

Abstract
The ability to precisely modify human genes has been made possible by the development of tools such as meganucleases, zinc finger nucleases, TALENs, and CRISPR/Cas. These now make it possible to generate targeted deletions, insertions, gene knock outs, and point variants; to modulate gene expression by targeting transcription factors or epigenetic machineries to DNA; or to target and modify RNA. Endogenous repair mechanisms are used to make the modifications required in DNA; they include non-homologous end joining, homology-directed repair, homology-independent targeted integration, microhomology-mediated end joining, base-excision repair, and mismatch repair. Off-target effects can be monitored using in silico prediction and sequencing and minimized using Cas proteins with higher accuracy, such as high-fidelity Cas9, enhanced-specificity Cas9, and hyperaccurate Cas9. Alternatives to Cas9 have been identified, including Cpf1, Cas12a, Cas12b, and smaller Cas9 orthologs such as CjCas9. Delivery of gene-editing components is performed ex vivo using standard techniques or in vivo using AAV, lipid nanoparticles, or cell-penetrating peptides. Clinical development of gene-editing technology is progressing in several fields, including immunotherapy in cancer treatment, antiviral therapy for HIV infection, and treatment of genetic disorders such as β-thalassemia, sickle cell disease, lysosomal storage disorders, and retinal dystrophy. Here we review these technological advances and the challenges to their clinical implementation.
Subject Areas: Genetics, Techniques in Genetics, Molecular Biology
Graphical Abstract
Genetics; Techniques in Genetics; Molecular Biology
Introduction
In recent years, various platforms for genetically engineering somatic and pluripotent stem cells have been developed. They include zinc finger nucleases (ZFN), transcription activator-like effector nucleases (TALENs), meganucleases (MNs), and clustered regularly interspaced short palindromic repeats (CRISPR) in combination with CRISPR-associated protein (CRISPR/Cas). From agriculture to biomedical science, these platforms are being explored in various fields and, more recently, in the first clinical trials (Barrangou and Doudna, 2016, Naldini, 2015, Rodriguez-Rodriguez et al., 2019, Zhan et al., 2019).
Several developments are taking place in parallel. First, technological improvements and variations on gene-editing strategies are being reported with unsurpassed speed. Second, many possible applications are being developed to address a wide variety of biomedical questions. Third, the first platforms for gene editing are entering the clinical testing stage. Here, we follow these themes to present an overview of these recent developments, focusing on the likely clinical implementation of gene-editing strategies as well as discussing recent technological advances, and aspects such as safety, efficacy, and delivery that are relevant to clinical implementation. We provide a short overview of the progress gene editing is making toward applications in the clinic.
Technological Advances
Basics of CRISPR/Cas
Genome editing depends on the ability to generate specific pre-designed alterations in the genome. Inducing double-strand breaks (DSBs), single-strand breaks (SSBs) (also termed “nicks”), or specific base changes result in the activation of endogenous repair mechanisms that can be used to alter the genome. MNs, ZFNs, and TALENs were the first tools for genome editing in mammalian cells. MNs are naturally occurring endonucleases and can be re-targeted to new sites. Gene editing with ZFNs and TALENS results from the fusion of the FokI nuclease domain to the DNA-binding modules of zinc finger proteins (in the case of ZFNs) or transcription activator-like effector proteins (TALEs) (in the case of TALENS). However, the design and construction of these gene-editing tools can be labor-intensive, and their efficiencies for performing gene-editing varies. The discovery of CRISPR/Cas as a new gene-editing platform introduced a fast, cheap, and relatively efficient genome-editing method that revolutionized genome engineering.
The mechanism of CRISPR/Cas is based on its role in adaptive immunity in prokaryotes, in which short stretches of invading foreign nucleic acids, so-called protospacers, are incorporated into the CRISPR locus of the bacterial or archaeal genome. After acquisition, CRISPR RNA (crRNA) is generated from the protospacers at the CRISPR locus. This crRNA can bind to complementary foreign nucleic acid and directs the Cas protein to recognize invading sequences. A second RNA known as the trans-activating CRISPR RNA (tracrRNA) is transcribed from a genomic locus upstream of the CRISPR locus and forms a complex with the crRNA. The crRNA:tracrRNA complex associates with a Cas protein (the nuclease) and creates an active ribonucleoprotein (RNP) complex that targets foreign nucleic acids for degradation (Mojica and Montoliu, 2016, Maeder and Gersbach, 2016).
For genome editing, the tracrRNA and crRNA are fused into a single guide RNA (sgRNA), which binds complementarily to a DNA target and guides the Cas protein to the desired target site, creating a DSB (Jinek et al., 2012). The target sequence is based on the presence of a protospacer adjacent motif (PAM), which is an absolute prerequisite for Cas protein to induce a DSB. The first publications on CRISPR arrays date back as early as 1987 (Ishino et al., 1987), Cas genes were discovered in 2002 (Jansen et al., 2002), and CRISPR/Cas was shown to cleave bacteriophage and plasmid DNA in vivo at specific sites in 2010 (Garneau et al., 2010). However, it was not until 2012 that two groups (Gasiunas et al., 2012, Jinek et al., 2012) adapted this system into a gene-editing tool. A review by Fernández and colleagues provides an extensive history of genome editing tools (Fernandez et al., 2017).
Its accessibility and relatively low costs have brought CRISPR/Cas a large number of applications in research worldwide and have catalyzed further research on understanding its mechanism of action, improving its functional capacities, and extending its biomedical applications. The CRISPR/Cas9 system originally applied has downsides that include a lower specificity than other gene-editing tools and a relatively large cargo size that hampers delivery to cells via vectors with limited cargo-size capacity (Fernandez et al., 2017, Guha and Edgell, 2017, Guha et al., 2017, Gupta and Musunuru, 2014, Zych et al., 2018). However, as we discuss below, innovative research has produced many adaptations to the original system that enhance its versatility and improve properties such as specificity and efficacy (Zhang et al., 2016, Wu et al., 2018).
Versatility of CRISPR/Cas-Mediated Gene Editing
As nucleases merely induce breaks in the DNA, the introduction of specific alterations in the genetic code requires the exploitation of distinct DNA-repair mechanisms. Intelligent engineering of the original genome-editing tools has broadened the scope of this toolkit, making it possible to achieve a great number of applications. As a result, CRISPR/Cas can be applied to interfere at multiple steps of gene-expression processes and can target processes at genomic and transcriptomic levels. Approaches include gene knock-out, precise correction of disease-associated variants, insertion of a cDNA in a safe harbor (a location in the genome—such as the AAVS1 locus—where there is no risk of insertional mutagenesis) (van der Wal et al., 2018, Sadelain et al., 2011); and manipulation of gene-expression regulatory elements such as promoter activity (Baliou et al., 2018) or splicing (Bergsma et al., 2018, Smith et al., 2018).
Targeted Deletion and Gene Knock-Out
The DNA repair pathway that is most commonly used to create deletions and specific gene knock-outs is non-homologous end joining (NHEJ). NHEJ repairs blunt or incompatible double strands, either by direct ligation or through mediation by microhomology of 5-25 nucleotides that flank the DSB to facilitate end joining (Chang et al., 2017a). NHEJ represents the major double-strand break repair system in mammalian cells and is active throughout the cell cycle (Chang et al., 2017a, Ranjha et al., 2018). Repair can be precise, leaving the target site intact for recleaving by the Cas nuclease. However, NHEJ-mediated DNA repair is rather error prone, as it can introduce random insertions or deletions of base pairs (indels) that will destroy the target site. These errors can result in a frameshift that inactivates gene products through mRNA decay (Ranjha et al., 2018). This effect is exploited in gene editing to create targeted gene knock-outs. Examples of clinical applications include knocking out disease-promoting genes such as oncogenes or restoring a reading frame by interfering with splice sites such as in Duchenne muscular dystrophy (Amoasii et al., 2018, Tabebordbar et al., 2016, Nelson et al., 2016).
Targeted Gene Editing and Knock-In
Traditionally, the homology-directed repair (HDR) pathway is used to achieve a precise knock-in. HDR recognizes DSBs and utilizes a homologous template—which, under physiological conditions, is the sister chromatid—to repair the defect (Ranjha et al., 2018). For precision gene editing, this pathway is exploited by introducing a donor template that is used in the repair of the DSB rather than the sister chromatid. This donor template has the desired alterations, which are flanked by 3′ and 5′ homology arms on both sites of the DSB. In this manner, it is possible to accurately correct a point variant (as in cystic fibrosis) (Schwank et al., 2013); a tandem repeat (as in Fragile X syndrome) (Xie et al., 2016); large inserts or deletions (as in Duchenne dystrophy) (Li et al., 2015); or other genetic defects.
Due to the use of a donor template, HDR is a very precise repair pathway that is less error-prone than NHEJ. However, HDR is less active in cells and therefore much less efficient in genome editing than NHEJ, which limits its clinical potential. To favor its activation over NHEJ induction, its efficiency would have to be increased. This has cast new light on research into the regulation of DNA repair pathways (Mateos-Gomez et al., 2017, Schimmel et al., 2017, Zelensky et al., 2017). Several strategies using small molecules to inhibit NHEJ have been followed with varying success (Chu et al., 2015, Maruyama et al., 2015, Song et al., 2016, Yu et al., 2015, Pinder et al., 2015). Recently, cold shock was found to increase HDR in cells in vitro (Guo et al., 2018a). As HDR is highly suppressed in the G1 phase and its activity is limited to the S and G2/M phases of the cell cycle, it is restricted to dividing cells (Ranjha et al., 2018), which, since many cells in the human body are non-dividing, greatly restricts the clinical potential of HDR-based gene editing. Recently, Orthwein et al. have shown reactivation of HDR in the G1 phase via the PALB2-BRCA1/CUL3/Keap1 pathway. Although this reactivation might allow therapeutic targeting of non-dividing cell types, the limited efficiency seen to date would indicate the need for further research to demonstrate that therapeutically relevant HDR levels can be achieved through G1 reactivation (Orthwein et al., 2015).
More recently, other pathways have been used to achieve targeted knock-in. Through a technique named homology-independent targeted integration (HITI), NHEJ can also be harnessed to create knock-ins (Suzuki and Izpisua Belmonte, 2018). After a donor vector containing the desired transgene flanked by a CRISPR target site has been introduced into the cell, the donor vector and the genomic target are cleaved by Cas9, generating blunt ends on both genomic target and donor vector. These blunt ends are utilized to induce NHEJ-mediated end-to-end ligation, allowing the integration of the donor sequence into the target location (Sawatsubashi et al., 2018). As the activity of NHEJ is higher than that of HDR, this approach can be more effective and can be used in non-dividing cell types, because NHEJ remains active in all phases of the cell cycle (Suzuki and Izpisua Belmonte, 2018).
Several disadvantages limit the clinical potential of NHEJ-mediated knock-in. First, the transgene is inserted in a random direction. Second, due to potential off-target effects of CRISPR/Cas9 system, the use of a donor template may give rise to nonspecific insertions of this template. Last, NHEJ can introduce random indels, possibly disrupting the target location. However, clever vector design and target site selection can minimize the number of non-specific insertions and insertions in the wrong orientation (Suzuki and Izpisua Belmonte, 2018, Sawatsubashi et al., 2018).
Besides the classical NHEJ and HDR pathways, another DNA repair pathway has emerged in the last two decades: microhomology-mediated end joining (MMEJ) (also known as alternative end joining (a-EJ)) (Frit et al., 2014). In MMEJ, microhomologies exist upstream and downstream of the DSB site on the two DNA strands. DSBs with microhomologies can result in annealing of the microhomologies and subsequent repair by MMEJ, which can result in short deletions (Kim et al., 2018b). This pathway has been used to integrate a gene of interest by using a precise integration into target chromosome (PITCh) system in cultured cells, zebrafish, silkworm, and frogs (Sakuma et al., 2016, Nakade et al., 2014, Hisano et al., 2015). With a reported knock-in efficiency that is 2.5 times higher than that of HR-assisted gene knock-in (Nakade et al., 2014), and with activity during all phases of the cell cycle (Taleei and Nikjoo, 2013, Truong et al., 2013), this pathway opens up the possibility of more efficiently targeted gene knock-in in different phases of the cell cycle. Further work will be necessary to demonstrate the applicability and the feasibility of MMEJ-mediated knock-ins for precision genome editing.
Base Editing in DNA
Incorrectly repaired DSBs and DSB at off-target sites are potentially pathogenic. Attempts to circumvent the generation of DSBs, and thereby to lower the risks associated with gene editing, have resulted in the development of the base-editing technique (Hess et al., 2017, Molla and Yang, 2019), which exploits the natural function of cytidine deaminases to convert cytidine to uridine in DNA. Eventually, the uridine is converted to thymidine by DNA duplication or via the DNA repair mechanisms base excision repair (BER) or mismatch repair (MMR) (Hess et al., 2017). Two different base-editing systems, BE (Komor et al., 2016) and target-AID (Nishida et al., 2016), have been developed by coupling cytidine deaminases to a catalytically deficient Cas9 protein (dCas9). Recently, a system with an incorporated adenosine deaminase was also developed. This enzyme hydrolyses adenosine into inosine, the base that pairs with cytidine. Inosine is thus replicated as guanine, thereby rendering an A to G change by DNA duplication (Gaudelli et al., 2017). These methods are clinically very interesting, as they open up opportunities for correcting monogenetic diseases in ways that reduce the risk of off-target effects. However, as most diseases are caused by various different disease-associated variants, this approach has to be tailored to each unique variant (Lessard et al., 2017). Another application of base editing is the introduction of a transcription-termination sequence to disrupt a gene in a highly specific manner (Kuscu et al., 2017, Billon et al., 2017).
Prime Editing
Recently, Anzalone et al. developed prime editing, a novel strategy for introducing deletions, inserting new genetic content or generating any of the 12 possible base-to-base conversions (Anzalone et al., 2019). For prime editing nicking Cas9 variant is fused to a reverse transcriptase, which, together with a prime editing extended guide RNA (pegRNA), makes gene editing possible. Prime editing offers a new range of possibilities in genome editing, with greater flexibility than the base editors, a greater reported efficiency than HDR, and no introduction of a DSB. Further research will determine the advantages and limitations of this promising novel concept.
