

Abstract
Excitation-contraction coupling (ECC) is a physiological process that links excitation of muscles by the nervous system to their mechanical contraction. In skeletal muscle, ECC is initiated with an action potential, generated by the somatic nervous system, which causes a depolarisation of the muscle fibre membrane (sarcolemma). This leads to a rapid change in the transmembrane potential, which is detected by the voltage-gated Ca2+ channel dihydropyridine receptor (DHPR) embedded in the sarcolemma. DHPR transmits the contractile signal to another Ca2+ channel, ryanodine receptor (RyR1), embedded in the membrane of the sarcoplasmic reticulum (SR), which releases a large amount of Ca2+ ions from the SR that initiate muscle contraction. Despite the fundamental role of ECC in skeletal muscle function of all vertebrate species, the molecular mechanism underpinning the communication between the two key proteins involved in the process (DHPR and RyR1) is still largely unknown. The goal of this work is to review the recent progress in our understanding of ECC in skeletal muscle from the point of view of the structure and interactions of proteins involved in the process, and to highlight the unanswered questions in the field.
Keywords: Excitation-contraction coupling, DHPR, CaV1.1, RyR1, STAC3
Overview of the excitation-contraction coupling in skeletal muscle
Excitation-contraction coupling (ECC) is an essential process in muscle physiology, responsible for linking electrical signals from the somatic nervous system (action potentials) to mechanical muscle contractions (Sandow 1952). In skeletal muscle, the ECC is initiated at the neuromuscular junction, where a motor neuron connects to a muscle fibre (a multinucleate cell), by the release of the neurotransmitter acetylcholine (ACh) from the axon terminal. Upon diffusing to the sarcolemma (muscle cell membrane), ACh binds to ligand-gated cation channels (ACh receptors) that initiate an action potential in the muscle fibre. The action potential travels down the specialised invaginations of the sarcolemma, called the transverse tubules (T tubules), causing a depolarisation of the T-tubular membrane. In skeletal muscle, the T tubules are flanked from their two sides by the terminal cisternae of the sarcoplasmic reticulum (SR), forming so-called triad junctions (Fig. 1). The rapid change in the electric potential across the T-tubular membrane is detected by the voltage-gated l-type Ca2+ channel CaV1.1 (also known as the dihydropyridine receptor, DHPR), which is embedded in the sarcolemma (Adams et al. 1990; Rios and Brum 1987; Tanabe et al. 1988). Upon changing its conformational state, DHPR communicates the contractile signal to the cation channel ryanodine receptor (RyR1), embedded in the membrane of SR, via mechanical interactions (Meissner and Lu 1995; Takeshima et al. 1994). This causes RyR1 to open up and release large amounts of Ca2+ from the SR into the sarcoplasm (Rebbeck et al. 2014). Finally, the released Ca2+ ions bind to troponin C, which changes conformation of the troponin complex. This, in turn, initiates the formation of cross-bridges between contractile proteins actin and myosin, causing them to slide along each other, leading to muscle contractions.
Fig. 1.

Schematic of a triad junction in a skeletal muscle fibre. The interaction between the dihydropyridine receptor (DHPR) and type 1 ryanodine receptor (RyR1) is central to the molecular mechanism of the excitation-contraction coupling in skeletal muscle. Structurally, a single RyR1 tetramer attached to the membrane of the sarcoplasmic reticulum aligns opposite to a tetrad of DHPR channels embedded in the sarcolemma, suggesting a mechanical nature of the interactions between these two Ca2+ channels (Block et al. 1988; Paolini et al. 2004)
The molecular mechanisms of the ECC in skeletal muscle, in terms of protein-protein interactions involved in this process, have been previously covered in a number of excellent reviews (Bannister 2007; Bannister 2016; Calderón et al. 2014; Dulhunty et al. 2002; Rebbeck et al. 2014). Structurally, clusters of four DHPR channels (tetrads) are formed in the T-tubular sections of the sarcolemma, with each DHPR in the tetrad facing one monomeric unit of the tetrameric RyR1 embedded in the SR membrane at the triad junctions (however, interestingly, only half of all RyR1 tetramers face DHPR tetrads) (Fig. 1). This specific arrangement suggests direct mechanical interactions between DHPR tetrads and RyR1 channels (Block et al. 1988; Paolini et al. 2004). However, even though the main sequence of events in the ECC process has been known for several decades, the molecular mechanisms underlying the precise nature of the interactions between DHPR and RyR1 has remained a mystery for many years (Beam and Bannister 2010; Rebbeck et al. 2014).
Hereditary and acquired defects in the ECC proteins are known to compromise muscle development and performance, thereby causing pathological conditions such as congenital myopathies (Jungbluth et al. 2018; Pancaroglu and Van Petegem 2018). For example, several mutations in RyR1 have been linked to malignant hyperthermia, central core disease and multi-minicore disease (Jungbluth et al. 2018). Our lack of understanding of the precise molecular mechanisms of the skeletal ECC impedes our ability to develop new ways of mitigating detrimental effects of these inherited disorders as well as age-related muscle weakness (sarcopenia). Thus, there is a strong motivation to advance our understanding of the ECC on the molecular level and how it affects other aspects of skeletal muscle biology. Recent developments in the field have made significant progress in resolving this long-standing research problem. Nevertheless, there are still major gaps in our understanding of the molecular mechanism of the skeletal ECC. The aim of this work is to review the known molecular interactions that underpin the ECC process in skeletal muscle and highlight the remaining unanswered questions, with a focus on the most recent developments.
Proteins involved in the skeletal excitation-contraction coupling
DHPR and RyR1 have been known as the main players in the skeletal ECC for several decades (Rebbeck et al. 2014). However, multiple attempts to resolve the molecular details of the interactions underpinning the communication between these two Ca2+ channels have essentially failed (Bannister 2007; Bannister 2016). Thus, many researchers speculated about the involvement of other molecules in mediating this interaction. Until a few years ago, it was unknown whether there are any other proteins (apart from subunits of DHPR/RyR1) that play a crucial role in the signal transmission from DHPR to RyR1 that is central to the mechanism of skeletal ECC. This situation changed in 2013, when a new essential component of the skeletal ECC was uncovered — an adaptor protein STAC3 (Horstick et al. 2013; Nelson et al. 2013). This revived an interest in uncovering the precise nature of the interactions between DHPR, STAC3 and RyR1 in skeletal muscle.
Furthermore, in a recent study, Perni et al. (2017) managed to fully reconstruct functional skeletal ECC in non-muscle cells. This was achieved by heterologous expression of α1s and β1a-subunits of DHPR, RyR1, STAC3 and junctophilin2 in human embryonic kidney cells tsA201, thus demonstrating that these five components constitute the complete set of partners required for a fully functional ECC. This recent report suggested that a detailed study of the interactions between this set of five proteins (DHPR-α1s, DHPR-β1a, RyR1, STAC3 and junctophilin2) will allow full resolution of the detailed molecular mechanism of the ECC in skeletal muscle. While it is possible that other proteins endogenously present in tsA201 cells might also be required for the skeletal ECC, this set of five partners appears to be absolutely essential for the process and thus each of them warrants an in-depth consideration. Accordingly, these individual ECC components are considered in turn in the following sections.
Dihydropyridine receptor
DHPR architecture
DHPR has been known as the critical component of the ECC machinery for > 30 years upon its identification as the ECC voltage sensor by Rios and Brum (1987). The main role of skeletal DHPR is to detect the action potential in the T-tubular part of the sarcolemma and communicate this signal to the RyR1, which then releases Ca2+ ions from the SR (Dulhunty et al. 2002; Paolini et al. 2004; Protasi 2002). Additionally, DHPR functions as a voltage-gated l-type Ca2+ channel (CaV1.1). However, its function as a Ca2+ channel has been shown to be irrelevant for skeletal ECC (Dirksen and Beam 1999) and, more recently, to muscle development and performance in general (Dayal et al. 2017). Unlike cardiomyocytes, in skeletal muscle, it is not the Ca2+ current via DHPR, but the conformational change of the protein upon depolarisation of the sarcolemma that is responsible for the transmission of the contractile signal to RyR1. This mechanical coupling between DHPR and RyR1 is bi-directional, as RyR1 is known to have an effect on Ca2+ currents via DHPR through the ‘retrograde’ coupling effect (Dirksen 2002; Nakai et al. 1996).
