

Abstract
Evidence over the last couple decades has comprehensively established that short, highly conserved, non-coding RNA species called microRNA (miRNA) exhibit the ability to regulate expression and function of host genes at the messenger RNA (mRNA) level. MicroRNAs play key regulatory roles in immune cell development, differentiation, and protective function. Intrinsic host immune response to invading pathogens rely on intricate orchestrated events in the development of innate and adaptive arms of immunity.
We discuss the involvement of miRNAs in regulating these processes against gram negative pathogens in this review.
Keywords: bacteria, gram negative, microRNAs, host immunity
INTRODUCTION
MicroRNA
Historically, Lin-4 was the first miRNA to be discovered and in Caenorhabditis elegans attributed to control of larval development [1]. Interestingly, this small RNA had the ability to regulate another mRNA (lin-14) and reduce the amount of lin-14 protein [2]. Discovery of lin-4 demonstrated a unique paring with the 3’untranslated region (3’UTR) of the gene and repressing lin-14 translation during larval stage cell division [1, 2]. Following lin-4, multiple such mRNA regulators have been found conserved across multiple organisms [3]. These small RNA single stranded ~22 nucleotide species were named microRNA (miRNA) [4]. Additionally, post-transcriptional regulation by miRNA was soon discovered on 5’UTR and coding sequences of target genes [5–7]. MiRNA binds target transcripts, which lead to either degradation or enhancement of mRNA allowing for additional control to environmental conditions [8–10].
The biogenesis of miRNA is a highly regulated process, miRNA are transcribed by RNA polymerase II into a long primary RNA (pri-miRNA) (Figure 1). These pri-miRNA can be longer than 1kb and contain a stem-loop structure [11]. Nuclear cleavage follows transcription of pri-miR, cleaving the stem-loop structure one helical turn (approximately 11 bp away from the basal junction, and 22 bps from the apical junction) into the stem leaving approximately 60 nucleotides [12]. Cleavage of the pri-miR is achieved by RNase III endonucleases Drosha along with RNA binding protein DGCR8 collectively called the Microprocessor complex [13–15]. Microprocessor cleavage leaves a staggered end 5’ phosphate and a small 2 nucleotide 3’ overhang called pre-miRNA. The pre-miRNA is exported out of the nucleus by the protein Exportin5 (EXP5) binding RAN-GTP and the pre-miRNA trans-locates outside of the nucleus. Once GTP is hydrolyzed it releases the pre-miRNA into the cytoplasm [11, 16].
Figure 1. A generic model for post-transcriptional miRNA expression.
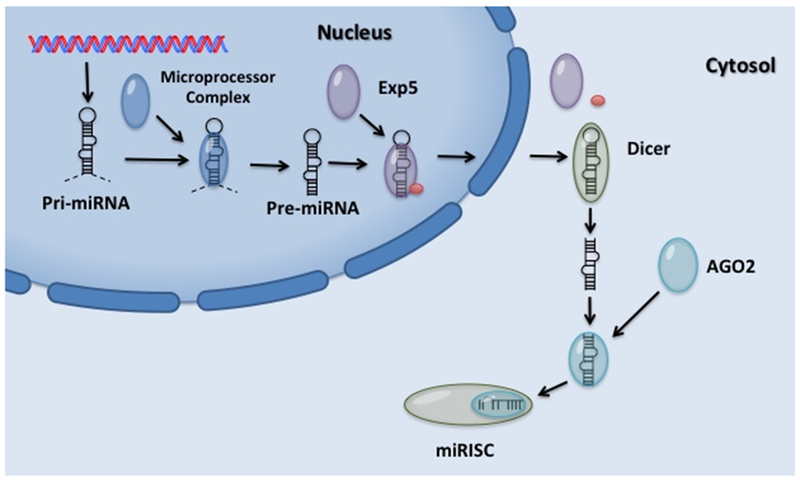
Full length pri-microRNA (pri-miRNA) containing a stem-loop structure initially transcribed by RNA polymerase II is cleaved by the microprocessor complex. Pre-miRNA transported out of the nucleus by Exportin 5 (Exp5) and identified by Dicer. Cleavage of pre-miRNA hairpin loop structure by Dicer leaves a mature double-stranded miRNA approximately 22 nucleotide in length. Argonaute (AGO2) preferentially loads one strand into microRNA-induced silencing complex (miRISC) which blocks translation or increases mRNA degradation.
In the cytoplasm, another RNase III type enzyme (Dicer) binds the 5’ end of the pre-miRNA and cleaves the double stranded RNA approximately two helical turns from the base of the hairpin [17]. After cleavage of the pre-miRNA by Dicer, the ~22 nucleotides double-stranded miRNA is identified by Argonaute (AGO2) protein and one strand is preferentially loaded into a microRNA-induced silencing complex (miRISC) [16, 18]. MiRISC targets conserved Watson-Crick pairing at the 5’ end of the miRNA centered on nucleotides 2-7 called the seed sequence and either blocks translation or increases mRNA degradation [4, 19, 20].
Disruption of Dicer1 in mouse embryonic stem (ES) cell lines display defects in differentiation showing the role of miRNA in development [21]. Complete loss of Dicer1 leads to early embryonic lethality which required researchers to develop conditional deletion of Dicer in order to further evaluate its role in immune signaling pathways [22]. Conditional deletion of Dicer in T cells impaired development and disrupts normal cytokine production [23]. These previous studies where key components of the miRNA biogenesis pathway and have implicated miRNA as an important regulator of immune response to gram-negative pathogens [23]. The ability of miRNA to coordinate precise protein expression in response to differentiation, apoptosis, inflammation and cell proliferation following stimulation has demonstrated their critical role in the immune system [10, 24, 25].
Innate Immunity
The immune system can be broadly characterized into two general components, the innate and adaptive immune systems. The innate immune system relies on dendritic cells, macrophages, monocytes, neutrophils, and NK cells. Both neutrophils and macrophages respond to microbial infection while other granulocytes such as basophils, mast cells, and eosinophils respond to parasites. More specifically, in response to the local environmental conditionals, macrophages are activated differently to respond to invading pathogens. Classically activated macrophages (M1) following microbial infection and alternatively activated macrophages (M2) to fungi and parasites further demonstrate the ability of the immune system to fine-tune a correct immune response to invading pathogens [26]. While this classification of macrophages does not exhibit the full spectrum of activated macrophages and can be thought of as oversimplification it shows how environmental conditions mediate pathogen resistance through differential activation. It has been shown that both monocytes and macrophages induce epigenetic changes, which leads to enhanced cytokine production and changes in cellular activation [27, 28]. Innate immunity is thought to be a nonspecific response to pathogens but has recently shown a great deal of specificity through epigenetic reprogramming [29]. These observations led to the hypothesis that certain infections or vaccines are able to induce reprogramming of the innate immune response, leading to nonspecific protection [30]. Different cell lineages with distinct chromatin signatures and transcriptional regulation help regulate innate cell function. Therefore, how cell lineage and the specific signaling pathways trigger activation in response to a particular pathogen needs to be considered [31].
Recognition of Lipopolysaccharide (LPS), an essential part of the outer membrane surface of most gram-negative bacterium, is a key detection target by host immune cells [32–34]. Modifications of its lipid-A portion of LPS allows gram-negative bacterium to evade host immune identification, enabling bacterium to survive in specific niches inside the host [35]. Host mucosal environment contains a high abundance of gram-negative commensal bacteria and it is essential that the host establish tolerance to these commensal bacterium [36]. Conserved targets, known as a pathogen-associated molecular pattern (PAMPs), are recognized by pattern-recognition receptors (PRR), which aid in further identification of gram-negative pathogens [37, 38]. Generalized recognition is the underlying premise for the innate immune system to be the first/ primary line of defense against invading organisms. Signaling through Toll-like receptors (TLRs), antigen presenting cells (APCs) elicit cytokine production among other early warning systems [39]. The TLR families are trans-membrane proteins that respond differently to surface markers on gram-negative bacteria [40]. Given that cytoplasmic domain of TLR proteins are homologous to IL-1R, these regions are referred to as Toll/IL-1R (TIR) domain. TLRs recruit adaptor molecule(s) containing TIR domain to its cytoplasmic TIR domain. One such adaptor, myeloid differentiation factor 88 (MyD88), associates with IL-1R-associated kinases (IRAKs) to mediate signaling to TNF-receptor-associated factor 6 (TRAF6), and stimulates IKKs and MAPKs. Ultimately, TLR signaling leads to activation of transcription factors such as NF-κB and AP-1, increasing clearance of a specific pathogen (Figure 2a) [37].
Figure 2. Role of microRNA (miR) in regulating Toll-like receptor (TLR) signaling.
