunculaceae: Caltha palustris L.). Proceedings of the Entomological Society of Washington, 118(4), 519–533. [Google Scholar]
- Ekrem, T. , Stur, E. , & Hebert, P. D. N. (2010). Females do count: Documenting Chironomidae (Diptera) species diversity using DNA barcoding. Organisms Diversity & Evolution, 10(5), 397–408. 10.1007/s13127-010-0034-y [DOI] [Google Scholar]
- Ekrem, T. , Willassen, E. , & Stur, E. (2007). A comprehensive DNA sequence library is essential for identification with DNA barcodes. Molecular Phylogenetics and Evolution, 43(2), 530–542. 10.1016/j.ympev.2006.11.021 [DOI] [PubMed] [Google Scholar]
- Elbrecht, V. , & Leese, F. (2015). Can DNA‐based ecosystem assessments quantify species abundance? Testing primer bias and biomass—sequence relationships with an innovative metabarcoding protocol. PLoS ONE, 10(7), e0130324 10.1371/journal.pone.0130324 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erwin, T. L. (1982). Tropical forests: Their richness in Coleoptera and other arthropod species. The Coleopterists Bulletin, 36(1), 74–75. [Google Scholar]
- Fontaine, B. , van Achterberg, K. , Alonso‐Zarazaga, M. A. , Araujo, R. , Asche, M. , Aspöck, H. , … Bouchet, P. (2012). New species in the Old World: Europe as a frontier in biodiversity exploration, a test bed for 21st Century taxonomy. PLoS ONE, 7(5), e36881 10.1371/journal.pone.0036881 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujita, M. K. , Leache, A. D. , Burbrink, F. T. , McGuire, J. A. , & Moritz, C. (2012). Coalescent‐based species delimitation in an integrative taxonomy. Trends in Ecology & Evolution, 27(9), 480–488. 10.1016/j.tree.2012.04.012 [DOI] [PubMed] [Google Scholar]
- Geiger, M. F. , Astrin, J. J. , Borsch, T. , Burkhardt, U. , Grobe, P. , Hand, R. , … Monje, C. (2016). How to tackle the molecular species inventory for an industrialized nation—lessons from the first phase of the German Barcode of Life initiative GBOL (2012–2015). Genome, 59(9), 661–670. [DOI] [PubMed] [Google Scholar]
- Geiger, M. F. , Moriniere, J. , Hausmann, A. , Haszprunar, G. , Wägele, W. , Hebert, P. D. N. , & Rulik, B. (2016). Testing the Global Malaise Trap Program – How well does the current barcode reference library identify flying insects in Germany? Biodiversity Data Journal, 4, e10671 10.3897/BDJ.4.e10671 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibson, J. , Shokralla, S. , Porter, T. M. , King, I. , van Konynenburg, S. , Janzen, D. H. , … Hajibabaei, M. (2014). Simultaneous assesssment of the macrobiome and microbiome in a bulk sample of tropical arthropods through DNA metasystematics. Proceedings of the National Academy of Sciences of the United States of America, 111, 8007–8012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gruttke, H. , Binot‐Hafke, M. , Balzer, S. , Haupt, H. , Hofbauer, N. , Ludwig, G. , & Ries, M. (2016). Rote Liste gefährdeter Tiere, Pflanzen und Pilze Deutschlands. Band 4: Wirbellose Tiere (Teil 2). Naturschutz Und Biologische Vielfalt, 70(4), 598. [Google Scholar]
- Gutiérrez, M. A. C. , Vivero, R. J. , Vélez, I. D. , Porter, C. H. , & Uribe, S. (2014). DNA barcoding for the identification of sand fly species (Diptera, Psychodidae, Phlebotominae) in Colombia. PLoS ONE, 9(1), e85496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gwiazdowski, R. A. , Foottit, R. G. , Maw, H. E. L. , & Hebert, P. D. N. (2015). The Hemiptera (Insecta) of Canada: Constructing a reference library of DNA barcodes. PLoS ONE, 10(4), e0125635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haarto, A. , & Ståhls, G. (2014). When mtDNA COI is misleading: Congruent signal of ITS2 molecular marker and morphology for North European Melanostoma Schiener, 1860 (Diptera, Syrphidae). ZooKeys, 431, 93–134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hajibabaei, M. , Shokralla, S. , Zhou, X. , Singer, G. A. , & Baird, D. J. (2011). Environmental barcoding: a next‐generation sequencing approach for biomonitoring applications using river benthos. PLoS ONE, 6(4), e17497 (7 pp). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hajibabaei, M. , Spall, J. L. , Shokralla, S. , & van Konynenburg, S. (2012). Assessing biodiversity of a freshwater benthic macroinvertebrate community through non‐destructive environmental barcoding of DNA from preservative ethanol. BMC Ecology, 12(1), 28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hallmann, C. A. , Sorg, M. , Jongejans, E. , Siepel, H. , Hofland, N. , Schwan, H. , … Goulson, D. (2017). More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE, 12(10), e0185809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hansson, C. , & Schmidt, S. (2018). Revision of the European species of Euplectrus Westwood (Hymenoptera, Eulophidae), with a key to European species of Euplectrini. Journal of Hymenoptera Research, 67, 900. [Google Scholar]
- Hardaluk, L. (in prep.). Metabarcoding in the Nationalpark Bayerischer Wald ‐ screening for invasive and pest invertebrates in bulk samples.
