

Abstract
Similarities between parent and offspring are widespread in psychology; however, shared genetic variants often confound causal inference for offspring outcomes. A polygenic score (PGS) derived from genome-wide association studies (GWAS) can be used to test for the presence of parental influence that controls for genetic variants shared across generations. We use a PGS for educational attainment (EA3; N ≈ 750 thousand) to predict offspring years of education in a sample of 2517 twins and both parents. We find that within families, the dizygotic twin with the higher PGS is more likely to attain higher education (unstandardized β = 0.32; p < 0.001). Additionally, however, we find an effect of parental genotype on offspring outcome that is independent of the offspring’s own genotype; this raises the variance explained in offspring years of education from 9.3 to 11.1% (ΔR2 = 0.018, p < 0.001). Controlling for parental IQ or socioeconomic status substantially attenuated or eliminated this effect of parental genotype. These findings suggest a role of environmental factors affected by heritable characteristics of the parents in fostering offspring years of education.
Introduction
Children typically develop traits and behaviors that are influenced by both the genes and the family environment they have inherited from their parents. Decades of twin and adoption studies have dramatically shifted our understanding of the factors that drive parent–offspring associations, including the discovery that the shared family environment provided by parents contributes less to the development of most behavioral traits than do the shared genetic factors [1, 2]. Despite the elusive nature of shared environment effects, pinpointing their sources remains a tantalizing challenge for many researchers. The ability to do so reliably has been bolstered by advances in quantitative genetics, which have made it possible to detect and measure genetic influences on complex behaviors at the molecular level. In particular, genome-wide association studies (GWAS) enable us to aggregate the tiny effects of many genetic variants into a polygenic score, which functions as an index of genetic propensity towards a phenotype of interest [3]. Here, we use polygenic scores to test the hypothesis that both parent and offspring genes contribute to offspring behaviors in a manner consistent with environmental mechanisms of intergenerational transmission.
Relevant to this transmission is the observation that genetic propensities are often associated with environmental experiences, a phenomenon known collectively as gene–environment correlation (rGE). Children usually grow up with relatives whose own genes help to create an environment that influences the development of the children’s traits or behaviors. For example, musically gifted children may inherit both genes and an environment conducive to developing musical ability—such as instruments and lessons—from their parents [4]. Evidence that parents drive such “passive” gene–environment correlation comes from studies of adopted families and of children of twins, which both control for genetic influence in different ways. Since adopted children do not share genes with their parents, a correlation between parent and offspring traits is likely mediated by the environment that the parents create.This approach has uncovered an effect of home environment on cognitive development [5] and educational attainment [6] of children in adopted families. The children-of-twins design compares the offspring of adult identical twins as a way to clarify whether measures of family environment directly influence child outcomes or are genetically mediated [7]. This method has produced evidence that the relationship between parental divorce and offspring drug use is genetically mediated, while divorce plays a direct environmental role in the development of offspring emotional problems [8].
While these are valuable methods, their utility is limited by a number of factors, including practical difficulties in assembling large samples of families with the relevant structure (e.g., adoption). GWAS, by contrast, provide a toolkit for direct quantification of genetic transmission from parents to offspring at the individual level. This is enabled by the massively polygenic architecture of many complex outcomes, and that individual scores based on aggregate single-nucleotide polymorphisms (SNPs) associated with these outcomes have substantial predictive value. One of the outcomes that has been studied most successfully in these ways is educational attainment (measured in total years of education, or “EduYears”). The most recent GWAS of educational attainment, which included 1.1 million individuals, identified 1271 significant SNPs [9]. A polygenic score (PGS) constructed from this large sample is now able to predict 11–13% of the variance in educational attainment and 7–10% of the variance in cognitive ability, exceeding all previous benchmarks [10, 11], and providing further evidence that educational attainment is a viable proxy for cognitive ability.
The PGS constructed from these studies are reliable enough to draw inferences about sources of environmental variation that are often inconclusive in twin and adoption studies. As of early 2018, these scores have been used to predict outcomes in offspring consistent with a causal role of the environment fostered by the parents. Kong et al. [12], the first to demonstrate “genetic nurture” with a PGS, found that almost 30% of the offspring PGS’s correlation with their educational attainment is due to the environment provided by their parents, as inferred from a significant effect of the non-transmitted portion of the parents’ genomes. Bates et al. [13] subsequently produced a similar finding in a sample of genotyped twins and their parents using a virtual-parent design, which additionally found that a family’s inherited socioeconomic status (SES) was significantly related to both parents’ and offspring’s polygenic scores. These studies were followed by Belsky et al. [14], which was the first to provide evidence for such environmental transmission using a PGS derived from EA3 data in a sample of siblings and their parents. Liu [15] replicated the genetic nurture effect of parents’ genome on offspring outcome and additionally examined this effect with grandparents, finding no evidence that the non-biological transmission of educational attainment persists beyond one generation.
These studies have shed valuable light on questions surrounding the non-genetic pathways of intergenerational transmission of outcomes, and in doing so have raised others of import and interest. For example, it remains largely unexplored whether and to what extent other social outcomes and parent/offspring phenotypes—for example, IQ and “soft” skills—follow a similar pattern of non-genetic transmission. Additionally, the family characteristics and processes that could mediate the effects of genetic nurture remain mostly unexplored.
We replicate the “genetic nurture” effect in a relatively rare kind of dataset consisting of both monozygotic and dizygotic twins and (typically) both parents, all of whom are genotyped. In doing so, we seek to address the outstanding questions about genetic nurture with the following logic. In the simplest quantitative-genetic model, the portion of the parent genotype that leads causally to the measured trait—their “true” genetic value—affects the educational attainment of the children through the true genetic value of the offspring, which is inherited fully from the parents. This model predicts that offspring polygenic scores would completely mediate the effect of the parent polygenic scores. However, a unique contribution of parental PGS to offspring outcomes would suggest that parental genotypes affect something about the environment that influences the offspring’s outcome; this causal path would therefore act apart from and in addition to the parents’ contribution to the offspring’s genotype. Other parental phenotypes, such as socioeconomic status, can be used as covariates to test for a significant reduction in the partial regression coefficient of parental PGS. This finding would be consistent with a causal role of parental PGS on these covariates, which may be good measures of the environmental variables that could in turn affect offspring outcomes (Fig. 1).
Fig. 1.