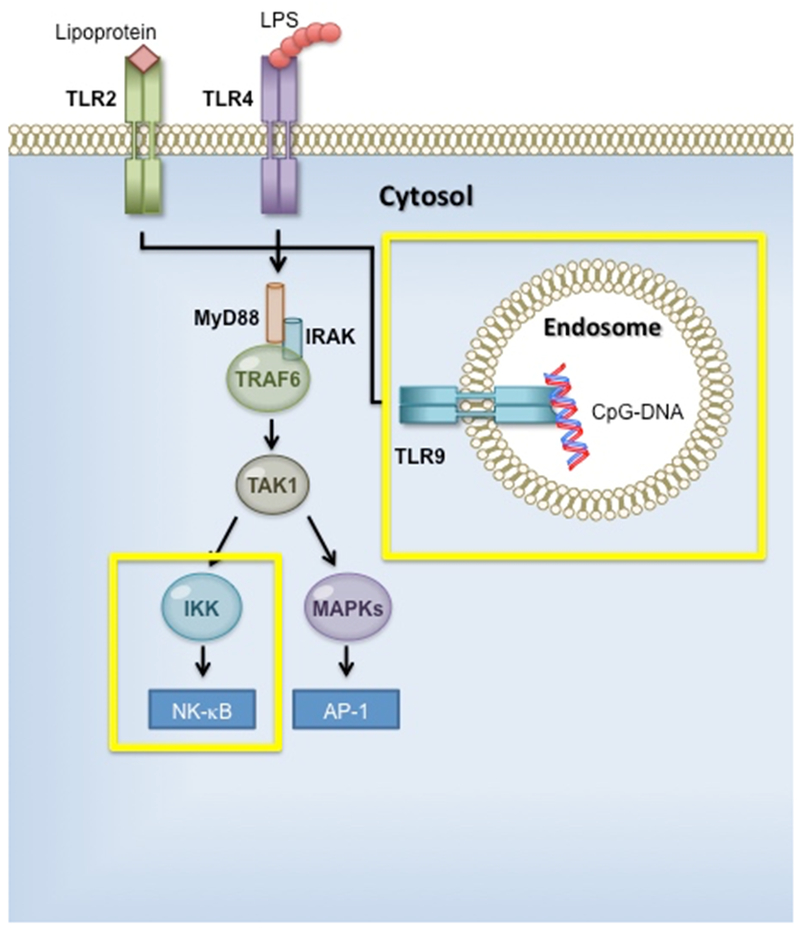
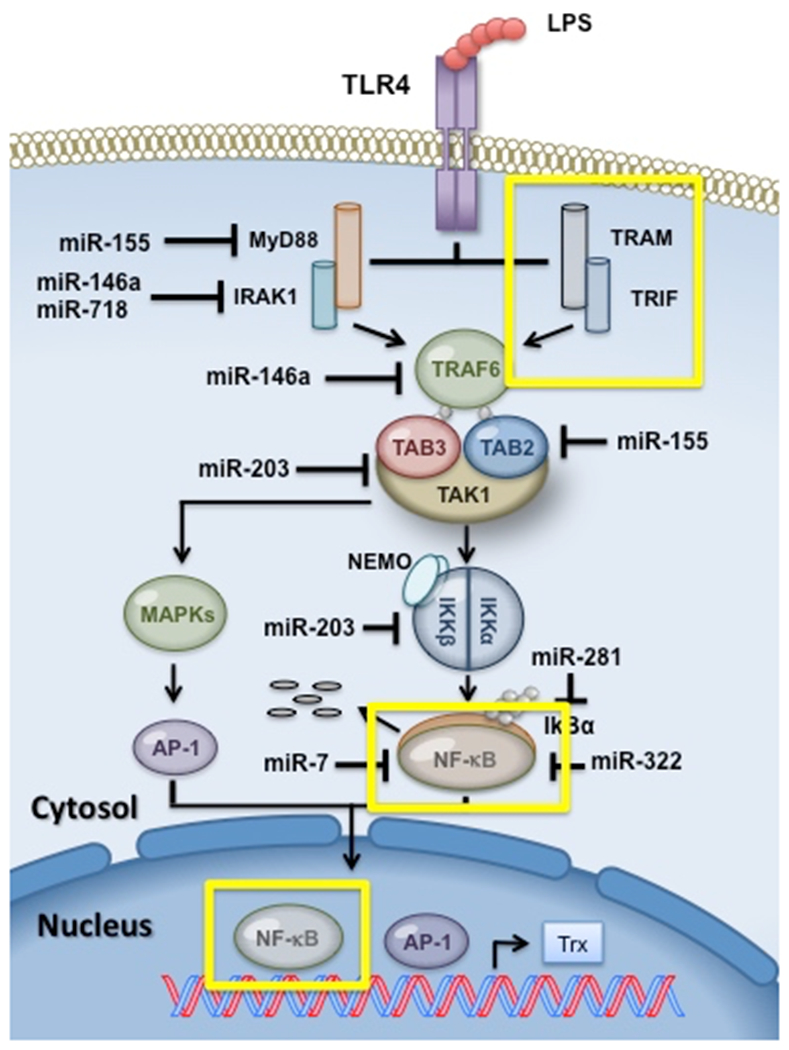
a. TLR signaling pathway. TLR families share similar signaling components, including MyD88, IRAK (IL-1R associated kinase),TRAF6 (TNF receptor-associated factor 6), and TAK1 (TGF-β-activated kinase). Activation leads to MAPK (mitogen-activated protein kinase) and nuclear factor-κB (NF-κB) translocation into the nucleus, targeting gene expression. b. Working model for miR mediated control of LPS-TLR4 signaling network.
TLR4 recognizes LPS and signals through MyD88 and TRIF pathway to downstream TRAF6 [38]. Multiple miRNA have been implicated in regulating TLR4 signaling following LPS stimulation (Figure 2b). Relative miRNA expression in chronic obstructive pulmonary disease (COPD) patients showed increased levels of miR-203 [41]. Bronchial epithelial cells from COPD patients stimulated with LPS have increased miR-203 and decreased Tak1 expression [41]. Immune suppression of Tak1 by miR-203 reduces NF-κB activity and may contribute to increased susceptibility to bacterial infection [41]. Treatment of A549 cells with LPS enhanced miR-381 in a dose dependent manner and identified IκBα as one of its targets [42]. Expression of miR-322 was suppressed in LPS treated macrophages, and regulated NF-κB p50s ability to fine-tune inflammatory responses following simulation [43]. LPS-induced acute lung injury in vivo and in LPS-treated mouse cells in vitro demonstrated regulation of VEGFA by miR-126-5p [44]. Overexpression of miR-126-5p attenuated LPS-induced regulation of alveolar fluid clearance [44]. LPS-induced acute lung inflammation apoptosis is mechanistically controlled by miR-181a regulating anti-apoptotic gene BCL-2 [45]. Induction of miR-375 and miR-106b following LPS simulation in Helicobacter pylori infection has been identified as putative regulators of JAK2/STAT3 pathway in gastric epithelial cells [46, 47].
TLR2 recognizes lipoproteins signaling by MyD88, similar to TLR4 MyD88 pathway signaling, and enables the translocation of NF-κB to the nucleus which activates target immune response genes [35]. THP-1 macrophage-derived foam cells infected with Chlamydia pneumoniae suppress ABCA1 and ABCA1-dependent cholesterol efflux, which is mediated by TLR2- NF-κB and miR-33 pathways [48]. MiR-146 expression following NF-κB translocation negatively regulates bacterial activated TLRs and cytokine receptor signaling [49]. TRAF6 and IRAK1 are direct targets for miR-146, which up-regulate following NF-κB activation [50]. Control of IRAK1 expression by miR-718 has also been identified [51]. The overlap of miR-718 with the 5’ UTR of IRAK1 suggests a role for miR-718 in controlling TLR signaling through a negative feedback loop [51]. MiR-155 is known to be a key role player in immunity [52, 53] and has also been identified as a regulator of LPS/TLR simulation [48, 54]. While miR-146a acts as a negative regulator, miR-155 establishes a pro-inflammatory condition through direct targeting TAB2, IKKε, and NIK [55]. Indirect regulation of RelA (p65) by miR-7 through targeting IKKε was observed in gastric cancer and shows feedback loops between miR-7 and NF-κB signaling [56]. Endosome located TLR9 identifies oligonucleotides containing unmethylated cytosine-phosphate-guanine (CPG) dinucleotides, and induces expression of IFN-α through transcription factor IRF7 [57]. NF-κB activation following TLR9 stimulation induces pro-inflammatory cytokines tumor necrosis factor alpha (TNF-α), IL-1β, and IL-6. TLR9 activation by CPG leads to up-regulation of miR-146 targeting Notch1 and repression of IL-12p70 within dendritic cells [58].
Aside from TLRs, other PRR such as the cytosolic NOD-like receptor (NLRs), the cell-surface C-type lectin receptors (CLRs) show the diversity of PRRs. Nucleotide-binding oligomerization domain-containing protein 1 (NOD1), a member of the NOD-like receptor protein family, regulates miR-155 in macrophages following recognition of H. pylori T4SS translocation [59]. In the absence of miR-155, bone marrow derived macrophages are less resistant to DNA damage induced apoptosis [60]. H. pylori regulation of apoptosis through miR-155 targets Trp53inp1, Bach1, Lpin1, Pmaip1, and Tspan14 allowing stabilization of macrophages in a hyper inflamed environment [59]. The variety of PRR ultimately delineate to a common intracellular signaling cascade involving NF-κB, mitogen-activated protein kinases (MAPKs), or interferon-regulatory factors (IRFs) [61]. A highly virulent type A strain of Francisella tularensis strain (Schu S4) has the ability to subvert host immune detection by regulating specific immune response pathways [62]. Upon engulfment, Schu S4 can escape from the phagosome [63] and when compared to less virulent stain F. tularensis novicida (F.n.) the regulation of miR-155 was substantial [64]. Increased NF-κB activity is seen in monocytes and macrophages following infection with the less virulent strain compared with Sshu S4[65]. SH-2 containing inositol 5’ polyphosphatase 1 (SHIP1), a negative regulator of PI3K/Akt pathway was down regulated following F.n. infection but did not change significantly following Sshu S4 [65]. MiR-155 is a direct target of SHIP1 and following miRNA regulation leads to increased activation of Akt [66]. Recruitment of neutrophils to lower genital tract following Chlamydia trachomatis genital infection in mice is mediated by intracellular adhesion molecule 1 (ICAM1) [67]. Recent evidence has shown that miR-214 mediates ICAM1 and up regulates miR-214, which in turn decreases neutrophil recruitment leading to greater pathology in mice [67, 68]. In the absence of IL-17A, miR-214 was increased and ICAM1 levels decreased with the subsequent abrogation of genital pathology in IL-17A−/− mice. IL-17A has been linked to neutrophil recruitment, hyper inflammation, and tissue damage eluding to a potential role for miR-214 mediated control of ICAM1 in neutrophils with the contribution of IL17A [67, 69, 70].
