

Abstract
The locus coeruleus is a pontine nucleus that produces much of the brain’s norepinephrine. Despite its small size, the locus coeruleus is critical for a myriad of functions and is involved in many neurodegenerative and neuropsychiatric disorders. In this review, we discuss the physiology and anatomy of the locus coeruleus system and focus on norepinephrine’s role in synaptic plasticity. We highlight Parkinson’s disease as a disorder with motor and neuropsychiatric symptoms that may be understood as aberrations in the normal functions of locus coeruleus.
Keywords: catecholamines, copper, neurodegenerative diseases, neuromodulation, neuronal circuits, neuropsychiatric symptoms, noradrenaline, synaptic plasticity
Introduction
The locus coeruleus (LC) is a norepinephrine-producing nucleus found in the dorsal pons of most vertebrates. The LC was first described in the late 1700s by Félix Vicq d’Azyr (Tubbs et al., 2011) although sources frequently credit Johann Christian Reil (Reil, 1908). Contemporary interest in the LC began with anatomical work by Fumio Sano and Glenn Russel, who independently concluded that the LC is a distinct nucleus with similar - though not identical - appearance across species (Maeda, 2000). Like other neuromodulatory structures, the LC contains an exceedingly small number of cells, yet projects to much of the brain. The human LC is estimated to contain approximately 30,000 neurons that provide norepinephrine to a substantial fraction of the brain’s 100 billion neurons (Mouton et al., 1994). The LC is therefore anatomically poised to modulate a wide range of functions, including homeostasis, sensory processing, motor behavior, and cognition. This review aims to summarize the basic physiology of the LC-norepinephrine system, its function in normal behavior, and its associated neural pathways. We then use this foundation to gain insight into pathophysiology, with an emphasis on Parkinson’s disease (PD). PD is a long-term neurodegenerative disease with both motor and neuropsychiatric symptoms (Rommelfanger and Weinshenker, 2017). PD is traditionally thought to be a disorder of the midbrain dopamine system, since it manifests with substantial dopaminergic neuron loss and is treated with L-DOPA administration, a dopamine precursor. In this review, we augment this perspective by highlighting recent work that emphasizes the contributions of norepinephrine dysfunction to pathogenesis.
We searched PubMed and Google Scholar using the keywords ‘locus coeruleus’ and ‘norepinephrine’ for basic science references and added ‘Parkinson’s disease’ for disease-specific references. We focused mainly on articles published between 2010 and 2019. We included older, seminal references when they were relevant to the scope of this review.
Basic Physiology
Seminal studies of LC physiology developed the idea that the system is a crucial node in arousal or attention. This key hypothesis has continued to shape the design and interpretation of modern experiments (Carter et al., 2010; Lovett-Barron et al., 2017). One of the first systematic studies to employ single-unit recordings in awake animals - rats and squirrel monkeys - found that LC neurons homogeneously increase stimulus-evoked firing in response to salient auditory, visual, and touch stimuli (Foote et al., 1980). This was an important finding, as previous studies in anesthetized animals found that only noxious stimuli could elicit robust increases in firing, which led to hypotheses that the system was primarily concerned with fear or nociception. It was also discovered that LC exhibit lower spontaneous firing during sleep, further cementing its role in arousal (Aston-Jones and Bloom, 1981).
Well-controlled behavioral experiments added to this view and began to paint a more nuanced picture. In macaques trained to respond to an oddball visual stimulus, target stimuli were found to drive robust phasic firing while non-target stimuli elicited no changes (Rajkowski et al., 1994). The phasic response scales with the expected value of the stimulus (Bouret and Richmond, 2015) and the effort required to produce the action (Varazzani et al., 2015). Further analyses revealed that this phasic response is better aligned to the action than the stimulus (Aston-Jones and Cohen, 2005; Bouret and Richmond, 2015). LC responses do not always precede overt motor behavior, as behavioral tasks designed to separate stimuli from actions have found phasic responses to both (Kalwani et al., 2014), and similar spontaneous movements (i.e., not in response to a cue) do not elicit changes in firing. Altogether, these findings have lent evidence to the hypothesis that LC is necessary to promote arousal or mobilize effort (Sara and Bouret, 2012; Varazzani et al., 2015; Xiang et al., 2019). However, one intriguing study trained monkeys to perform a countermanding saccade task and found that - although LC responds to go cues and to saccades - it does not respond when saccades are appropriately cancelled (Kalwani et al., 2014). Although the full implications of this finding have yet to be thoroughly understood, it challenges the most straightforward interpretation of the arousal/effort hypotheses, since inhibiting a planned saccade is an effortful and demanding action.
LC responds vigorously to novel stimuli. However, if the stimuli are not behaviorally relevant, the response decays rapidly. Activity reappears during reversal learning or extinction (Sara and Segal, 1991), and after reversal, the LC response to stimuli can be expressed before behavioral expression of reversal (Aston-Jones et al., 1997). Additionally, LC neurons are sensitive to changes in task states in monkeys performing a decision-making task (Jahn et al., 2019). These studies lent evidence to the idea that LC is important for learning new behavioral contingencies.
The LC is now appreciated to have at least two modes of firing, with consequences for behavioral performance: a background tonic mode, and a short-timescale phasic mode (Varazzani et al., 2015). The first hints of a relationship between LC and behavioral performance came from primate studies which found that low tonic activity is correlated with task disengagement and drowsiness. Elevated tonic/low phasic firing is also correlated with task disengagement, although monkeys are distractible in this regime (Varazzani et al., 2015). A sweet spot in the middle, with low tonic activity and high phasic activity, is related to good task engagement. These modes of firing elicit different release profiles of norepinephrine and distinct modulation of downstream circuits (McBurney-Lin et al., 2019).
Role in Learning and Memory
Manipulation of the LC-norepinephrine system has profound consequences for cognitive behavior. Lesion studies have reported gross deficits in learning, with LC-ablated animals taking much longer to learn to run down a runway for food reward (Anlezark et al., 1973). This was likely not due to gross motivation deficits, as control and lesioned animals had similar body weights and exploratory behavior in an open field. Vicarious trial-and-error, an index of learning, is disrupted with target-specific pharmacological manipulation of norepinephrine (Amemiya et al., 2016). LC-norepinephrine manipulation affects distinct phases of memory processing, depending on task conditions and details of manipulation (Khakpour-Taleghani et al., 2009). Other studies have pointed to a more nuanced role in learning when task contingencies change. Recent optogenetic manipulation experiments have argued that disparate effects on learning may be a consequence of projection-target specificity and downstream computations (Uematsu et al., 2017).
Unified Accounts of Locus Coeruleus Function
One of the earliest general theories of LC hypothesized that norepinephrine release from LC facilitates learning associations between stimuli and outcomes (Kety, 1970). This theory has received support over the years, but the exact role of LC with learning is nuanced and remains to be fully explored. Modern theories of LC function have taken into account numerous findings since Kety’s hypothesis was formulated. One theory holds that LC encodes for unexpected uncertainty - that is, when the world changes in an unpredictable manner (Dayan and Yu, 2006). The functional effect is to influence inference and mediate changes in synaptic plasticity to allow for rapid learning. This theory has received support from human pupillometry studies (Nassar et al., 2012). Another similar theory, the adaptive-gain hypothesis, argues for an inverted-U relationship between tonic LC activity and behavioral performance (Varazzani et al., 2015). Low and high tonic LC are associated with task disengagement, with high tonic LC favoring distractibility and exploratory behavior. When tonic LC is in between these extremes, then task engagement is high. The function of these LC patterns is to optimize reward/utility, which is hypothesized to be under the control of prefrontal value circuitry. The adaptive-gain hypothesis has likewise received support from human pupillometry studies (Gilzenrat et al., 2010; Jepma and Nieuwenhuis, 2011). Another similar account holds that LC signals higher-order prediction errors to enable fast-timescale behavioral change (Sales et al., 2019). Finally, since the effects of LC manipulation are occasionally specific to extinction learning, we propose that LC is involved in the generation of latent states (Gershman et al., 2017). This theory predicts that during extinction learning (after a cue has been associated with an outcome), the brain does not degrade the original memory but rather associates the extinction memory with a new latent state. The original cue-outcome memory remains in the brain, largely unaffected. If LC is over- or underactive, the brain may create too many or too few latent states. Dysfunction of the LC-norepinephrine system may therefore result in cognitive deficits, particularly in learning, memory, and decision making.