Several years ago, the group of Nieng Yan determined the structure of rabbit DHPR at a resolution of 4.2 Å (Wu et al. 2015) and 3.6 Å (Wu et al. 2016) using single-particle cryo-electron microscopy (cryo-EM). This provided some important details about the principal architecture of the protein (Wu et al. 2016). Skeletal DHPR consists of four principal subunits: α1s, α2δ, β1a and γ (Catterall 2011). α1s is the major, membrane-embedded pore-forming subunit also referred to as CaV1.1. The other three subunits are auxiliary: γ is membrane-associated and forms contacts with one of the transmembrane domains of α1s (Wu et al. 2016), α2δ is extracellular and interacts with the extended extracellular loops of α1s (Wu et al. 2016), while β1a is bound to α1s from the intracellular side (Chen et al. 2004; Opatowsky et al. 2004; Van Petegem et al. 2004). Both α1s and β1a are known to be critical for the ECC (Coronado et al. 2004), while α2δ and γ subunits play roles in membrane targeting and regulatory functions of DHPR, but they are not essential for the ECC (Obermair et al. 2008).
DHPR-α1s
α1s subunit of DHPR is critical for the ECC as it contains the voltage sensor that changes its conformation upon sensing depolarisation of the membrane. The architecture of α1s is similar to that of other CaV channels: it is composed of four homologous membrane-spanning motifs designated as I, II, III and IV (Fig. 2). Each of these motifs contains six transmembrane helices, denoted as S1–S6. It is the fourth helix S4 that contains several positively charged residues (Arg and Lys) participating in the voltage sensing mechanism. Additionally, there are five major cytosolic regions of CaV1.1: the N-terminal domain (NTD), the linkers between repeats I and II, II and III, III and IV, and the C-terminal domain (CTD) (Fig. 2).
Fig. 2.

Topology of the membrane-embedded pore-forming α1s-subunit of the skeletal muscle dihydropyridine receptor (CaV1.1). Each of the membrane-spanning motifs I–IV is composed of six transmembrane helices S1–S6. The five cytosolic regions of the protein include the N-terminal domain (NTD), I–II loop, II–III loop, III–IV loop and the C-terminal domain (CTD)
Apart from the voltage sensor in the core of α1s, the linker between domains II and III (II–III loop) is known to be critical for the skeletal ECC (Tanabe et al. 1990). The central portion of the II–III loop (designated as ‘peptide C’ (El-Hayek et al. 1995)) contains an amino acid sequence that was identified to have the stretch of residues (720–765) essential for skeletal ECC (Grabner et al. 1999; Nakai et al. 1998b; Wilkens et al. 2001). In fact, Kugler et al. (2004) demonstrated that substitution of only four critical residues in this portion of the loop (A739, F741, P742 and D744) suppresses ECC.
Later biophysical studies demonstrated that the N-terminal part of the II–III loop of DHPR-α1s (residues 671–690) interacts with RyR1 (Cui et al. 2009; Tae et al. 2009). This part of the II–III loop (‘peptide A’) was previously shown to activate RyR1 channels in vitro (El-Hayek et al. 1995), and a later study determined the minimum motif (681–690) required for this interaction (El-Hayek and Ikemoto 1998). Subsequent studies showed that this N-terminal part of the II-III loop is not essential for the ECC (Bannister et al. 2009; Proenza et al. 2000b; Wilkens et al. 2001); nevertheless, the interaction of the N-terminal part of the II–III loop with RyR1 was reported to have at least some significance for the ECC process (Ahern et al. 2001). Thus, the physiological role of the N-terminal part of the II–III loop with respect to ECC remains ill-defined.
Overall, the fine details of how the II–III loop is involved in the ECC are still unknown. It is missing in the recent cryo-EM structures of DHPR (Wu et al. 2016; Wu et al. 2015), most likely due to its inherently disordered nature (Casarotto et al. 2006; Cui et al. 2009). It is possible, however, that the structure of the II–III loop changes upon the conformational change in the core α1s during the action potential (Polster et al. 2012). Thus, a structural change in the II–III loop might be responsible for the transmission of the contractile signal to RyR1. However, despite an extensive search, the corresponding interacting residues of RyR1 have not been unambiguously identified (Rebbeck et al. 2014). In fact, there is still a lack of strong evidence that the II–III loop interacts directly with the RyR1 in a manner that is important for the ECC. This raised the possibility of other proteins directly participating in the coupling between DHPR and RyR1. Indeed, recent studies identified interactions of the critical peptide from the II–III loop with STAC3, which suggested a direct role of this adaptor protein in the transmission of the ECC signal in skeletal muscle (see below the section for STAC3 protein).
The linker between domains I and II of α1s binds another ECC-critical subunit of DHPR (β1a) (Pragnell et al. 1994), and this interaction between β1a and the I–II loop is required for targeting of β1a to the triad junctions (Neuhuber et al. 1998). The III–IV linker forms a globular domain in the complex with the C-terminal domain (CTD) of α1s (Wu et al. 2016) and influences ECC indirectly through its ability to modulate channel gating (Bannister et al. 2008; Weiss et al. 2004), while the CTD is also required for targeting of DHPR to the triad junctions (Flucher et al. 2000; Proenza et al. 2000a). The N-terminal cytosolic part of α1s is not essential for the ECC (Bannister and Beam 2005).
DHPR-β1a
As introduced above, the cytosolic β1a subunit binds with a high affinity to a sequence in the I–II loop of DHPR-α1s (residues 357–374) that forms an α-helix designated as the AID (alpha-interacting domain). Multiple reports identified that β1a is critical for the ECC process (Beurg et al. 1999; Gregg et al. 1996; Schredelseker et al. 2009; Strube et al. 1996). β1a-knockout mice die perinatally due to their inability to breathe (Gregg et al. 1996), as β1a is required for correct assembly of the DHPR arrays in the triad junction (Schredelseker et al. 2005) and β1a-null muscles do not elicit RyR1-mediated Ca2+ transients upon electric stimulation (Strube et al. 1996).
In mammals, there are four distinct isoforms for the β-subunits (β1–β4) (Buraei and Yang 2010), but β1a is specific to skeletal muscle and the only isoform capable of mediating the skeletal-type ECC (Schredelseker et al. 2009). Structurally, β-subunits belong to a family of membrane-associated guanylate kinase (MAGUK) proteins (Karunasekara et al. 2009; Norris et al. 2017; Van Petegem et al. 2004). They consist of conserved and structured Src homology 3 (SH3) and guanylate kinase (GK) domains, which interact with each other via hydrogen bonds and van der Waals interactions, thus forming the stable core of the protein (Opatowsky et al. 2003). β-Subunits also contain less conserved and mostly disordered N-terminal and C-terminal domains, and the so-called HOOK region which forms a long loop within the SH3 domain (Dolphin 2003) (Fig. 3).
Fig. 3.
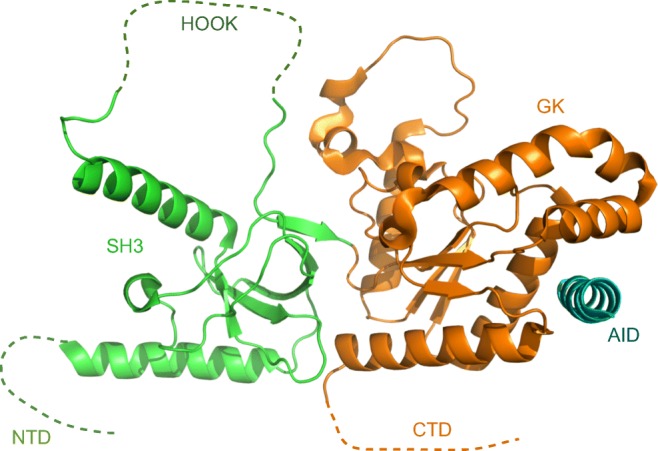
Structure of the β1a-subunit of the skeletal muscle dihydropyridine receptor in complex with the alpha-interacting domain (AID) peptide. The five principal domains of β1a are indicated: the N-terminal domain (NTD), Src homology 3 (SH3), the HOOK region, guanylate kinase (GK) and the C-terminal domain (CTD). Adapted from the PDB structure 4ZW2 (Norris et al. 2017)
The SH3 domains are known to bind proline-rich motifs thus mediating protein-protein interactions (Mayer 2001). However, the canonical binding site in the SH3 domains of β-subunits is occluded by an α-helix leading to the HOOK region, thus making direct interactions of this domain with other proteins unlikely (Chen et al. 2004; Van Petegem et al. 2004). Nevertheless, the SH3 domain and the proximal C-terminus of β1a were found to work cooperatively to enable α1s to act as a voltage sensor, thus playing an important role in the ECC (Dayal et al. 2013). The GK domain is structurally related to the nucleotide monophosphate kinase family of proteins; however, in β-subunits, this domain is catalytically inactive. It is the GK domain that contains a hydrophobic groove that binds the AID sequence of the I–II loop of the DHPR-α1s with a nanomolar affinity (Richards et al. 2004; Van Petegem et al. 2004), with the AID domain being sandwiched in between the α1s and β1a domains of the DHPR complex (Wu et al. 2016).