- Haszprunar, G. (2009). Barcoding Fauna Bavarica–eine Chance für die Entomologie. Nachrichtenblatt Der Bayerischen Entomologen Bayer Ent, 58(1/2), 45. [Google Scholar]
- Hausmann, A. , Godfray, H. C. J. , Huemer, P. , Mutanen, M. , Rougerie, R. , van Nieukerken, E. J. , … Hebert, P. D. N. (2013). Genetic patterns in European geometrid moths revealed by the Barcode Index Number (BIN) system. PLoS ONE, 8(12), e84518 10.1371/journal.pone.0084518 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hausmann, A. , Haszprunar, G. , & Hebert, P. D. N. (2011). DNA barcoding the geometrid fauna of Bavaria (Lepidoptera): Successes, surprises, and questions. PLoS ONE, 6(2), e17134 10.1371/journal.pone.0017134 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hausmann, A. , Haszprunar, G. , Segerer, A. H. , Speidel, W. , Behounek, G. , & Hebert, P. D. N. (2011). Now DNA‐barcoded: The butterflies and larger moths of Germany. Spixiana, 34(1), 47–58. [Google Scholar]
- Havemann, N. , Gossner, M. M. , Hendrich, L. , Morinière, J. , Niedringhaus, R. , Schäfer, P. , & Raupach, M. J. (2018). From water striders to water bugs: The molecular diversity of aquatic Heteroptera (Gerromorpha, Nepomorpha) of Germany based on DNA barcodes. PeerJ, 6, e4577 10.7717/peerj.4577 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawlitschek, O. , Fernández‐González, A. , Balmori‐de la Puente, A. , & Castresana, J. (2018). A pipeline for metabarcoding and diet analysis from fecal samples developed for a small semi‐aquatic mammal. PLoS ONE, 13(8), e0201763 10.1371/journal.pone.0201763 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawlitschek, O. , Morinière, J. , Lehmann, G. U. C. , Lehmann, A. W. , Kropf, M. , Dunz, A. , … Haszprunar, G. (2017). DNA barcoding of crickets, katydids and grasshoppers (Orthoptera) from Central Europe with focus on Austria. Germany and Switzerland. Molecular Ecology Resources, 17(5), 1037–1053. 10.1111/1755-0998.12638 [DOI] [PubMed] [Google Scholar]
- Hebert, P. D. N. , Cywinska, A. , Ball, S. L. , & Dewaard, J. R. (2003). Biological identifications through DNA barcodes. Proceedings of the Royal Society of London. Series B: Biological Sciences, 270(1512), 313–321. 10.1098/rspb.2002.2218 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hebert, P. D. N. , Ratnasingham, S. , Zakharov, E. V. , Telfer, A. C. , Levesque‐Beaudin, V. , Milton, M. A. , … Jannetta, P. (2016). Counting animal species with DNA barcodes: Canadian insects. Philosophical Transactions of the Royal Society B: Biological Sciences, 371(1702), 20150333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heller, K. , Köhler, A. , Menzel, F. , Olsen, K. M. , & Gammelo, Ø. (2016). Two formerly unrecognized species of Sciaridae (Diptera) revealed by DNA barcoding. Norwegian Journal of Entomology, 63(1), 96–115. [Google Scholar]
- Heller, K. , & Rulik, B. (2016). Ctenosciara alexanderkoenigi sp. n. (Diptera: Sciaridae), an exotic invader in Germany? Biodiversity Data Journal, 4, e6460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hendrich, L. , Morinière, J. , Haszprunar, G. , Hebert, P. D. N. , Hausmann, A. , Köhler, F. , & Balke, M. (2015). A comprehensive DNA barcode database for Central European beetles with a focus on Germany: Adding more than 3500 identified species to BOLD. Molecular Ecology Resources, 15(4), 795–818. 10.1111/1755-0998.12354 [DOI] [PubMed] [Google Scholar]
- Hernández‐Triana, L. M. , Prosser, S. W. , Rodríguez‐Perez, M. A. , Chaverri, L. G. , Hebert, P. D. N. , & Ryan Gregory, T . (2014). Recovery of DNA barcodes from blackfly museum specimens (Diptera: Simuliidae) using primer sets that target a variety of sequence lengths. Molecular Ecology Resources, 14(3), 508–518. [DOI] [PubMed] [Google Scholar]