Synaptic Plasticity Mechanisms of Noradrenergic Signaling
The role of LC in learning, memory, and behavioral flexibility has stimulated interest in noradrenaline’s mechanism of action. Synaptic plasticity mechanisms are particularly well-suited to re-adapt neural circuitry to process new sensory inputs and task contingencies to produce desired actions. Initial work from Bear and Singer (1986) demonstrated that synergy of both adrenergic and cholinergic neurotransmission is required for experience-dependent plasticity in the primary visual cortex. Later studies showed that norepinephrine has independent effects on plasticity throughout the brain including visual cortex (van den Pol et al., 2002; Huang et al., 2013), frontal cortex (Lim et al., 2010), primary auditory cortex (Martins and Froemke, 2015), hippocampus (Takeuchi et al., 2016) and amygdala (Johansen et al., 2014). In mice, chemogenetic stimulation of LC induces rapid increases in brain-wide functional connectivity in regions involved in salience processing (Zerbi et al., 2019). Importantly, these findings appear to translate to the human brain, as fMRI studies have shown changes in brain-wide connectivity with increased catecholamine concentration (van den Brink et al., 2016).
Noradrenergic signaling can produce both synaptic weakening as well strengthening. This is, in part, due to heterogeneity of downstream receptors. Norepinephrine can have an excitatory effect on its target neurons through α1 (Gq-coupled) and β (Gs-coupled) adrenergic receptors and inhibitory effects through α2 (Gi-coupled) adrenergic receptors (McBurney-Lin et al., 2019). An interesting theory suggests that, given the same history of synaptic activity, a synapse can be primed for either potentiation or depression depending on the G-coupled receptor activated by neuromodulators (van den Pol et al., 2002). This bidirectional plasticity has been shown in visual cortex (Seol et al., 2007; Huang et al., 2013; He et al., 2015) and could be the answer for rapid modulation of synaptic connectivity by neuromodulators. Further, He et al. (2015) suggested particular neuromodulators can prime synapses for a specific polarity of plasticity. Norepinephrine dependent regulation of the AMPA receptor population could allow for widespread plasticity through the cerebral cortex (Hu et al., 2007). At a circuit level, the ability of norepinephrine to differentially regulate inhibitory cells (through α2 receptors) and excitatory cells (through α1 and β receptors), as observed in the basal forebrain and cortex, could produce a change in excitatory/inhibitory drive onto their downstream targets. This would trigger plasticity mechanisms in target neurons in order to maintain excitatory/inhibitory balance (McBurney-Lin et al., 2019).
Afferent Pathways of the Locus Coeruleus
The myriad of LC functions can be explained, in part, by its functional connectivity. Modern studies have exploited the specificity of viral transgenic tools to label the LC afferent population and revealed that LC receives inputs from a remarkably diverse number of regions (Schwarz et al., 2015; Uematsu et al., 2017). Almost all areas of the neocortex project to the LC (Schwarz et al., 2015), with strong glutamatergic projections from the prefrontal cortex (Jodo and Aston-Jones, 1997) and corticotropin-releasing factor projections from the amygdala (Szabadi, 2013). Sleep-promoting GABAergic neurons in ventrolateral preoptic nucleus, a region of the hypothalamus, inhibit LC during slow wave sleep (Szymusiak and McGinty, 2008) and orexinergic neurons in lateral hypothalamus/perifornical area send strong excitation to LC to promote wakefulness (Szabadi, 2013). LC neurons also receive excitatory input from dopaminergic neurons in the ventral tegmental area. Because infusion of dopamine into LC inhibits sleep (Crochet and Sakai, 2003), this pathway is thought to be part of the wakefulness promoting pathway. Given the important role of the midbrain dopamine system in motivated behavior, this pathway may also be critical for LC’s function in reward-based learning. Other reviews provide detailed information regarding further important input pathways (Szabadi, 2013; Schwarz and Luo, 2015).
Efferent Pathways of the Locus Coeruleus
Since individual LC neurons have little molecular and morphological diversity (compared to neuromodulators like the dorsal raphe serotonergic system), the diverse nature of LC function may be explained by projection profiles of neuronal subsets (Seo and Bruchas, 2017). The classical view holds that LC broadly and nonspecifically modulates neuronal function throughout the brain via volumetric transmission. However, these findings were mostly inferred from experiments using non-specific labeling techniques. This view has been recently challenged with the development of molecular labeling techniques. There is evidence for some modularity, but there is extensive collateralization of many output pathways (Schwarz and Luo, 2015; Kebschull et al., 2016). Overall, there are at least three major efferent pathways originating from LC (Figure 1): 1) ascending pathway projections to the cortex, 2) cerebellar pathway and 3) descending pathway projections to the spinal cord (Szabadi, 2013). The ascending pathway is mostly involved in arousal, behavioral flexibility, and brain state mediation. The descending pathway is thought to control lower-level motor actions. The cerebellar pathway is remarkably understudied and a distinct function is unclear. However, the cerebellar cortex has robust expression of noradrenergic receptors and disruption of this signaling impairs motor function. We intentionally limit the scope of this review but other references provide more details (Szabadi, 2013; Schwarz et al., 2015). We will focus on the ascending and descending pathways.
Figure 1.
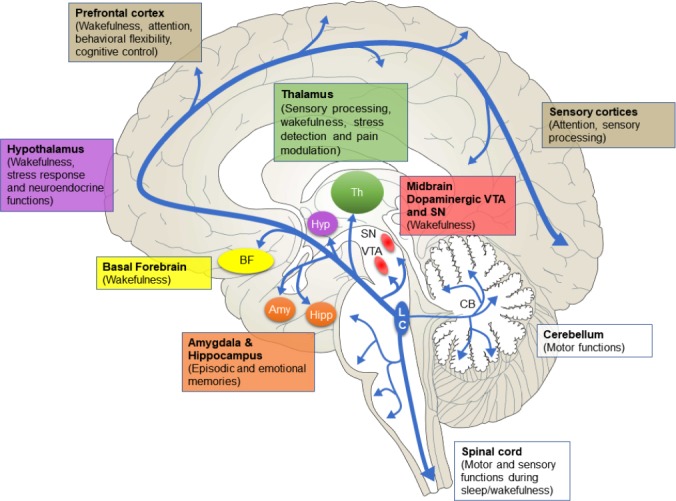
Locus coeruleus (LC) efferent pathways and relevant functions.
LC projects throughout the brain in three major pathways with widespread collateralization. The ascending pathway consists of the ventral tegmental area (VTA) and substantia nigra (SN), amygdala (Amy), hippocampus (Hipp), hypothalamus (Hyp), thalamus (Th), basal forebrain (BF), prefrontal cortex and sensory cortices. The cerebellar pathway and descending pathway to the spinal cord are involved in motor functions. Dysfunction in these pathways may lead to PD symptoms. Note: The size of each area is not to scale.