While it is possible that the ECC signal is transmitted from DHPR-α1s to RyR1 via the DHPR-β1a/AID complex, it is unknown whether the structural change in α1s might confer an allosteric change in DHPR-β1a via its interaction with AID. Even though crystal structures of homologous β2–β4 were determined > 15 years ago (Chen et al. 2004; Opatowsky et al. 2004; Van Petegem et al. 2004), a high-resolution structure of the β1a-subunit in a complex with AID was determined only recently (Fig. 3), and the detailed structure of β1a in the absence of AID still remains unknown (Norris et al. 2017). In fact, it is quite likely that β1a is distinct among the β-subunits in that its three-dimensional fold is significantly altered by the interaction with AID, as it has an 8 °C lower melting point in the absence of the peptide (Norris et al. 2017). A perturbation in the conformation or position of AID during the ECC is quite likely as it is separated from the transmembrane helix S6 of the DHPR repeat I by only a small number of residues (Fig. 2). In turn, any perturbation in the conformation or position of AID during the ECC might cause an allosteric change in β1a that is subsequently transmitted to RyR1. However, as a high-resolution structure of DHPR-β1a on its own (without AID) has not been determined yet, the structural role of the DHPR-β1a/AID interaction in the ECC process remains unclear.
Several studies suggested that the C-terminal domain of β1a binds directly to RyR1 in an ECC-relevant interaction. In particular, the distal C-terminal part of β1a has been proposed to be critical (Coronado et al. 2004), as deletion of the 35-residue C-terminal tail (residues 490–524) resulted in a 5-fold reduction in the ECC efficiency (Beurg et al. 1999). In later studies, affinity chromatography showed that this 35-residue peptide can pull-down the whole RyR1 and activate the channel in planar lipid bilayers (Karunasekara et al. 2012; Rebbeck et al. 2011). Moreover, more recent studies showed that a shorter β1a peptide from the same region (490–508) was able to activate the RyR1 (Hernández-Ochoa et al. 2014), and residues 489–503 were determined to be critical for communication between DHPR and RyR1 (Eltit et al. 2014); nevertheless, the corresponding interacting interface of RyR1 has not been determined. In another study, Sheridan et al. (2004) reported that simultaneous mutation of the three hydrophobic residues forming a ‘heptad repeat’ within the C-terminal sequence of β1a (L478A, V485A, V492A) suppressed the skeletal ECC. However, a later study showed that the same combined mutations did not affect the ECC in β1a-null zebrafish myotubes (Dayal et al. 2010). Overall, despite a consensus regarding the importance of the C-terminal portion of DHPR-β1a in the ECC process, the corresponding interacting residues in RyR1 have not been determined, and the exact structural function of the β1a-subunit in the signal transmission to RyR1 remains unresolved (Bannister 2016).
In summary, while the role of the core DHPR-α1s as the voltage sensor in the ECC is well established, it is still unknown whether DHPR-β1a or the II–III loop plays an important role in the transmission of the signal to RyR1 in the ECC process.
Ryanodine receptor
Ryanodine receptor is the largest known ion channel in mammals: it is composed of four identical subunits, each with the molecular weight of ~ 560 kDa (Lanner et al. 2010). In 2015, three groups simultaneously reported cryo-EM structures of RyR1 with an overall resolution of < 5 Å (Efremov et al. 2015; Yan et al. 2015; Zalk et al. 2015). RyR1 has a pore structure similar to that of K+ channels, and it is permeable to both monovalent and divalent cations. The channel has a mushroom-like appearance, with the N-terminal domain forming 90% of the structure which is located in the sarcoplasmic gap between the terminal cisternae of the SR and the sarcolemma, while the remaining C-terminal domain is embedded in the SR membrane and forms the channel pore (Fig. 4).
Fig. 4.

Overview of the proteins and interactions involved in the excitation-contraction coupling in skeletal muscle. The cartoon is based on the cryo-EM structure of the DHPR complex (PDB 5GJV) (Wu et al. 2016), cryo-EM structure of RyR1 (PDB 3J8E) (Zalk et al. 2015), X-ray structure of the tandem SH3 domain of STAC2 in the complex with the II–III loop peptide (PDB 6B27) (Yuen et al. 2017) and the NMR structure of the C1 domain of STAC3 (PDB 2DB6). The established interactions include those between the GK domain of DHPR-β1a and the AID peptide of the DHPR-α1s I–II loop (Norris et al. 2017), between the C1 domain of STAC3 and the C-terminal domain (CTD) of DHPR-α1s (Campiglio et al. 2018a; Campiglio and Flucher 2017), and the interaction between the central part of the II–III loop and the first SH3 domain of STAC3 (Polster et al. 2018; Yuen et al. 2017). Interactions of RyR1 with either DHPR or STAC3 remain ambiguous
In mammals, there are two other isoforms of ryanodine receptors called RyR2 and RyR3 which are primarily expressed in the cardiac and neuronal tissues, respectively (Franzini-Armstrong and Protasi 1997). However, the three isoforms are sufficiently different and only RyR1 is capable of mediating the skeletal-type ECC (Nakai et al. 1997). This allowed RyR1/RyR2 and RyR1/RyR3 chimaera studies using dyspedic myotubes (naturally lacking RyR) to identify regions in RyR1 that are important for the ECC in skeletal muscle. In an early study, Nakai et al. (1998a) suggested that residues 1635–2636 of RyR1 are important for the bi-directional coupling with DHPR, while Yamazawa et al. (1997) reported that deletion of a small region of RyR1 that is poorly conserved between the three RyR isoforms (1303–1406) abolishes the ECC in skeletal muscle. In line with the latter study, Sheridan et al. (2006) demonstrated that residues 1272–1455 of RyR1 were important for DHPR tetrad formation; however, a chimaera containing residues 1–1681 of RyR1 on the background of RyR3 only partially restored the ECC, indicating the importance of multiple interactions between DHPR and RyR1 (Sheridan et al. 2006).
In another study, a weak interaction between the critical region of the II–III loop of DHPR-α1s and residues 1835–2154 of RyR1 was identified (Proenza et al. 2002). However, the chimaera of these residues of RyR1 on the background of RyR2 restored only a weak ECC (Proenza et al. 2002), suggesting a requirement for additional sequences in RyR1 for the proper ECC. Indeed, further studies showed that a much larger sequence of residues 1635–3720 was necessary for full restoration of skeletal ECC, again leading to a conclusion that multiple regions of RyR1 are essential for this process (Perez et al. 2003; Protasi et al. 2002; Sheridan et al. 2006). Furthermore, an ECC-enhancing interaction between DHPR-β1a and a short sequence of RyR1 (3495–3502) was reported (Cheng et al. 2005). Nevertheless, when the proposed interacting residues in RyR1 have been substituted, the interaction persisted (Rebbeck et al. 2014).
In summary, multiple interactions between DHPR and RyR1 during the ECC process might be at play, as several regions of RyR1 appear to be essential for the ECC. Moreover, as introduced above, STAC3 might also play an important role in mediating these interactions, which is discussed in the next section.