The innate immune system is very effective in recognizing PAMPs, but an uncontrolled immune response could lead to hyper inflammation or sepsis followed by multiple organ dysfunction and potentially death [57]. Klebsiella pneumoniae is one of the most prevalent gram-negative bacteria found in intensive care units [71]. A common cause of surgical sepsis, patients with peritonitis lack the ability to mount an effective immune response, which leads to a persistent infection, and an overactive immune response [72]. It is vital that the host counteracts hyper inflammation, and through the development of tolerance it’s able to prevent endotoxin shock. Mice can be desensitized with a non-lethal amount of LPS prior to a lethal does and leads to a reduction in LPS related immune response [73]. Following tolerance, monocytes increase their ability to phagocytose pathogens by up-regulation of CD64 while down regulating major histocompatibility complex class II (MHCII) gene CIITA [73]. Early expression of miR-132 with LPS-pretreatment down-regulated IL-1β [72]. A confirmed target of miR-21, PDCD4 shows lower mortality rates by LPS and lower IL-6 while increasing IL-10 production [72]. Selective and transient silencing of inflammatory genes at the level of chromatin plays a critical role in LPS tolerant macrophages [74]. During H. pylori infections of ASG cell lines, decreased miR-218 expression elevates ECOP inducing NF-κB activation [75]. TLR4 signaling can be reduced by direct targeting of IKKε or MyD88 by miR-155, and allows a decrease in rapid response to pathogens without inducing hyper inflammation. Salmonella sp. infection prevents septic shock and apoptosis through macrophage mediation of let-7 [76]. A common regulator in gram-negative immune response, let-7 decreases following TLR-4 signaling by LPS and correlates to an increase in IL-6, IL-8, and IL-10 [76, 77]. Multiple binding sites on IL-10 by let-7 suggest an important role for let-7 mediation of IL-10 [76].
Macrophages deficient in miR-146 show higher levels of phagocytic activity and reduced levels of bacteria burden [78]. IL-10 or TGF-β up-regulation suppress MHC Class II presentation albeit through different pathways and reduces pro-inflammatory immune responses. IL-10 down regulates miR-155 while up-regulating miR-146 further eluding to the fact that miR-146 is an anti-inflammatory gene while miR-155 favors a pro-inflammatory phenotype. Inhibition of miR-21 reduced severe steroid-insensitive allergic airway disease induced by gram-negative bacteria through regulation of PTEN, PI3K, and HDAC2 potentially identifying a novel treatment fonr asthma [79]. During wound healing (efferocytosis), engulfment of apoptotic cells by macrophages induces an anti-inflammatory condition by up-regulating both miRNA and interleukins. IL-10 up regulation and PTEN down-regulation is seen following efferocytosis and has been identified as targets for miR-21 [80]. PTEN repression and enhanced PI3K/Akt signaling by miR-718 in macrophages infection with LPS demonstrates multiple miRNA involvement in PTEN regulation [51]. Furthermore, Anti-miR-718 macrophages exhibited control over N. gonorrhoeae replication by reducing the number of bacteria recovered compared to miR-718 macrophages [51]. Silencing of key tight junction proteins Zona occludens-1,2 (ZO-1, ZO-2) by both miR-106b and miR-146a has been shown to lead to compromised wound closure following infiltration by Acinetobacter sp. [81].
Adaptive Immunity
The recognition of conserved regions by the innate immune system allows for a rapid response to pathogens and their antigens. The ensuing adaptive immune response provides a more robust and antigen specific immune defense against invading pathogens. Immune effector cells called antigen presenting cells (APC) such as dendritic cells or macrophages present antigens to the adaptive immune arm of lymphocytes via MHC on the APC along with other co-stimulating signals [82]. Antigen recognition by T cells though T Cell Receptors (TCR) or B cells through B Cell Receptors (BCR) allow antigens to be recognized by the adaptive immune system. Hematopoiesis in the bone marrow and later in respective organs; thymus for T cells and spleen by B cells on surface receptors are generated [83]. Following MHC Class I binding by TCRs a commitment to Cytotoxic CD8+ T cell (CD8+) lineage is established. Conversely, when TCR binds to MHC Class II it promotes T cell differentiation into T regulatory, Th1, Th2, Th9, or Th17 lineages [84–86]. The identification of self and non-self is an important step that leads to accurate pathogen identification while reducing the about of damage to the host [82, 83]. During development in the thymus, T cells that recognize self-proteins are destroyed and a similar mechanism is put into place in the periphery to eliminate mature T cells that develop a propensity toward self. Importantly, immune response to pathogens by miRNA ultimately leads to fate-determining steps within T cells (Figure 3). Following activation, T cells produce IL-2 through the binding of IL-2/IL-2R cascade. Conditional deletion of Dicer in CD4+ cells disrupts the ability of mTOR to discriminate between activation and anergy, allowing CD4+ cells to escape inactive states [87–89]. H. pylori regulation of SOX4 through decreased miR-204 expression aids in T cell differentiation [90]. Induced by TGF-β, SOX4 inhibits GATA-3 function leading to a decrease in Th2 differentiation [91–93]. MiR-155 deficient mice exhibit an inability to initiate an adaptive immune response to S. typhimurium or H. pylori through impaired Ag-specific Th1 or Th17 [94, 95]. T cell proliferation experiments with miR-155 showed little change in Th1 production of IFN-γ while elevated Th2 producing cytokines IL-4, IL-5, and IL-10 demonstrate an intrinsic bias toward Th2 differentiation [94]. A transactivator of IL-4 promoter, c-MAF was identified as a match for miR-155 further supporting increased Th2 cytokine production seen in knockdown mice [94]. Additionally, mice produced significantly reduced amounts of immunoglobulin M (IgM), Indicative of impaired B cell response [94]. B cells that lack miR-155 fail to produce high affinity IgG1 antibodies due to decreased regulation of miR-155 direct binding of PU.1 [96]. Th1 mediated protection via miRs- 182 and miR-183, expressed in the same operon [97], have been reported in genital tract infection with C. trachomatis intravaginally infected mice [98]. Importantly, IFN-γ secretion in infected mice was synergistically regulated by miR-155 and miR-182 [98]. Similar to this study, a report published simultaneously demonstrated a role for miR-182/ 183 cluster in regulation in Th17 lineage specific cells [99, 100]. Regulation of IFN-γ mRNA has also been shown to be controlled by miR-29, miR-29 downregulation in Th1, Th2, Th17, CD8+, along with NK cells facilitated IFN-γ production [101].
Figure 3. CD4+ T cell differentiation mediated by microRNA.
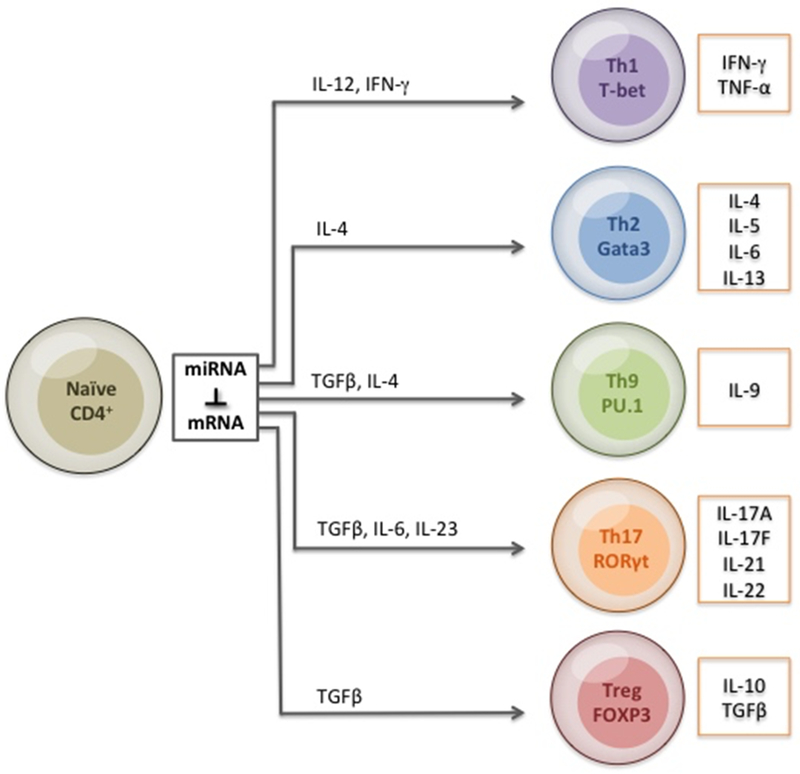
MiRNA regulates lineage determination and differentiation by fine-tuning signal strength, duration, and cytokine expression following gram-negative bacterial infection.
