

Nitrogen is the most limiting macronutrient for plant growth, and rhizobia are important bacteria for agriculture because they can fix atmospheric nitrogen and make it available to legumes through the establishment of a symbiotic relationship with their host plants. In this work, we studied the nitrogen fixation process in the microsymbiont Sinorhizobium fredii at the genome level. A metabolic model was built using genome annotation and literature to reconstruct the symbiotic form of S. fredii. Genes controlling the nitrogen fixation process were identified by simulating gene knockouts. Additionally, the nitrogen-fixing capacities of S. fredii CCBAU45436 in symbiosis with cultivated and wild soybeans were evaluated. The predictions suggested an outperformance of S. fredii with cultivated soybean, consistent with published experimental evidence. The reconstruction presented here will help to understand and improve nitrogen fixation capabilities of S. fredii and will be beneficial for agriculture by reducing the reliance on fertilizer applications.
KEYWORDS: bacteroid, metabolic model, nitrogen fixation, rhizobia, symbiosis
ABSTRACT
Rhizobia are soil bacteria able to establish symbiosis with diverse host plants. Specifically, Sinorhizobium fredii is a soil bacterium that forms nitrogen-fixing root nodules in diverse legumes, including soybean. The strain S. fredii CCBAU45436 is a dominant sublineage of S. fredii that nodulates soybeans in alkaline-saline soils in the Huang-Huai-Hai Plain region of China. Here, we present a manually curated metabolic model of the symbiotic form of Sinorhizobium fredii CCBAU45436. A symbiosis reaction was defined to describe the specific soybean-microsymbiont association. The performance and quality of the reconstruction had a 70% score when assessed using a standardized genome-scale metabolic model test suite. The model was used to evaluate in silico single-gene knockouts to determine the genes controlling the nitrogen fixation process. One hundred forty-one of 541 genes (26%) were found to influence the symbiotic process, wherein 121 genes were predicted as essential and 20 others as having a partial effect. Transcriptomic profiles of CCBAU45436 were used to evaluate the nitrogen fixation capacity in cultivated versus in wild soybean inoculated with the microsymbiont. The model quantified the nitrogen fixation activities of the strain in these two hosts and predicted a higher nitrogen fixation capacity in cultivated soybean. Our results are consistent with published data demonstrating larger amounts of ureides and total nitrogen in cultivated soybean than in wild soybean. This work presents the first metabolic network reconstruction of S. fredii as an example of a useful tool for exploring the potential benefits of microsymbionts to sustainable agriculture and the ecosystem.
IMPORTANCE Nitrogen is the most limiting macronutrient for plant growth, and rhizobia are important bacteria for agriculture because they can fix atmospheric nitrogen and make it available to legumes through the establishment of a symbiotic relationship with their host plants. In this work, we studied the nitrogen fixation process in the microsymbiont Sinorhizobium fredii at the genome level. A metabolic model was built using genome annotation and literature to reconstruct the symbiotic form of S. fredii. Genes controlling the nitrogen fixation process were identified by simulating gene knockouts. Additionally, the nitrogen-fixing capacities of S. fredii CCBAU45436 in symbiosis with cultivated and wild soybeans were evaluated. The predictions suggested an outperformance of S. fredii with cultivated soybean, consistent with published experimental evidence. The reconstruction presented here will help to understand and improve nitrogen fixation capabilities of S. fredii and will be beneficial for agriculture by reducing the reliance on fertilizer applications.
INTRODUCTION
Rhizobia are diazotrophic Gram-negative bacteria able to infect and form nitrogen-fixing nodules on compatible host plants (1). These soil bacteria are distributed across six genera: Rhizobium, Sinorhizobium/Ensifer, Bradyrhizobium, Allorhizobium, Azorhizobium, and Mesorhizobium (2). Symbiotic nitrogen-fixing bacteria provide a major source of nitrogen to plants, especially legumes. Most of the species within the family Leguminosae (Fabaceae) can establish a symbiotic association with rhizobia to obtain biological nitrogen (3–5). The interactions between the host plant and rhizobia occur in three stages: recognition and production of specific signal molecules by rhizobia (6), formation of root nodules and newly differentiated tissue in the roots of host plants (7, 8), and differentiation of rhizobia into their symbiotic form, bacteroids, which are able to reduce atmospheric nitrogen to ammonia inside the nodules (5, 9, 10). The symbiotic association between legume and rhizobium involves the release of ammonia to the plant cell cytosol by the nitrogen-fixing bacteroid in exchange for organic acids provided by the host plant. This symbiotic relationship contributes to nitrogen enrichment of the soil, improves plant growth, and reduces the need for chemical fertilizers (11). These features highlight the huge potential of exploiting this interaction for sustainable agriculture. However, the details and degree of the symbiosis are very strain specific and host specific (12), and deeper understanding of the interactions at the metabolic level is required to optimize it.
Among rhizobia, Sinorhizobium fredii is able to form nitrogen-fixing nodules with important legume food crops such as soybeans (Glycine max and Glycine soja), pigeon pea (Cajanus cajan), and cowpea (Vigna unguiculata) (13). In particular, S. fredii is a fast-growing soybean rhizobium and dominates in alkaline-saline soil in the Huang-Huai-Hai Plain region in China (14, 15). The S. fredii strain CCBAU45436 represents one of the dominant sublineages of S. fredii that nodulate soybeans in northern China (16). Recently, the complete genome sequence for CCBAU45436 was fully assembled (17). The CCBAU45436 multipartite genome consists of a chromosome (cSF45436, 4.16 Mb), a chromid (pSF45436b, 1.96 Mb), a symbiotic plasmid (pSF45436a, 0.42 Mb), and two accessory plasmids (pSF45436d, 0.20 Mb, and pSF45436e, 0.17 Mb). Evidence suggests a distinct role for each of these forms of DNA in rhizobial adaptations to symbiotic conditions (17–19). CCBAU45436 is able to induce effective nodules on the roots of the wild soybean accession W05 (Glycine soja W05) and the cultivated soybean accession C08 (Glycine max C08). Symbiotic nitrogen fixation by rhizobia in legume root nodules involves several simultaneous biological processes, including biological nitrogen fixation carried out by bacteroids, carbon-nitrogen metabolism, and exchange of nutrients and cofactors between the host plant and bacteroid (20). A better understanding of the nitrogen fixation process is required to facilitate the optimization of symbiotic nitrogen fixation to further increase legume productivity. The differential nitrogen-fixing capacity of the representative S. fredii strain, CCBAU45436, in symbiosis with Glycine max C08 versus with Glycine soja W05 (21) provides a unique opportunity to analyze the host-specific nitrogen-fixing metabolism of S. fredii. The constraint-based modeling approach has been successfully used to integrate genomic and high-throughput metabolomic data to systematically analyze the metabolic capabilities of other rhizobial species to get new insights into the complex process of symbiotic nitrogen fixation carried out in mature nodules (19, 22–25).
Here, we present a metabolic network reconstruction, iCC541, for the symbiotic form of the soybean microsymbiont, S. fredii CCBAU45436. This is the first manually curated reconstruction of the S. fredii bacteroid. The performance and quality of the reconstruction were examined using a standardized genome-scale metabolic model test suite (26). Consistency between model predictions and available experimental observations was evaluated. Flux balance analysis (27) was used to analyze the physiological capabilities of the bacterium under different symbiotic nitrogen fixation conditions. Symbiotic genes were identified and compared with experimental evidence. Finally, the transcriptome profiles of CCBAU45436 at the symbiotic stage within the nodules of cultivated (C08) and wild (W05) soybeans were used to evaluate the nitrogen fixation capacity of S. fredii bacteroids.
RESULTS
Formulation of metabolic network reconstruction of the symbiotic form of S. fredii CCBAU45436.
A metabolic reconstruction (iCC541) was developed to represent the symbiotic form of the microsymbiont S. fredii CCBAU45436. Metabolic reconstructions previously generated (19, 24) were used as a starting point, with 303 reactions selected in total from both models. Reactions used as the template are indicated in Table S1 in the supplemental material. The draft model was manually curated using information from literature, the S. fredii strain pathway database in Kyoto Encyclopedia of Genes and Genomes (KEGG) (28), automated reconstruction databases (29), and other available databases to ensure that it captures the biochemical and physiological knowledge available on nitrogen fixation. A schematic representation of the nutrient and cofactor exchanges between the host plant and the bacteroid is shown in Fig. 1.
FIG 1.

Schematic representation of main metabolic pathways and transport systems in soybean microsymbiont S. fredii CCBAU45436. Carbon sources in the plant cytosol are transported across the symbiosome and bacteroid membranes in exchange for fixed nitrogen. Malate and succinate enter the TCA cycle to be metabolized. Transport of cofactors required for nitrogen fixation is indicated. Dct, C4-dicarboxylate transport system; ala-L, alanine; asp-L, aspartate; OAA, oxaloacetate; AKG, 2-oxoglutarate; G3P, glyceraldehyde-3-phosphate; PHB, poly-3-hydroxybutyrate.
List of reactions included in the reconstruction model with their description, associated genes, and their reference numbers in various databases. Download Table S1, XLSX file, 0.1 MB (84.1KB, xlsx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
A symbiosis reaction was defined to integrate the essential and characteristic elements of the soybean-rhizobia symbiosis association. Specifically, the legume-rhizobium symbiosis determines the accumulation of metabolites inside the microsymbionts related to carbon and energy storage. S. fredii is able to nodulate different legumes and form determinate or indeterminate nodules depending on the host species (30). In particular, poly-3-hydroxybutyrate (PHB), glycogen, and lipids were included in the symbiosis reaction, as S. fredii has been shown to accumulate these compounds in determinate soybean nodules (31–34). The synthesis of storage compounds is important for managing the carbon source and reducing power supplied by the host plant. However, the role of carbon storage compounds in the symbiotic process differs depending on the rhizobium strain (12, 20). Furthermore, bacteroid development and nitrogen fixation are dependent on the amino acids supplied by the host plant (35, 36). Therefore, the uptake and export of various amino acids such as glutamate, alanine, aspartate, and arginine between the bacteroid and host plant were also considered in the symbiosis reaction. Bacteroids isolated from soybean nodules possess an active uptake system for glutamate supplied by the plant cytosol (37, 38). Experimental evidence shows that alanine and aspartate can be supplied to the plant by the bacteroid when malate is provided as a carbon source (39). Meanwhile, arginine also plays a role in the symbiotic process, since the decarboxylation products of arginine are the precursors of polyamines (40, 41). Legume root nodules can contain high levels of polyamines, and it plays a role in the regulation of nutrient exchange (42, 43). Transcriptome data have shown that genes associated with polyamine biosynthesis are upregulated in the symbiosis stage (44). We have also included valine and isoleucine in modeling the symbiotic capacity of S. fredii, because their biosynthetic pathways have been indicated as essential for effective symbiosis in Sinorhizobium and Rhizobium (45), and two transposon Tn5-induced isoleucine and valine auxotrophs of S. fredii HH303 were unable to form effective nodules on soybean as well (46). Moreover, nitrogenous base production has also been demonstrated to be essential for the formation of effective soybean nodules, and S. fredii purine and pyrimidine mutants form ineffective nodules in their hosts (42, 43, 46–49). Therefore, purine and pyrimidine metabolism also constitute a component of the reconstruction. However, symbiosis-defective mutants were not genetically characterized, and further experiments are required to explain this phenotype.