STAC3
STAC3 is a member of a small group of STAC (SH3 and cysteine-rich domain-containing) proteins, consisting of STAC1, STAC2 and STAC3 (Flucher and Campiglio 2018). STAC1 and STAC2 are predominantly located in neural tissues, while the STAC3 isoform is specific to skeletal muscle (Nelson et al. 2013). In the context of muscle physiology, STAC3 was initially reported as an important regulator of myotube formation and myogenic differentiation (Bower et al. 2012; Ge et al. 2014), with homozygous STAC3-knockout mice dying at birth (Reinholt et al. 2013). In 2013, the knockout studies in zebrafish (Horstick et al. 2013) and mice (Nelson et al. 2013) revealed STAC3 as an essential component of the skeletal muscle ECC.
STAC3 is a 364-residue protein consisting of a mostly disordered N-terminal domain which contains a sequence of 12 consecutive glutamate residues, a protein kinase C-like domain (PKC C1) which is rich in cysteines, the flexible linker, and the tandem-SH3 domains at the C-terminus (designated as SH3_1 and SH3_2) (Fig. 4). The structure of the Zn2+-binding PKC C1 domain was determined by NMR spectroscopy and deposited to the PDB database in 2006 (PDB ID: 2DB6). Recently, crystal structures of STAC1 and STAC2 tandem-SH3 domains were determined by X-ray crystallography to the resolution of 2.4 and 1.2 Å, respectively (Yuen et al. 2017). However, STAC3 tandem-SH3 resisted crystallisation, and only the structure of the individual 2nd SH3 domain of STAC3 (SH3_2) was determined (Yuen et al. 2017). The structure of SH3_2 was similar for all three STAC proteins, and it was speculated that the structure of the tandem SH3 domains of STAC3 would be quite similar to those of STAC1 and STAC2 (Yuen et al. 2017).
Several recent reports strongly suggested that the DHPR-RyR1 interactions in the ECC process are in fact mediated by STAC3 (Flucher and Campiglio 2018). The initial zebrafish study showed that STAC3 co-immunoprecipitates with both DHPR and RyR1 (Horstick et al. 2013), and it is required for voltage-dependent Ca2+ release from the SR (Nelson et al. 2013). Moreover, a mutation in SH3_1 of STAC3 (W284S) results in misregulation of the DHPR channels (Linsley et al. 2017), disruption of ECC (Polster et al. 2016) and the direct cause of a severe muscle disease known as Native American myopathy (NAM) (Horstick et al. 2013).
Several years ago, Polster et al. (2015) demonstrated that STAC3 facilitates expression and membrane targeting of CaV1.1 in tsA201 cells, and later, this function of STAC3 was shown to be due to the interaction of the PKC C1 domain of STAC3 with the C-terminal domain of the DHPR-α1s (Campiglio et al. 2018a; Campiglio and Flucher 2017). This interaction was also shown to play a role in the inhibition of calcium-dependent inactivation of CaV channels (Campiglio et al. 2018a; Niu et al. 2018a; Niu et al. 2018b); however, it does not appear to be directly relevant to the ECC. In a follow-up work, Polster et al. (2016) showed that the STAC3 is not absolutely required for membrane targeting of CaV1.1 in tsA201 cells, as this role could also be accomplished by the γ-subunit of DHPR; however, STAC3-null myotubes with properly membrane-targeted CaV1.1 did not have functional ECC, thus implicating the direct role of STAC3 in mediating the interactions between DHPR and RyR1 (Polster et al. 2016).
Recently, it has been revealed that the II–III loop of DHPR interacts with STAC3 (Yuen et al. 2017), and this interaction appears to be critical for the ECC (Polster et al. 2018). According to the determined crystal structure of STAC2 with a peptide from the II–III loop of DHPR-α1s (residues 747–760), the interaction is mostly with SH3_1. Notably, mutation of the three critical residues in the II–III loop peptide (I752A, P753A and R757A) abolished the interaction (Yuen et al. 2017). The binding also involves the critical tryptophan residue which corresponds to the residue in STAC3 mutated in NAM (W284) (Yuen et al. 2017). Nevertheless, more recent co-immunoprecipitation studies showed that the NAM mutation does not significantly compromise association between STAC3 and DHPR (Zaharieva et al. 2018), possibly due to an intact interaction via the C1 domain of STAC3. As the canonical binding site of SH3_2 remains unoccupied, it might potentially interact with other proteins, including RyR1 (Yuen et al. 2017). Indeed, recent indirect evidence hints that STAC3 might interact with RyR1 as it incorporates into triad junctions independently from the DHPR (Campiglio et al. 2018b). Nevertheless, a direct interaction between STAC3 and RyR1 is yet to be reported.
In summary, there are several established interactions of STAC3 with DHPR (PKC C1 domain of STAC3 with the CTD of DHPR-α1s and SH3_1 with the II–III loop of DHPR-α1s). There is good amount of evidence suggesting that the interaction of STAC3 with the II–III loop of DHPR-α1s is critical for the ECC, confirming an essential role of STAC3 in mediating the interactions between DHPR and RyR1 (Campiglio et al. 2018b; Yuen et al. 2017). However, the lack of the available structural information on the tandem-SH3 domains of the STAC3 isoform (either in the absence or presence of the II–III loop peptide) prevents the understanding of any direct implication of this interaction in the ECC process. Moreover, any unambiguous evidence for the interaction between STAC3 and RyR1 is still missing. Thus, despite these significant recent advances, the exact functional role of STAC3 in skeletal ECC remains unknown.
Junctophilins
While DHPR, RyR1 and STAC3 appear to be directly responsible for the transduction of the action potential to the release of Ca2+ from the SR in the ECC process, the primary role of junctophilins (JPs) is to maintain the structure of junctions between the plasma membrane and sarcoplasmic reticulum (Takeshima et al. 2000). There are two isoforms of junctophilins (JPs) expressed in skeletal muscles — JP1 and JP2 (Nishi et al. 2000). The N-terminal part of the JPs contains the so-called MORN (membrane occupation and recognition nexus) motif that binds the sarcolemma, while the C-terminal part of JPs binds to the membrane of the SR. This allows formation of junctions between the T-tubular membranes and the terminal cisternae of the SR, and proteolysis of JPs was shown to cause disruption of the triad junctions and suppression of ECC (Murphy et al. 2013). JP1 has been reported to co-immunoprecipitate with RyR1 (Phimister et al. 2007), and both JP1 and JP2 were reported to interact with the DHPR (Golini et al. 2011). Recently, the interaction of junctophilins with the C-terminus of DHPR has been shown to be essential for muscle contraction (Nakada et al. 2018). Overall, these interactions of JPs appear to be important for the formation of the triad junctions, and thus, they are relevant for the maintenance of the functional ECC.
Summary and outlook
In summary, it has been recently revealed that there are five core requisite components in the skeletal ECC machinery: α1s and β1a subunits of DHPR, RyR1, STAC3 and junctophilin2 (Perni et al. 2017). The major established and putative interactions are summarised in Fig. 4. While binding of β1a-subunit to DHPR-α1s via the AID peptide is well established (Norris et al. 2017; Wu et al. 2015), its functional relevance in the transduction of the ECC signal remains ambiguous. The other interaction that appears to be important for the ECC is the interaction between the first SH3 domain of STAC3 and the II–III loop of DHPR-α1s. However, there are still no definitive data clearly showing ECC-relevant interactions of RyR1 with either STAC3 or any of the subunits of DHPR. Thus, overall, despite ~ 30 years of research after the identification of DHPR and RyR1 as key ECC partners, the precise nature of molecular interactions between these two proteins in skeletal muscle remains mostly unresolved (Bannister 2016; Rebbeck et al. 2014; Yuen et al. 2017). As discussed, one of the major hurdles for further progress in our understanding of the ECC mechanism is the fact that the exact three-dimensional structures of DHPR-β1a (without AID) and STAC3 (tandem-SH3 domains) are still unavailable. Another factor that makes this research problem difficult is the fact that ryanodine receptor is a huge protein and multiple interactions of RyR1 appear to be at play, making it difficult to dissect the exact interface or amino acid residues that participate in the ECC. Future structural, biophysical and physiological studies should provide better understanding of the molecular mechanism underlying the interactions between the ECC proteins. The holy grail of the field is to determine a high-resolution structure of a complex containing DHPR and RyR1, potentially together with STAC3 and any of the junctophilins or other components. The rapidly evolving field of cryo-EM might be instrumental in resolving this long-standing issue. Delineation of the molecular details of the communications between DHPR and RyR1 in the ECC process will not only answer a major outstanding question in field, but will have a strong impact in designing rational therapies for the ECC-related pathological conditions.