- Hubert, N. , & Hanner, R. (2015). DNA barcoding, species delineation and taxonomy: A historical perspective. DNA Barcodes, 3(1), 44–58. 10.1515/dna-2015-0006 [DOI] [Google Scholar]
- Ivanova, N. V. , Dewaard, J. R. , & Hebert, P. D. N. (2006). An inexpensive, automation‐friendly protocol for recovering high‐quality DNA. Molecular Ecology Notes, 6(4), 998–1002. 10.1111/j.1471-8286.2006.01428.x [DOI] [Google Scholar]
- Jaschhof, M. (2009). Eine aktualisierte Artenliste der Holzmücken Deutschlands, unter besonderer Berücksichtigung der Fauna Bayerns (Diptera, Cecidomyiidae, Lestremiinae). Spixiana, 32(1), 139–151. [Google Scholar]
- Ji, Y. , Ashton, L. , Pedley, S. M. , Edwards, D. P. , Tang, Y. , Nakamura, A. , … Yu, D. W. (2013). Reliable, verifiable and efficient monitoring of biodiversity via metabarcoding. Ecology Letters, 16(10), 1245–1257. 10.1111/ele.12162 [DOI] [PubMed] [Google Scholar]
- Jordaens, K. , Goergen, G. , Virgilio, M. , Backeljau, T. , Vokaer, A. , & De Meyer, M. (2015). DNA barcoding to improve the taxonomy of the Afrotropical hoverflies (Insecta: Diptera: Syrphidae). PLoS ONE, 10(10), e0140264 10.1371/journal.pone.0140264 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jordaens, K. , Sonet, G. , Braet, Y. , De Meyer, M. , Backeljau, T. , Goovaerts, F. , … Desmyter, S. (2013). DNA barcoding and the differentiation between North American and West European Phormia regina (Diptera, Calliphoridae, Chrysomyinae). ZooKeys, 365, 149–174. 10.3897/zookeys.365.6202 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karlsson, D. , Pape, T. , Johansson, K. A. , Liljeblad, J. , & Ronquist, F. (2005). The Swedish Malaise Trap Project, or how many species of Hymenoptera and Diptera are there in Sweden? Entomologisk Tidsskrift, 126, 43–53. [Google Scholar]
- Klausnitzer, B. (2006). Stiefkinder der Entomologie in Mitteleuropa. Beiträge Zur Entomologie, 56, 360–368. [Google Scholar]
- Krüger, A. , Strüven, L. , Post, R. J. , & Faulde, M. (2011). The sandflies (Diptera: Psychodidae, Phlebotominae) in military camps in northern Afghanistan (2007–2009), as identified by morphology and DNA ‘barcoding’. Annals of Tropical Medicine & Parasitology, 105(2), 163–176. 10.1179/136485911X12899838683241 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar, N. P. , Rajavel, A. R. , Natarajan, R. , & Jambulingam, P. (2007). DNA barcodes can distinguish species of Indian mosquitoes (Diptera: Culicidae). Journal of Medical Entomology, 44(1), 01–07. 10.1093/jmedent/41.5.01 [DOI] [PubMed] [Google Scholar]
- Kumar, N. P. , Srinivasan, R. , & Jambulingam, P. (2012). DNA barcoding for identification of sand flies (Diptera: Psychodidae) in India. Molecular Ecology Resources, 12(3), 414–420. 10.1111/j.1755-0998.2012.03117.x [DOI] [PubMed] [Google Scholar]
- Latibari, M. H. , Moravvej, G. , Heller, K. , Rulik, B. , & Namaghi, H. S. (2015). New records of Black Fungus Gnats (Diptera: Sciaridae) from Iran, including the reinstatement of Bradysia cellarum Frey. Studia Dipterologica, 22(1), 39–44. [Google Scholar]
- Leray, M. , & Knowlton, N. (2015). DNA barcoding and metabarcoding of standardized samples reveal patterns of marine benthic diversity. Proceedings of the National Academy of Sciences USA, 112(7), 2076–2081. 10.1073/pnas.1424997112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leray, M. , Yang, Y. J. , Meyer, C. P. , Mills, S. C. , Agudelo, N. , Ranwez, V. , … Machida, R. J. (2013). A new versatile primer set targeting a short fragment of the mitochondrial COI region for metabarcoding metazoan diversity: Application for characterizing coral reef fish gut contents. Frontiers in Zoology, 10(1), 34 10.1186/1742-9994-10-34 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lister, B. C. , & Garcia, A. (2018). Climate‐driven declines in arthropod abundance restructure a rainforest food web. Proceedings of the National Academy of Sciences, 115(44), E10397–E10406. 10.1073/pnas.1722477115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin, M. (2011). Cutadapt removes adapter sequences from high‐throughput sequencing reads. EMBnet.journal, 17(1), 10 10.14806/ej.17.1.200 [DOI] [Google Scholar]