Transient Modifications
All previous modifications result in permanent modifications at the genomic level. As an alternative approach, gene editing has been used to introduce transient modifications.
CRISPR interference (CRISPRi) was developed to inhibit gene transcription. Depending on the nature of the targeted locus, this approach is designed to interfere with transcription initiation or elongation (Bikard et al., 2013, Qi et al., 2013). The fusion of dCas9 to transcriptional activators or repressors has further sophisticated to the gene-editing toolkit for regulating transcription by CRISPR/Cas systems (Mahas et al., 2018).
The CRISPR/Cas system has also been engineered to modify epigenetic states by coupling dCas9 to several epigenetic modifiers such as P300 Core (Hilton et al., 2015), KRAB (Thakore et al., 2015), LSD1, Tet1, and Dnmt3 (Liao et al., 2017). Possible applications of this technique include the reversal of pathological epigenetic changes in conditions such as Fragile X syndrome, caused by silencing of the FMR1 gene, which is associated with hypermethylation of the CGG expansion in the region encoding the FMR1 5′ UTR. Recently, Liu and colleagues showed that by using dCas9-Tet1 the CGG expansion can be demethylated, leading to reactivation of FMR1 (Liu et al., 2018b). Future research is needed to test whether this is a valuable approach to treating fragile X syndrome and other epigenetic diseases.
Various studies over the years have shown that it is possible to target RNA with Cas13, an analogous member of the Cas family that is also referred to as C2c2 (O'Connell et al., 2014, East-Seletsky et al., 2016, Abudayyeh et al., 2016, Batra et al., 2017). One of these studies described the development of the RNA-editing platform known as RNA Editing for Programmable A to I Replacement (REPAIR) (Cox et al., 2017). For REPAIR a catalytically inactive Cas13 nuclease is coupled to a modified deaminase domain of adenosine deaminase that acts on RNA type 2 (ADAR2), which swaps A bases for I in RNA sequences. Additional modifications in the Cas13-ADAR2 fusion increased the targeted specificity of the REPAIR system almost a thousand-fold. In another study, protein engineering and characterization of different Cas13d orthologs generated a ribonuclease effector called CasRx (Konermann et al., 2018). The CasRx system has been shown to be highly effective in transcript knockdown or repression and in splice isoform manipulation.
Potentially, transcript editing could play a role in the therapy of diseases that cause transient changes in gene expression, such as local inflammation. As no definitive changes are made in the genome, but rather at transcriptional level, these approaches might represent a safer therapeutic strategy. However, the transient nature of these modifications will require repeated administration of the therapeutic agents.
Challenges in Bringing Genome Editing to the Clinic: Safety, Efficacy, and Delivery
Many conditions have to be met before genome-editing techniques can be considered for clinical development. Their efficiency and delivery must be great enough to attain a clinically significant result. Adverse events produced by permanent unintended variants should be minimized. Below, we highlight the targeting specificities and efficacies of natural and engineered variants and the broad diversity of strategies for delivering gene-editing tools.
Specificity
Mechanisms of Undesired On-Target and Off-Target Effects
The development of precise gene-editing tools to treat genetic disorders in the clinic requires careful consideration of the medical implications of permanent modifications in the genome. Nucleases should provide sufficient targeted specificity to prevent potentially detrimental effects derived from DSBs and the subsequent DNA repair mechanism. These effects can include undesired small insertions and deletions, point variants, aberrant chromosomal rearrangements, and large deletions in edited cells (Kosicki and Bradley, 2018, Hsu et al., 2013); they occur at or around the targeted location (“on-target”), as well as at more distant locations in the genome (“off-target”).
Influence of p53
p53 has been reported to cause a targeting selection bias in engineered cells: Cas9-gene targeting showed higher editing efficiency in cells with an altered p53 gene (Haapaniemi et al., 2018, Ihry et al., 2018), and transient p53 inhibition was shown to increase editing efficiency in human pluripotent stem cells, retinal pigment epithelial cells, and, more recently, in hematopoietic stem and progenitor cells (HSPCs) (Schiroli et al., 2019). However, due to the role of p53 in multiple DNA damage-response mechanisms, transient p53 inhibition may also leave cells more vulnerable to off-target mutagenesis. This idea has been challenged by others, who analyzed a large number of datasets derived from CRISPR screens (Brown et al., 2019, Ihry et al., 2019, Mair et al., 2019). These studies failed to show differences between p53-deficient and wild-type cells with respect to the enrichment of essential genes, thereby arguing against selection for clones that have a defective DNA damage-response pathway in CRISPR screens. Recently, guidelines were proposed for the performance of CRISPR screens with respect to monitoring and ensuring the quality of the screening performance (Brown et al., 2019).
R-Loop Formation
In addition to the risk of cleavage at an undesired location, the formation of R-loop-linked mutagenesis poses a risk. CRISPR/Cas cleavage is initiated by forming an R-loop, i.e., RNA-guided DNA unwinding to form an RNA-DNA hybrid with a displaced DNA strand inside the Cas protein (Jiang et al., 2016, Szczelkun et al., 2014). Although this R-loop is vital for stable binding and cleavage by the Cas protein, recent evidence in yeast indicates that the R-loop itself promotes mutagenesis on both on- and off-target sites (Laughery et al., 2019). This implies that undesirable mutagenesis may occur in many applications based on dCas9-fusions that were previously presumed to be safe, such as epigenome editing, transcriptional repression, and activation.
Effects on Transcription and Translation
Another risk is imposed by the introduction of indels via NHEJ, which can have unanticipated impacts on the regulation of RNA products and the translation of the protein it encodes. This includes promotion of internal ribosomal entry and alternative spliced mRNAs that lead to alternative products with a gain-of-function or a partially functional protein (Thomas et al., 2019, Mou et al., 2017, Tuladhar et al., 2019). mRNA decay can also trigger transcriptional adaptation. Via this genetic compensation mechanism, the expression of targeted or related genes can be upregulated independently or through protein feedback loops (El-Brolosy et al., 2019). Because the underlying mechanisms involved in splicing regulation and genetic compensation remain poorly understood, it remains a challenge to anticipate the translational and transcriptional changes induced by NHEJ-mediated indels.
Off-Target Effects of Base Editing
Because base editors do not produce DSBs in the genome, they do not pose risks of unintended damage triggered by DSBs. However, recent studies have reported the production of other types of off-target effects by adenine and cytosine base editors. In mouse embryos, variants using the APOBEC1 cytosine deaminase have been reported to generate multiple single-nucleotide variants (SNVs) at frequencies 20 times higher than the spontaneous mutation rate (Zuo et al., 2019). Other studies observed similar results, not only with the cytosine deaminases but also with the adenine editors based on the TadA deaminase, albeit at much lower frequencies (Kim et al., 2019). To reduce the rate of mutagenesis, Kim and colleagues coupled TadA with engineered Cas9 variants, successfully reducing their off-target activity (Kim et al., 2019). In addition, tens of thousands of SNVs were recently identified in RNA transcripts induced by both adenine and cytosine editors (Grunewald et al., 2019a, Zhou et al., 2019). These studies raised concerns about potentially detrimental variations not only in the genome but also in the transcriptome. Appropriate monitoring strategies will therefore be needed to evaluate the DNA and RNA of engineered cells that are created with base editors. To enable the safe use of base editors in the clinic, further research on novel variants or on recently engineered deaminases (Grunewald et al., 2019b) may reduce their off-target effects.
Monitoring Undesired On-Target and Off-Target Effects
Off-target mutations are more difficult to detect than on-target variants, as they can be present anywhere in the genome. The most common workflow applied to date is to design gRNA sequences with minimal predicted off-target effects and perform targeted sequencing to those sites with a high predicted value. Many tools are publicly available, the choice depending on the model organism of interest (Listgarten et al., 2018). A recent comprehensive study by Allen and colleagues investigated the outcome of more than 40,000 Cas9-edited DNA sequences using different cell types (Allen et al., 2018), showing how different cell types preferentially use specific repair mechanisms for certain DNA sequences. This research led to the development of a prediction tool (FORECasT) that could be used to predict off-target effects.
Common monitoring methods for detecting on-target mutations are based on polymerase chain reaction (PCR) (reviewed in Zischewski et al. [2017]). Each assay is designed according to the mutation introduced into the host genome, and the method selected must detect the difference in the heteroduplex DNA in order to determine the editing outcome. Mismatch cleavage assays, such as T7E1 or Surveyor, are commonly used to identify indels in bulk and single-cell preparations. Although these methods are simple and effective in detecting small indels, they underrate the detection of large deletions or insertions. Additional methods that may provide detailed information about introduced mutations are based on Sanger sequencing.
Considering our limited knowledge of the mechanisms involved in CRISPR/Cas off-target activity, unbiased methods should be included that do not rely on predictions. Although whole-genome sequencing offers an unbiased high-throughput assessment of unintended variants, it is expensive and only provides information on bulk genomes. When applied to cell clones that have been edited ex vivo, whole-genome sequencing provides a valuable option. However, when applied to cell populations without clonal expansion, whole-genome sequencing is less suitable, as it may fail to detect low-abundant events that might eventually lead to oncogenic transformation. Other methods for detecting off-target effects are based on the fact that nucleases generate breaks that are repaired by endogenous DNA repair mechanisms. These have been exploited to develop techniques for quantifying the DSBs that have occurred in vitro (BLESS, GUIDE-seq, Digenome-seq) (Zischewski et al., 2017).
The selection of the most suitable detection method will depend on the sensitivity, throughput, limitations, and cost of the technique, and ultimately on the editing strategy (ex vivo or in vivo) and delivery system used (viral or non-viral). A recent study by Akcakaya and colleagues describes a robust evaluation of off-targets using in vitro assessment of potential off-targets via CIRCLE-seq, followed by in vivo targeted deep sequencing of engineered organs (VIVO) (Akcakaya et al., 2018). In a more recent study, Wienert and colleagues developed an unbiased monitoring strategy (DISCOVER-Seq) to evaluate potential off-target DSBs in cells and tissues (Wienert et al., 2019). Because these approaches are based on pre-repair mechanisms, additional detection methods will be required to evaluate their editing outcome and cytotoxic effect. In human diseases, robust standardized assessment guidelines will therefore be essential for precise genome engineering.
Increasing Specificity
Research has made great advances in the characterization of novel CRISPR/Cas9 orthologs and homologs from various species and in the generation of engineered enhanced-specificity Cas enzymes and sgRNAs. The most commonly used CRISPR/Cas system in research exploits the Cas9 protein derived from streptococcus pyogenes. However, many other orthologs offer different targeting abilities and specificities (Karvelis et al., 2017). Other natural homologs discovered recently include Cas12a (Cpf1) and Cas12b (C2c1), which can also be engineered for enhanced DNA specificity (Wu et al., 2018).
Recently, many engineered forms of Cas9 proteins with improved and broad targeting specificities have been developed. For example, Cas9 nickases (Ran et al., 2013) or Fok I fused to dead Cas9 nucleases (Tsai et al., 2014) use two different sgRNAs to perform ssDNA breaks, which can significantly reduce the off-target effects of WT Cas9 variants. Cas9 nickases have been used to generate paired single-strand DNA breaks in donor plasmids and at genomic target sites, thereby increasing gene-targeting efficiency, specificity, and fidelity in human cells when compared with nicking targeting DNA alone (Chen et al., 2017b).
More recently, new engineered Cas9 proteins with enhanced targeted specificity have raised the possibility of using smart-designed nucleases for precise genome editing. The high-fidelity Cas9 (Cas9-HF1) (Kleinstiver et al., 2016) was designed by altering the composition of four residues involved in non-specific interactions of Cas9 with its target DNA. These modifications reduced the generation of mismatches with minimal loss of on-target efficiency. Using GUIDE-seq to monitor the frequency of off-target effects showed that editing with Cas9-HF1 resulted in only one single mismatch compared with the 65 detected for WT Cas9 using eight different sgRNAs toward four human genes in U2OS cells. The enhanced specificity Cas9 (eSpCas9) (Slaymaker et al., 2016) was created by modifying positively charged residues involved in the unwinding of the non-complementary DNA strand. Weakening this interaction reduced Cas9 off-target activity 10 times compared with that of WT Cas9, while maintaining on-target efficiency in human embryonic kidney cells. The evolved Cas9 (evoCas9) was generated through directed evolution in yeast (Casini et al., 2018), which involved the screening of random variants to identify beneficial variants in the REC3 domain involved in the recognition of the sgRNA and DNA heteroduplex. The combination of four beneficial variants generated the evoCas9 nuclease: a high-fidelity Cas9 variant with minimal loss of on-target activity and a reported 98.7% reduction in off-target activity relative to that in WT Cas9 when analyzed via GUIDE-seq (Casini et al., 2018).
The lack of mechanistic insight regarding target recognition of Cas9-HF1 and eSpCas9 led to the development of the hyper-accurate Cas9 (HypaCas9) (Chen et al., 2017a). It was shown that raising the energy requirements for the conformational activation of the HNH domain, which acts as a Cas9 editing checkpoint, reduced off-target activity in these variants. HypaCas9 was designed by modifying four amino acids involved in this process. Genome-wide specificity of this variant was significantly better than that of the WT Cas9 and equivalent to that of the Cas9-HF1 and eSpCas9 nucleases. On-target activity was at least >70% that of WT Cas9 when tested in U2OS cells.