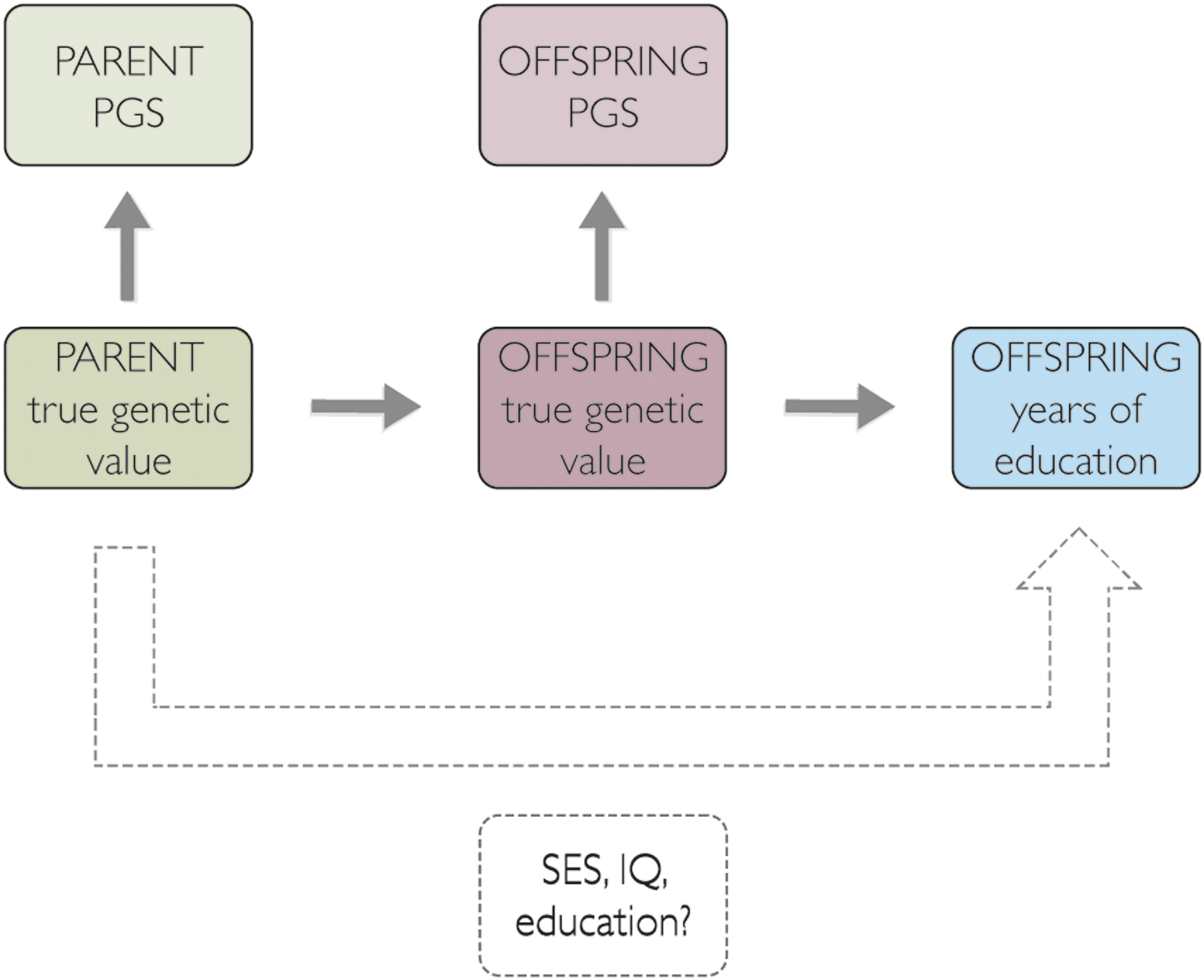
Path diagram representing hypothesized causal and mediating pathways. The true genetic value of the parents, which is imperfectly captured in a polygenic score, causally influences the offspring’s genotype. The parents’ genes may influence the offspring trait through parental characteristics such as socioeconomic status, IQ, and education
To conduct a direct test of this model, we use EA3-derived polygenic scores from pairs of monozygotic and dizygotic twins and both of their parents, enabling the investigation of two fundamental questions. First, do polygenic scores predict educational attainment and related phenotypes within families, such that the dizygotic twin with the higher polygenic score will also tend to have attained the highest education? If genetic nurture is operating, then a population GWAS will overestimate a given SNP’s “true” regression coefficient—that is, the regression coefficient induced by its average effect of gene substitution [16, 17] and linkage disequilibrium with other causal sites. The reason for the overestimation is that an individual’s SNP genotype is confounded by the genotypes of the parents, which affect some parental phenotype that in turn affects the trait being studied in the GWAS. In a within-family setting, however, two siblings with different PGS values are expected to differ phenotypically by an amount given solely by the true effects; this is because the confounding factor of parent genotype is fixed to be the same for all siblings begotten by a given pair of parents. Thus, if the PGS is derived from a GWAS of a trait subject to genetic nurture, the predictive power of the PGS should decline when it is applied within families.
Second, does accounting for parental PGS improve the prediction of offspring years of education over and above how well offspring PGS predicts this outcome, and do environmental factors correlated with parental PGS explain this increment in prediction?
We address these research questions in an investigation of genetic nurture for years of education using moleculargenetic techniques on large samples.
Methods
Sample
The current project is based on the results of a genome-wide association study conducted by the Minnesota Center for Twin and Family Research (MCTFR). The sample under investigation represents 4478 genotyped individuals of European ancestry with polygenic scores (PGS) for years of education [18]. Comprising this sample are sets of parents and offspring from a total of 1223 families, including 2032 parents (1093 mothers and 939 fathers) and 2446 twins (830 dizygotic, 55% female; 1616 monozygotic, 53% female). Parent predictors and outcomes are calculated as the mean of mother and father on a given variable score; e.g., EduYears PGS.
Measures
Participants were genotyped on 527,829 SNPs using the Illumina Human660W-Quad array (see ref. [18] for additional details on sampling, assessment, quality control, and imputation performed on the MCTFR sample). A polygenic score (PGS), often called the polygenic risk score in disease prediction, is calculated from a set of SNPs that are tested in the initial sample for association with a trait of interest (see Supplementary Online Materials Section 1) [19]. Parent and offspring polygenic scores for years of education are used to predict outcomes using multivariate regressions, as was done by Belsky et al. and Liu. A finding of the parent PGS having a significant partial regression coefficient is equivalent in interpretation to the finding of non-transmitted alleles having a significant partial regression coefficient in the method of Kong et al. and Bates et al. (Supplementary Online Materials Section 7). Note that such a finding cannot be the result of ameliorating the noisiness of the offspring PGS, as the weights in both the parent and offspring PGS are derived from the same GWAS with the same sampling errors (Supplementary Online Materials Section 4 and accompanying Supplementary Fig. S1).
