

Summary
Photosynthesis is currently a focus for crop improvement. The majority of this work has taken place and been assessed in leaves, and limited consideration has been given to the contribution that other green tissues make to whole‐plant carbon assimilation. The major focus of this review is to evaluate the impact of non‐foliar photosynthesis on carbon‐use efficiency and total assimilation. Here we appraise and summarize past and current literature on the substantial contribution of different photosynthetically active organs and tissues to productivity in a variety of different plant types, with an emphasis on fruit and cereal crops. Previous studies provide evidence that non‐leaf photosynthesis could be an unexploited potential target for crop improvement. We also briefly examine the role of stomata in non‐foliar tissues, gas exchange, maintenance of optimal temperatures and thus photosynthesis. In the final section, we discuss possible opportunities to manipulate these processes and provide evidence that Triticum aestivum (wheat) plants genetically manipulated to increase leaf photosynthesis also displayed higher rates of ear assimilation, which translated to increased grain yield. By understanding these processes, we can start to provide insights into manipulating non‐foliar photosynthesis and stomatal behaviour to identify novel targets for exploitation in continuing breeding programmes.
Keywords: photosynthesis, Calvin–Benson cycle, sink capacity, wheat ears, non‐foliar organs, stomata
Significance Statement
Measurements of photosynthesis focus on leaves, however non‐foliar green tissue can make significant contributions to total plant carbon assimilation. This review evaluate photosynthesis and stomatal behavior in non‐foliar organs with a view to manipulate these processes for improved yield or quality.
Introduction
Photosynthesis in leaves is a well‐established and extremely well researched process whereby plants harvest the energy from sunlight and use this to convert CO2 into soluble carbohydrates, which are subsequently used for plant growth (Calvin and Benson, 1948; Bassham and Calvin, 1960; Raines, 2003; Biel and Fomina, 2015). Photosynthesis is responsible, therefore, either directly (through plant growth) or indirectly (through the food chain), for all food consumed worldwide. The majority of studies on photosynthesis often only consider photosynthesis in leaves, with little appreciation of potential carbon assimilation in other green non‐foliar tissue and its contribution to overall yield. With the predicted requirement to double food production by the year 2020 (WorldBank, 2008; RSOL, 2009; Tilman and Clark, 2015; FAO, 2017) and the fact that annual genetic gains in yield, using current breeding approaches, are reducing or slowing for many crops (Ray et al., 2012; Ray et al., 2013), research into photosynthesis and the processes associated with it are being increasingly recognized as potential novel targets for improving crop yield. Crop yield is determined by the cumulative rate of photosynthesis over the growing season. The maximum yield obtained (yield potential), defined as the yield obtainable when a crop is grown in optimal conditions with no biotic or abiotic stress (Evans and Fischer, 1999), is the result of three key determinants: (i) light capture; (ii) radiation use efficiency (RUE) or energy conversion efficiency (the product of which is biomass); and (iii) harvest index (HI, the partition of harvestable produce relative to plant biomass) (Reynolds et al., 2009). Significant gains in both HI and light interception have been made over the last several decades, with considerable increases in HI following the green revolution and the introduction of dwarfing (Rht) genes (Gale and Youssefian, 1985; Calderini et al., 1995). The current focus is on RUE (Reynolds et al., 2009; Parry et al., 2011), which is primarily photosynthesis and the conversion of light energy into fixed carbon. Several recent studies have demonstrated that improving diverse aspects of photosynthesis in leaf tissue, including altering key enzymes within the Calvin–Benson cycle (CBC) (Lefebvre et al., 2005; Simkin et al., 2015; Driever et al., 2017; Simkin et al., 2017a), electron transport (Chida et al., 2007; Simkin et al., 2017b; Yadav et al., 2018; Ermakova et al., 2019), photorespiration (Timm et al., 2012; López‐Calcagno et al., 2018) and the kinetics of non‐photochemical quenching (NPQ) (Kromdijk et al., 2016; Glowacka et al., 2018) can improve yield potential in both glasshouse‐ and field‐grown plants (Simkin, 2019; Simkin et al., 2019). Leaves are not the only location within the plant where photosynthesis occurs, however, with evidence that petioles and stems (Hibberd and Quick, 2002), seeds (Schwender et al., 2004), fruit (Hetherington et al., 1998; Carrara et al., 2001; Hiratsuka et al., 2015; Sui et al., 2017), Triticum aestivum (wheat) ears (Maydup et al., 2010), and the husks of Zea mays (corn) (Pengelly et al., 2011) all photosynthesize and may provide significant and alternative sources of the photoassimlates, essential for optimal yield. Figure 1 illustrates chlorophyll fluorescence imaging of the operating efficiency of photosystem II (PSII) photochemistry () in non‐leaf tissues, which is indicative of functional electron transport in these green non‐leaf organs. To date little data exist on how potential manipulation of photosynthetic processes may impact these chlorophyll‐containing tissues.
Figure 1.
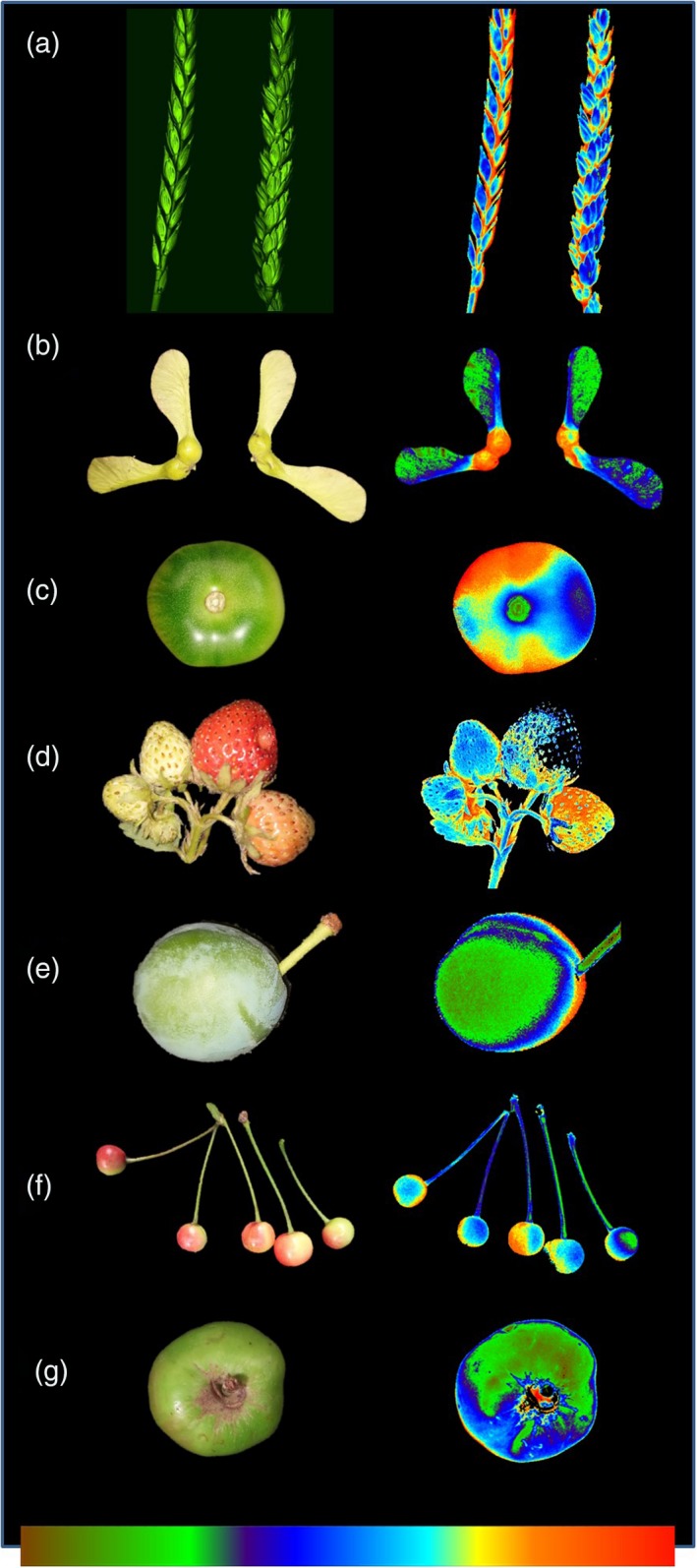
Chlorophyll fluorescence (CF) images of photosystem II (PSII) operating efficiency () in green non‐leaf tissue was used to demonstrate photosynthetic electron transport. CF images: (a) wheat ear; (b) sycamore seed pods; (c) tomato fruit; (d) strawberry fruit; (e) greengage; (f) cherries; and (g) apples. Colour scale bar represents an of: (a) 0.45–0.75; (b) 0.30–0.55; (c) 0.50–0.70; (d) 0.50–0.70; (e) 0.5–0.75; (f) 0.5–0.80; and (g) 0.45–0.70.
The majority of studies that have examined photosynthesis in non‐foliar tissue have assumed and described a photosynthetic pathway similar to that of the mesophyll. One key difference in non‐foliar tissue photosynthesis is the fact that there are two potential major sources of CO2. First, ribulose‐1,5‐bisphosphate carboxylase (Rubisco) assimilates atmospheric CO2 that diffuses into the cells through the stomatal pores, leading to the production of sugars via the CBC, similar to the CO2 pathway in leaf (C3) tissue. Second, CO2 released by mitochondrial respiration can be the main supply of CO2 and is refixed (recycling photosynthesis; Aschan and Pfanz, 2003; Millar et al., 2011), and there is limited diffusion and supply of external CO2. Although stomata are present in various numbers on some non‐foliar tissues their function has not been fully evaluated, and the amount of photosynthesis that relies on the atmospheric supply of CO2 through these pores is not currently known. In this review we focus on photosynthesis in non‐foliar tissues and the potential contribution to yield, as well as the role of stomata in this process. Before discussing the possibility to manipulate non‐foliar photosynthesis for improved productivity or nutritional quality, we first provide an overview of what is known about photosynthesis in various organs, focusing on stems and fruits as well as various parts of cereals.
Photosynthesis in stems
Stems act as temporary storage sites for photoassimilates from leaves and carry out photosynthesis in their own right (Aschan and Pfanz, 2003). In Solanum lycopersicum (tomato), chlorophyll levels were found to be higher in the upper parts of the stem than in the lower parts of the stem (Xu et al., 1997), and a comparison of the photosynthetic activity of various plant parts found the entire stem accounted for up to 4% of photosynthetic activity (Hetherington et al., 1998). The contribution of stem photosynthesis to yield has been demonstrated in Gossypium hirsutum (cotton) by Hu et al. (2012) who reported that keeping the main stem in darkness reduced seed weight by 16% (Hu et al., 2012). These findings were supported by Simbo et al. (2013), who showed that when light was excluded from the stem of defoliated Adansonia digitata L. (African baobab) and Ricinus communis (castor bean) a reduction in bud dry weight was observed, providing further evidence for the importance of the stem for providing photoassimilates for plant development and growth. In some plants, such as Justicia californica, flowers and fruits develop in the absence of leaves, where the stem is the only photosynthetically active tissue (Tinoco‐Ojanguren, 2008; Ávila et al., 2014), also highlighting the role of stem photosynthesis for reproductive success. This is emphasized further by a reported stem photosynthesis equivalent to 130% of leaf levels in this species (Tinoco‐Ojanguren, 2008), whereas in other species, rates of between 16 and 60% relative to leaf levels have been reported (Ehleringer et al., 1987; Ávila et al., 2014). In the woody plant Eucalyptus photosynthesis in chlorophyll‐containing tissue, chlorenchyma, located beneath the periderm layer (Pfanz et al., 2002; Manetas, 2004), known as corticular photosynthesis (CP), contributed 11% of total photosynthate to plant growth, demonstrating the contribution of CP to eucalyptus growth (Cernusak and Hutley, 2011).