Acknowledgements
Investigator Grant fellowship (GNT1173015) to DS from the National Health & Medical Research Council (NHMRC) of Australia is acknowledged. The author would like to thank A/Prof. Marco G. Casarotto and Prof. Angela F. Dulhunty for proofreading the manuscript and many useful discussions on the topic of the review.
Compliance with ethical standards
Conflict of interest
The author declares that he has no conflict of interest.
Ethical approval
This article does not contain any studies with human participants or animals performed by the author.
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- Adams BA, Tanabe T, Mikami A, Numa S, Beam KG. Intramembrane charge movement restored in dysgenic skeletal muscle by injection of dihydropyridine receptor cDNAs. Nature. 1990;346:569–572. doi: 10.1038/346569a0. [DOI] [PubMed] [Google Scholar]
- Ahern CA, Bhattacharya D, Mortenson L, Coronado R. A component of excitation-contraction coupling triggered in the absence of the T671-L690 and L720-Q765 regions of the II-III loop of the dihydropyridine receptor α1s pore subunit. Biophys J. 2001;81:3294–3307. doi: 10.1016/S0006-3495(01)75963-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bannister RA. Bridging the myoplasmic gap: recent developments in skeletal muscle excitation–contraction coupling. J Muscle Res Cell M. 2007;28:275–283. doi: 10.1007/s10974-007-9118-5. [DOI] [PubMed] [Google Scholar]
- Bannister RA. Bridging the myoplasmic gap II: more recent advances in skeletal muscle excitation–contraction coupling. J Exp Biol. 2016;219:175–182. doi: 10.1242/jeb.124123. [DOI] [PubMed] [Google Scholar]
- Bannister RA, Beam KG. The α1S N-terminus is not essential for bi-directional coupling with RyR1. Biochem Bioph Res Commun. 2005;336:134–141. doi: 10.1016/j.bbrc.2005.08.047. [DOI] [PubMed] [Google Scholar]
- Bannister Roger A., Grabner Manfred, Beam Kurt G. The α1SIII-IV Loop Influences 1,4-Dihydropyridine Receptor Gating but Is Not Directly Involved in Excitation-Contraction Coupling Interactions with the Type 1 Ryanodine Receptor. Journal of Biological Chemistry. 2008;283(34):23217–23223. doi: 10.1074/jbc.M804312200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bannister RA, Papadopoulos S, Haarmann CS, Beam KG. Effects of inserting fluorescent proteins into the α1S II–III loop: insights into excitation–contraction coupling. J Gen Physiol. 2009;134:35–51. doi: 10.1085/jgp.200910241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beam KG, Bannister RA. Looking for answers to EC coupling’s persistent questions. J Gen Physiol. 2010;136:7–12. doi: 10.1085/jgp.201010461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beurg M, Ahern CA, Vallejo P, Conklin MW, Powers PA, Gregg RG, Coronado R. Involvement of the carboxy-terminus region of the dihydropyridine receptor β1a subunit in excitation-contraction coupling of skeletal muscle. Biophys J. 1999;77:2953–2967. doi: 10.1016/S0006-3495(99)77128-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Block BA, Imagawa T, Campbell KP, Franzini-Armstrong C. Structural evidence for direct interaction between the molecular components of the transverse tubule/sarcoplasmic reticulum junction in skeletal muscle. J Cell Biol. 1988;107:2587–2600. doi: 10.1083/jcb.107.6.2587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bower NI, de la Serrana DG, Cole NJ, Hollway GE, Lee H-T, Assinder S, Johnston IA. Stac3 is required for myotube formation and myogenic differentiation in vertebrate skeletal muscle. J Biol Chem. 2012;287:43936–43949. doi: 10.1074/jbc.M112.361311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buraei Z, Yang J. The β subunit of voltage-gated Ca2+ channels. Physiol Rev. 2010;90:1461–1506. doi: 10.1152/physrev.00057.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calderón JC, Bolaños P, Caputo C. The excitation–contraction coupling mechanism in skeletal muscle. Biophys Rev. 2014;6:133–160. doi: 10.1007/s12551-013-0135-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campiglio Marta, Costé de Bagneaux Pierre, Ortner Nadine J., Tuluc Petronel, Van Petegem Filip, Flucher Bernhard E. STAC proteins associate to the IQ domain of CaV1.2 and inhibit calcium-dependent inactivation. Proceedings of the National Academy of Sciences. 2018;115(6):1376–1381. doi: 10.1073/pnas.1715997115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campiglio M, Flucher BE. STAC3 stably interacts through its C1 domain with CaV1.1 in skeletal muscle triads. Sci Rep. 2017;7:41003. doi: 10.1038/srep41003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campiglio M, Kaplan MM, Flucher BE. STAC3 incorporation into skeletal muscle triads occurs independent of the dihydropyridine receptor. J Cell Physiol. 2018;233:9045–9051. doi: 10.1002/jcp.26767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casarotto MG, Cui Y, Karunasekara Y, Harvey PJ, Norris N, Board PG, Dulhunty AF. Structural and functional characterization of interactions between the dihydropyridine receptor II-III loop and the ryanodine receptor. Clin Exp Pharmacol Physiol. 2006;33:1114–1117. doi: 10.1111/j.1440-1681.2006.04501.x. [DOI] [PubMed] [Google Scholar]
- Catterall WA. Voltage-gated calcium channels. Cold Spring Harb Perspect Biol. 2011;3:a003947. doi: 10.1101/cshperspect.a003947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Y-H, Li M-H, Zhang Y, He L-l, Yamada Y, Fitzmaurice A, Shen Y, Zhang H, Tong L, Yang J. Structural basis of the α1–β subunit interaction of voltage-gated Ca2+ channels. Nature. 2004;429:675–680. doi: 10.1038/nature02641. [DOI] [PubMed] [Google Scholar]
- Cheng W, Altafaj X, Ronjat M, Coronado R. Interaction between the dihydropyridine receptor Ca2+ channel β-subunit and ryanodine receptor type 1 strengthens excitation-contraction coupling. Proc Natl Acad Sci U S A. 2005;102:19225–19230. doi: 10.1073/pnas.0504334102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coronado R, Ahern CA, Sheridan DC, Cheng W, Carbonneau L, Bhattacharya D. Functional equivalence of dihydropyridine receptor a1S and b1a subunits in triggering excitation-contraction coupling in skeletal muscle. Biol Res. 2004;37:565–575. doi: 10.4067/s0716-97602004000400010. [DOI] [PubMed] [Google Scholar]
- Cui Y, Tae H-S, Norris NC, Karunasekara Y, Pouliquin P, Board PG, Dulhunty AF, Casarotto MG. A dihydropyridine receptor α1s loop region critical for skeletal muscle contraction is intrinsically unstructured and binds to a SPRY domain of the type 1 ryanodine receptor. Int J Biochem Cell Biol. 2009;41:677–686. doi: 10.1016/j.biocel.2008.08.004. [DOI] [PubMed] [Google Scholar]
- Dayal A, Bhat V, Franzini-Armstrong C, Grabner M. Domain cooperativity in the β1a subunit is essential for dihydropyridine receptor voltage sensing in skeletal muscle. Proc Natl Acad Sci U S A. 2013;110:7488–7493. doi: 10.1073/pnas.1301087110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dayal A, Schredelseker J, Franzini-Armstrong C, Grabner M. Skeletal muscle excitation–contraction coupling is independent of a conserved heptad repeat motif in the C-terminus of the DHPRβ1a subunit. Cell Calcium. 2010;47:500–506. doi: 10.1016/j.ceca.2010.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dayal A, Schrötter K, Pan Y, Föhr K, Melzer W, Grabner M. The Ca2+ influx through the mammalian skeletal muscle dihydropyridine receptor is irrelevant for muscle performance. Nat Commun. 2017;8:475. doi: 10.1038/s41467-017-00629-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dirksen RT. Bi-directional coupling between dihydropyridine receptors and ryanodine receptors. Front Biosci. 2002;7:d659–d670. doi: 10.2741/A802. [DOI] [PubMed] [Google Scholar]