Additionally, symbiotic cofactors are key for the activity of nitrogenase and were therefore added to the symbiosis reaction. Nitrogenase is the enzyme responsible for the reduction of atmospheric nitrogen supplied by the host plant to ammonia. The nitrogenase complex consists of two enzymes: dinitrogenase reductase and dinitrogenase (5, 10). Dinitrogenase reductase is a dimeric iron (Fe) protein and dinitrogenase is a tetrameric iron-molybdenum (FeMo) protein. Nitrogenase activities depend on the availability of energy (ATP), low-potential electrons (iron-sulfur clusters), and microaerobic conditions (5, 50). One of these cofactors is homocitrate, which is a component of the iron-molybdenum cofactor of nitrogenase. From the genome sequence of CCBAU45436, no homocitrate synthase has been annotated; thus, the host plant has to provide homocitrate to the bacteroid to synthesize nitrogenase (51). MucR1 has been identified as a transcription regulator associated with the expression of ion transporters linked to phosphate, molybdenum, zinc, iron, and sulfur. Symbiotic defects were observed in S. fredii mucR1 mutants (52). Additional evidence highlights the role of phosphate in the nitrogen fixation process (33). After nitrogen, phosphate is the most limiting macronutrient, and a high demand for it has been reported in the nodules of nitrogen-fixing legumes (53). Heme and nicotinate are also synthesized by bacteroids, which are supplied to the houseplant for the synthesis of leghemoglobin to maintain the microaerobic condition for nitrogen fixation (36, 46, 47). Other cofactors include pyridoxine (19, 24), magnesium (54), and glutathione (55, 56). The symbiosis equation is a combination of the components described above and it is provided in Table S1. The element balance was checked, and the chemical formulae of the symbiotic product was determined to ensure a molecular weight (MW) of 1 g mmol−1 (57).
Evaluation of iCC541 reconstruction.
iCC541 describes 538 reactions encoded by 541 genes as well as 508 metabolites. Table 1 summarizes the main properties of iCC541. iCC541 was evaluated using three methods. First, the reconstruction was validated using experimental data reported in the literature to ensure the model is able to capture unique characteristics of the nitrogen fixation capacity of S. fredii. The conditions tested and captured by the model can be found in Table 2. Published experimental data represent the current knowledge status of an organism, and the validation procedure tests the biological accuracy of the reconstruction. Strain-specific metabolic capabilities included carbon sources and phenotypes of mutant strains.
TABLE 1.
iCC541 statistics
| Parameter | Value |
|---|---|
| No. of ORFsa | 541 |
| Chromosome (n [%]) | 441 (82) |
| Chromid (n [%]) | 81 (15) |
| Symbiotic plasmid (n [%]) | 11 (2) |
| Accessory plasmids (n [%]) | 7 (1) |
| Total reactions (n) | 535 |
| Metabolic conversions (n) | 491 |
| Reactions with ORF assignments (n) | 474 |
| % of reactions with ORF | 96 |
| Reactions without ORF (n) | 17 |
| % of reactions without ORF | 4 |
| Transport reactions (n) | 24 |
| Transport reactions with ORF assignments (n) | 17 |
| % of transport reactions with ORF | 71 |
| Exchange reactions (n) | 23 |
| No. of metabolites | 508 |
ORF, open reading frame.
TABLE 2.
Published experimental data used during the validation process
| Phenotype or metabolic characteristics | Reference(s) |
|---|---|
| Reduction of nitrogen to ammonia by nitrogenase reaction | 15 |
| Malate as carbon source | 86 |
| Succinate as carbon source | 86 |
| Alanine production by bacteroid | 64 |
| Production of PHB | 51 |
| pyrC mutant: Fix− | 42 |
| pyrF mutant: Fix− | 48 |
| C4-dicarboxylate requirement | 62 |
| purL mutant: Fix− | 47, 49 |
| purQ mutant: Fix− | 47 |
| idhA mutant: decrease nitrogen fixation | 66 |
| Glutamate requirement: enhances N2 fixation | 38 |
| znuA mutant: reduce nitrogen fixation | 17 |
| ptsSCAB mutant: reduce nitrogen fixation | 52 |
| cobO mutant: reduce number of nodules | 43 |
Additionally, the iCC541 formulation was assessed using a standardized genome-scale metabolic model test suite called memote (26). This metabolic model test approach provides a series of comparisons to benchmark metabolic models and facilitates quality control during model reconstruction. Scores are assigned to properties such as consistency (stoichiometry, mass and charge balance, and metabolite connectivity) and annotation (database cross-references and ontology terms). Scores represent the degree of completeness with respect to the tested property, with a maximum achievable score of 100%. An overall score of 70% was assigned to iCC541 by memote (Fig. 2a). Consistency, metabolites, reactions, genes, and systems biology ontology (sbo) annotations achieved scores of 85%, 68%, 69%, 33%, and 62%, respectively (Fig. 2b). The subsections under consistency, namely, stoichiometric consistency, mass balance, charge balance, and metabolite connectivity, were assigned scores of 100%, 99.2%, 96.9%, and 100%, respectively (Fig. 2c). A maximum score in each of these subsections is desirable to ensure reliable model predictions. These scores are comparable to those for well-curated models of Escherichia coli (E. coli core model and iAF1260b) and Saccharomyces cerevisiae (iMM904) from the Biochemical, Genetic, and Genomic (BiGG) knowledge base (58). iCC541 has significantly improved quality in terms of model consistency and annotations compared to that of the published Sinorhizobium meliloti reconstructions, iGD1575 and iHZ565 (25, 59), which were also tested by memote in this study using the published Systems Biology Markup Language (SBML) files (Fig. 2). The full report returned by memote for iCC541 and the two S. meliloti models can be found in Fig. S1.
FIG 2.

Scores and consistency assigned to iCC541, iHZ565, and iGD1575 by memote. (a) Total scores. (b) Scores by main categories: consistency and annotations (systems biology ontology [sbo], reactions, metabolites, and genes). (c) Consistency across subsections.
Snapshot of reports obtained from memote for comparison of iCC541 with S. meliloti models in HTML format. Model tests are divided in functionality libraries to evaluate core and organism-specific model properties. Core tests are independent of the modeled organism and complexity involved in the model formulation. Each test is divided into scored and unscored sections. The maximum achievable score is 100%. Unscored sections are related to general statistics and specific characteristics of some models such as the quality of the biomass reaction. Download FIG S1, PDF file, 0.5 MB (531.6KB, pdf) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Finally, the simulation performance of iCC541 was compared against the S. meliloti models. Inconsistencies reported by memote were solved in order to facilitate cross-comparisons between models. For that purpose, mass and charge imbalances and energy-generating cycles (ATP) (60) were fixed for the S. meliloti models. The symbiosis reactions were scaled to make sure the total mass of the material through the symbiosis reactions was the same in all three models in order to compare symbiosis yields (symbiosis product in grams per millimole carbon) (Table 3). The symbiosis reaction flux was maximized under microaerobic conditions for each model: 0.0107, 0.0145, and 0.0130 g mmol−1 were obtained for iCC541, iHZ565, and iGD1575, respectively. Similar yields relative to nitrogen uptake were observed for iCC541 and iGD1575: 0.0747 and 0.0906 g mmol−1 nitrogen, respectively, whereas iHZ565 predicts a higher yield with 0.1774 g mmol−1. C/N ratios obtained under standard nitrogen fixation conditions (defined according to literature) were comparable in iCC541 and iGD1575, with iHZ565 showing a lower ratio. The number of share reactions, metabolites, and genes between iCC541 and the S. meliloti models are indicated in Table 4.
TABLE 3.
Comparison of symbiosis yields for S. fredii (iCC541) and S. meliloti (iHZ565 and iGD1575)
| Model | Symbiosis yield (g product/mmol) on: |
C/N | |
|---|---|---|---|
| Carbon | Nitrogen | ||
| iCC541 | 0.0107 | 0.0747 | 0.1427 |
| iHZ565 | 0.0145 | 0.1774 | 0.1431 |
| iGD1575 | 0.0130 | 0.0906 | 0.0815 |
TABLE 4.
Comparison of iCC541 against iHZ565 and iGD1575
| Property | iCC541 (n) | No. (%)a
|
|
|---|---|---|---|
| Shared by iHZ565 | Shared by iGD1575 | ||
| Genes | 541 | 407 (75) | 400 (74) |
| Metabolic conversions | 491 | 461 (94) | 414 (84) |
| Metabolic conversions with same directionality | 491 | 455 (93) | 291 (59) |
| Transport reactions | 24 | 16 (67) | 16 (67) |
| Exchange reactions | 23 | 20 (87) | 23 (100) |
| Metabolites | 508 | 488 (96) | 434 (85) |
Percentage of coverage in iCC541.
Symbiotic pathway utilization analyses.