Stem photosynthesis is particularly important in deciduous species. In the summer‐deciduous, green‐stemmed Mediterranean shrub Calicotome villosa, the total branch photosynthesis is higher in the summer because of an absence of leaves, and green‐stem photosynthesis outcompetes leaf photosynthesis on an annual basis (Yiotis et al., 2008). In the desert ephemeral Erigonum inflatum substantial photosynthesis was demonstrated in the inflated stems, despite the fact that these contained only half the chlorophyll and nitrogen content of the leaves (Osmond et al., 1987). Internal CO2 concentrations in these stems was reported to be extremely high. Interestingly, fixation of this internal CO2 was between six and 10 times less than the fixation of atmospheric CO2; however, although small, this additional internal CO2 pool facilitated high water‐use efficiency (WUE, measured as water lost relative to carbon gained) as a result of no water loss through stomata for this carbon gain. Greater WUE was further enhanced in this species by smaller stem stomata that are more responsive to temperature and high vapour pressure deficit (VPD), compared with their leaf counterparts (Osmond et al., 1987). The importance of stem photosynthesis in desert species is supported by a more recent study by Avila‐Lovera et al. (2017), who examined 11 green‐stemmed desert plants and revealed coordination between stem photosynthesis and hydraulics similar to that observed in leaves, with an even tighter relationship during the dry season, facilitating additional carbon gain and potential mechanisms for enhanced drought tolerance. Furthermore, stem photosynthetic rates were higher during the dry season when leaves were lost and light interception by the stems was increased, due to the absence of foliage (Avila‐Lovera et al., 2017). Together these studies illustrate the importance and annual contribution of stem photosynthesis to overall carbon gain, which not only contributes to the survival of plants growing in dry and hot environments, but also supports the notion that stem photosynthesis may contribute significantly to yield, and that this contribution may be more important under conditions such as reduced water availability, high temperatures and high VPD. To date, however, there have been limited studies that have evaluated the importance of stem photosynthesis to yield in key crop species. Therefore, although stem photosynthesis may represent a potential novel target to support enhanced photosynthetic carbon gain, particular under conditions of water stress (such as those predicted under climate change for certain agricultural areas), more quantitative information on stem performance in crops is needed to evaluate and fully exploit this process.
Fruit photosynthesis
Fruit photosynthesis is particularly interesting, as many species (e.g. tomato) undergo a shift from green photosynthetic (or partial photosynthetic) to fully heterotrophic metabolism on ripening (Lytovchenko et al., 2011). As early as 1974, Tanaka and coworkers conducted shading experiments on tomato fruits and showed that fruit photosynthesis contributes to net sugar accumulation and growth (Tanaka et al., 1974), and from this work concluded that photosynthesis contributed between 10 and 15% of the total fixed carbon, which was later confirmed by Hetherington et al. (1998) and Obiadalla‐Ali et al. (2004). In addition to showing a similar photosynthetic function to leaves, developing tomato fruit have also been reported to have approximately 41% of the photosynthetic electron transport capacity of leaf tissue (Piechulla et al., 1987). Recent proteomic analysis has demonstrated that all of the components of the CBC and photorespiratory cycle accumulate at the protein level in tomato fruit (Barsan et al., 2010; Barsan et al., 2012). The major light‐harvesting proteins, including the thylakoid membrane light‐harvesting complex proteins of PSI (psaA) and PSII (psbA), and the chlorophyll a/b binding proteins, have also been observed (Piechulla et al., 1986; Lemaire‐Chamley et al., 2005), in conjunction with plastocyanin, cytochrome f, cytochrome b, ferredoxins, Rieske iron sulphur protein (Piechulla et al., 1987; Livne and Gepstein, 1988; Cheung et al., 1993; Aoki et al., 1998) and the CBC proteins, Rubisco and fructose 1,6‐bisphophate aldolase (FBPaldolase) (Barsan et al., 2010; Steinhauser et al., 2010). Rubisco assays have also demonstrated that the enzyme is active in tomato fruit (Willmer and Johnston, 1976; Bravdo et al., 1977; Laval‐Martin et al., 1977; Piechulla et al., 1987; Sugita and Gruissem, 1987).
Despite the fact that transcriptomic and metabolomic analyses have revealed high expression levels of many of these photosynthetic genes in tomato fruit, and have shown that photosynthetic carbon assimilation in these organs makes an important contribution to early fruit development (Wang et al., 2009), many studies do not agree that these fruit are net assimilators of CO2 (see Blanke and Lenz, 1989; Carrara et al., 2001). Lytovchenko et al. (2011) used antisense technology to reduce expression of the chlorophyll biosynthesis gene glutamate 1‐semialdehyde aminotransferase, which resulted in a reduced photosynthetic rate; however, fruit size and metabolite levels remained unchanged. These authors suggested that transport of photosynthate from leaves compensated for any reduction in fruit localized photosynthetic rates and proposed that fruit photosynthesis is dispensable. However, a delay in seed development was observed, suggesting that localized CO2 fixation/re‐assimilation may be important for seed formation (Lytovchenko et al., 2011). In contrast, another study demonstrated that decreased expression of fruit chloroplastic fructose‐1,6‐bisphosphatase (FBPase) resulted in a 15–20% negative impact on fruit development (Obiadalla‐Ali et al., 2004). Lytovchenko et al. (2011) suggested that these contradictory results could be explained by different promotor specificity and/or the impact of reduced FBPase activity later in the development of the fruit.
Although it is evident that photosynthesis occurs in fruits, the extent and importance is not clear. The fact that tomato fruit lack stomata (Vogg et al., 2004) (Figure 2) implies that photosynthesis in these organs relies exclusively on CO2 liberated from mitochondria, that no ‘new’ carbon is fixed and that photosynthesis functions to re‐assimilate CO2 (recycling photosynthesis) that would otherwise be lost. This is supported by the reported accumulation of transcripts in tomato loculare tissue associated with photosynthesis, clearly demonstrating photosynthetic capacity, but alongside high measured respiration rates (Lemaire‐Chamley et al., 2005). CO2 generated by the oxidative pentose pathway is re‐assimilated by the CBC in a manner previously reported in green seeds of Brassica napus (oilseed rape) (Schwender et al., 2004). It has been reported that these photosynthesis‐specific transcripts are regulated by transcription factors in a similar way to those in leaf tissue (Hetherington et al., 1998; Carrara et al., 2001); however, a number of authors have reported the existence of some fruit‐specific regulation of photosynthetic genes (Piechulla et al., 1987; Piechulla and Gruissem, 1987; Sugita and Gruissem, 1987; Manzara et al., 1993), and Cocaliadis et al. (2014) suggested that this is likely to optimize photosynthetic function for fruit development. This specificity therefore provides a potential route for manipulating key photosynthetic genes specifically in fruit to enhance development, yield or nutritional quality.
Figure 2.

Example of epidermal impressions taken from tomato (b) and cucumber (d). Photographs of the fruit are presented in (a) and (c). Stomata were absent from the epidermis of tomato (b), whereas a relatively high stomatal density is illustrated in cucumber (d, with the inset showing a magnified stomatal complex).
In summary, it appears that photosynthetic carbon assimilation does take place in green immature tomato fruit and that this relies almost exclusively on respired CO2, and that any reductions in the rate of photosynthesis in these organs can be compensated for by the upregulation of leaf photosynthesis (Nunes‐Nesi et al., 2005; Araújo et al., 2011) and increased imported photoassimilates from leaves. Such import cannot compensate for the losses of fruit photosynthesis for seed set, establishment and development (Lytovchenko et al., 2011). Therefore, altering fruit photosynthesis could provide advantages of early seed set, as well as maintaining yield, particularly under conditions of stress when leaf photosynthesis may be compromised.
Tomato photosynthesis is restricted to the green phases of development up until chloroplast‐to‐chloroplast differentiation, which is marked by the loss of chlorophyll, the degradation of the thylakoid membranes, and a strong decrease in the levels of photosynthesis‐associated transcripts and proteins (Harris and Spurr, 1969a; Harris and Spurr, 1969b; Cheung et al., 1993; Barsan et al., 2012), after which the fruit continues to develop and ripen. This is similar for other fruits such as Capsicum annum (pepper) (Steer and Pearson, 1976), Citrus unshiu (satsuma mandarin) (Hiratsuka et al., 2015), blueberry (Birkhold et al., 1992); coffee (Coffea arabica) (Cannell, 1985; Lopez et al., 2000); Prunus tomentova (plum) (Aoyagi and Bassham, 1984); the ornamental plant Arum italicum (Ferroni et al., 2013) and Jatropha curcas (Ranjan et al., 2012). In satsuma mandarin, it has been demonstrated that photosynthesis occurs in these fruits, is greater at low irradiances, and increases with increasing [CO2] supplied through fully developed stomata in the rind of satsuma (Hiratsuka et al., 2015). The fact that stomata can be found in densities of about 72 mm−2 in immature Jatropha curcas fruit suggests that new carbon can be assimilated through these tissues (Ranjan et al., 2012). In this case, given the importance of fruit photosynthesis in the absence of leaves, increasing the stomatal density could increase CO2 uptake and boost photosynthetic rates in fruit, with a positive impact on yield.
Cucumis sativus (cucumber) is fundamentally different to tomato and other coloured fruit, remaining green through to full maturity, with a surface area equivalent to a fully expanded leaf (Sui et al., 2017). An analysis of gene expression found a number of CBC enzymes (SBPase, FBPase, rbcL, rbcS) and light‐harvesting complex proteins of PSI (Lhca) and PSII (Lhcb) expressed in the exocarp (Sui et al., 2017). Interestingly, unlike tomato, stomata are found on the epidermis of cucumbers (Figure 2), although Sui et al. (2017) reported a layer of epicuticular waxes around the guard cells that may reduce function. However, the presence of these pores on the fruit surface suggests, in the case of cucumber at least, that these fruits are capable of assimilating some CO2 directly from the atmosphere. Their physiology also suggests that photosynthesis can occur from the re‐assimilation of respiratory CO2. Cucumber fruits have been shown to have both high photosynthetic and respiratory rates (Todd et al., 1961), and a recent study demonstrated that fruit photosynthesis contributed 9.4% of its own carbon requirements whereas 88% of respiratory CO2 in fruit was refixed (Sui et al., 2017). Improving photosynthetic efficiency in fruit, therefore, has the potential to increase the fruit carbon contribution for growth through both recycling respiratory CO2 and atmospheric assimilation, which could in turn directly impact WUE. The need to maintain or increase fruit yield (or fruit size) whilst using less water cannot be underestimated given current environmental changes.
Are stomata important in fruit photosynthesis?