- Matthews, R. W. , & Matthews, J. R. (1971). The Malaise trap: Its utility and potential for sampling insect populations. The Great Lakes Entomologist, 4(4), 4. [Google Scholar]
- May, R. M. (1988). How many species are there on earth? Science, 241(4872), 1441–1449. [DOI] [PubMed] [Google Scholar]
- Meier, R. , Shiyang, K. , Vaidya, G. , & Ng, P. K. (2006). DNA barcoding and taxonomy in Diptera: A tale of high intraspecific variability and low identification success. Systematic Biology, 55(5), 715–728. 10.1080/10635150600969864 [DOI] [PubMed] [Google Scholar]
- Mengual, X. , Ståhls, G. , Vujić, A. , & Marcos‐Garcia, M. A. (2006). Integrative taxonomy of Iberian Merodon species (Diptera, Syrphidae). Zootaxa, 1377, 900–26. [Google Scholar]
- Meyer, H. , & Stark, A. (2015). Verzeichnis und Bibliografie der Tanzfliegenverwandten Deutschlands (Diptera: Empidoidea: Atelestidae, Brachystomatidae, Dolichopodidae s. l., Empididae, Hybotidae, “Iteaphila‐Gruppe”, Oreogetonidae). Studia Dipterologica Supplement 19.
- Montagna, M. , Mereghetti, V. , Lencioni, V. , & Rossaro, B. (2016). Integrated taxonomy and DNA barcoding of alpine midges (Diptera: Chironomidae). PLoS ONE, 11(3), e0149673 10.1371/journal.pone.0149673 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morinière, J. , Cancian de Araujo, B. , Lam, A. W. , Hausmann, A. , Balke, M. , Schmidt, S. , … Haszprunar, G. (2016). Species identification in Malaise trap samples by DNA barcoding based on NGS technologies and a scoring matrix. PLoS ONE, 11(5), e0155497 10.1371/journal.pone.0155497 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morinière, J. , Hendrich, L. , Balke, M. , Beermann, A. J. , König, T. , Hess, M. , … Haszprunar, G. (2017). A DNA barcode library for Germany′s mayflies, stoneflies and caddisflies (Ephemeroptera, Plecoptera and Trichoptera). Molecular Ecology Resources, 17(6), 1293–1307. 10.1111/1755-0998.12683 [DOI] [PubMed] [Google Scholar]
- Morinière, J. , Hendrich, L. , Hausmann, A. , Hebert, P. , Haszprunar, G. , & Gruppe, A. (2014). Barcoding Fauna Bavarica: 78% of the Neuropterida fauna barcoded!. PLoS ONE, 9(10), e109719 10.1371/journal.pone.0109719 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mutanen, M. , Kivelä, S. M. , Vos, R. A. , Doorenweerd, C. , Ratnasingham, S. , Hausmann, A. , … Godfray, H. C. J. (2016). Species‐level para‐and polyphyly in DNA barcode gene trees: Strong operational bias in European Lepidoptera. Systematic Biology, 65(6), 1024–1040. 10.1093/sysbio/syw044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagy, Z. T. , Sonet, G. , Mortelmans, J. , Vandewynkel, C. , & Grootaert, P. (2013). Using DNA barcodes for assessing diversity in the family Hybotidae (Diptera, Empidoidea). ZooKeys, 365, 263–278. 10.3897/zookeys.365.6070 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson, L. A. , Lambkin, C. L. , Batterham, P. , Wallman, J. F. , Dowton, M. , Whiting, M. F. , … Cameron, S. L. (2012). Beyond barcoding: A mitochondrial genomics approach to molecular phylogenetics and diagnostics of blowflies (Diptera: Calliphoridae). Gene, 511(2), 131–142. 10.1016/j.gene.2012.09.103 [DOI] [PubMed] [Google Scholar]
- Nelson, L. A. , Wallman, J. F. , & Dowton, M. (2007). Using COI barcodes to identify forensically and medically important blowflies. Medical and Veterinary Entomology, 21(1), 44–52. 10.1111/j.1365-2915.2007.00664.x [DOI] [PubMed] [Google Scholar]
- Normark, B. B. (2003). The evolution of alternative genetic systems in insects. Annual Review of Entomology, 48(1), 397–423. [DOI] [PubMed] [Google Scholar]