CONCLUSION
Overall, accumulating evidence has demonstrated that miRNA play important roles in several host processes [102–105] including the innate and adaptive immune signaling network following bacterial infections [106–108]. It may however be misleading to consider individual cell pathways or populations as having a predominant effect during a gram-negative infection. Immune cell activation may be individually customized by single or cohorts of miRNAs in response to specific gram-negative pathogens [109–111]. Compromised immune systems from severe steroid-insensitive allergic airway disease [112], septic shock induced tolerance [76], or wound healing [80] has the potential to controlled miRNA regulation and restore the balance between key regulators of the immune system. Harnessing the intrinsic functionality of miRNA has led to the appreciation of the previously untapped therapeutic potential [113, 114] and resulted in patents [115–118], and advancements in various areas including cervical cancers [119, 120], tissue transplantation [121–123], and infections such as influenza [118, 124, 125], HCV [126, 127], HBV [128], Salmonella [129], Mycobacterium sp. [106, 130] and Helicobacter sp. [131]. The role of miRNA in vaccination has also been reported [113, 132, 133]. For example, co-involvement of miR-462/731 in IFN-mediated protection was conferred by TLR-3 agonist (poly I:C; polyinosinic: polycytidylate) in vaccinated fish challenged with viral hemorrhagic septicemia [134]. The immune augmenting capability of miRNA was demonstrated in vivo by overexpression of miR-155 in hepatotropic adeno-associated virus (AAV)-8 vectors thereby enhancing the protective capacity of genetically attenuated malarial parasites in mice prime-booster vaccination regime with a prime only regime [135]. Similarly, AAV-based miR-26a replacement therapy was used for a liver cancer mouse model [136], in vivo targeted delivery of a let7g-aptamer led to reduction in lung adenocarcinomas [137]. MiR-22 impaired the anti-tumor ability of dendritic cells by targeting p38 in a B16 melanoma murine model [138], while miR-182 reduced in vivo glioblastoma multiforme xenografts growth [139]. The exosome-associated miR-150 has been found as a sensor for vaccination against 2009 pandemic flu (H1N1) in cohorts of adults and children [140]. Importantly, the success of MRX-34 [141, 142], a liposome formulation of miR-34 mimic for hepatic carcinomas and miravirsen[126, 143], a locked nucleic acid (LNA)-anti-miR-122 alone or in combinatorial regimes with telaprevir and ribavirin controls HCV [144, 145] and has led to phase I / II trials showing promise in miR mediated therapy [113, 146, 147]. Additionally, miRNA-based RNA interference [148] or combination miRNA-based strategies, have been reported to control influenza [149] and non-small cell lung cancers [150], respectively. To this end, specific miRNA that may affect critical host functions and directly or indirectly contribute to antigen specific immunity hold further promise (Table 1) [50, 151–182]. Hence as our current understanding of miRNA in gram-negative pathogens continues to grow, the possibly that miRNA could be used as new therapeutics [183–185] to regulate dysfunctional immune networks would allow medical professionals additional approaches for identification and treatment for gram-negative bacteria.
Table 1.
Regulation of MicroRNAs and Gram Negative Bacteria
Figure 4. Gram-negative host miRNA immune regulation working model.
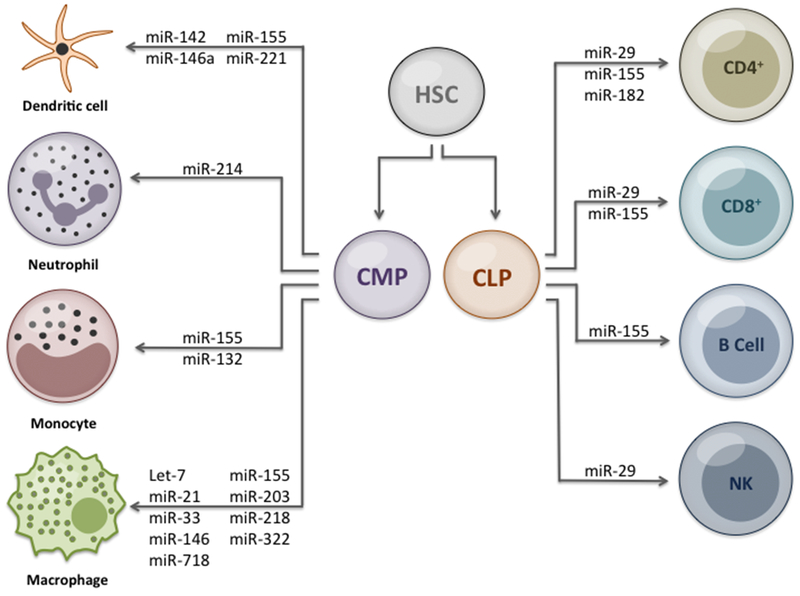
Representation of miRNA regulation following commitment of cell lineage in response to gram-negative bacteria infection. Hematopoetic stem cells (HSC); common myeloid progenitor (CMP); and common lymphoid progenitor (CLP).
ACKNOWLEDGEMENTS
This work was supported by National Institutes of Health Grant (1RO3AI11771401A1), Army Research Office, Department of Defense Contract No. W911NF-11-1-0136 to BPA and National Institute of Child Health and Human Development Grant (HD061580) to LKC.
Footnotes
DISCLOSURE
The authors declare that they have no competing interests.
REFERENCES
- [1].Lee RC, Feinbaum RL, and Ambros V, The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell, 1993. 75(5):843–54. [DOI] [PubMed] [Google Scholar]
- [2].Wightman B, Ha I, and Ruvkun G, Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell, 1993. 75(5):855–62. [DOI] [PubMed] [Google Scholar]
- [3].Lagos-Quintana M, Rauhut R, Lendeckel W, et al. , Identification of novel genes coding for small expressed RNAs. Science, 2001. 294(5543):853–8. [DOI] [PubMed] [Google Scholar]
- [4].Bartel DP, MicroRNAs: target recognition and regulatory functions. Cell, 2009. 136(2):215–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Lytle JR, Yario TA, and Steitz JA, Target mRNAs are repressed as efficiently by microRNA-binding sites in the 5’ UTR as in the 3’ UTR. Proc Natl Acad Sci U S A, 2007. 104(23):9667–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Moretti F, Thermann R, and Hentze MW, Mechanism of translational regulation by miR-2 from sites in the 5’ untranslated region or the open reading frame. RNA, 2010. 16(12):2493–502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Lee I, Ajay SS, Yook JI, et al. , New class of microRNA targets containing simultaneous 5’-UTR and 3’-UTR interaction sites. Genome Res, 2009. 19(7):1175–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Tsai NP, Lin YL, and Wei LN, MicroRNA mir-346 targets the 5’-untranslated region of receptor-interacting protein 140 (RIP140) mRNA and up-regulates its protein expression. Biochem J, 2009. 424(3):411–8. [DOI] [PubMed] [Google Scholar]
- [9].Vasudevan S, Posttranscriptional upregulation by microRNAs. Wiley Interdiscip Rev RNA, 2012. 3(3):311–30. [DOI] [PubMed] [Google Scholar]
- [10].Mehta A and Baltimore D, MicroRNAs as regulatory elements in immune system logic. Nat Rev Immunol, 2016. 16(5):279–94. [DOI] [PubMed] [Google Scholar]
- [11].Shivdasani RA, MicroRNAs: regulators of gene expression and cell differentiation. Blood, 2006. 108(12):3646–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Han J, Lee Y, Yeom KH, et al. , Molecular basis for the recognition of primary microRNAs by the Drosha-DGCR8 complex. Cell, 2006. 125(5):887–901. [DOI] [PubMed] [Google Scholar]
- [13].Faller M, Toso D, Matsunaga M, et al. , DGCR8 recognizes primary transcripts of microRNAs through highly cooperative binding and formation of higher-order structures. RNA, 2010. 16(8):1570–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Denli AM, Tops BB, Plasterk RH, et al. , Processing of primary microRNAs by the Microprocessor complex. Nature, 2004. 432(7014):231–5. [DOI] [PubMed] [Google Scholar]
- [15].Gregory RI, Yan KP, Amuthan G, et al. , The Microprocessor complex mediates the genesis of microRNAs. Nature, 2004. 432(7014):235–40. [DOI] [PubMed] [Google Scholar]
- [16].Ha M and Kim VN, Regulation of microRNA biogenesis. Nat Rev Mol Cell Biol, 2014. 15(8):509–24. [DOI] [PubMed] [Google Scholar]
- [17].Lee Y, Kim M, Han J, et al. , MicroRNA genes are transcribed by RNA polymerase II. EMBO J, 2004. 23(20):4051–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Mourelatos Z, Dostie J, Paushkin S, et al. , miRNPs: a novel class of ribonucleoproteins containing numerous microRNAs. Genes Dev, 2002. 16(6):720–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Brennecke J, Stark A, Russell RB, et al. , Principles of microRNA-target recognition. PLoS Biol, 2005. 3(3):e85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Lewis BP, Burge CB, and Bartel DP, Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell, 2005. 120(1):15–20. [DOI] [PubMed] [Google Scholar]
- [21].Kanellopoulou C, Muljo SA, Kung AL, et al. , Dicer-deficient mouse embryonic stem cells are defective in differentiation and centromeric silencing. Genes Dev, 2005. 19(4):489–501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Bernstein E, Kim SY, Carmell MA, et al. , Dicer is essential for mouse development. Nat Genet, 2003. 35(3):215–7. [DOI] [PubMed] [Google Scholar]
- [23].Muljo SA, Ansel KM, Kanellopoulou C, et al. , Aberrant T cell differentiation in the absence of Dicer. J Exp Med, 2005. 202(2):261–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Sonkoly E and Pivarcsi A, microRNAs in inflammation. Int Rev Immunol, 2009. 28(6):535–61. [DOI] [PubMed] [Google Scholar]
- [25].Sonkoly E and Pivarcsi A, Advances in microRNAs: implications for immunity and inflammatory diseases. J Cell Mol Med, 2009. 13(1):24–38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Gordon S and Martinez FO, Alternative activation of macrophages: mechanism and functions. Immunity, 2010. 32(5):593–604. [DOI] [PubMed] [Google Scholar]
- [27].Netea MG, Latz E, Mills KH, et al. , Innate immune memory: a paradigm shift in understanding host defense. Nat Immunol, 2015. 16(7):675–9. [DOI] [PubMed] [Google Scholar]
- [28].Netea MG, Quintin J, and van der Meer JW, Trained immunity: a memory for innate host defense. Cell Host Microbe, 2011. 9(5):355–61. [DOI] [PubMed] [Google Scholar]