Polygenic scores for two non-behavioral phenotypes, derived using the same prediction methods as for EduYears, are used as negative controls to test the assumption that the hypothesized effect of genetic nurture on years of education is unique to its PGS, and is not simply the result of attenuating the noise in the offspring PGS due to GWAS sampling error. Therefore, if the addition of parent PGS for EduYears is found to add significant incremental variance to offspring years of education, we would predict that a non-behavioral parent PGS would add a nonsignificant increment in predicting their accompanying offspring phenotypes. We used a physical PGS for body-mass index (BMI; body mass divided by the square of body height, kg/m2) and height for all participants in our sample. These scores are derived from UK Biobank summary statistics [20].
Outcome variables are represented by a variety of behavioral and physical phenotype data for both parents and offspring. Behavioral phenotypes include educational attainment, assessed as years of education for offspring (ranging from 11 to 20) and level of education for parents (coded on a 1–5 scale with 1 = less than high-school up to 5 = professional degree), self-reported high-school grades reported on a 0 to 4 grade point average (GPA) scale, IQ scores for both parents and offspring, family socioeconomic status (SES; represented by a composite made up of family income, parent education level, and parent occupation level on a z-score scale), and soft skills (represented by a composite of personality measures of conscientiousness, capacity to be hard-working, and self-control on a z-score scale). This soft skill measure is included to explore to what extent a polygenic score for years of education significantly predicts a non-cognitive behavioral phenotype that is plausibly related to educational success. Socioeconomic status z-scores were available for 1221 families. Offspring were assessed for some phenotypes at different ages. Years of education were recorded at an age typical for having completed education (M age = 29, SD = 1.4). High-school GPA was assessed at age 17 for all offspring. IQ scores were assessed using an abbreviated form of either the Weschler Adult Intelligence Scale-Revised (WAIS-R [21]) for participants age 16 years and older (47.1% of sample), or the Weschler Intelligence Scale for Children-Revised (WISC-R [22]) for those younger than 16(52.9% of sample), for an overall mean IQ assessment age of 14.4 (SD = 2.9) for offspring in the sample. BMI and height were measured directly for both parents and offspring in the MCTFR sample, and offspring were assessed at age 17.
Phenotype data and polygenic scores are standardized for all analyses. The Supplementary Online Materials (Section 2) detail our exclusion criteria, standardization, and bootstrap resampling.
Results
Polygenic scores as predictors of individual outcome
The pattern of correlations found among IQ, years of education, and family socioeconomic status for parents and offspring (Table 1) is consistent with existing literature on the relationships between these and related variables [23–25], though the causality underlying these relationships is still debated [26, 27].
Table 1.
Descriptive statistics and correlations among polygenic scores and all analyzed phenotypes
| Variable | M | SD | Range | Correlations | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | ||||
| 1. Socioeconomic status (SES) | 0.00 | 1.00 | −2.48–2.26 | 0.37 [0.33, 0.41] | 0.55 [0.52, 0.58] | 0.83 [0.82, 0.84] | 0.24 [0.20, 0.28] | 0.30 [0.27, 0.34] | 0.44 [0.41, 0.47] | 0.27 [0.23, 0.31] | 0.20 [0.17, 0.24] |
| 2. Parental EduYears PGS | 0.02 | 0.75 | −2.48–2.41 | — | 0.34 [0.30, 0.38] | 0.39 [0.35, 0.42] | 0.73 [0.71, 0.75] | 0.24 [0.20, 0.29] | 0.31 [0.27, 0.35] | 0.23 [0.19, 0.28] | 0.15 [0.11, 0.20] |
| 3. Parental IQ score | 104.4 | 11.69 | 73.0–144.5 | — | 0.60 [0.57, 0.62] | 0.24 [0.20, 0.28] | 0.48 [0.45, 0.51] | 0.29 [0.26, 0.33] | 0.23 [0.19, 0.27] | 0.08 [0.04, 0.13] | |
| 4. Parental educationa | 2.65 | 0.91 | 1.00–5.00 | — | 0.25 [0.21, 0.29] | 0.33 [0.30, 0.37] | 0.42 [0.39, 0.45] | 0.27 [0.24, 0.31] | 0.17 [0.13, 0.21] | ||
| 5. Offspring EduYears PGS | −0.05 | 0.99 | −3.46–3.35 | — | 0.29 [0.25, 0.32] | 0.31 [0.27, 0.34] | 0.26 [0.22, 0.30] | 0.16 [0.12, 0.20] | |||
| 6. Offspring IQ score | 102 | 14.19 | 50.0–156.0 | — | 0.36 [0.32, 0.39] | 0.35 [0.31, 0.39] | 0.17 [0.13, 0.21] | ||||
| 7. Offspring educationb | 14.68 | 1.88 | 11.0-–20.0 | — | 0.53 [0.50, 0.56] | 0.43 [0.39, 0.46] | |||||
| 8. Offspring GPA | 2.98 | 0.88 | 0.00–4.00 | — | 0.58 [0.55, 0.60] | ||||||
| 9. Offspring soft skills | 0.00 | 1.00 | −3.38–2.53 | — |
All correlations are significant at the p < 0.001 level, and 95% confidence intervals are given in brackets. All parental values represent mean of mother and father. Polygenic scores, SES, and soft skills are standardized for all prediction models. Soft skills score is represented by a z-score composite of conscientiousness, capacity to be hard-working, and self-control scores
Parental education is coded as “level of education”, with 1 = less than high school and 5 = professional degree (see Methods: Phenotypic measures)
Offspring education is coded as “total years of education”
Individual-level predictions of EduYears polygenic scores on the four outcome phenotypes are robust and significant at the p < 0.001 level. Variance explained by polygenic scores in years of education (R2 = 0.093), IQ (R2 = 0.082) and GPA (R2 = 0.071) is similar to predictions made for cognitive phenotypes in validation samples studied by Lee et al. Although the effect of PGS on soft skills (R2 = 0.028) is modest, it nevertheless demonstrates that EduYears PGS is predictive for non-cognitive behavioral phenotypes, a phenomenon noted in an earlier study [28]. An intercept for years of education of 14.79 indicates that the average PGS in the sample is associated with 14.79 years of education (equivalent to 2.79 years of college), and a slope of 0.51 (SE = 0.06) represents a gain of about one-half year of education for each standard deviation increase in PGS.
Predictions within twinships
EduYears PGS is a reliable predictor for individual outcomes, but within-family predictions can account for possible effects of the shared environment by examining whether sibling genetic differences are associated with differences in sibling outcomes. Dizygotic twins in the present sample (total pairs = 415) are the same age, the same sex, and grew up in the same family environment, but only share an average of half of their genetic material identically by descent, and therefore have different polygenic scores for educational attainment. This makes these pairs useful for determining whether the random difference in variants associated with educational attainment inherited from the parents is a significant predictor of difference in outcome, thereby controlling for population stratification and passive gene–environment correlation effects [29]. Within-twinship predictions are made for EduYears polygenic score on outcome phenotypes years of education, IQ, GPA, and soft skills by using the difference score of each pair of dizygotic twins’ PGS as the predictor for the difference score of each of the four outcome variables. For independence of comparisons, the results of these regressions are compared against individual predictions for monozygotic twins only (total pairs in sample = 808).