- Nzelu, C. O. , Cáceres, A. G. , Arrunátegui‐Jiménez, M. J. , Lañas‐Rosas, M. F. , Yañez‐Trujillano, H. H. , Luna‐Caipo, D. V. , … Kato, H. (2015). DNA barcoding for identification of sand fly species (Diptera: Psychodidae) from leishmaniasis‐endemic areas of Peru. Acta Tropica, 145, 45–51. 10.1016/j.actatropica.2015.02.003 [DOI] [PubMed] [Google Scholar]
- Ødegaard, F. (2000). How many species of arthropods? Erwin's estimate revised. Biological Journal of the Linnean Society, 71(4), 583–597. 10.1111/j.1095-8312.2000.tb01279.x [DOI] [Google Scholar]
- Oliverio, A. M. , Gan, H. , Wickings, K. , & Fierer, N. (2018). A DNA metabarcoding approach to characterize soil arthropod communities. Soil Biology and Biochemistry, 125, 37–43. 10.1016/j.soilbio.2018.06.026 [DOI] [Google Scholar]
- Oosterbroek, P. (2006). The European Families of the Diptera. Uitgeverij: KNNV‐Vereniging voor Veldbiologie. [Google Scholar]
- Packer, L. , Gibbs, J. , Sheffield, C. , & Hanner, R. (2009). DNA barcoding and the mediocrity of morphology. Molecular Ecology Resources, 9(Supplement 1), 42–50. 10.1111/j.1755-0998.2009.02631.x [DOI] [PubMed] [Google Scholar]
- Padial, J. M. , Miralles, A. , De la Riva, I. , & Vences, M. (2010). The integrative future of taxonomy. Frontiers in Zoology, 7(1), 16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Page, R. D. M. (2016). DNA barcoding and taxonomy: Dark taxa and dark texts. Philosophical Transactions of the Royal Society B: Biological Sciences, 371(1702), 20150334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pante, E. , Schoelinck, C. , & Puillandre, N. (2014). From integrative taxonomy to species description: One step beyond. Systematic Biology, 64(1), 152–160. [DOI] [PubMed] [Google Scholar]
- Pape, T. (2009). Palaearctic Diptera ‐ from tundra to desert In Pape T., Bickel D., & Meier R. (Eds.), Diptera diversity: Status, challenges and tools (pp. 121–154). Leiden, The Netherlands: Brill. [Google Scholar]
- Pape, T. , Blagoderov, V. , & Mostovski, M. B. (2011). Order Diptera Linnaeus, 1758 In Zhang Z.‐Q. (Ed.), Animal biodiversity: An outline of higher‐level classification and survey of taxonomic richness (pp. 222–229). Woodcroft, South Australia: Magnolia Press. [Google Scholar]
- Papp L., & Darvas B. (Eds.) (1997). Contribution to a Manual of Palaearctic Diptera. Vol. 2, Nematocera and Lower Brachycera. Budapest, Hungary: Science Herald. [Google Scholar]
- Papp L., & Darvas B. (Eds.) (1998). Contribution to a Manual of Palaearctic Diptera. Vol. 3, Higher Brachycera. Budapest, Hungary: Science Herald. [Google Scholar]
- Papp L., & Darvas B. (Eds.) (2000a). Contribution to a Manual of Palaearctic Diptera. Vol. 1, General and Applied Dipterology. Budapest, Hungary: Science Herald. [Google Scholar]
- Papp L., & Darvas B. (Eds.) (2000b). Contribution to a Manual of Palaearctic Diptera. Appendix. Budapest, Hungary: Science Herald. [Google Scholar]
- Pfenninger, M. , Nowak, C. , Kley, C. , Steinke, D. , & Streit, B. (2007). Utility of DNA taxonomy and barcoding for the inference of larval community structure in morphologically cryptic Chironomus (Diptera) species. Molecular Ecology, 16(9), 1957–1968. [DOI] [PubMed] [Google Scholar]
- Pohjoismäki, J. L. , Kahanpää, J. , & Mutanen, M. (2016). DNA barcodes for the northern European tachinid flies (Diptera: Tachinidae). PLoS ONE, 11(11), e0164933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Potts, S. G. , Biesmeijer, J. C. , Kremen, C. , Neumann, P. , Schweiger, O. , & Kunin, W. E. (2010). Global pollinator declines: Trends, impacts and drivers. Trends in Ecology & Evolution, 25(6), 345–353. [DOI] [PubMed] [Google Scholar]