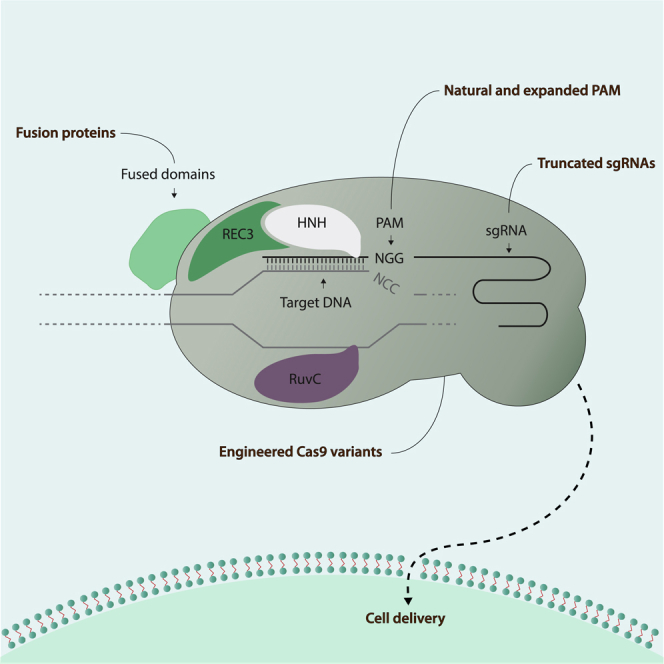
Other Cas engineering strategies focused on increasing the targeting capacities of CRISPR/Cas nucleases by altering their PAM recognition sites without losing on-target specificity (Figure 1) (Kleinstiver et al., 2015, Hu et al., 2018). Directed evolution was used to create different variants of Cas9 nucleases with altered PAM targeting properties; the VQR and VRER variants demonstrated enhanced specificity in human cells. Other variants include xCas9, which recognizes a broader range of PAM sequences than natural Cas9. Recently, Nishimasu and colleagues designed a SpCas9-NG nuclease with a targeting capacity similar to that of the xCas9 variant. However, comparative studies showed that the editing efficiency of the SpCas9-NG variant was greater than that of xCas9. To convert C-to-T bases at NG PAM sites in human cells, the authors further combined SpCas9-NG with a cytidine deaminase, thereby providing new engineered Cas9 variants with many capacities beyond improved specificity (Nishimasu et al., 2018). Other homologs, such as Cas12a, have also been engineered to act on different PAM sequences instead of acting on their natural binding sites (Gao et al., 2017).
Figure 1.

SpCas9 Ribonucleoprotein Variants with Altered Targeting Capacities
Engineered SpCas9 nuclease variants with potential therapeutic advantages. Variants with expanded PAM recognition sequences include natural Cas9 and its well-described homologs Cas12a and Cas12b. Designed engineered nucleases with improved specificity include the split-Cas9 variant, which was created to improve the packaging of Cas9 proteins in AAV vehicles. Cas9 nucleases have also been fused with multiple functional domains to allow for targeted epigenetic modifications, single nucleotide modifications, single-strand nicking activity, or temporal regulation of CRISPR/Cas activity. Truncated versions of sgRNAs have been successfully used to increase targeting specificity. SpCas9, streptococcus pyogenes Cas9.
In addition, sgRNAs have been altered to enhance their targeting specificities. By modifying their length, secondary structure, or chemical composition, several studies demonstrated that off-target mutagenesis can be reduced (Fu et al., 2014, Hendel et al., 2015, Kocak et al., 2019). However, these changes may also influence their on-target efficiency. Subsequent studies found that partial replacement of RNA nucleotides with DNA could further reduce the off-target activity while retaining on-target efficiency (Yin et al., 2018). The optimal targeting specificity of sgRNAs will ultimately depend on the in silico design. Many software programs are publicly available for this purpose. However, it is important to consider the genetic variations between different individuals, as it can substantially alter the on-target and off-targeting activities of precise genome engineering (Lessard et al., 2017).
Spatiotemporal Control of CRISPR/Cas Activity
The capacity of nucleases to generate permanent genome changes has prompted researchers to seek novel methods of exerting spatiotemporal control over CRISPR/Cas activity. Such control is important: when targeting DNA to reduce undesirable off-target effects, any intervention in a desired tissue or organ should be brief.
The spatial localization of precision gene-editing tools can be manipulated by using different delivery vehicles. For instance, different serotypes of viral particles such as lentivirus or adeno-associated virus (AAV) have higher affinities for specific cell types. By using tissue-specific promoters, the expression of CRISPR/Cas components can be limited to target tissues or organs. Other organs can be targeted through local delivery with limited distribution into other tissues, such as the eye. However, for efficient targeting, diseases involving larger organs or tissues (such as muscle or skin) might require systemic administration.
Off-target effects are also magnified by extended gene-editing activity. Several approaches have therefore been developed to influence the spatiotemporal activity of CRISPR/Cas systems. Many natural Cas9 enzymes have been engineered into switchable Cas9 nucleases, either to be light sensitive, to be controlled by allosteric regulation, or to express specific localization signals (Richter et al., 2017). However, the clinical potential of these engineered forms can be limited, as some switchable nucleases respond to toxic regulators such as doxycyclin or UV light. Nevertheless, the characterization of novel natural inhibitors for Cas9 and Cas12a variants holds great promise for the therapeutic regulation of CRISPR/Cas activity in a clinical setting (Pawluk et al., 2016, Watters et al., 2018, Jiang et al., 2019, Uribe et al., 2019). Another approach that holds great potential for controlling the CRISPR/Cas system and off-target activity is the use of anti-CRISPRs. Discovered in phages to overcome CRISPR immunity, these anti-CRISPRs can be used, repurposed, and structurally engineered to control the CRISPR/Cas9 system (Pawluk et al., 2018).
Immunogenicity and Safety
Gene targeting must be performed with minimal toxicity and immunogenicity. The immunogenic properties of gene-editing components have recently been described in several studies (Charlesworth et al., 2019, Kim et al., 2018a, Wagner et al., 2019, Chew et al., 2016). CRISPR/Cas systems are protein complexes derived from bacteria and archaea, some of which, such as S. aureus (SaCas9) or S. pyogenes (SpCas9) are common infectious agents in the human population. The risks associated with immune responses against Cas9 proteins (or other Cas homologs) and sgRNAs should be considered in clinical trials. A recent study by Charlesworth and colleagues on the presence of preexisting antibodies reported that 78% of donors involved in the study presented antibodies against SaCas9 and 58% presented antibodies against SpCas9. In addition, analysis of blood samples indicated that 78% had anti-SaCas9 T cells and 67% had anti-SpCas9 T cells (Charlesworth et al., 2019). The detection of anti-Cas9 T cells might pose a serious challenge to precision genomic engineering, as exposure to a cytotoxic T cell response might eliminate engineered cells. Because the assays used in this study were not intended to investigate cellular immune responses against CRISPR/Cas components, this issue should be addressed in future studies.
Gene-editing tools used in vivo will be exposed to the host immune system. Potential immune reactions will be influenced by the type of delivery vehicle used. For instance, viral-based vectors could lead to long-term expression of gene-editing tools. These could lead to sustained nuclease activity, which, depending on the variant used, might in turn lead to extensive damage to the human genome and a prolonged immune response. In addition, immune responses to viral-based vectors such as AAV are known for their capacity to neutralize the therapy, including an induction of a cytotoxic T cell response that eliminates cells that have been targeted by the AAV vector (Mingozzi and High, 2011, Mingozzi and High, 2013). This is highly relevant, as AAV is a naturally occurring virus against which ∼40% or an even higher percentage of people already carry antibodies to AAV capsid proteins.
To minimize the effects of potential immune reactions, researchers can follow various strategies. In one approach, gene-editing activity can be spatiotemporally controlled, which has great potential for clinical use (see above for the section on Spatiotemporal control of CRISPR/Cas activity). Gene-editing components can for instance be directed to immune-privileged organs such as the eye or to tolerogenic organs such as the liver. Strategies involving immune modulation might prevent side effects derived from bacterial Cas proteins or from antigens derived from the delivery vehicles. An appropriate evaluation of candidate approaches and potential detrimental effects will be essential for the safe use of precision genome engineering in the clinic.
Delivery Strategies
The development of efficient and safe delivery systems is one of the most challenging aspects of introducing precise genome editing to the clinic. The delivery approach to be used will be influenced by the type of gene-editing material. So far, CRISPR/Cas systems have been successfully administered in naked or encapsulated plasmid DNA, mRNA, or functional (ribonucleo)protein complexes both in vivo and ex vivo (Figure 2).
Figure 2.

Delivery Strategies Used in Preclinical Studies
CRISPR/Cas gene-editing tools have been delivered as ribonucleoprotein complexes, plasmid DNA, or RNA. They can be delivered as naked components using chemical or physical methods or in delivery vehicles including virus and/or different types of nanocomplexes. There are two main strategies for delivering precision gene-editing platforms. In multiple organs and tissues in animal models, in vivo delivery strategies have been successfully used, some of which have been clinically approved for application in humans. Ex vivo delivery strategies are being extensively used in T cell engineering, hematopoietic stem-cell gene editing, and iPSCs modeling. iTOP, induced transduction by osmocytosis and propanebetaine; CPP, cell-penetrating peptide; LNP, lipid nanoparticle; GNP, gold nanoparticle, HSPCs, hematopoietic stem and progenitor cells; iPSCs, induced pluripotent stem cells.
In Vivo Delivery
In many clinical and pre-clinical studies, adeno-associated viruses are commonly used delivery vehicles in vivo. Different AAV serotypes provide increased delivery efficiencies for specific cell types, thereby allowing tissue/organ targeting (Colella et al., 2018). In vivo delivery of gene-editing machinery using AAV has been successfully used in multiple animal models of metabolic diseases (Pankowicz et al., 2016, Yin et al., 2016, Villiger et al., 2018, Rossidis et al., 2018), human immunodeficiency virus (HIV) infection (Yin et al., 2017a), muscle dystrophies (Amoasii et al., 2018), brain disease (Nishiyama et al., 2017), retinal disorders (Suzuki et al., 2016, Huang et al., 2017, Maeder et al., 2019), degenerative disorders (Beyret et al., 2019, Santiago-Fernandez et al., 2019), and diabetes and kidney malignancies (Liao et al., 2017). More recently, it has been used to upregulate the expression of endogenous genes in haploinsufficiencies such as obesity (Matharu et al., 2018).
Due to its high efficiency, viral delivery through AAVs offers promising results for precision genome-editing medicine. However, AAVs elicit immune responses that may limit the therapeutic potential of genome-engineering tools. If the gene-editing strategy should be administered repeatedly over time, this is especially relevant, as patients will develop antibodies against the AAV virus after the first administration, precluding any subsequent treatment using AAV as delivery vehicle. Furthermore, as a significant proportion of the population has pre-existing antibodies against the AAV virus, it is not eligible for AAV-based treatment (Louis Jeune et al., 2013). However, some of these limitations may be overcome by combined immunosuppressive therapies. AAV delivery can also drive long-term transgene expression and might result in low incidence of transgene integration. This promotes long-lasting gene editing in tissues and thus poses a high risk of off-target events. Methods for inactivating Cas activity would be required to prevent the long-lasting introduction of double-strand breaks in vivo and to reduce the risks of chromosomal abnormalities.
The large size of Cas proteins hampers their packaging into AAV plasmids. To circumvent this, researchers have successfully treated eye diseases using smaller Cas9 orthologs derived from campylobacter jejuni (CjCas9) (Kim et al., 2017). The study in question described how researchers were able to package the DNA sequence of the CjCas9, sgRNAs, and a donor template in a single AAV vector. Others have designed a flexible AAV-split-Cas9 platform to compact the Cas9 DNA sequence into AAV vectors (Chew et al., 2016). By manipulating the WT protein sequence of the Cas9 nuclease into fused functional domains, it was possible to shorten the newly engineered Cas9 DNA sequence by more than 2 kb.
Over recent years, different types of nanomaterial have been developed with encouraging results in multiple diseases (reviewed in Li et al. [2018]). Unlike AAVs, lipid-based nanoparticles are able to transfer Cas9 plasmids or proteins without the risk of genomic integration; some of them have received FDA approval for therapeutic use (Glass et al., 2018). In mice, lipid nanoparticles are currently used to deliver Cas9 components locally to the brain (Wang et al., 2016) or inner ear (Gao et al., 2018) and systemically to the liver (Yin et al., 2017b, Finn et al., 2018). However, for the delivery of Cas9 and a donor template for HDR, both components must be encapsulated in distinct lipid nanoparticles, which might affect its editing efficiency in vivo. More recently, newly developed gold nanoparticles have been successfully used in rodent models to simultaneously deliver Cas9 ribonucleoproteins and donor templates to treat Duchenne muscular dystrophy (Lee et al., 2017) and fragile X syndrome (Lee et al., 2018). Park and colleagues have shown successful genome editing of post-mitotic neurons in different Alzheimer disease mouse models using amphiphilic nanocomplexes generated using the R7L10 peptide together with the Cas9 nuclease and the sgRNAs (Park et al., 2019a).
Ex Vivo Delivery
Many vehicles are being exploited to deliver precision gene editing ex vivo. Depending on the type of cell or stem cell used for ex vivo editing, the gene-editing machinery may be delivered via electroporation, microinjection, chemical methods such as cell-penetrating peptides and nanoparticles (Wu et al., 2018), and virus-based vehicles such as AAVs or lentiviruses (Figure 2). The efficacy with which various types of immune cells can be engineered ex vivo has been investigated in multiple preclinical models, greatly stimulating research in several fields, particularly hematology and cancer therapeutics (Bak et al., 2018, Huang et al., 2018). Pluripotent stem cells such as induced pluripotent stem cells (iPSCs) have also been used extensively for genome engineering ex vivo for cell-based regenerative approaches and for disease modeling (Jang and Ye, 2016). iPSCs offer great potential for disease modeling, as they can be differentiated into any cell type that is relevant for the disease, such as cardiomyocytes (Brandao et al., 2017, Devalla and Passier, 2018), skeletal muscle cells (van der Wal et al., 2018, Magli and Perlingeiro, 2017, Chal and Pourquie, 2017), neuronal cells (Bordoni et al., 2018, Compagnucci et al., 2014), hepatocytes (Fiorotto et al., 2019, Hannoun et al., 2016), and many other cell types. Isogenic controls that correct for genetic background effects can be generated in iPSCs using CRISPR/Cas; this correction for background effects is considered important due to the large variation in many parameters among individuals. The risk of acquiring variants during the reprogramming of somatic cells into iPSCs has imposed the need for caution in the use of iPSC-derived cells for cell-based therapy. In addition, the development of affordable clinical treatment is inhibited by the cost associated with the quality control of patient-specific iPSCs. As a possible solution, the generation of iPS-cell banks covering the majority of HLA isotypes known globally is currently in progress. This would make a validated iPSC line available for cell-based therapy for almost each individual patient without the need to generate patient-specific iPSCs (Ben Jehuda et al., 2018, Mandai et al., 2017).