It is important to note that although stomata are routinely found on the surface of some fruit and are of a similar size to stomata found on respective leaves, the numbers are generally significantly lower compared with those found in leaf tissue (Blanke, 1998). For example, Blanke and Lenz (1998) reported that the number of stomata on mature Malus domestica (apple) fruit was 30 times less abundant than the stomata found on apple leaves. Stomatal numbers are fixed at anthesis and as the fruit expands during growth, they become more dispersed (Hieke et al., 2002; Hetherington and Woodward, 2003). Although it has been reported that stomatal density in fruit typically represents 1–10% of the frequency found in corresponding leaf tissue (Sánchez et al., 2013), these numbers can vary greatly depending on the species. In Persea americana (avocado), the number of stomata on the fruit represent 14% of the number on the leaf (Blanke, 1992), whilst in green coffee fruit this number is 13–23% (Cannell, 1985), whereas in oranges the stomatal densities can reach up to 30% of those found on leaves (Moreshet and Green, 1980). To date, most studies have focused on the presence of stomata on various fruit tissue but have not fully demonstrated the functionality. If functional, however, the presence and stomatal densities reported above suggest that under certain conditions, in certain plants at least, stomata may play a role in gas exchange and therefore manipulating stomatal numbers through developmental mechanisms or transgenic approaches has the potential to change CO2 assimilation rates and yields. In other plants, however, the contribution of stomata to assimilation appears to be negligible compared with recycling photosynthesis. In these plants, we cannot rule out that the role of stomata is primarily for evaporative cooling. Although not directly related to CO2 uptake, this process may help maintain fruit temperature at an optimal level for recycling photosynthesis, thereby maximising CO2 recovery.
Seed and embryo photosynthesis
The fruit pericarp is not the only non‐foliar green tissue that is capable of photosynthesis. The embryos of many taxa contain significant quantities of chlorophyll, which persists until maturity (Yakovlev and Zhukova, 1980; Simkin et al., 2010; Puthur et al., 2013; Smolikova and Medvedev, 2016). This group includes model species (Arabidopsis thaliana) and important crops such as Cicer arietinum L. (chickpeas), coffee, cotton, Glycine max L. (soybean), oilseed rape, Pisum sativum L. (peas), and Vicia faba L. (broad beans). These embryos, first referred to as chloroembryos by Palanisamy and Vivekanandan (1986), contain all the photosynthetic complexes of PSI and PSII, cytochrome b 6 f complex and ATP synthase (Weber et al., 2005; Allorent et al., 2015; Kohzuma et al., 2017). Chloroembryos have been shown to photosynthesize (Smolikova and Medvedev, 2016; Smolikova et al., 2017), and confirmation of carbon fixation is supported by the activity of the CBC enzymes NADP‐glyceraldehyde‐3‐phosphate dehydrogenase (GAPDH) in the chloroembryo chloroplasts of oilseed rape and pea (Smith et al., 1990; Eastmond et al., 1996) and fructose‐1,6‐bisphosphatase (FBPase) in oilseed rape (Kang and Rawsthorne, 1996). Furthermore, Rubisco has also been shown to be active in the seeds of soybean (Allen et al., 2009), oilseed rape (Hills, 2004; Ruuska et al., 2004), broad bean (Willmer and Johnston, 1976) and Trigonella foenum‐graecum (Willmer and Johnston, 1976).
The contribution of photosynthesis in embryos may be different to that described above for fruit, as it has been reported that embryo photosynthesis contributes a significant amount of oxygen, which fuels energy‐generating biochemical pathways, including respiration and glycolysis (Ruuska et al., 2004; Borisjuk et al., 2005; Tschiersch et al., 2011; Galili et al., 2014). The role of photosynthesis in chloroembryos has also been associated with the rapid synthesis of ATP and NADPH for the synthesis of complex carbohydrates, fatty acids and proteins (Asokanthan et al., 1997; Wu et al., 2014). It has been reported that a key source of carbon is sucrose, imported from the leaves (Asokanthan et al., 1997), which is respired by the seed, releasing CO2 (Ruuska et al., 2004; Smolikova and Medvedev, 2016) within chloroembryos, which is subsequently efficiently re‐assimilated and thus directly affects the carbon economy of the seed (Puthur et al., 2013).
In oilseed rape, seed photosynthesis plays a role in the accumulation of storage lipids (Eastmond et al., 1996; Ruuska et al., 2004). Interestingly, Rubisco acts in a distinctive context, without the CBC, to increase the carbon‐use efficiency for the synthesis of oil (Schwender et al., 2004). This unique pathway generates 20% more acetyl‐CoA than glycolysis, reducing the loss of CO2 and increasing the availability of acetyl‐CoA for fatty acid biosynthesis (Schwender et al., 2004). In the embryos of legumes, including pea, the main CO2‐refixing enzyme is phosphoenol pyruvate (PEP) carboxylase (Golombek et al., 1999) suggesting that CO2 is refixed at the site of origin. In the case of pea, a small spherical seed with a green embryo within a seed pod, only a fraction of light reaches the photosynthetically active tissue. The light is attenuated by the pod, reflecting or absorbing as much as 75% of the sunlight. Only 32% of the remaining sunlight (approximately 8% of photosynthetic active radiation (PAR)), penetrates the pod and seed coat to reach the surface of the embryo; however, this is enough to drive photosynthesis with the highest electron transport rates reported in the seed coat (Tschiersch et al., 2011). In addition to seed photosynthesis, pea pods also photosynthesize in two distinct layers. First, the outer layer, comprising chlorenchyma and mesocarp, assimilates CO2 from the atmosphere and second, the inner epidermis lining of the pod cavity reassimilates the CO2 released by the embryonic respiration into the pod cavity (Atkins et al., 1977). Rubisco activity has also been detected in the pod wall of pea embryos, although this activity is 10–100 times lower than that detected in the leaf tissue (Hedley et al., 1975).
Importance of photosynthesis in non‐foliar cereal organs
In cereals, although leaf photosynthesis plays a central role in biomass accumulation and yield formation over the entire growing season (Fischer et al., 1998; Gu et al., 2014), the photosynthetic activity of the ear has been shown to dramatically contribute to the pool of carbohydrates translocated to the developing grains over the post‐anthesis stages (Tambussi et al., 2005; Tambussi et al., 2007; Maydup et al., 2010; Sanchez‐Bragado et al., 2014). Although on an area basis, the ear CO2 assimilation rate is lower than that of the flag leaf (Tambussi et al., 2005; Tambussi et al., 2007; Zhou et al., 2016), experimental evidence suggests that in bread and durum wheat, ear photosynthesis can contribute to the individual grain weight yield component by up to 70% in a large range of genotypes (Maydup et al., 2010) and contrasting environments (Sanchez‐Bragado et al., 2014). Similarly to wheat, in Hordeum vulgare (barley), shading experiments revealed a significant contribution of the ear (up to 50%) to grain weight and therefore yield (Bort et al., 1994). In the next few sections we focus on different aspects of ear photosynthesis and the challenges in assessing photosynthesis in non‐foliar organs.
Photosynthetically active ear components
The ear bracts (which consist of glume, lemma and palea) contain chlorophyll and possess stomata (Figure 3), and therefore have potential to fix atmospheric CO2 (Tambussi et al., 2007). Genotypic variation in ear photosynthetic CO2 assimilation per unit area and contribution of ear photosynthesis to grain weight have been reported in the literature (Maydup et al., 2014; Sanchez‐Bragado et al., 2014). The exploitation of this variation might be of pivotal importance for cereal improvement. Several ear bracts have been considered putative locations of photosynthetic activity, with glumes, lemmas and awns considered the most photosynthetically active (Tambussi et al., 2007; Hu et al., 2019). In particular, the floral‐derived awns have been targeted as a trait to increase wheat yield owing to their high photosynthetic capacity of 7–35 µmol m−2 sec−1 (Hein et al., 2016), and especially in view of the limited possibility to further increase assimilates partitioning to grains by manipulating the harvest index (Maydup et al., 2014).
Figure 3.
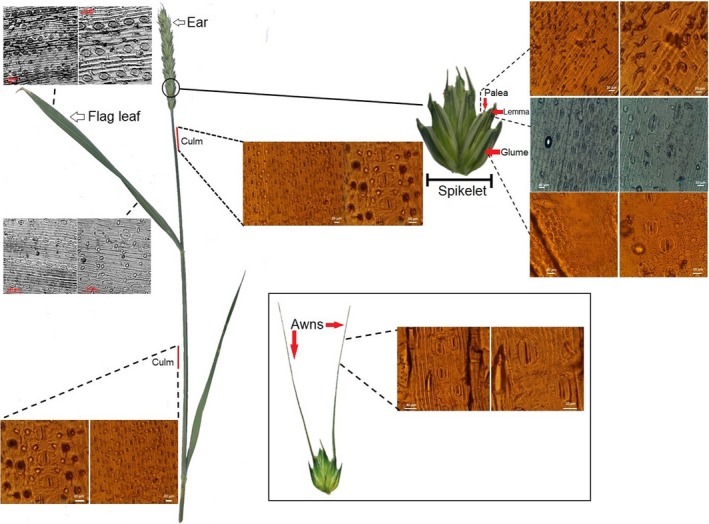
Schematic diagram and images of epidermal impressions illustrating stomatal anatomy and density in different components of wheat leaves (flag leaf), culm (stem) and ear (i.e. external surfaces of the glume, lemma and palea). The insert box provides an example of the stomatal density found on the awns of Soissons wheat.
The seasonality of the post‐anthesis stages in cereals are often associated with increases in environmental stresses and severe water deficit conditions, leading to reduced yield. Numerous studies provide strong evidence that the ear possesses an elevated drought tolerance when compared with the flag leaf and highlight the ear as the main potential buffer for photoassimilate production under disadvantageous environments (Jia et al., 2015). Additionally, the ear shows a lower transpiration rate than the flag leaf and a higher intrinsic WUE, confirmed by less negative δ13C values (Araus et al., 1993; Tambussi et al., 2007; Sanchez‐Bragado et al., 2014; Vicente et al., 2018). Xeromorphic characteristics in glumes, lemmas and awns of durum wheat have been observed, such as sclerenchymatous tissue and thick walls (Tambussi et al., 2005). The same authors observed a higher osmotic adjustment and relative water content of the ear compared with the flag leaf under reduced water availability, leading to a sustained chlorophyll fluorescence signal. Similarly, ear bracts in barley maintained higher relative water content and gas exchange under water stress, compared with the leaf, as well as greater osmotic adjustment (Hein et al., 2016).
Comparing awned and awnless lines under stress conditions showed higher ear intrinsic WUE (mainly driven by high photosynthetic activity for similar stomatal conductance (g s per unit area) and photosynthetic capacity when awns were present, suggesting that awn photosynthesis also plays an important role when foliar tissue is reduced as a result of stress (Weyhrich, 1994; Weyhrich et al., 1995). However, no differences in whole‐plant WUE and grain weight were found between these lines. Therefore, in this investigation, the higher photosynthetic capacity in the awns failed to contribute to yield. In contrast, a multi‐location field study on the effect of awns on wheat yield components showed that the presence of awns increased the grain size; however, this increase was compensated by a reduction in grain number (Rebetzke et al., 2016), which was mainly attributed to the cost of awn setting. Assimilate partitioning to the floret is decreased in awned varieties through allocation to the rapidly growing awns, potentially followed by an associated reduction in floret fertility (Guo and Schnurbusch, 2016; Rebetzke et al., 2016). It was concluded that awns are mainly useful under terminal drought conditions, owing to their elevated water stress tolerance that facilitates the maintenance of grain weight and a reduced number of shrivelled grains (screenings), compared with awnless lines, thus potentially providing higher economic yield and commercial value under such conditions. This was also confirmed by Maydup et al. (2014), who showed that awned varieties have higher ear photosynthesis, water status and ear water conductance compared with awnless varieties under water‐stress conditions in the field.