In Table 2, the intercept and slope for each outcome variable, regressed on EduYears PGS, is shown alongside the standard error (SE) for both individual (MZ twins only) and within-twinship (DZ twins only) predictions. The PGS of individual (MZ) twins is a similarly robust predictor of each phenotype as in the full sample of all twins. Interestingly, the within-family coefficient for high-school GPA tends to be larger than the individual-level coefficient. This might be because DZ twins will mostly attend the same school, and GPA comparisons between students who attend the same school are more meaningful than comparisons between students who attend different schools. Only the within-twinship prediction for soft skills fails to reach sta tistical significance. This indicates that within the same dizygotic twinship, the twin with the higher EduYears PGS is significantly more likely to have a higher GPA, more years of education, and a higher IQ. These findings are highlighted in Fig. 2, which compares the size and significance of the standardized regression coefficients for an individual’s PGS in predicting years of education, GPA, IQ, and soft skills to the within-twinship coefficients for the same predictor and outcomes. These findings are consistent with similar within-twinship comparisons conducted in an earlier study [14]. The Supplementary Online Materials (Section 6) provide further discussion of these within-family comparisons and their interpretations (Supplementary Fig. S2).
Table 2.
Individual and within-twinship regression coefficients (unstandardized) for EduYears PGS on four behavioral outcome phenotypes
| Phenotype | Intercept (SE) | β-estimate (SE) |
|---|---|---|
| Individual predictions | ||
| Years of education | 14.79 (0.06) | 0.51 (0.06) |
| IQ | 102.73 (0.43) | 4.16 (0.47) |
| GPA | 3.04 (0.03) | 0.21 (0.04) |
| Soft skills | 0.04 (0.04) | 0.15 (0.03) |
| Within-twinship predictions | ||
| Years of eduction | 0.32 (0.11) | |
| IQ | 2.24 (0.72) | |
| GPA | 0.24 (0.05) | |
| Soft skills | 0.10 (0.07) | |
Individual predictions are based on the subset of the sample comprising only monozygotic twins (N pairs = 808), and within-twinship predictions on dizygotic twins (N pairs = 415), to ensure independence of observations. Within-twinship predictions are based on difference scores and have intercepts forced to zero
Fig. 2.
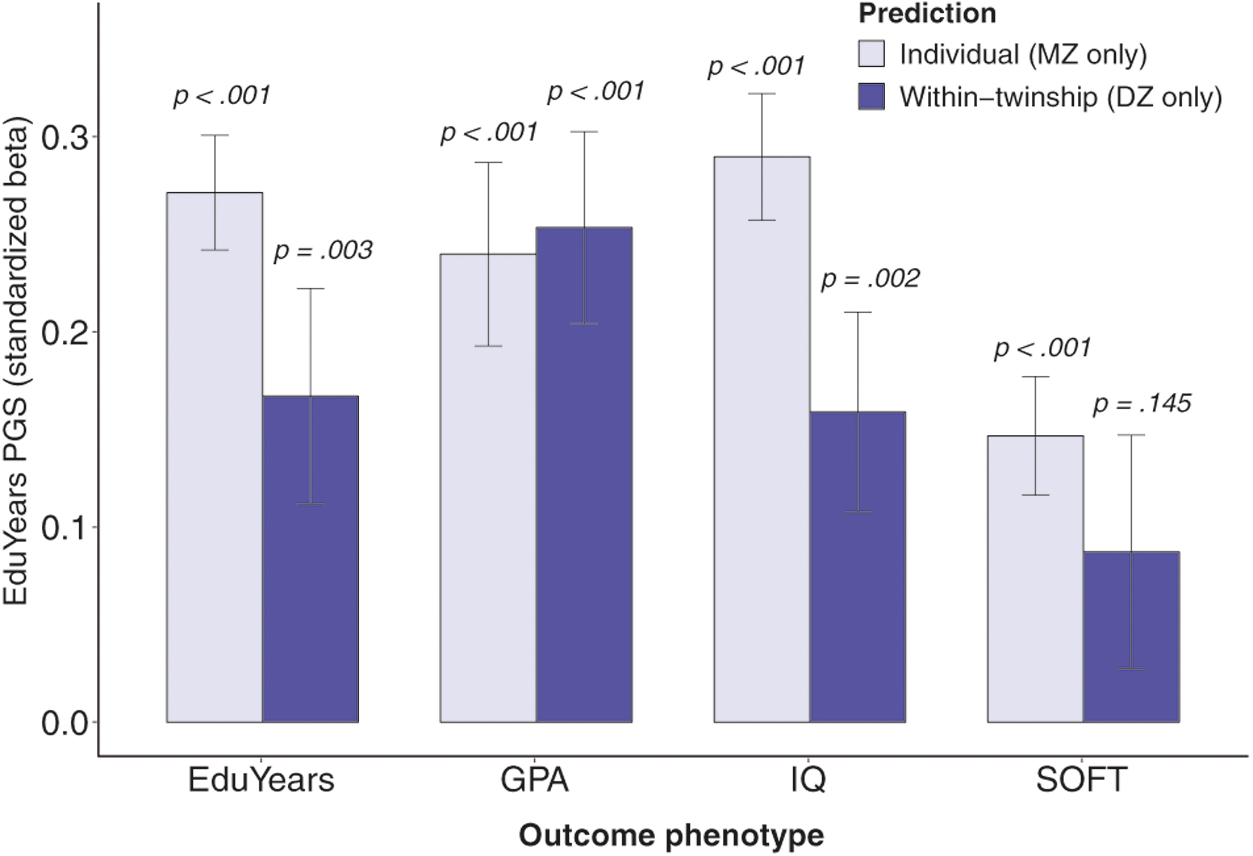
Comparison of β-coefficients of offspring polygenic score (PGS) on outcomes between individual monozygotic twins (N pairs = 808) and within dyzgotic twinship (N pairs = 415) for years of education, high-school GPA, IQ score, and soft skills. Error bars represent ± 1 standard error. All values are standardized
Parent polygenic scores and offspring outcomes
To test for direct effects of parental influence on offspring outcomes, we include parental EduYears PGS (mean of mother and father) along with offspring EduYears PGS as predictors of offspring outcomes. We found that parental EduYears PGS adds significant incremental R2 to offspring EduYears PGS in predicting actual years of education attained by the offspring, raising total variance explained in outcome from 9.3 to 11.1% (ΔR2 = 0.018, p < 0.001; Table 3). While the offspring’s β-coefficient of 0.566 (SE =0.06) on its own translates to ~6.8 months of education gained for every standard deviation gained in the offspring’s EduYears PGS, the parents’ β-estimate of 0.477 (SE =0.099) in the full model predicts a gain of nearly 6 months in educational attainment of the offspring for every standard deviation of parents’ EduYears PGS. IQ, GPA, and soft skills show a more modest association with parent PGS (all ΔR2 < 0.009). These results indicate that parents’ genetic value for educational attainment uniquely predicts their children’s educational achievement, in a way that is not explained by children’s own genetic value, consistent with the hypothesized operation of genetic nurture. Our estimate of genetic nurture affecting EduYears is larger than that of other studies (Supplementary Table S2), but not by enough to suggest substantial heterogeneity, as all studies are consistent with a ratio of direct causal effect to nurture-confounded coefficient exceeding 0.6. Note that such a ratio is also consistent with the ratio of our within-family and individual-level coefficients (Fig. 2) and the estimate of the EA3 authors, who used a more indirect method.