- [29].Saeed S, Quintin J, Kerstens HH, et al. , Epigenetic programming of monocyte-to-macrophage differentiation and trained innate immunity. Science, 2014. 345(6204):1251086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Blok BA, Arts RJ, van Crevel R, et al. , Trained innate immunity as underlying mechanism for the long-term, nonspecific effects of vaccines. J Leukoc Biol, 2015. 98(3):347–56. [DOI] [PubMed] [Google Scholar]
- [31].Rivera A, Siracusa MC, Yap GS, et al. , Innate cell communication kick-starts pathogen-specific immunity. Nat Immunol, 2016. 17(4):356–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Maldonado RF, Sa-Correia I, and Valvano MA, Lipopolysaccharide modification in Gram-negative bacteria during chronic infection. FEMS Microbiol Rev, 2016. 40(4):480–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Whitfield C and Trent MS, Biosynthesis and export of bacterial lipopolysaccharides. Annu Rev Biochem, 2014. 83:99–128. [DOI] [PubMed] [Google Scholar]
- [34].Rhee SH, Lipopolysaccharide: basic biochemistry, intracellular signaling, and physiological impacts in the gut. Intest Res, 2014. 12(2):90–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Needham BD and Trent MS, Fortifying the barrier: the impact of lipid A remodelling on bacterial pathogenesis. Nat Rev Microbiol, 2013. 11(7):467–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Sansonetti PJ, War and peace at mucosal surfaces. Nat Rev Immunol, 2004. 4(12):953–64. [DOI] [PubMed] [Google Scholar]
- [37].Medzhitov R, Toll-like receptors and innate immunity. Nat Rev Immunol, 2001. 1(2):135–45. [DOI] [PubMed] [Google Scholar]
- [38].Molteni M, Gemma S, and Rossetti C, The Role of Toll-Like Receptor 4 in Infectious and Noninfectious Inflammation. Mediators Inflamm, 2016. 2016:6978936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Mogensen TH, Pathogen recognition and inflammatory signaling in innate immune defenses. Clin Microbiol Rev, 2009. 22(2):240–73, [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Satoh T and Akira S, Toll-Like Receptor Signaling and Its Inducible Proteins. Microbiol Spectr, 2016. 4(6). [DOI] [PubMed] [Google Scholar]
- [41].Shi L, Xin Q, Chai R, et al. , Ectopic expressed miR-203 contributes to chronic obstructive pulmonary disease via targeting TAK1 and PIK3CA. Int J Clin Exp Pathol, 2015. 8(9):10662–70. [PMC free article] [PubMed] [Google Scholar]
- [42].Xu Z, Dong D, Chen X, et al. , MicroRNA-381 Negatively Regulates TLR4 Signaling in A549 Cells in Response to LPS Stimulation. Biomed Res Int, 2015. 2015:849475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Zhang K, Song F, Lu X, et al. , MicroRNA-322 inhibits inflammatory cytokine expression and promotes cell proliferation in LPS-stimulated murine macrophages by targeting NF-kappaB1 (p50). Biosci Rep, 2017. 37(1). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Tang R, Pei L, Bai T, et al. , Down-regulation of microRNA-126-5p contributes to overexpression of VEGFA in lipopolysaccharide-induced acute lung injury. Biotechnol Lett, 2016. 38(8):1277–84. [DOI] [PubMed] [Google Scholar]
- [45].Li W, Qiu X, Jiang H, et al. , Downregulation of miR-181a protects mice from LPS-induced acute lung injury by targeting Bcl-2. Biomed Pharmacother, 2016. 84:1375–1382. [DOI] [PubMed] [Google Scholar]
- [46].Ye F, Tang C, Shi W, et al. , A MDM2-dependent positive-feedback loop is involved in inhibition of miR-375 and miR-106b induced by Helicobacter pylori lipopolysaccharide. Int J Cancer, 2015. 136(9):2120–31. [DOI] [PubMed] [Google Scholar]
- [47].Miao L, Liu K, Xie M, et al. , miR-375 inhibits Helicobacter pylori-induced gastric carcinogenesis by blocking JAK2-STAT3 signaling. Cancer Immunol Immunother, 2014. 63(7):699–711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Zhao GJ, Mo ZC, Tang SL, et al. , Chlamydia pneumoniae negatively regulates ABCA1 expression via TLR2-Nuclear factor-kappa B and miR-33 pathways in THP-1 macrophage-derived foam cells. Atherosclerosis, 2014. 235(2):519–25. [DOI] [PubMed] [Google Scholar]
- [49].Taganov KD, Boldin MP, Chang KJ, et al. , NF-kappaB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc Natl Acad Sci U S A, 2006. 103(33):12481–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Liu M, John CM, and Jarvis GA, Induction of endotoxin tolerance by pathogenic Neisseria is correlated with the inflammatory potential of lipooligosaccharides and regulated by microRNA-146a. J Immunol, 2014. 192(4):1768–77. [DOI] [PubMed] [Google Scholar]
- [51].Kalantari P, Harandi OF, Agarwal S, et al. , miR-718 Represses Pro-inflammatory Cytokine Production Through Targeting PTEN. J Biol Chem, 2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [52].Tili E, Croce CM, and Michaille JJ, miR-155: on the crosstalk between inflammation and cancer. Int Rev Immunol, 2009. 28(5):264–84. [DOI] [PubMed] [Google Scholar]
- [53].Lu LF, Gasteiger G, Yu IS, et al. , A Single miRNA-mRNA Interaction Affects the Immune Response in a Context- and Cell-Type-Specific Manner. Immunity, 2015. 43(1):52–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].O’Neill LA, Sheedy FJ, and McCoy CE, MicroRNAs: the fine-tuners of Toll-like receptor signalling. Nat Rev Immunol, 2011. 11(3):163–75. [DOI] [PubMed] [Google Scholar]
- [55].Schulte LN, Westermann AJ, and Vogel J, Differential activation and functional specialization of miR-146 and miR-155 in innate immune sensing. Nucleic Acids Res, 2013. 41(1):542–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Zhao XD, Lu YY, Guo H, et al. , MicroRNA-7/NF-kappaB signaling regulatory feedback circuit regulates gastric carcinogenesis. J Cell Biol, 2015. 210(4):613–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [57].Nahid MA, Satoh M, and Chan EK, MicroRNA in TLR signaling and endotoxin tolerance. Cell Mol Immunol, 2011. 8(5):388–403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Bai Y, Qian C, Qian L, et al. , Integrin CD11b negatively regulates TLR9-triggered dendritic cell cross-priming by upregulating microRNA-146a. J Immunol, 2012. 188(11):5293–302. [DOI] [PubMed] [Google Scholar]
- [59].Koch M, Mollenkopf HJ, Klemm U, et al. , Induction of microRNA-155 is TLR- and type IV secretion system-dependent in macrophages and inhibits DNA-damage induced apoptosis. Proc Natl Acad Sci U S A, 2012. 109(19):E1153–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Lu C, Huang X, Zhang X, et al. , miR-221 and miR-155 regulate human dendritic cell development, apoptosis, and IL-12 production through targeting of p27kip1, KPC1, and SOCS-1. Blood, 2011. 117(16):4293–303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [61].Blander JM and Sander LE, Beyond pattern recognition: five immune checkpoints for scaling the microbial threat. Nat Rev Immunol, 2012. 12(3):215–25. [DOI] [PubMed] [Google Scholar]
- [62].Bosio CM, Bielefeldt-Ohmann H, and Belisle JT, Active suppression of the pulmonary immune response by Francisella tularensis Schu4. J Immunol, 2007. 178(7):4538–47. [DOI] [PubMed] [Google Scholar]
- [63].Jones BD, Faron M, Rasmussen JA, et al. , Uncovering the components of the Francisella tularensis virulence stealth strategy. Front Cell Infect Microbiol, 2014. 4:32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [64].Bandyopadhyay S, Long ME, and Allen LA, Differential expression of microRNAs in Francisella tularensis-infected human macrophages: miR-155-dependent downregulation of MyD88 inhibits the inflammatory response. PLoS One, 2014. 9(10):e109525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [65].Cremer TJ, Ravneberg DH, Clay CD, et al. , MiR-155 induction by F. novicida but not the virulent F. tularensis results in SHIP down-regulation and enhanced pro-inflammatory cytokine response. PLoS One, 2009. 4(12):e8508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [66].O’Connell RM, Chaudhuri AA, Rao DS, et al. , Inositol phosphatase SHIP1 is a primary target of miR-155. Proc Natl Acad Sci U S A, 2009. 106(17):7113–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [67].Arkatkar T, Gupta R, Li W, et al. , Murine MicroRNA-214 regulates intracellular adhesion molecule (ICAM1) gene expression in genital Chlamydia muridarum infection. Immunology, 2015. 145(4):534–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [68].Nakae S, Komiyama Y, Nambu A, et al. , Antigen-specific T cell sensitization is impaired in IL-17-deficient mice, causing suppression of allergic cellular and humoral responses. Immunity, 2002. 17(3):375–87. [DOI] [PubMed] [Google Scholar]
- [69].Andrew DW, Cochrane M, Schripsema JH, et al. , The duration of Chlamydia muridarum genital tract infection and associated chronic pathological changes are reduced in IL-17 knockout mice but protection is not increased further by immunization. PLoS One, 2013. 8(9):e76664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [70].Gu C, Wu L, and Li X, IL-17 family: cytokines, receptors and signaling. Cytokine, 2013. 64(2):477–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [71].Barnett RE, Keskey RC, Rao JM, et al. , Poor outcome in bacterial peritonitis is associated with dysregulated microRNAs and an increased inflammatory response. Surgery, 2013. 154(3):521–7. [DOI] [PubMed] [Google Scholar]