Ascending pathway
The ascending pathway is made up of projections to the limbic system, midbrain, thalamus, basal forebrain and all of the neocortex (Figure 1). This pathway is thought to be involved in behavioral flexibility, cognitive control, wakefulness, formation and retrieval of episodic and emotional memories, modulation of pain, response to stress, cardiovascular regulation, nociception, pupillary light reflex, and some sympathetic and parasympathetic functions.
The neocortex and basal forebrain are heavily innervated by LC (Szabadi, 2013; Schwarz and Luo, 2015; McBurney-Lin et al., 2019). Expression of excitatory noradrenergic receptors (α1 and β) on excitatory neurons and inhibitory receptors (α2) on GABAergic neurons allows LC to exert excitatory influence on these areas (Szabadi, 2013; Schwarz and Luo, 2015), which would prime LC to modulate cortical arousal and cognition.
LC extensively projects to the thalamus, primarily to excitatory neurons expressing the excitatory α1 receptor (Szabadi, 2013). Through the thalamus, LC affects sensory processing, wakefulness, stress detection and pain modulation (Szabadi, 2013; Beas et al., 2018; Rodenkirch et al., 2019). Outside of the thalamus, LC has bidirectional connectivity with the hypothalamus; this circuitry is part of sleep-wake circuitry through ventrolateral preoptic area and lateral hypothalamic/perifornical areas, stress response through the paraventricular nucleus, and neuroendocrine functions through the arcuate nucleus (Szabadi, 2013; Schwarz and Luo, 2015). In the midbrain, dopaminergic neurons in the ventral tegmental area and substantia nigra receive noradrenergic innervation from LC (as well as other brainstem adrenergic regions) (Rommelfanger and Weinshenker, 2007; Mejias-Aponte, 2016). This particular pathway highlights the fact that neuromodulators influence each other, suggesting a need to understand their combinatorial functions, so we may better understand physiology and pathophysiology.
The role of LC in learning and memory is thought to depend in part on the amygdala and hippocampus. The amygdala, known to mediate fear and anxiety responses (Sah, 2017), contains all major noradrenergic receptors; these have been implicated in formation and retrieval of emotional memories (Szabadi, 2013; Uematsu et al., 2017). LC is the exclusive source of noradrenaline in the hippocampus and, like the amygdala, all major adrenoreceptors have been reported (Szabadi, 2013). The LC-hippocampus pathway is critical for the formation, consolidation and retrieval of memories (Sara and Devauges, 1988; Takeuchi et al., 2016). Recent studies have revealed a special case for LC neurotransmission. Dopamine, a precursor of norepinephrine, is released from LC terminals in the dorsal hippocampus to support the formation of novelty-based memory (Kempadoo et al., 2016; Takeuchi et al., 2016; Wagatsuma et al., 2018).
Projections from LC to the brainstem are thought to be critical for parasympathetic and sympathetic functions (Szabadi, 2013).
Descending pathway
The coeruleo-spinal pathway is formed by widespread projects from dorsal and caudal LC neurons to the spinal cord, with some collateralization in the brainstem (Figure 1). The targets are sensory neurons in the dorsal horn, motor neurons in the ventral horn and preganglionic neurons in intermediolateral nuclei (Szabadi, 2013). LC norepinephrine largely inhibits sensory neurons in the dorsal horn, as these cells express inhibitory α2 receptors. One primary function is pain desensitization, as activation of this receptor subtype provides analgesia/hyperalgesia. The net effect of norepinephrine in the ventral horn is excitation, due to expression of α1 receptors. This is consistent with the finding that LC activity is low during REM sleep, since muscle tone is lost (Szabadi, 2013; Sara, 2017).
Locus Coeruleus Dysfunction in Parkinson’s Disease – Motor and Neuropsychiatric Symptoms
Given LC’s role in a number of basic functions, one may expect that dysfunction should similarly lead to a wide range of symptoms. Indeed, significant neuronal loss in LC is associated with neurodegenerative disorders, such as PD and Alzheimer’s disease (German et al., 1992; Hoogendijk et al., 1995; Marien et al., 2004), as well as psychiatric diseases such as depression (Ordway and Klimek, 2001). In order to link the basic findings of LC function with disease, we will specifically focus on PD, a disorder with both motor and psychiatric symptoms (Gold and Chrousos, 2002; Marien et al., 2004). Although the literature lacks clear mechanistic explanations, we attempt to link symptoms of PD with LC dysfunction and highlight plausible mechanistic links.
PD is primarily thought to be a disease of dopaminergic cell loss in the substantia nigra. However, recent evidence has pointed to LC norepinephrine as a critical component of this disease (Rommelfanger and Weinshenker, 2007). In PD, LC cell loss occurs throughout the nucleus and extends into the peri-LC subcoeruleus region. The remaining neurons exhibit significant shrinkage and have an altered phenotype (Hoogendijk et al., 1995). In animal models of this disease, noradrenergic signaling from LC provides protection from dopaminergic cell loss in the substantia nigra (Srinivasan and Schmidt, 2003; Marien et al., 2004). One study confirmed dopaminergic cell death in the substantia nigra of PD patients and observed dramatic cell loss in the LC (Zarow et al., 2003). Furthermore, in a mouse model of PD (mutation of the PARK2 gene), there is cell loss from the LC while the nigrostriatal system is unaffected (Von Coelln et al., 2004). LC’s projections to motor cortices and the spinal cord could provide crucial insight for understanding wide-ranging symptoms in PD.
Neuropsychiatric Symptoms in Parkinson’s Disease
LC is associated with early non-motor symptoms of PD such as depression (Remy et al., 2005) and anxiety (Rahman et al., 2009; McCall et al., 2015; Zhu et al., 2017). A useful biomarker for neurodegeneration in PD is neuromelanin – a byproduct of dopamine and norepinephrine synthesis - which can be accessed through MRI. Neuromelanin is decreased in both the substantia nigra and LC of PD patients and LC neuron loss is exaggerated in patients with depressive symptoms (Wang et al., 2018). One link between LC and depression is the observation that stressful episodes can potentiate depressive symptoms. This is possibly due to interactions between the stress-induced corticotropin-releasing hormone system and LC (Gold and Chrousos, 2002; Gold et al., 2015), as corticotropin-releasing factor (potentially released from the amygdala (Reyes et al., 2008) is increased in the LC of patients with depression (Bissette et al., 2003). The link between stress and depression is strengthened by the observation that in post-traumatic stress disorder, 50% of patients develop depression (Pitman et al., 2012). In addition, patients with melancholic depression suffer from hyperarousal, anxiety, and sleep disturbances and have elevated norepinephrine levels in the plasma and cerebrospinal fluid (Wong et al., 2000). The treatment of depression frequently requires targeting the LC-norepinephrine system with serotonin-norepinephrine reuptake inhibitors and norepinephrine reuptake inhibitors (Zhou, 2004).
Other disorders like chronic neuropathic pain can induce depression associated with noradrenergic impairment (Alba-Delgado et al., 2013). In pain-related anxiety, corticotropin-releasing hormone induces activation of extracellular signal-regulated kinase 1/2 signaling to upregulate LC function (Borges et al., 2015). Stress is known to increase the firing of LC neurons (Bingham et al., 2011), which, in turn, induces arousal and can precipitate anxiety and aversion (McCall et al., 2015).