Preclinical Studies and Clinical Trials
Therapeutic gene editing by ZFN, TALEN, or CRISPR/Cas9 platforms is already being explored in a number of clinical trials registered at clinicaltrials.gov. As we discuss briefly below, these target cancer, genetic disorders, and HIV/AIDS (Figure 3).
Figure 3.

Examples of Gene-Editing Strategies in Current Clinical Trials
(A) In cancer, gene-editing targets PD-1 or CD7 in T cells to enhance immune responses.
(B) In patients with HIV, CCR5 is targeted in HSPCs to prevent HIV entry.
(C) In β-thalassemia and sickle cell disease, induction of fetal hemoglobin or correction of adult hemoglobin in HSPCs is used.
(D) AAV-mediated gene editing in the liver provides circulating enzymes in lysosomal storage diseases.
Cancer
In HPV-related cervical cancer, CRISPR/CAS9, ZFNs, or TALENs are applied topically to precancerous lesions in order to disrupt viral oncogenes E6 or E7 in vivo (Ding et al., 2014, Hu et al., 2014, Hu et al., 2015, Kennedy et al., 2014, Shankar et al., 2017, Zhen et al., 2014). In addition to induction of (viral) oncogene knock-out, gene editing for the treatment of various types of cancer is also being investigated in cellular immunotherapies or adoptive cell therapy (ACT). To overcome evasion of immune surveillance by cancer cells, PD-1, an immune checkpoint molecule, was knocked out ex vivo in autologous T cells using CRISPR/Cas9 before reinfusion into patients (Beane et al., 2015, Menger et al., 2016, Su et al., 2016, Zhao et al., 2018). PD-1 knock-out have been deployed to improve redirected T cells (Guo et al., 2018b, Hu et al., 2019a, Hu et al., 2019b, Lu et al., 2019, Ouchi et al., 2018, Rupp et al., 2017, Su et al., 2017), which were genetically altered to express a transgenic T cell receptor (tTCR) or a chimeric antigen receptor (CAR) targeted at a disease-associated antigen. To enable ACT for T cell malignancies expressing CD7, CRISPR/Cas9 has also been used ex vivo to knock out the CD7 gene in anti-CD7 CAR-T cells, preventing these CAR-T cells from killing each other (Cooper et al., 2018, Gomes-Silva et al., 2017). Finally, to circumvent the necessity for custom-made autologous therapy for each patient, ex vivo gene editing by CRISPR/CAS9 or TALENs in CAR-T cells manufactured from donor-derived T cells is being applied to generate universal CAR-T cells (Torikai and Cooper, 2016, Yang et al., 2015). This involves the disruption of the endogenous TCR to prevent graft-versus-host disease and of HLA components to prevent graft rejection by the host immune system.
HIV Infection/AIDS
In patients infected with HIV, T cells or HSPCs are harvested and the CCR5 gene is knocked out in vitro by CRISPR/Cas9 or ZFNs before reinfusion into the patient (Holt et al., 2010, Perez et al., 2008, Tebas et al., 2014, Wang et al., 2014, Xu et al., 2017, Yu et al., 2018). As CCR5 is essential for HIV invasion of T cells, this intervention should prevent spreading of the virus into the engineered cells (Brelot and Chakrabarti, 2018).
Genetic Disorders
Sickle cell disease (SCD) and β-thalassemia are caused by disease-associated variants in the HBB gene, which encodes the beta subunit of hemoglobin. One strategy involves inducing the production of fetal hemoglobin (HbF), which can compensate for malformed adult hemoglobin (in SCD) or reduced levels of adult hemoglobin (in β-thalassemia). This can be accomplished by ex vivo disruption of the intronic erythroid-specific enhancer of BCL11A in HSPCs by CRISPR/Cas9 or ZFN (Chang et al., 2017b, Psatha et al., 2018). After reinfusion, these HSPCs will produce erythrocytes with reduced BCL11A levels. As BCL11A represses transcription from HBG (Liu et al., 2018a), which encodes the gamma subunit of HbF, the level of the gamma subunit and thus HbF will increase. In another approach, one clinical trial is currently investigating direct gene restoration by CRISPR/Cas9 ex vivo in iPSC-derived HSPCs for the treatment of β-thalassemia (Cai et al., 2018, Martin et al., 2019, Park et al., 2019b, Wattanapanitch et al., 2018).
Clinical trials have started for the monogenic disorders hemophilia B and mucopolysaccharidosis (MPS) types I and II. Hemophilia B is caused by a deficiency of clotting factor IX, whereas MPS I and II are lysosomal storage disorders caused by deficiency of enzymes involved in the lysosomal degradation of glycosaminoglycans. These trials use systemic injection of AAV-expressing ZFN to achieve the targeted integration of a functional copy of the deficient gene into the albumin locus in liver cells, which secrete the enzyme into the circulation (Laoharawee et al., 2018, Sharma et al., 2015).
Finally, gene editing is being investigated in one of the subtypes of Leber's congenital amaurosis (LCA), a congenital retinal dystrophy. The subtype LCA10 is most commonly caused by an intronic variant in the CEP290 gene that generates a cryptic splice site, resulting in defective protein production (Xu et al., 2018). A CRISPR/Cas9-based gene-editing strategy has been developed that removes the cryptic splice site; it is administered in vivo via subretinal injection using AAV as delivery vehicle (Maeder et al., 2019, Ruan et al., 2017).
Future Challenges
Among the most challenging aspects of gene editing to date are its delivery, specificity, and efficacy. The delivery strategy that is most appropriate for gene-editing intervention will depend on the cell-targeting approach that is most suitable for treating a specific disease. Ex vivo strategies can be conducted in a controlled environment, are amenable to efficient engineering using multiple methods, and may be a more straightforward way of bringing precision gene-editing medicine to the clinic. For example, when applied to stem cells (including adult stem cells or iPSCs) ex vivo gene editing can be subjected to quality control for genotoxicity before it is decided to engraft the cells in question. The use of ex vivo protein transduction ensures a transient exposure to nucleases, as opposed to the long-term exposure with in vivo approaches that use DNA constructs. However, ex vivo strategies face other issues associated with ex vivo cell manipulation. Engineered cells have to engraft efficiently into host individuals and should evade immune rejections if they are derived from a different donor. On the other hand, in vivo strategies will face the challenge of uncontrolled off-target events. In such cases, the selection of a suitable delivery vehicle, of a highly specific gene-targeting tool, and of possibly a strategy for spatiotemporal control, will all be especially relevant to preventing unintended variants and reducing potential immune reactions. In vivo gene-editing strategies that use persistent CAS9 expression—such as AAV—as a delivery vehicle may pose a risk of long-term exposure to gene-editing events, increasing the risk for off-target effects. Despite these challenges, the ongoing iteration of the CRISPR/Cas system into versions with improved specificity and efficacy holds great promise for a wide range of clinical applications. It will be important to remain cautious and, when the time comes for clinical testing, to evaluate the advantages against the possible risks. To fully understand the long-term effects of potential new treatments involving precise genomic engineering, thorough preclinical work is required.
Acknowledgments
This work has received funding from Texnet, Zeldzame Ziekten Fonds/WE Foundation, Metakids, and Stofwisselkracht. The collaboration project is co-funded by the PPP Allowance made available by Health ∼ Holland, Top Sector Life Sciences & Health, to stimulate public-private partnerships (project numbers LSHM16008 and LSHM19015). The collaboration project is co-initiated by the Prinses Beatrix Spierfonds.
Author Contributions
M.B., P.H.H., M.E., and W.W.M.P.P. wrote and revised the manuscript. M.B., P.H.H., and M.E. performed the graphical abstract and figures. A.T.v.d.P contributed to the review and editing of the manuscript. A.T.v.d.P and W.W.M.P.P. secured funding.
Declaration of Interests
A.T.v.d.P. has provided consulting services for various industries in the field of Pompe disease under an agreement between these industries and Erasmus MC, Rotterdam, The Netherlands.
References
- Abudayyeh O.O., Gootenberg J.S., Konermann S., Joung J., Slaymaker I.M., Cox D.B., Shmakov S., Makarova K.S., Semenova E., Minakhin L. C2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector. Science. 2016;353:aaf5573. doi: 10.1126/science.aaf5573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akcakaya P., Bobbin M.L., Guo J.A., Lopez J.M., Clement M.K., Garcia S.P., Fellows M.D., Porritt M.J., Firth M.A., Carreras A. In vivo CRISPR-Cas gene editing with no detectable genome-wide off-target mutations. Nature. 2018;561:416–419. doi: 10.1038/s41586-018-0500-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allen F., Crepaldi L., Alsinet C., Strong A.J., Kleshchevnikov V., De Angeli P., Palenikova P., Kosicki M., Bassett A.R., Harding H. Mutations generated by repair of Cas9-induced double strand breaks are predictable from surrounding sequence. Nat. Biotechnol. 2018;37:64–72. doi: 10.1038/nbt.4317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amoasii L., Hildyard J.C.W., Li H., Sanchez-Ortiz E., Mireault A., Caballero D., Harron R., Stathopoulou T.R., Massey C., Shelton J.M. Gene editing restores dystrophin expression in a canine model of Duchenne muscular dystrophy. Science. 2018;362:86–91. doi: 10.1126/science.aau1549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anzalone A.V., Randolph P.B., Davis J.R., Sousa A.A., Koblan L.W., Levy J.M., Chen P.J., Wilson C., Newby G.A., Raguram A., Liu D.R. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature. 2019;576:149–157. doi: 10.1038/s41586-019-1711-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bak R.O., Dever D.P., Porteus M.H. CRISPR/Cas9 genome editing in human hematopoietic stem cells. Nat. Protoc. 2018;13:358–376. doi: 10.1038/nprot.2017.143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baliou S., Adamaki M., Kyriakopoulos A.M., Spandidos D.A., Panayiotidis M., Christodoulou I., Zoumpourlis V. Role of the CRISPR system in controlling gene transcription and monitoring cell fate (Review) Mol. Med. Rep. 2018;17:1421–1427. doi: 10.3892/mmr.2017.8099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barrangou R., Doudna J.A. Applications of CRISPR technologies in research and beyond. Nat. Biotechnol. 2016;34:933–941. doi: 10.1038/nbt.3659. [DOI] [PubMed] [Google Scholar]
- Batra R., Nelles D.A., Pirie E., Blue S.M., Marina R.J., Wang H., Chaim I.A., Thomas J.D., Zhang N., Nguyen V. Elimination of toxic microsatellite repeat expansion RNA by RNA-targeting Cas9. Cell. 2017;170:899–912.e10. doi: 10.1016/j.cell.2017.07.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beane J.D., Lee G., Zheng Z., Mendel M., Abate-Daga D., Bharathan M., Black M., Gandhi N., Yu Z., Chandran S. Clinical scale zinc finger nuclease-mediated gene editing of PD-1 in tumor infiltrating lymphocytes for the treatment of metastatic melanoma. Mol. Ther. 2015;23:1380–1390. doi: 10.1038/mt.2015.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ben Jehuda R., Shemer Y., Binah O. Genome editing in induced pluripotent stem cells using CRISPR/Cas9. Stem Cell Rev. Rep. 2018;14:323–336. doi: 10.1007/s12015-018-9811-3. [DOI] [PubMed] [Google Scholar]
- Bergsma A.J., van der Wal E., Broeders M., van der Ploeg A.T., Pim Pijnappel W.W.M. Alternative splicing in genetic diseases: improved diagnosis and novel treatment options. Int. Rev. Cell Mol. Biol. 2018;335:85–141. doi: 10.1016/bs.ircmb.2017.07.008. [DOI] [PubMed] [Google Scholar]
- Beyret E., Liao H.K., Yamamoto M., Hernandez-Benitez R., Fu Y., Erikson G., Reddy P., Izpisua Belmonte J.C. Single-dose CRISPR-Cas9 therapy extends lifespan of mice with Hutchinson-Gilford progeria syndrome. Nat. Med. 2019;25:419–422. doi: 10.1038/s41591-019-0343-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bikard D., Jiang W., Samai P., Hochschild A., Zhang F., Marraffini L.A. Programmable repression and activation of bacterial gene expression using an engineered CRISPR-Cas system. Nucleic Acids Res. 2013;41:7429–7437. doi: 10.1093/nar/gkt520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Billon P., Bryant E.E., Joseph S.A., Nambiar T.S., Hayward S.B., Rothstein R., Ciccia A. CRISPR-Mediated base editing enables efficient disruption of Eukaryotic genes through induction of STOP codons. Mol. Cell. 2017;67:1068–1079.e4. doi: 10.1016/j.molcel.2017.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bordoni M., Rey F., Fantini V., Pansarasa O., Di Giulio A.M., Carelli S., Cereda C. From neuronal differentiation of iPSCs to 3D neuro-organoids: modelling and therapy of neurodegenerative diseases. Int. J. Mol. Sci. 2018;19:3972. doi: 10.3390/ijms19123972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brandao K.O., Tabel V.A., Atsma D.E., Mummery C.L., Davis R.P. Human pluripotent stem cell models of cardiac disease: from mechanisms to therapies. Dis. Model. Mech. 2017;10:1039–1059. doi: 10.1242/dmm.030320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brelot A., Chakrabarti L.A. CCR5 revisited: how mechanisms of HIV entry govern AIDS pathogenesis. J. Mol. Biol. 2018;430:2557–2589. doi: 10.1016/j.jmb.2018.06.027. [DOI] [PubMed] [Google Scholar]