- Dirksen RT, Beam KG. Role of calcium permeation in dihydropyridine receptor function: insights into channel gating and excitation–contraction coupling. J Gen Physiol. 1999;114:393–404. doi: 10.1085/jgp.114.3.393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dolphin AC. β subunits of voltage-gated calcium channels. J Bioenerg Biomembr. 2003;35:599–620. doi: 10.1023/b:jobb.0000008026.37790.5a. [DOI] [PubMed] [Google Scholar]
- Dulhunty AF, Haarmann CS, Green D, Laver DR, Board PG, Casarotto MG. Interactions between dihydropyridine receptors and ryanodine receptors in striated muscle. Prog Biophys Mol Biol. 2002;79:45–75. doi: 10.1016/s0079-6107(02)00013-5. [DOI] [PubMed] [Google Scholar]
- Efremov RG, Leitner A, Aebersold R, Raunser S. Architecture and conformational switch mechanism of the ryanodine receptor. Nature. 2015;517:39–43. doi: 10.1038/nature13916. [DOI] [PubMed] [Google Scholar]
- El-Hayek R, Antoniu B, Wang J, Hamilton SL, Ikemoto N. Identification of calcium release-triggering and blocking regions of the II-III loop of the skeletal muscle dihydropyridine receptor. J Biol Chem. 1995;270:22116–22118. doi: 10.1074/jbc.270.38.22116. [DOI] [PubMed] [Google Scholar]
- El-Hayek Roque, Ikemoto Noriaki. Identification of the Minimum Essential Region in the II-III Loop of the Dihydropyridine Receptor α1Subunit Required for Activation of Skeletal Muscle-Type Excitation−Contraction Coupling†. Biochemistry. 1998;37(19):7015–7020. doi: 10.1021/bi972907o. [DOI] [PubMed] [Google Scholar]
- Eltit JM, Franzini-Armstrong C, Perez CF. Amino acid residues 489–503 of dihydropyridine receptor (DHPR) β1a subunit are critical for structural communication between the skeletal muscle DHPR complex and type 1 ryanodine receptor. J Biol Chem. 2014;289:36116–36124. doi: 10.1074/jbc.M114.615526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flucher BE, Campiglio M. STAC proteins: the missing link in skeletal muscle EC coupling and new regulators of calcium channel function. BBA-Mol Cell Res. 2018;1866:1101–1110. doi: 10.1016/j.bbamcr.2018.12.004. [DOI] [PubMed] [Google Scholar]
- Flucher BE, Kasielke N, Grabner M. The triad targeting signal of the skeletal muscle calcium channel is localized in the COOH terminus of the α1S subunit. J Cell Biol. 2000;151:467–478. doi: 10.1083/jcb.151.2.467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franzini-Armstrong C, Protasi F. Ryanodine receptors of striated muscles: a complex channel capable of multiple interactions. Physiol Rev. 1997;77:699–729. doi: 10.1152/physrev.1997.77.3.699. [DOI] [PubMed] [Google Scholar]
- Ge X, Zhang Y, Park S, Cong X, Gerrard DE, Jiang H. Stac3 inhibits myoblast differentiation into myotubes. PLoS One. 2014;9:e95926. doi: 10.1371/journal.pone.0095926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golini L, Chouabe C, Berthier C, Cusimano V, Fornaro M, Bonvallet R, Formoso L, Giacomello E, Jacquemond V, Sorrentino V. Junctophilin 1 and 2 proteins interact with the L-type Ca2+ channel dihydropyridine receptors (DHPRs) in skeletal muscle. J Biol Chem. 2011;286:43717–43725. doi: 10.1074/jbc.M111.292755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grabner M, Dirksen RT, Suda N, Beam KG. The II-III loop of the skeletal muscle dihydropyridine receptor is responsible for the bi-directional coupling with the ryanodine receptor. J Biol Chem. 1999;274:21913–21919. doi: 10.1074/jbc.274.31.21913. [DOI] [PubMed] [Google Scholar]
- Gregg RG, Messing A, Strube C, Beurg M, Moss R, Behan M, Sukhareva M, Haynes S, Powell JA, Coronado R, Powers PA. Absence of the β subunit (cchb1) of the skeletal muscle dihydropyridine receptor alters expression of the α1 subunit and eliminates excitation-contraction coupling. Proc Natl Acad Sci U S A. 1996;93:13961–13966. doi: 10.1073/pnas.93.24.13961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hernández-Ochoa EO, Olojo RO, Rebbeck RT, Dulhunty AF, Schneider MF. β1a490–508, a 19-residue peptide from C-terminal tail of Cav1.1 β1a subunit, potentiates voltage-dependent calcium release in adult skeletal muscle fibers. Biophys J. 2014;106:535–547. doi: 10.1016/j.bpj.2013.11.4503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horstick EJ, Linsley JW, Dowling JJ, Hauser MA, McDonald KK, Ashley-Koch A, Saint-Amant L, Satish A, Cui WW, Zhou W, Sprague SM, Stamm DS, Powell CM, Speer MC, Franzini-Armstrong C, Hirata H, Kuwada JY. Stac3 is a component of the excitation–contraction coupling machinery and mutated in Native American myopathy. Nat Commun. 2013;4:1952. doi: 10.1038/ncomms2952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jungbluth H, Treves S, Zorzato F, Sarkozy A, Ochala J, Sewry C, Phadke R, Gautel M, Muntoni F. Congenital myopathies: disorders of excitation–contraction coupling and muscle contraction. Nat Rev Neurol. 2018;14:151–167. doi: 10.1038/nrneurol.2017.191. [DOI] [PubMed] [Google Scholar]
- Karunasekara Y, Dulhunty AF, Casarotto MG. The voltage-gated calcium-channel β subunit: more than just an accessory. Eur Biophys J. 2009;39:75–81. doi: 10.1007/s00249-009-0467-4. [DOI] [PubMed] [Google Scholar]
- Karunasekara Y, Rebbeck RT, Weaver LM, Board PG, Dulhunty AF, Casarotto MG. An α-helical C-terminal tail segment of the skeletal L-type Ca2+ channel β1a subunit activates ryanodine receptor type 1 via a hydrophobic surface. FASEB J. 2012;26:5049–5059. doi: 10.1096/fj.12-211334. [DOI] [PubMed] [Google Scholar]
- Kugler G, Weiss RG, Flucher BE, Grabner M. Structural requirements of the dihydropyridine receptor α1S II-III loop for skeletal-type excitation-contraction coupling. J Biol Chem. 2004;279:4721–4728. doi: 10.1074/jbc.M307538200. [DOI] [PubMed] [Google Scholar]
- Lanner JT, Georgiou DK, Joshi AD, Hamilton SL. Ryanodine receptors: structure, expression, molecular details, and function in calcium release. Cold Spring Harb Perspect Biol. 2010;2:a003996. doi: 10.1101/cshperspect.a003996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Linsley JW, Hsu I-U, Groom L, Yarotskyy V, Lavorato M, Horstick EJ, Linsley D, Wang W, Franzini-Armstrong C, Dirksen RT, Kuwada JY. Congenital myopathy results from misregulation of a muscle Ca2+ channel by mutant Stac3. Proc Natl Acad Sci U S A. 2017;114:E228–E236. doi: 10.1073/pnas.1619238114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayer BJ. SH3 domains: complexity in moderation. J Cell Sci. 2001;114:1253–1263. doi: 10.1242/jcs.114.7.1253. [DOI] [PubMed] [Google Scholar]
- Meissner G, Lu X. Dihydropyridine receptor-ryanodine receptor interactions in skeletal muscle excitation-contraction coupling. Biosci Rep. 1995;15:399–408. doi: 10.1007/BF01788371. [DOI] [PubMed] [Google Scholar]
- Murphy RM, Dutka TL, Horvath D, Bell JR, Delbridge LM, Lamb GD. Ca2+-dependent proteolysis of junctophilin-1 and junctophilin-2 in skeletal and cardiac muscle. J Physiol. 2013;591:719–729. doi: 10.1113/jphysiol.2012.243279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakada T, Kashihara T, Komatsu M, Kojima K, Takeshita T, Yamada M. Physical interaction of junctophilin and the CaV1.1 C terminus is crucial for skeletal muscle contraction. Proc Natl Acad Sci U S A. 2018;115:4507–4512. doi: 10.1073/pnas.1716649115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakai J, Dirksen RT, Nguyen HT, Pessah IN, Beam KG, Allen PD. Enhanced dihydropyridine receptor channel activity in the presence of ryanodine receptor. Nature. 1996;380:72–75. doi: 10.1038/380072a0. [DOI] [PubMed] [Google Scholar]