Patients with PD commonly suffer from sleep disturbances, a non-motor symptom thought to be related to LC pathology (Braak et al., 2003; Abbott et al., 2005). As previously mentioned, the LC norepinephrine system is part of the sleep-wake cycle through projections to wake-promoting regions (Szabadi, 2013; Schwarz, 2015). Almost all PD patients suffer from sleep disturbances with individual variability, which include disordered breathing, vivid dreaming and excessive daytime sleepiness (Chaudhuri et al., 2006; Verbaan et al., 2008). The LC system is known to be involved in chemoreception to maintain normal breathing, a function that is disrupted in PD (Oliveira et al., 2017). PD patients also suffer from cognitive impairment, even in the early stages of the disease (Weintraub et al., 2015). We suggest that these impairments may be understood as disruptions in LC’s computational functions (adaptive gain, unexpected uncertainty, generation of latent states).
Motor Symptoms in Parkinson’s Disease
Motor symptoms of PD arise in humans when ~80% of dopamine neurons are lost in the substantia nigra (Rommelfanger and Weinshenker, 2007). Interestingly, this may not be sufficient to generate motor symptoms. Pharmacological application of MPTP, a drug that selectively destroys dopamine cells, causes significant dopamine loss in the nigrostriatal system yet does not generally result in profound motor symptoms. When MPTP is combined with pharmacological ablation of the LC, the classic motor symptoms of PD emerge (Marien et al., 1993; Rommelfanger and Weinshenker, 2007). Mice that lack the norepinephrine transporter gene are partially protected from MPTP toxicity, suggesting that extracellular norepinephrine may be protective against dopamine cell death (Rommelfanger et al., 2004). Interestingly, ascending LC projections to the forebrain are not the only contributors to PD, as dysfunction of the descending cerulospinal tract may also play a key role in rigidity (Paulus and Jellinger, 1991).
Typical motor symptoms of PD become apparent when α-synucleinopathy can be detected and substantia nigra deterioration occurs (Braak et al., 2003). α-Synuclein, a presynaptic protein thought to regulate neurotransmission, is overexpressed in PD and believed to contribute to dysregulation of homeostasis, cell death, and may contribute to disease propagation (Stefanis, 2012). Interestingly, α-synuclein plaques are not only involved in compartmentalization of dopamine in the substantia nigra, but also alter the storage of norepinephrine in the dentate gyrus (Yavich et al., 2006). It has been suggested that α-synucleinopathy might interfere with an antioxidant role for norepinephrine. Both extracellular dopamine and norepinephrine can prevent free radical formation and act to protect neurons against oxidative stress (Troadec et al., 2002; Traver et al., 2005). This protective role could explain why norepinephrine transporter knockout protects against MPTP toxicity. The antioxidative role of catecholamines is further supported by the localization of dopamine β-hydroxylase, the enzyme that catalyzes the hydroxylation of dopamine to norepinephrine, to the mitochondrial membranes of LC neurons (Issidorides et al., 2004).
Copper-Rich Food for Thought
Copper is an essential cofactor for dopamine β-hydroxylase (Schmidt et al., 2018). Under normal conditions, copper levels are enriched in the LC (Schmidt et al., 2019) and are higher than in the substantia nigra (Zecca et al., 2004). In particular, copper is thought to play a protective role in these brain regions, and copper dysregulation could contribute to neuronal cell death. One hypothesis is that the protective effect of copper could be due to the anti-oxidative role of superoxide dismutase 1, a copper-dependent enzyme (Genoud et al., 2017; Trist et al., 2017). The malfunction of superoxide dismutase 1 in the substantia nigra of PD patients may contribute to dopaminergic cell loss (Trist et al., 2019). Consistent with this idea, there is a decrease in copper levels and expression of copper transporter Ctr1 in both the substantia nigra and LC of PD patients (Davies et al., 2014; Genoud et al., 2017).
Interestingly, in Wilson’s disease, a classic disorder of copper accumulation caused by a mutation in the copper transporter ATP7B, patients often manifest with PD-like symptoms of tremor and gait impairment. These patients typically have abnormal serum catecholamine levels and suffer from similar neuropsychiatric symptoms as PD patients (Benhamla et al., 2007). A recent study suggested that the role of ATP7B is to sequester intracellular copper, which is required for secretion of dopamine β-hydroxylase, and thus provides regulation of intracellular and extracellular catecholamine (Schmidt et al., 2018). For these reasons, we speculate that aberrance in the copper-regulated catecholamine balance may contribute to symptoms in PD.
Conclusion
In summary, the LC-norepinephrine system is a pontine neuromodulatory nucleus with broad projections throughout the forebrain, cerebellum, and spinal cord. Classic studies have implicated LC in a myriad of functions such as arousal, behavioral flexibility, learning, memory, and wakefulness. LC neurons are remarkably homogeneous, especially when compared to other neuromodulatory structures. It is likely that functions of LC may be understood in terms of the relevant efferent pathways, target structures, and local receptor heterogeneity. Mechanistically, LC-norepinephrine has long been studied as a modulator of synaptic plasticity, allowing local circuits to dynamically adapt in the face of new inputs. The many functions of LC may help explain symptoms of PD, a neurodegenerative disease that is now understood to be a dysfunction of both the dopamine and norepinephrine system. LC-norepinephrine is neuroprotective in PD and its loss may contribute to both motor-related and non-motor-related symptoms. Thanks to modern research using cell-type-specific techniques, the circuit logic of LC is slowly becoming clear, with implications for understanding and treating disease.
Acknowledgments
We thank Jeremiah Y. Cohen and Hongdian Yang for comments on the manuscript.
Footnotes
Conflicts of interest: The authors declare no conflicts of interest.
Financial support: This work was supported by the National Institutes of Health grant F30MH110084 (to BAB).
Copyright license agreement: The Copyright License Agreement has been signed by all authors before publication.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Open peer reviewer: Cristoforo Comi, University of Eastern Piedmont, Italy.
Funding: This work was supported by the National Institutes of Health grant F30MH110084 (to BAB).