Genotypic variation of ear water‐stress tolerance has also been demonstrated by Li et al. (2017), where a stress‐tolerant wheat variety displayed a conservative water‐use strategy during post‐anthesis by reducing leaf transpiration while maintaining high levels of ear gas exchange. Vicente et al. (2018) postulated that water stress in wheat reduced the expression of photosynthetic genes (e.g. ATPase) in the flag leaf but not in the ear, and that the upregulation of respiration‐related genes, such as phosphoenolpyruvate carboxylase (PEPCase), 2‐oxoglutarate dehydrogenase complex (OGDC), alternative oxidase (AOX) and pyruvate kinase, was associated with the increased refixed CO2 in the ear organs. An observed upregulation of dehydrins (Abebe et al., 2010), increased transcript levels of antioxidant enzyme genes (Vicente et al., 2018), followed by high levels of antioxidant enzymes and low levels of ROS (Kong et al., 2015) confirmed the higher drought tolerance of the ear and its importance as a main contributor to grain weight and, more broadly, grain yield under disadvantageous environmental conditions.
Wheat endosperm and pericarp
Caley et al. (1990), followed by Tambussi et al. (2005), also proposed a possible role of the green pericarp in CO2 refixation. Although stomata are almost absent in the growing endosperm, suggesting limited gas‐exchange capacity, immunocytochemical analysis showed chloroplasts and Rubisco co‐localization in the green pericarp with elevated photosynthetic capacity (Kong et al., 2016), which can account for up to 42% of the total photosynthetic activity of the ear (Evans and Rawson, 1970). Recent work reported that genes specific for the C4 pathways such as PEPC, NAD‐ME and NADP‐MDH are expressed in the cross and tube‐cell layer of the pericarp (Rangan et al., 2016), agreeing with earlier studies that had already suggested the presence of C4 or C3–C4 intermediate metabolism in the ear (Ziegler‐Jöns, 1989; Imaizumi et al., 1990; Li et al., 2004; Jia et al., 2015), potentially induced under water‐stress conditions. On the other hand, the following observations suggest limited evidence for a C4 pathway in the green pericarp and other ear organs: (i) oxygen sensitivity of CO2 assimilation rate of the ear (increased by up to 45% under conditions of 2% O2; Tambussi et al., 2005; Tambussi et al., 2007); (ii) high rates of CO2 assimilation through the CBC rather than conversion into C4 malate or aspartate (Bort et al., 1995); and (iii) a lack of the specific C4 anatomy (Tambussi et al., 2005), although future analyses are required to confirm this and it remains a topic of debate.
The importance of stomata for ear photosynthesis
Several studies have demonstrated that the stomatal density in the flag leaf of wheat varies between 40 and 90 mm−2 (e.g. Faralli et al., 2019a), and that in ear organs the stomatal density can be either higher (Kong et al., 2015) or drastically lower (Tambussi et al., 2005) than in the leaf. Furthermore, different stomatal densities and distributions have been reported on both the ventral and the dorsal sides of the glume and the lemma (Figure 3), with the lemma showing variable density depending on the shading area of the neighbouring glume (Tambussi et al., 2005). As the growing endosperm releases respired CO2, the presence of stomata in the internal surface of glumes and lemmas is evidence of CO2 recycling capacity. As reported for fruit (see above), several studies have demonstrated large rates of refixation of respiratory CO2 in the ear (Bort et al., 1996), which can contribute up to 79% of the sucrose accumulated in bracts (Gebbing and Schnyder, 2001). The refixation capacity has several advantages, in particular: (i) respiratory CO2 losses are minimized; and (ii) photosynthetic metabolism is fully independent of the environment.
Genotypic variation in stomatal distribution in glumes and lemmas, and on the different sides, also exists in current elite bread wheat cultivars (Figures 3 and 4), which suggests different strategies for atmospheric CO2 assimilation or CO2 refixation that could be further exploited for ear gas‐exchange optimization. In general, high stomatal densities are reported on the external side of glumes (up to 32 mm−2) and awns (up to 70 mm−2), with lower numbers found in lemmas (between 20 and 10 mm−2) and absent in paleas (Figure 4). The stomatal density on the internal surfaces are comparable for glumes and lemmas (between 20 and 9 mm−2), but are almost absent in paleas. It has been reported that stomatal functionality may be strongly limited in the ear by: (i) the mechanical constraint induced by the growing grains inside the florets; and (ii) by the accumulation of waxes preventing guard cells opening and closing (Araus et al., 1993) and hence limiting photosynthetic CO2 uptake, especially during the late grain filling stage. Figure 5 shows thermal images from the ear and flag leaf of two wheat varieties, and reveals that although the temperature regulation of the ear is significantly lower than that of the flag leaf (i.e. lower transpiration rate), the ear stomata are responsive and open when subjected to a transition from low to high light conditions. In the ear the two cultivars also differ in the magnitude and rapidity of stomatal opening (Faralli et al., 2019b), suggesting potential genotypic variation, driven by either differences in wax accumulation (Araus et al., 1993) or variation in stomatal size, density and distribution, as well as functional differences. Indeed, in glasshouse experiments, six recombinant inbred lines grown under conditions of heat and water stress showed the presence of cooling capacity in the ear at early anthesis (i.e. before pollen release) (Steinmeyer et al., 2013). With the elevated sensitivity of pollen to high temperatures, ear stomatal dynamics and the overall evaporative cooling capacity may be important novel traits for increasing stress tolerance by protecting pollen viability and minimizing floret damage at anthesis. Indeed, at the reproductive stage, stress tolerance in crops is based on both the ability to produce viable pollen and to ‘shield’ the pollen from environmental stresses (i.e. reducing the temperature of reproductive organs with a high transpiration rate) (Steinmeyer et al., 2013). In addition, enhancing stomatal regulation and transpiration may increase assimilate translocation to the developing grains and the remobilization of resources, and could be considered as an additional target for increasing yield potential.
Figure 4.
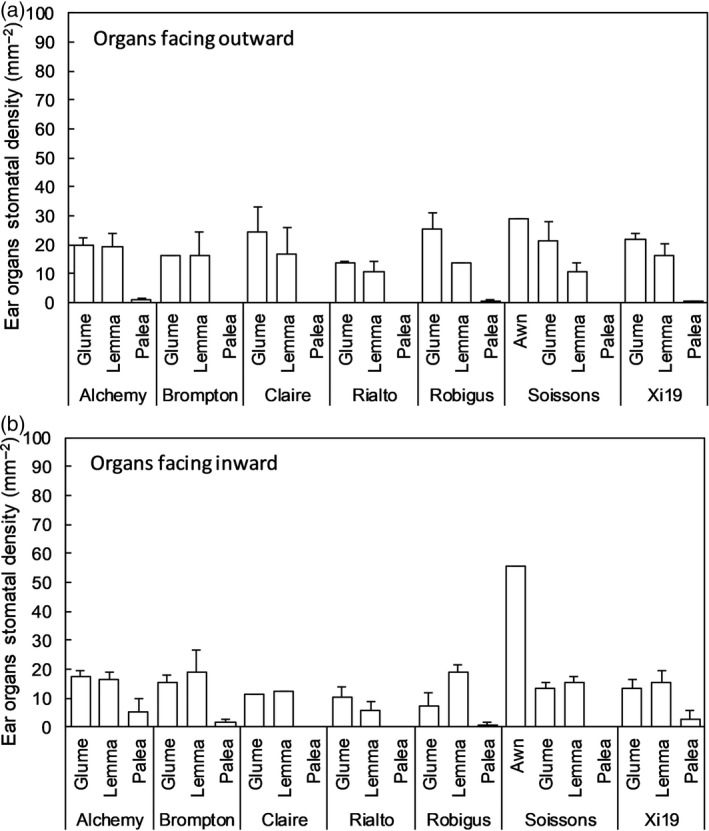
Stomatal density of ear organs (glume, lemma, palea and awns, when present) for seven bread wheat elite cultivars collected after anthesis (a and b). Wheat plants were grown in a glasshouse and ears were harvested at the end of anthesis (i.e. GS69). Stomatal analysis was carried out as described by Faralli et al. 2019b. Data are means ± standard errors of the mean (n = 2–7).
Figure 5.
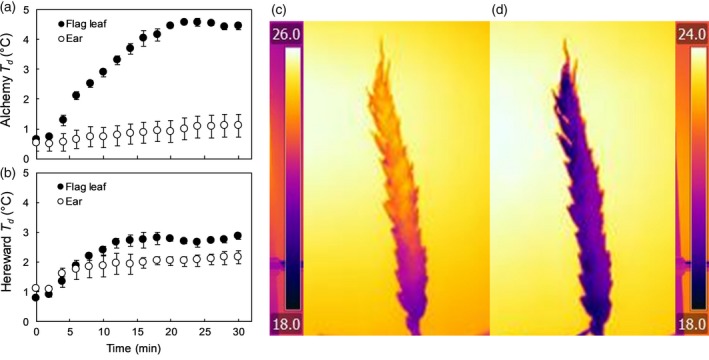
Temperature differential between the dry reference and either the flag leaf or the ear of two bread wheat varieties grown in glasshouse conditions (n = 4 cv. Alchemy and Hereward) subjected to a step change in light (from 100 to 1000 µmol m−2 s−1) and maintained under high light conditions for 30 min. Thermal images of a wheat ears (c, d) following the step increase in light intensity shows significant temperature differences in plants subjected to 10 min (c) or 25 min (d) of illumination, illustrating stomatal functioning in increased evaporative cooling.
Challenges associated with measuring photosynthesis in non‐foliar tissue
Further experimental evidence is needed to fully understand the mechanisms involved in photosynthetic activity of the ear and other non‐foliar photosynthetic organs. There are challenges associated with measuring photosynthesis in non‐laminar tissues using the standard approaches used for leaves. For example, most leaf‐level measurements of CO2 uptake are conducted using infrared gas analysis (IRGA), which requires the material to be enclosed in a sealed chamber, with the differences in gas fluxes in and out of the chamber being assessed. Using such approaches for non‐leaf material represents challenges, including: (i) the small size of commercially available leaf gas‐exchange chambers; (ii) the complication of refixation of respiratory CO2 in determining gas differentials; and (iii) the complexity of ear architecture in wheat, making the normalization of gas‐exchange data per unit area particularly difficult and leading to strong uncertainty in the absolute values. New methodologies are needed and should be implemented to assess ear gas exchange and organ contribution to grain weight. For instance, 3D scanners help to refine the estimation of area, in particular in view of the consistent underestimation (and thus gas‐exchange overestimation) that occurs with standard techniques (e.g. using a ruler; Figure 6). Additionally, the design and development of bespoke chambers is required to enclose an entire ear or fruit to allow the assessment of whole‐organ gas exchange. Such chambers present further challenges that arise from the large volumes required, which can lead to slow gas mixing and difficult temperature control. In addition, although saturated light can be provided in large cuvettes for all surfaces, the shading effects from neighbouring organs, e.g. spikelet morphology and distance between spikelets, may lead to additional sources of error. Chlorophyll fluorescence has been shown to be a good candidate for ear photosynthetic assessment (Tambussi et al., 2005; Maydup et al., 2012), and combined with gas exchange (McAusland et al., 2013) may help to dissect the proportion of photosynthesis relying on the refixation of respiratory CO2 from atmospheric CO2, as well as determining differences in the O2 sensitivity of various genotypes.
Figure 6.