- Puillandre, N. , Lambert, A. , Brouillet, S. , & Achaz, G. (2012). ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Molecular Ecology, 21(8), 1864–1877. 10.1111/j.1365-294X.2011.05239.x [DOI] [PubMed] [Google Scholar]
- R Core Team . (2018). R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; http://www.R-project.org/ [Google Scholar]
- Ratnasingham, S. , & Hebert, P. D. N. (2007). BOLD: The Barcode of Life Data System (http://www.barcodinglife.org). Molecular Ecology Notes, 7(3), 355–364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ratnasingham, S. , & Hebert, P. D. N. (2013). A DNA‐based registry for all animal species: The Barcode Index Number (BIN) system. PLoS ONE, 8(7), e66213 10.1371/journal.pone.0066213 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raupach, M. J. , Hannig, K. , Moriniere, J. , & Hendrich, L. (2016). A DNA barcode library for ground beetles (Insecta, Coleoptera, Carabidae) of Germany: The genus Bembidion Latreille, 1802 and allied taxa. ZooKeys, 592, 121–141. 10.3897/zookeys.592.8316 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raupach, M. J. , Hannig, K. , Morinière, J. , & Hendrich, L. (2018). A DNA barcode library for ground beetles of Germany: The genus Amara Bonelli, 1810 (Insecta, Coleoptera, Carabidae). ZooKeys, 759, 57–80. 10.3897/zookeys.759.24129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raupach, M. J. , Hendrich, L. , Küchler, S. M. , Deister, F. , Morinière, J. , & Gossner, M. M. (2014). Building‐up of a DNA barcode library for true bugs (Insecta: Hemiptera: Heteroptera) of Germany reveals taxonomic uncertainties and surprises. PLoS ONE, 9(9), e106940 10.1371/journal.pone.0106940 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reibe, S. , Schmitz, J. , & Madea, B. (2009). Molecular identification of forensically important blowfly species (Diptera: Calliphoridae) from Germany. Parasitology Research, 106(1), 257–261. 10.1007/s00436-009-1657-9 [DOI] [PubMed] [Google Scholar]
- Reimann, B. , & Rulik, B. (2015). Dasiops calvus Morge (Diptera: Lonchaeidae), a lance fly new to the German fauna, revealed by the GBOL‐project. Studia Dipterologica, 21(2), 283–287. [Google Scholar]
- Renaud, A. K. , Savage, J. , & Adamowicz, S. J. (2012). DNA barcoding of Northern Nearctic Muscidae (Diptera) reveals high correspondence between morphological and molecular species limits. BMC Ecology, 12(1), 24 10.1186/1472-6785-12-24 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riedel, A. , Sagata, K. , Suhardjono, Y. R. , Tänzler, R. , & Balke, M. (2013). Integrative taxonomy on the fast track‐towards more sustainability in biodiversity research. Frontiers in Zoology, 10(1), 15 10.1186/1742-9994-10-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rivera, J. , & Currie, D. C. (2009). Identification of Nearctic black flies using DNA barcodes (Diptera: Simuliidae). Molecular Ecology Resources, 9, 224–236. 10.1111/j.1755-0998.2009.02648.x [DOI] [PubMed] [Google Scholar]
- Rognes, T. , Flouri, T. , Nichols, B. , Quince, C. , & Mahé, F. (2016). VSEARCH: A versatile open source tool for metagenomics. PeerJ, 4, e2584 10.7717/peerj.2584 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rojo, S. , Ståhls, G. , Pérez‐Bañón, C. , & Marcos‐García, M. Á. (2006). Testing molecular barcodes: Invariant mitochondrial DNA sequences vs the larval and adult morphology of West Palaearctic Pandasyopthalmus species (Diptera: Syrphidae: Paragini). European Journal of Entomology, 103(2), 443 10.14411/eje.2006.058 [DOI] [Google Scholar]
- Rulik, B. , Eberle, J. , von der Mark, L. , Thormann, J. , Jung, M. , Köhler, F. , … Ahrens, D. (2017). Using taxonomic consistency with semiautomated data preprocessing for high quality DNA barcodes. Methods in Ecology and Evolution, 8(12), 1878–1887. 10.1111/2041-210X.12824 [DOI] [Google Scholar]