P-Reviewer: Comi C; C-Editors: Zhao M, Li JY; T-Editor: Jia Y
References
- 1.Abbott RD, Ross GW, White LR, Tanner CM, Masaki KH, Nelson JS, Curb JD, Petrovitch H. Excessive daytime sleepiness and subsequent development of Parkinson disease. Neurology. 2005;65:1442–1446. doi: 10.1212/01.wnl.0000183056.89590.0d. [DOI] [PubMed] [Google Scholar]
- 2.Alba-Delgado C, Llorca-Torralba M, Horrillo I, Ortega JE, Mico JA, Sanchez-Blazquez P, Meana JJ, Berrocoso E. Chronic pain leads to concomitant noradrenergic impairment and mood disorders. Biol Psychiatry. 2013;73:54–62. doi: 10.1016/j.biopsych.2012.06.033. [DOI] [PubMed] [Google Scholar]
- 3.Amemiya S, Kubota N, Umeyama N, Nishijima T, Kita I. Noradrenergic signaling in the medial prefrontal cortex and amygdala differentially regulates vicarious trial-and-error in a spatial decision-making task. Behav Brain Res. 2016;297:104–111. doi: 10.1016/j.bbr.2015.09.002. [DOI] [PubMed] [Google Scholar]
- 4.Anlezark GM, Crow TJ, Greenway AP. Impaired learning and decreased cortical norepinephrine after bilateral locus coeruleus lesions. Science. 1973;181:682–684. doi: 10.1126/science.181.4100.682. [DOI] [PubMed] [Google Scholar]
- 5.Aston-Jones G, Bloom FE. Activity of norepinephrine-containing locus coeruleus neurons in behaving rats anticipates fluctuations in the sleep-waking cycle. J Neurosci. 1981;1:876–886. doi: 10.1523/JNEUROSCI.01-08-00876.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Aston-Jones G, Cohen JD. An integrative theory of locus coeruleus-norepinephrine function: adaptive gain and optimal performance. Annu Rev Neurosci. 2005;28:403–450. doi: 10.1146/annurev.neuro.28.061604.135709. [DOI] [PubMed] [Google Scholar]
- 7.Aston-Jones G, Rajkowski J, Kubiak P. Conditioned responses of monkey locus coeruleus neurons anticipate acquisition of discriminative behavior in a vigilance task. Neuroscience. 1997;80:697–715. doi: 10.1016/s0306-4522(97)00060-2. [DOI] [PubMed] [Google Scholar]
- 8.Bear MF, Singer W. Modulation of visual cortical plasticity by acetylcholine and noradrenaline. Nature. 1986;320:172–176. doi: 10.1038/320172a0. [DOI] [PubMed] [Google Scholar]
- 9.Beas BS, Wright BJ, Skirzewski M, Leng Y, Hyun JH, Koita O, Ringelberg N, Kwon HB, Buonanno A, Penzo MA. The locus coeruleus drives disinhibition in the midline thalamus via a dopaminergic mechanism. Nat Neurosci. 2018;21:963–973. doi: 10.1038/s41593-018-0167-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Benhamla T, Tirouche YD, Abaoub-Germain A, Theodore F. The onset of psychiatric disorders and Wilson’s disease. Encephale. 2007;33:924–932. doi: 10.1016/j.encep.2006.08.009. [DOI] [PubMed] [Google Scholar]
- 11.Bingham B, McFadden K, Zhang X, Bhatnagar S, Beck S, Valentino R. Early adolescence as a critical window during which social stress distinctly alters behavior and brain norepinephrine activity. Neuropsychopharmacology. 2011;36:896–909. doi: 10.1038/npp.2010.229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bissette G, Klimek V, Pan J, Stockmeier C, Ordway G. Elevated concentrations of CRF in the locus coeruleus of depressed subjects. Neuropsychopharmacology. 2003;28:1328–1335. doi: 10.1038/sj.npp.1300191. [DOI] [PubMed] [Google Scholar]
- 13.Borges GP, Mico JA, Neto FL, Berrocoso E. Corticotropin-releasing factor mediates pain-induced anxiety through the ERK1/2 signaling cascade in locus coeruleus neurons. Int J Neuropsychopharmacol. 2015 doi: 10.1093/ijnp/pyv019. doi:101093/ijnp/pyv019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bouret S, Richmond BJ. Sensitivity of locus ceruleus neurons to reward value for goal-directed actions. J Neurosci. 2015;35:4005–4014. doi: 10.1523/JNEUROSCI.4553-14.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Braak H, Del Tredici K, Rub U, de Vos RA, Jansen Steur EN, Braak E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol Aging. 2003;24:197–211. doi: 10.1016/s0197-4580(02)00065-9. [DOI] [PubMed] [Google Scholar]
- 16.Carter ME, Yizhar O, Chikahisa S, Nguyen H, Adamantidis A, Nishino S, Deisseroth K, de Lecea L. Tuning arousal with optogenetic modulation of locus coeruleus neurons. Nat Neurosci. 2010;13:1526–1533. doi: 10.1038/nn.2682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chaudhuri KR, Healy DG, Schapira AH. Non-motor symptoms of Parkinson’s disease: diagnosis and management. Lancet Neurol. 2006;5:235–245. doi: 10.1016/S1474-4422(06)70373-8. [DOI] [PubMed] [Google Scholar]
- 18.Crochet S, Sakai K. Dopaminergic modulation of behavioral states in mesopontine tegmentum: a reverse microdialysis study in freely moving cats. Sleep. 2003;26:801–806. doi: 10.1093/sleep/26.7.801. [DOI] [PubMed] [Google Scholar]
- 19.Davies KM, Bohic S, Carmona A, Ortega R, Cottam V, Hare DJ, Finberg JP, Reyes S, Halliday GM, Mercer JF, Double KL. Copper pathology in vulnerable brain regions in Parkinson’s disease. Neurobiol Aging. 2014;35:858–866. doi: 10.1016/j.neurobiolaging.2013.09.034. [DOI] [PubMed] [Google Scholar]
- 20.Dayan P, Yu AJ. Phasic norepinephrine: a neural interrupt signal for unexpected events. Network. 2006;17:335–350. doi: 10.1080/09548980601004024. [DOI] [PubMed] [Google Scholar]
- 21.Foote SL, Aston-Jones G, Bloom FE. Impulse activity of locus coeruleus neurons in awake rats and monkeys is a function of sensory stimulation and arousal. Proc Natl Acad Sci U S A. 1980;77:3033–3037. doi: 10.1073/pnas.77.5.3033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Genoud S, Roberts BR, Gunn AP, Halliday GM, Lewis SJG, Ball HJ, Hare DJ, Double KL. Subcellular compartmentalisation of copper, iron, manganese, and zinc in the Parkinson’s disease brain. Metallomics. 2017;9:1447–1455. doi: 10.1039/c7mt00244k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.German DC, Manaye KF, White CL, 3rd, Woodward DJ, McIntire DD, Smith WK, Kalaria RN, Mann DM. Disease-specific patterns of locus coeruleus cell loss. Ann Neurol. 1992;32:667–676. doi: 10.1002/ana.410320510. [DOI] [PubMed] [Google Scholar]
- 24.Gershman SJ, Monfils MH, Norman KA, Niv Y. The computational nature of memory modification. Elife. 2017 doi: 10.7554/eLife.23763. doi:107554/eLife23763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gilzenrat MS, Nieuwenhuis S, Jepma M, Cohen JD. Pupil diameter tracks changes in control state predicted by the adaptive gain theory of locus coeruleus function. Cogn Affect Behav Neurosci. 2010;10:252–269. doi: 10.3758/CABN.10.2.252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gold PW, Chrousos GP. Organization of the stress system and its dysregulation in melancholic and atypical depression: high vs low CRH/NE states. Mol Psychiatry. 2002;7:254–275. doi: 10.1038/sj.mp.4001032. [DOI] [PubMed] [Google Scholar]