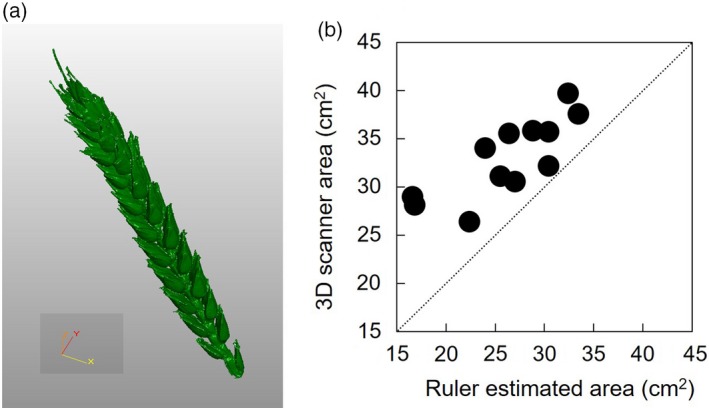
(a) Example of a detailed assessment of wheat ear area and volume using a 3D scanner approach. (b) Example of the underestimation of ear area using a ruler‐based approach compared with a detailed 3D scanner estimation. Wheat plants (cv. Cadenza) were grown in a glasshouse and primary and secondary ears were harvested at different times and over three periods after anthesis. The area was estimated with a ruler by measuring ear length and width of all the four surfaces and then the same ear was assessed with a 3D scanner (n = 4 for each harvest).
Defoliation, inhibition of photosynthesis through shading and herbicide application are some of the most commonly used approaches to evaluate the contribution of ear photosynthesis to yield (Sanchez‐Bragado et al., 2016). Although these approaches may be useful to evaluate genotypic variation, they are likely to induce compensatory mechanisms (and potentially overestimations). Sanchez‐Bragado et al. (2016) suggested carbon isotope discrimination as an alternative for assessing ear photosynthetic traits. In addition, owing to the Rubisco discrimination of 13C and because of the lack of carbon discrimination in PEPC, the isotopic signature may help to discern potential variation between the C3 and C4 pathways (Hu et al., 2019). It must be recognized that almost all the approaches outlined above lack the advantage of high throughput and are generally considered time consuming and laborious, and this therefore limits their use for screening large populations or samples for ear photosynthetic phenotypes. There is no doubt that improvement in experimental procedures along with further advances in high‐throughput approaches for screening ear photosynthesis will increase our understanding of ear photosynthetic activity and therefore help to design new cereal varieties with elevated yield potential and stability.
Conclusion
Although most studies examining photosynthesis have focused on leaf‐level measurements, including current approaches to improve photosynthesis, the contribution that other green tissues make to total photoassimilates has largely been ignored. As highlighted above, these green tissues contribute significantly to plant development, growth and yield, and therefore present novel opportunities for exploitation to improve productivity. The fact that the full spectrum of light harvest, electron transport and CBC proteins and transcripts are found in non‐foliar tissues (Barsan et al., 2010; Barsan et al., 2012; Sui et al., 2017; Vicente et al., 2018) offers the potential to manipulate non‐foliar photosynthetic pathways to increase rates of photosynthesis using similar approaches to those currently being employed in leaves (for a review, see Simkin, 2019 and Simkin et al., 2019). For example, recent experiments in transgenic wheat with increased activity of the CBC enzyme SBPase, driven by a constitutive promotor (Driever et al., 2017), revealed increased gross photosynthesis in the ears of mutant plants relative to the wild‐type control (Figure 7). It is therefore possible that the overall increase in yield of plants overexpressing SBpase reported by Driever et al. (2017) may have been achieved in part by an increase in ear‐derived assimilates, although this would require further investigation. Such studies highlight the potential benefits of improving photosynthesis in organs other than leaves for improving crop productivity and yield. Furthermore, as photosynthesis provides the building blocks for many downstream products and metabolites, modifying photosynthetic processes in fruits, for example, offers the potential to alter fruit quality and nutritional value.
Figure 7.

Gross assimilation rate calculated as the sum of light‐saturated assimilation rate and dark respiration of wheat ears (n = 5) of control cv. Cadenza plants and transgenic plants overexpressing SBPase (Driever et al., 2017). Data were collected post‐anthesis in glasshouse‐grown plants with a Licor 6400XT mounted with a bespoke cuvette ensuring saturating light (1000 μmol m−2 sec−1) and a 25°C block temperature.
A major difference between leaf and non‐leaf tissues is the primary source of CO2 for CBC (atmospheric versus respiratory), and therefore the manipulation of stomatal density or function presents an additional avenue to manipulate photosynthetic processes in some tissues, e.g. wheat ears. For example, increasing stomatal density or aperture could result in increasing assimilation by removing diffusional constraints and increasing the flux of atmospheric CO2 to the site of carboxylation; however, such an approach would also facilitate the leakage of respiratory CO2 (Sui et al., 2017), which has been demonstrated to be of greater importance in some organs. Alternatively, increased stomatal density in wheat ears could improve evaporative cooling, thereby maintaining assimilation rates under elevated temperatures, assuming a similar temperature sensitivity of photosynthesis in wheat ears and leaves (Scafaro et al., 2012; Scafaro et al., 2016; Perdomo et al., 2017). On the other hand, this ‘risky’ behaviour might increase the possibility of early ear dehydration under severe terminal stress conditions, although further experimental evidence is required to support this theory. Stomatal behaviour and transpiration in ears may also provide a key role in the translocation of photoassimilates to the ear, and therefore altering gs could assist with sink–source relationships. Although stomatal behaviour is important for photosynthesis, it should be acknowledged that stomatal pores are also an important component of non‐leaf tissues to facilitate drying, which is essential for the dispersal of spores and seeds (e.g. stomata in the spore capsules of moss; Merced and Renzaglia, 2013; Chater et al., 2016). Before such novel targets for improved photosynthesis can be exploited, a better understanding of the contribution of non‐foliar photosynthesis to yield and quality (particularly under conditions of stress) and the role of stomata in these processes is needed.
Conflict of interest
There are no conflicts of interest to declare.
Author contributions
AJS, MF and TL all wrote the article and contributed to the figures. Data presented on the SBPase wheat are from work carried out by MF, AJS, TL and Christine Raines.
Acknowledgements
We acknowledge Christine Raines for providing the wheat SBPase overexpression lines. Joe Chadwick, Alexandra Milliken and Tanja Hofmann are acknowledged for assistance with assembling the figures. M.F. was supported by BBSRC (grant no. BB/N016831/1), awarded to T.L.
Data availability statement
Data presented within this review are example data sets that are not publicly available; please contact T.L. to request access to any data.
References
- Abebe, T. , Melmaiee, K. , Berg, V. and Wise, R.P. (2010) Drought response in the spikes of barley: gene expression in the lemma, palea, awn, and seed. Funct. Integr. Genomics, 10, 191–205. [DOI] [PubMed] [Google Scholar]
- Allen, D.K. , Ohlrogge, J.B. and Shachar‐Hill, Y. (2009) The role of light in soybean seed filling metabolism. Plant J. 58, 220–234. [DOI] [PubMed] [Google Scholar]
- Allorent, G. , Osorio, S. , Ly, Vu et al . (2015) Adjustments of embryonic photosynthetic activity modulate seed fitness in Arabidopsis thaliana . New Phytol. 205, 707–719. [DOI] [PubMed] [Google Scholar]
- Aoki, K. , Yamamoto, M. and Wada, K. (1998) Photosynthetic and heterotrophic ferredoxin isoproteins are colocalized in fruit plastids of tomato. Plant Physiol. 118, 439–449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aoyagi, K. and Bassham, J.A. (1984) Pyruvate Orthophosphate Dikinase of C3 seeds and leaves as compared to the enzyme from maize. Plant Physiol. 75, 387–392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Araújo, W.L. , Nunes‐Nesi, A. , Osorio, S. et al . (2011) Antisense inhibition of the iron‐sulphur subunit of succinate dehydrogenase enhances photosynthesis and growth in tomato via an organic acid–mediated effect on stomatal aperture. Plant Cell, 23(2), 600–627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Araus, J.L. , Brown, H.R. , Febrero, A. , Bort, J. and Serret, M.D. (1993) Ear photosynthesis, carbon isotope discrimination and the contribution of respiratory CO2 to differences in grain mass in durum wheat. Plant, Cell Environ. 16, 383–392. [Google Scholar]
- Aschan, G. and Pfanz, H. (2003) Non‐foliar photosynthesis – a strategy of additional carbon acquisition. Flora, 198, 81–97. [Google Scholar]
- Asokanthan, P.S. , Johnson, R.W. , Griffith, M. and Krol, M. (1997) The photosynthetic potential of canola embryos. Physiol Plantarum, 101, 353–360. [Google Scholar]
- Atkins, C.R. , Kuo, J. , Pate, J.S. , Flinn, A.M. and Steel, T.W. (1977) Photosynthetic pod wall of pea (Pisum sativum L.): distribution of carbon dioxide‐fixing enzymes in relation to pod structure. Plant Physiol. 60, 779–786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ávila, E. , Herrera, A. and Tezara, W. (2014) Contribution of stem CO2 fixation to whole‐plant carbon balance in nonsucculent species. Photosynthetica, 52, 3–15. [Google Scholar]
- Avila‐Lovera, E. , Zerpa, A.J. and Santiago, L.S. (2017) Stem photosynthesis and hydraulics are coordinated in desert plant species. New Phytol. 216, 1119–1129. [DOI] [PubMed] [Google Scholar]
- Barsan, C. , Sanchez‐Bel, P. , Rombaldi, C. , Egea, I. , Rossignol, M. , Kuntz, M. , Zouine, M. , Latche, A. , Bouzayen, M. and Pech, J.C. (2010) Characteristics of the tomato chromoplast revealed by proteomic analysis. J. Exp. Bot. 61, 2413–2431. [DOI] [PubMed] [Google Scholar]
- Barsan, C. , Zouine, M. , Maza, E. et al . (2012) Proteomic analysis of chloroplast‐to‐chromoplast transition in tomato reveals metabolic shifts coupled with disrupted thylakoid biogenesis machinery and elevated energy‐production components. Plant Physiol. 160, 708–725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassham, J.A. and Calvin, M. (1960) The path of carbon in photosynthesis In Die CO2‐Assimilation / The Assimilation of Carbon Dioxide: In 2 Teilen/2 Parts (Pirson A. ed). Berlin, Heidelberg: Springer, Berlin Heidelberg, pp. 884–922. [Google Scholar]
- Biel, K. and Fomina, I. (2015) Benson‐Bassham‐Calvin cycle contribution to the organic life on our planet. Photosynthetica, 53, 161–167. [Google Scholar]
- Birkhold, K.T. , Koch, K.E. and Darnell, R.L. (1992) Carbon and nitrogen economy of developing Rabbiteye blueberry fruit. J. Am. Soc. Hortic. Sci. 117, 139–145. [Google Scholar]
- Blanke, M.M. (1992) Photosynthesis of Avocado fruit. Proceedings of 2nd world avocado congress, 179–189.