- Santos, D. , Sampronha, S. , & Santos, C. M. D. (2017). Advances on dipterology in the 21st century and extinction rates. Papéis Avulsos De Zoologia, 57(33), 433–444. 10.11606/0031-1049.2017.57.33 [DOI] [Google Scholar]
- Schlick‐Steiner, B. C. , Arthofer, W. , & Steiner, F. M. (2014). Take up the challenge! Opportunities for evolution research from resolving conflict in integrative taxonomy. Molecular Ecology, 23(17), 4192–4194. 10.1111/mec.12868 [DOI] [PubMed] [Google Scholar]
- Schlick‐Steiner, B. C. , Steiner, F. M. , Seifert, B. , Stauffer, C. , Christian, E. , & Crozier, R. H. (2010). Integrative taxonomy: A multisource approach to exploring biodiversity. Annual Review of Entomology, 55, 421–438. 10.1146/annurev-ento-112408-085432 [DOI] [PubMed] [Google Scholar]
- Schmid‐Egger, C. , Straka, J. , Ljubomirov, T. , Blagoev, G. A. , Morinière, J. , & Schmidt, S. (2019). DNA barcodes identify 99 per cent of apoid wasp species (Hymenoptera: Ampulicidae, Crabronidae, Sphecidae) from the Western Palearctic. Molecular Ecology Resources, 19(2), 476–484. 10.1111/1755-0998.12963 [DOI] [PubMed] [Google Scholar]
- Schmidt, S. , Schmid‐Egger, C. , Morinière, J. , Haszprunar, G. , & Hebert, P. D. N. (2015). DNA barcoding largely supports 250 years of classical taxonomy: Identifications for Central European bees (Hymenoptera, Apoidea partim). Molecular Ecology Resources, 15(4), 985–1000. [DOI] [PubMed] [Google Scholar]
- Schmidt, S. , Taeger, A. , Morinière, J. , Liston, A. , Blank, S. M. , Kramp, K. , … Stahlhut, J. (2017). Identification of sawflies and horntails (Hymenoptera, ‘Symphyta’) through DNA barcodes: Successes and caveats. Molecular Ecology Resources, 17(4), 670–685. 10.1111/1755-0998.12614 [DOI] [PubMed] [Google Scholar]
- Schumann, H. (2002). Erster Nachtrag zur „Checkliste der Dipteren Deutschlands “. Studia Dipterologica, 9(2), 437–445. [Google Scholar]
- Schumann, H. (2004). Zweiter Nachtrag zur „Checkliste der Dipteren Deutschlands “. Studia Dipterologica, 11(2), 619–630. [Google Scholar]
- Schumann, H. (2010). Dritter Nachtrag zur „Checkliste der Dipteren Deutschlands“. Studia Dipterologica, 16(1/2), 17–27. [Google Scholar]
- Schumann, H. , Bährmann, R. , & Stark, A. (1999). Checkliste der Dipteren Deutschlands. Entomofauna Germanica 2 . Studia Dipterologica Supplement, 2, 900–354. [Google Scholar]
- Schumann, H. , Doczkal, D. , & Ziegler, J. (2011). Diptera ‐ Zweiflügler In: Klausnitzer B. (Ed.), Stresemann, Exkursionsfauna von Deutschland. Vol. 2, Wirbellose: Insekten. 11 (pp. 832–932). Auflage: Spektrum Akademischer Verlag. [Google Scholar]
- Serrana, J. M. , Miyake, Y. , Gamboa, M. , & Watanabe, K. (2018). Comparison of DNA metabarcoding and morphological identification for stream macroinvertebrate biodiversity assessment and monitoring. bioRxiv, 436162 10.1101/436162 [DOI] [Google Scholar]
- Ševčík, J. , Kaspřák, D. , & Rulik, B. (2016). A new species of Docosia Winnertz from Central Europe, with DNA barcoding based on four gene markers (Diptera, Mycetophilidae). ZooKeys, 549, 127–143. 10.3897/zookeys.549.6925 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shokralla, S. , Spall, J. , Gibson, J. , & Hajibabaei, M. (2012). Next‐generation sequencing technologies for environmental DNA research. Molecular Ecology, 21(8), 1794–1805. 10.1111/j.1365-294X.2012.05538.x [DOI] [PubMed] [Google Scholar]
- Sinclair, C. S. , & Gresens, S. E. (2008). Discrimination of Cricotopus species (Diptera: Chironomidae) by DNA barcoding. Bulletin of Entomological Research, 98(6), 555–563. 10.1017/S0007485308005865 [DOI] [PubMed] [Google Scholar]
- Sorg, M. , Schwan, H. , Stenmans, W. , & Müller, A. (2013). Ermittlung der Biomassen flugaktiver Insekten im Naturschutzgebiet Orbroicher Bruch mit Malaise Fallen in den Jahren 1989 und 2013. Mitteilungen Entomologischer Verein Krefeld, 1, 900–5. [Google Scholar]