Table 3.
Comparison of variance explained in six phenotypes by offspring polygenic score to variance explained by offspring PGS in addition to parent PGS
| Offspring outcomes | Model | ||
|---|---|---|---|
| Offspring PGS alone | Offspring + parent PGS | ||
| Total R2 | Parental ΔR2 (p-value) | Total R2 | |
| Behavioral phenotypes | |||
| Years of education | 0.093 | 0.018 (<0.001) | 0.111 |
| IQ | 0.082 | 0.004 (0.041) | 0.085 |
| GPA | 0.071 | 0.008 (0.004) | 0.078 |
| Soft skillsa | 0.028 | 0.005 (0.012) | 0.033 |
| Physical phenotypes | |||
| Heighta | 0.334 | <0.001 (0.665) | 0.334 |
| BMFa | 0.134 | <0.001 (0.551) | 0.134 |
Behavioral phenotypes are predicted for offspring and parent using EduYears PGS. Physical phenotypes are predicted for offspring and parent using height and BMI PGS, respectively. Parent values represent the mean of mother and father. Total R2 for both offspring-only model and offspring + midparent model are significant at the p < 0.001 level
Indicates standardized phenotypes (z-scores)
Polygenic scores and their accompanying phenotypes for two non-behavioral traits, BMI and height, are used as negative controls. While it is in-tuitively plausible that parents’ genetic value associated with a complex behavioral phenotype could change their children’s environment in important ways, it would be difficult to explain an effect of similar size on a physical property such as height. Offspring polygenic scores for height explain ~33.4% of the variance in actual height measured at age 17 in this sample of twins, and polygenic scores for BMI explain ~13.4% of the variance in BMI. In contrast to the 1.8% added by parental EduYears PGS, the parental PGS for both height and BMI add essentially 0.0% of incremental variance to offspring outcomes (coefficient p = 0.665 and coefficient p = 0.511 for height and BMI, respectively). A comparison of offspring and offspring + parent PGS models for height and BMI is shown in Fig. 3. This pattern of results, where the increment added to offspring outcome by parent PGS is much larger and less likely to be due to chance for EduYears than it is for BMI or height, provides further evidence that the parents’ genes associated with educational attainment are influencing aspects of the offspring’s environment that lead to differences in actual educational attainment.
Fig. 3.
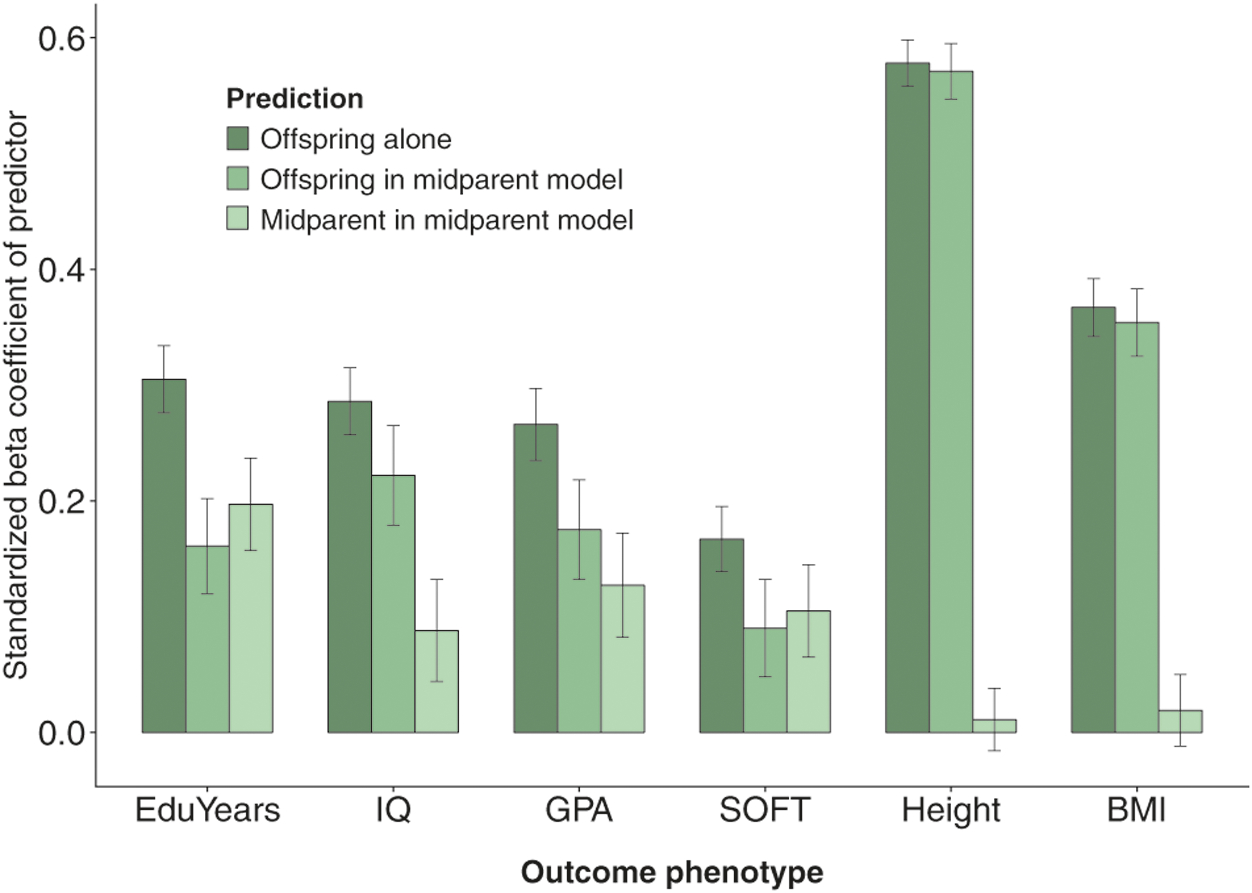
Bar plot showing the standardized beta coefficients of offspring polygenic score (PGS) alone on outcome phenotypes, offspring PGS when midparent PGS is added to the regression model, and midparent PGS in this multivariate model. EduYears PGS is predictor for EduYears, IQ, GPA, and SOFT phenotypes; height PGS and body-mass index (BMI) PGS are predictors for height and BMI phenotypes, respectively. Error bars represent ± 1 standard error
Socioeconomic status and other parental phenotypes as covariates
If parents’ genes are changing something about their offspring’s environment, it may be possible to detect this by adding covariates to the model and determining whether the coefficient of the parent PGS declines, as expected if the covariates are on the causal path from the parent PGS to offspring EduYears [29, 30]. Family SES and parent IQ are two possible mediators. When each of these variables is added to the model where offspring PGS and parent PGS are both significant predictors of offspring years of education, the β-coefficient declines and the p-value increases for the parent PGS coefficient, but not for the offspring’s own PGS. The effect is particularly dramatic for family socioeconomic status, which reduces both the coefficient and the ΔR2 of parent PGS to close to zero on its own. We again used the physical phenotypes of height and BMI as negative controls. Figure 4 shows that for both height and BMI, the addition of the parental characteristics as covariates does not appreciably change the effect size and significance of parental PGS as a predictor, consistent with the use of these as negative controls detailed in Fig. 3. Supplementary Table S1 details the persistence of parental PGS as a significant predictor of offspring outcomes across a number of different parental characteristics as covariates.