- Nakai J, Ogura T, Protasi F, Franzini-Armstrong C, Allen PD, Beam KG. Functional nonequality of the cardiac and skeletal ryanodine receptors. Proc Natl Acad Sci U S A. 1997;94:1019–1022. doi: 10.1073/pnas.94.3.1019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakai J, Sekiguchi N, Rando TA, Allen PD, Beam KG. Two regions of the ryanodine receptor involved in coupling with L-type Ca2+ channels. J Biol Chem. 1998;273:13403–13406. doi: 10.1074/jbc.273.22.13403. [DOI] [PubMed] [Google Scholar]
- Nakai J, Tanabe T, Konno T, Adams B, Beam KG. Localization in the II-III loop of the dihydropyridine receptor of a sequence critical for excitation-contraction coupling. J Biol Chem. 1998;273:24983–24986. doi: 10.1074/jbc.273.39.24983. [DOI] [PubMed] [Google Scholar]
- Nelson BR, Wu F, Liu Y, Anderson DM, McAnally J, Lin W, Cannon SC, Bassel-Duby R, Olson EN. Skeletal muscle-specific T-tubule protein STAC3 mediates voltage-induced Ca2+ release and contractility. Proc Natl Acad Sci U S A. 2013;110:11881–11886. doi: 10.1073/pnas.1310571110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neuhuber B, Gerster U, Döring F, Glossmann H, Tanabe T, Flucher BE. Association of calcium channel α1S and β1a subunits is required for the targeting of β1a but not of α1S into skeletal muscle triads. Proc Natl Acad Sci U S A. 1998;95:5015–5020. doi: 10.1073/pnas.95.9.5015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishi M, Mizushima A, K-i N, Takeshima H. Characterization of human junctophilin subtype genes. Biochem Bioph Res Commun. 2000;273:920–927. doi: 10.1006/bbrc.2000.3011. [DOI] [PubMed] [Google Scholar]
- Niu J, Dick IE, Yang W, Bamgboye MA, Yue DT, Tomaselli G, Inoue T, Ben-Johny M. Allosteric regulators selectively prevent Ca2+-feedback of CaV and NaV channels. eLife. 2018;7:e35222. doi: 10.7554/eLife.35222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niu Jacqueline, Yang Wanjun, Yue David T., Inoue Takanari, Ben-Johny Manu. Duplex signaling by CaM and Stac3 enhances CaV1.1 function and provides insights into congenital myopathy. The Journal of General Physiology. 2018;150(8):1145–1161. doi: 10.1085/jgp.201812005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Norris NC, Joseph S, Aditya S, Karunasekara Y, Board PG, Dulhunty AF, Oakley AJ, Casarotto MG. Structural and biophysical analyses of the skeletal dihydropyridine receptor β subunit β1a reveal critical roles of domain interactions for stability. J Biol Chem. 2017;292:8401–8411. doi: 10.1074/jbc.M116.763896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Obermair GJ, Tuluc P, Flucher BE. Auxiliary Ca2+ channel subunits: lessons learned from muscle. Curr Opin Pharmacol. 2008;8:311–318. doi: 10.1016/j.coph.2008.01.008. [DOI] [PubMed] [Google Scholar]
- Opatowsky Y, Chen C-C, Campbell KP, Hirsch JA. Structural analysis of the voltage-dependent calcium channel β subunit functional core and its complex with the α1 interaction domain. Neuron. 2004;42:387–399. doi: 10.1016/s0896-6273(04)00250-8. [DOI] [PubMed] [Google Scholar]
- Opatowsky Y, Chomsky-Hecht O, Kang M-G, Campbell KP, Hirsch JA. The voltage-dependent calcium channel β subunit contains two stable interacting domains. J Biol Chem. 2003;278:52323–52332. doi: 10.1074/jbc.M303564200. [DOI] [PubMed] [Google Scholar]
- Pancaroglu R, Van Petegem F. Calcium channelopathies: structural insights into disorders of the muscle excitation–contraction complex. Annu Rev Genet. 2018;52:373–396. doi: 10.1146/annurev-genet-120417-031311. [DOI] [PubMed] [Google Scholar]
- Paolini C, Fessenden JD, Pessah IN, Franzini-Armstrong C. Evidence for conformational coupling between two calcium channels. Proc Natl Acad Sci U S A. 2004;101:12748–12752. doi: 10.1073/pnas.0404836101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perez CF, Voss A, Pessah IN, Allen PD. RyR1/RyR3 chimeras reveal that multiple domains of RyR1 are involved in skeletal-type E-C coupling. Biophys J. 2003;84:2655–2663. doi: 10.1016/S0006-3495(03)75071-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perni S, Lavorato M, Beam KG. De novo reconstitution reveals the proteins required for skeletal muscle voltage-induced Ca2+ release. Proc Natl Acad Sci U S A. 2017;114:13822–13827. doi: 10.1073/pnas.1716461115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phimister AJ, Lango J, Lee EH, Ernst-Russell MA, Takeshima H, Ma J, Allen PD, Pessah IN. Conformation-dependent stability of junctophilin 1 (JP1) and ryanodine receptor type 1 (RyR1) channel complex is mediated by their hyper-reactive thiols. J Biol Chem. 2007;282:8667–8677. doi: 10.1074/jbc.M609936200. [DOI] [PubMed] [Google Scholar]
- Polster A, Nelson BR, Olson EN, Beam KG. Stac3 has a direct role in skeletal muscle-type excitation–contraction coupling that is disrupted by a myopathy-causing mutation. Proc Natl Acad Sci U S A. 2016;113:10986–10991. doi: 10.1073/pnas.1612441113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polster A, Nelson BR, Papadopoulos S, Olson EN, Beam KG. Stac proteins associate with the critical domain for excitation–contraction coupling in the II–III loop of CaV1.1. J Gen Physiol. 2018;150:613–624. doi: 10.1085/jgp.201711917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polster A, Ohrtman JD, Beam KG, Papadopoulos S. Fluorescence resonance energy transfer (FRET) indicates that association with the type I ryanodine receptor (RyR1) causes reorientation of multiple cytoplasmic domains of the dihydropyridine receptor (DHPR) α1S subunit. J Biol Chem. 2012;287:41560–41568. doi: 10.1074/jbc.M112.404194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polster A, Perni S, Bichraoui H, Beam KG. Stac adaptor proteins regulate trafficking and function of muscle and neuronal L-type Ca2+ channels. Proc Natl Acad Sci U S A. 2015;112:602–606. doi: 10.1073/pnas.1423113112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pragnell M, De Waard M, Mori Y, Tanabe T, Snutch TP, Campbell KP. Calcium channel β-subunit binds to a conserved motif in the I–II cytoplasmic linker of the α1-subunit. Nature. 1994;368:67–70. doi: 10.1038/368067a0. [DOI] [PubMed] [Google Scholar]
- Proenza C, O'Brien J, Nakai J, Mukherjee S, Allen PD, Beam KG. Identification of a region of RyR1 that participates in allosteric coupling with the α1S (CaV1.1) II–III loop. J Biol Chem. 2002;277:6530–6535. doi: 10.1074/jbc.M106471200. [DOI] [PubMed] [Google Scholar]
- Proenza C, Wilkens C, Lorenzon NM, Beam KG. A carboxyl-terminal region important for the expression and targeting of the skeletal muscle dihydropyridine receptor. J Biol Chem. 2000;275:23169–23174. doi: 10.1074/jbc.M003389200. [DOI] [PubMed] [Google Scholar]
- Proenza C, Wilkens CM, Beam KG. Excitation-contraction coupling is not affected by scrambled sequence in residues 681–690 of the dihydropyridine receptor II-III loop. J Biol Chem. 2000;275:29935–29937. doi: 10.1074/jbc.C000464200. [DOI] [PubMed] [Google Scholar]