In symbiotic nitrogen fixation nodules, the host plant provides C4-dicarboxylic acids (malate or succinate) to bacteroids as the major energy source, while the microsymbiont returns organic fixation products to the host. The primary carbon source, malate, is transported into bacteroids to be metabolized through the tricarboxylic acid (TCA) cycle to provide reducing equivalents, energy, and precursors for biosynthetic pathways (12, 61, 62). The TCA cycle together with oxidative phosphorylation generates the energy and conditions required for the nitrogen fixation process. Oxygen levels must be maintained at the minimum to avoid inhibition of the nitrogenase activity, since it acts as a regulator of nitrogen fixation genes (nif and fix genes) (12, 20, 63). To examine the extent to which the model can capture these metabolic characteristics, flux balance analysis (FBA) was performed by maximizing the symbiosis reaction with malate as the primary carbon source and succinate and inositol as secondary carbon sources, with a small amount of oxygen available (maximum uptake rates being constrained by experimental data) (see Table S1). Figure 3 describes the flux distribution predicted by the model. In detail, the simulated exchange profile of the model shows the active uptake of glutamate and cofactors (including homocitrate, molybdenum, thiamine, cobalamin, zinc, iron, myo-inositol, phosphate, magnesium, and sulfate) as expected from the formulation of the symbiosis reaction (62, 64, 65). Oxygen is utilized by cytochrome oxidase to generate ATP. Ammonia, alanine, and aspartate are transported to the plant cell cytosol (39, 87). In addition, the predicted internal flux distribution describes the metabolic route of malate. Malate is actively transported into the cell and converted to pyruvate and CO2 via malate dehydrogenase. Then, pyruvate is decarboxylated by pyruvate dehydrogenase to form acetyl coenzyme A (acetyl-CoA) and enters the TCA cycle. The full oxidative TCA cycle is utilized by the bacteroid during the process, according to the model predictions (nonzero fluxes confirmed by flux variability analysis). Enzymes participating in the TCA cycle appear to have an active role in the nitrogen fixation process in S. fredii. TCA cycle mutants of S. meliloti, Rhizobium tropici, and Rhizobium leguminosarum have the common phenotype of being unable to fix nitrogen (68–70). The model, predicting the use of malate to produce energy and products for symbiosis through the TCA cycle, is therefore able to capture this important aspect of rhizobium-legume symbiosis. However, the role of the TCA cycle varies among rhizobia, with high metabolic plasticity observed in Bradyrhizobium japonicum (71, 72). PHB biosynthesis and gluconeogenesis pathways are active according to the simulation results and the definition of the symbiosis reaction. The use of these pathways is validated by the fact that both of these carbon storage compounds have been found to be present in determinate nodules of soybean (31–34). The model also predicts a low but nonzero flux through some reactions of the pentose phosphate pathway. This pathway generates ribose 5-phosphate, which is a precursor of purines and pyrimidines. Reduced symbiotic properties have been observed in pyrimidine and purine mutants of S. fredii (42, 43, 46–49). According to our reconstruction formulation and in silico results, the ammonia exported to the plant cell cytosol originates from the ammonia produced by the nitrogenase reaction (Fig. 3).
FIG 3.

Flux distribution during symbiotic nitrogen fixation in S. fredii. Reactions are colored according to a scale of purple (flux of 0.04 mmol g−1 [dry weight] h−1) to red (flux of 1.8 mmol g−1 [dry weight] h−1 or greater). Reactions carrying zero flux or less than 0.04 mmol g−1 (dry weight) h−1 are colored gray. Colored arrowheads indicate the directionality of the flux. Flux metabolic networks were constructed using the Web tool Escher.
Identification of genes involved in symbiosis.
Next, the model was used to identify symbiotic genes under standard nitrogen fixation conditions. As defined by the gene-protein-reaction (GPR) association, a gene was classified as a symbiotic gene if one or more reactions associated with that gene need to carry a nonzero flux to satisfy the symbiotic objective function. Results of simulations were categorized according to the effect of the gene deletion compared to that of the wild-type strain (see Table S2). According to the simulations, 141 of 541 genes (26%) were predicted to affect the symbiotic process, wherein 121 genes were classified as essential and 20 genes as having a partial effect, where their deletions resulted in a reduction in the capacity of S. fredii to fix atmospheric nitrogen. Most of these genes (84%) were located in the chromosome, while the others were present in the symbiosis plasmid, accessory plasmids, and chromid (Table S2). Coexpression patterns in the replicon of S. fredii strains have been observed under both free-living and symbiotic conditions (17).
Predicted symbiotic rate ratios between single-gene deletion mutants and wild-type strain. Download Table S2, XLSX file, 0.1 MB (40KB, xlsx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Many genetic studies have been conducted to examine the legume-rhizobium symbiosis, providing important insights into the symbiotic capacity of rhizobia. A detailed comparative genomic study of rhizobia that nodulate soybean was used to validate the predicted symbiotic genes from our model (16). Genes predicted to affect the symbiotic process were compared to a list of known symbiosis genes. The analysis matched 50 of the 141 predicted symbiotic genes to previously identified genes in the literature. Based on our model, mutations in these genes had a negative impact on the symbiotic phenotypes of rhizobial strains. For example, symbiotic genes related to oxidative phosphorylation were identified. According to the simulation in our model, oxidative phosphorylation must be active in vivo to effectively fix nitrogen, as previously identified by pathway analysis (63). A complete list of these genes and the references providing the corresponding validating evidence are available in Table S3. Among the 91 remaining predicted symbiotic genes, 58 are orthologs to essential genes identified by iHZ565 (24). These genes have already been listed in the Database of Essential Genes (DEG) (73) and therefore have been shown to perform core functions that are required under both free-living and symbiotic conditions. The remaining 33 genes have not been shown to be essential to the symbiotic process. Their locations and predicted functions are provided in Table S2. Most of these genes are linked to the purine, pyrimidine, TCA, and amino acid pathways. One example is the gene AB395_00003474 (sucB). sucB and sucA encode subunits of the α-ketoglutarate dehydrogenase which catalyzes the conversion of α-ketoglutarate to succinyl-CoA in the TCA cycle. R. leguminosarum mutants form ineffective nodules (69). However, B. japonicum sucA mutants only showed a delay in the development of bacteroids, but their nitrogen fixation rates were similar to those of the wild type (72). On the other hand, AB395_00003483, which encodes a succinate dehydrogenase flavoprotein subunit, was predicted here to be a symbiotic gene. A gene encoding succinate dehydrogenase A (sdhA) has been identified as essential in Bradyrhizobium sp. strain ORS278 (74). Another symbiotic gene predicted by our model, AB395_00004794, is involved in the isoleucine and valine biosynthesis pathway. Tn5 experiments have successfully identified this pathway as being related to the symbiotic process in S. fredii HH303, but the genes have not been characterized (46). The predicted gene is an ilv gene that encodes a dihydroxy-acid dehydratase. A BLASTP search of the amino acid sequence of the dihydroxy-acid dehydratases from S. meliloti 1021 (NP_384214, NP_436655, and NP_386934) against the S. fredii genome revealed identities of 89.37%, 64%, and 31.67%, respectively, with AB395_00004794 at the amino acid level. Mutations of ilv genes have been reported to result in phenotypes ranging from no nodule formation (Nod−) (35) to being unable to fix nitrogen (Nod delayed, Fix−) (75). In addition, genes related to the transport of cofactors such as sulfate were also predicted as novel symbiotic genes by our model. Additional experiments are required to confirm the functions and relevance of these genes in the TCA cycle, branched-amino-acid biosynthesis, and transport systems to the symbiotic processes in S. fredii. As stated above, myo-inositol uptake was active according to the model predictions. Simulations showed a slight reduction in the fixation rate, and shadow price analyses indicated it was a rate-limiting substrate. This is qualitatively but not quantitatively consistent with a previous finding that a mutation in the myo-inositol dehydrogenase gene (idhA) caused a much more significant reduction in the nitrogen fixation rate (66). Then, false-negative cases were evaluated. These cases represent discrepancies with experimentally observed phenotypes and indicate potential errors or knowledge gaps in the model. The symbiotic genes predictions exhibited 45 (8%) false negatives. The list of these genes can be found in Table S4. Analysis of these genes revealed active roles in processes such as nodulation, bacteroid development, microoxic respiration regulation, osmoregulation, and the inositol pathway. Regulation processes and steps previous to the symbiotic stage (e.g., root nodule formation) were not included in the current scope of the model, which explains this result. Additional false-negative cases occurred with knockout of genes involved in inositol catabolism, including iolA (AB395_0000388), iolC (AB395_00002655), and iolD (AB395_00002654) (76, 77). Shadow price analyses identified inositol as a rate-limiting step as stated above. pstC (AB395_0000141) and pit (AB395_00003896) involved in phosphate transport were classified as nonessential (78). FBA may use any transport reaction in the network to fix nitrogen under the standard nitrogen fixation condition. Additional constraints are required to simulate phosphate transport via high- and low-affinity systems.
List of predicted genes that influence symbiosis and the corresponding references for the associated experimental evidence. Download Table S3, DOCX file, 0.1 MB (38.9KB, docx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
List of false-negative predicted genes by the model. Download Table S4, DOCX file, 0.1 MB (35.7KB, docx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Nitrogen fixation capacity of S. fredii bacteroids in wild versus in cultivated soybean.