- 27.Gold PW, Machado-Vieira R, Pavlatou MG. Clinical and biochemical manifestations of depression: relation to the neurobiology of stress. Neural Plast. 2015;2015:581976. doi: 10.1155/2015/581976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.He K, Huertas M, Hong SZ, Tie X, Hell JW, Shouval H, Kirkwood A. Distinct eligibility traces for LTP and LTD in cortical synapses. Neuron. 2015;88:528–538. doi: 10.1016/j.neuron.2015.09.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Hoogendijk WJ, Pool CW, Troost D, van Zwieten E, Swaab DF. Image analyser-assisted morphometry of the locus coeruleus in Alzheimer’s disease, Parkinson’s disease and amyotrophic lateral sclerosis. Brain. 1995;118:131–143. doi: 10.1093/brain/118.1.131. [DOI] [PubMed] [Google Scholar]
- 30.Hu H, Real E, Takamiya K, Kang MG, Ledoux J, Huganir RL, Malinow R. Emotion enhances learning via norepinephrine regulation of AMPA-receptor trafficking. Cell. 2007;131:160–173. doi: 10.1016/j.cell.2007.09.017. [DOI] [PubMed] [Google Scholar]
- 31.Huang S, Huganir RL, Kirkwood A. Adrenergic gating of Hebbian spike-timing-dependent plasticity in cortical interneurons. J Neurosci. 2013;33:13171–13178. doi: 10.1523/JNEUROSCI.5741-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Issidorides MR, Havaki S, Arvanitis DL, Chrysanthou-Piterou M. Noradrenaline storage function of species-specific protein bodies, markers of monoamine neurons in human locus coeruleus demonstrated by dopamine-beta-hydroxylase immunogold localization. Prog Neuropsychopharmacol Biol Psychiatry. 2004;28:829–847. doi: 10.1016/j.pnpbp.2004.05.034. [DOI] [PubMed] [Google Scholar]
- 33.Jahn CI, Varazzani C, Sallet J, Walton ME, Bouret S. Noradrenergic but not dopaminergic neurons signal task state changes and predict re-engagement after a failure. bioRxiv. 2019:686428. doi: 10.1093/cercor/bhaa089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Jepma M, Nieuwenhuis S. Pupil diameter predicts changes in the exploration-exploitation trade-off: evidence for the adaptive gain theory. J Cogn Neurosci. 2011;23:1587–1596. doi: 10.1162/jocn.2010.21548. [DOI] [PubMed] [Google Scholar]
- 35.Jodo E, Aston-Jones G. Activation of locus coeruleus by prefrontal cortex is mediated by excitatory amino acid inputs. Brain Res. 1997;768:327–332. doi: 10.1016/s0006-8993(97)00703-8. [DOI] [PubMed] [Google Scholar]
- 36.Johansen JP, Diaz-Mataix L, Hamanaka H, Ozawa T, Ycu E, Koivumaa J, Kumar A, Hou M, Deisseroth K, Boyden ES, LeDoux JE. Hebbian and neuromodulatory mechanisms interact to trigger associative memory formation. Proc Natl Acad Sci U S A. 2014;111:E5584–5592. doi: 10.1073/pnas.1421304111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kalwani RM, Joshi S, Gold JI. Phasic activation of individual neurons in the locus ceruleus/subceruleus complex of monkeys reflects rewarded decisions to go but not stop. J Neurosci. 2014;34:13656–13669. doi: 10.1523/JNEUROSCI.2566-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kebschull JM, Garcia da Silva P, Reid AP, Peikon ID, Albeanu DF, Zador AM. High-throughput mapping of single-neuron projections by sequencing of barcoded RNA. Neuron. 2016;91:975–987. doi: 10.1016/j.neuron.2016.07.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kempadoo KA, Mosharov EV, Choi SJ, Sulzer D, Kandel ER. Dopamine release from the locus coeruleus to the dorsal hippocampus promotes spatial learning and memory. Proc Natl Acad Sci U S A. 2016;113:14835–14840. doi: 10.1073/pnas.1616515114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kety SS. The biogenic amines in the central nervous system: Their possible roles in arousal, emotion and learning. In: Schmitt FO, editor. The Neurosciences: Second Study Program. New York: Rockefeller Press; 1970. [Google Scholar]
- 41.Khakpour-Taleghani B, Lashgari R, Motamedi F, Naghdi N. Effect of reversible inactivation of locus ceruleus on spatial reference and working memory. Neuroscience. 2009;158:1284–1291. doi: 10.1016/j.neuroscience.2008.11.001. [DOI] [PubMed] [Google Scholar]
- 42.Lim EP, Tan CH, Jay TM, Dawe GS. Locus coeruleus stimulation and noradrenergic modulation of hippocampo-prefrontal cortex long-term potentiation. Int J Neuropsychopharmacol. 2010;13:1219–1231. doi: 10.1017/S1461145709991131. [DOI] [PubMed] [Google Scholar]
- 43.Lovett-Barron M, Andalman AS, Allen WE, Vesuna S, Kauvar I, Burns VM, Deisseroth K. Ancestral circuits for the coordinated modulation of brain state. Cell. 2017;171:1411–1423. doi: 10.1016/j.cell.2017.10.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Maeda T. The locus coeruleus: history. J Chem Neuroanat. 2000;18:57–64. doi: 10.1016/s0891-0618(99)00051-4. [DOI] [PubMed] [Google Scholar]
- 45.Marien M, Briley M, Colpaert F. Noradrenaline depletion exacerbates MPTP-induced striatal dopamine loss in mice. Eur J Pharmacol. 1993;236:487–489. doi: 10.1016/0014-2999(93)90489-5. [DOI] [PubMed] [Google Scholar]
- 46.Marien MR, Colpaert FC, Rosenquist AC. Noradrenergic mechanisms in neurodegenerative diseases: a theory. Brain Res Brain Res Rev. 2004;45:38–78. doi: 10.1016/j.brainresrev.2004.02.002. [DOI] [PubMed] [Google Scholar]
- 47.Martins AR, Froemke RC. Coordinated forms of noradrenergic plasticity in the locus coeruleus and primary auditory cortex. Nat Neurosci. 2015;18:1483–1492. doi: 10.1038/nn.4090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.McBurney-Lin J, Lu J, Zuo Y, Yang H. Locus coeruleus-norepinephrine modulation of sensory processing and perception: A focused review. Neurosci Biobehav Rev. 2019;105:190–199. doi: 10.1016/j.neubiorev.2019.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.McCall JG, Al-Hasani R, Siuda ER, Hong DY, Norris AJ, Ford CP, Bruchas MR. CRH Engagement of the locus coeruleus noradrenergic system mediates stress-induced anxiety. Neuron. 2015;87:605–620. doi: 10.1016/j.neuron.2015.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Mejias-Aponte CA. Specificity and impact of adrenergic projections to the midbrain dopamine system. Brain Res. 2016;1641:258–273. doi: 10.1016/j.brainres.2016.01.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Mouton PR, Pakkenberg B, Gundersen HJ, Price DL. Absolute number and size of pigmented locus coeruleus neurons in young and aged individuals. J Chem Neuroanat. 1994;7:185–190. doi: 10.1016/0891-0618(94)90028-0. [DOI] [PubMed] [Google Scholar]
- 52.Nassar MR, Rumsey KM, Wilson RC, Parikh K, Heasly B, Gold JI. Rational regulation of learning dynamics by pupil-linked arousal systems. Nat Neurosci. 2012;15:1040–1046. doi: 10.1038/nn.3130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Oliveira LM, Tuppy M, Moreira TS, Takakura AC. Role of the locus coeruleus catecholaminergic neurons in the chemosensory control of breathing in a Parkinson’s disease model. Exp Neurol. 2017;293:172–180. doi: 10.1016/j.expneurol.2017.04.006. [DOI] [PubMed] [Google Scholar]