Fig. 4.
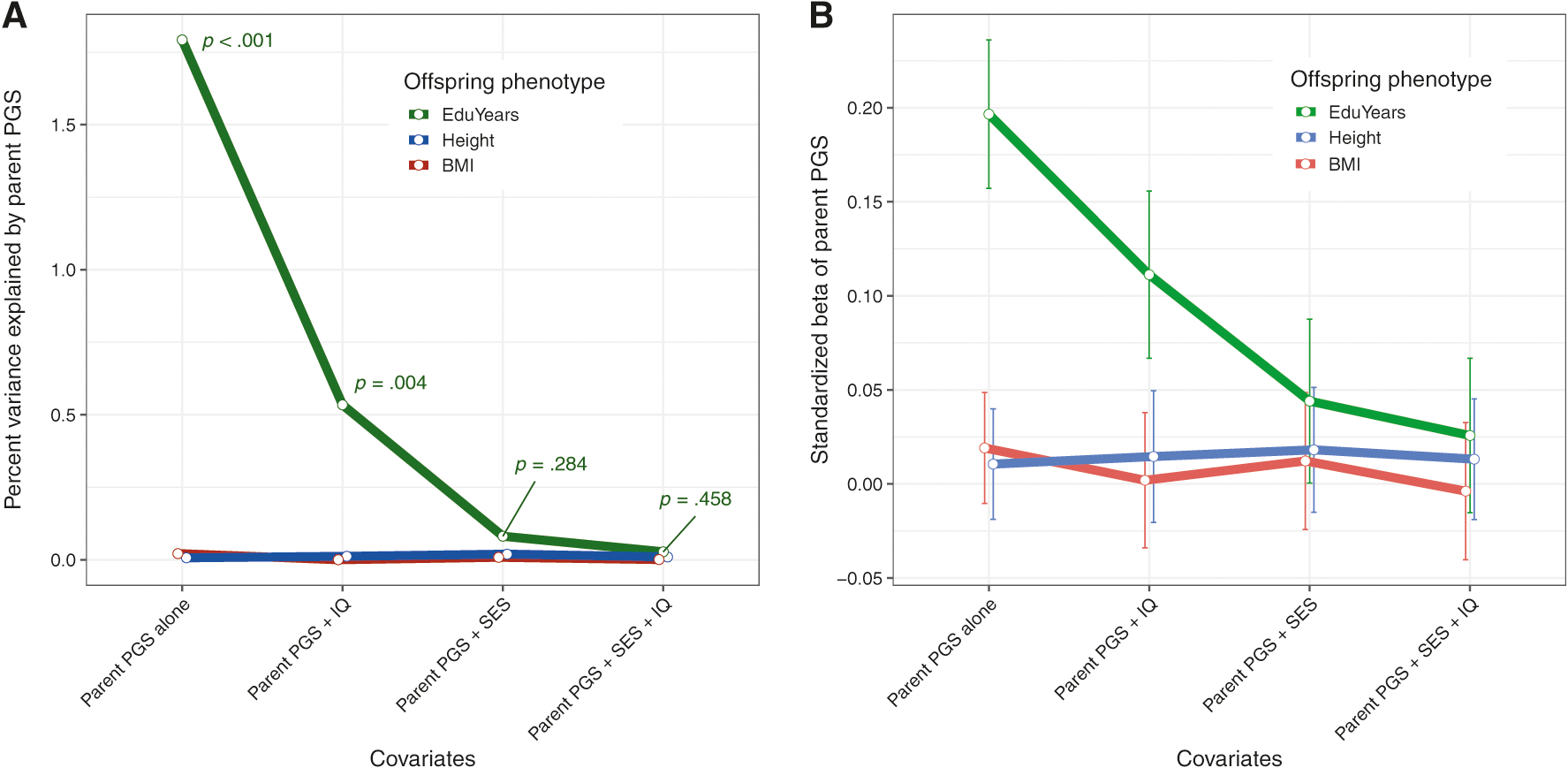
Line graphs comparing the effects of parent characteristics as covariates on a the incremental variance explained and b size of the standardized β-coefficient of parental polygenic score (PGS). Each regression model used the parent and offspring PGS specific to its dependent variable (EduYears, height, or body-mass index (BMI)). The error bars in panel b represent ± 1 standard error
Discussion
Through the use of polygenic scores constructed from the largest GWAS of educational attainment yet conducted [9], we have provided evidence of causal mechanisms underlying the relationship between genotype and phenotype by examining the effects of these scores in a sample of families. While it is well known that genes can affect an individual’s environment [31, 32], and that observed environmental effects on phenotypes may be at least partially under genetic control [33], the study of these effects has historically been confined to traditional twin and family designs. The application of GWAS results to data from family units of parents and their twin offspring permits causal inference to be drawn from within-family prediction models that are robust against confounding. This design also enables us to use mean polygenic scores of mothers and fathers to predict offspring outcomes, thereby testing for the presence of genetic nurture. These techniques, which make use of “triangulation” [34] to elucidate an effect, have produced evidence consistent with environmental causation.