- Brown K.R., Mair B., Soste M., Moffat J. CRISPR screens are feasible in TP53 wild-type cells. Mol. Syst. Biol. 2019;15:e8679. doi: 10.15252/msb.20188679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cai L., Bai H., Mahairaki V., Gao Y., He C., Wen Y., Jin Y.C., Wang Y., Pan R.L., Qasba A. A universal approach to correct various HBB gene mutations in human stem cells for gene therapy of beta-thalassemia and sickle cell disease. Stem Cells Transl. Med. 2018;7:87–97. doi: 10.1002/sctm.17-0066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casini A., Olivieri M., Petris G., Montagna C., Reginato G., Maule G., Lorenzin F., Prandi D., Romanel A., Demichelis F. A highly specific SpCas9 variant is identified by in vivo screening in yeast. Nat. Biotechnol. 2018;36:265–271. doi: 10.1038/nbt.4066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chal J., Pourquie O. Making muscle: skeletal myogenesis in vivo and in vitro. Development. 2017;144:2104–2122. doi: 10.1242/dev.151035. [DOI] [PubMed] [Google Scholar]
- Chang H.H.Y., Pannunzio N.R., Adachi N., Lieber M.R. Non-homologous DNA end joining and alternative pathways to double-strand break repair. Nat. Rev. Mol. Cell Biol. 2017;18:495–506. doi: 10.1038/nrm.2017.48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang K.H., Smith S.E., Sullivan T., Chen K., Zhou Q., West J.A., Liu M., Liu Y., Vieira B.F., Sun C. Long-Term engraftment and fetal globin induction upon BCL11A gene editing in bone-marrow-derived CD34(+) hematopoietic stem and progenitor cells. Mol. Ther. Methods Clin. Dev. 2017;4:137–148. doi: 10.1016/j.omtm.2016.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charlesworth C.T., Deshpande P.S., Dever D.P., Camarena J., Lemgart V.T., Cromer M.K., Vakulskas C.A., Collingwood M.A., Zhang L., Bode N.M. Identification of preexisting adaptive immunity to Cas9 proteins in humans. Nat. Med. 2019;25:249–254. doi: 10.1038/s41591-018-0326-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen J.S., Dagdas Y.S., Kleinstiver B.P., Welch M.M., Sousa A.A., Harrington L.B., Sternberg S.H., Joung J.K., Yildiz A., Doudna J.A. Enhanced proofreading governs CRISPR-Cas9 targeting accuracy. Nature. 2017;550:407–410. doi: 10.1038/nature24268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen X., Janssen J.M., Liu J., Maggio I., T'Jong A.E.J., Mikkers H.M.M., Gonçalves M.A.F.V. In trans paired nicking triggers seamless genome editing without double-stranded DNA cutting. Nat. Commun. 2017;8:657. doi: 10.1038/s41467-017-00687-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chew W.L., Tabebordbar M., Cheng J.K.W., Mali P., Wu E.Y., Ng A.H.M., Zhu K., Wagers A.J., Church G.M. A multifunctional AAV-CRISPR-Cas9 and its host response. Nat. Methods. 2016;13:868–874. doi: 10.1038/nmeth.3993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chu V.T., Weber T., Wefers B., Wurst W., Sander S., Rajewsky K., Kuhn R. Increasing the efficiency of homology-directed repair for CRISPR-Cas9-induced precise gene editing in mammalian cells. Nat. Biotechnol. 2015;33:543–548. doi: 10.1038/nbt.3198. [DOI] [PubMed] [Google Scholar]
- Colella P., Ronzitti G., Mingozzi F. Emerging issues in AAV-mediated in vivo gene therapy. Mol. Ther. Methods Clin. Dev. 2018;8:87–104. doi: 10.1016/j.omtm.2017.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Compagnucci C., Nizzardo M., Corti S., Zanni G., Bertini E. In vitro neurogenesis: development and functional implications of iPSC technology. Cell Mol. Life Sci. 2014;71:1623–1639. doi: 10.1007/s00018-013-1511-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooper M.L., Choi J., Staser K., Ritchey J.K., Devenport J.M., Eckardt K., Rettig M.P., Wang B., Eissenberg L.G., Ghobadi A. An "off-the-shelf" fratricide-resistant CAR-T for the treatment of T cell hematologic malignancies. Leukemia. 2018;32:1970–1983. doi: 10.1038/s41375-018-0065-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cox D.B.T., Gootenberg J.S., Abudayyeh O.O., Franklin B., Kellner M.J., Joung J., Zhang F. RNA editing with CRISPR-Cas13. Science. 2017;358:1019–1027. doi: 10.1126/science.aaq0180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devalla H.D., Passier R. Cardiac differentiation of pluripotent stem cells and implications for modeling the heart in health and disease. Sci. Transl. Med. 2018;10:eaah5457. doi: 10.1126/scitranslmed.aah5457. [DOI] [PubMed] [Google Scholar]
- Ding W., Hu Z., Zhu D., Jiang X., Yu L., Wang X., Zhang C., Wang L., Ji T., Li K. Zinc finger nucleases targeting the human papillomavirus E7 oncogene induce E7 disruption and a transformed phenotype in HPV16/18-positive cervical cancer cells. Clin. Cancer Res. 2014;20:6495–6503. doi: 10.1158/1078-0432.CCR-14-0250. [DOI] [PubMed] [Google Scholar]
- East-Seletsky A., O'Connell M.R., Knight S.C., Burstein D., Cate J.H., Tjian R., Doudna J.A. Two distinct RNase activities of CRISPR-C2c2 enable guide-RNA processing and RNA detection. Nature. 2016;538:270–273. doi: 10.1038/nature19802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El-Brolosy M.A., Kontarakis Z., Rossi A., Kuenne C., Gunther S., Fukuda N., Kikhi K., Boezio G.L.M., Takacs C.M., Lai S.L. Genetic compensation triggered by mutant mRNA degradation. Nature. 2019;568:193–197. doi: 10.1038/s41586-019-1064-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandez A., Josa S., Montoliu L. A history of genome editing in mammals. Mamm. Genome. 2017;28:237–246. doi: 10.1007/s00335-017-9699-2. [DOI] [PubMed] [Google Scholar]
- Finn J.D., Smith A.R., Patel M.C., Shaw L., Youniss M.R., van Heteren J., Dirstine T., Ciullo C., Lescarbeau R., Seitzer J. A single administration of CRISPR/Cas9 lipid nanoparticles achieves robust and persistent in vivo genome editing. Cell Rep. 2018;22:2455–2468. doi: 10.1016/j.celrep.2018.02.014. [DOI] [PubMed] [Google Scholar]
- Fiorotto R., Amenduni M., Mariotti V., Fabris L., Spirli C., Strazzabosco M. Liver diseases in the dish: iPSC and organoids as a new approach to modeling liver diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2019;1865:920–928. doi: 10.1016/j.bbadis.2018.08.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frit P., Barboule N., Yuan Y., Gomez D., Calsou P. Alternative end-joining pathway(s): bricolage at DNA breaks. DNA Repair (Amst.) 2014;17:81–97. doi: 10.1016/j.dnarep.2014.02.007. [DOI] [PubMed] [Google Scholar]
- Fu Y., Sander J.D., Reyon D., Cascio V.M., Joung J.K., Unit P., Hospital M.G., Biology I., Hospital M.G., Hospital M.G. Improving CRISPR-Cas nuclease specificity using truncated guide RNAs. Nat. Biotechnol. 2014;32:279–284. doi: 10.1038/nbt.2808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao L., Cox D.B.T., Yan W.X., Manteiga J.C., Schneider M.W., Yamano T., Nishimasu H., Nureki O., Crosetto N., Zhang F. Engineered Cpf1 variants with altered PAM specificities increase genome targeting range. Nat. Biotechnol. 2017;35:789–792. doi: 10.1038/nbt.3900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao X., Tao Y., Lamas V., Huang M., Yeh W.H., Pan B., Hu Y.J., Hu J.H., Thompson D.B., Shu Y. Treatment of autosomal dominant hearing loss by in vivo delivery of genome editing agents. Nature. 2018;553:217–221. doi: 10.1038/nature25164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garneau J.E., Dupuis M.E., Villion M., Romero D.A., Barrangou R., Boyaval P., Fremaux C., Horvath P., Magadan A.H., Moineau S. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature. 2010;468:67–71. doi: 10.1038/nature09523. [DOI] [PubMed] [Google Scholar]
- Gasiunas G., Barrangou R., Horvath P., Siksnys V. Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proc. Natl. Acad. Sci. U S A. 2012;109:E2579–E2586. doi: 10.1073/pnas.1208507109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaudelli N.M., Komor A.C., Rees H.A., Packer M.S., Badran A.H., Bryson D.I., Liu D.R. Programmable base editing of A*T to G*C in genomic DNA without DNA cleavage. Nature. 2017;551:464–471. doi: 10.1038/nature24644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glass Z., Lee M., Li Y., Xu Q. Engineering the delivery system for CRISPR-based genome editing. Trends Biotechnol. 2018;36:173–185. doi: 10.1016/j.tibtech.2017.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomes-Silva D., Srinivasan M., Sharma S., Lee C.M., Wagner D.L., Davis T.H., Rouce R.H., Bao G., Brenner M.K., Mamonkin M. CD7-edited T cells expressing a CD7-specific CAR for the therapy of T-cell malignancies. Blood. 2017;130:285–296. doi: 10.1182/blood-2017-01-761320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grunewald J., Zhou R., Garcia S.P., Iyer S., Lareau C.A., Aryee M.J., Joung J.K. Transcriptome-wide off-target RNA editing induced by CRISPR-guided DNA base. Nature. 2019;569:433–437. doi: 10.1038/s41586-019-1161-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grunewald J., Zhou R., Iyer S., Lareau C.A., Garcia S.P., Aryee M.J., Joung J.K. CRISPR DNA base editors with reduced RNA off-target and self-editing activities. Nat. Biotechnol. 2019;37:1041–1048. doi: 10.1038/s41587-019-0236-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guha T.K., Edgell D.R. Applications of alternative nucleases in the age of CRISPR/Cas9. Int. J. Mol. Sci. 2017;18:2565. doi: 10.3390/ijms18122565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guha T.K., Wai A., Hausner G. Programmable genome editing tools and their regulation for efficient genome engineering. Comput. Struct. Biotechnol. J. 2017;15:146–160. doi: 10.1016/j.csbj.2016.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo Q., Mintier G., Ma-Edmonds M., Storton D., Wang X., Xiao X., Kienzle B., Zhao D., Feder J.N. Cold shock' increases the frequency of homology directed repair gene editing in induced pluripotent stem cells. Sci. Rep. 2018;8:2080. doi: 10.1038/s41598-018-20358-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo X., Jiang H., Shi B., Zhou M., Zhang H., Shi Z., Du G., Luo H., Wu X., Wang Y. Disruption of PD-1 enhanced the anti-tumor activity of chimeric antigen receptor T cells against hepatocellular carcinoma. Front. Pharmacol. 2018;9:1118. doi: 10.3389/fphar.2018.01118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gupta R.M., Musunuru K. Expanding the genetic editing tool kit: ZFNs, TALENs, and CRISPR-Cas9. J. Clin. Invest. 2014;124:4154–4161. doi: 10.1172/JCI72992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haapaniemi E., Botla S., Persson J., Schmierer B., Taipale J. CRISPR–Cas9 genome editing induces a p53-mediated DNA damage response. Nat. Med. 2018:1. doi: 10.1038/s41591-018-0049-z. [DOI] [PubMed] [Google Scholar]
- Hannoun Z., Steichen C., Dianat N., Weber A., Dubart-Kupperschmitt A. The potential of induced pluripotent stem cell derived hepatocytes. J. Hepatol. 2016;65:182–199. doi: 10.1016/j.jhep.2016.02.025. [DOI] [PubMed] [Google Scholar]
- Hendel A., Bak R.O., Clark J.T., Kennedy A.B., Ryan D.E., Roy S., Steinfeld I., Lunstad B.D., Kaiser R.J., Alec B. Chemically modified guide RNAs enhance CRISPR-Cas genome editing in human primary cells. Nat. Biotechnol. 2015;33:985–989. doi: 10.1038/nbt.3290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hess G.T., Tycko J., Yao D., Bassik M.C. Methods and applications of CRISPR-mediated base editing in Eukaryotic genomes. Mol. Cell. 2017;68:26–43. doi: 10.1016/j.molcel.2017.09.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hilton I.B., D'Ippolito A.M., Vockley C.M., Thakore P.I., Crawford G.E., Reddy T.E., Gersbach C.A. Epigenome editing by a CRISPR-Cas9-based acetyltransferase activates genes from promoters and enhancers. Nat. Biotechnol. 2015;33:510–517. doi: 10.1038/nbt.3199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hisano Y., Sakuma T., Nakade S., Ohga R., Ota S., Okamoto H., Yamamoto T., Kawahara A. Precise in-frame integration of exogenous DNA mediated by CRISPR/Cas9 system in zebrafish. Sci. Rep. 2015;5:8841. doi: 10.1038/srep08841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holt N., Wang J., Kim K., Friedman G., Wang X., Taupin V., Crooks G.M., Kohn D.B., Gregory P.D., Holmes M.C., Cannon P.M. Human hematopoietic stem/progenitor cells modified by zinc-finger nucleases targeted to CCR5 control HIV-1 in vivo. Nat. Biotechnol. 2010;28:839–847. doi: 10.1038/nbt.1663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsu P.D., Scott D.A., Weinstein J.A., Ran F.A., Konermann S., Agarwala V., Li Y., Fine E.J., Wu X., Shalem O. DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. 2013;31:827–832. doi: 10.1038/nbt.2647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu Z., Yu L., Zhu D., Ding W., Wang X., Zhang C., Wang L., Jiang X., Shen H., He D. Disruption of HPV16-E7 by CRISPR/Cas system induces apoptosis and growth inhibition in HPV16 positive human cervical cancer cells. Biomed. Res. Int. 2014;2014:612823. doi: 10.1155/2014/612823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu Z., Ding W., Zhu D., Yu L., Jiang X., Wang X., Zhang C., Wang L., Ji T., Liu D. TALEN-mediated targeting of HPV oncogenes ameliorates HPV-related cervical malignancy. J. Clin. Invest. 2015;125:425–436. doi: 10.1172/JCI78206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu J.H., Miller S.M., Geurts M.H., Tang W., Chen L., Sun N., Zeina C.M., Gao X., Rees H.A., Lin Z., Liu D.R. Evolved Cas9 variants with broad PAM compatibility and high DNA specificity. Nature. 2018;556:57–63. doi: 10.1038/nature26155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu B., Zou Y., Zhang L., Tang J., Niedermann G., Firat E., Huang X., Zhu X. Nucleofection with plasmid DNA for CRISPR/Cas9-Mediated inactivation of programmed cell death protein 1 in CD133-specific CAR T cells. Hum. Gene Ther. 2019;30:446–458. doi: 10.1089/hum.2017.234. [DOI] [PubMed] [Google Scholar]