Finally, transcriptome profiles were used to determine the nitrogen fixation capacity of CCBAU45436 with two different soybean cultivars, the wild soybean accession W05 and the cultivated soybean accession C08. Condition-specific models were built from iCC541 using the E-Flux approach (79). Gene expressions were used to constrain reaction flux bounds to assess the impact of the host plant on nitrogen fixation. Tables S5 and S6 list the flux ranges for each condition and raw expression data, respectively. Flux distributions are shown in Fig. 4. Exchange profiles of the models showed carbon and essential nutrient requirements. Additionally, certain secretion patterns overlap one another. As expected, malate is transported inside the cell and pyruvate is generated by the reaction catalyzed by malic enzyme. Pyruvate is further decarboxylated by pyruvate dehydrogenase to form acetyl-CoA. In both soybean cultivars, soybean bacteroid diverts acetyl-CoA into the TCA cycle and into the production of the lipid-like polymer PHB, both of which are pathways known for electron allocation in nitrogen-fixing bacteroids (80). Although similitudes are observed in the flux distributions, there are also differences in the use of some reactions and/or their magnitudes (Fig. 4c). To fully assess the impact on nitrogen fixation capacity, yields were calculated based on the model predictions (Table 5); 0.0032 and 0.0097 g symbiosis product per mmol of carbon uptake were obtained in the symbiosis with W05 and C08, respectively. Additionally, there was a difference in the flux of fixed ammonia with 0.0038 and 0.0275 mmol g−1 (dry weight) h−1 transported from the bacteroid to W05 and C08, respectively. Simulations predicted a higher nitrogen fixation capacity of CCBAU45436 with C08 than with W05. C/N ratios of 0.1435 and 0.0426 were calculated for the C08 and W05 symbiotic conditions, respectively. The ammonia produced by nitrogen fixation in bacteroids is transported back to the host plant and assimilated into glutamate and glutamine by glutamine synthetase and glutamate synthase (20). In determinate nodules, such as those in soybean roots, glutamine enters the purine biosynthetic pathway to be further converted to ureides, allantoin, and allantoic acid (65, 81). Ureides, products of nitrogen fixation, are then transported from the root to the shoot. Thus, a higher ureides and nitrogen content should be expected in the root nodules of C08 compared to those of W05 when soybean is inoculated with CCBAU45436. Ureides and nitrogen content in root nodules are commonly used as an indicator of nitrogen fixation capacity and were previously investigated by our group using the same symbiotic conditions, with W05 and C08 being inoculated with S. fredii CCBAU45436. The results corroborate this model prediction (21). Experimental observations showed cultivated soybean outperformed wild soybeans on all the tested nitrogen fixation-related traits, in particular, the total nitrogen and total ureides accumulation (21). An imbalance in carbon and nitrogen metabolism in wild soybean was suggested as the cause of the lower capacity for nitrogen fixation. This experimental evidence validates the model predictions and indicates a consequence of the process of domestication and human selection in soybean. Furthermore, a simple evolutionary sensitivity analysis was performed to identify possible reactions (Fig. 4c) responsible for the higher performance of S. fredii with the cultivated soybean, by starting from the W05 condition (i.e., the model with flux bounds inferred from the E-Flux method used in the previous analysis), iteratively imposing bounds for the C08 condition randomly and fixing the bounds that increase nitrogen fixation until no further increase could be achieved (Fig. 5a). Across 10,000 simulations, different activity levels in the inositol pathway, cytochrome oxidase reaction, and iron transport were consistently identified to be the determinants for the predicted phenotypes (Fig. 5b). The inositol pathway is a lower-yielding pathway having a marginal effect on improving nitrogen fixation under the W05 condition (increased by 20%), while the cytochrome oxidase reaction appears to be a major bottleneck. However, a gene encoding an ATP-binding cassette (ABC)-type metal ion transport system (AB395_00003450) seemed to limit the nitrogen fixation rate under the C08 condition because of its lower expression level under the C08 condition than the W05 condition. Potential explanations for this include the existence of regulatory or resource allocation constraints that are not accounted for by the model. The proportionality assumption between maximum enzymatic activities and transcript levels applied to the model can serve as a proxy for estimating relative activities across conditions in the absence of additional data, but it is probably an oversimplification and may underestimate the activity of the metal ion transport reaction under the C08 condition or vice versa. Further in vitro and in vivo testing of these potential targets will help resolve this. Overall, a fine tuning of the energy, reducing factors, and regulations of gene expression are required to optimize the nitrogen fixation process.
FIG 4.


Nitrogen fixation capacity of S. fredii CCBAU45436 in wild soybean accession W05 (a) and cultivated soybean accession C08 (b). (c) The difference between the flux distributions of C08 and W05. Flux metabolic networks were constructed using the Web tool Escher. Reactions are colored according to a scale of purple (median flux or greater) to red (maximum flux). Colored arrowheads indicate the directionality of the flux. Reactions carrying zero flux or less than the median are colored gray.
TABLE 5.
Comparison of nitrogen fixation capacity of S. fredii with W05 and C08 soybeans
| Condition | Symbiosis flux yield (g mmol−1 carbon) | Fixed NH3 (mmol g−1 [dry wt] h− 1) | C/N |
|---|---|---|---|
| W05 | 0.0032 | 0.0038 | 0.0426 |
| C08 | 0.0097 | 0.0275 | 0.1435 |
FIG 5.
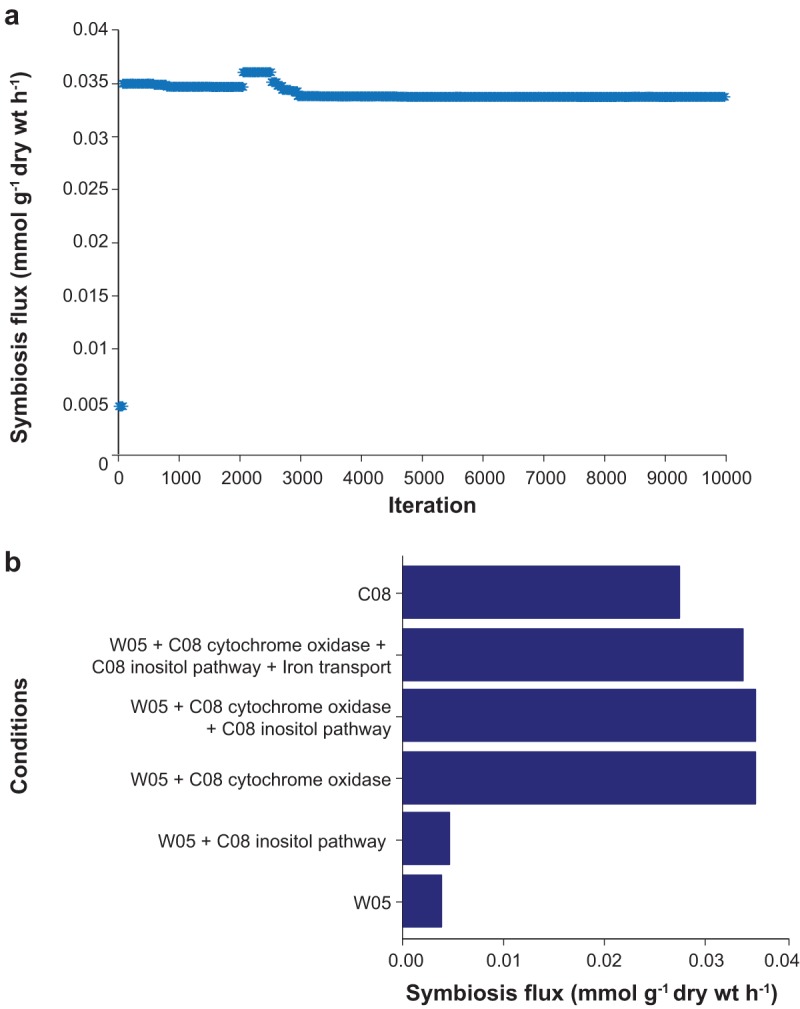
Evolutionary sensitivity analysis. (a) Symbiosis fluxes were determined by simulations starting from the W05 condition where bounds of the C08 condition were randomly imposed. (b) Reactions were identified where S. fredii inoculated in cultivated soybean outperformed that in wild soybean in nitrogen fixation.
Upper and lower boundaries of each reaction in iCC541 of bacteroids isolated from nodules of G. soja (W05) and G. max (C08) as determined by gene expression data. Download Table S5, XLSX file, 0.1 MB (31.3KB, xlsx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Average RPKM values of each gene in iCC541 under both symbiotic conditions (bacteroids isolated from G. soja W05 and G. max C08). Download Table S6, XLSX file, 0.1 MB (39.3KB, xlsx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
DISCUSSION
In this study, we developed a metabolic model for the fast-growing Sinorhizobium fredii strain CCBAU45436. This represents the first metabolic reconstruction of the symbiotic form of S. fredii for exploring the metabolic capabilities of this strain in symbiosis with soybean, one of the most important food crops worldwide. This legume-rhizobium association can fix atmospheric nitrogen, which can be a great benefit to sustainable agriculture. A symbiosis equation was formulated to capture and describe the unique physiological characteristics of this interaction and the symbiotic nitrogen fixation. Each host plant-rhizobium interaction as well as their metabolic interactions are association specific. Extensive literature reviews must be conducted to include the distinctive characteristics that define a specific interaction, such as amino acids, nitrogenous bases, and cofactor requirements, since general rules have proved to be not universally applicable. All the current metabolic models available for rhizobia define a distinct symbiosis reaction. The establishment of this reaction is relevant to the accurate description of the biological processes, and new reconstructions should revise, adapt, or propose metabolic requirements as this area of research progresses. The symbiosis reaction must describe symbiotic phenotypes supporting nitrogen fixation, and the reaction should be modified if new experimental evidence shows that the phenotypes are a consequence of the rhizobia being unable to differentiate or infect the host plant. The symbiosis reaction of S. fredii described in this model, iCC541, allowed us to mimic symbiotic nitrogen fixation and bacteroid physiology in a way that is consistent with experimental data. Specifically, our model predicts the utilization of important pathways involved in nitrogen fixation, such as the TCA cycle, oxidative phosphorylation, gluconeogenesis, and PHB. Additionally, the effects of single-gene deletions on nitrogen fixation helped us identify the essential genes, with 77% of them in agreement with experimental evidence. Thirty-three novel symbiotic genes were predicted and can be useful as targets for selecting better performing rhizobium strains. False negatives gave insight about the missing regulatory factors and scope of the reconstruction. Functional differences can be identified using metabolic networks by examining cellular metabolism under different conditions. Taking advantage of the flexibility to integrate high-throughput data in the constraint-based modeling approach, transcriptomic data were used to assess nitrogen fixation capacity in wild and cultivated soybean populations inoculated with S. fredii CCBAU45436. Condition-specific models were used to predict and analyze symbiotic capabilities. The contextualized models predicted a higher performance of cultivated soybean compared to that of wild soybean under symbiosis. In addition, the inositol pathway and cytochrome oxidase reaction were identified as potential targets for strain design experiments. Evidence indicates that artificial selection during the domestication of soybean had a key role in the performance of bacteroids (21). In the future, new findings can be used to modify the reconstruction and adapt it to better represent the microsymbiont, such as experimental assessment of the ATP requirements for maintaining the bacteroid to further tune the model. Proteomic data of the S. fredii bacteroid would also help to complement and further improve the predictions made here with the model.
iCC541 only covers the metabolism of the bacteroid, the differentiated form of S. fredii, which is able to fix biological nitrogen for the host plant. As the next step, iCC541’s metabolic pathways should be extended to represent the free-living form of the rhizobium to study other niche conditions. Bulk soil and rhizosphere are part of the life cycle of rhizobia, and they must be colonized to induce nodule formation and allow the final differentiation of rhizobia. All current metabolic reconstructions for rhizobia describe the symbiotic nitrogen fixation process with the subsequent metabolic exchange between the microsymbiont and its host plant (19, 22–25). However, only two of the reconstructions available on rhizobia cover the full spectrum of metabolism. For example, phenotypic microarrays have been successfully used to test the metabolic capabilities of rhizobia at the free-living stage (19, 25). Expanding the scope of the model will require additional experimental data for validation.