- [72].Kanaan Z, Barnett R, Gardner S, et al. , Differential microRNA (miRNA) expression could explain microbial tolerance in a novel chronic peritonitis model. Innate Immun, 2013. 19(2):203–12. [DOI] [PubMed] [Google Scholar]
- [73].Biswas SK and Lopez-Collazo E, Endotoxin tolerance: new mechanisms, molecules and clinical significance. Trends Immunol, 2009. 30(10):475–87. [DOI] [PubMed] [Google Scholar]
- [74].Foster SL, Hargreaves DC, and Medzhitov R, Gene-specific control of inflammation by TLR-induced chromatin modifications. Nature, 2007. 447(7147):972–8. [DOI] [PubMed] [Google Scholar]
- [75].Gao C, Zhang Z, Liu W, et al. , Reduced microRNA-218 expression is associated with high nuclear factor kappa B activation in gastric cancer. Cancer, 2010. 116(1):41–9. [DOI] [PubMed] [Google Scholar]
- [76].Schulte LN, Eulalio A, Mollenkopf HJ, et al. , Analysis of the host microRNA response to Salmonella uncovers the control of major cytokines by the let-7 family. EMBO J, 2011. 30(10):1977–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [77].Teng GG, Wang WH, Dai Y, et al. , Let-7b is involved in the inflammation and immune responses associated with Helicobacter pylori infection by targeting Toll-like receptor 4. PLoS One, 2013. 8(2):e56709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [78].Lochhead RB, Ma Y, Zachary JF, et al. , MicroRNA-146a provides feedback regulation of lyme arthritis but not carditis during infection with Borrelia burgdorferi. PLoS Pathog, 2014. 10(6):e1004212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [79].Kim RY, Horvat JC, Pinkerton JW, et al. , MicroRNA-21 drives severe, steroid-insensitive experimental asthma by amplifying phosphoinositide 3-kinase-mediated suppression of histone deacetylase 2. J Allergy Clin Immunol, 2016. [DOI] [PubMed] [Google Scholar]
- [80].Das A, Ganesh K, Khanna S, et al. , Engulfment of apoptotic cells by macrophages: a role of microRNA-21 in the resolution of wound inflammation. J Immunol, 2014. 192(3):1120–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [81].Roy S, Elgharably H, Sinha M, et al. , Mixed-species biofilm compromises wound healing by disrupting epidermal barrier function. J Pathol, 2014. 233(4):331–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [82].Mantegazza AR, Magalhaes JG, Amigorena S, et al. , Presentation of phagocytosed antigens by MHC class I and II. Traffic, 2013. 14(2):135–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [83].Germain RN, T-cell development and the CD4-CD8 lineage decision. Nat Rev Immunol, 2002. 2(5):309–22. [DOI] [PubMed] [Google Scholar]
- [84].Jeker LT and Bluestone JA, MicroRNA regulation of T-cell differentiation and function. Immunol Rev, 2013. 253(1):65–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [85].Kaplan MH, Th9 cells: differentiation and disease. Immunol Rev, 2013. 252(1):104–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [86].Schmitt E, Klein M, and Bopp T, Th9 cells, new players in adaptive immunity. Trends Immunol, 2014. 35(2):61–8. [DOI] [PubMed] [Google Scholar]
- [87].Schwartz RH, T cell anergy. Annu Rev Immunol, 2003. 21:305–34. [DOI] [PubMed] [Google Scholar]
- [88].Marcais A, Blevins R, Graumann J, et al. , microRNA-mediated regulation of mTOR complex components facilitates discrimination between activation and anergy in CD4 T cells. J Exp Med, 2014. 211(11):2281–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [89].Merkenschlager M and Marcais A, microRNAs calibrate T cell responses by regulating mTOR. Oncotarget, 2015. 6(33):34059–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [90].Zhou X, Li L, Su J, et al. , Decreased miR-204 in H. pylori-associated gastric cancer promotes cancer cell proliferation and invasion by targeting SOX4. PLoS One, 2014. 9(7):e101457. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- [91].Kuwahara M, Yamashita M, Shinoda K, et al. , The transcription factor Sox4 is a downstream target of signaling by the cytokine TGF-beta and suppresses T(H)2 differentiation. Nat Immunol, 2012. 13(8):778–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [92].Li N, Xie C, and Lu NH, Transforming growth factor-beta: an important mediator in Helicobacter pylori-associated pathogenesis. Front Cell Infect Microbiol, 2015. 5:77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [93].Kivrak Salim D, Sahin M, Koksoy S, et al. , Local Immune Response in Helicobacter pylori Infection. Medicine (Baltimore), 2016. 95(20):e3713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [94].Rodriguez A, Vigorito E, Clare S, et al. , Requirement of bic/microRNA-155 for normal immune function. Science, 2007. 316(5824):608–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [95].Oertli M, Engler DB, Kohler E, et al. , MicroRNA-155 is essential for the T cell-mediated control of Helicobacter pylori infection and for the induction of chronic Gastritis and Colitis. J Immunol, 2011. 187(7):3578–86. [DOI] [PubMed] [Google Scholar]
- [96].Vigorito E, Perks KL, Abreu-Goodger C, et al. , microRNA-155 regulates the generation of immunoglobulin class-switched plasma cells. Immunity, 2007. 27(6):847–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [97].Tang T, Wong HK, Gu W, et al. , MicroRNA-182 plays an onco-miRNA role in cervical cancer. Gynecol Oncol, 2013. 129(1):199–208. [DOI] [PubMed] [Google Scholar]
- [98].Gupta R, Arkatkar T, Keck J, et al. , Antigen specific immune response in Chlamydia muridarum genital infection is dependent on murine microRNAs-155 and -182. Oncotarget, 2016. 7(40):64726–64742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [99].Kanellopoulou C and Muljo SA, Fine-Tuning Th17 Cells: To Be or Not To Be Pathogenic? Immunity, 2016. 44(6):1241–3. [DOI] [PubMed] [Google Scholar]
- [100].Ichiyama K, Gonzalez-Martin A, Kim BS, et al. , The MicroRNA-183-96-182 Cluster Promotes T Helper 17 Cell Pathogenicity by Negatively Regulating Transcription Factor Foxo1 Expression. Immunity, 2016. 44(6):1284–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [101].Ma F, Xu S, Liu X, et al. , The microRNA miR-29 controls innate and adaptive immune responses to intracellular bacterial infection by targeting interferon-gamma. Nat Immunol, 2011. 12(9):861–9. [DOI] [PubMed] [Google Scholar]
- [102].Aalaei-Andabili SH and Rezaei N, MicroRNAs (MiRs) Precisely Regulate Immune System Development and Function in Immunosenescence Process. Int Rev Immunol, 2016. 35(1):57–66. [DOI] [PubMed] [Google Scholar]
- [103].Fitzgerald JB, George J, and Christenson LK, Non-coding RNA in Ovarian Development and Disease. Adv Exp Med Biol, 2016. 886:79–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [104].Navakanitworakul R, Hung WT, Gunewardena S, et al. , Characterization and Small RNA Content of Extracellular Vesicles in Follicular Fluid of Developing Bovine Antral Follicles. Sci Rep, 2016. 6:25486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [105].Ma X, Kong L, and Zhu S, Reprogramming cell fates by small molecules. Protein Cell, 2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [106].Staedel C and Darfeuille F, MicroRNAs and bacterial infection. Cell Microbiol, 2013. 15(9):1496–507. [DOI] [PubMed] [Google Scholar]
- [107].Eulalio A, Schulte L, and Vogel J, The mammalian microRNA response to bacterial infections. RNA Biol, 2012. 9(6):742–50. [DOI] [PubMed] [Google Scholar]
- [108].Maudet C, Mano M, and Eulalio A, MicroRNAs in the interaction between host and bacterial pathogens. FEBS Lett, 2014. 588(22):4140–7. [DOI] [PubMed] [Google Scholar]
- [109].Igietseme JU, Omosun Y, Stuchlik O, et al. , Role of Epithelial-Mesenchyme Transition in Chlamydia Pathogenesis. PLoS One, 2015. 10(12):e0145198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [110].Yeruva L, Pouncey DL, Eledge MR, et al. , MicroRNAs Modulate Pathogenesis Resulting from Chlamydial Infection in Mice. Infect Immun, 2017. 85(1). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [111].Peng W, Lu DQ, Li GF, et al. , Two distinct interferon-gamma genes in Tetraodon nigroviridis: Functional analysis during Vibrio parahaemolyticus infection. Mol Immunol, 2016. 70:34–46. [DOI] [PubMed] [Google Scholar]
- [112].Kim RY, Horvat JC, Pinkerton JW, et al. , MicroRNA-21 drives severe, steroid-insensitive experimental asthma by amplifying phosphoinositide 3-kinase-mediated suppression of histone deacetylase 2. J Allergy Clin Immunol, 2017. 139(2):519–532. [DOI] [PubMed] [Google Scholar]
- [113].Brown BD and Naldini L, Exploiting and antagonizing microRNA regulation for therapeutic and experimental applications. Nat Rev Genet, 2009. 10(8):578–85. [DOI] [PubMed] [Google Scholar]
- [114].Ling H, Fabbri M, and Calin GA, MicroRNAs and other non-coding RNAs as targets for anticancer drug development. Nat Rev Drug Discov, 2013. 12(11):847–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [115].Baltimore D, Boldin M, and Taganov K, Modulating immune system development and function through microrna mir-146. 2009, Google Patents.