- Hu W., Zi Z., Jin Y., Li G., Shao K., Cai Q., Ma X., Wei F. CRISPR/Cas9-mediated PD-1 disruption enhances human mesothelin-targeted CAR T cell effector functions. Cancer Immunol. Immunother. 2019;68:365–377. doi: 10.1007/s00262-018-2281-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang X., Zhou G., Wu W., Duan Y., Ma G., Song J., Xiao R., Vandenberghe L., Zhang F., D'Amore P.A., Lei H. Genome editing abrogates angiogenesis in vivo. Nat. Commun. 2017;8:4–11. doi: 10.1038/s41467-017-00140-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang C.-h., Lee K.-c., Doudna J.A. Applications of CRISPR-Cas enzymes in cancer therapeutics and detection. Trends Cancer. 2018;4:499–512. doi: 10.1016/j.trecan.2018.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ihry R.J., Worringer K.A., Salick M.R., Frias E., Ho D., Theriault K., Kommineni S., Chen J., Sondey M., Ye C. p53 inhibits CRISPR – Cas9 engineering in human pluripotent stem cells. Nat. Med. 2018:1–8. doi: 10.1038/s41591-018-0050-6. [DOI] [PubMed] [Google Scholar]
- Ihry R.J., Salick M.R., Ho D.J., Sondey M., Kommineni S., Paula S., Raymond J., Henry B., Frias E., Wang Q. Genome-Scale CRISPR screens identify human pluripotency-specific genes. Cell Rep. 2019;27:616–630.e6. doi: 10.1016/j.celrep.2019.03.043. [DOI] [PubMed] [Google Scholar]
- Ishino Y., Shinagawa H., Makino K., Amemura M., Nakata A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. J. Bacteriol. 1987;169:5429–5433. doi: 10.1128/jb.169.12.5429-5433.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jang Y.Y., Ye Z. Gene correction in patient-specific iPSCs for therapy development and disease modeling. Hum. Genet. 2016;135:1041–1058. doi: 10.1007/s00439-016-1691-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jansen R., Embden J.D., Gaastra W., Schouls L.M. Identification of genes that are associated with DNA repeats in prokaryotes. Mol. Microbiol. 2002;43:1565–1575. doi: 10.1046/j.1365-2958.2002.02839.x. [DOI] [PubMed] [Google Scholar]
- Jiang F., Taylor D.W., Chen J.S., Kornfeld J.E., Zhou K., Thompson A.J., Nogales E., Doudna J.A. Structures of a CRISPR-Cas9 R-loop complex primed for DNA cleavage. Science. 2016;351:867–871. doi: 10.1126/science.aad8282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang F., Liu J.J., Osuna B.A., Xu M., Berry J.D., Rauch B.J., Nogales E., Bondy-Denomy J., Doudna J.A. Temperature-Responsive competitive inhibition of CRISPR-Cas9. Mol. Cell. 2019;73:601–610.e5. doi: 10.1016/j.molcel.2018.11.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jinek M., Chylinski K., Fonfara I., Hauer M., Doudna J.A., Charpentier E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science. 2012;337:816–821. doi: 10.1126/science.1225829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karvelis T., Gasiunas G., Siksnys V. Harnessing the natural diversity and in vitro evolution of Cas9 to expand the genome editing toolbox. Curr. Opin. Microbiol. 2017;37:88–94. doi: 10.1016/j.mib.2017.05.009. [DOI] [PubMed] [Google Scholar]
- Kennedy E.M., Kornepati A.V., Goldstein M., Bogerd H.P., Poling B.C., Whisnant A.W., Kastan M.B., Cullen B.R. Inactivation of the human papillomavirus E6 or E7 gene in cervical carcinoma cells by using a bacterial CRISPR/Cas RNA-guided endonuclease. J. Virol. 2014;88:11965–11972. doi: 10.1128/JVI.01879-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim E., Koo T., Park S.W., Kim D., Kim K., Cho H.Y., Song D.W., Lee K.J., Jung M.H., Kim S. In vivo genome editing with a small Cas9 orthologue derived from Campylobacter jejuni. Nat. Commun. 2017;8:1–12. doi: 10.1038/ncomms14500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim S., Koo T., Jee H.-G., Cho H.-Y., Lee G., Lim D.-G., Shin H.S., Kim J.-S. CRISPR RNAs trigger innate immune responses in human cells. Genome Res. 2018;28:367–373. doi: 10.1101/gr.231936.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim S.I., Matsumoto T., Kagawa H., Nakamura M., Hirohata R., Ueno A., Ohishi M., Sakuma T., Soga T., Yamamoto T., Woltjen K. Microhomology-assisted scarless genome editing in human iPSCs. Nat. Commun. 2018;9:939. doi: 10.1038/s41467-018-03044-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim D., Kim D.E., Lee G., Cho S.I., Kim J.S. Genome-wide target specificity of CRISPR RNA-guided adenine base editors. Nat. Biotechnol. 2019;37:430–435. doi: 10.1038/s41587-019-0050-1. [DOI] [PubMed] [Google Scholar]
- Kleinstiver B.P., Prew M.S., Tsai S.Q., Topkar V.V., Nguyen N.T., Zheng Z., Gonzales A.P.W., Li Z., Peterson R.T., Yeh J.R.J. Engineered CRISPR-Cas9 nucleases with altered PAM specificities. Nature. 2015;523:481–485. doi: 10.1038/nature14592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kleinstiver B.P., Pattanayak V., Prew M.S., Tsai S.Q., Nguyen N., Zheng Z., Joung J.K., Unit P., Biology I., Hospital M.G., Kong H. High-fidelity CRISPR-Cas9 variants with undetectable genome-wide off-targets. Nature. 2016;529:490–495. doi: 10.1038/nature16526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kocak D.D., Josephs E.A., Bhandarkar V., Adkar S.S., Kwon J.B., Gersbach C.A. Increasing the specificity of CRISPR systems with engineered RNA secondary structures. Nat. Biotechnol. 2019;37:657–666. doi: 10.1038/s41587-019-0095-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Komor A.C., Kim Y.B., Packer M.S., Zuris J.A., Liu D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature. 2016;533:420–424. doi: 10.1038/nature17946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Konermann S., Lotfy P., Brideau N.J., Oki J., Shokhirev M.N., Hsu P.D. Transcriptome engineering with RNA-targeting type VI-D CRISPR effectors. Cell. 2018;173:665–676.e14. doi: 10.1016/j.cell.2018.02.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kosicki M., Bradley A. Repair of CRISPR–Cas9-induced double-stranded breaks leads to large deletions and complex rearrangements. Nat. Biotechnol. 2018;36:765–771. doi: 10.1038/nbt.4192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuscu C., Parlak M., Tufan T., Yang J., Szlachta K., Wei X., Mammadov R., Adli M. CRISPR-STOP: gene silencing through base-editing-induced nonsense mutations. Nat. Methods. 2017;14:710–712. doi: 10.1038/nmeth.4327. [DOI] [PubMed] [Google Scholar]
- Laoharawee K., DeKelver R.C., Podetz-Pedersen K.M., Rohde M., Sproul S., Nguyen H.O., Nguyen T., St Martin S.J., Ou L. Dose-dependent prevention of metabolic and neurologic disease in murine MPS II by ZFN-mediated in vivo genome editing. Mol. Ther. 2018;26:1127–1136. doi: 10.1016/j.ymthe.2018.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laughery M.F., Mayes H.C., Pedroza I.K., Wyrick J.J. R-loop formation by dCas9 is mutagenic in Saccharomyces cerevisiae. Nucleic Acids Res. 2019;47:2389–2401. doi: 10.1093/nar/gky1278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee K., Conboy M., Park H.M., Jiang F., Kim H.J., Dewitt M.A., Mackley V.A., Chang K., Rao A., Skinner C. Nanoparticle delivery of Cas9 ribonucleoprotein and donor DNA in vivo induces homology-directed DNA repair. Nat. Biomed. Eng. 2017;1:889–901. doi: 10.1038/s41551-017-0137-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee B., Lee K., Panda S., Gonzales-Rojas R., Chong A., Bugay V., Park H.M., Brenner R., Murthy N., Lee H.Y. Nanoparticle delivery of CRISPR into the brain rescues a mouse model of fragile X syndrome from exaggerated repetitive behaviours. Nat. Biomed. Eng. 2018;2:497–507. doi: 10.1038/s41551-018-0252-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lessard S., Francioli L., Alfoldi J., Tardif J.-C., Ellinor P.T., MacArthur D.G., Lettre G., Orkin S.H., Canver M.C. Human genetic variation alters CRISPR-Cas9 on- and off-targeting specificity at therapeutically implicated loci. Proc. Natl. Acad. Sci. U S A. 2017;114:E11257–E11266. doi: 10.1073/pnas.1714640114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li H.L., Fujimoto N., Sasakawa N., Shirai S., Ohkame T., Sakuma T., Tanaka M., Amano N., Watanabe A., Sakurai H. Precise correction of the dystrophin gene in duchenne muscular dystrophy patient induced pluripotent stem cells by TALEN and CRISPR-Cas9. Stem Cell Reports. 2015;4:143–154. doi: 10.1016/j.stemcr.2014.10.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L., Hu S., Chen X. Non-viral delivery systems for CRISPR/Cas9-based genome editing: challenges and opportunities. Biomaterials. 2018;171:207–218. doi: 10.1016/j.biomaterials.2018.04.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liao H.K., Hatanaka F., Araoka T., Reddy P., Wu M.Z., Sui Y., Yamauchi T., Sakurai M., O'Keefe D.D., Nunez-Delicado E. In vivo target gene activation via CRISPR/Cas9-Mediated trans-epigenetic modulation. Cell. 2017;171:1495–1507.e15. doi: 10.1016/j.cell.2017.10.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Listgarten J., Weinstein M., Kleinstiver B.P., Sousa A.A., Joung J.K., Crawford J., Gao K., Hoang L., Elibol M., Doench J.G., Fusi N. Prediction of off-target activities for the end-to-end design of CRISPR guide RNAs. Nat. Biomed. Eng. 2018;2:38–47. doi: 10.1038/s41551-017-0178-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu N., Hargreaves V.V., Zhu Q., Kurland J.V., Hong J., Kim W., Sher F., Macias-Trevino C., Rogers J.M., Kurita R. Direct promoter repression by BCL11A controls the fetal to adult hemoglobin switch. Cell. 2018;173:430–442.e17. doi: 10.1016/j.cell.2018.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu X.S., Wu H., Krzisch M., Wu X., Graef J., Muffat J., Hnisz D., Li C.H., Yuan B., Xu C. Rescue of fragile X syndrome neurons by DNA methylation editing of the FMR1 gene. Cell. 2018;172:979–991.e6. doi: 10.1016/j.cell.2018.01.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Louis Jeune V., Joergensen J.A., Hajjar R.J., Weber T. Pre-existing anti–adeno-associated virus antibodies as a challenge in AAV gene therapy. Hum. Gene Ther. Methods. 2013;24:59–67. doi: 10.1089/hgtb.2012.243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu S., Yang N., He J., Gong W., Lai Z., Xie L., Tao L., Xu C., Wang H., Zhang G. Generation of cancer-specific cytotoxic PD-1(-) T cells using liposome-encapsulated CRISPR/Cas system with dendritic/tumor fusion cells. J. Biomed. Nanotechnol. 2019;15:593–601. doi: 10.1166/jbn.2019.2712. [DOI] [PubMed] [Google Scholar]
- Maeder M.L., Gersbach C.A. Genome-editing technologies for gene and cell therapy. Mol. Ther. 2016;24:430–446. doi: 10.1038/mt.2016.10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maeder M.L., Stefanidakis M., Wilson C.J., Baral R., Barrera L.A., Bounoutas G.S., Bumcrot D., Chao H., Ciulla D.M., DaSilva J.A. Development of a gene-editing approach to restore vision loss in Leber congenital amaurosis type 10. Nat. Med. 2019;25:229–233. doi: 10.1038/s41591-018-0327-9. [DOI] [PubMed] [Google Scholar]
- Magli A., Perlingeiro R.R.C. Myogenic progenitor specification from pluripotent stem cells. Semin. Cell Dev. Biol. 2017;72:87–98. doi: 10.1016/j.semcdb.2017.10.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mahas A., Neal Stewart C., Jr., Mahfouz M.M. Harnessing CRISPR/Cas systems for programmable transcriptional and post-transcriptional regulation. Biotechnol. Adv. 2018;36:295–310. doi: 10.1016/j.biotechadv.2017.11.008. [DOI] [PubMed] [Google Scholar]
- Mair B., Tomic J., Masud S.N., Tonge P., Weiss A., Usaj M., Tong A.H.Y., Kwan J.J., Brown K.R., Titus E. Essential gene profiles for human pluripotent stem cells identify uncharacterized genes and substrate dependencies. Cell Rep. 2019;27:599–615.e12. doi: 10.1016/j.celrep.2019.02.041. [DOI] [PubMed] [Google Scholar]