The number of metabolic reconstructions is increasing as well as the number of their applications (82). At present, no minimum standards for publishing metabolic reconstructions have been clearly defined. This makes it difficult to assess quality and undermines reproducibility and transferability. New tools have been developed in the modeling community to solve this problem, and their use should be encouraged during the peer review process (26). This requirement will help to improve the usage of available models by both experts and nonexperts alike, which is one of the main goals for the development of new reconstructions. Also, the use of these tools will help with the curation of larger reconstructions. A snapshot report of our S. fredii reconstruction has been included in this study to help future users to understand, adapt, or improve the current reconstruction.
In summary, iCC541 provides a framework for studying the symbiotic nitrogen fixation capabilities of S. fredii within different host plants.
MATERIALS AND METHODS
Metabolic network reconstruction.
To create a metabolic reconstruction to represent the soybean microsymbiont S. fredii, the genome sequence of CCBAU45436 was used to identify orthologs. The genomes of six closely related rhizobial strains with high-quality draft genomes were selected to build an ortholog table. Ortholog predictions of rhizobia against S. fredii CCBAU45436 were identified by comparing two proteomes using KBase narrative interface methods (83). A minimum suboptimal best bidirectional hit (BBH) ratio of 90% was used. The ortholog table is available in Table S7 in the supplemental material.
List of orthologs of predicted symbiosis-related soybean genes used to build iCC541. The values represent best bidirectional hits (BBHs) percent. Download Table S7, XLSX file, 0.4 MB (422.1KB, xlsx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Reactions from previously generated metabolic reconstructions of Sinorhizobium meliloti (19, 24) were added to iCC541 (model in this study) according to the ortholog table. Only reactions from iGD1575 (published model constructed on S. meliloti) associated with the symbiotic process were considered (19). Biomass and associated reactions of iGD1575 were not considered. Reactions associated with missing orthologs were not included in the preliminary draft. Gene-protein-reaction (GPR) associations were rewritten for each reaction based on the identified orthologs. GPR associations provide a link between different levels of information. Specifically, they indicate which protein is encoded by each gene and link the functional proteins to one or more enzymatic reactions. Additional reactions and gene contents from the Kyoto Encyclopedia of Genes and Genomes (KEGG) database (28) were added using the ortholog table. KEGG has no information related to S. fredii CCBAU45436, but it contains other rhizobium strains such as S. fredii NGR234 and HH103. In addition, reactions from a draft model automatically generated using KBase (29) were evaluated and added to the reconstruction. The poly-3-hydroxybutyrate (PHB) synthesis pathway was extracted from the iOR363 model constructed on Rhizobium etli (22). The draft reconstruction was manually inspected, curated, and validated to include main metabolic pathways related to the stages of symbiotic nitrogen fixation and to permit the computation of steady-state properties. An established reference protocol for the reconstruction of metabolic networks was followed throughout the process to maintain the consistency of the model formulation (67).
Literature and databases (e.g., KEGG, UniProtKB, Brenda, and NCBI) were used to obtain information for gene annotations, carbon-nitrogen pathway utilization, and other physiological and phenotypic properties during the stages of nitrogen fixation. GenBank locus tags were used as gene identifiers (IDs). Biochemical, Genetic, and Genomic (BiGG) and MetaNetX databases were used to standardize metabolite identifiers (58, 84). Lists of the reactions and metabolites included in the reconstruction are available in Tables S1 and S8. All genome sequences were downloaded from GenBank (85) on 20 July 2018. A published protocol was used to guarantee mass and charge balances (57). Briefly, the element balance of internal reactions was checked, including that of the symbiosis reaction. Inconsistencies were solved by finding the set of minimal formulae to solve the metabolites with unknown molecular groups and the metabolites with generic side groups. Elemental hydrogen balances were achieved through the addition of missing protons. ATP cycles were solved by checking the reversibility constraints.
List of metabolites included in the reconstruction of iCC541, with their corresponding charges, chemical formulae, and reference IDs in various databases Download Table S8, XLSX file, 0.1 MB (51KB, xlsx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Symbiosis reaction.
A symbiosis reaction was designed to represent the specific legume-rhizobium association studied in this work. Essential components were determined from the literature, and their fractional contributions were adapted from a previously published study (24). A biomass equation was not added to the reconstruction, since growth-associated pathways are switched off in nitrogen-fixing bacteroids (33). The symbiosis reaction and publications supporting its formulation are provided in Tables S1 and S9, respectively. The chemical formulae of symbiotic components were scaled to ensure that the symbiotic product produced by the model has a molecular weight (MW) of 1 g mmol−1 (57).
List of references used to formulate the symbiosis reactions. Download Table S9, DOCX file, 0.1 MB (43.3KB, docx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
In silico symbiotic conditions.
Flux balance analysis (FBA) simulations (27) were performed to predict nitrogen fixation rates and metabolite productions by S. fredii in soybean nodule cells. The symbiosis equation was used as the objective function of the optimization problem assuming steady-state conditions. Predictions were matched qualitatively to the experimental results reported in the literature.
Legume-rhizobium symbiosis involves the exchange of metabolites and cofactors to obtain biological nitrogen. Metabolism and transport in bacteroids have been reviewed in detail (20, 51). The exchange of nutrients was simulated by allowing an unlimited uptake of phosphate, sulfate, cobalamin, molybdate, thiamine, iron, N2, H+, H2O, Mg2+, and Zn2+. The lower bounds of the respective exchange reactions were set to −1,000 mmol g−1 (dry weight) h−1. Uptake rates of C4-dicarboxylate acids supplied by the host plant, i.e., malate and succinate, were set to 1.44 mmol g−1 (dry weight) h−1 and 1.38 mmol g−1 (dry weight) h−1, respectively (86). Oxygen consumption rate was limited to 1.26 mmol g−1 (dry weight) h−1 (87) to mimic the low-oxygen environment provided by root nodule cells (50). Lower bounds of glutamate and inositol were set according to a reference study (19). A flux of nongrowth-associated ATP maintenance (ATPm) was fixed at 4 mmol g−1 (dry weight) h−1 (25). Ammonia generated by the reaction catalyzed by the nitrogenase enzyme is exported to the plant cytoplasmic compartment (19). Nutrient and essential cofactor exchanges between the host plant and the bacteroid are schematically represented in Fig. 1.
The dual problem of the linear optimization described above was solved to determine the dual variables or shadow prices (88). Shadow prices measure the sensitivity of the objective function to changes in the availability of each metabolite. The symbiosis reaction was used as the objective function.
For the identification of symbiotic genes, in silico single-gene knockout simulations were performed by setting bounds of the corresponding reaction(s) to 0 mmol g−1 (dry weight) h−1 if a single gene was associated with one or multiple reactions (89). Reactions catalyzed by multiple noninteracting genes were not removed by a single-gene deletion. A symbiotic rate ratio between the deletion strain and wild type was computed to identify the set of essential genes for nitrogen fixation. A gene was classified as a symbiotic gene if the symbiotic rate ratio was less than the cutoff set at 0.05 (90). All simulations were performed using the MATLAB-based COBRA Toolbox (82). Gurobi Optimizer (v8.1.0) was employed as a linear programming solver. Escher was used for the visualization of the reconstruction and analysis (91). The E. coli core map was used as a template to build a map for S. fredii central metabolism. A model in JSON format was generated using COBRApy (92).
Condition-specific models.
To evaluate the capacity of CCBAU45436 to fix atmospheric nitrogen in the nodules of different soybean cultivars, a transcriptome data set was incorporated into the analysis. Raw transcriptome sequencing (RNA-seq) data of S. fredii CCBAU45436 bacteroid at the symbiotic stage within the nodules of cultivated soybean C08 and wild soybean W05 were retrieved from a previous publication for reanalysis (17). In brief, the clean reads in fastq files were mapped to the reference genomes of S. fredii CCBAU45436 using HISAT2 (default parameters) (93). The counting of mapped reads for protein-coding genes from the sam files were performed by HTseq-count (94). The read counts were normalized with DESeq2 using the medians-of-ratios method (95). Gene expression data were preprocessed, and gene scores were calculated according to transcriptomic data integration methods (96, 97). The expression data were then used to tailor the metabolic reconstruction using the E-Flux approach (79). Gene scores were mapped to the model by parsing the GPR rules associated with each reaction (98). No gene expression data and orphan reactions were given an expression value of −1. Lower and upper bounds of the reactions were determined by flux variability analysis (99). The minimum percentage of the optimal solution was set to zero. Under each condition, i.e., W05 and C08, the minimum values between expression reactions were identified and bounds were scaled down by the corresponding expression ratio between W05 and C08 (100).
Flux distributions of bacteroids within the nodules of C08 and W05 were compared by simultaneously solving two FBA problems and minimize the 1-norm of the flux vectors in the MATLAB-based COBRA Toolbox (82). An evolutionary sensitivity analysis was performed to evaluate these flux distributions and identify which gene expression changes are critical under these two conditions. The condition with the lower yield (W05) was selected as the starting point (i.e., model imposed with the flux bounds inferred using the aforementioned E-Flux method). A reaction was randomly selected, and the flux bound was changed randomly toward the bound for the same reaction under the condition with higher yield (C08) until the resultant yield stabilized and reached the level under condition C08.
Data availability.
The reconstruction, memote reports, and scripts are available at https://github.com/cacontad/SfrediiScripts.
ACKNOWLEDGMENTS
We thank Min Xie and Man-Wah Li of CUHK for their constructive suggestions for the manuscript. Jee Yan Chu copyedited the manuscript.
This work was supported by the Hong Kong Research Grants Council General Research Fund (14108014) and Area of Excellence Scheme (AoE/M-403/16), CUHK VC Discretionary Fund VCF2014004, and the Lo Kwee-Seong Biomedical Research Fund to H.-M.L.