- Blanke, M.M. (1998) Fruit Photosynthesis. Dordrecht: Springer. [Google Scholar]
- Blanke, M.M. and Lenz, F. (1989) Fruit photosynthesis. Plant, Cell Environ. 12, 31–46. [Google Scholar]
- Borisjuk, L. , Nguyen, T.H. , Neuberger, T. et al . (2005) Gradients of lipid storage, photosynthesis and plastid differentiation in developing soybean seeds. New Phytol. 167, 761–776. [DOI] [PubMed] [Google Scholar]
- Bort, J. , Febrero, A. , Amaro, T. and Araus, J.L. (1994) Role of awns in ear water‐use efficiency and grain weight in barley. Agronomie, 14, 133–139. [Google Scholar]
- Bort, J. , Brown, R.H. and Araus, J.L. (1995) Lack of C4 photosynthetic metabolism in ears of C3 cereals. Plant, Cell Environ. 18, 697–702. [Google Scholar]
- Bort, J. , Brown, R.H. and Araus, J.L. (1996) Refixation of respiratory CO2 in the ears of C3 cereals. J. Exp. Bot. 47, 1567–1575. [Google Scholar]
- Bravdo, B.‐A. , Palgi, A. , Lurie, S. and Frenkel, C. (1977) Changing ribulose diphosphate carboxylase/oxygenase activity in ripening tomato fruit. Plant Physiol. 60, 309–312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calderini, D.F. , Dreccer, M.F. and Slafer, G.A. (1995) Genetic improvement in wheat yield and associated traits. A re‐examination of previous results and the latest trends. Plant Breeding, 114, 108–112. [Google Scholar]
- Caley, C.Y. , Duffus, C.M. and Jeffcoat, B. (1990) Photosynthesis in the Pericarp of Developing Wheat Grains. J. Exp. Bot. 41, 303–307. [Google Scholar]
- Calvin, M. and Benson, A.A. (1948) The path of carbon in photosynthesis. Science, 107, 476–480. [DOI] [PubMed] [Google Scholar]
- Cannell, M.G.R. (1985) Physiology of the Coffee Crop. Boston, MA: Springer. [Google Scholar]
- Carrara, S. , Pardossi, A. , Soldatini, G.F. , Tognoni, F. and Guidi, L. (2001) Photosynthetic activity of ripening tomato fruit. Photosynthetica, 39, 75–78. [Google Scholar]
- Cernusak, L.A. and Hutley, L.B. (2011) Stable Isotopes reveal the contribution of corticular photosynthesis to growth in branches of Eucalyptus miniata . Plant Physiol. 155, 515–523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chater, C.C. , Caine, R.S. , Tomek, M. et al . (2016) Origin and function of stomata in the moss Physcomitrella patens. Nat Plants, 2, 16179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheung, A.Y. , McNellis, T. and Piekos, B. (1993) Maintenance of chloroplast components during chromoplast differentiation in the tomato mutant green flesh. Plant Physiol. 101, 1223–1229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chida, H. , Nakazawa, A. , Akazaki, H. et al . (2007) Expression of the algal cytochrome c 6 gene in Arabidopsis enhances photosynthesis and growth. Plant Cell Physiol. 48, 948–957. [DOI] [PubMed] [Google Scholar]
- Cocaliadis, M.F. , Fernández‐Muñoz, R. , Pons, C. , Orzaez, D. and Granell, A. (2014) Increasing tomato fruit quality by enhancing fruit chloroplast function. A double‐edged sword? J. Exp. Bot. 65, 4589–4598. [DOI] [PubMed] [Google Scholar]
- Driever, S.M. , Simkin, A.J. , Alotaibi, S. et al . (2017) Increased SBPase activity improves photosynthesis and grain yield in wheat grown in greenhouse conditions. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 372, 1730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eastmond, P. , Koláčná, L. and Rawsthorne, S. (1996) Photosynthesis by developing embryos of oilseed rape (Brassica napus L.). J. Exp. Bot. 47, 1763–1769. [Google Scholar]
- Ehleringer, J.R. , Comstock, J.P. and Cooper, T.A. (1987) Leaf‐twig carbon isotope ratio differences in photosynthetic‐twig desert shrubs. Oecologia, 71, 318–320. [DOI] [PubMed] [Google Scholar]
- Ermakova, M. , Lopez‐Calcagno, P.E. , Raines, C.A. , Furbank, R.T. and von Caemmerer, S. (2019) Overexpression of the Rieske FeS protein of the Cytochrome b6f complex increases C4 photosynthesis in Setaria viridis . Commun. Biol. 2, 1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans, T.L. and Fischer, R.A. (1999) Yield potential: its definition, measurement, and significance. Crop Sci. 39, 1544–1551. [Google Scholar]
- Evans, L.T. and Rawson, H.M. (1970) Photosynthesis and respiration by the flag leaf and components of the ear during grain development in wheat. Aust. J. Biol. Sci. 23, 245–254. [Google Scholar]
- FAO (2017) The future of food and agriculture – trends and challenges. Rome: http://www.fao.org/publications/fofa/en/ [Google Scholar]
- Faralli, M. , Cockram, J. , Ober, E. , Wall, S. , Galle, A. , Van Rie, J. , Raines, C. and Lawson, T. (2019a) Genotypic, developmental and environmental effects on the rapidity of gs in Wheat: impacts on carbon gain and water‐use efficiency. Front. Plant Sci. 10, 492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faralli, M. , Matthews, J. and Lawson, T. (2019b) Exploiting natural variation and genetic manipulation of stomatal conductance for crop improvement. Curr. Opin. Plant Biol. 49, 1–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferroni, L. , Pantaleoni, L. , Baldisserotto, C. , Aro, E. and Pancaldi, S. (2013) Low photosynthetic activity is linked to changes in the organization of photosystem II in the fruit of Arum italicum . Plant Physiol. Biochem. 63, 140–150. [DOI] [PubMed] [Google Scholar]
- Fischer, R.A. , Rees, D. , Sayre, K.D. , Lu, Z.M. , Condon, A.G. and Saavedra, A.L. (1998) Wheat yield progress associated with higher stomatal conductance and photosynthetic rate, and cooler canopies. Crop Sci. Soc. Am. 38, 1467–1475. [Google Scholar]
- Gale, M.D. and Youssefian, S. (1985) Dwarfing Genes in Wheat. London: Butterworth and Co. [Google Scholar]
- Galili, G. , Avin‐Wittenberg, T. , Angelovici, R. and Fernie, A.R. (2014) The role of photosynthesis and amino acid metabolism in the energy status during seed development. Front. Plant Sci. 5, 447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gebbing, T. and Schnyder, H. (2001) 13C labeling kinetics of sucrose in glumes indicates significant refixation of respiratory CO2 in the wheat ear. Funct. Plant Biol. 28, 1047–1053. [Google Scholar]
- Glowacka, K. , Kromdijk, J. , Kucera, K. , Xie, J. , Cavanagh, A.P. , Leonelli, L. , Leakey, A.D.B. , Ort, D.R. , Niyogi, K.K. and Long, S.P. (2018) Photosystem II subunit S overexpression increases the efficiency of water use in a field‐grown crop. Nat. Commun. 9, 868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golombek, S. , Heim, U. , Horstmann, C. , Wobus, U. and Weber, H. (1999) Phosphoenolpyruvate carboxylase in developing seeds of Vicia faba L.: gene expression and metabolic regulation. Planta, 208, 66–72. [DOI] [PubMed] [Google Scholar]
- Gu, J. , Yin, X. , Stomph, T.J. and Struik, P.C. (2014) Can exploiting natural genetic variation in leaf photosynthesis contribute to increasing rice productivity? Plant Cell Environ. 37, 22–34. [DOI] [PubMed] [Google Scholar]
- Guo, Z. and Schnurbusch, T. (2016) Costs and benefits of awns. J. Exp. Bot. 67, 2533–2535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris, W.M. and Spurr, A.R. (1969a) Chromoplasts of tomato fruits: I. Ultrastructure of low‐pigment and high‐beta mutants: carotene analyses. Am. J. Bot. 56, 369–379. [Google Scholar]
- Harris, W.M. and Spurr, A.R. (1969b) Chromoplasts of tomato fruits: II. The red tomato. Am. J. Bot. 56, 380–389. [Google Scholar]
- Hedley, C.L. , Harvey, D.M. and Keely, R.J. (1975) Role of PEP carboxylase during seed development in Pisum sativum . Nature, 258, 352–354. [Google Scholar]
- Hein, J.A. , Sherrard, M.E. , Manfredi, K.P. and Abebe, T. (2016) The fifth leaf and spike organs of barley (Hordeum vulgare L.) display different physiological and metabolic responses to drought stress. BMC Plant Biol. 16, 248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hetherington, A.M. and Woodward, F.I. (2003) The role of stomata in sensing and driving environmental change. Nature, 424, 901–908. [DOI] [PubMed] [Google Scholar]
- Hetherington, S.E. , Smillie, R.M. and Davies, W.J. (1998) Photosynthetic activities of vegetative and fruiting tissues of tomato. J. Exp. Bot. 49, 1173–1181. [Google Scholar]
- Hibberd, J.M. and Quick, W.P. (2002) Characteristics of C4 photosynthesis in stems and petioles of C3 flowering plants. Nature, 415, 451–454. [DOI] [PubMed] [Google Scholar]
- Hieke, S. , Menzel, C.M. and Ludders, P. (2002) Effects of leaf, shoot and fruit development on photosynthesis of lychee trees (Litchi chinensis). Tree Physiol. 22, 955–961. [DOI] [PubMed] [Google Scholar]
- Hills, M.J. (2004) Control of storage‐product synthesis in seeds. Curr. Opin. Plant. Biol. 7, 302–308. [DOI] [PubMed] [Google Scholar]
- Hiratsuka, S. , Suzuki, M. , Nishimura, H. and Nada, K. (2015) Fruit photosynthesis in Satsuma mandarin . Plant Sci. 241, 65–69. [DOI] [PubMed] [Google Scholar]
- Hu, Y.‐Y. , Zhang, Y.‐L. , Luo, H.‐H. , Li, W. , Oguchi, R. , Fan, D.‐Y. , Chow, W.S. and Zhang, W.‐F. (2012) Important photosynthetic contribution from the non‐foliar green organs in cotton at the late growth stage. Planta, 235, 325–336. [DOI] [PubMed] [Google Scholar]
- Hu, Y.‐Y. , Zhang, Y.‐L. , Xia, H. , Fan, S. , Song, J. , Lv, X. and Kong, L. (2019) Photosynthetic characteristics of non‐foliar organs in main C3 cereals. Physiol Plant. 166, 226–239. [DOI] [PubMed] [Google Scholar]
- Imaizumi, N. , Usuda, H. , Nakamoto, H. and Ishihara, K. (1990) Changes in the rate of photosynthesis during grain filling and the enzymatic activities associated with the photosynthetic carbon metabolism in rice panicles. Plant Cell Physiol. 31, 835–844. [Google Scholar]
- Jia, S. , Lv, J. , Jiang, S. , Liang, T. , Liu, C. and Jing, Z. (2015) Response of wheat ear photosynthesis and photosynthate carbon distribution to water deficit. Photosynthetica, 53(1), 95–109. [Google Scholar]
- Kang, F. and Rawsthorne, S. (1996) Metabolism of glucose‐6‐phosphate and utilization of multiple metabolites for fatty acid synthesis by plastids from developing oilseed rape embryos. Planta, 199, 321–327. [Google Scholar]