- 54.Ordway GA, Klimek V. Noradrenergic pathology in psychiatric disorders: postmortem studies. CNS Spectr. 2001;6:697–703. doi: 10.1017/s1092852900001395. [DOI] [PubMed] [Google Scholar]
- 55.Paulus W, Jellinger K. The neuropathologic basis of different clinical subgroups of Parkinson’s disease. J Neuropathol Exp Neurol. 1991;50:743–755. doi: 10.1097/00005072-199111000-00006. [DOI] [PubMed] [Google Scholar]
- 56.Pitman RK, Rasmusson AM, Koenen KC, Shin LM, Orr SP, Gilbertson MW, Milad MR, Liberzon I. Biological studies of post-traumatic stress disorder. Nat Rev Neurosci. 2012;13:769–787. doi: 10.1038/nrn3339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Rahman MK, Rahman F, Rahman T, Kato T. Dopamine-beta-hydroxylase (DBH), its cofactors and other biochemical parameters in the serum of neurological patients in Bangladesh. Int J Biomed Sci. 2009;5:395–401. [PMC free article] [PubMed] [Google Scholar]
- 58.Rajkowski J, Kubiak P, Aston-Jones G. Locus coeruleus activity in monkey: phasic and tonic changes are associated with altered vigilance. Brain Res Bull. 1994;35:607–616. doi: 10.1016/0361-9230(94)90175-9. [DOI] [PubMed] [Google Scholar]
- 59.Reil JC. Untersuchungen über den Bau des grossen Gehirns im Menschen. Arch Physiol. 1908;9:136–208. [Google Scholar]
- 60.Remy P, Doder M, Lees A, Turjanski N, Brooks D. Depression in Parkinson’s disease: loss of dopamine and noradrenaline innervation in the limbic system. Brain. 2005;128:1314–1322. doi: 10.1093/brain/awh445. [DOI] [PubMed] [Google Scholar]
- 61.Reyes BA, Drolet G, Van Bockstaele EJ. Dynorphin and stress-related peptides in rat locus coeruleus: contribution of amygdalar efferents. J Comp Neurol. 2008;508:663–675. doi: 10.1002/cne.21683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Rodenkirch C, Liu Y, Schriver BJ, Wang Q. Locus coeruleus activation enhances thalamic feature selectivity via norepinephrine regulation of intrathalamic circuit dynamics. Nat Neurosci. 2019;22:120–133. doi: 10.1038/s41593-018-0283-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Rommelfanger KS, Weinshenker D. Norepinephrine: The redheaded stepchild of Parkinson’s disease. Biochem Pharmacol. 2007;74:177–190. doi: 10.1016/j.bcp.2007.01.036. [DOI] [PubMed] [Google Scholar]
- 64.Rommelfanger KS, Weinshenker D, Miller GW. Reduced MPTP toxicity in noradrenaline transporter knockout mice. J Neurochem. 2004;91:1116–1124. doi: 10.1111/j.1471-4159.2004.02785.x. [DOI] [PubMed] [Google Scholar]
- 65.Sah P. Fear, anxiety, and the amygdala. Neuron. 2017;96:1–2. doi: 10.1016/j.neuron.2017.09.013. [DOI] [PubMed] [Google Scholar]
- 66.Sales AC, Friston KJ, Jones MW, Pickering AE, Moran RJ. Locus Coeruleus tracking of prediction errors optimises cognitive flexibility: An Active Inference model. PLoS Comput Biol. 2019;15:e1006267. doi: 10.1371/journal.pcbi.1006267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Samuele A, Mangiagalli A, Armentero MT, Fancellu R, Bazzini E, Vairetti M, Ferrigno A, Richelmi P, Nappi G, Blandini F. Oxidative stress and pro-apoptotic conditions in a rodent model of Wilson’s disease. Biochim Biophys Acta. 2005;1741:325–330. doi: 10.1016/j.bbadis.2005.06.004. [DOI] [PubMed] [Google Scholar]
- 68.Sara SJ. Sleep to Remember. J Neurosci. 2017;37:457–463. doi: 10.1523/JNEUROSCI.0297-16.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Sara SJ, Devauges V. Priming stimulation of locus coeruleus facilitates memory retrieval in the rat. Brain Res. 1988;438:299–303. doi: 10.1016/0006-8993(88)91351-0. [DOI] [PubMed] [Google Scholar]
- 70.Sara SJ, Segal M. Plasticity of sensory responses of locus coeruleus neurons in the behaving rat: implications for cognition. Prog Brain Res. 1991;88:571–585. doi: 10.1016/s0079-6123(08)63835-2. [DOI] [PubMed] [Google Scholar]
- 71.Sara SJ, Bouret S. Orienting and reorienting: the locus coeruleus mediates cognition through arousal. Neuron. 2012;76:130–141. doi: 10.1016/j.neuron.2012.09.011. [DOI] [PubMed] [Google Scholar]
- 72.Schmidt K, Ralle M, Schaffer T, Jayakanthan S, Bari B, Muchenditsi A, Lutsenko S. ATP7A and ATP7B copper transporters have distinct functions in the regulation of neuronal dopamine-beta-hydroxylase. J Biol Chem. 2018;293:20085–20098. doi: 10.1074/jbc.RA118.004889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Schmidt K, Bari B, Ralle M, Washington-Hughes C, Muchenditsi A, Maxey E, Lutsenko S. Localization of the locus coeruleus in the mouse brain. J Vis Exp. 2019 doi: 10.3791/58652. doi:103791/58652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Schwarz LA, Luo L. Organization of the locus coeruleus-norepinephrine system. Curr Biol. 2015;25:R1051–1056. doi: 10.1016/j.cub.2015.09.039. [DOI] [PubMed] [Google Scholar]
- 75.Schwarz LA, Miyamichi K, Gao XJ, Beier KT, Weissbourd B, DeLoach KE, Ren J, Ibanes S, Malenka RC, Kremer EJ, Luo L. Viral-genetic tracing of the input-output organization of a central noradrenaline circuit. Nature. 2015;524:88–92. doi: 10.1038/nature14600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Seo DO, Bruchas MR. Polymorphic computation in locus coeruleus networks. Nat Neurosci. 2017;20:1517–1519. doi: 10.1038/nn.4663. [DOI] [PubMed] [Google Scholar]
- 77.Seol GH, Ziburkus J, Huang S, Song L, Kim IT, Takamiya K, Huganir RL, Lee HK, Kirkwood A. Neuromodulators control the polarity of spike-timing-dependent synaptic plasticity. Neuron. 2007;55:919–929. doi: 10.1016/j.neuron.2007.08.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Srinivasan J, Schmidt WJ. Potentiation of parkinsonian symptoms by depletion of locus coeruleus noradrenaline in 6-hydroxydopamine-induced partial degeneration of substantia nigra in rats. Eur J Neurosci. 2003;17:2586–2592. doi: 10.1046/j.1460-9568.2003.02684.x. [DOI] [PubMed] [Google Scholar]
- 79.Stefanis L. α-Synuclein in Parkinson’s disease. Cold Spring Harb Perspect Med. 2012;2:a009399. doi: 10.1101/cshperspect.a009399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Szabadi E. Functional neuroanatomy of the central noradrenergic system. J Psychopharmacol. 2013;27:659–693. doi: 10.1177/0269881113490326. [DOI] [PubMed] [Google Scholar]
- 81.Szymusiak R, McGinty D. Hypothalamic regulation of sleep and arousal. Ann N Y Acad Sci. 2008;1129:275–286. doi: 10.1196/annals.1417.027. [DOI] [PubMed] [Google Scholar]
- 82.Takeuchi T, Duszkiewicz AJ, Sonneborn A, Spooner PA, Yamasaki M, Watanabe M, Smith CC, Fernandez G, Deisseroth K, Greene RW, Morris RG. Locus coeruleus and dopaminergic consolidation of everyday memory. Nature. 2016;537:357–362. doi: 10.1038/nature19325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Traver S, Salthun-Lassalle B, Marien M, Hirsch EC, Colpaert F, Michel PP. The neurotransmitter noradrenaline rescues septal cholinergic neurons in culture from degeneration caused by low-level oxidative stress. Mol Pharmacol. 2005;67:1882–1891. doi: 10.1124/mol.104.007864. [DOI] [PubMed] [Google Scholar]