- Protasi F. Structural interaction between RYRs and DHPRs in calcium release units of cardiac and skeletal muscle cells. Front Biosci. 2002;7:d650–d658. doi: 10.2741/A801. [DOI] [PubMed] [Google Scholar]
- Protasi F, Paolini C, Nakai J, Beam KG, Franzini-Armstrong C, Allen PD. Multiple regions of RyR1 mediate functional and structural interactions with α1S-dihydropyridine receptors in skeletal muscle. Biophys J. 2002;83:3230–3244. doi: 10.1016/S0006-3495(02)75325-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rebbeck RT, Karunasekara Y, Board PG, Beard NA, Casarotto MG, Dulhunty AF. Skeletal muscle excitation–contraction coupling: who are the dancing partners? Int J Biochem Cell Biol. 2014;48:28–38. doi: 10.1016/j.biocel.2013.12.001. [DOI] [PubMed] [Google Scholar]
- Rebbeck RT, Karunasekara Y, Gallant EM, Board PG, Beard NA, Casarotto MG, Dulhunty AF. The β1a subunit of the skeletal DHPR binds to skeletal RyR1 and activates the channel via its 35-residue C-terminal tail. Biophys J. 2011;100:922–930. doi: 10.1016/j.bpj.2011.01.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reinholt BM, Ge X, Cong X, Gerrard DE, Jiang H. Stac3 is a novel regulator of skeletal muscle development in mice. PLoS One. 2013;8:e62760. doi: 10.1371/journal.pone.0062760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richards MW, Butcher AJ, Dolphin AC. Ca2+ channel β-subunits: structural insights AID our understanding. Trends Pharmacol Sci. 2004;25:626–632. doi: 10.1016/j.tips.2004.10.008. [DOI] [PubMed] [Google Scholar]
- Rios E, Brum G. Involvement of dihydropyridine receptors in excitation–contraction coupling in skeletal muscle. Nature. 1987;325:717–720. doi: 10.1038/325717a0. [DOI] [PubMed] [Google Scholar]
- Sandow A. Excitation-contraction coupling in muscular response. Yale J Biol Med. 1952;25:176–201. [PMC free article] [PubMed] [Google Scholar]
- Schredelseker J, Dayal A, Schwerte T, Franzini-Armstrong C, Grabner M. Proper restoration of excitation-contraction coupling in the dihydropyridine receptor β1-null zebrafish relaxed is an exclusive function of the β1a subunit. J Biol Chem. 2009;284:1242–1251. doi: 10.1074/jbc.M807767200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schredelseker J, Di Biase V, Obermair GJ, Felder ET, Flucher BE, Franzini-Armstrong C, Grabner M. The β1a subunit is essential for the assembly of dihydropyridine-receptor arrays in skeletal muscle. Proc Natl Acad Sci U S A. 2005;102:17219–17224. doi: 10.1073/pnas.0508710102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheridan DC, Cheng W, Carbonneau L, Ahern CA, Coronado R. Involvement of a heptad repeat in the carboxyl terminus of the dihydropyridine receptor β1a subunit in the mechanism of excitation-contraction coupling in skeletal muscle. Biophys J. 2004;87:929–942. doi: 10.1529/biophysj.104.043810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheridan DC, Takekura H, Franzini-Armstrong C, Beam KG, Allen PD, Perez CF. Bidirectional signaling between calcium channels of skeletal muscle requires multiple direct and indirect interactions. Proc Natl Acad Sci U S A. 2006;103:19760–19765. doi: 10.1073/pnas.0609473103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strube C, Beurg M, Powers PA, Gregg RG, Coronado R. Reduced Ca2+ current, charge movement, and absence of Ca2+ transients in skeletal muscle deficient in dihydropyridine receptor β1 subunit. Biophys J. 1996;71:2531–2543. doi: 10.1016/S0006-3495(96)79446-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tae H-S, Norris NC, Cui Y, Karunasekara Y, Board PG, Dulhunty AF, Casarotto MG. Molecular recognition of the disordered dihydropyridine receptor II-III loop by a conserved SPRY domain of the type 1 ryanodine receptor. Clin Exp Pharmacol Physiol. 2009;36:346–349. doi: 10.1111/j.1440-1681.2008.05130.x. [DOI] [PubMed] [Google Scholar]
- Takeshima H, Iino M, Takekura H, Nishi M, Kuno J, Minowa O, Takano H, Noda T. Excitation-contraction uncoupling and muscular degeneration in mice lacking functional skeletal muscle ryanodine-receptor gene. Nature. 1994;369:556–559. doi: 10.1038/369556a0. [DOI] [PubMed] [Google Scholar]
- Takeshima H, Komazaki S, Nishi M, Iino M, Kangawa K. Junctophilins: a novel family of junctional membrane complex proteins. Mol Cell. 2000;6:11–22. doi: 10.1016/s1097-2765(00)00003-4. [DOI] [PubMed] [Google Scholar]
- Tanabe T, Beam KG, Adams BA, Niidome T, Numa S. Regions of the skeletal muscle dihydropyridine receptor critical for excitation–contraction coupling. Nature. 1990;346:567–569. doi: 10.1038/346567a0. [DOI] [PubMed] [Google Scholar]
- Tanabe Tsutomu, Beam Kurt G., Powell Jeanne A., Numa Shosaku. Restoration of excitation—contraction coupling and slow calcium current in dysgenic muscle by dihydropyridine receptor complementary DNA. Nature. 1988;336(6195):134–139. doi: 10.1038/336134a0. [DOI] [PubMed] [Google Scholar]
- Van Petegem F, Clark KA, Chatelain FC, Minor DL., Jr Structure of a complex between a voltage-gated calcium channel β-subunit and an α-subunit domain. Nature. 2004;429:671–675. doi: 10.1038/nature02588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weiss RG, O’Connell KMS, Flucher BE, Allen PD, Grabner M, Dirksen RT. Functional analysis of the R1086H malignant hyperthermia mutation in the DHPR reveals an unexpected influence of the III-IV loop on skeletal muscle EC coupling. Am J Physiol: Cell Physiol. 2004;287:C1094–C1102. doi: 10.1152/ajpcell.00173.2004. [DOI] [PubMed] [Google Scholar]
- Wilkens CM, Kasielke N, Flucher BE, Beam KG, Grabner M. Excitation–contraction coupling is unaffected by drastic alteration of the sequence surrounding residues L720–L764 of the α1S II-III loop. Proc Natl Acad Sci U S A. 2001;98:5892–5897. doi: 10.1073/pnas.101618098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu J, Yan Z, Li Z, Qian X, Lu S, Dong M, Zhou Q, Yan N. Structure of the voltage-gated calcium channel CaV1.1 at 3.6 Å resolution. Nature. 2016;537:191–196. doi: 10.1038/nature19321. [DOI] [PubMed] [Google Scholar]
- Wu J, Yan Z, Li Z, Yan C, Lu S, Dong M, Yan N. Structure of the voltage-gated calcium channel CaV1.1 complex. Science. 2015;350:aad2395. doi: 10.1126/science.aad2395. [DOI] [PubMed] [Google Scholar]
- Yamazawa T, Takeshima H, Shimuta M, Iino M. A region of the ryanodine receptor critical for excitation-contraction coupling in skeletal muscle. J Biol Chem. 1997;272:8161–8164. doi: 10.1074/jbc.272.13.8161. [DOI] [PubMed] [Google Scholar]
- Yan Z, X-c B, Yan C, Wu J, Li Z, Xie T, Peng W, C-c Y, Li X, Scheres SH, Shi Y, Yan N. Structure of the rabbit ryanodine receptor RyR1 at near-atomic resolution. Nature. 2015;517:50–55. doi: 10.1038/nature14063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuen SMWK, Campiglio M, Tung C-C, Flucher BE, Van Petegem F. Structural insights into binding of STAC proteins to voltage-gated calcium channels. Proc Natl Acad Sci U S A. 2017;114:E9520–E9528. doi: 10.1073/pnas.1708852114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaharieva IT, et al. STAC3 variants cause a congenital myopathy with distinctive dysmorphic features and malignant hyperthermia susceptibility. Hum Mutat. 2018;39:1980–1994. doi: 10.1002/humu.23635. [DOI] [PubMed] [Google Scholar]
- Zalk R, Clarke OB, des Georges A, Grassucci RA, Reiken S, Mancia F, Hendrickson WA, Frank J, Marks AR. Structure of a mammalian ryanodine receptor. Nature. 2015;517:44–49. doi: 10.1038/nature13950. [DOI] [PMC free article] [PubMed] [Google Scholar]