- Kohzuma, K. , Sato, Y. , Ito, H. et al . (2017) The non‐mendelian green cotyledon gene in Soybean encodes a small subunit of photosystem II. Plant Physiol. 173, 2138–2147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kong, L. , Sun, M. , Xie, Y. , Wang, F. and Zhao, Z. (2015) Photochemical and antioxidative responses of the glume and flag leaf to seasonal senescence in wheat. Front. Plant Sci. 6, 358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kong, L.A. , Xie, Y. , Sun, M.Z. , Si, J.S. and Hu, L. (2016) Comparison of the photosynthetic characteristics in the pericarp and flag leaves during wheat (Triticum aestivum L.) caryopsis development. Photosynthetica, 54, 40–46. [Google Scholar]
- Kromdijk, J. , Glowacka, K. , Leonelli, L. , Gabilly, S.T. , Iwai, M. , Niyogi, K.K. and Long, S.P. (2016) Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science, 354, 857–861. [DOI] [PubMed] [Google Scholar]
- Laval‐Martin, D. , Farineau, J. and Diamond, J. (1977) Light versus dark carbon metabolism in cherry tomato fruits: I. occurrence of photosynthesis. Study of the Intermediates. Plant Physiol. 60, 872–876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lefebvre, S. , Lawson, T. , Fryer, M. , Zakhleniuk, O.V. , Lloyd, J.C. and Raines, C.A. (2005) Increased sedoheptulose‐1,7‐bisphosphatase activity in transgenic tobacco plants stimulates photosynthesis and growth from an early stage in development. Plant Physiol. 138, 451–460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lemaire‐Chamley, M. , Petit, J. , Garcia, V. , Just, D. , Baldet, P. , Germain, V. , Fagard, M. , Mouassite, M. , Cheniclet, C. and Rothan, C. (2005) Changes in transcriptional profiles are associated with early fruit tissue specialization in tomato. Plant Physiol. 139, 750–769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, X. , Hou, J. , Bai, K. , Yang, X. , Lin, J. , Li, Z. and Kuang, T. (2004) Activity and distribution of carbonic anhydrase in leaf and ear parts of wheat (Triticum aestivum L.). Plant Sci. 166, 627–632. [Google Scholar]
- Li, Y. , Li, H. , Li, Y. and Zhang, S. (2017) Improving water‐use efficiency by decreasing stomatal conductance and transpiration rate to maintain higher ear photosynthetic rate in drought‐resistant wheat. Crop J. 5, 231–239. [Google Scholar]
- Livne, A. and Gepstein, S. (1988) Abundance of the major chloroplast polypeptides during development and ripening of tomato fruits. Plant Physiol. 87, 239–243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez, Y. , Riaño, N. , Mosquera, P. , Cadavid, A. and Arcila, J. (2000) Activities of phosphoenolpyruvate carboxylase and ribulose‐1,5‐bisphosphate carboxylase/oxygenase in leaves and fruit pericarp tissue of different coffee (Coffea sp.) genotypes. Photosynthetica, 38, 215–220. [Google Scholar]
- López‐Calcagno, P.E. , Fisk, S.J. , Brown, K. , Bull, S.E. , South, P.F. and Raines, C.A. (2018) Overexpressing the H‐protein of the glycine cleavage system increases biomass yield in glasshouse and field‐grown transgenic tobacco plants. Plant Biotechnol. J. 17, 141–151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lytovchenko, A. , Eickmeier, I. , Pons, C. et al . (2011) Tomato fruit photosynthesis is seemingly unimportant in primary metabolism and ripening but plays a considerable role in seed development. Plant Physiol. 157, 1650–1663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manetas, Y. (2004) Probing corticular photosynthesis through in vivo chlorophyll fluorescence measurements: evidence that high internal CO levels suppress electron flow and increase the risk of photoinhibition. Physiol. Plant, 120, 509–517. [DOI] [PubMed] [Google Scholar]
- Manzara, T. , Carrasco, P. and Gruissem, W. (1993) Developmental and organ‐specific changes in DNA‐protein interactions in the tomato rbcS1, rbcS2 and rbcS3A promoter regions. Plant Mol. Biol. 21, 69–88. [DOI] [PubMed] [Google Scholar]
- Maydup, M.L. , Antonietta, M. , Guiamet, J.J. , Graciano, C. , Lopez, J.R. and Tambussi, E.A. (2010) The contribution of ear photosynthesis to grain filling in bread wheat. Field. Crop. Res. 119, 48–58. [Google Scholar]
- Maydup, M.L. , Antonietta, M. , Guiamet, J.J. and Tambussi, E.A. (2012) The contribution of green parts of the ear to grain filling in old and modern cultivars of bread wheat (Triticum aestivum L.): evidence for genetic gains over the past century. Field. Crop. Res. 134, 208–215. [Google Scholar]
- Maydup, M.L. , Antonietta, M. , Graciano, C. , Guiamet, J.J. and Tambussi, E.A. (2014) The contribution of the awns of bread wheat (Triticum aestivum L.) to grain filling: responses to water deficit and the effects of awns on ear temperature and hydraulic conductance. Field. Crop. Res. 167, 102–111. [Google Scholar]
- McAusland, L. , Davey, P.A. , Kanwal, N. , Baker, N.R. and Lawson, T. (2013) A novel system for spatial and temporal imaging of intrinsic plant water use efficiency. J. Exp. Bot. 64, 4993–5007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Merced, A. and Renzaglia, K.S. (2013) Moss stomata in highly elaborated Oedipodium (Oedipodiaceae) and highly reduced Ephemerum (Pottiaceae) sporophytes are remarkably similar. Am. J. Bot. 100, 2318–2327. [DOI] [PubMed] [Google Scholar]
- Millar, A.H. , Whelan, J. , Soole, K.L. and Day, D.A. (2011) Organization and regulation of mitochondrial respiration in plants. Annu. Rev. Plant Biol. 62, 79–104. [DOI] [PubMed] [Google Scholar]
- Moreshet, S. and Green, G.C. (1980) Photosynthesis and diffusion conductance of the valencia orange fruit under field conditions. J. Exp. Bot. 31, 15–27. [Google Scholar]
- Nunes‐Nesi, A. , Carrari, F. , Lytovchenko, A. , Smith, A.M.O. , Ehlers Loureiro, M. , Ratcliffe, R.G. , Sweetlove, L.J. and Fernie, A.R. (2005) Enhanced photosynthetic performance and growth as a consequence of decreasing mitochondrial malate dehydrogenase activity in transgenic tomato plants. Plant Physiol. 137, 611–622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Obiadalla‐Ali, H. , Fernie, A.R. , Lytovchenko, A. , Kossmann, J. and Lloyd, J.R. (2004) Inhibition of chloroplastic fructose 1,6‐bisphosphatase in tomato fruits leads to decreased fruit size, but only small changes in carbohydrate metabolism. Planta, 219, 533–540. [DOI] [PubMed] [Google Scholar]
- Osmond, C.B. , Smith, S.D. , Gui‐Ying, B. and Sharkey, T.D. (1987) Stem photosynthesis in a desert ephemeral, Eriogonum inflatum . Oecologia, 72, 542–549. [DOI] [PubMed] [Google Scholar]
- Palanisamy, K. and Vivekanandan, M. (1986) Photosynthetic Functions and induction of etiolation in chloroembryos of Dolichos lablab L. J. Plant Physiol. 123, 395–399. [Google Scholar]
- Parry, M.A. , Reynolds, M. , Salvucci, M.E. , Raines, C. , Andralojc, P.J. , Zhu, X.G. , Price, G.D. , Condon, A.G. and Furbank, R.T. (2011) Raising yield potential of wheat. II. Increasing photosynthetic capacity and efficiency. J. Exp. Bot. 62, 453–467. [DOI] [PubMed] [Google Scholar]
- Pengelly, J.J.L. , Kwasny, S. , Bala, S. , Evans, J.R. , Voznesenskaya, E.V. , Koteyeva, N.K. , Edwards, G.E. , Furbank, R.T. and von Caemmerer, S. (2011) Functional analysis of corn husk photosynthesis. Plant Physiol. 156, 503–513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perdomo, J.A. , Capo‐Bauca, S. , Carmo‐Silva, E. and Galmes, J. (2017) Rubisco and rubisco activase play an important role in the biochemical limitations of photosynthesis in rice, wheat, and maize under high temperature and water deficit. Front. Plant Sci. 8, 490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfanz, H. , Aschan, G. , Langenfeld‐Heyser, R. , Wittmann, C. and Loose, M. (2002) Ecology and ecophysiology of tree stems: corticular and wood photosynthesis. Naturwissenschaften, 89, 147–162. [DOI] [PubMed] [Google Scholar]
- Piechulla, B. and Gruissem, W. (1987) Diurnal mRNA fluctuations of nuclear and plastid genes in developing tomato fruits. EMBO J. 6, 3593–3599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piechulla, B. , Pichersky, E. , Cashmore, A.R. and Gruissem, W. (1986) Expression of nuclear and plastid genes for photosynthesis‐specific proteins during tomato fruit development and ripening. Plant Mol. Biol. 7, 367–376. [DOI] [PubMed] [Google Scholar]
- Piechulla, B. , Glick, R.E. , Bahl, H. , Melis, A. and Gruissem, W. (1987) Changes in photosynthetic capacity and photosynthetic protein pattern during tomato fruit ripening. Plant Physiol. 84, 911–917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Puthur, J.T. , Shackira, A.M. , Saradhi, P.P. and Bartels, D. (2013) Chloroembryos: a unique photosynthesis system. J. Plant Physiol. 170, 1131–1138. [DOI] [PubMed] [Google Scholar]
- Raines, C.A. (2003) The Calvin cycle revisited. Photosynth. Res. 75, 1–10. [DOI] [PubMed] [Google Scholar]
- Rangan, P. , Furtado, A. and Henry, R.J. (2016) New evidence for grain specific C4 photosynthesis in wheat. Sci. Rep. 6, 31721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ranjan, S. , Singh, R. , Soni, D.K. , Pathre, U.V. and Shirke, P.A. (2012) Photosynthetic performance of Jatropha curcas fruits. Plant Physiol. Biochem. 52, 66–76. [DOI] [PubMed] [Google Scholar]
- Ray, D.K. , Ramankutty, N. , Mueller, N.D. , West, P.C. and Foley, J.A. (2012) Recent patterns of crop yield growth and stagnation. Nat. Commun. 3, 1293. [DOI] [PubMed] [Google Scholar]
- Ray, D.K. , Mueller, N.D. , West, P.C. and Foley, J.A. (2013) Yield trends are insufficient to double global crop production by 2050. PLoS ONE, 8, e66428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rebetzke, G.J. , Bonnett, D.G. and Reynolds, M.P. (2016) Awns reduce grain number to increase grain size and harvestable yield in irrigated and rainfed spring wheat. J. Exp. Bot. 67, 2573–2586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reynolds, M. , Foulkes, M.J. , Slafer, G.A. , Berry, P. , Parry, M.A.J. , Snape, J.W. and Angus, W.J. (2009) Raising yield potential in wheat. J. Exp. Bot. 60, 1899–1918. [DOI] [PubMed] [Google Scholar]
- RSOL (2009) Royal Society of London, Reaping the Benefits: Science and the Sustainable Intensification of Global Agriculture. London: Royal Society. [Google Scholar]
- Ruuska, S.A. , Schwender, J. and Ohlrogge, J.B. (2004) The capacity of green oilseeds to utilize photosynthesis to drive biosynthetic processes. Plant Physiol. 136, 2700–2709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sánchez, C. , Fischer, G. and Sanjuanelo, D.W. (2013) Stomatal behavior in fruits and leaves of the purple passion fruit (Passiflora edulis Sims) and fruits and cladodes of the yellow pitaya [Hylocereus megalanthus (K. Schum. ex Vaupel) Ralf Bauer]. Agronomía Colombiana, 31, 38–47. [Google Scholar]