- 84.Trist BG, Davies KM, Cottam V, Genoud S, Ortega R, Roudeau S, Carmona A, De Silva K, Wasinger V, Lewis SJG, Sachdev P, Smith B, Troakes C, Vance C, Shaw C, Al-Sarraj S, Ball HJ, Halliday GM, Hare DJ, Double KL. Amyotrophic lateral sclerosis-like superoxide dismutase 1 proteinopathy is associated with neuronal loss in Parkinson’s disease brain. Acta Neuropathol. 2017;134:113–127. doi: 10.1007/s00401-017-1726-6. [DOI] [PubMed] [Google Scholar]
- 85.Trist BG, Hare DJ, Double KL. Oxidative stress in the aging substantia nigra and the etiology of Parkinson’s disease. Aging Cell. 2019 doi: 10.1111/acel.13031. doi:101111/acel13031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Troadec JD, Marien M, Mourlevat S, Debeir T, Ruberg M, Colpaert F, Michel PP. Activation of the mitogen-activated protein kinase (ERK(1/2)) signaling pathway by cyclic AMP potentiates the neuroprotective effect of the neurotransmitter noradrenaline on dopaminergic neurons. Mol Pharmacol. 2002;62:1043–1052. doi: 10.1124/mol.62.5.1043. [DOI] [PubMed] [Google Scholar]
- 87.Tubbs RS, Loukas M, Shoja MM, Mortazavi MM, Cohen-Gadol AA. Felix Vicq d’Azyr (1746-1794): early founder of neuroanatomy and royal French physician. Childs Nerv Syst. 2011;27:1031–1034. doi: 10.1007/s00381-011-1424-y. [DOI] [PubMed] [Google Scholar]
- 88.Uematsu A, Tan BZ, Ycu EA, Cuevas JS, Koivumaa J, Junyent F, Kremer EJ, Witten IB, Deisseroth K, Johansen JP. Modular organization of the brainstem noradrenaline system coordinates opposing learning states. Nat Neurosci. 2017;20:1602–1611. doi: 10.1038/nn.4642. [DOI] [PubMed] [Google Scholar]
- 89.van den Brink RL, Pfeffer T, Warren CM, Murphy PR, Tona KD, van der Wee NJ, Giltay E, van Noorden MS, Rombouts SA, Donner TH, Nieuwenhuis S. Catecholaminergic neuromodulation shapes intrinsic MRI functional connectivity in the human brain. J Neurosci. 2016;36:7865–7876. doi: 10.1523/JNEUROSCI.0744-16.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.van den Pol AN, Ghosh PK, Liu RJ, Li Y, Aghajanian GK, Gao XB. Hypocretin (orexin) enhances neuron activity and cell synchrony in developing mouse GFP-expressing locus coeruleus. J Physiol. 2002;541:169–185. doi: 10.1113/jphysiol.2002.017426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Varazzani C, San-Galli A, Gilardeau S, Bouret S. Noradrenaline and dopamine neurons in the reward/effort trade-off: a direct electrophysiological comparison in behaving monkeys. J Neurosci. 2015;35:7866–7877. doi: 10.1523/JNEUROSCI.0454-15.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Verbaan D, van Rooden SM, Visser M, Marinus J, van Hilten JJ. Nighttime sleep problems and daytime sleepiness in Parkinson’s disease. Mov Disord. 2008;23:35–41. doi: 10.1002/mds.21727. [DOI] [PubMed] [Google Scholar]
- 93.Von Coelln R, Thomas B, Savitt JM, Lim KL, Sasaki M, Hess EJ, Dawson VL, Dawson TM. Loss of locus coeruleus neurons and reduced startle in parkin null mice. Proc Natl Acad Sci U S A. 2004;101:10744–10749. doi: 10.1073/pnas.0401297101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Wagatsuma A, Okuyama T, Sun C, Smith LM, Abe K, Tonegawa S. Locus coeruleus input to hippocampal CA3 drives single-trial learning of a novel context. Proc Natl Acad Sci U S A. 2018;115:E310–316. doi: 10.1073/pnas.1714082115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Wang J, Li Y, Huang Z, Wan W, Zhang Y, Wang C, Cheng X, Ye F, Liu K, Fei G, Zeng M, Jin L. Neuromelanin-sensitive magnetic resonance imaging features of the substantia nigra and locus coeruleus in de novo Parkinson’s disease and its phenotypes. Eur J Neurol. 2018;25:949–973. doi: 10.1111/ene.13628. [DOI] [PubMed] [Google Scholar]
- 96.Weintraub D, Simuni T, Caspell-Garcia C, Coffey C, Lasch S, Siderowf A, Aarsland D, Barone P, Burn D, Chahine LM, Eberling J, Espay AJ, Foster ED, Leverenz JB, Litvan I, Richard I, Troyer MD, Hawkins KA Parkinson’s Progression Markers I. Cognitive performance and neuropsychiatric symptoms in early, untreated Parkinson’s disease. Mov Disord. 2015;30:919–927. doi: 10.1002/mds.26170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Wong ML, Kling MA, Munson PJ, Listwak S, Licinio J, Prolo P, Karp B, McCutcheon IE, Geracioti TD, DeBellis MD, Rice KC, Goldstein DS, Veldhuis JD, Chrousos GP, Oldfield EH, McCann SM, Gold PW. Pronounced and sustained central hypernoradrenergic function in major depression with melancholic features: relation to hypercortisolism and corticotropin-releasing hormone. Proc Natl Acad Sci U S A. 2000;97:325–330. doi: 10.1073/pnas.97.1.325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Xiang L, Harel A, Gao H, Pickering AE, Sara SJ, Wiener SI. Behavioral correlates of activity of optogenetically identified locus coeruleus noradrenergic neurons in rats performing T-maze tasks. Sci Rep. 2019;9:1361. doi: 10.1038/s41598-018-37227-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Yavich L, Jakala P, Tanila H. Abnormal compartmentalization of norepinephrine in mouse dentate gyrus in alpha-synuclein knockout and A30P transgenic mice. J Neurochem. 2006;99:724–732. doi: 10.1111/j.1471-4159.2006.04098.x. [DOI] [PubMed] [Google Scholar]
- 100.Zarow C, Lyness SA, Mortimer JA, Chui HC. Neuronal loss is greater in the locus coeruleus than nucleus basalis and substantia nigra in Alzheimer and Parkinson diseases. Arch Neurol. 2003;60:337–341. doi: 10.1001/archneur.60.3.337. [DOI] [PubMed] [Google Scholar]
- 101.Zecca L, Stroppolo A, Gatti A, Tampellini D, Toscani M, Gallorini M, Giaveri G, Arosio P, Santambrogio P, Fariello RG, Karatekin E, Kleinman MH, Turro N, Hornykiewicz O, Zucca FA. The role of iron and copper molecules in the neuronal vulnerability of locus coeruleus and substantia nigra during aging. Proc Natl Acad Sci U S A. 2004;101:9843–9848. doi: 10.1073/pnas.0403495101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Zerbi V, Floriou-Servou A, Markicevic M, Vermeiren Y, Sturman O, Privitera M, von Ziegler L, Ferrari KD, Weber B, De Deyn PP, Wenderoth N, Bohacek J. Rapid reconfiguration of the functional connectome after chemogenetic locus coeruleus activation. Neuron. 2019;103:702–718e5. doi: 10.1016/j.neuron.2019.05.034. [DOI] [PubMed] [Google Scholar]
- 103.Zhou J. Norepinephrine transporter inhibitors and their therapeutic potential. Drugs Future. 2004;29:1235–1244. doi: 10.1358/dof.2004.029.12.855246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Zhu K, van Hilten JJ, Marinus J. Onset and evolution of anxiety in Parkinson’s disease. Eur J Neurol. 2017;24:404–411. doi: 10.1111/ene.13217. [DOI] [PubMed] [Google Scholar]