Our results indicate a difference in magnitude of the effects of “genetic nurture” on different outcomes. For example, while polygenic score associations with education and IQ are of similar magnitude, trans-generational effects of parental genome on offspring outcomes—that is, “genetic nurture”—are smaller for IQ than for education. Although this is possibly a chance difference, this pattern of influence is consistent with the body of literature on how families affect children’s outcomes, particularly the well-established finding from 50 years of twin studies that shared environment effects are substantial for educational attainment but are often negligible for IQ. The 2015 twin study meta-analysis conducted by Polderman et al. [2], for example, reports shared environment as explaining an average of 27% of the variance in educational attainment. By contrast, they report the average shared environment effect of “intellectual functions” at 12.3%. At the same time, it is noteworthy that the within-twinship association of PGS with IQ is attenuated by roughly 50% of the individual-level effect (Fig. 2). Given that our polygenic scores are constructed from a GWAS of years of education rather than IQ, it is unclear how this ought to be interpreted from this dataset. This finding should be investigated analytically and empirically in future research, taking note of changes in the magnitudes of heritable and environmental influences on IQ with age.
Socioeconomic status (i.e., a composite of parental education, income, and occupational status) emerges as a plausible explanation for the effect parents are able to have on their children’s educational attainment. The finding that socioeconomic status and its correlates have substantial effect on the stratification of cognitive and educational outcomes is well-established in the behavior genetics literature, and they have recently been found to account for the majority of shared environment variance in education and IQ [35]. Moreover, there are some plausible pathways between parental SES and offspring educational attainment: higher-SES parents may, among other things, be able to provide financial assistance to their offspring in obtaining a college education.
The use of BMI and height as negative controls adds further credence to the genetic nurture interpretation of the effect of parental genome on offspring years of education. The near-zero effect sizes for genetic nurture on these phenotypes as compared to years of education is consistent with the latter being a real effect. The causal pathway from parental attributes to offspring education is plausible, straightforward, and consistent with our covariate analysis of SES and other parental traits that may mediate genetic nurture. A similar pathway for BMI or height is difficult to conceive and certainly less straightforward: It is hard to imagine what mechanism would connect parental genetics to the height or BMI of their offspring other than simple genetic inheritance. For example, the height of a parent might change over time, since people tend to become slightly shorter as they age, but the propagation of any such change to the offspring’s height seems highly unlikely. However, it is important to note that the existence of such a pathway might be more plausible in a population that has not developed in a modern environment where caloric intake is arguably excessive, and very likely at saturation in its ability to influence development. As such, nutrition is unlikely to be a mitigating issue for this finding in this population of Minnesotans, but care should be taken in generalizing the utility of these negative controls to other populations.
Although genomic methods can mitigate certain draw-backs of twin and adoption studies, these tools have their own limitations. Missing heritability is a perennial problem in GWA studies, and while each subsequent GWAS has shown that sample size dovetails with predictive value, we are still only able to explain a maximum of 13% of the variance in educational attainment with a polygenic score at this time. True causal inference is always difficult to establish in correlational genetic studies and our presented results, while consistent with a causal explanation of parental environment on offspring outcome, are no exception. Socioeconomic status as a mediating variable of this posited influence is a complex phenotype and could be correlated with any number of variables that contribute to a causal effect on offspring outcome, such as parents’ level of education. Additionally, the generalizability of our findings is limited by the single point in time at which phenotypes such as IQ were measured in offspring. It remains an open question whether and to what extent the effects of “genetic nurture” persist or change across childhood into adulthood, with previous literature suggesting that development plays a complex role in establishing genetic and environmental contributions to cognitive phenotypes [36].
The use of genome-wide association studies has opened up new possibilities in testing for the presence of environmental effects on phenotypes, and EA3 polygenic scores are our best genetic predictors of years of education yet constructed. By examining the net effect of genetic variants across generations, we can now directly test decades-old predictions about the passive interplay between genes and environment [4, 29, 31, 37]. The available data may present further opportunities to detect gene–environment interaction, reciprocal sibling effects, the effects of ontogenetic development on genetic nurture, and bidirectional parent–offspring transmission, as well as to further understand the nature of parental phenotypes, such as socioeconomic status, and whether and to what extent they causally influence offspring phenotypes. In sum, we have provided direct quantitative-genetic evidence consistent with an enduring effect that parents can have on the educational outcomes of their children.
Supplementary Material
Acknowledgements
Thanks to the Minnesota Center for Twin and Family Research staff for their expert management and synthesis of the data used in this study.
Funding The research reported here and the preparation of this manuscript were supported by grants from the U.S. National Institute on Alcohol Abuse and Alcoholism (AA09367, AA11886), the National Institute of Mental Health (MH066140), and the National Institute on Drug Abuse (DA05147, DA013240).
Footnotes
Conflict of interest The authors declare that they have no conflict of interest.
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information The online version of this article (https://doi.org/10.1038/s41380-019-0494-1) contains supplementary material, which is available to authorized users.