REFERENCES
- 1.Rosenberg E, DeLong E, Lory S, Stackebrandt E, Thompson F. 2014. The prokaryotes, 4th ed Springer-Verlag, Berlin, Germany. [Google Scholar]
- 2.Moreira F, Siqueira J. 2006. Microbiologia e bioquímica do solo, p 488 UFLA, Lavras, Brazil. [Google Scholar]
- 3.Andrews M, Andrews ME. 2017. Specificity in legume-rhizobia symbioses. Int J Mol Sci 18:E705. doi: 10.3390/ijms1804070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Sprent JI, Ardley J, James EK. 2017. Biogeography of nodulated legumes and their nitrogen-fixing symbionts. New Phytol 215:40–56. doi: 10.1111/nph.14474. [DOI] [PubMed] [Google Scholar]
- 5.Rascio N, La Rocca N. 2013. Biological nitrogen fixation, p 1–16. Reference module in earth systems and environmental sciences. Elsevier B.V, Amsterdam, Netherlands. [Google Scholar]
- 6.Spaink HP. 2000. Root Nodulation and infection factors produced by rhizobial bacteria. Annu Rev Microbiol 54:257–288. doi: 10.1146/annurev.micro.54.1.257. [DOI] [PubMed] [Google Scholar]
- 7.Timmers ACJ, Soupene E, Auriac M-C, de Billy F, Vasse J, Boistard P, Truchet G. 2000. Saprophytic intracellular rhizobia in alfalfa nodules. Mol Plant Microbe Interact 13:1204–1213. doi: 10.1094/MPMI.2000.13.11.1204. [DOI] [PubMed] [Google Scholar]
- 8.Vasse J, De Billy F, Camut S, Truchet G. 1990. Correlation between ultrastructural differentiation of bacteroids and nitrogen fixation in alfalfa nodules. J Bacteriol 172:4295–4306. doi: 10.1128/jb.172.8.4295-4306.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Poole P, Ramachandran V, Terpolilli J. 2018. Rhizobia: from saprophytes to endosymbionts. Nat Rev Microbiol 16:291–303. doi: 10.1038/nrmicro.2017.171. [DOI] [PubMed] [Google Scholar]
- 10.Rubio LM, Ludden PW. 2008. Biosynthesis of the iron-molybdenum cofactor of nitrogenase. Annu Rev Microbiol 62:93–111. doi: 10.1146/annurev.micro.62.081307.162737. [DOI] [PubMed] [Google Scholar]
- 11.Herridge DF, Peoples MB, Boddey RM. 2008. Global inputs of biological nitrogen fixation in agricultural systems. Plant Soil 311:1–18. doi: 10.1007/s11104-008-9668-3. [DOI] [Google Scholar]
- 12.Terpolilli JJ, Hood GA, Poole PS. 2012. What determines the efficiency of N2-fixing Rhizobium-legume symbioses? Adv Microb Physiol 60:326–389. [DOI] [PubMed] [Google Scholar]
- 13.Brenner DJ, Krieg NR, Staley JT. (ed) 2005. Bergey’s manual of systematic bacteriology. Vol 2: the Proteobacteria. Second ed. Springer, New York, NY. [Google Scholar]
- 14.Han LL, Wang ET, Han TX, Liu J, Sui XH, Chen WF, Chen WX. 2009. Unique community structure and biogeography of soybean rhizobia in the saline-alkaline soils of Xinjiang, China. Plant Soil 324:291–305. doi: 10.1007/s11104-009-9956-6. [DOI] [Google Scholar]
- 15.Zhang YM, Li Y, Chen WF, Wang ET, Tian CF, Li QQ, Zhang YZ, Sui XH, Chen WX. 2011. Biodiversity and biogeography of rhizobia associated with soybean plants grown in the North China Plain. Appl Environ Microbiol 77:6331–6342. doi: 10.1128/AEM.00542-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Tian CF, Zhou YJ, Zhang YM, Li QQ, Zhang YZ, Li DF, Wang S, Wang J, Gilbert LB, Li YR, Chen WX. 2012. Comparative genomics of rhizobia nodulating soybean suggests extensive recruitment of lineage-specific genes in adaptations. Proc Natl Acad Sci U S A 109:8629–8634. doi: 10.1073/pnas.1120436109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Jiao J, Ni M, Zhang B, Zhang Z, Young JPW, Chan T-F, Chen WX, Lam H-M, Tian CF. 2018. Coordinated regulation of core and accessory genes in the multipartite genome of Sinorhizobium fredii. PLoS Genet 14:e1007428. doi: 10.1371/journal.pgen.1007428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.diCenzo GC, Maclean AM, Milunovic B, Golding GB, Finan TM. 2014. Examination of prokaryotic multipartite genome evolution through experimental genome reduction. PLoS Genet 10:e1004742. doi: 10.1371/journal.pgen.1004742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.diCenzo GC, Checcucci A, Bazzicalupo M, Mengoni A, Viti C, Dziewit L, Finan TM, Galardini M, Fondi M. 2016. Metabolic modelling reveals the specialization of secondary replicons for niche adaptation in Sinorhizobium meliloti. Nat Commun 7:12219–12210. doi: 10.1038/ncomms12219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Liu A, Contador CA, Fan K, Lam H-M. 2018. Interaction and regulation of carbon, nitrogen, and phosphorus metabolisms in root nodules of legumes. Front Plant Sci 9:1860. doi: 10.3389/fpls.2018.01860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Muñoz N, Qi X, Li M-W, Xie M, Gao Y, Cheung M-Y, Wong FL, Lam H-M. 2016. Improvement in nitrogen fixation capacity could be part of the domestication process in soybean. Heredity (Edinb) 117:84–93. doi: 10.1038/hdy.2016.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Resendis-Antonio O, Reed JL, Encarnación S, Collado-Vides J, Palsson B. 2007. Metabolic reconstruction and modeling of nitrogen fixation in Rhizobium etli. PLoS Comput Biol 3:1887–1895. doi: 10.1371/journal.pcbi.0030192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Resendis-Antonio O, Hernández M, Mora Y, Encarnación S. 2012. Functional modules, structural topology, and optimal activity in metabolic networks. PLoS Comput Biol 8:e1002720. doi: 10.1371/journal.pcbi.1002720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Zhao H, Li M, Fang K, Chen W, Wang J. 2012. In silico insights into the symbiotic nitrogen fixation in Sinorhizobium meliloti via metabolic reconstruction. PLoS One 7:e31287. doi: 10.1371/journal.pone.0031287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Yang Y, Hu XP, Ma BG. 2017. Construction and simulation of the Bradyrhizobium diazoefficiens USDA110 metabolic network: a comparison between free-living and symbiotic states. Mol Biosyst 13:607–620. doi: 10.1039/c6mb00553e. [DOI] [PubMed] [Google Scholar]
- 26.Lieven C, Beber ME, Olivier BG, Bergmann FT, Parizad B, Bartell J, Blank LM, Chauhan S, Correia K, Diener C, Dräger A, Ebert BE, Edirisinghe JN, Fleming RMT, García-Jiménez B, van Helvoirt W, Henry CS, Hermjakob H, Herrgård MJ, Kim HU, King Z, Koehorst JJ, Klamt S, Klipp E, Lakshmanan M, Le Novère N, Lee DY, Lee SY, Lee S, Lewis NE, Ma H, et al. 21 June 2018. Memote : a community driven effort towards a standardized genome-scale metabolic model test suite. bioRxiv doi: 10.1101/350991. [DOI]
- 27.Orth JD, Thiele I, Palsson BØ. 2010. What is flux balance analysis? Nat Biotechnol 28:245–248. doi: 10.1038/nbt.1614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kanehisa M, Furumichi M, Tanabe M, Sato Y, Morishima K. 2017. KEGG: new perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res 45:D353–D361. doi: 10.1093/nar/gkw1092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Arkin AP, Cottingham RW, Henry CS, Harris NL, Stevens RL, Maslov S, Dehal P, Ware D, Perez F, Canon S, Sneddon MW, Henderson ML, Riehl WJ, Murphy-Olson D, Chan SY, Kamimura RT, Kumari S, Drake MM, Brettin TS, Glass EM, Chivian D, Gunter D, Weston DJ, Allen BH, Baumohl J, Best AA, Bowen B, Brenner SE, Bun CC, Chandonia J-M, Chia J-M, Colasanti R, Conrad N, Davis JJ, Davison BH, DeJongh M, Devoid S, Dietrich E, Dubchak I, Edirisinghe JN, Fang G, Faria JP, Frybarger PM, Gerlach W, Gerstein M, Greiner A, Gurtowski J, Haun HL, He F, Jain R, et al. 2018. KBase: the United States Department of Energy systems biology knowledgebase. Nat Biotechnol 36:566–569. doi: 10.1038/nbt.4163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Li YZ, Wang D, Feng XY, Jiao J, Chen WX, Tian CF. 2016. Genetic analysis reveals the essential role of nitrogen phosphotransferase system components in Sinorhizobium fredii CCBAU 45436 symbioses with soybean and pigeonpea plants. Appl Environ Microbiol 82:1305–1315. doi: 10.1128/AEM.03454-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Tombolini R, Nuti MP. 1989. Poly(β-hydroyxalkanoate) biosynthesis and accumulation by different Rhizobium species. FEMS Microbiol Lett 60:299–304. [Google Scholar]
- 32.Lodwig E, Poole P. 2003. Metabolism of Rhizobium bacteroids. CRC Crit Rev Plant Sci 22:37–78. doi: 10.1080/713610850. [DOI] [Google Scholar]
- 33.Li Y, Tian CF, Chen WF, Wang L, Sui XH, Chen WX. 2013. High-resolution transcriptomic analyses of Sinorhizobium sp. NGR234 bacteroids in determinate nodules of Vigna unguiculata and indeterminate nodules of Leucaena leucocephala. PLoS One 8:e70531. doi: 10.1371/journal.pone.0070531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Johnson GV, Evans HJ, Ching T. 1966. Enzymes of the glyoxylate cycle in rhizobia and nodules of legumes. Plant Physiol 41:1330–1336. doi: 10.1104/pp.41.8.1330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Prell J, White JP, Bourdes A, Bunnewell S, Bongaerts RJ, Poole PS. 2009. Legumes regulate Rhizobium bacteroid development and persistence by the supply of branched-chain amino acids. Proc Natl Acad Sci U S A 106:12477–12482. doi: 10.1073/pnas.0903653106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Randhawa GS, Hassani R. 2002. Role of rhizobial biosynthetic pathways of amino acids, nucleotide bases and vitamins in symbiosis. Indian J Exp Biol 40:755–764. [PubMed] [Google Scholar]
- 37.Salminen S, Streeter JG. 1987. Involvement of glutamate in the respiratory metabolism of Bradyrhizobium japonicum bacteroids. J Bacteriol 169:495–499. doi: 10.1128/jb.169.2.495-499.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kouchi H, Fukai K, Kihara A. 1991. Metabolism of glutamate and aspartate in bacteroids isolated from soybean root nodules. J Gen Microbiol 137:2901–2910. doi: 10.1099/00221287-137-12-2901. [DOI] [Google Scholar]
- 39.Rosendahl L, Dilworth M, Glenn AR. 1992. Exchange of metabolites across the peribacteroid membrane in pea root nodules. J Plant Physiol 139:636–638. [Google Scholar]