- Sanchez‐Bragado, R. , Molero, G. , Reynolds, M.P. and Araus, J.L. (2014) Relative contribution of shoot and ear photosynthesis to grain filling in wheat under good agronomical conditions assessed by differential organ δ13C. J. Exp. Bot. 65, 5401–5413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanchez‐Bragado, R. , Molero, G. , Reynolds, M.P. and Araus, J.L. (2016) Photosynthetic contribution of the ear to grain filling in wheat: a comparison of different methodologies for evaluation. J. Exp. Bot. 67, 2787–2798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scafaro, A.P. , Yamori, W. , Carmo‐Silva, A.E. , Salvucci, M.E. , von Caemmerer, S. and Atwell, B.J. (2012) Rubisco activity is associated with photosynthetic thermotolerance in a wild rice (Oryza meridionalis). Physiol. Plant. 146, 99–109. [DOI] [PubMed] [Google Scholar]
- Scafaro, A.P. , Galle, A. , Van Rie, J. , Carmo‐Silva, E. , Salvucci, M.E. and Atwell, B.J. (2016) Heat tolerance in a wild Oryza species is attributed to maintenance of Rubisco activation by a thermally stable Rubisco activase ortholog. New Phytol. 211, 899–911. [DOI] [PubMed] [Google Scholar]
- Schwender, J. , Goffman, F. , Ohlrogge, J.B. and Shachar‐Hill, Y. (2004) Rubisco without the Calvin cycle improves the carbon efficiency of developing green seeds. Nature, 432, 779–782. [DOI] [PubMed] [Google Scholar]
- Simbo, D.J. , Van den Bilcke, N. and Samson, R. (2013) Contribution of corticular photosynthesis to bud development in African baobab (Adansonia digitata L.) and Castor bean (Ricinus communis L.) seedlings. Environ. Exp. Bot. 95, 1–5. [Google Scholar]
- Simkin, A.J. (2019) Genetic engineering for global food security: Photosynthesis and biofortification. Plants, 8, 586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simkin, A.J. , Kuntz, M. , Moreau, H. and McCarthy, J. (2010) Carotenoid profiling and the expression of carotenoid biosynthetic genes in developing coffee grain. Plant Physiol. Biochem. 48, 434–442. [DOI] [PubMed] [Google Scholar]
- Simkin, A.J. , McAusland, L. , Headland, L.R. , Lawson, T. and Raines, C.A. (2015) Multigene manipulation of photosynthetic carbon assimilation increases CO2 fixation and biomass yield in tobacco. J. Exp. Bot. 66, 4075–4090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simkin, A.J. , Lopez‐Calcagno, P.E. , Davey, P.A. , Headland, L.R. , Lawson, T. , Timm, S. , Bauwe, H. and Raines, C.A. (2017a) Simultaneous stimulation of sedoheptulose 1,7‐bisphosphatase, fructose 1,6‐bisphophate aldolase and the photorespiratory glycine decarboxylase H‐protein increases CO2 assimilation, vegetative biomass and seed yield in Arabidopsis. Plant Biotechnol. J. 15, 805–816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simkin, A.J. , McAusland, L. , Lawson, T. and Raines, C.A. (2017b) Over‐expression of the RieskeFeS protein increases electron transport rates and biomass yield. Plant Physiol. 175, 134–145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simkin, A.J. , Lopez‐Calcagno, P.E. and Raines, C.A. (2019) Feeding the world: improving photosynthetic efficiency for sustainable crop production. J. Exp. Bot. 70, 1119–1140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith, A.M. , Quinton‐Tulloch, J. and Denyer, K. (1990) Characteristics of plastids responsible for starch synthesis in developing pea embryos. Planta, 180, 517–523. [DOI] [PubMed] [Google Scholar]
- Smolikova, G.N. and Medvedev, S.S. (2016) Photosynthesis in the seeds of chloroembryophytes. Russ. J. Plant Physiol. 63, 1–12. [Google Scholar]
- Smolikova, G. , Dolgikh, E. , Vikhnina, M. , Frolov, A. and Medvedev, S. (2017) Genetic and hormonal regulation of chlorophyll degradation during maturation of seeds with green embryos. Int. J. Mol. Sci. 18, 1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steer, B.T. and Pearson, C.J. (1976) Photosynthate translocation in Capsicum annuum . Planta, 128, 155. [DOI] [PubMed] [Google Scholar]
- Steinhauser, M.‐C. , Steinhauser, D. , Koehl, K. , Carrari, F. , Gibon, Y. , Fernie, A.R. and Stitt, M. (2010) Enzyme activity profiles during fruit development in tomato cultivars and Solanum pennellii . Plant Physiol. 153, 80–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinmeyer, F.T. , Lukac, M. , Reynolds, M.P. and Jones, H.E. (2013) Quantifying the relationship between temperature regulation in the ear and floret development stage in wheat (Triticum aestivum L.) under heat and drought stress. Funct. Plant Biol. 40, 700–707. [DOI] [PubMed] [Google Scholar]
- Sugita, M. and Gruissem, W. (1987) Developmental, organ‐specific, and light‐dependent expression of the tomato ribulose‐1,5‐bisphosphate carboxylase small subunit gene family. Proc. Natl Acad. Sci. USA, 84, 7104–7108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sui, X. , Shan, N. , Hu, L. , Zhang, C. , Yu, C. , Ren, H. , Turgeon, R. and Zhang, Z. (2017) The complex character of photosynthesis in cucumber fruit. J. Exp. Bot. 68, 1625–1637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tambussi, E.A. , Nogués, S. and Araus, J.L. (2005) Ear of durum wheat under water stress: water relations and photosynthetic metabolism. Planta, 221, 446–458. [DOI] [PubMed] [Google Scholar]
- Tambussi, E.A. , Bort, J. , Guiamet, J.J. , Nogués, S. and Araus, J.L. (2007) The photosynthetic role of ears in C3 cereals: metabolism, water use efficiency and contribution to grain yield. Crit. Rev. Plant Sci. 26, 1–16. [Google Scholar]
- Tanaka, A. , Fujita, K. and Kikuchi, K. (1974) Nutrio‐physiological studies on the tomato plant. J. Soil Sci. Plant Nutr. 20, 57–68. [Google Scholar]
- Tilman, D. and Clark, M. (2015) Food, agriculture and the environment: can we feed the world and save the Earth? Daedalus, 144, 8–23. [Google Scholar]
- Timm, S. , Florian, A. , Arrivault, S. , Stitt, M. , Fernie, A.R. and Bauwe, H. (2012) Glycine decarboxylase controls photosynthesis and plant growth. FEBS Lett. 586, 3692–3697. [DOI] [PubMed] [Google Scholar]
- Tinoco‐Ojanguren, C. (2008) Diurnal and seasonal patterns of gas exchange and carbon gain contribution of leaves and stems of Justicia californica in the Sonoran Desert. J. Arid Environ. 72, 127–140. [Google Scholar]
- Todd, G.W. , Bean, R.C. and Propst, B. (1961) Photosynthesis & respiration in developing fruits. II. Comparative rates at various stages of development. Plant Physiol. 36, 69–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tschiersch, H. , Borisjuk, L. , Rutten, T. and Rolletschek, H. (2011) Gradients of seed photosynthesis and its role for oxygen balancing. Biosystems, 103, 302–308. [DOI] [PubMed] [Google Scholar]
- Vicente, R. , Vergara‐Díaz, O. , Medina, S. , Chairi, F. , Kefauver, S.C. , Bort, J. , Serret, M.D. , Aparicio, N. and Araus, J.L. (2018) Durum wheat ears perform better than the flag leaves under water stress: gene expression and physiological evidence. Environ. Exp. Bot. 153, 271–285. [Google Scholar]
- Vogg, G. , Fischer, S. , Leide, J. , Emmanuel, E. , Jetter, R. , Levy, A.A. and Riederer, M. (2004) Tomato fruit cuticular waxes and their effects on transpiration barrier properties: functional characterization of a mutant deficient in a very‐long‐chain fatty acid b‐ketoacyl‐CoA synthase. Journal of Experimental Botany, 55, 1401–1410. [DOI] [PubMed] [Google Scholar]
- Wang, H. , Schauer, N. , Usadel, B. , Frasse, P. , Zouine, M. , Hernould, M. , Latche, A. , Pech, J.C. , Fernie, A.R. and Bouzayen, M. (2009) Regulatory features underlying pollination‐dependent and ‐independent tomato fruit set revealed by transcript and primary metabolite profiling. Plant Cell, 21, 1428–1452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weber, H. , Borisjuk, L. and Wobus, U. (2005) Molecular physiology of legume seed development. Annu. Rev. Plant Biol. 56, 253–279. [DOI] [PubMed] [Google Scholar]
- Weyhrich, R.A. (1994) Effects of awn suppression on grain yield and agronomic traits in hard red winter wheat. Crop Sci. 34, 965–969. [Google Scholar]
- Weyhrich, R.A. , Carver, B.F. and Martin, B.C. (1995) Photosynthesis and water‐use efficiency of awned and awnletted near‐isogenic lines of hard winter wheat. Crop Sci. 35, 172–176. [Google Scholar]
- Willmer, C.M. and Johnston, W.R. (1976) Carbon dioxide assimilation in some aerial plant organs and tissues. Planta, 130, 33–37. [DOI] [PubMed] [Google Scholar]
- WorldBank (2008) World Development Report 2008: Agriculture for Development. Washington, DC: World Bank. [Google Scholar]
- Wu, X.‐L. , Liu, Z.‐H. , Hu, Z.‐H. and Huang, R.‐Z. (2014) BnWRI1 coordinates fatty acid biosynthesis and photosynthesis pathways during oil accumulation in rapeseed. J. Integr. Plant Biol. 56, 582–593. [DOI] [PubMed] [Google Scholar]
- Xu, H. , Gauthier, L. , Desjardins, Y. and Gosselin, A. (1997) Photosynthesis in leaves, fruits, stem and petioles of greenhouse‐grown tomato plants. Photosynthetica, 33, 113–123. [Google Scholar]
- Yadav, S.K. , Khatri, K. , Rathore, M.S. and Jha, B. (2018) Introgression of UfCyt c6, a thylakoid lumen protein from a green seaweed Ulva fasciata Delile enhanced photosynthesis and growth in tobacco. Mol. Biol. Rep. 45(6), 1745–1758. [DOI] [PubMed] [Google Scholar]
- Yakovlev, M.S. and Zhukova, G.Y. (1980) Chlorophyll in embryos of angiosperm seeds, a review. Bot. Not. Ser. 133, 323–336. [Google Scholar]
- Yiotis, C. , Psaras, G.K. and Manetas, Y. (2008) Seasonal photosynthetic changes in the green‐stemmed Mediterranean shrub Calicotome villosa: a comparison with leaves. Photosynthetica, 46, 262. [Google Scholar]
- Zhou, B. , Serret, M.D. , Elazab, A. , Bort Pie, J. , Araus, J.L. , Aranjuelo, I. and Sanz‐Sáez, Á. (2016) Wheat ear carbon assimilation and nitrogen remobilization contribute significantly to grain yield. J. Integr. Plant Biol. 58, 914–926. [DOI] [PubMed] [Google Scholar]
- Ziegler‐Jöns, A. (1989) Gas‐exchange of ears of cereals in response to carbon dioxide and light. Planta, 178, 164–117. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data presented within this review are example data sets that are not publicly available; please contact T.L. to request access to any data.