- [116].Meyers R, Bettencourt B, WONG JE, et al. , Viral attenuation and vaccine production. 2013, Google Patents.
- [117].Nair V and Lambeth L, Vector. 2009, Google Patents.
- [118].Tenoever B, Live attenuated influenza virus vaccines comprising microrna response elements. 2012, Google Patents.
- [119].Guo J, Lv J, Liu M, et al. , MiR-346 Up-regulates Argonaute 2 (AGO2) Protein Expression to Augment the Activity of Other MiRNAs and Contributes to Cervical Cancer Cell Malignancy. J Biol Chem, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [120].Sharma G, Dua P, and Agarwal SM, A Comprehensive Review of Dysregulated miRNAs Involved in Cervical Cancer. Curr Genomics, 2014. 15(4):310–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [121].van den Akker EK, Dor FJ, JN IJ, et al. , MicroRNAs in Kidney Transplantation: Living up to Their Expectations? J Transplant, 2015. 2015:354826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [122].Raschzok N, Sallmon H, Pratschke J, et al. , MicroRNAs in liver tissue engineering - New promises for failing organs. Adv Drug Deliv Rev, 2015. 88:67–77. [DOI] [PubMed] [Google Scholar]
- [123].Fishman JA, What’s new and what’s hot? Basic science at the American Transplant Congress 2012. Am J Transplant, 2013. 13(2):275–80. [DOI] [PubMed] [Google Scholar]
- [124].Vela EM, Kasoji MD, Wendling MQ, et al. , MicroRNA expression in mice infected with seasonal H1N1, swine H1N1 or highly pathogenic H5N1. J Med Microbiol, 2014. 63(Pt 9):1131–42. [DOI] [PubMed] [Google Scholar]
- [125].Perez JT, Pham AM, Lorini MH, et al. , MicroRNA-mediated species-specific attenuation of influenza A virus. Nat Biotechnol, 2009. 27(6):572–6. [DOI] [PubMed] [Google Scholar]
- [126].Janssen HL, Reesink HW, Lawitz EJ, et al. , Treatment of HCV infection by targeting microRNA. N Engl J Med, 2013. 368(18):1685–94. [DOI] [PubMed] [Google Scholar]
- [127].Thibault PA and Wilson JA, Targeting miRNAs to treat Hepatitis C Virus infections and liver pathology: Inhibiting the virus and altering the host. Pharmacol Res, 2013. 75:48–59. [DOI] [PubMed] [Google Scholar]
- [128].Xiong Y, Chen S, Liu L, et al. , Increased serum microRNA-155 level associated with nonresponsiveness to hepatitis B vaccine. Clin Vaccine Immunol, 2013. 20(7):1089–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [129].Zhang B, Liu Y, Wang X, et al. , A novel recombinant Salmonella vaccine enhances the innate immunity of NK cells against acute myeloid leukaemia cells Kasumi-1 in vitro. Cell Biol Int, 2013. 37(12):1320–9. [DOI] [PubMed] [Google Scholar]
- [130].Singh Y, Kaul V, Mehra A, et al. , Mycobacterium tuberculosis controls microRNA-99b (miR-99b) expression in infected murine dendritic cells to modulate host immunity. J Biol Chem, 2013. 288(7):5056–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [131].Koch M, Meyer TF, and Moss SF, Inflammation, immunity, vaccines for Helicobacter pylori infection. Helicobacter, 2013. 18 Suppl 1:18–23. [DOI] [PubMed] [Google Scholar]
- [132].Lauring AS, Jones JO, and Andino R, Rationalizing the development of live attenuated virus vaccines. Nat Biotechnol, 2010. 28(6):573–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [133].Holmstrom K, Pedersen AW, Claesson MH, et al. , Identification of a microRNA signature in dendritic cell vaccines for cancer immunotherapy. Hum Immunol, 2010. 71(1):67–73. [DOI] [PubMed] [Google Scholar]
- [134].Bela-ong DB, Schyth BD, Zou J, et al. , Involvement of two microRNAs in the early immune response to DNA vaccination against a fish rhabdovirus. Vaccine, 2015. 33(28):3215–22. [DOI] [PubMed] [Google Scholar]
- [135].Hentzschel F, Hammerschmidt-Kamper C, Borner K, et al. , AAV8-mediated in vivo overexpression of miR-155 enhances the protective capacity of genetically attenuated malarial parasites. Mol Ther, 2014. 22(12):2130–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [136].Kota J, Chivukula RR, O’Donnell KA, et al. , Therapeutic microRNA delivery suppresses tumorigenesis in a murine liver cancer model. Cell, 2009. 137(6):1005–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [137].Esposito CL, Cerchia L, Catuogno S, et al. , Multifunctional aptamer-miRNA conjugates for targeted cancer therapy. Mol Ther, 2014. 22(6):1151–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [138].Liang X, Liu Y, Mei S, et al. , MicroRNA-22 impairs anti-tumor ability of dendritic cells by targeting p38. PLoS One, 2015. 10(3):e0121510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [139].Kouri FM, Hurley LA, Daniel WL, et al. , miR-182 integrates apoptosis, growth, and differentiation programs in glioblastoma. Genes Dev, 2015. 29(7):732–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [140].de Candia P, Torri A, Pagani M, et al. , Serum microRNAs as Biomarkers of Human Lymphocyte Activation in Health and Disease. Front Immunol, 2014. 5:43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [141].Bader AG, miR-34 - a microRNA replacement therapy is headed to the clinic. Front Genet, 2012. 3:120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [142].Misso G, Di Martino MT, De Rosa G, et al. , Mir-34: a new weapon against cancer? Mol Ther Nucleic Acids, 2014. 3:e194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [143].Lieberman J and Sarnow P, Micromanaging hepatitis C virus. N Engl J Med, 2013. 368(18):1741–3. [DOI] [PubMed] [Google Scholar]
- [144].Jopling CL, Norman KL, and Sarnow P, Positive and negative modulation of viral and cellular mRNAs by liver-specific microRNA miR-122. Cold Spring Harb Symp Quant Biol, 2006. 71:369–76. [DOI] [PubMed] [Google Scholar]
- [145].Jopling CL, Yi M, Lancaster AM, et al. , Modulation of hepatitis C virus RNA abundance by a liver-specific MicroRNA. Science, 2005. 309(5740):1577–81. [DOI] [PubMed] [Google Scholar]
- [146].Jain CK, Gupta A, Dogra N, et al. , MicroRNA therapeutics: the emerging anticancer strategies. Recent Pat Anticancer Drug Discov, 2014. 9(3):286–96. [DOI] [PubMed] [Google Scholar]
- [147].Li Z and Rana TM, Therapeutic targeting of microRNAs: current status and future challenges. Nat Rev Drug Discov, 2014. 13(8):622–38. [DOI] [PubMed] [Google Scholar]
- [148].Fellmann C and Lowe SW, Stable RNA interference rules for silencing. Nat Cell Biol, 2014. 16(1):10–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [149].Xu F, Liu G, Liu Q, et al. , RNA interference of influenza A virus replication by microRNA-adapted lentiviral loop short hairpin RNA. J Gen Virol, 2015. 96(10):2971–81. [DOI] [PubMed] [Google Scholar]
- [150].Kasinski AL, Kelnar K, Stahlhut C, et al. , A combinatorial microRNA therapeutics approach to suppressing non-small cell lung cancer. Oncogene, 2015. 34(27):3547–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [151].Cheng SF, Li L, and Wang LM, miR-155 and miR-146b negatively regulates IL6 in Helicobacter pylori (cagA+) infected gastroduodenal ulcer. Eur Rev Med Pharmacol Sci, 2015. 19(4):607–13. [PubMed] [Google Scholar]
- [152].Craig VJ, Cogliatti SB, Rehrauer H, et al. , Epigenetic silencing of microRNA-203 dysregulates ABL1 expression and drives Helicobacter-associated gastric lymphomagenesis. Cancer Res, 2011. 71(10):3616–24. [DOI] [PubMed] [Google Scholar]