- 40.Fujihara S, Abe H, Minakawa Y, Akao S, Yoneyama T. 1994. Polyamines in nodules from various plant-microbe symbiotic associations. Plant Cell Physiol 35:1127–1134. doi: 10.1093/oxfordjournals.pcp.a078705. [DOI] [Google Scholar]
- 41.Whitehead LF, Tyerman SD, Day DA. 2001. Polyamines as potential regulators of nutrient exchange across the peribacteroid membrane in soybean root nodules. Aust J Plant Physiol 28:677–683. doi: 10.1071/PP01025. [DOI] [Google Scholar]
- 42.Wang D, Wang YC, Wu LJ, Liu JX, Zhang P, Jiao J, Yan H, Liu T, Tian CF, Chen WX. 2016. Construction and pilot screening of a signature-tagged mutant library of Sinorhizobium fredii. Arch Microbiol 198:91–99. doi: 10.1007/s00203-015-1161-9. [DOI] [PubMed] [Google Scholar]
- 43.Medina C, Crespo-Rivas JC, Moreno J, Espuny MR, Cubo MT. 2009. Mutation in the cobO gene generates auxotrophy for cobalamin and methionine and impairs the symbiotic properties of Sinorhizobium fredii HH103 with soybean and other legumes. Arch Microbiol 191:11–21. doi: 10.1007/s00203-008-0424-0. [DOI] [PubMed] [Google Scholar]
- 44.Resendis-Antonio O, Hernández M, Salazar E, Contreras S, Batallar GM, Mora Y, Encarnación S. 2011. Systems biology of bacterial nitrogen fixation: high-throughput technology and its integrative description with constraint-based modeling. BMC Syst Biol 5:120. doi: 10.1186/1752-0509-5-120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Dunn MF. 2015. Key roles of microsymbiont amino acid metabolism in rhizobia-legume interactions. Crit Rev Microbiol 41:411–451. doi: 10.3109/1040841X.2013.856854. [DOI] [PubMed] [Google Scholar]
- 46.Kim CH, Kuykendall LD, Shah KS, Keister DL. 1988. Induction of symbiotically defective auxotrophic mutants of Rhizobium fredii HH303 by transposon mutagenesis. Appl Environ Microbiol 54:423–427. doi: 10.1128/AEM.54.2.423-427.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Newman JD, Diebold RJ, Schultz BW, Noel KD. 1994. Infection of soybean and pea nodules by Rhizobium spp. purine auxotrophs in the presence of 5-aminoimidazole-4-carboxamide riboside. J Bacteriol 176:3286–3294. doi: 10.1128/jb.176.11.3286-3294.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Crespo-Rivas JC, Margaret I, Pérez-Montaño F, López-Baena FJ, Vinardell JM, Ollero FJ, Moreno FJ, Ruiz-Sainz JE, Buendía-Clavería AM. 2007. A pyrF auxotrophic mutant of Sinorhizobium fredii HH103 impaired in its symbiotic interactions with soybean and other legumes. Int Microbiol 10:169–176. [PubMed] [Google Scholar]
- 49.Buendía-Clavería AM, Moussaid A, Ollero FJ, Vinardell JM, Torres A, Moreno J, Gil-Serrano AM, Rodríguez-Carvajal MA, Tejero-Mateo P, Peart JL, Brewin NJ, Ruiz-Sainz JE. 2003. A purL mutant of Sinorhizobium fredii HH103 is symbiotically defective and altered in its lipopolysaccharide. Microbiology 149:1807–1818. doi: 10.1099/mic.0.26099-0. [DOI] [PubMed] [Google Scholar]
- 50.Kuzma MM, Hunt S, Layzell DB. 1993. Role of oxygen in the limitation and inhibition of nitrogenase activity and respiration rate in individual soybean nodules. Plant Physiol 101:161–169. doi: 10.1104/pp.101.1.161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Udvardi M, Poole PS. 2013. Transport and metabolism in legume-rhizobia symbioses. Annu Rev Plant Biol 64:781–805. doi: 10.1146/annurev-arplant-050312-120235. [DOI] [PubMed] [Google Scholar]
- 52.Jiao J, Wu LJ, Zhang B, Hu Y, Li Y, Zhang XX, Guo HJ, Liu LX, Chen WX, Zhang Z, Tian CF. 2016. MucR is required for transcriptional activation of conserved ion transporters to support nitrogen fixation of Sinorhizobium fredii in soybean nodules. Mol Plant Microbe Interact 29:352–361. doi: 10.1094/MPMI-01-16-0019-R. [DOI] [PubMed] [Google Scholar]
- 53.Gunawardena S, Danso SKA, Zapata F. 1992. Phosphorus requirements and nitrogen accumulation by three mungbean (Vigna radiata (L) Welzek) cultivars. Plant Soil 147:267–274. doi: 10.1007/BF00029078. [DOI] [Google Scholar]
- 54.Karunakaran R, Ramachandran VK, Seaman JC, East AK, Mouhsine B, Mauchline TH, Prell J, Skeffington A, Poole PS. 2009. Transcriptomic analysis of Rhizobium leguminosarum biovar viciae in symbiosis with host plants Pisum sativum and Vicia cracca. J Bacteriol 191:4002–4014. doi: 10.1128/JB.00165-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.El Msehli S, Lambert A, Baldacci-Cresp F, Hopkins J, Boncompagni E, Aschi Smiti S, Hérouart D, Frendo P. 2011. Crucial role of (homo)glutathione in nitrogen fixation in Medicago truncatula nodules. New Phytol 191:496–506. doi: 10.1111/j.1469-8137.2011.03810.x. [DOI] [PubMed] [Google Scholar]
- 56.Dalton DA, Post CJ, Langeberg L. 1991. Effects of ambient oxygen and of fixed nitrogen on concentrations of glutathione, ascrobate, and associated enzymes in soybean root nodules. Plant Physiol 96:812–818. doi: 10.1104/pp.96.3.812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Chan SHJ, Cai J, Wang L, Simons-Senftle MN, Maranas CD. 2017. Standardizing biomass reactions and ensuring complete mass balance in genome-scale metabolic models. Bioinformatics 33:3603–3609. doi: 10.1093/bioinformatics/btx453. [DOI] [PubMed] [Google Scholar]
- 58.King ZA, Lu J, Dräger A, Miller P, Federowicz S, Lerman JA, Ebrahim A, Palsson BO, Lewis NE. 2016. BiGG models: a platform for integrating, standardizing and sharing genome-scale models. Nucleic Acids Res 44:D515–D522. doi: 10.1093/nar/gkv1049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.diCenzo GC, Benedict AB, Fondi M, Walker GC, Finan TM, Mengoni A, Griffitts JS. 2018. Robustness encoded across essential and accessory replicons of the ecologically versatile bacterium Sinorhizobium meliloti. PLoS Genet 14:e1007357. doi: 10.1371/journal.pgen.1007357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Schellenberger J, Lewis NE, Palsson BØ. 2011. Elimination of thermodynamically infeasible loops in steady-state metabolic models. Biophys J 100:544–553. doi: 10.1016/j.bpj.2010.12.3707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.White J, Prell J, James EK, Poole P. 2007. Nutrient sharing between symbionts. Plant Physiol 144:604–614. doi: 10.1104/pp.107.097741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Mitsch MJ, diCenzo GC, Cowie A, Finan TM. 2017. Succinate transport is not essential for symbiotic nitrogen fixation by Sinorhizobium meliloti or Rhizobium leguminosarum. Appl Environ Microbiol 84:e01561-17. doi: 10.1128/AEM.01561-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Preisig O, Anthamatten D, Hennecke H. 1993. Genes for a microaerobically induced oxidase complex in Bradyrhizobium japonicum are essential for a nitrogen-fixing endosymbiosis. Proc Natl Acad Sci U S A 90:3309–3313. doi: 10.1073/pnas.90.8.3309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Li Y, Parsons R, Day DA, Bergersen FJ. 2002. Reassessment of major products of N2 fixation by bacteroids from soybean root nodules. Microbiology 148:1959–1966. doi: 10.1099/00221287-148-6-1959. [DOI] [PubMed] [Google Scholar]
- 65.Werner AK, Medina-Escobar N, Zulawski M, Sparkes IA, Cao F, Witte C-P. 2013. The ureide-degrading reactions of purine ring catabolism employ three amidohydrolases and one aminohydrolase in Arabidopsis, soybean, and rice. Plant Physiol 163:672–681. doi: 10.1104/pp.113.224261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Jiang G, Krishnan AH, Kim Y, Wacek TJ, Krishnan HB, Acteriol JB. 2001. A functional myo-inositol dehydrogenase gene is required for efficient nitrogen fixation and competitiveness of Sinorhizobium fredii USDA191 to nodulate soybean (Glycine max [L.] Merr.). J Bacteriol 183:2595–2604. doi: 10.1128/JB.183.8.2595-2604.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Thiele I, Palsson BØ. 2010. A protocol for generating a high-quality genome-scale metabolic reconstruction. Nat Protoc 5:93–121. doi: 10.1038/nprot.2009.203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Duncan MJ, Fraenkel DG. 1979. α-Ketoglutarate dehydrogenase mutant of Rhizobium meliloti. J Bacteriol 137:415–419. doi: 10.1128/JB.137.1.415-419.1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Walshaw DL, Wilkinson A, Mundy M, Smith M, Poole PS. 1997. Regulation of the TCA cycle and the general amino acid permease by overflow metabolism in Rhizobium leguminosarum. Microbiology 143:2209–2221. doi: 10.1099/00221287-143-7-2209. [DOI] [PubMed] [Google Scholar]
- 70.Pardo M, Lagunez J, Miranda J, Martinez E. 1994. Nodulating ability of Rhizobium tropici is conditioned by a plasmid‐encoded citrate synthase. Mol Microbiol 11:315–321. doi: 10.1111/j.1365-2958.1994.tb00311.x. [DOI] [PubMed] [Google Scholar]
- 71.Acuña G, Ebeling S, Hennecke H. 1991. Cloning, sequencing, and mutational analysis of the Bradyrhizobium japonicum fumC-like gene: evidence for the existence of two different fumarases. J Gen Microbiol 137:991–1000. doi: 10.1099/00221287-137-4-991. [DOI] [PubMed] [Google Scholar]
- 72.Green LS, Emerich DW. 1997. The formation of nitrogen fixing bacteroids is delayed but not abolished in soybean infected by an alpha-ketoglutarate dehydrogenase deficient mutant of Bradyrhizobium japonicum. Plant Physiol 114:1359–1368. doi: 10.1104/pp.114.4.1359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Zhang R, Lin Y. 2009. DEG 5.0, a database of essential genes in both prokaryotes and eukaryotes. Nucleic Acids Res 37:D455–D458. doi: 10.1093/nar/gkn858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Bonaldi K, Gourion B, Fardoux J, Hannibal L, Cartieaux F, Boursot M, Vallenet D, Chaintreuil C, Prin Y, Nouwen N, Giraud E. 2010. Large-scale transposon mutagenesis of photosynthetic Bradyrhizobium sp. strain ORS278 reveals new genetic loci putatively important for nod-independent symbiosis with Aeschynomene indica. Mol Plant Microbe Interact 23:760–770. doi: 10.1094/MPMI-23-6-0760. [DOI] [PubMed] [Google Scholar]