References
- 1.Turkheimer E. Three laws of behavior genetics and what they mean. Curr Directions Psychol Sci. 2000;9:160–4. 10.1111/1467-8721.00084 [DOI] [Google Scholar]
- 2.Polderman TJ, Benyamin B, De Leeuw CA, Sullivan PF, Van Bochoven A, Visscher PM, et al. Meta-analysis of the heritability of human traits based on fifty years of twin studies. Nat Genet. 2015;47:702–9. 10.1038/ng.3285 [DOI] [PubMed] [Google Scholar]
- 3.Dudbridge F. Power and predictive accuracy of polygenic risk scores. PLoS Genet. 2013;9:e1003348. 10.1371/journal.pgen.1003348 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Plomin R, DeFries JC, Loehlin JC. Genotype–environment interaction and correlation in the analysis of human behavior. Psychol Bull. 1977;84:309–22. 10.1037/0033-2909.84.2.309 [DOI] [PubMed] [Google Scholar]
- 5.Plomin R, Loehlin JC, DeFries JC. Genetic and environmental components of “environmental” influences. Dev Psychol. 1985;21:391–402. 10.1037/0012-1649.21.3.391 [DOI] [Google Scholar]
- 6.Sacerdote B. How large are the effects from changes in family environment? A study of Korean American adoptees. Q J Econ. 2007;122:119–57. 10.1162/qjec.122.1.119 [DOI] [Google Scholar]
- 7.D’Onofrio BM, Turkheimer EN, Eaves LJ, Corey LA, Berg K, Solaas MH, et al. The role of the children of twins design in elucidating causal relations between parent characteristics and child outcomes. J Child Psychol Psychiatry Allied Discip. 2003;44:1130–44. 10.1111/1469-7610.00196 [DOI] [PubMed] [Google Scholar]
- 8.D’Onofrio BM, Turkheimer E, Emery RE, Slutske WS, Heath AC, Madden PA, et al. A genetically informed study of the processes underlying the association between parental marital instability and offspring adjustment. Dev Psychology. 2006;42:486–99. 10.1037/0012-1649.42.3.486 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lee JJ, Wedow R, Okbay A, Kong E, Maghzian O, Zacher M, et al. Gene discovery and polygenic prediction from a genome-wide association study of educational attainment in 1.1 million individuals. Nat Genet. 2018;50:1112–21. 10.1038/s41588-018-0147-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Selzam S, Krapohl E, Von Stumm S, O’Reilly PF, Rimfeld K, Kovas Y, et al. Predicting educational achievement from DNA. Mol Psychiatry. 2017;22:267–72. 10.1038/mp.2016.107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sniekers S, Stringer S, Watanabe K, Jansen PR, Coleman JR, Krapohl E, et al. Genome-wide association meta-analysis of 78,308 individuals identifies new loci and genes influencing human intelligence. Nat. Genet 2017; 49. 10.1038/ng.3869 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kong A, Thorleifsson G, Frigge ML, Vilhjálmsson BJ, Young AI, Thorgeirsson TE, et al. The nature of nurture: effects of parental genotypes. Science. 2018;359:424–8. 10.1126/science.aan6877 [DOI] [PubMed] [Google Scholar]
- 13.Bates TC, Maher BS, Medland SE, McAloney K, Wright MJ, Hansell NK, et al. The nature of nurture: Using a virtual-parent design to test parenting effects on children’s educational attainment in genotyped families. Twin Res Hum Genet. 2018;21:73–83. 10.1017/thg.2018.11 [DOI] [PubMed] [Google Scholar]
- 14.Belsky DW, Domingue BW, Wedow R, Arseneault L, Boardman JD, Caspi A, et al. Genetic analysis of social-class mobility in five longitudinal studies. Proc Natl Acad Sci. 2018;115:E7275–84. 10.1073/pnas.1801238115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Liu H. Social and genetic pathways in multigenerational transmission of educational attainment. Am Sociol Rev. 2018;83:278–304. 10.1177/0003122418759651 [DOI] [Google Scholar]
- 16.Fisher RA. Average excess and average effect of a gene substitution. Ann Eugenics. 1941;11:53–63. 10.1111/j.1469-1809.1941.tb02272.x [DOI] [Google Scholar]
- 17.Lee JJ, Chow CC. The causal meaning of Fisher’s average effect. Genet Res. 2013;95:89–109. 10.1017/S0016672313000074 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Miller MB, Basu S, Cunningham J, Eskin E, Malone SM, Oetting WS, et al. The Minnesota Center for Twin and Family Research genome-wide association study. Twin Res Hum Genet. 2012;15:767–74. 10.1017/thg.2012.62 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Vilhjálmsson BJ, Yang J, Finucane HK, Gusev A, Lindström S, Ripke S, et al. Modeling linkage disequilibrium increases accuracy of polygenic risk scores. Am J Hum Genet. 2015;97:576–92. 10.1016/j.ajhg.2015.09.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Loh PR, Kichaev G, Gazal S, Schoech AP, Price AL. Mixed-model association for biobank-scale datasets. Nat Genet. 2018;50:906–8. 10.1038/s41588-018-0144-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wechsler D. Wechsler adult intelligence scale-revised. San Antonio, TX: Psychological Corporation; 1981. [Google Scholar]
- 22.Wechsler D. Manual for the Wechsler Intelligence Scale for Children–Revised. New York, NY: Psychological Corporation; 1974. [Google Scholar]
- 23.Schmidt FL, Hunter J. General mental ability in the world of work: Occupational attainment and job performance. J Pers Soc Psychol. 2004;86:162–73. 10.1037/0022-3514.86.1.162 [DOI] [PubMed] [Google Scholar]
- 24.Gottfredson LS. Why g matters: the complexity of everyday life. Intelligence. 1997;24:79–132. 10.1016/S0160-2896(97)90014-3 [DOI] [Google Scholar]
- 25.McGue M, Rustichini A, Iacono WG. Cognitive, noncognitive, and family background contributions to college attainment: a behavioral genetic perspective. J Personal. 2017;85:65–78. 10.1111/jopy.1223 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Stumm S, von, Plomin R. Socioeconomic status and the growth of intelligence from infancy through adolescence. Intelligence. 2015;48:30–36. 10.1016/j.intell.2014.10.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Strenze T. Intelligence and socioeconomic success: A meta-analytic review of longitudinal research. Intelligence. 2007;35:401–26. 10.1016/j.intell.2006.09.004 [DOI] [Google Scholar]
- 28.Belsky DW, Moffitt TE, Corcoran DL, Domingue B, Harrington H, Hogan S, et al. The genetics of success: How single-nucleotide polymorphisms associated with educational attainment relate to life-course development. Psychol Sci. 2016;27:957–72. 10.1177/0956797616643070 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lee JJ. Correlation and causation in the study of personality. Eur J Personal. 2012;26:372–90. 10.1002/per.1863 [DOI] [Google Scholar]
- 30.Pearl J. Causality: models, reasoning, and inference. 2nd ed. New York, NY: Cambridge University Press, 2009. [Google Scholar]
- 31.Plomin R, Bergeman CS. The nature of nurture: Genetic influence on “environmental” measures. Behav Brain Sci. 1991;14:373–86. 10.1017/S0140525X00070278 [DOI] [Google Scholar]
- 32.Plomin R. Genetics and experience: the interplay between nature and nurture. Thousand Oaks, CA, US: Sage Publications, Inc; 1994. [Google Scholar]
- 33.Vinkhuyzen AA, Van Der Sluis S, De Geus EJ, Boomsma DI, Posthuma D. Genetic influences on “environmental” factors. Genes Brain Behav. 2010;9:276–87. 10.1111/j.1601-183X.2009.00554.x [DOI] [PubMed] [Google Scholar]
- 34.Munafò MR, Smith DG. Repeating experiments is not enough. Nature. 2018;553:399–401. 10.1038/d41586-018-01023-3 [DOI] [Google Scholar]
- 35.Engelhardt LE, Church JA, Harden KP, Tucker-Drob EM. Accounting for the shared environment in cognitive abilities and academic achievement with measured socioecological contexts. Dev Sci. 2019;22:e12699. 10.1111/desc.12699 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Tucker-Drob EM, Briley DA, Harden KP. Genetic and environmental influences on cognition across development and context. Curr Dir Psychol Sci. 2013;22:349–55. 10.1177/0963721413485087 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Scarr S, McCartney K. How people make their own environments: a theory of genotype–environment effects. Child Dev. 1983;54:424. 10.2307/1129703 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.