- [153].Derrick T, Last AR, Burr SE, et al. , Inverse relationship between microRNA-155 and -184 expression with increasing conjunctival inflammation during ocular Chlamydia trachomatis infection. BMC Infect Dis, 2016. 16:60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [154].Derrick T, Roberts C, Rajasekhar M, et al. , Conjunctival MicroRNA expression in inflammatory trachomatous scarring. PLoS Negl Trop Dis, 2013. 7(3):e2117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [155].Fassi Fehri L, Koch M, Belogolova E, et al. , Helicobacter pylori induces miR-155 in T cells in a cAMP-Foxp3-dependent manner. PLoS One, 2010. 5(3):e9500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [156].Feng Y, Wang L, Zeng J, et al. , FoxM1 is overexpressed in Helicobacter pylori-induced gastric carcinogenesis and is negatively regulated by miR-370. Mol Cancer Res, 2013. 11(8):834–44. [DOI] [PubMed] [Google Scholar]
- [157].Gupta R, Arkatkar T, Yu JJ, et al. , Chlamydia muridarum infection associated host MicroRNAs in the murine genital tract and contribution to generation of host immune response. Am J Reprod Immunol, 2015. 73(2):126–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [158].Hishida A, Matsuo K, Goto Y, et al. , Combined effect of miR-146a rs2910164 G/C polymorphism and Toll-like receptor 4 +3725 G/C polymorphism on the risk of severe gastric atrophy in Japanese. Dig Dis Sci, 2011. 56(4):1131–7. [DOI] [PubMed] [Google Scholar]
- [159].Huang WT, Kuo SH, Cheng AL, et al. , Inhibition of ZEB1 by miR-200 characterizes Helicobacter pylori-positive gastric diffuse large B-cell lymphoma with a less aggressive behavior. Mod Pathol, 2014. 27(8):1116–25. [DOI] [PubMed] [Google Scholar]
- [160].Ishimoto T, Izumi D, Watanabe M, et al. , Chronic inflammation with Helicobacter pylori infection is implicated in CD44 overexpression through miR-328 suppression in the gastric mucosa. J Gastroenterol, 2015. 50(7):751–7. [DOI] [PubMed] [Google Scholar]
- [161].Kaakoush NO, Deshpande NP, Man SM, et al. , Transcriptomic and proteomic analyses reveal key innate immune signatures in the host response to the gastrointestinal pathogen Campylobacter concisus. Infect Immun, 2015. 83(2):832–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [162].Kiga K, Mimuro H, Suzuki M, et al. , Epigenetic silencing of miR-210 increases the proliferation of gastric epithelium during chronic Helicobacter pylori infection. Nat Commun, 2014. 5:4497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [163].Lochhead RB, Zachary JF, Dalla Rosa L, et al. , Antagonistic Interplay between MicroRNA-155 and IL-10 during Lyme Carditis and Arthritis. PLoS One, 2015. 10(8):e0135142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [164].Lv X, Song H, Yang J, et al. , A multi-epitope vaccine CTB-UE relieves Helicobacter pylori-induced gastric inflammatory reaction via up-regulating microRNA-155 to inhibit Th17 response in C57/BL6 mice model. Hum Vaccin Immunother, 2014. 10(12):3561–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [165].Lv X, Yang J, Song H, et al. , Therapeutic efficacy of the multi-epitope vaccine CTB-UE against Helicobacter pylori infection in a Mongolian gerbil model and its microRNA-155-associated immuno-protective mechanism. Vaccine, 2014. 32(41):5343–52. [DOI] [PubMed] [Google Scholar]
- [166].Nayar G, Gauna A, Chukkapalli S, et al. , Polymicrobial infection alter inflammatory microRNA in rat salivary glands during periodontal disease. Anaerobe, 2016. 38:70–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [167].Nosho K, Igarashi H, Nojima M, et al. , Association of microRNA-31 with BRAF mutation, colorectal cancer survival and serrated pathway. Carcinogenesis, 2014. 35(4):776–83. [DOI] [PubMed] [Google Scholar]
- [168].Ordas A, Kanwal Z, Lindenberg V, et al. , MicroRNA-146 function in the innate immune transcriptome response of zebrafish embryos to Salmonella typhimurium infection. BMC Genomics, 2013. 14:696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [169].Qi Y, Cui L, Ge Y, et al. , Altered serum microRNAs as biomarkers for the early diagnosis of pulmonary tuberculosis infection. BMC Infect Dis, 2012. 12:384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [170].Qi Y, Zhu Z, Shi Z, et al. , Dysregulated microRNA expression in serum of non-vaccinated children with varicella. Viruses, 2014. 6(4):1823–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [171].Saito Y, Suzuki H, Tsugawa H, et al. , Overexpression of miR-142–5p and miR-155 in gastric mucosa-associated lymphoid tissue (MALT) lymphoma resistant to Helicobacter pylori eradication. PLoS One, 2012. 7(11):e47396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [172].Shen J, Xiao Z, Wu WK, et al. , Epigenetic silencing of miR-490-3p reactivates the chromatin remodeler SMARCD1 to promote Helicobacter pylori-induced gastric carcinogenesis. Cancer Res, 2015. 75(4):754–65. [DOI] [PubMed] [Google Scholar]
- [173].Suzuki R, Yamamoto E, Nojima M, et al. , Aberrant methylation of microRNA-34b/c is a predictive marker of metachronous gastric cancer risk. J Gastroenterol, 2014. 49(7):1135–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [174].Uribe JH, Collado-Romero M, Zaldivar-Lopez S, et al. , Transcriptional analysis of porcine intestinal mucosa infected with Salmonella Typhimurium revealed a massive inflammatory response and disruption of bile acid absorption in ileum. Vet Res, 2016. 47:11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [175].Voinnet O, Micro-balancing innate immunity to Salmonella. EMBO J, 2011. 30(10):1877–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [176].Wang W, Stassen FR, Surcel HM, et al. , Analyses of polymorphisms in the inflammasome-associated NLRP3 and miRNA-146A genes in the susceptibility to and tubal pathology of Chlamydia trachomatis infection. Drugs Today (Barc), 2009. 45 Suppl B:95–103. [PubMed] [Google Scholar]
- [177].Wu K, Yang L, Li C, et al. , MicroRNA-146a enhances Helicobacter pylori induced cell apoptosis in human gastric cancer epithelial cells. Asian Pac J Cancer Prev, 2014. 15(14):5583–6. [DOI] [PubMed] [Google Scholar]
- [178].Wu K, Zhu C, Yao Y, et al. , MicroRNA-155-enhanced autophagy in human gastric epithelial cell in response to Helicobacter pylori. Saudi J Gastroenterol, 2016. 22(1):30–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [179].Xiao B, Liu Z, Li BS, et al. , Induction of microRNA-155 during Helicobacter pylori infection and its negative regulatory role in the inflammatory response. J Infect Dis, 2009. 200(6):916–25. [DOI] [PubMed] [Google Scholar]
- [180].Xiao B, Zhu ED, Li N, et al. , Increased miR-146a in gastric cancer directly targets SMAD4 and is involved in modulating cell proliferation and apoptosis. Oncol Rep, 2012. 27(2):559–66. [DOI] [PubMed] [Google Scholar]
- [181].Yan H, Chen Y, Zhou S, et al. , Expression Profile Analysis of miR-221 and miR-222 in Different Tissues and Head Kidney Cells of Cynoglossus semilaevis, Following Pathogen Infection. Mar Biotechnol (NY), 2016. 18(1):37–48. [DOI] [PubMed] [Google Scholar]
- [182].Yao M, Gao W, Tao H, et al. , Regulation signature of miR-143 and miR-26 in porcine Salmonella infection identified by binding site enrichment analysis. Mol Genet Genomics, 2016. 291(2):789–99. [DOI] [PubMed] [Google Scholar]
- [183].Adams BD, Parsons C, Walker L, et al. , Targeting noncoding RNAs in disease. J Clin Invest, 2017. 127(3):761–771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [184].Rupaimoole R and Slack FJ, MicroRNA therapeutics: towards a new era for the management of cancer and other diseases. Nat Rev Drug Discov, 2017. 16(3):203–222. [DOI] [PubMed] [Google Scholar]
- [185].Paul P, Chakraborty A, Sarkar D, et al. , Interplay between miRNAs and Human Diseases: A Review. J Cell Physiol, 2017. [DOI] [PubMed] [Google Scholar]