- Mandai M., Watanabe A., Kurimoto Y., Hirami Y., Morinaga C., Daimon T., Fujihara M., Akimaru H., Sakai N., Shibata Y. Autologous induced stem-cell-derived retinal cells for macular degeneration. N. Engl. J. Med. 2017;376:1038–1046. doi: 10.1056/NEJMoa1608368. [DOI] [PubMed] [Google Scholar]
- Martin R.M., Ikeda K., Cromer M.K., Uchida N., Nishimura T., Romano R., Tong A.J., Lemgart V.T., Camarena J., Pavel-Dinu M. Highly efficient and marker-free genome editing of human pluripotent stem cells by CRISPR-Cas9 RNP and AAV6 donor-mediated homologous recombination. Cell Stem Cell. 2019;24:821–828.e5. doi: 10.1016/j.stem.2019.04.001. [DOI] [PubMed] [Google Scholar]
- Maruyama T., Dougan S.K., Truttmann M.C., Bilate A.M., Ingram J.R., Ploegh H.L. Increasing the efficiency of precise genome editing with CRISPR-Cas9 by inhibition of nonhomologous end joining. Nat. Biotechnol. 2015;33:538–542. doi: 10.1038/nbt.3190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mateos-Gomez P.A., Kent T., Deng S.K., McDevitt S., Kashkina E., Hoang T.M., Pomerantz R.T., Sfeir A. The helicase domain of Poltheta counteracts RPA to promote alt-NHEJ. Nat. Struct. Mol. Biol. 2017;24:1116–1123. doi: 10.1038/nsmb.3494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matharu N., Rattanasopha S., Tamura S., Maliskova L., Wang Y., Bernard A., Hardin A., Eckalbar W.L., Vaisse C., Ahituv N. CRISPR-mediated activation of a promoter or enhancer rescues obesity caused by haploinsufficiency. Science. 2018;0629:eaau0629. doi: 10.1126/science.aau0629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menger L., Sledzinska A., Bergerhoff K., Vargas F.A., Smith J., Poirot L., Pule M., Hererro J., Peggs K.S., Quezada S.A. TALEN-Mediated inactivation of PD-1 in tumor-reactive lymphocytes promotes intratumoral T-cell persistence and rejection of established tumors. Cancer Res. 2016;76:2087–2093. doi: 10.1158/0008-5472.CAN-15-3352. [DOI] [PubMed] [Google Scholar]
- Mingozzi F., High K.A. Therapeutic in vivo gene transfer for genetic disease using AAV: progress and challenges. Nat. Rev. Genet. 2011;12:341–355. doi: 10.1038/nrg2988. [DOI] [PubMed] [Google Scholar]
- Mingozzi F., High K.A. Immune responses to AAV vectors: overcoming barriers to successful gene therapy. Blood. 2013;122:23–36. doi: 10.1182/blood-2013-01-306647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mojica F.J., Montoliu L. On the origin of CRISPR-Cas technology: from prokaryotes to mammals. Trends Microbiol. 2016;24:811–820. doi: 10.1016/j.tim.2016.06.005. [DOI] [PubMed] [Google Scholar]
- Molla K.A., Yang Y. CRISPR/Cas-Mediated base editing: technical considerations and practical applications. Trends Biotechnol. 2019;37:1121–1142. doi: 10.1016/j.tibtech.2019.03.008. [DOI] [PubMed] [Google Scholar]
- Mou H., Smith J.L., Peng L., Yin H., Moore J., Zhang X.O., Song C.Q., Sheel A., Wu Q., Ozata D.M. CRISPR/Cas9-mediated genome editing induces exon skipping by alternative splicing or exon deletion. Genome Biol. 2017;18:108. doi: 10.1186/s13059-017-1237-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakade S., Tsubota T., Sakane Y., Kume S., Sakamoto N., Obara M., Daimon T., Sezutsu H., Yamamoto T., Sakuma T., Suzuki K.T. Microhomology-mediated end-joining-dependent integration of donor DNA in cells and animals using TALENs and CRISPR/Cas9. Nat. Commun. 2014;5:5560. doi: 10.1038/ncomms6560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naldini L. Gene therapy returns to centre stage. Nature. 2015;526:351–360. doi: 10.1038/nature15818. [DOI] [PubMed] [Google Scholar]
- Nelson C.E., Hakim C.H., Ousterout D.G., Thakore P.I., Moreb E.A., Castellanos Rivera R.M., Madhavan S., Pan X., Ran F.A. In vivo genome editing improves muscle function in a mouse model of Duchenne muscular dystrophy. Science. 2016;351:403–407. doi: 10.1126/science.aad5143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishida K., Arazoe T., Yachie N., Banno S., Kakimoto M., Tabata M., Mochizuki M., Miyabe A., Araki M., Hara K.Y. Targeted nucleotide editing using hybrid prokaryotic and vertebrate adaptive immune systems. Science. 2016;353:aaf8729. doi: 10.1126/science.aaf8729. [DOI] [PubMed] [Google Scholar]
- Nishimasu H., Shi X., Ishiguro S., Gao L., Hirano S., Okazaki S., Noda T., Abudayyeh O.O., Gootenberg J.S., Mori H. Engineered CRISPR-Cas9 nuclease with expanded targeting space. Science. 2018;361:1259–1262. doi: 10.1126/science.aas9129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishiyama J., Mikuni T., Yasuda R. Virus-Mediated genome editing via homology-directed repair in mitotic and postmitotic cells in mammalian brain. Neuron. 2017;96:755–768.e5. doi: 10.1016/j.neuron.2017.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Connell M.R., Oakes B.L., Sternberg S.H., East-Seletsky A., Kaplan M., Doudna J.A. Programmable RNA recognition and cleavage by CRISPR/Cas9. Nature. 2014;516:263–266. doi: 10.1038/nature13769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orthwein A., Noordermeer S.M., Wilson M.D., Landry S., Enchev R.I., Sherker A., Munro M., Pinder J., Salsman J., Dellaire G. A mechanism for the suppression of homologous recombination in G1 cells. Nature. 2015;528:422–426. doi: 10.1038/nature16142. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Ouchi Y., Patil A., Tamura Y., Nishimasu H., Negishi A., Paul S.K., Takemura N., Satoh T., Kimura Y., Kurachi M. Generation of tumor antigen-specific murine CD8+ T cells with enhanced anti-tumor activity via highly efficient CRISPR/Cas9 genome editing. Int. Immunol. 2018;30:141–154. doi: 10.1093/intimm/dxy006. [DOI] [PubMed] [Google Scholar]
- Pankowicz F.P., Barzi M., Legras X., Hubert L., Mi T., Tomolonis J.A., Ravishankar M., Sun Q., Yang D., Borowiak M. Reprogramming metabolic pathways in vivo with CRISPR/Cas9 genome editing to treat hereditary tyrosinaemia. Nat. Commun. 2016;7:1–6. doi: 10.1038/ncomms12642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park H., Oh J., Shim G., Cho B., Chang Y., Kim S., Baek S., Kim H., Shin J., Choi H. In vivo neuronal gene editing via CRISPR-Cas9 amphiphilic nanocomplexes alleviates deficits in mouse models of Alzheimers disease. Nat. Neurosci. 2019;22:524–528. doi: 10.1038/s41593-019-0352-0. [DOI] [PubMed] [Google Scholar]
- Park S.H., Lee C.M., Dever D.P., Davis T.H., Camarena J., Srifa W., Zhang Y., Paikari A., Chang A.K., Porteus M.H. Highly efficient editing of the beta-globin gene in patient-derived hematopoietic stem and progenitor cells to treat sickle cell disease. Nucleic Acids Res. 2019;47:7955–7972. doi: 10.1093/nar/gkz475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pawluk A., Amrani N., Zhang Y., Garcia B., Hidalgo-Reyes Y., Lee J., Edraki A., Shah M., Sontheimer E.J., Maxwell K.L., Davidson A.R. Naturally occurring off-switches for CRISPR-Cas9. Cell. 2016;167:1829–1838.e9. doi: 10.1016/j.cell.2016.11.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pawluk A., Davidson A.R., Maxwell K.L. Anti-CRISPR: discovery, mechanism and function. Nat. Rev. Microbiol. 2018;16:12–17. doi: 10.1038/nrmicro.2017.120. [DOI] [PubMed] [Google Scholar]
- Perez E.E., Wang J., Miller J.C., Jouvenot Y., Kim K.A., Liu O., Wang N., Lee G., Bartsevich V.V., Lee Y.L. Establishment of HIV-1 resistance in CD4+ T cells by genome editing using zinc-finger nucleases. Nat. Biotechnol. 2008;26:808–816. doi: 10.1038/nbt1410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pinder J., Salsman J., Dellaire G. Nuclear domain 'knock-in' screen for the evaluation and identification of small molecule enhancers of CRISPR-based genome editing. Nucleic Acids Res. 2015;43:9379–9392. doi: 10.1093/nar/gkv993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Psatha N., Reik A., Phelps S., Zhou Y., Dalas D., Yannaki E., Levasseur D.N., Urnov F.D., Holmes M.C., Papayannopoulou T. Disruption of the BCL11A erythroid enhancer reactivates fetal hemoglobin in erythroid cells of patients with beta-thalassemia major. Mol. Ther. Methods Clin. Dev. 2018;10:313–326. doi: 10.1016/j.omtm.2018.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qi L.S., Larson M.H., Gilbert L.A., Doudna J.A., Weissman J.S., Arkin A.P., Lim W.A. Repurposing CRISPR as an RNA-guided platform for sequence-specific control of gene expression. Cell. 2013;152:1173–1183. doi: 10.1016/j.cell.2013.02.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ran F.A., Hsu P.D., Lin C.-y., Gootenberg J.S., Trevino A., Scott D.A., Inoue A., Matoba S. Double nicking by RNA-guided CRISPR Cas9 for enhanced genome editing specificity. Cell. 2013;154:1380–1389. doi: 10.1016/j.cell.2013.08.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ranjha L., Howard S.M., Cejka P. Main steps in DNA double-strand break repair: an introduction to homologous recombination and related processes. Chromosoma. 2018;127:187–214. doi: 10.1007/s00412-017-0658-1. [DOI] [PubMed] [Google Scholar]
- Richter F., Fonfara I., Gelfert R., Nack J., Charpentier E., Möglich A. Switchable Cas9. Curr. Opin. Biotechnol. 2017;48:119–126. doi: 10.1016/j.copbio.2017.03.025. [DOI] [PubMed] [Google Scholar]
- Rodriguez-Rodriguez D.R., Ramirez-Solis R., Garza-Elizondo M.A., Garza-Rodriguez M.L., Barrera-Saldana H.A. Genome editing: a perspective on the application of CRISPR/Cas9 to study human diseases (Review), Int. J. Mol. Med. 2019;43:1559–1574. doi: 10.3892/ijmm.2019.4112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossidis A.C., Stratigis J.D., Chadwick A.C., Hartman H.A., Ahn N.J., Li H., Singh K., Coons B.E., Li L., Lv W. In utero CRISPR-mediated therapeutic editing of metabolic genes. Nat. Med. 2018;24:1513–1518. doi: 10.1038/s41591-018-0184-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruan G.X., Barry E., Yu D., Lukason M., Cheng S.H., Scaria A. CRISPR/Cas9-Mediated genome editing as a therapeutic approach for leber congenital amaurosis 10. Mol. Ther. 2017;25:331–341. doi: 10.1016/j.ymthe.2016.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rupp L.J., Schumann K., Roybal K.T., Gate R.E., Ye C.J., Lim W.A., Marson A. CRISPR/Cas9-mediated PD-1 disruption enhances anti-tumor efficacy of human chimeric antigen receptor T cells. Sci. Rep. 2017;7:737. doi: 10.1038/s41598-017-00462-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sadelain M., Papapetrou E.P., Bushman F.D. Safe harbours for the integration of new DNA in the human genome. Nat. Rev. Cancer. 2011;12:51–58. doi: 10.1038/nrc3179. [DOI] [PubMed] [Google Scholar]
- Sakuma T., Nakade S., Sakane Y., Suzuki K.T., Yamamoto T. MMEJ-assisted gene knock-in using TALENs and CRISPR-Cas9 with the PITCh systems. Nat. Protoc. 2016;11:118–133. doi: 10.1038/nprot.2015.140. [DOI] [PubMed] [Google Scholar]
- Santiago-Fernandez O., Osorio F.G., Quesada V., Rodriguez F., Basso S., Maeso D., Rolas L., Barkaway A., Nourshargh S., Folgueras A.R. Development of a CRISPR/Cas9-based therapy for Hutchinson-Gilford progeria syndrome. Nat. Med. 2019;25:423–426. doi: 10.1038/s41591-018-0338-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sawatsubashi S., Joko Y., Fukumoto S., Matsumoto T., Sugano S.S. Development of versatile non-homologous end joining-based knock-in module for genome editing. Sci. Rep. 2018;8:593. doi: 10.1038/s41598-017-18911-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schimmel J., Kool H., van Schendel R., Tijsterman M. Mutational signatures of non-homologous and polymerase theta-mediated end-joining in embryonic stem cells. EMBO J. 2017;36:3634–3649. doi: 10.15252/embj.201796948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schiroli G., Conti A., Ferrari S., Della Volpe L., Jacob A., Albano L., Beretta S., Calabria A., Vavassori V., Gasparini P. Precise gene editing preserves hematopoietic stem cell function following transient p53-mediated DNA damage response. Cell Stem Cell. 2019;24:551–565.e8. doi: 10.1016/j.stem.2019.02.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwank G., Koo B.K., Sasselli V., Dekkers J.F., Heo I., Demircan T., Sasaki N., Boymans S., Cuppen E., van der Ent C.K. Functional repair of CFTR by CRISPR/Cas9 in intestinal stem cell organoids of cystic fibrosis patients. Cell Stem Cell. 2013;13:653–658. doi: 10.1016/j.stem.2013.11.002. [DOI] [PubMed] [Google Scholar]