- Spelda, J. , Reip, H. S. , Oliveira Biener, U. , & Melzer, R. R. (2011). Barcoding Fauna Bavarica: Myriapoda – a contribution to DNA sequence‐based identifications of centipedes and millipedes (Chilopoda, Diplopoda). ZooKeys, 115, 123–139. 10.3897/zookeys.156.2176 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ssymank, A. , Doczkal, D. , Rennwald, K. , & Dziock, F. (2011). Rote Liste und Gesamtartenliste der Schwebfliegen (Diptera: Syrphidae) Deutschlands. Naturschutz Und Biologische Vielfalt, 70(3), 13–83. [Google Scholar]
- Ssymank, A. , Sorg, M. , Doczkal, D. , Rulik, B. , Merkel‐Wallner, G. , & Vischer‐Leopold, M. (2018). Praktische Hinweise und Empfehlungen zur Anwendung von Malaisefallen für Insekten in der Biodiversitätserfassung und im Monitoring. Series Naturalis, 1, 900–12. [Google Scholar]
- Stur, E. , & Borkent, A. (2014). When DNA barcoding and morphology mesh: Ceratopogonidae diversity in Finnmark, Norway. ZooKeys, 463, 95–131. 10.3897/zookeys.463.7964 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stur, E. , & Ekrem, T. (2011). Exploring unknown life stages of Arctic Tanytarsini (Diptera: Chironomidae) with DNA barcoding. Zootaxa, 2743(1), 27–39. 10.11646/zootaxa.2743.1.2 [DOI] [Google Scholar]
- Taberlet, P. , Coissac, E. , Pompanon, F. , Brochmann, C. , & Willerslev, E. (2012). Towards next‐generation biodiversity assessment using DNA metabarcoding. Molecular Ecology, 21(8), 2045–2050. 10.1111/j.1365-294X.2012.05470.x [DOI] [PubMed] [Google Scholar]
- Vanbergen, A. J. , & Insect Pollinators Initiative . (2013). Threats to an ecosystem service: Pressures on pollinators. Frontiers in Ecology and the Environment, 11(5), 251–259. 10.1890/120126 [DOI] [Google Scholar]
- Versteirt, V. , Nagy, Z. T. , Roelants, P. , Denis, L. , Breman, F. C. , Damiens, D. , … Van Bortel, W. (2015). Identification of Belgian mosquito species (Diptera: Culicidae) by DNA barcoding. Molecular Ecology Resources, 15(2), 449–457. 10.1111/1755-0998.12318 [DOI] [PubMed] [Google Scholar]
- Völkl, W. , Blick, T. , Kornacker, P. M. , & Martens, H. (2004). Quantitativer Überblick über die rezente Fauna von Deutschland. Natur Und Landschaft, 79(7), 293–295. [Google Scholar]
- Wang, G. , Li, C. , Guo, X. , Xing, D. , Dong, Y. , Wang, Z. , … Zhao, T. (2012). Identifying the main mosquito species in China based on DNA barcoding. PLoS ONE, 7(10), e47051 10.1371/journal.pone.0047051 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wesener, T. , Voigtländer, K. , Decker, P. , Oeyen, J. P. , Spelda, J. , & Lindner, N. (2015). First results of the German Barcode of Life (GBOL)– Myriapoda project: Cryptic lineages in German Stenotaenia linearis (Koch, 1835) (Chilopoda, Geophilomorpha). ZooKeys, 510, 15–928. 10.3897/zookeys.510.8852 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wheeler, Q. D. , Raven, P. H. , & Wilson, E. O. (2004). Taxonomy: Impediment or expedient? Science, 303, 285–285. 10.1126/science.303.5656.285 [DOI] [PubMed] [Google Scholar]
- Whitworth, T. L. , Dawson, R. D. , Magalon, H. , & Baudry, E. (2007). DNA barcoding cannot reliably identify species of the blowfly genus Protocalliphora (Diptera: Calliphoridae). Proceedings of the Royal Society B: Biological Sciences, 274(1619), 1731–1739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolff, D. , Gebel, M. , & Geller‐Grimm, F. (2018). Die Raubfliegen Deutschlands. Quelle & Meyer Bestimmungsbücher. [Google Scholar]
- Yu, D. W. , Ji, Y. , Emerson, B. C. , Wang, X. , Ye, C. , Yang, C. , & Ding, Z. (2012). Biodiversity soup: Metabarcoding of arthropods for rapid biodiversity assessment and biomonitoring. Methods in Ecology and Evolution, 3(4), 613–623. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All specimen data have been made publicly available within the BOLD workbench ‐ a DOI for the dataset has been added.