- 75.de las Nieves Peltzer M, Roques N, Poinsot V, Aguilar OM, Batut J, Capela D. 2008. Auxotrophy accounts for nodulation defect of most Sinorhizobium meliloti mutants in the branched-chain amino acid biosynthesis pathway. Mol Plant Microbe Interact 21:1232–1241. doi: 10.1094/MPMI-21-9-1232. [DOI] [PubMed] [Google Scholar]
- 76.Kohler PRA, Zheng JY, Schoffers E, Rossbach S. 2010. Inositol catabolism, a key pathway in Sinorhizobium meliloti for competitive host nodulation. Appl Environ Microbiol 76:7972–7980. doi: 10.1128/AEM.01972-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Fry J, Wood M, Poole PS. 2001. Investigation of myo-inositol catabolism in Rhizobium leguminosarum bv. viciae and its effect on nodulation competitiveness. Mol Plant Microbe Interact 14:1016–1025. doi: 10.1094/MPMI.2001.14.8.1016. [DOI] [PubMed] [Google Scholar]
- 78.Mao C, Qiu J, Wang C, Charles TC, Sobral BW. 2005. NodMutDB: a database for genes and mutants involved in symbiosis. Bioinformatics 21:2927–2929. doi: 10.1093/bioinformatics/bti427. [DOI] [PubMed] [Google Scholar]
- 79.Colijn C, Brandes A, Zucker J, Lun DS, Weiner B, Farhat MR, Cheng TY, Moody DB, Murray M, Galagan JE. 2009. Interpreting expression data with metabolic flux models: predicting Mycobacterium tuberculosis mycolic acid production. PLoS Comput Biol 5:e1000489. doi: 10.1371/journal.pcbi.1000489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Terpolilli JJ, Masakapalli SK, Karunakaran R, Webb IUC, Green R, Watmough NJ, Kruger NJ, Ratcliffe RG, Poole PS. 2016. Lipogenesis and redox balance in nitrogen-fixing pea bacteroids. J Bacteriol 198:2864–2875. doi: 10.1128/JB.00451-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Collier R, Tegeder M. 2012. Soybean ureide transporters play a critical role in nodule development, function and nitrogen export. Plant J 72:355–367. doi: 10.1111/j.1365-313X.2012.05086.x. [DOI] [PubMed] [Google Scholar]
- 82.Heirendt L, Arreckx S, Pfau T, Mendoza SN, Richelle A, Heinken A, Haraldsdóttir HS, Wachowiak J, Keating SM, Vlasov V, Magnusdóttir S, Ng CY, Preciat G, Žagare A, Chan SHJ, Aurich MK, Clancy CM, Modamio J, Sauls JT, Noronha A, Bordbar A, Cousins B, El Assal DC, Valcarcel LV, Apaolaza I, Ghaderi S, Ahookhosh M, Ben Guebila M, Kostromins A, Sompairac N, Le HM, Ma D, Sun Y, Wang L, Yurkovich JT, Oliveira MAP, Vuong PT, El Assal LP, Kuperstein I, Zinovyev A, Hinton HS, Bryant WA, Aragón Artacho FJ, Planes FJ, Stalidzans E, Maass A, Vempala S, Hucka M, Saunders MA, Maranas CD, et al. 2019. Creation and analysis of biochemical constraint-based models: the COBRA Toolbox v3.0. Nat Protoc 14:639–702. doi: 10.1038/s41596-018-0098-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Kellis M, Patterson N, Birren B, Berger B, Lander ES. 2004. Methods in comparative genomics: genome correspondence, gene identification and regulatory motif discovery. J Comput Biol 11:319–355. doi: 10.1089/1066527041410319. [DOI] [PubMed] [Google Scholar]
- 84.Moretti S, Martin O, Du Tran TV, Bridge A, Morgat A, Pagni M. 2016. MetaNetX/MNXref–reconciliation of metabolites and biochemical reactions to bring together genome-scale metabolic networks. Nucleic Acids Res 44:D523–D526. doi: 10.1093/nar/gkv1117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Benson DA, Clark K, Karsch-Mizrachi I, Lipman DJ, Ostell J, Sayers EW. 2015. GenBank. Nucleic Acids Res 43:D30–D35. doi: 10.1093/nar/gku1216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Udvardi MK, Price GD, Gresshoff PM, Day DA. 1988. A dicarboxylate transporter on the peribacteroid membrane of soybean nodules. FEBS Lett 231:36–40. doi: 10.1016/0014-5793(88)80697-5. [DOI] [Google Scholar]
- 87.Bergensen F. 1997. Physiological and biochemical aspects of nitrogen fixation by bacteroids in soybean nodule cells. Soil Biol Biochem 29:875–880. doi: 10.1016/S0038-0717(96)00220-9. [DOI] [Google Scholar]
- 88.Varma A, Palsson BO. 1994. Metabolic flux balancing: basic concepts, scientific and practical use. Nat Biotechnol 12:994–998. doi: 10.1038/nbt1094-994. [DOI] [Google Scholar]
- 89.Becker SA, Feist AM, Mo ML, Hannum G, Palsson BØ, Herrgard MJ. 2007. Quantitative prediction of cellular metabolism with constraint-based models: the COBRA Toolbox. Nat Protoc 2:727–738. doi: 10.1038/nprot.2007.99. [DOI] [PubMed] [Google Scholar]
- 90.Contador CA, Rodríguez V, Andrews BA, Asenjo JA. 2019. Use of genome-scale models to get new insights into the marine actinomycete genus Salinispora. BMC Syst Biol 13:11. doi: 10.1186/s12918-019-0683-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.King ZA, Dräger A, Ebrahim A, Sonnenschein N, Lewis E, Palsson BO. 2015. Escher: a web application for building, sharing, and embedding data-rich visualizations of biological pathways. PLoS Comput Biol 11:e1004321. doi: 10.1371/journal.pcbi.1004321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Ebrahim A, Lerman JA, Palsson BO, Hyduke DR. 2013. COBRApy: constraints-based reconstruction and analysis for Python. BMC Syst Biol 7:74. doi: 10.1186/1752-0509-7-74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Kim D, Langmead B, Salzberg S. 2015. HISAT: a fast spliced aligner with low memory requirements. Nat Methods 12:357–360. doi: 10.1038/nmeth.3317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Anders S, Pyl P, Huber W. 2015. HTSeq: a Python framework to work with high-throughput sequencing data. Bioinformatics 31:166–169. doi: 10.1093/bioinformatics/btu638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Love M, Huber W, Anders S. 2014. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol 15:550. doi: 10.1186/s13059-014-0550-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Richelle A, Chiang AWT, Kuo C, Lewis NE. 2019. Increasing consensus of context-specific metabolic models by integrating data-inferred cell functions. PLoS Comput Biol 15:e1006867. doi: 10.1371/journal.pcbi.1006867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Richelle A, Joshi C, Lewis NE. 2019. Assessing key decisions for transcriptomic data integration in biochemical networks. PLoS Comput Biol 15:e1007185. doi: 10.1371/journal.pcbi.1007185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Jensen PA, Lutz KA, Papin JA. 2011. TIGER: toolbox for integrating genome-scale metabolic models, expression data, and transcriptional regulatory networks. BMC Syst Biol 5:147. doi: 10.1186/1752-0509-5-147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Gudmundsson S, Thiele I. 2010. Computationally efficient flux variability analysis. BMC Bioinformatics 11:489. doi: 10.1186/1471-2105-11-489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Sarkar D, Mueller TJ, Liu D, Pakrasi HB, Maranas CD. 2019. A diurnal flux balance model of Synechocystis sp. PCC 6803 metabolism. PLoS Comput Biol 15:e1006692. doi: 10.1371/journal.pcbi.1006692. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
List of reactions included in the reconstruction model with their description, associated genes, and their reference numbers in various databases. Download Table S1, XLSX file, 0.1 MB (84.1KB, xlsx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Snapshot of reports obtained from memote for comparison of iCC541 with S. meliloti models in HTML format. Model tests are divided in functionality libraries to evaluate core and organism-specific model properties. Core tests are independent of the modeled organism and complexity involved in the model formulation. Each test is divided into scored and unscored sections. The maximum achievable score is 100%. Unscored sections are related to general statistics and specific characteristics of some models such as the quality of the biomass reaction. Download FIG S1, PDF file, 0.5 MB (531.6KB, pdf) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Predicted symbiotic rate ratios between single-gene deletion mutants and wild-type strain. Download Table S2, XLSX file, 0.1 MB (40KB, xlsx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
List of predicted genes that influence symbiosis and the corresponding references for the associated experimental evidence. Download Table S3, DOCX file, 0.1 MB (38.9KB, docx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
List of false-negative predicted genes by the model. Download Table S4, DOCX file, 0.1 MB (35.7KB, docx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Upper and lower boundaries of each reaction in iCC541 of bacteroids isolated from nodules of G. soja (W05) and G. max (C08) as determined by gene expression data. Download Table S5, XLSX file, 0.1 MB (31.3KB, xlsx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Average RPKM values of each gene in iCC541 under both symbiotic conditions (bacteroids isolated from G. soja W05 and G. max C08). Download Table S6, XLSX file, 0.1 MB (39.3KB, xlsx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
List of orthologs of predicted symbiosis-related soybean genes used to build iCC541. The values represent best bidirectional hits (BBHs) percent. Download Table S7, XLSX file, 0.4 MB (422.1KB, xlsx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
List of metabolites included in the reconstruction of iCC541, with their corresponding charges, chemical formulae, and reference IDs in various databases Download Table S8, XLSX file, 0.1 MB (51KB, xlsx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
List of references used to formulate the symbiosis reactions. Download Table S9, DOCX file, 0.1 MB (43.3KB, docx) .
Copyright © 2020 Contador et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Data Availability Statement
The reconstruction, memote reports, and scripts are available at https://github.com/cacontad/SfrediiScripts.