- Shankar S., Prasad D., Sanawar R., Das A.V., Pillai M.R. TALEN based HPV-E7 editing triggers necrotic cell death in cervical cancer cells. Sci. Rep. 2017;7:5500. doi: 10.1038/s41598-017-05696-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma R., Anguela X.M., Doyon Y., Wechsler T., DeKelver R.C., Sproul S., Paschon D.E., Miller J.C., Davidson R.J., Shivak D. In vivo genome editing of the albumin locus as a platform for protein replacement therapy. Blood. 2015;126:1777–1784. doi: 10.1182/blood-2014-12-615492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slaymaker I.M., Gao L., Zetsche B., Scott D.A., Yan W.X., Zhang F. Rationally engineered Cas9 nucleases with improved specificity. Science. 2016;351:84–88. doi: 10.1126/science.aad5227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith J.L., Mou H., Xue W. Understanding and repurposing CRISPR-mediated alternative splicing. Genome Biol. 2018;19:184. doi: 10.1186/s13059-018-1565-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song J., Yang D., Xu J., Zhu T., Chen Y.E., Zhang J. RS-1 enhances CRISPR/Cas9- and TALEN-mediated knock-in efficiency. Nat. Commun. 2016;7:10548. doi: 10.1038/ncomms10548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Su S., Hu B., Shao J., Shen B., Du J., Du Y., Zhou J., Yu L., Zhang L., Chen F. CRISPR-Cas9 mediated efficient PD-1 disruption on human primary T cells from cancer patients. Sci. Rep. 2016;6:20070. doi: 10.1038/srep20070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Su S., Zou Z., Chen F., Ding N., Du J., Shao J., Li L., Fu Y., Hu B., Yang Y. CRISPR-Cas9-mediated disruption of PD-1 on human T cells for adoptive cellular therapies of EBV positive gastric cancer. Oncoimmunology. 2017;6:e1249558. doi: 10.1080/2162402X.2016.1249558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki K., Izpisua Belmonte J.C. In vivo genome editing via the HITI method as a tool for gene therapy. J. Hum. Genet. 2018;63:157–164. doi: 10.1038/s10038-017-0352-4. [DOI] [PubMed] [Google Scholar]
- Suzuki K., Tsunekawa Y., Hernandez-Benitez R., Wu J., Zhu J., Kim E.J., Hatanaka F., Yamamoto M., Araoka T., Li Z. In vivo genome editing via CRISPR/Cas9 mediated homology-independent targeted integraion. Nature. 2016;540:144–149. doi: 10.1038/nature20565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szczelkun M.D., Tikhomirova M.S., Sinkunas T., Gasiunas G., Karvelis T., Pschera P., Siksnys V., Seidel R. Direct observation of R-loop formation by single RNA-guided Cas9 and Cascade effector complexes. Proc. Natl. Acad. Sci. U S A. 2014;111:9798–9803. doi: 10.1073/pnas.1402597111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tabebordbar M., Zhu K., Cheng J.K.W., Chew W.L., Widrick J.J., Yan W.X., Maesner C., Wu E.Y., Xiao R., Ran F.A. In vivo gene editing in dystrophic mouse muscle and muscle stem cells. Science. 2016;351:407–411. doi: 10.1126/science.aad5177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taleei R., Nikjoo H. Biochemical DSB-repair model for mammalian cells in G1 and early S phases of the cell cycle. Mutat. Res. 2013;756:206–212. doi: 10.1016/j.mrgentox.2013.06.004. [DOI] [PubMed] [Google Scholar]
- Tebas P., Stein D., Tang W.W., Frank I., Wang S.Q., Lee G., Spratt S.K., Surosky R.T., Giedlin M.A., Nichol G. Gene editing of CCR5 in autologous CD4 T cells of persons infected with HIV. N. Engl. J. Med. 2014;370:901–910. doi: 10.1056/NEJMoa1300662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thakore P.I., D'Ippolito A.M., Song L., Safi A., Shivakumar N.K., Kabadi A.M., Reddy T.E., Crawford G.E., Gersbach C.A. Highly specific epigenome editing by CRISPR-Cas9 repressors for silencing of distal regulatory elements. Nat. Methods. 2015;12:1143–1149. doi: 10.1038/nmeth.3630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas M., Burgio G., Adams D.J., Iyer V. Collateral damage and CRISPR genome editing. PLoS Genet. 2019;15:e1007994. doi: 10.1371/journal.pgen.1007994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torikai H., Cooper L.J. Translational implications for off-the-shelf immune cells expressing chimeric antigen receptors. Mol. Ther. 2016;24:1178–1186. doi: 10.1038/mt.2016.106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Truong L.N., Li Y., Shi L.Z., Hwang P.Y., He J., Wang H., Razavian N., Berns M.W., Wu X. Microhomology-mediated End Joining and Homologous Recombination share the initial end resection step to repair DNA double-strand breaks in mammalian cells. Proc. Natl. Acad. Sci. U S A. 2013;110:7720–7725. doi: 10.1073/pnas.1213431110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsai S.Q., Wyvekens N., Khayter C., Foden J.A., Thapar V., Reyon D., Goodwin M.J., Aryee M.J., Joung K., Unit P., Hospital M.G. Dimeric CRISPR RNA-guided FokI nucleases for highly specific genome editing. Nat. Biotechnol. 2014;32:569–576. doi: 10.1038/nbt.2908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tuladhar R., Yeu Y., Tyler Piazza J., Tan Z., Rene Clemenceau J., Wu X., Barrett Q., Herbert J., Mathews D.H., Kim J. CRISPR-Cas9-based mutagenesis frequently provokes on-target mRNA misregulation. Nat. Commun. 2019;10:4056. doi: 10.1038/s41467-019-12028-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uribe R.V., van der Helm E., Misiakou M.A., Lee S.W., Kol S., Sommer M.O.A. Discovery and characterization of Cas9 inhibitors disseminated across seven bacterial phyla. Cell Host Microbe. 2019;25:233–241.e5. doi: 10.1016/j.chom.2019.01.003. [DOI] [PubMed] [Google Scholar]
- Villiger L., Grisch-Chan H.M., Lindsay H., Ringnalda F., Pogliano C.B., Allegri G., Fingerhut R., Häberle J., Matos J., Robinson M.D. Treatment of a metabolic liver disease by in vivo genome base editing in adult mice. Nat. Med. 2018;24:1519–1525. doi: 10.1038/s41591-018-0209-1. [DOI] [PubMed] [Google Scholar]
- Wagner D.L., Amani L., Wendering D.J., Reinke P., Volk H.-D., Schmueck-Henneresse M. High prevalence of Streptococcus pyogenes Cas9- reactive T cells within the adult human population. Nat. Med. 2019;25:242–248. doi: 10.1038/s41591-018-0204-6. [DOI] [PubMed] [Google Scholar]
- van der Wal E., Herrero-Hernandez P., Wan R., Broeders M., In 't Groen S.L.M., van Gestel T.J.M., van IJcken W.F.J., Cheung T.H., van der Ploeg A.T., Schaaf G.J., Pijnappel W. Large-Scale expansion of human iPSC-derived skeletal muscle cells for disease modeling and cell-based therapeutic strategies. Stem Cell Reports. 2018;10:1975–1990. doi: 10.1016/j.stemcr.2018.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang W., Ye C., Liu J., Zhang D., Kimata J.T., Zhou P. CCR5 gene disruption via lentiviral vectors expressing Cas9 and single guided RNA renders cells resistant to HIV-1 infection. PLoS One. 2014;9:e115987. doi: 10.1371/journal.pone.0115987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang M., Zuris J.A., Meng F., Rees H., Sun S., Deng P., Han Y., Gao X., Pouli D., Wu Q. Efficient delivery of genome-editing proteins using bioreducible lipid nanoparticles. Proc. Natl. Acad. Sci. U S A. 2016;113:2868–2873. doi: 10.1073/pnas.1520244113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wattanapanitch M., Damkham N., Potirat P., Trakarnsanga K., Janan M., U-Pratya Y., Kheolamai P., Klincumhom N., Issaragrisil S. One-step genetic correction of hemoglobin E/beta-thalassemia patient-derived iPSCs by the CRISPR/Cas9 system. Stem Cell Res. Ther. 2018;9:46. doi: 10.1186/s13287-018-0779-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watters K.E., Fellmann C., Bai H.B., Ren S.M., Doudna J.A. Systematic discovery of natural CRISPR-Cas12a inhibitors. Science. 2018;5138:1–8. doi: 10.1126/science.aau5138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wienert B., Wyman S.K., Richardson C.D., Yeh C.D., Akcakaya P., Porritt M.J., Morlock M., Vu J.T., Kazane K.R., Watry H.L. Unbiased detection of CRISPR off-targets in vivo using DISCOVER-Seq. Science. 2019;364:286–289. doi: 10.1126/science.aav9023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu W.Y., Lebbink J.H.G., Kanaar R., Geijsen N., van der Oost J. Genome editing by natural and engineered CRISPR-associated nucleases. Nat. Chem. Biol. 2018;14:642–651. doi: 10.1038/s41589-018-0080-x. [DOI] [PubMed] [Google Scholar]
- Xie N., Gong H., Suhl J.A., Chopra P., Wang T., Warren S.T. Reactivation of FMR1 by CRISPR/Cas9-Mediated deletion of the expanded CGG-repeat of the fragile X chromosome. PLoS One. 2016;11:e0165499. doi: 10.1371/journal.pone.0165499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu L., Yang H., Gao Y., Chen Z., Xie L., Liu Y., Liu Y., Wang X., Li H., Lai W. CRISPR/Cas9-Mediated CCR5 ablation in human hematopoietic stem/progenitor cells confers HIV-1 resistance in vivo. Mol. Ther. 2017;25:1782–1789. doi: 10.1016/j.ymthe.2017.04.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu C.L., Cho G.Y., Sengillo J.D., Park K.S., Mahajan V.B., Tsang S.H. Translation of CRISPR genome surgery to the bedside for retinal diseases. Front. Cell Dev. Biol. 2018;6:46. doi: 10.3389/fcell.2018.00046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Y., Jacoby E., Fry T.J. Challenges and opportunities of allogeneic donor-derived CAR T cells. Curr. Opin. Hematol. 2015;22:509–515. doi: 10.1097/MOH.0000000000000181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yin H., Song C.-q., Dorkin J.R., Zhu L.J., Li Y., Wu Q., Park A., Yang J., Suresh S., Bizhanova A. Therapeutic genome editing by combined viral and non-viral delivery of CRISPR system components in vivo. Nat. Biotechnol. 2016;34:328–333. doi: 10.1038/nbt.3471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yin C., Zhang T., Qu X., Zhang Y., Putatunda R., Xiao X., Li F., Xiao W., Zhao H., Dai S. In vivo excision of HIV-1 provirus by saCas9 and multiplex single-guide RNAs in animal models. Mol. Ther. 2017;25:1168–1186. doi: 10.1016/j.ymthe.2017.03.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yin H., Song C.-q., Suresh S., Wu Q., Walsh S., Rhym L.H., Mintzer E., Bolukbasi M.F., Zhu L.J., Kauffman K. Structure-guided chemical modification of guide RNA enables potent non-viral in vivo genome editing. Nat. Biotechnol. 2017;35:1179–1187. doi: 10.1038/nbt.4005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yin H., Song C.Q., Suresh S., Kwan S.Y., Wu Q., Walsh S., Ding J., Bogorad R.L., Zhu L.J., Wolfe S.A. Partial DNA-guided Cas9 enables genome editing with reduced off-target activity. Nat. Chem. Biol. 2018;14:311–316. doi: 10.1038/nchembio.2559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu C., Liu Y., Ma T., Liu K., Xu S., Zhang Y., Liu H., La Russa M., Xie M., Ding S., Qi L.S. Small molecules enhance CRISPR genome editing in pluripotent stem cells. Cell Stem Cell. 2015;16:142–147. doi: 10.1016/j.stem.2015.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu A.Q., Ding Y., Lu Z.Y., Hao Y.Z., Teng Z.P., Yan S.R., Li D.S., Zeng Y. TALENs-mediated homozygous CCR5Delta32 mutations endow CD4+ U87 cells with resistance against HIV1 infection. Mol. Med. Rep. 2018;17:243–249. doi: 10.3892/mmr.2017.7889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zelensky A.N., Schimmel J., Kool H., Kanaar R., Tijsterman M. Inactivation of Pol theta and C-NHEJ eliminates off-target integration of exogenous DNA. Nat. Commun. 2017;8:66. doi: 10.1038/s41467-017-00124-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhan T., Rindtorff N., Betge J., Ebert M.P., Boutros M. CRISPR/Cas9 for cancer research and therapy. Semin. Cancer Biol. 2019;55:106–119. doi: 10.1016/j.semcancer.2018.04.001. [DOI] [PubMed] [Google Scholar]
- Zhang J.H., Adikaram P., Pandey M., Genis A., Simonds W.F. Optimization of genome editing through CRISPR-Cas9 engineering. Bioengineered. 2016;7:166–174. doi: 10.1080/21655979.2016.1189039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao Z., Shi L., Zhang W., Han J., Zhang S., Fu Z., Cai J. CRISPR knock out of programmed cell death protein 1 enhances anti-tumor activity of cytotoxic T lymphocytes. Oncotarget. 2018;9:5208–5215. doi: 10.18632/oncotarget.23730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhen S., Hua L., Takahashi Y., Narita S., Liu Y.H., Li Y. In vitro and in vivo growth suppression of human papillomavirus 16-positive cervical cancer cells by CRISPR/Cas9. Biochem. Biophys. Res. Commun. 2014;450:1422–1426. doi: 10.1016/j.bbrc.2014.07.014. [DOI] [PubMed] [Google Scholar]
- Zhou C., Sun Y., Yan R., Liu Y., Zuo E., Gu C., Han L., Wei Y., Hu X., Zeng R. Off-target RNA mutation induced by DNA base editing and its elimination by mutagenesis. Nature. 2019;571:275–278. doi: 10.1038/s41586-019-1314-0. [DOI] [PubMed] [Google Scholar]
- Zischewski J., Fischer R., Bortesi L. Detection of on-target and off-target mutations generated by CRISPR/Cas9 and other sequence-specific nucleases. Biotechnol. Adv. 2017;35:95–104. doi: 10.1016/j.biotechadv.2016.12.003. [DOI] [PubMed] [Google Scholar]
- Zuo E., Sun Y., Wei W., Yuan T., Ying W., Sun H., Yuan L., Steinmetz L.M., Li Y., Yang H. Cytosine base editor generates substantial off-target single-nucleotide variants in mouse embryos. Science. 2019;364:289–292. doi: 10.1126/science.aav9973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zych A.O., Bajor M., Zagozdzon R. Application of genome editing techniques in immunology. Arch. Immunol. Ther. Exp. (Warsz) 2018;66:289–298. doi: 10.1007/s00005-018-0504-z. [DOI] [PMC free article] [PubMed] [Google Scholar]